late quaternary benthic foraminifera and the orinoco plume
TRANSCRIPT

Marine Micropaleontology 121 (2015) 85–96
Contents lists available at ScienceDirect
Marine Micropaleontology
j ourna l homepage: www.e lsev ie r .com/ locate /marmicro
Late Quaternary benthic foraminifera and the Orinoco Plume
Brent Wilson a,⁎, Lee-Ann C. Hayek b
a Petroleum Geoscience Programme, Department of Chemical Engineering, The University of the West Indies, St. Augustine, Trinidad and Tobagob Smithsonian Institution, Mathematics and Statistics, NMNH, MRC-121, Washington, DC, USA
⁎ Corresponding author.E-mail address: [email protected] (B. Wilson)
http://dx.doi.org/10.1016/j.marmicro.2015.11.0040377-8398/© 2015 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 14 April 2015Received in revised form 13 October 2015Accepted 2 November 2015Available online 05 November 2015
Keywords:Statistical analysisYounger DryasTrinidad and TobagoInter-tropical convergence zoneFundamental niche
The bathyal benthic foraminiferal palaeoecology east of Trinidad is currently unknown. The area is oceanograph-ically complex, comprising a pro-delta deep-sea fan building across a transpressional plate boundary. Outflowfrom the Orinoco River forms a high-productivity surface plume that abuts low-productivitywater of the tropicalwestern Atlantic Ocean across a laterally extensive but sharp front. The plume's areal extent differs between therainy and dry seasons, which are governed by the position of the inter-tropical convergence zone (ITCZ). UpperQuaternary benthic foraminifera are examined in two ~4m-long piston cores, taken near the eastern edge of theplume. CHIRP profiles show that core BGT086 (water depth 626m) comprised in situmaterial throughout, whilethe lower part of core BGT096 (water depth 