lecozotan (sra-333): a selective serotonin 1a receptor antagonist that enhances the stimulated...
TRANSCRIPT

Lecozotan (SRA-333): A Selective Serotonin 1A ReceptorAntagonist That Enhances the Stimulated Release ofGlutamate and Acetylcholine in the Hippocampus andPossesses Cognitive-Enhancing Properties
L. E. Schechter, D. L. Smith, S. Rosenzweig-Lipson, S. J. Sukoff, L. A. Dawson,1
K. Marquis, D. Jones, M. Piesla, T. Andree, S. Nawoschik, J. A. Harder, M. D. Womack,J. Buccafusco, A. V. Terry, B. Hoebel, P. Rada, M. Kelly,2 M. Abou-Gharbia,J. E. Barrett,3 and W. ChildersWyeth, Neuroscience Discovery Research, Princeton, New Jersey (L.E.S., D.L.S., S.R.-L., S.J.S., L.A.D., K.M., D.J., M.P., T.A.,S.N., J.E.B.); School of Pharmacy, University of Bradford, Bradford, West Yorkshire, United Kingdom (J.A.H., M.D.W.); Programin Clinical and Experimental Therapeutics, University of Georgia College of Pharmacy, Medical College of Georgia, Augusta,Georgia (J.B., A.V.T.); Princeton University; Princeton, New Jersey (B.H., P.R.); and Wyeth, Chemical and Screening Sciences,Princeton, New Jersey (M.K., M.A.-G., W.C.)
Received March 17, 2005; accepted June 9, 2005
ABSTRACTRecent data has suggested that the 5-hydroxytryptamine (5-HT)1Areceptor is involved in cognitive processing. A novel 5-HT1Areceptor antagonist, 4-cyano-N-{2R-[4-(2,3-dihydrobenzo[1,4]-dioxin-5-yl)-piperazin-1-yl]-propyl}-N-pyridin-2-yl-benzamide HCl(lecozotan), which has been characterized in multiple in vitro andin vivo pharmacological assays as a drug to treat cognitive dys-function, is reported. In vitro binding and intrinsic activity determi-nations demonstrated that lecozotan is a potent and selective5-HT1A receptor antagonist. Using in vivo microdialysis, lecozotan(0.3 mg/kg s.c.) antagonized the decrease in hippocampal extra-cellular 5-HT induced by a challenge dose (0.3 mg/kg s.c.) of8-hydroxy-2-dipropylaminotetralin (8-OH-DPAT) and had no ef-fects alone at doses 10-fold higher. Lecozotan significantly poten-tiated the potassium chloride-stimulated release of glutamate andacetylcholine in the dentate gyrus of the hippocampus. Chronicadministration of lecozotan did not induce 5-HT1A receptor toler-
ance or desensitization in a behavioral model indicative of 5-HT1Areceptor function. In drug discrimination studies, lecozotan(0.01–1 mg/kg i.m.) did not substitute for 8-OH-DPAT and pro-duced a dose-related blockade of the 5-HT1A agonist discrimina-tive stimulus cue. In aged rhesus monkeys, lecozotan produceda significant improvement in task performance efficiency at anoptimal dose (1 mg/kg p.o.). Learning deficits induced by theglutamatergic antagonist MK-801 [(�)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-imine maleate] (assessed byperceptually complex and visual spatial discrimination) and byspecific cholinergic lesions of the hippocampus (assessedby visual spatial discrimination) were reversed by lecozotan (2mg/kg i.m.) in marmosets. The heterosynaptic nature of the effectsof lecozotan imbues this compound with a novel mechanism ofaction directed at the biochemical pathologies underlying cogni-tive loss in Alzheimer’s disease.
The multiplicity of biological data associated with the 5-hy-droxytryptamine (5-HT)1A receptor subtype, since its discov-ery by radioligand binding in 1981 (Pedigo et al., 1981) andsubsequent cloning in 1988 (Fargin et al., 1988), implicatesthis receptor in numerous behavioral and physiological func-tions, including cognition, psychosis, feeding/satiety, temper-
This research was supported by Wyeth Research (Princeton, NJ and Colle-geville, PA).
1 Current address: Psychiatry Centre of Excellence, GlaxoSmithKline, NewFrontiers Science Park, Harlow Essex, UK; 2 Current address: Renovis, SouthSan Francisco, CA; 3 Current address: Adolor Corporation, Exton, PA.
Article, publication date, and citation information can be found athttp://jpet.aspetjournals.org.
doi:10.1124/jpet.105.086363.
ABBREVIATIONS: 5-HT, 5-hydroxytryptamine; 8-OH-DPAT, 8-hydroxy-2-dipropylaminotetralin; WAY-100135, N-tert-butyl-3-[4-(2-methoxyphenyl)piperazin-1-yl]-2-phe-nylpropanamide; WAY-100635, [O-methyl-3H]-N-(2-(4-(2-methoxyphenyl)-1-piperazinyl)ethyl)-N-(2-pyridinyl)cyclohexanecarboxamide trihydrochloride; NAD-299, (R)-3-N,N-dicyclobutylamino-8-fluoro-3,4-dihydro-2H-1-benzopyran-5-carboxamide hydrogen (2R,3R)-tartrate monohydrate; AD, Alzheimer’s disease; NMDA, N-methyl-D-aspartate;SRA-333, 4-cyano-N-{2R-[4-(2,3-dihydrobenzo[1,4]dioxin-5-yl)-piperazin-1-yl]-propyl}-N-pyridin-2-yl-benzamide HCl (lecozotan); CHO, Chinese hamster ovary; DA, dopa-mine; GTP�S, guanosine 5�-O-(3-thio)triphosphate; aCSF, artificial cerebrospinal fluid; HPLC, high-performance liquid chromatography; ANOVA, analysis of variance; FR,fixed ratio; 5-MeODMT, 5-methoxy-dimethyltryptamine; DMTS, delayed matching-to-sample; MK-801, (�)-5-methyl-10,11-dihydro-5H-dibenzo[a,d]cyclohepten-5,10-iminemaleate; VBD, vertical diagonal band; WGTA, Wisconsin General Test Apparatus; DRN, dorsal raphe neuronal; CI, confidence interval; BMY-7378, 8-[2-[4-(2-methoxyphenyl)-1-piperazinyl]ethyl]-8-azaspiro[4.5]decane-7,9-dione; NAN-190, 1-(2-methoxyphenyl)-4-[4-(2-phthalimido)butyl]piperazine.
0022-3565/05/3143-1274–1289$20.00THE JOURNAL OF PHARMACOLOGY AND EXPERIMENTAL THERAPEUTICS Vol. 314, No. 3Copyright © 2005 by The American Society for Pharmacology and Experimental Therapeutics 86363/3048068JPET 314:1274–1289, 2005 Printed in U.S.A.
1274

ature regulation, anxiety, depression, sleep, pain perception,and sexual activity. The development of 8-hydroxy-2-dipro-pylaminotetralin (8-OH-DPAT) as a selective agonist for the5-HT1A receptor has been instrumental in defining the phys-iological role of the 5-HT1A receptor (Arvidsson et al., 1981)as well as its anatomic localization (Gozlan et al., 1983).However, it was not until the development of a “silent”5-HT1A receptor antagonist that systematic pharmacologicalstudies explored the consequences of blocking this receptor inthe brain.
The first breakthrough in this area was reported in 1993with the development of the arylpiperazine compound WAY-100135, which had moderate potency (Fletcher et al., 1993).The more potent and selective congener WAY-100635 wasidentified shortly thereafter (Forster et al., 1995). Since then,it has been demonstrated that WAY-100635 lacks intrinsicactivity in multiple assay systems that are sensitive tothe agonist effects of 5-HT1A compounds. Accordingly, WAY-100635 antagonizes the responses of 8-OH-DPAT at 5-HT1A
somatodendritic autoreceptors to inhibit the firing rate ofdorsal raphe neurons and antagonizes the ability of 8-OH-DPAT to decrease the accumulation of cAMP at postsynaptic5-HT1A receptors in the hippocampus (Fletcher et al., 1996).Although WAY-100635 remains an important pharmacolog-ical tool in defining 5-HT1A receptor function, other 5-HT1A
antagonists have been developed over the past decade, in-cluding WAY-405 (Schechter et al., 2000), which is anotheraryl-piperazine analog, and NAD-299 (Johansson et al.,1997) and LY-426965 [(2S)-1-cyclohexyl-4-[4-(2-methoxyphe-nyl)-1-piperazinyl]2-methyl-2-phenyl-1-butanone monohy-drochloride] (Rasmussen et al., 2000), which are structuralanalogs of 8-OH-DPAT and pindolol, respectively.
One of the most intriguing areas of research surrounding5-HT1A receptor antagonists is the potential of these com-pounds to enhance cognitive ability. AD has been describedas a neurodegenerative disorder characterized by multipledeficits in neurotransmitter function. Although many studieshave focused upon the cholinergic hypothesis, it has becomeapparent that not all patients can be characterized by deficitsin this system alone, as shown by the moderate efficacyproduced by acetylcholinesterase inhibitors (Morris et al.,1986; Bliss and Collingridge, 1993). In fact, recent researchsuggests that glutamatergic deficits may occur before thoseobserved in the cholinergic system (Bliss and Collingridge,1993). Furthermore, and in contrast to glutamatergic andcholinergic neurotransmission, the serotonergic system maybe hyperactive in the disease as a result of the enhancedturnover of serotonin (Kowall and Beal, 1991), which ulti-mately would reduce the firing of the cortical pyramidal-associated pathways through stimulation of 5-HT1A autore-ceptors. It was this finding of enhanced serotonergic turnoverthat lead Bowen et al. (1994) to the hypothesis that 5-HT1A
receptor antagonists may be effective in treating the cogni-tive loss associated with AD.
A compelling rationale has been proposed for treating thedementia associated with AD based on data that antagonistsof 5-HT1A receptors have a facilitatory effect on glutamater-gic transmission (Bliss and Collingridge, 1993; Bowen et al.,1994). N-Methyl-D-aspartate (NMDA)-induced glutamate re-lease from pyramidal neurons is potentiated by a 5-HT1A
receptor antagonist (Dijk et al., 1995). The 5-HT1A antago-nist WAY-100635 can alleviate cognitive deficits induced by
both glutamatergic dysfunction (dizocilpine treatment-in-duced cognitive deficits) and cholinergic dysfunction (fornixtransaction surgery) in primates (Harder et al., 1996; Harderand Ridley, 2000). The blockade of 5-HT1A receptors alsoenhances glutamate release from rat hippocampal slices(Van den Hooff and Galvan, 1991). Other data suggest that5-HT1A receptor antagonists can inhibit the tonic hyperpo-larizing action of serotonin on pyramidal neurons in both thecortex and hippocampus (Araneda and Andrade, 1991; Vanden Hooff and Galvan, 1992), which ultimately would resultin an enhancement of glutamatergic neurotransmission andsignaling. Thus, it is possible that 5-HT1A receptor antago-nists may improve cognition by removing the inhibitory ef-fects of endogenous serotonin on pyramidal neurons and thusenhance glutamatergic activation and ensuing signal trans-duction.
The purpose of this study was to investigate the pharma-cological characteristics of SRA-333 (lecozotan), a novel, se-lective, and potent antagonist of 5-HT1A receptors. Lecozotanspecifically antagonizes functional responses associated withstimulating the 5-HT1A receptor both in vitro and in vivo.The ability of lecozotan to potentiate the release of acetylcho-line and glutamate in vivo as well as its positive role inrelevant biological models for cognition supports the utility ofthis compound as a novel strategy to treat the cognitivedeficits associated with AD.
Materials and MethodsRadioligand Binding
Membrane Preparations. 5-HT1A receptor. The polymerasechain reaction cloning of the human 5-HT1A receptor subtype from ahuman genomic library has been described previously (Chanda et al.,1993). A stable Chinese hamster ovary cell line (CHO-K1) expressingthe human 5-HT1A receptor subtype (h5-HT1A CHO cells) was usedthroughout this study. Cells were maintained in Dulbecco’s modifiedEagle’s medium supplemented with 10% fetal calf serum, nonessen-tial amino acids, and penicillin/streptomycin. Cells were grown to 95to 100% confluence as a monolayer before membranes were har-vested for binding studies. Cells were gently scraped from the cultureplates, transferred to centrifuge tubes, and washed twice by centrif-ugation (2000 rpm for 10 min at 4°C) in buffer (50 mM Tris, pH 7.5).The resulting pellets were aliquoted and placed at �80°C. On theday of assay, the cells were thawed on ice and resuspended in buffer.
Dopamine receptors. The binding profile of lecozotan was assessedin stable CHO-K1 cells containing dopamine D2 (DA)2, dopamine D3
(DA3), and dopamine D4 (DA4) receptor subtypes. The CHO-K1 cellswere maintained in Dulbecco’s modified Eagle’s medium supple-mented with 10% fetal calf serum, nonessential amino acids, andpenicillin/streptomycin. The cells were grown to 95% confluence as amonolayer and were harvested by scraping. The harvested cells werecentrifuged at low speed (1000g) for 10 min to remove the culturemedia. The harvested cells were suspended in half-volume of freshphysiological phosphate-buffered saline solution and recentrifugedat the same speed. This operation was repeated once more. Thecollected cells were then homogenized in 10 volumes of 50 mMTris-HCl, pH 7.4, and 0.5 mM EDTA. The homogenate was centri-fuged at 40,000g for 30 min, and the precipitate was collected. Thepellet was resuspended in 10 volumes of Tris-HCl buffer and recen-trifuged at the same speed. The final pellet was suspended in a smallvolume of Tris-HCl buffer, and the tissue protein content was deter-mined in aliquots of 10- to 25-�l volumes. Bovine serum albumin wasused as the standard in the protein determination by the method ofLowry et al. (1951). The volume of the suspended cell membraneswas adjusted to give a tissue protein concentration of 1.0 mg/ml
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1275

suspension. The prepared membrane suspension (10 times concen-trated) was aliquoted in 1.0-ml volumes and stored at �70°C untilused in subsequent binding experiments.
�1 Adrenergic receptor. Cortical tissue from male Sprague-Dawleyrats was homogenized on ice in 50 mM Tris-HCl, pH 7.4, buffer witha Polytron homogenizer. The homogenate was then centrifuged at25,000g for 10 min, and the supernatant was discarded. The pelletwas resuspended in fresh buffer, centrifuged, and then resuspended.After a third wash of the membranes, the pellet was resuspended inbuffer, aliquoted into separate vials, and stored at �70°C untilsubsequent assay.
Radioligand Binding Protocols. 5-HT1A receptor binding.[3H]WAY-100635 and [3H]8-OH-DPAT were used to label 5-HT1A
receptors. Experiments were conducted in 96-well microtiter platesin a total volume of 250 �l of buffer (50 mM Tris-HCl, pH 7.4).Nonspecific binding was defined with 10 �M methiothepin for[3H]WAY-100635 binding. Competition studies were conducted us-ing 1 nM ligand. For [3H]8-OH-DPAT binding, nonspecific bindingwas defined with 10 �M serotonin. Competition studies were con-ducted using 1.50 nM of the ligand. The binding assays were initi-ated by the addition of 50 �l of the harvested stabile transfected5-HT1A cells (0.05 mg/sample) and were incubated at 25°C for 30min. The reaction was terminated by vacuum filtration throughpresoaked (0.5% polyethyleneimine) Whatman GF/B filter paper(Brandel Inc., Gaithersburg, MD) using a Brandel 96-cell harvester.Filters were washed with ice-cold buffer (50 mM Tris-HCl, pH 7.4)and transferred to scintillation vials, to which 5 ml of Opti-Fluor(PerkinElmer Life and Analytical Sciences, Boston, MA) was added.Radioactivity was measured by liquid scintillation counting using aBeckman LS 6000TA liquid scintillation counter (Beckman CoulterInc., Fullerton, CA). Protein concentrations were determined by themethod of Bradford using bovine serum albumin as the standard.Measurements were taken at 595 nm with a PerkinElmer Lambda3B spectrophotometer (PerkinElmer Life and Analytical Sciences).Binding data were analyzed by ReceptorFit (Lundon Software,Cleveland Heights, OH), a computer-assisted nonlinear regressionanalysis program.
Dopamine receptor binding. The radioligand [3H]spiperone wasused at 0.5 nM to label DA2, DA3, and DA4 receptors. Nonspecificbinding was determined in the presence of 10 �M d-butaclamol;7-hydroxy-DPAT; or clozapine for DA2, DA3, and DA4, respectively.Binding experiments were performed in a 96-well microtiter plateformat, in a total volume of 200 �l. Competition experiments wereperformed using seven to nine concentrations of lecozotan, added in20-�l volume. To each well was added 80 �l of incubation buffermade in 50 mM Tris-HCl buffer, pH 7.4, containing 10 mM MgCl2and 0.5 mM EDTA, and 20 �l of radioisotope. The reaction wasinitiated by the final addition of 100 �l of tissue suspension. Thereaction proceeded in the dark for 120 min at room temperature, atwhich time the bound ligand-receptor complex was filtered off on a96-well unifilter with a Filtermate 196 harvester (PerkinElmer Lifeand Analytical Sciences). The radioactivity of the bound complexcaught on the filter disk was measured after drying and addition of40 �l of Microscint-20 scintillant to each well in a TopCount(PerkinElmer Life and Analytical Sciences) equipped with six pho-tomultiplier detectors. The unifilter plate was heat sealed andcounted in a TopCount with efficiencies of 31.0 and 60% for tritiumand [125I], respectively.
�1 Adrenergic receptor binding. Membranes were thawed, placedin buffer, and incubated with [3H]prazosin (0.2 nM) for 30 min at25°C. All tubes contained either vehicle, test compound (one to eightconcentrations), or a saturating concentration of phentolamine (10�M) to define specific binding. All reactions were terminated by theaddition of ice-cold Tris buffer followed by rapid filtration using aTomTech filtration device (TomTech, Hamden, CT) to separatebound from free [3H]prazosin. Bound radioactivity was quantitatedusing a Wallac 1205 Beta Plate counter (PerkinElmer Life andAnalytical Sciences).
Nova Screen binding profile. A Nova Screen binding profile wasdetermined for lecozotan at 61 neurotransmitter receptor, reuptake,ion channel, and enzyme sites. [3H]8-OH-DPAT and [3H]WAY-100635 were used as the 5-HT1A agonist radioligand and the 5-HT1A
antagonist radioligand, respectively. Displacement of specific radio-ligand binding at these sites was determined at three concentrationsof lecozotan (1, 100, and 10,000 nM).
Determination of Intrinsic Activity Using a cAMPAccumulation Assay
Assays were performed by incubating the cells with Dulbecco’smodified Eagle’s medium containing 25 mM HEPES, 5 mM theo-phylline, and 10 �M pargyline for a period of 20 min at 37°C.Functional activity was assessed by treating the cells with forskolin(1 �M final concentration) followed immediately by a test compound(six concentrations) for an additional 10 min at 37°C. In separateexperiments, six concentrations of antagonist were preincubated for20 min before the addition of 10 nM 8-OH-DPAT (Sigma/RBI,Natick, MA) and forskolin. The reaction was terminated by removalof the media and addition of 0.5 ml of ice-cold assay buffer. Plateswere stored at �20°C before the assessment of cAMP formation by acAMP SPA assay (Amersham Biosciences Inc., Piscataway, NJ).
Assessment of Intrinsic Activity Using [35S]GTP�S Binding
The guanosine 5�-O-(3-thio)triphosphate ([35S]GTP�S) binding as-say was similar to that used by Lazareno and Birdsall (1993). Briefly,5-HT1A-cloned receptor membrane fragments (as used for 5-HT1A
receptor binding assays) were stored at �70°C until needed. Whenneeded, membranes were rapidly thawed, centrifuged at 40,000g for10 min, and resuspended at 4°C for 10 min in assay buffer [25 mMHEPES, 3 mM MgCl2, 100 mM NaCl, 1 mM EDTA, 10 �M GDP, and500 mM dithiothreitol (Cleland’s Reagent), pH 8.0]. These mem-branes were then incubated for 30 min at 30°C with [35S]GTP�S (1nM) in the presence of vehicle, test compound (one to eight concen-trations), or excess 8-OH-DPAT to define maximum agonist re-sponse. All reactions were terminated by the addition of ice-cold Trisbuffer followed by rapid filtration using a TomTech filtration deviceto separate bound from free [35S]GTP�S. Agonists produced an in-crease in the amount of [35S]GTP�S bound, whereas antagonistsproduced no increase in binding. Bound radioactivity was countedand analyzed as described above.
In Vivo Microdialysis: 5-HT and Glutamate
Subjects and Surgery. Male Sprague-Dawley rats (280–350 g;Charles River Laboratories, Inc., Wilmington, MA) were used in allexperiments. Animals were housed as a group with food and wateravailable ad libitum and maintained on a 12-h light/dark cycle(lights on at 6:00 AM) with all work performed during the lightphase. After surgery, the animals were housed separately in Plexi-glas cages (45 � 45 � 30 cm) with free access to food and water.
Microdialysis Procedures. After induction of anesthesia withgaseous administration of halothane (2%) (Fluothane; Zeneca,Cheshire, UK), the animals were secured in a stereotaxic frame withear and incisor bars. Anesthesia was maintained by continuousadministration of halothane (1 to 2%). A microdialysis probe guidecannula (CMA/Microdialysis, Stockholm, Sweden) was implantedinto either the ventral hippocampus or the dentate gyrus. Coordi-nates for the both brain regions were taken from Paxinos et al.(1985): RC, �4.8; L, �5.0; and V, �4.4 and RC, �3.8; L, �1.4, and V,�3.6, respectively; reference points for microdialysis cannula im-plantation were taken from bregma (RC and L) and the dura (V). Acannula was also implanted s.c. at this time between the animal’sshoulders. Both cannulae were secured to the skull using dentalacrylic (Plastics One, Roanoke, VA). The wound was sutured and theanimals left to recover for 24 h in their home cages with free accessto food and water.
A pre-equilibrated microdialysis probe (optical density 0.5 mm,
1276 Schechter et al.

membrane length 4 or 1 mm; CMA/Microdialysis), perfused over-night in artificial cerebrospinal fluid, was inserted, via the guidecannula, into either the ventral hippocampus or the dentate gyrus,respectively, of the unrestrained rat 24 h after surgery. The probewas perfused with aCSF (125 mM NaCl, 3.0 mM KCl, 0.75 mMMgSO4, and 1.2 mM CaCl2, pH 7.4) at a flow rate of 1 �l/min. A 3-hstabilization period was allowed after probe implantation, afterwhich time microdialysis sampling was carried out by a modificationof the method of Dawson and Routledge (1995). For 5-HT analysis,20-min samples were taken throughout the experimental period, andfour preinjection samples were taken before drug administration.Alternatively, for glutamate determinations, a 5-min sampling reg-imen was used throughout the experimental period, and six presa-mples were taken before drug or vehicle injection to achieve a steadybaseline. All subsequent samples were expressed as a percentage ofthese baseline concentrations. Lecozotan or vehicle was adminis-tered via the cannula (s.c.). Stimulation of neurotransmitter releasewas brought about by infusion of aCSF containing 100 mM KCl (28mM NaCl, 100 mM KCl, 0.75 mM MgSO4, and 1.2 mM CaCl2, pH 7.4)through the microdialysis probe.
Analysis of Microdialysates. 5-HT analysis. 5-HT was sepa-rated by reverse-phase high-performance liquid chromatography(HPLC) (C18 ODS2 column, 100 � 3.0 mm; Metachem, Torrance,CA) and detected using an ANTEC electrochemical detector (AN-TEC, Leiden, The Netherlands) set at a potential of 0.65 V versus aAg/AgCl reference electrode. Mobile phase was delivered by a JascoPU980 HPLC pump (Jasco Ltd., Essex, UK) at 0.6 ml/min andcontained 0.15 M NaH2PO4 buffer at pH 4.3, 0.25 mM EDTA, 1.5 mM1-octane sodium sulfonic acid, and 5% isopropanol.
Glutamate analysis. Measurement of glutamate was performedusing a Crystal 310 capillary electrophoresis system (Thermo Bio-Analysis, Sante Fe, NM) with a Zeta laser-induced fluorescencedetector (ZETA Technology, Toulouse, France) coupled with a Om-nichrome helium-cadmium laser (emission wavelength 442 nm;Melles Griot, Carlsbad, CA). All samples were prederivatized withnaphthalene 2,3-dicarboxaldehyde by a modification of the method ofHernandez et al. (1993). Microdialysate or standard samples (3 �l)were mixed with 50 mM boric acid buffer, pH 9.5, containing 20 mMsodium cyanide (5 �l) and 30 mM naphthalene 2,3-dicarboxaldehydein methanol (1 �l). Samples were allowed to react for 3 min at roomtemperature before injection. Separations were performed accordingto Dawson et al. (1997) in fused silica capillaries (75 �m i.d., 375 �mo.d., 47 cm; Polymicro Technologies, Phoenix, AZ) with an appliedvoltage of 0.6 kV/cm. Samples (5 nl) were applied to the capillary viaa high-pressure injection system. Separations used 30 mM boric acid,pH 9.5 (pH was adjusted using 1 M NaOH). The capillary was rinsedwith 0.1 M NaOH (1.5 min) and running buffer (1.5 min) betweenanalyses. All data were acquired using the Atlas software package(Thermo Labsystems, Gulph Mills, PA) for the PC.
The femtomoles per micromolar perfusate values of 5-HT/gluta-mate for the first four baseline samples were averaged, and thisvalue was denoted as 100%. Results were analyzed by analysis ofvariance (ANOVA) with repeated measures followed by pairwisecomparisons using Bonferroni adjustment for multiple comparisonsusing the Statview software application (Abacus Concepts, Berkeley,CA) for the PC.
In Vivo Microdialysis: Acetylcholine
Subjects and Surgery. Male Sprague-Dawley rats weighing be-tween 350 to 400 g were maintained in individual cages. Animalswere anesthetized with a combination of ketamine (50 mg/kg i.p.)and xylazine (10 mg/kg i.p.), and a single guide shaft aimed at thehippocampus (equal right and left side implants; region CA1) wasimplanted (coordinates RC, �5.6; L, 5.0; and V, 3.0 mm). Rats wereallowed to recover at least a week before experimentation.
Microdialysis Procedure. A concentric microdialysis probe witha 4-mm cellulose tip was inserted in the hippocampus of each rat 14 hbefore the experiment. The probe was perfused with a modified
Ringer’s solution (142 mM NaCl, 3.9 mM KCl, 1.2 mM CaCl2, 1.0 mMMgCl2, and 10 mM bicarbonate, pH 7.4) at a flow rate of 0.5 �l/minovernight and 1 �l/min starting 1 h before the experiment. To en-hance acetylcholine recovery, it was necessary to add a very smallconcentration of neostigmine (0.3 �M) to the Ringer’s solution. Sam-ples were collected every 20 min before and after injection of lecozo-tan.
Analysis of microdialysates. Acetylcholine was measured by re-verse-phase high-performance liquid chromatography with electro-chemical detection using an ESA model 580 pump and mobile phaseof 200 mM potassium phosphate, pH 8.0, at a flow rate of 0.6 ml/min(Rada et al., 2001). Microdialysates were injected into a 20-�l loopleading to a 10-cm C18 analytical column to separate acetylcholine,which was converted to betaine and hydrogen peroxide by an immo-bilized enzyme reactor. Detection was accomplished with an ampero-metric detector (model 400; PerkinElmer Life and Analytical Sci-ences) that oxidized the hydrogen peroxide on a platinum electrode(BAS Bioanalytical Systems, West Lafayette, IN) set at 0.5 V withrespect to a Ag/AgCl reference electrode.
In Vivo Electrophysiology
Male Sprague-Dawley rats (240–265 g; Laboratories, Inc.) werehoused in groups on a 12-h light/dark cycle with ad libitum access tofood and water. On the day of the experiments, rats were initiallyanesthetized with halothane in an induction chamber. Anesthesiawas subsequently maintained by continuously administering halo-thane (in oxygen at 1 to 3%) through a nose cone using a Fluotechalothane vaporizer. A heating pad was placed beneath the animalto maintain the body temperature at 37.5°C.
Rats were placed in a stereotaxic frame and the surface of theskull was exposed. Glass micropipettes (filled with pontamine skyblue) with an impedance of 3 to 6 megaohms were lowered througha craniotomy to a depth of 0.5 mm above the dorsal raphe nucleus (onthe midline and �0.6 mm from the interaural zero). A hydraulicdrive was used to further advance the recording electrode. Neuronalaction potentials were passed through a high-input impedance am-plifier and were monitored on an oscilloscope. Action potentials werediscriminated from background noise and used to trigger an output,which was counted and recorded by an online computer system(DataWave Technology, Longmont, CO).
Dorsal raphe neurons were identified by characteristic wave formactivity and then spontaneous activity was monitored for approxi-mately 10 min to establish a baseline rate of neuronal firing. Apredetermined dose of the test compound was then administeredsubcutaneously and firing rate was monitored for an additional 3min to observe any effect of the test compound. 8-OH-DPAT was thenadministered s.c. in cumulative doses at 3-min intervals. Only oneneuron was studied in each rat to avoid residual effects. At thetermination of the experiment, the pontamine sky blue was depos-ited for 20 min by a 10-�A anodal current for histological confirma-tion of the recording site. Each animal was then perfused intracar-dially with Mirsky’s formula, after which the brain was removed,sectioned at 64 �m, and counterstained with neutral red. Data fromrecording sites not found within the histological boundaries of thedorsal raphe were discarded.
All drugs were put into solution with 0.9% saline. The singlepretreatment dose of lecozotan was in a volume concentration of 300�g/kg/0.1 ml. 8-OH-DPAT was injected in cumulative doses startingat 25 �g/kg in a volume of 0.05 ml. All doses were expressed on thebasis of active moiety. Firing rates for the baseline period and thatafter administration of lecozotan were compared using a paired t test(p � 0.05).
Fixed Ratio (FR) Operant Studies in Rats
Male Sprague-Dawley rats weighing 300 to 350 g were housedindividually and maintained at 85% of their free-feeding body
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1277

weights by food presented during the session and by postsessionfeeding. Water was freely available in the animal’s home cage.
Experimental sessions were conducted in a standard operant con-ditioning chamber placed inside a ventilated sound-attenuating shellthat was equipped with white noise to mask extraneous sounds(MED Associates, Georgia, VT). A response lever and a food troughwere on the front panel of the chamber. Bioserv (Frenchtown, NJ)45-mg precision dustless pellets could be delivered to food troughs toserve as reinforcers. The operant chambers were controlled andmonitored by computers with software from MED Associates.
Rats were trained to respond on the right lever under an FR30schedule of food presentation. Experimental sessions consisted ofthree 10-min components, each preceded by a 10-min time-out periodduring which drugs were administered. During the time-out period,the chamber was dark, and there were no programmed conse-quences. During the response component, the house light was illu-minated, and lever pressing was associated with an audible feedbackclick. 8-OH-DPAT (0.03–0.3 mg/kg) was administered cumulativelys.c. at the start of the time-out periods. Lecozotan was administeredas a 30-min pretreatment before a cumulative 8-OH-DPAT dose-effect curve. The ED50 was defined as the dose of 8-OH-DPAT thatproduced a 50% reduction in the response rate in the presence orabsence of lecozotan. Data from multiple administrations were over-lapped and combined. Rates of responding were calculated sepa-rately in each of the three components of the session by dividing thetotal number of responses by the total time the component was ineffect for each animal.
Fixed Ratio Operant Studies in Squirrel Monkeys
Four adult male squirrel monkeys, Saimiri sciureus, were housedin individual cages except during experimental sessions. Each mon-key had unlimited access to water and received a nutritionally bal-anced diet of Purina Monkey Chow, fresh fruits, vegetables, andvitamin supplements.
Experimental sessions were conducted in ventilated sound-atten-uating chambers (MED Associates) that were provided with whitenoise to mask extraneous sounds. Monkeys sat in a Plexiglas chairsimilar to one used by Kelleher et al. (1972) and faced a panel onwhich a response lever and colored stimulus lamps were mounted.Each press of the lever with a minimal downward force of 0.25 N wasrecorded as a response. A shaved portion of the tail of the monkeywas secured in a stock beneath brass electrodes. Electrode pasteinsured a low-resistance contact between the electrodes and the tail.A brief, low-intensity electric shock (200 ms; 3–5 mA) could bedelivered through the electrodes to the tail.
Responding was maintained by termination of a visual stimulusassociated with an electric shock (Kelleher et al., 1972). In thepresence of a red light, the completion of a 10-response fixed ratiounit (FR10) turned off the red light and initiated a 30-s time-outperiod. If the FR was not completed within a 10-s period, an electricshock was delivered every 10 s; a maximum of 10 shocks was deliv-ered. Daily sessions consisted of five components. Each componentconsisted of a 10-min time-out period during which drugs could beadministered, followed by a 5-min response period during which theFR10 schedule was in effect. Responding during the time-out periodhad no programmed consequences.
Incremental doses of 8-OH-DPAT were administered cumulativelyi.m. at the start of the 10-min time-out period that preceded each ofthe five sequential components. Lecozotan was administered 30 mini.m. or 60 min p.o. before the start of the first response component.
Rates of responding were calculated separately in each of the fivecomponents of the session by dividing the total number of responsesby the total time the component was in effect for each animal. ED50
values were determined for the effects of 8-OH-DPAT alone and afterpretreatment with lecozotan.
Discriminative Stimulus Effects in Pigeons
Six male white Carneaux pigeons, approximately 1-year old, wereobtained from the Palmetto Pigeon Plant (Sunter, SC). All pigeonswere experimentally naive and were maintained individually incages that were provided with continuously available water and grit.Lighting in the temperature- and-humidity-controlled vivarium wason a 12-h light/dark cycle. All pigeons were reduced to approximately85% of their free-feeding body weights before to key peck trainingand were maintained at this weight for the duration of the study.
Experimental sessions were conducted in a standard operant con-ditioning chamber placed inside a ventilated sound-attenuating shellthat was equipped with white noise to mask extraneous sounds(MED Associates). The front panel of the chamber contained threeresponse keys. The keys could be transilluminated with differentcolors. The left and right keys were lit white and used in the presentstudy. Pecks that exceeded approximately 0.15 N on the key oper-ated a feedback relay behind the front wall and were counted as aresponse. Below the center key was a rectangular opening (4.5 � 10cm) that provided access to a solenoid-driven food magazine contain-ing mixed grain. During food delivery, the magazine was illumi-nated.
Pigeons were trained to discriminate 8-OH-DPAT (0.1 mg/kg i.m.)from saline using a two-key grain-reinforced drug discriminationprocedure. Pigeons were initially trained to key peck on a FR sched-ule of food presentation (only one key illuminated at a time) with agradual increase in the FR (final FR � 30) over the training sessions.Pigeons were then trained to respond on one key after a saline (s)injection (i.m.) and on the other key after an 8-OH-DPAT (d) injec-tion (i.m.) using a double-alternation daily injection schedule (s, d, d,s, s, d). When the injection schedule started, both keys were lit, andthe FR was dropped to 5, with a gradual increase to FR30. At thefinal schedule, completion of 30 consecutive responses on the injec-tion-appropriate key resulted in 3-s access to mixed grain. Respond-ing on the incorrect key reset the FR for the injection-appropriatekey. Training and test sessions were 30 min in duration. Criteria forestablishing 8-OH-DPAT as a discriminative stimulus were that thefirst ratio (30 consecutive responses) was completed on the injection-appropriate key and that there was �90% injection-appropriate re-sponse for the total session for five consecutive sessions.
Antagonism studies were conducted once or twice a week in indi-vidual pigeons. For antagonism studies, lecozotan (0.01–1.0 mg/kgi.m.) was administered as a pretreatment (40 min before test session)followed by an injection (i.m.) of the training dose of 8-OH-DPAT (0.1mg/kg 30 min before test session). On test days, completion of 30consecutive responses on either key resulted in access to mixedgrain. Test sessions were conducted only if a pigeon met the criteria(first ratio correct and �90% responding on the injection-appropriatekey) on the previous day and on four of the five previous trainingdays. Training sessions did not take place on days after test sessionsto allow for a washout period for test compounds.
Rates of responding were calculated by dividing the total numberof responses by the total time of the session. Baseline drug key rateswere calculated by averaging rates of drug injection days precedinga test day. Baseline saline key rates were calculated by averagingrates of saline injection days preceding a test day. The percentage ofdrug lever responding was calculated by dividing the number ofresponses on the drug-appropriate key by the total number of re-sponses in the session multiplied by 100. Compounds were consid-ered to fully attenuate the effects of the training dose of 8-OH-DPATin an individual animal if drug key responding was reduced to �20%.Compounds were considered to partially attenuate the effects of thetraining dose of 8-OH-DPAT in an individual animal if the drug keyresponding was reduced to 20 to 60%. If an animal did not receive areinforcer during a test session, the percentage of drug key respond-ing was not analyzed. However, the rate of responding was includedin the analysis.
1278 Schechter et al.

Antagonism of 5-Methoxy-N,N-dimethyltryptamine(5-MeODMT)-Induced Syndrome
Male Sprague-Dawley rats (200–350 g) were housed in groups(four/cage) under a 12-h light/dark cycle with food and water avail-able ad libitum. Serotonin syndrome tests were conducted accordingto a modification of the procedure by Smith and Peroutka (1986).Scoring the serotonin syndrome consisted of rating each of the fol-lowing behaviors: 1) forepaw treading, 2) headweaving, 3) tremor, 4)hindlimb abduction, 5) flattened body posture, and 6) Straub tail ona four-point (0 to 3) ranked intensity scale (maximum score 18). Toevaluate acute effects, lecozotan or vehicle was administered (p.o.) torats (n � 8/dose) placed individually in observation cages (9.5 inwidth � 7.5 in height � 17.5 in length) lined with corncob bedding.Agonist activity was evaluated by scoring subjects for the presence ofthe serotonin syndrome during the final 15 min of the 1-h drugpretreatment interval. Antagonist activity was then determined byscoring for the presence of the serotonin syndrome for 15 min afterchallenging (i.p.) the subjects with 5-MeODMT (5.6 mg/kg), a non-selective 5-HT agonist. This agonist was used because it produced amore robust syndrome that could be completely blocked by full an-tagonists of the 5-HT1A receptor. In a separate experiment, lecozotan(10 mg/kg) or vehicle (1 ml/kg) was administered (p.o.) once daily forseven consecutive days. Twenty-four hours after the last dose, thepotency of 5-MeO-DMT (0.3–5.6 mg/kg; n � 8 per dose) given i.p. wasdetermined in a dose-response study in rats receiving either chronicvehicle or chronic lecozotan. In addition, the potency of lecozotan(1–17 mg/kg; n � 8 per group) given p.o. to antagonize a challengedose of 5-MeO-DMT (5.6 mg/kg i.p.) was evaluated in a separategroup of rats receiving p.o. chronic vehicle or lecozotan (10 mg/kg).
Statistically significant effects were determined by ANOVA fol-lowed by Dunnett’s test (p � 0.05). ED50 values were determinedusing nonlinear regression analysis with inverse prediction.
Aged Rhesus Monkey Task Performance
Male (two) and female (three) rhesus monkeys aged 21 to 29 yearswere well trained (�100 individual sessions) in the delayed match-ing-to-sample (DMTS) task. The animals were maintained on tapwater (unlimited) and standard laboratory monkey chow supple-mented with fruits and vegetables. They were maintained on afeeding schedule such that approximately 15% of their normal daily(except weekends) food intake was derived from 300-mg reinforce-ment food pellets (commercial composition of standard monkey chowand banana flakes) obtained during experimental sessions. The re-mainder was made available after each test session. On weekends,the animals were fed twice per day. The monkeys were maintainedon a 12-h light/dark cycle and were tested each weekday between9:00 AM and 2:00 PM. Room temperature and humidity was main-tained at 72 � 1°C and 52 � 2%, respectively. The aged femaleanimals in the study were perimenopausal, i.e., still cycling, butinfrequently. Testing was performed between menstrual cycles.
Animals were tested simultaneously in their home cages using acomputer-automated training and testing system. Daily sessionsconsisted of 96 trials. Each test unit included four press-key/pelletdispenser units (MED Associates) mounted in lightweight aluminumchassis that could be attached to the home cage. A trial began by theillumination of the sample press key located in the upper center ofthe panel. Monkeys were trained to press the illuminated sample(red, green, or yellow) area to initiate a trial. This action also extin-guished the sample during a computer-specified delay interval. Afterthe delay interval, the choice press keys (two), but not the sample,were illuminated. One of the two choices matched the sample,whereas the other (incorrect) choice was presented as one of the tworemaining colors. Correct trials (matches) were rewarded by thedelivery of a reinforcement food pellet (intertrial interval 5 s). Thevarious combinations of stimulus color were arranged so that eachoccurred an equal number of times as a sample, each color occurredan equal number of times as choices, and each color occurred an
equal number of times in combination with each other color. Like-wise, when two colors occurred in combination, each color was coun-terbalanced between left and right sides of the screen in a nonpre-dictable pattern. Finally, all stimulus-counterbalancing procedureswere matched to length of delay interval. For each subject, baselineaccuracy was normalized by adjusting the length of delay intervals toprovide: zero delay (85–100% correct); short delay (75–84% correct);medium delay (65–74% correct), and long delay (55–64% correct).Baseline data were obtained after the administration of a drugvehicle (sterile saline).
Data for percentage of correct were subdivided according to delayinterval for each 24-trial delay component of the session. All statis-tical analyses were performed on raw data (percentage of trialscorrect). Data were analyzed by a multifactorial ANOVA with re-peated measures (SAS; JMP statistical software package). An or-thogonal multicomparison t test was used to compare individualmeans. Differences between means from experimental groups wereconsidered significant at the p � 0.05 level (two-sided test).
Cognitive-Enhancing Profile of Lecozotan in Marmosets
Animals. Common marmosets (Callithrix jacchus), bred andhoused in the primate colony at the University of Bradford School ofPharmacy, were used for these studies. All marmosets were housedin mixed sex pairs, in which the male animals were vasectomized atleast 3 months before any behavioral studies. Marmosets were fedand watered ad libitum throughout the studies. Their diet consistedof a mixture of commercially available primate “rings” and primate“forage mix” (Harlan UK Limited, Bicester, Oxon, UK), plus variedfruits, multiseed bread, and occasional sweet treats in the form ofgum feeders.
Treatments. All drugs were administered (i.m.) with saline as thevehicle. Injection volumes were kept at a maximum of 0.1 ml permarmoset per drug per day. MK-801 (dizocilpine maleate) was ob-tained from Sigma-Aldrich (St. Louis, MO). Both drugs were dilutedin 0.9% sterile saline. Dizocilpine was administered at a dose of 15�g/kg. Lecozotan was administered at doses of 1 and 2 mg/kg. Dizo-cilpine was administered as a 30-min pretreatment on each daywhen animals were learning tasks under either dizocilpine treat-ment alone or dizocilpine � lecozotan treatment. Lecozotan wasadministered as a 20-min pretreatment on days when animals werelearning tasks under either lecozotan treatment alone or under dizo-cilpine � lecozotan treatment.
All animals received all treatments in a counter-balanced order.Dosing was semichronic; e.g., when an animal took 2 weeks to learna task, that animal would receive 2 weeks of daily injections duringthe period when the task was being acquired. If an animal took amonth to learn a task, then drugs were administered throughout themonth of testing on a daily basis.
Lesion Surgery. We have previously demonstrated that a signif-icant and highly specific lesion of the marmoset vertical diagonalband (VDB), the source of the majority of cholinergic input to thehippocampus, is possible using a p75-saporin conjugate (Harder etal., 1998). An antibody to the p75 human low-affinity nerve growthfactor receptor, thought to be found mainly on cholinergic cell bodies,is conjugated to the neurotoxin saporin. This preparation is commer-cially available through Advanced Targeting Systems as ME 20.4-SAP. The preparation was diluted to 2 �g/�l in sterile 0.9% saline.
Marmosets were sedated with 0.05 ml of ketamine solution (Veta-lar; Aveco Co. Inc., Fort Dodge, IA), before being weighed and anes-thetized with Saffan. Marmosets were placed in a pouch of bubblewrap on a warmed operating table with their heads placed in astereotaxic frame. A small (approximately 2.5 cm) incision from thecenter of the forehead running posteriorly to the back of the head wasmade, and the skin retracted on either side. The stereotaxic framewas centered to bregma, and burr holes were drilled using a smalldental drill to allow placement of 1.5 �l of immunotoxin in eachhemisphere at the following coordinates: AP, �1.15 cm; L, �0.08 cm;V, �0.83 cm and AP, �1.20 cm; L, �0.05 cm; V, �0.98 cm. After
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1279

suturing and recovery in a heated secure box, marmosets were re-turned to the home cage for behavioral testing, beginning 2 weekspostoperatively.
Lesion Verification. Upon completion of the study the lesionsadministered to the subjects were verified to ensure bilateral lesionshad been achieved in the marmosets. Lesion verification wasachieved exactly as described by the method of Ridley et al. (1999).Briefly, after behavioral testing was completed the lesion procedurewas validated using an antibody stain for the p75 nerve growthfactor expressed on cholinergic neurons. Sections were examinedunder a light microscope at 25� magnification. All animals that weretested in the behavioral procedure were validated and had a com-plete bilateral loss of cholinergic staining in the hippocampi, whichcorrelated with the cognitive deficits observed.
Behavioral Testing. Previous studies in many laboratories havemade extensive use of the Wisconsin General Test Apparatus(WGTA) adapted for use with marmosets. Two WGTA task typesused in these studies are described below. Their neural substrateshave been well delineated (Harder et al., 1998; Ridley et al., 1999).
Cortical task: the perceptually complex visual discrimination task.Previously, we have shown that acquisition of perceptually simplevisual discrimination tasks is not robustly affected by dizocilpine,but if the simple visual discrimination task is made more perceptu-ally difficult, a robust deficit is seen (Harder et al., 1998). Hence, inthis study marmosets were only tested on perceptually complexversion tasks. Objects were painted matte black using car spraypaint, such that only the shape could be used to distinguish betweendifferent objects (color and textural differences being eliminatedthrough the painting).
Two different objects were presented to the marmoset in theWGTA: one situated over the right food well, and the other over theleft food well. For every task, two different objects were used, and ineach case, only one of the objects was associated with a reward. Thus,for example, object A might be the cap of a pen, painted black, andobject B might be the small plastic toy from a cereal box, also paintedblack. On any single trial, the marmoset would see the two objects, Aand B, positioned according to a pseudorandom schedule over theright and left food wells. On trial 1, A (the pen cap) might be on theright, and B (the toy) on the left. On trial 2, B might be on the rightand A on the left.
Marmosets chose an object by moving the object to reveal thereward underneath, if the correct object was chosen. In the case ofvisual discrimination tasks, the task is “evaluative”; i.e., it involvesthe formation of an association between a particular object andreward (Ridley et al., 1999). Thus, in the example above, the pen capwas always situated over the food reward, whether presented on theright or the left, and the toy was never situated over the reward.
Over a number of trials, marmosets acquired the association, andeventually selected the “correct” object (A, the pen cap) with suffi-cient accuracy (90% correct) to “pass” the task and move onto thenext task in the experimental design schedule.
Hippocampal task: the visuospatial discrimination task. Visuospa-tial discrimination tasks require a different type of learning from thevisual discrimination task described above. Visuospatial tasks re-quire “conditional” (rather than associative) learning to take placefor successful completion (Ridley and Baker, 1991), because all ob-jects are equally associated with reward, and a “rule” must belearned instead of a simple association.
Pairs of identical objects are used in this instance. For every task,two pairs of identical objects were used, and in each trial, twoidentical objects were seen by the marmoset: one over the left foodwell and the other over the right food well. For example, objects Yand Y may be identical colored key rings, whereas objects Z and Zmight be identical Eppendorf tubes. On any single trial, the marmo-set could see either the two key rings, or the two Eppendorf tubes.The choice of which pair was seen was again determined by a pseu-dorandom schedule.
Marmosets choose an object by moving it to reveal the reward (or
lack thereof) hidden underneath. The conditional rule that must beadhered to was that when objects Y and Y were seen, the rewardwould be hidden under the rightmost of the two identical objects;when objects Z and Z were seen, the reward would be hidden underthe leftmost example of object Z. So in the example given above,when the key rings were seen, food was under the right key ring, andwhen the Eppendorf tubes were seen, food was under the left tube.Marmosets had to reach a criterion of 90% correct to pass the taskand move onto the next.
Study Design. The first stage of the study was to “shape” themarmosets, i.e., to take experimentally naive animals and trainthem to respond to objects presented in the WGTA. All animals weretrained with an identical series of tasks: two perceptually simplevisual discrimination tasks; the first taken to a criterion of 90 correctof 100 consecutive trials (90/100) and the second to a criterion of 27correct of 30 consecutive trials (27/30). This was followed by twovisuospatial discrimination tasks; the first to a criterion of 90/100and the second to a criterion of 27/30. Last, animals learned twoperceptually complex (black object) visual discrimination tasks to acriterion level of 27/30 in both cases.
Once these tasks had been successfully mastered, marmosets wereassigned to either the perceptually complex visual discriminationstudy or the visuospatial discrimination study, such that there wasan equal spread of learning ability across the two studies [by bal-ancing learning scores (number of trials to achieve criterion) betweengroups]. Other marmosets, after completing the same training, wereassigned to either the surgery or the control group, in such a waythat there was an equal spread of learning ability between the twogroups before surgery took place.
In all experiments, the marmosets learned a series of consecutivetasks using new objects for each task. All tasks were presented in thesame order, such that interference between stimuli was kept identi-cal for all monkeys. Treatments were given daily for the duration oflearning task (as described above). Learning scores (the number oftrials before the 27/30 correct criterion was reached) were averagedfor each animal under each treatment to balance for practice effectsacross the tasks. At the end of the study, mean data for each animalunder each treatment were analyzed, and the control group wascompared with the lesion group.
Data Analysis
A repeated measures ANOVA was used to analyze the data, usinga Statview II software package on an Apple Macintosh computer.The mean learning scores for each marmoset under each treatmentprovided the raw data. Post hoc analyses included Dunnett’s t test(for comparing various treatments to control/vehicle data) andScheffe’s F test (for comparing different treatments to one another).In all cases, p � 0.05 indicated a significant difference.
All animal procedures were approved by the Animal Care Com-mittees of Princeton University and Wyeth Research and conformedwith the Guide for the Care and Use of Laboratory Animals asadopted and promulgated by the National Institutes of Health.
Materials
All chemicals were of analytical grade and were purchased fromAldrich Chemical Co. (Milwaukee, WI). 8-OH-DPAT was purchasedfrom Sigma/RBI. Lecozotan was synthesized by Chemical Sciences(Wyeth Research, Princeton, NJ).
ResultsIn Vitro Profile and Structure
The chemical structure of lecozotan is shown in Fig. 1.Lecozotan displayed high-affinity binding at the cloned hu-man 5-HT1A receptor (Ki � 1.6 � 0.3 and 4.5 � 0.6 nM) using[3H]8-OH-DPAT as the 5-HT1A agonist radioligand and[3H]WAY-100635 as the 5-HT1A antagonist radioligand, re-
1280 Schechter et al.
spectively (Table 1). Lecozotan was selective for the 5-HT1A
receptor over other monoamine receptors that have previ-ously been shown to be cross-reactive with the aryl-pipera-zine pharmacophore. A Nova Screen profile revealed thatgreater than 50% displacement was only recorded at thehighest concentration of 10,000 nM at 22 sites, indicatingthat lecozotan demonstrates significant selectivity for the5-HT1A receptor. Lecozotan did not exhibit significant activ-ity (�50% inhibition at any concentration) at any of the other61 binding sites that were evaluated at that concentrationexcept for dopamine D4 (inhibition 63%). Most notably, thecompound was not active at various �-adrenergic (1A, 1B, 2A,2B, and 2C), �-adrenergic (1 and 2), adenosine (1–3), dopa-mine (1, 2, 3, and 5), histamine (1–3), muscarinic (1–5), orserotonin (1B, 1D, 2A, 2C, and 3–7) receptors.
In a clonal CHO cell line expressing the human 5-HT1A
receptor, lecozotan antagonized the ability of 8-OH-DPAT toinhibit forskolin-stimulated cAMP production. In this study,the response to 10 nM 8-OH-DPAT, representing a submaxi-mal concentration, was evaluated in the presence of increas-ing concentrations of antagonist. The calculated IC50 for leco-zotan was 25.1 � 5.3 nM (Table 1). These results werecorroborated using a [35S]GTP�S assay where lecozotan alsoblocked the agonist effects of 8-OH-DPAT (IC50 � 36.7 � 0.74nM). Lecozotan alone did not induce any agonist-like activityunder any assay conditions.
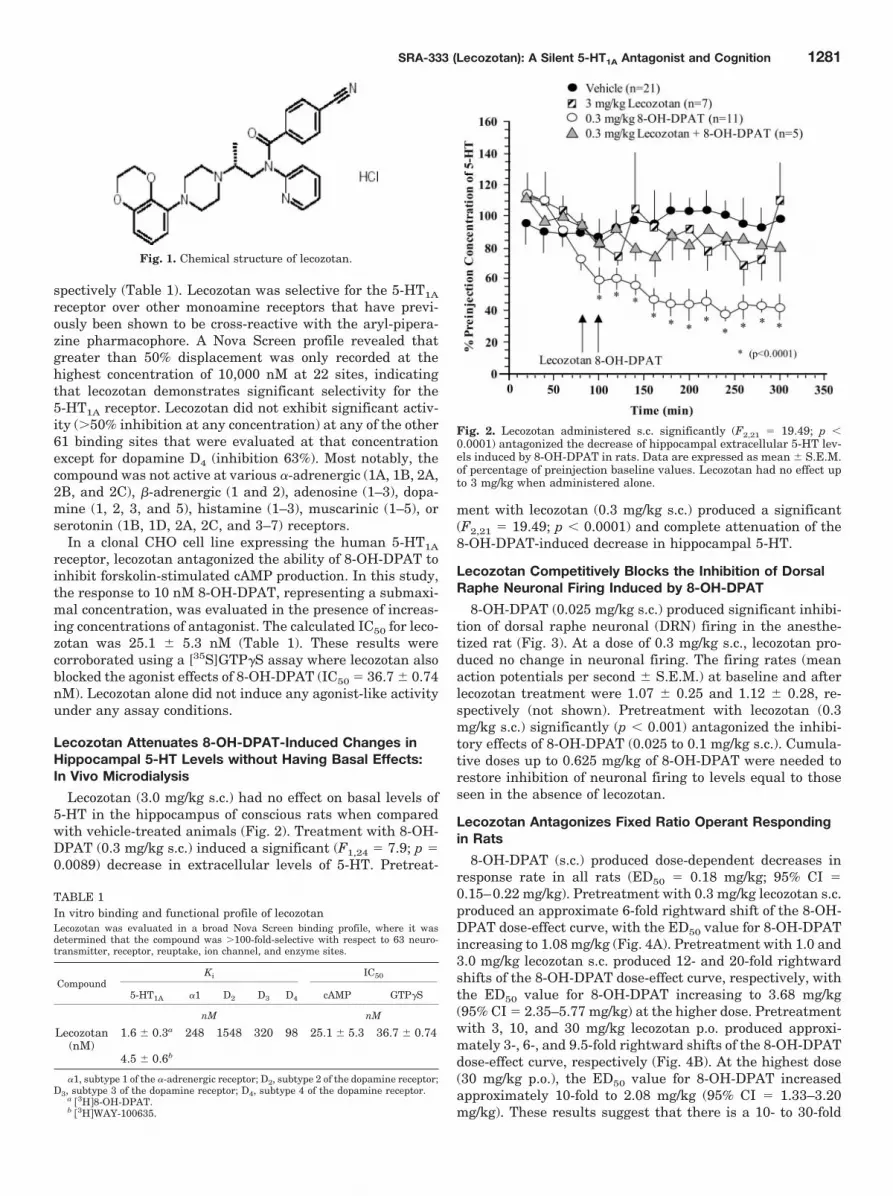
Lecozotan Attenuates 8-OH-DPAT-Induced Changes inHippocampal 5-HT Levels without Having Basal Effects:In Vivo Microdialysis
Lecozotan (3.0 mg/kg s.c.) had no effect on basal levels of5-HT in the hippocampus of conscious rats when comparedwith vehicle-treated animals (Fig. 2). Treatment with 8-OH-DPAT (0.3 mg/kg s.c.) induced a significant (F1,24 � 7.9; p �0.0089) decrease in extracellular levels of 5-HT. Pretreat-
ment with lecozotan (0.3 mg/kg s.c.) produced a significant(F2,21 � 19.49; p � 0.0001) and complete attenuation of the8-OH-DPAT-induced decrease in hippocampal 5-HT.
Lecozotan Competitively Blocks the Inhibition of DorsalRaphe Neuronal Firing Induced by 8-OH-DPAT
8-OH-DPAT (0.025 mg/kg s.c.) produced significant inhibi-tion of dorsal raphe neuronal (DRN) firing in the anesthe-tized rat (Fig. 3). At a dose of 0.3 mg/kg s.c., lecozotan pro-duced no change in neuronal firing. The firing rates (meanaction potentials per second � S.E.M.) at baseline and afterlecozotan treatment were 1.07 � 0.25 and 1.12 � 0.28, re-spectively (not shown). Pretreatment with lecozotan (0.3mg/kg s.c.) significantly (p � 0.001) antagonized the inhibi-tory effects of 8-OH-DPAT (0.025 to 0.1 mg/kg s.c.). Cumula-tive doses up to 0.625 mg/kg of 8-OH-DPAT were needed torestore inhibition of neuronal firing to levels equal to thoseseen in the absence of lecozotan.
Lecozotan Antagonizes Fixed Ratio Operant Respondingin Rats
8-OH-DPAT (s.c.) produced dose-dependent decreases inresponse rate in all rats (ED50 � 0.18 mg/kg; 95% CI �0.15–0.22 mg/kg). Pretreatment with 0.3 mg/kg lecozotan s.c.produced an approximate 6-fold rightward shift of the 8-OH-DPAT dose-effect curve, with the ED50 value for 8-OH-DPATincreasing to 1.08 mg/kg (Fig. 4A). Pretreatment with 1.0 and3.0 mg/kg lecozotan s.c. produced 12- and 20-fold rightwardshifts of the 8-OH-DPAT dose-effect curve, respectively, withthe ED50 value for 8-OH-DPAT increasing to 3.68 mg/kg(95% CI � 2.35–5.77 mg/kg) at the higher dose. Pretreatmentwith 3, 10, and 30 mg/kg lecozotan p.o. produced approxi-mately 3-, 6-, and 9.5-fold rightward shifts of the 8-OH-DPATdose-effect curve, respectively (Fig. 4B). At the highest dose(30 mg/kg p.o.), the ED50 value for 8-OH-DPAT increasedapproximately 10-fold to 2.08 mg/kg (95% CI � 1.33–3.20mg/kg). These results suggest that there is a 10- to 30-fold
Fig. 1. Chemical structure of lecozotan.
TABLE 1In vitro binding and functional profile of lecozotanLecozotan was evaluated in a broad Nova Screen binding profile, where it wasdetermined that the compound was �100-fold-selective with respect to 63 neuro-transmitter, receptor, reuptake, ion channel, and enzyme sites.
CompoundKi IC50
5-HT1A �1 D2 D3 D4 cAMP GTP�S
nM nM
Lecozotan(nM)
1.6 � 0.3a 248 1548 320 98 25.1 � 5.3 36.7 � 0.74
4.5 � 0.6b
�1, subtype 1 of the �-adrenergic receptor; D2, subtype 2 of the dopamine receptor;D3, subtype 3 of the dopamine receptor; D4, subtype 4 of the dopamine receptor.
a 3H8-OH-DPAT.b 3HWAY-100635.
Fig. 2. Lecozotan administered s.c. significantly (F2,21 � 19.49; p �0.0001) antagonized the decrease of hippocampal extracellular 5-HT lev-els induced by 8-OH-DPAT in rats. Data are expressed as mean � S.E.M.of percentage of preinjection baseline values. Lecozotan had no effect upto 3 mg/kg when administered alone.
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1281

separation between the effects of subcutaneously and orallyadministered lecozotan.
In an experiment to investigate the pharmacodynamictime course of antagonism, it was determined that significant(p � 0.05) blockade of 8-OH-DPAT by lecozotan was presentas long as 4 h after administration of lecozotan (1 mg/kg s.c.),data not shown. A single dose of 0.3 mg/kg 8-OH-DPAT s.c.decreased the response rate to 0.74 responses/s in the firstcomponent of the session (10 to 20 min after administrationof 8-OH-DPAT). Rates of responding gradually returned tobaseline rates across the 1-h session with a response rate of2.33 responses/s in the third component of the session (50 to60 min after administration of 8-OH-DPAT). After a 2- or 4-hpretreatment with lecozotan (1.0 mg/kg s.c.), the decreases in
response rates observed after 0.3 mg/kg 8-OH-DPAT alonewere completely antagonized for the entire session, with re-sponse rates greater than 2 responses/s in all three responsecomponents.
Lecozotan Antagonizes Fixed Ratio Operant Respondingin Squirrel Monkeys
8-OH-DPAT i.m. produced a dose-dependent decrease inresponse rates in all monkeys (ED50 value of 0.07 mg/kg; 95%CI � 0.04–0.11 mg/kg). Lecozotan administered at doses of0.01 and 0.1 mg/kg i.m. produced dose-dependent rightwardshifts of the dose-effect curve for 8-OH-DPAT (Fig. 5A). Atthe highest dose (0.1 mg/kg), the ED50 value for 8-OH-DPATwas shifted approximately 17-fold to 1.21 mg/kg (95% CI �0.45–3.25 mg/kg). Similar shifts in the 8-OH-DPAT dose-effect curves were produced by the oral administration oflecozotan at doses of 0.1, 0.3, and 1.0 mg/kg (Fig. 5B). At thehighest dose (1.0 mg/kg), the ED50 value for 8-OH-DPAT wasshifted approximately 10-fold to 0.68 mg/kg (95% CI � 0.27–1.72 mg/kg).
Lecozotan Blocks the Discriminative Stimulus Effects of8-OH-DPAT in Pigeons
Once animals were trained to the stimulus cue, animalsresponded almost exclusively (�98%) on the drug-appropri-ate key after an injection of 0.1 mg/kg 8-OH-DPAT and on thesaline-appropriate key (�2% drug-key responses) after aninjection of saline (Fig. 6). Subsequently, lecozotan was as-sessed for its ability to block the stimulus cue evoked by anacute injection of 8-OH-DPAT. Lecozotan was administeredas a pretreatment (0.01–1 mg/kg i.m.) to the training dose of0.1 mg/kg 8-OH-DPAT i.m. Lecozotan dose dependently de-creased the percentage of responses on the 8-OH-DPAT ap-propriate key, consistent with its actions as a 5-HT1A recep-tor antagonist (Fig. 6). Response completely shifted to thesaline-appropriate key after administration of 1.0 mg/kg leco-zotan. Response rates were unaffected by lecozotan whengiven alone at doses of 0.01 to 1.0 (data not shown).
Fig. 3. Effect of lecozotan on 8-OH-DPAT-induced inhibition of DRNfiring in the anesthetized rat. After lecozotan pretreatment (0.3 mg/kg),the dose of 8-OH-DPAT needed to demonstrate comparable inhibitionincreased to a cumulative dose of 0.625 mg/kg. �, p � 0.001 vs. 8-OHD-PAT alone.
Fig. 4. 8-OH-DPAT (s.c.) produced decreasesin fixed ratio operant response rates in allrats. Pretreatment with lecozotan adminis-tered s.c. (A) and p.o. (B) shifted the 8-OH-DPAT dose effect curve to the right. Valuesare means � S.E.M.
1282 Schechter et al.

Lecozotan Antagonizes the 5-Methoxy-dimethyltryptamine-Induced Syndrome in the Rat
The 5-HT syndrome is mediated by postsynaptic 5-HT1A
receptors and is considered a model of central 5-HT1A recep-tor stimulation. The 5-HT syndrome for these studies wasinduced by 5-MeODMT at a dose of 5.6 mg/kg i.p. Lecozotan(10 mg/kg) or vehicle (1 ml/kg) were given p.o. once daily forseven consecutive days to rats (Tables 2 and 3). Twenty-fourhours after the last dose, the antagonist potency was deter-mined using the 5-MeODMT-induced syndrome. ID50 was4.26 mg/kg (95% CI � 2.23–8.13) after chronic lecozotanadministration, which did not differ significantly from the
acute potency or the potency after chronic vehicle (4.92 mg/kg; 95% CI � 2.70–9.00). Lecozotan exhibited no agonistactivity by itself when administered up to 17 mg/kg. Thus,chronic lecozotan treatment did not alter the antagonist po-tency in this model.
In separate experiments, the agonist potency of5-MeODMT was evaluated 24 h after the last dose. An ED50
value of 1.83 mg/kg (95% CI � 1.37–2.43) was calculatedafter i.p. administration of chronic vehicle. An ED50 value of1.68 mg/kg (95% CI � 1.00–2.82) was observed after chronici.p. administration of lecozotan (10 mg/kg). Thus, chroniclecozotan treatment did not alter the sensitivity of 5-HT1A
receptors.
Effects of Lecozotan on Basal and Stimulated GlutamateRelease from the Dentate Gyrus
Lecozotan produced a significant (F2,17 � 4.52; p � 0.027)augmentation of K�-induced increase in extracellular gluta-mate concentrations within the dentate gyrus of the rat.
Fig. 5. 8-OH-DPAT (i.m.) produced decreases in fixed ratio operant response rates in all squirrel monkeys. Pretreatment with lecozotan administeredi.m. (A) and p.o. (B) shifted the 8-OH-DPAT dose effect curve to the right. Values are means � S.E.M.
Fig. 6. In a discriminative stimulus effects study using pigeons, lecozotan(0.01–1.0 mg/kg) dose dependently decreased the percentage of responseson the 8-OH-DPAT-appropriate key, consistent with its actions as a5-HT1A receptor antagonist. Values are means � S.E.M. percentage ofdrug key responding. 8-OH-DPAT (i.m.) training dose was 0.1 mg/kg.Saline was used as a vehicle.
TABLE 2Potency of lecozotan to antagonize 5-MeODMT-induced syndrome afterchronic lecozotan
Chronic Treatment ID50 (95% CI)
mg/kg
Vehicle (1 ml/kg p.o.) 4.92 (2.70–9.00)Lecozotan (10 mg/kg p.o.) 4.26 (2.23–8.13)
TABLE 3Potency of 5-MeODMT to induced syndrome after chronic lecozotan
Chronic Treatment ED50 (95% CI)
mg/kg
Vehicle (1 ml/kg i.p.) 1.83 (1.37–2.43)Lecozotan (10 mg/kg i.p.) 1.68 (1.00–2.82)
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1283

Administration s.c. of lecozotan (1.0 mg/kg) alone producedno change in basal levels of glutamate within the dentategyrus (Fig. 7). Infusion of aCSF containing 100 mM KClproduced a small but nonsignificant (p � 0.053) increase inextracellular glutamate levels, reaching a maximum value of153.4 � 51.6% of preinfusion control levels. Pretreatment s.c.with a single dose of lecozotan (0.3 mg/kg) produced a signif-icant potentiation of K�-induced increases in extracellularglutamate with a maximum value of 471.5 � 83.1% of pre-infusion levels (p � 0.012).
Effects of Lecozotan on Basal and StimulatedAcetylcholine Release from the CA1 Region of theHippocampus
Lecozotan administered s.c. at a dose of 1 mg/kg had asmall but significant effect on basal acetylcholine levels(146 � 5%) in the CA1 region of the hippocampus comparedwith vehicle (125 � 7%). When a dose of 1 mg/kg lecozotanwas administered s.c. before a 10-min infusion of KCl (100mM), extracellular acetylcholine levels increased (p � 0.05)to 275 � 27% over basal levels (Fig. 8).
Lecozotan Enhances Cognitive Performance in the AgedRhesus Monkey
Vehicle treatment was associated with a significant (F3,40
� 35.7; p � 0.001) delay-dependent reduction in task accu-racy that approximated the target accuracies indicated underMaterials and Methods. Figure 9A shows the composite dataset for the effects of lecozotan i.m. on DMTS performanceefficiency. Statistical analysis of the data revealed no signif-icant effect of treatment alone (F4,40 � 1.03; p � 0.40), and nosignificant effect of treatment’s interaction with delay inter-val (p � 0.49). There also were no significant effects of treat-ment for sessions run 24 h after drug administration (datanot shown). On average, however, improvement in task per-formance efficiency was maintained for the 0.3 and the 1mg/kg doses for trials associated with short delay intervals.
The optimal (best) dose was defined as the dose thatevoked the greatest task improvement independent of delayinterval. Three subjects received overall (all 96 trials) benefit(overall task accuracy greater than respective vehicle level)from two of the five doses. One subject received benefit fromthree of the five doses, and one subject received benefit fromonly one dose. Therefore, none of the subjects could be con-sidered a nonresponder under the conditions of the experi-ment. Most of the average task improvement was noted inthe middle of the dose-response relationship: two animalsreceived most improvement from the 1 mg/kg dose, and theother three animals’ best doses were 0.1, 0.3, and 1 mg/kg.The average best dose for the study group was 1.1 mg/kg.
To reduce the likelihood of false positives, the best dosewas repeated on one additional occasion. Both sessions wereaveraged, and the data are presented in Fig. 9B. There was asignificant effect of drug treatment, independent of delayinterval (F1,44 � 7.31; p � 0.01). When improvement in taskefficiency was calculated as the percentage increase frombaseline levels of performance, after best dose, accuracy in-creased by 16.5% of control for trials associated with shortdelay intervals. Although the treatment times delay effectwas not statistically significant, the improvement in accu-racy associated with short delay trials accounted for 45% ofthe total task improvement.
In addition to task accuracies, two response latencies weremeasured: one to initiate the trial and one to initiate a choice.Drug treatment did not significantly affect (F2,40 � 2.19; p �
Fig. 7. Effects of lecozotan on basal and K�-stimulated glutamate levelsin the dentate gyrus of rats. Data are expressed as means � S.E.M. ofpercentage of preinjection levels of glutamate; n � 6 to 8 per study group.Lecozotan significantly (F2,17 � 4.5; p � 0.027) augmented the K�-stim-ulated increases in extracellular glutamate within the dentate gyrusreaching a maximum value of 471.5 � 83.1% at 120 min (p � 0.012).Asterisk (�) denotes statistical (p � 0.05) difference between K�-stimu-lated versus lecozotan � K�-stimulated responses.
Fig. 8. Lecozotan (1.0 mg/kg s.c.) produced a significantpotentiation of 100 mM K�-induced increases in extra-cellular acetylcholine levels in the CA1 region of thehippocampus. Values are means � S.E.M. Asterisk (�)indicates significant difference (p � 0.05) comparedwith vehicle.
1284 Schechter et al.
0.13) the duration of either response latency (data notshown).
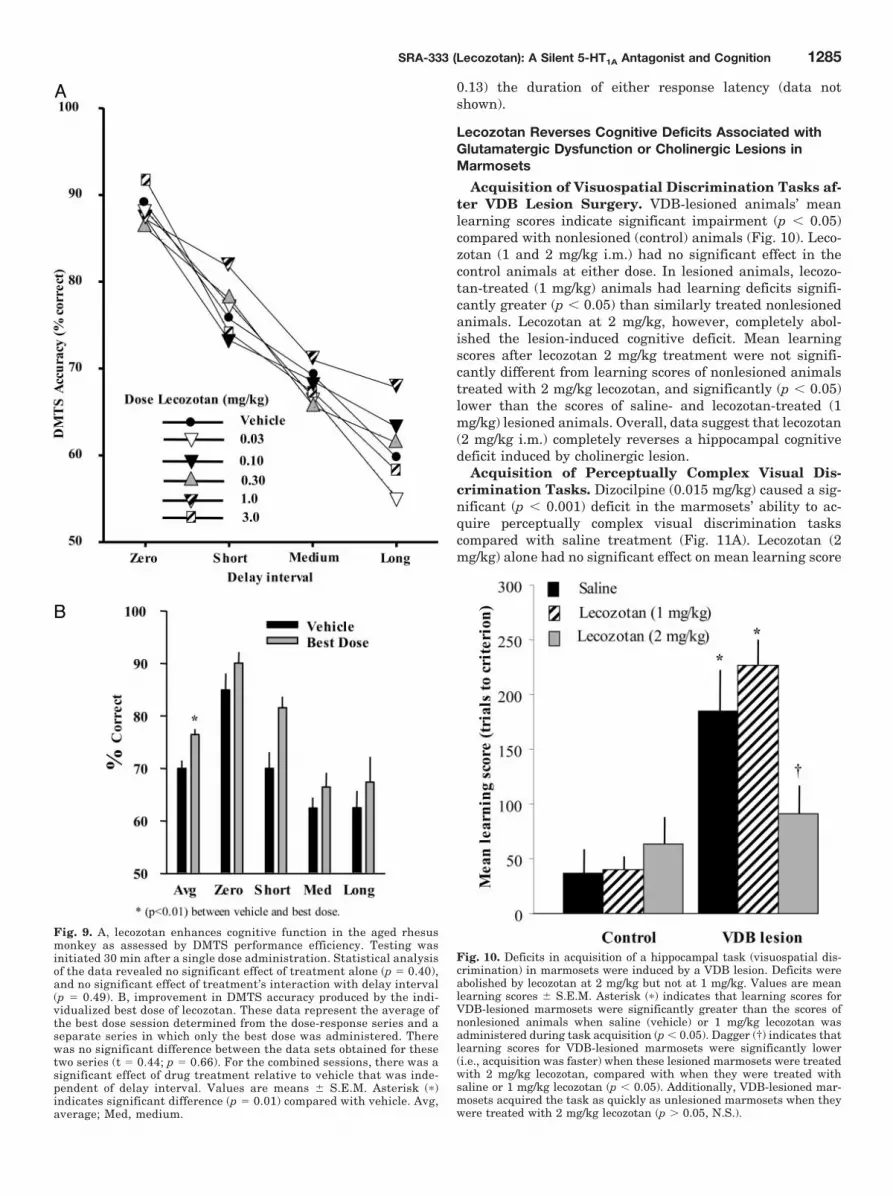
Lecozotan Reverses Cognitive Deficits Associated withGlutamatergic Dysfunction or Cholinergic Lesions inMarmosets
Acquisition of Visuospatial Discrimination Tasks af-ter VDB Lesion Surgery. VDB-lesioned animals’ meanlearning scores indicate significant impairment (p � 0.05)compared with nonlesioned (control) animals (Fig. 10). Leco-zotan (1 and 2 mg/kg i.m.) had no significant effect in thecontrol animals at either dose. In lesioned animals, lecozo-tan-treated (1 mg/kg) animals had learning deficits signifi-cantly greater (p � 0.05) than similarly treated nonlesionedanimals. Lecozotan at 2 mg/kg, however, completely abol-ished the lesion-induced cognitive deficit. Mean learningscores after lecozotan 2 mg/kg treatment were not signifi-cantly different from learning scores of nonlesioned animalstreated with 2 mg/kg lecozotan, and significantly (p � 0.05)lower than the scores of saline- and lecozotan-treated (1mg/kg) lesioned animals. Overall, data suggest that lecozotan(2 mg/kg i.m.) completely reverses a hippocampal cognitivedeficit induced by cholinergic lesion.
Acquisition of Perceptually Complex Visual Dis-crimination Tasks. Dizocilpine (0.015 mg/kg) caused a sig-nificant (p � 0.001) deficit in the marmosets’ ability to ac-quire perceptually complex visual discrimination taskscompared with saline treatment (Fig. 11A). Lecozotan (2mg/kg) alone had no significant effect on mean learning score
Fig. 9. A, lecozotan enhances cognitive function in the aged rhesusmonkey as assessed by DMTS performance efficiency. Testing wasinitiated 30 min after a single dose administration. Statistical analysisof the data revealed no significant effect of treatment alone (p � 0.40),and no significant effect of treatment’s interaction with delay interval(p � 0.49). B, improvement in DMTS accuracy produced by the indi-vidualized best dose of lecozotan. These data represent the average ofthe best dose session determined from the dose-response series and aseparate series in which only the best dose was administered. Therewas no significant difference between the data sets obtained for thesetwo series (t � 0.44; p � 0.66). For the combined sessions, there was asignificant effect of drug treatment relative to vehicle that was inde-pendent of delay interval. Values are means � S.E.M. Asterisk (�)indicates significant difference (p � 0.01) compared with vehicle. Avg,average; Med, medium.
Fig. 10. Deficits in acquisition of a hippocampal task (visuospatial dis-crimination) in marmosets were induced by a VDB lesion. Deficits wereabolished by lecozotan at 2 mg/kg but not at 1 mg/kg. Values are meanlearning scores � S.E.M. Asterisk (�) indicates that learning scores forVDB-lesioned marmosets were significantly greater than the scores ofnonlesioned animals when saline (vehicle) or 1 mg/kg lecozotan wasadministered during task acquisition (p � 0.05). Dagger (†) indicates thatlearning scores for VDB-lesioned marmosets were significantly lower(i.e., acquisition was faster) when these lesioned marmosets were treatedwith 2 mg/kg lecozotan, compared with when they were treated withsaline or 1 mg/kg lecozotan (p � 0.05). Additionally, VDB-lesioned mar-mosets acquired the task as quickly as unlesioned marmosets when theywere treated with 2 mg/kg lecozotan (p � 0.05, N.S.).
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1285

compared with saline treatment. When lecozotan (2 mg/kg)was given concurrently with dizocilpine, the dizocilpine-in-duced deficit in task acquisition was abolished (p � 0.001).Overall, data suggest that lecozotan i.m. reverses cognitivedeficits induced by glutamatergic blockade in a corticallymediated task.
Acquisition of Visuospatial Discrimination Tasks.Dizocilpine at 0.015 mg/kg caused a significant (p � 0.05)deficit in the marmosets’ ability to acquire visuospatial dis-crimination tasks compared with saline treatment (Fig. 11B).Lecozotan at 2 mg/kg had no significant effect on mean learn-ing score compared with saline treatment. When 2 mg/kglecozotan was given concurrently with dizocilpine, the dizo-cilpine-induced deficit in task acquisition was abolished (p �0.05). Overall, data suggest that lecozotan i.m. completelyreverses a dizocilpine-induced cognitive deficit in hippocam-pally mediated tasks.
DiscussionIn-depth pharmacological studies in this report demon-
strate that lecozotan is a novel 5-HT1A receptor antagonistwith intriguing procognitive properties. In vitro and in vivoassays designed to test the intrinsic activity of lecozotandemonstrate that it is a potent and selective 5-HT1A receptorantagonist. This inclusive series of studies has determinedthe intrinsic activity of lecozotan in models indicative of pre-versus postsynaptic 5-HT1A receptor function. Molecules thatpossess intrinsic activity at the presynaptic 5-HT1A autore-ceptor lead to decreases in serotonergic cell firing andsubsequent reductions in 5-HT levels in terminal regions.Lecozotan demonstrated an antagonist profile, because ad-ministration of relatively high doses resulted in no change ineither extracellular 5-HT levels in terminal projection areasof the serotonergic cell bodies or any change in serotonergicneuronal firing in the DRN. Instead, lecozotan completely
antagonized 8-OH-DPAT-induced decreases in hippocampal5-HT, indicating that lecozotan seemed to act as a 5-HT1A
receptor antagonist in vivo. These data contrast with thoseobserved for the 5-HT1A receptor partial agonists BMY-7378,NAN-190, and buspirone, which can antagonize postsynaptic5-HT1A-mediated responses but act as agonists at the soma-todendritic 5-HT1A autoreceptors (Yocca, 1990; Greuel andGlaser, 1992).
Lecozotan demonstrated potent in vivo antagonist activityin rodents, nonhuman primates, and pigeons. Pharmacolog-ical studies using fixed ratio responding in rats and squirrelmonkeys demonstrated that the compound is a competitiveantagonist. Drug discrimination studies in pigeons revealedthat lecozotan blocked the discriminative stimulus effects ofthe prototypical 5-HT1A receptor agonist 8-OH-DPAT, but itdid not substitute for the discriminative stimulus effects of8-OH-DPAT. Chronic administration of lecozotan did notseem to alter the sensitivity of 5-HT1A receptors in that therewas no significant shift in the potency of an agonist to induceserotonin syndrome in male rats after continuous adminis-tration of lecozotan. Together, these data show lecozotan tobe a potent, selective, and orally bioavailable competitive5-HT1A receptor antagonist in vivo that produces no evidenceof tolerance to its antagonist effects or alters the sensitivityof 5-HT1A receptors.
Alzheimer’s disease is a neurodegenerative disorder char-acterized by various deficits in neurotransmitter function. Itis widely thought that loss of these functions contributes tothe loss in memory that is the hallmark of this disease. Thehippocampus is one of the major brain structures shown toplay a critical role in both mnemonic and cognitive function(Eichenbaum et al., 1992; Jarrard, 1993). Degeneration of thepyramidal cells in the corticohippocampal and corticocorticalsystems is one of the earliest changes in AD (Braak andBraak, 1991). Whereas many studies have concentrated onthe cholinergic deficits observed in this disease, glutamater-
Fig. 11. A, deficits in acquisition of a hippocam-pal task in marmosets induced by dizocilpine(NMDA antagonist) are completely reversed bylecozotan (i.m.) as assessed using complexvisuospatial discrimination. Marmosets underdizocilpine treatment had significant (��, p �0.001) deficits in learning scores compared withtheir scores under saline treatment. Lecozotan(2.0 mg/kg) significantly (†, p � 0.001) reversedthe dizocilpine-induced deficits. Values aremeans � S.E.M. B, deficits in acquisition of aneocortical task in marmosets induced by dizo-cilpine (NMDA antagonist) are completely re-versed by lecozotan (2 mg/kg) as assessed usingperceptually complex visual discrimination.Marmosets under dizocilpine treatment hadsignificant (�, p � 0.05) deficits in learningscores compared with their scores under salinetreatment. Lecozotan (2.0 mg/kg) significantly(†, p � 0.05) reversed the dizocilpine-induceddeficits. Values are means � S.E.M.
1286 Schechter et al.

gic dysfunction in Alzheimer’s disease has received increas-ing interest as an alternative to the well established cholin-ergic hypothesis (Maragos et al., 1987; Proctor et al., 1988;Francis et al., 1993; Myhrer, 1993).
Anatomical data indicate that 5-HT1A receptors might con-trol the excitability and propagation of information transmit-ted by the pyramidal cells to subcortical structures alongglutamate pathways (Czyrak et al., 2003). Serotonin hasbeen shown to exhibit inhibitory effects on the glutamatergicprojections of the brain, in particular those of the cortex (Dijket al., 1995; Grunschlag et al., 1997) and hippocampus(Schmitz et al., 1995; Matsuyama et al., 1996). The cellularmechanisms underlying these processes seem to involve ac-tivation of K� conductance (Grunschlag et al., 1997) and/orinhibition of presynaptic Ca2� entry (Schmitz et al., 1995;Strosznajder et al., 1996). Other data suggest that 5-HT1A
receptor antagonists can inhibit the tonic hyperpolarizingaction of serotonin on pyramidal neurons in both the cortexand hippocampus (Araneda and Andrade, 1991; Van denHooff and Galvan, 1992). Thus, it is possible that 5-HT1A
receptor antagonists may improve cognition by removing theinhibitory effects of endogenous serotonin on pyramidal neu-rons and enhancing the glutamatergic activation in both thecortex and hippocampus. An important aspect of this hypoth-esis is supported by the suggestion that a functionally hyper-responsive serotonin system in AD may contribute to cogni-tive disturbances (McLoughlin et al., 1994).
To test the hypothesis that a 5-HT1A receptor antagonistwould enhance glutamatergic neurotransmission, we usedlecozotan to examine excitatory transmission under basaland stimulated conditions within the terminal regions of theperforant pathway (i.e., granular layer of the dentate gyrus)using in vivo microdialysis. Lecozotan alone produced nochange in basal levels of glutamate within the dentate gyrus,but it induced significant augmentation of dentate gyrusglutamate levels when administered in the presence of aninfusion of K� in a way similar to that observed previouslywith WAY-101405 (Schechter et al., 2000). Furthermore,these in vivo data confirm the in vitro observations of Mat-suyama et al. (1996), who demonstrated that NAN-190 couldaugment NMDA-induced increases in glutamate in theguinea pig dentate gyrus. Serotonergic modulation of gluta-matergic neurotransmission via the 5-HT1A receptor has alsobeen observed in other brain regions (Dijk et al., 1995;Schmitz et al., 1995). Thus, lecozotan-enhanced stimulatedrelease of glutamate in the hippocampus supports the con-tention that 5-HT1A receptor antagonists can modulate stim-ulated glutamate release as part of the mechanism related toenhancing cognitive function.
The “cholinergic hypothesis”, which claims that AD is pri-marily the result of substantial cholinergic abnormalities,has received widespread attention in the development oftherapeutic strategies directed at this psychopathological ill-ness (Bartus, 2000). A major feature of the neuropathology ofAD is the degeneration of the cholinergic magnocellular neu-rons in the basal forebrain. A high density of 5-HT1A bindingsites has been reported in the region of cholinergic cell bodieswithin the nucleus basalis magnocellularis. WAY-100635 re-verses the choice accuracy deficit in rats with �-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid-induced lesionsof the nucleus basalis magnocellularis (Balducci et al., 2003).Our data indicated that in addition to the ability of lecozotan
to enhance stimulated glutamate efflux, the 5-HT1A receptorantagonist is also able to enhance stimulated acetylcholinelevels and reverse cognitive deficits induced by cholinergiclesions.
Serotonin-1A receptor antagonists potentiate NMDA-in-duced glutamate release from pyramidal neurons. Further-more, a 5-HT1A receptor antagonist has the ability to signif-icantly elevate glutamate release when administered alone(Araneda and Andrade, 1991). Since cognitive deficits havepreviously been shown to be susceptible to cognition enhanc-ing treatments in the marmoset monkey (Harder and Ridley,2000), the use of the NMDA receptor antagonist dizocilpine isappropriate for the evaluation of putative cognitive enhanc-ers targeted at dementia. Degeneration of the cholinergiccells of the basal forebrain is the other major feature that haslong been linked to cognitive dysfunction. In our marmosets,a specific and selective lesion of the cholinergic cell bodies ofthe VDB eliminated the majority of the cholinergic input tothe hippocampus and produced the expected cognitive deficitin visuospatial learning. Because the deficit obtained hasbeen shown to be susceptible to cognitive-enhancing agents,this “model” is also appropriate for evaluating dementiatreatments. The fact that the 5-HT1A antagonist lecozotanwas able to overcome both a VDB lesion and the effects of anNMDA receptor antagonist compound suggests that it is apotentially valuable cognitive-enhancing agent. In addition,WAY-100635 antagonizes the extinction of recognition mem-ory and counteracts scopolamine-induced performance defi-cits in the normal rat (Pitsikas et al., 2003). WAY-101405, anorally active 5-HT1A antagonist, can reverse the deficits in-duced by MK-801 in rats in the radial arm maze (Boast et al.,1999). In contrast to the cognitive-enhancing properties of5-HT1A antagonists, the prototypical 5-HT1A agonist 8-OH-DPAT can impair passive avoidance performance in the rat(Santucci and Shaw, 2003; Winsauer et al., 1999). WAY-100635 has been shown to reverse the cognitive deficits inmarmosets induced by a fornix lesion and reverses the cog-nitive deficits induced by the glutamatergic antagonist MK-801 and the muscarinic antagonist scopolamine as testedusing visuospatial and visual discrimination tasks in mar-mosets (Harder et al., 1996; Harder and Ridley, 2000). Thesestudies, which tested different aspects of cognition and cog-nitive impairment induced by various neurochemical andneuroanatomic insults, strongly suggest that 5-HT1A recep-tor antagonism is a viable drug strategy for restoring themultiple deficits associated with AD, and as such, will rep-resent a novel and unique treatment approach. This poten-tial therapeutic utility is further supported by the data inaged monkeys. Here, the effects of lecozotan administrationon DMTS task accuracy did not rise to statistical signifi-cance, owing primarily to individual variations in drug re-sponsiveness. However, the dose-response study revealedthat, at least on average, the 1 mg/kg dose increased accu-racy, particularly during short-delay trials. Selection andreadministration of the individualized best dose was consis-tent with the dose-response analysis in that the average bestdose was 1.1 mg/kg and that there was an overall significantimprovement in accuracy, which again was primarily associ-ated with short-delay trials.
The degree of cognitive improvement induced by lecozotanin aged rhesus monkeys using DMTS, as indicated by the16.5% increase in task accuracy above baseline, represents a
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1287

very good level of efficacy compared with other compoundstested under similar conditions in this setting (Buccafuscoand Terry, 2000). The observation that on average the effectwas largely relegated to trials associated with short-delayintervals suggests the possibility that lecozotan may have aselective action to improve attentional aspects of cognition(Paule et al., 1998). Further studies using behavioral para-digms more suited to assessing drug effects on attentionshould be used to confirm this contention. The lack of effecton task response latencies was consistent with no effect of thedrug on psychomotor speed. No untoward effects of the drugwere noted before or after testing. Together, these data dem-onstrate the efficacy of 5-HT1A receptor antagonists in en-hancing cognitive function in both rodents and primates.
Together, the heterosynaptic nature of the effects of leco-zotan imbues this compound with a novel therapeutic ap-proach to the treatment of the cognitive deficits associatedwith AD, and possibly other psychopathological diseasestates in need of improved therapies to treat cognitive- ormemory-associated loss. Lecozotan has completed phase 1clinical trials and is continuing clinical investigation in phase2 as an agent to reverse cognitive loss in AD.
Acknowledgments
The assistance of Phil Vinall (Wyeth Research) in the preparationof the manuscript is gratefully appreciated.
ReferencesAraneda R and Andrade R (1991) 5-Hydroxytryptamine 2 and 5-hydroxytryptamine
1A receptors mediate opposing responses on membrane excitability in rat associ-ation cortex. Neuroscience 40:399–412.
Arvidsson LE, Hacksell U, Nilsson JL, Hjorth S, Carlsson A, Lindberg P, Sanchez D,and Wikstrom H (1981) 8-Hydroxy-2-(di-n-propylamino)tetralin, a new centrallyacting 5-hydroxytryptamine receptor agonist. J Med Chem 24:921–923.
Balducci C, Nurra M, Pietropoli A, and Samanin R (2003) Reversal of visual atten-tion dysfunction after AMPA lesions of the nucleus basalis magnocellularis (NBM)by the cholinesterase inhibitor donepezil and by a 5-HT1A receptor antagonistWAY 100635. Psychopharmacology 167:28–36.
Bartus R (2000) On neurodegenerative diseases, models and treatment strategies:lessons learned and lessons forgotten a generation following the cholinergic hy-pothesis. Exp Neur 163:495–529.
Bliss TV and Collingridge GL (1993) A synaptic model of memory: long-term poten-tiation in the hippocampus. Nature (Lond) 361:31–39.
Boast C, Bartolomeo AC, Morris H, and Moyer JA (1999) 5HT antagonists attenuateMK801-impaired radial arm maze performance in rats. Neurobiol Learn Mem71:259–271.
Bowen DM, Francis PT, Chessell IP, and Webster MT (1994) Neurotransmission-thelink integrating Alzheimer research? Trends Neurosci 17:149–150.
Braak H and Braak E (1991) Alzheimer’s disease affects limbic nuclei of the thala-mus. Acta Neuropathol 82:239–259.
Buccafusco JJ and Terry AV Jr (2000) Multiple CNS targets for eliciting beneficialeffects on memory and cognition. J Pharmacol Exp Ther 295:438–446.
Chanda PK, Minchin MC, Davis AR, Greenberg L, Reilly Y, McGregor WH, Bhat R,Lubeck MD, Mizutani S, and Hung PP (1993) Identification of residues importantfor ligand binding to the human 5-hydroxytryptamine1A serotonin receptor. MolPharmacol 43:516–520.
Czyrak A, Czepiel K, Mackowiak M, Chocyk A, and Wcdzony K (2003) Serotonin5-HT1A receptors might control the output of cortical glutamatergic neurons in ratcingulated cortex. Brain Res 989:42–51.
Dawson LA and Routledge C (1995) Differential effects of potassium channel block-ers on extracellular concentrations of dopamine and 5-HT in the striatum ofconscious rats. Br J Pharmacol 116:3260–3264.
Dawson LA, Stow JM, and Palmer AM (1997) An improved method for the measure-ment of glutamate and aspartate using capillary electrophoresis with laser in-duced fluorescence detection and its application to brain microdialysis. J Chro-matogr B 694:455–460.
Dijk SN, Francis PT, Stratmann GC, and Bowen DM (1995) NMDA-induced gluta-mate and aspartate release from rat cortical pyramidal neurons: evidence formodulation by a 5-HT1A antagonist. Br J Pharmacol 115:1169–1174.
Eichenbaum H, Otto T, and Cohen EJ (1992) The hippocampus–what does it do?Behav Neural Biol 57:2–36.
Fargin A, Raymond JR, Lohse MJ, Kobilka BK, Caron MG, and Lefkowitz RJ (1988)The genomic clone G-21 which resembles a beta-adrenergic receptor sequenceencodes the 5-HT1A receptor. Nature (Lond) 335:358–360.
Fletcher A, Bill DJ, Bill SJ, Cliffe IA, Dover GM, Forster EA, Haskins JT, Jones D,Mansell HL, and Reilly Y (1993) WAY-100135: a novel, selective antagonist atpresynaptic and postsynaptic 5-HT1A receptors. Eur J Pharmacol 237:283–291.
Fletcher A, Forster EA, Bill DJ, Brown G, Cliffe IA, Hartley JE, Jones DE, McLena-
chan A, Stanhope KJ, Critchley DJ, et al. (1996) Electrophysiological, biochemical,neurohormonal and behavioral studies with WAY-100635, a potent, selective andsilent 5-HT1A receptor antagonist. Behav Brain Res 73:337–353.
Forster EA, Cliffe IA, Bill DJ, Dover GM, Jones D, Reilly Y, and Fletcher A (1995) Apharmacological profile of the selective silent 5-HT1A receptor antagonist, WAY-100635. Eur J Pharmacol 281:81–88.
Francis PT, Sims NR, Proctor AW, and Bowen DM (1993) Cortical pyramidal neuronloss may cause glutamatergic hypoactivity and cognitive impairment in Alzhei-mer’s disease. J Neurochem 60:1589–1604.
Gozlan H, El Mestikawy S, Pichat L, Glowinski J, and Hamon M (1983) Identifica-tion of presynaptic serotonin autoreceptors using a new ligand: 3H-PAT. Nature(Lond) 305:140–142.
Greuel JM and Glaser T (1992) The putative 5-HT1A receptor antagonists NAN-190and BMY 7378 are partial agonists in the rat dorsal raphe nucleus in vitro. EurJ Pharmacol 211:211–219.
Grunschlag CR, Haas HL, and Stevens DR (1997) 5-HT inhibits lateral entorhinalcortical neurons of the rat in vitro by activation of potassium channel-coupled5-HT1A receptors. Brain Res 770:10–17.
Harder JA, Baker HF, and Ridley RM (1998) The role of the central cholinergicprojections in cognition: implications of the effects of scopolamine on discrimina-tion learning by monkeys. Brain Res Bull 45:319–326.
Harder JA, Maclean CJ, Alder JT, Francis PT, and Ridley RM (1996) The 5-HT1Aantagonist, WAY-100635, ameliorates the cognitive impairment induced by fornixtransection in the marmoset. Psychopharmacology 127:245–254.
Harder JA and Ridley RM (2000) The 5-HT1A antagonist, WAY-100635, alleviatescognitive impairments induced by dizocilpine (MK-801) in monkeys. Neurophar-macology 39:547–552.
Hernandez L, Tucci S, Gusman N, and Paez X (1993) In vivo monitoring of glutamatein the brain by microdialysis and capillary electrophoresis with laser-inducedfluorescence detection. J Chromatogr A 652:393–398.
Jarrard LE (1993) On the role of the hippocampus in learning and memory in the rat.Behav Neural Biol 60:9–26.
Johansson L, Sohn D, Thorberg S-O, Jackson DM, Kelder D, Larsson L-G, Renyi L,Ross SB, Wallsten C, Eriksson H, et al. (1997) The pharmacological characteriza-tion of a novel selective 5-hydroxytryptamine1A receptor antagonist, NAD-299.J Pharmacol Exp Ther 283:216–225.
Kelleher RT, Morse WH, and Herd JA (1972) Effects of propranolol, phentolamineand methyl atropine on cardiovascular function in the squirrel monkey duringbehavioral experiments. J Pharmacol Exp Ther 182:204–217.
Kowall NW and Beal MF (1991) Glutamate-, glutaminase- and taurine-immunoreactive neurons develop neurofibrillary tangles in Alzheimer’s disease.Ann Neurol 29:162–167.
Lazareno S and Birdsall NJ (1993) Pharmacological characterization of acetylcho-line-stimulated [35S]-GTP gamma S binding mediated by human muscarinicm1–m4 receptors: antagonist studies. Br J Pharmacol 109:1120–1127.
Lowry OH, Rosenbrough NJ, Farr AL, and Randall RJ (1951) Protein measurementswith folin phenol reagent. J Biol Chem 193:265–275.
Maragos WF, Greenamyre T, Penney JB Jr, and Young AB (1987) Glutamatedysfunction in Alzheimer’s disease: a hypothesis. Trends Neurosci 10:65–67.
Matsuyama S, Nei K, and Tanaka C (1996) Regulation of glutamate release viaNMDA and 5-HT1A receptors in guinea pig dentate gyrus. Brain Res 728:175–180.
McLoughlin DM, Lucey JV, and Dinan G (1994) Central serotonergic hyperrespon-sivity in late-onset Alzheimer’s disease. Am J Psychiatry 151:1701–1703.
Morris RG, Anderson E, Lynch GS, and Baudry M (1986) Selective impairment oflearning and blockade of long term potentiation by an N-methyl-D-aspartatereceptor antagonist, AP5. Nature (Lond) 319:774–776.
Myhrer T (1993) Animal models of Alzheimer’s Disease: glutamatergic denervationsan alternative approach to cholinergic denervation. Neurosci Behav Rev 17:195–202.
Paule MG, Bushnell PJ, Maurissen JPJ, Wenger GR, Buccafusco JJ, Chelonis JJ,and Elliott R (1998) Symposium overview: the use of delayed matching-to-sampleprocedures in studies of short-term memory in animals and humans. NeurotoxicolTeratol 20:493–502.
Paxinos G, Watson C, Pennisi M, and Topple A (1985) Bregma, lambda and theinteraural midpoint in stereotaxic surgery with rats of different sex, strain andweight. J Neurosci Meth 13:139–143.
Pedigo NW, Yamamura HI, and Nelson DL (1981) Discrimination of multiple [3H]5-hydroxytryptamine binding sites by the neuroleptic spiperone in rat brain. J Neu-rochem 36:220–226.
Pitsikas N, Rigamonti AE, Cella SG, and Muller EE (2003) The 5-HT1A receptorantagonist WAY 100635 improves rat’s performance in different models of amne-sia evaluated by the object recognition task. Brain Res 983:215–222.
Proctor AW, Palmer AM, Francis PT, Lowe SL, Neary D, Murphy E, Doshi R, andBowen DM (1988) Evidence of glutamatergic denervation and possible abnormalmetabolism in Alzheimer’s Disease. J Neurochem 50:790–802.
Rada P, Jensen K, and Hoebel BG (2001) Effects of nicotine and mecamylamine-induced withdrawal on extracellular dopamine and acetylcholine in the rat nu-cleus accumbens. Psychopharmacology 157:105–110.
Rasmussen K, Calligaro DO, Czachura JF, Dreshfield-Ahmad LJ, Evans DC, Hem-rick-Luecke SK, Kallman MJ, Kendrick WT, Leander JD, Nelson DL, et al. (2000)The novel 5-hydroxytryptamine1A antagonist LY426965: effects on nicotine with-drawal and interactions with fluoxetine. J Pharmacol Exp Ther 294:688–700.
Ridley RM and Baker HF (1991) A critical evaluation of monkey models of amnesiaand dementia. Brain Res Rev 16:15–37.
Ridley RM, Barefoot HC, Maclean CJ, Pugh P, and Baker HF (1999) Severe learningimpairment caused by combined immunotoxic lesion of the cholinergic projectionsto the cortex and hippocampus in monkeys. Brain Res 836:120–138.
Santucci AC and Shaw C (2003) Peripheral 8-OH-DPAT and scopolamine infused
1288 Schechter et al.

into the frontal cortex produce passive avoidance retention impairments in rats.Neurobiol Learn Mem 79:136–141.
Schechter LE, Smith DL, Rosenzweig-Lipson S, Marquis K, Jones D, Nguyen HQ,Dawson LA, and Kelly MG (2000) Pharmacological characterization of an orallyactive 5-HT1A receptor antagonist. Soc Neurosci 26:392.
Schmitz D, Empson RM, and Heinemann U (1995) Serotonin and 8-OH-DPATreduced excitatory transmission in rat hippocampal area CA1 via reduction inpresumed presynaptic Ca2� entry. Brain Res 701:249–254.
Smith LM and Peroutka SJ (1986) Differential effects of 5-hydroxytryptamine 1Aselective drugs on the 5-HT behavioral syndrome. Pharmacol Biochem Behav24:1513–1519.
Strosznajder J, Chalimoniuk M, and Samochocki M (1996) Activation of serotonergic5-HT1A receptor reduces Ca2�- and glutamatergic receptor-evoked arachidonicacid and NO/cGMP release in adult hippocampus. Neurochemistry 28:439–444.
Van den Hooff P and Galvan M (1991) Electrophysiology of the 5-HT1A ligand MDL73005EF in the rat hippocampal slice. Eur J Pharmacol 196:291–298.
Van den Hooff P and Galvan M (1992) Actions of 5-hydroxytryptamine and 5-HT1Areceptor ligands on rat dorso-lateral septal neurons in vitro. Br J Pharmacol106:893–899.
Winsauer PJ, Rodriguez FH, Cha AE, and Moerschbaecher JM (1999) Full andpartial 5-HT1A receptor agonists disrupt learning and performance in rats.J Pharmacol Exp Ther 288:335–347.
Yocca FD (1990) Novel anxiolytic agents: actions on specific subtypes of central 5-HTreceptors. Prog Clin Biol Res 361:145–177.
Address correspondence to: Dr. Lee Schechter, Neuroscience Discovery,Wyeth Research, Princeton, NJ 08543. E-mail: [email protected]
SRA-333 (Lecozotan): A Silent 5-HT1A Antagonist and Cognition 1289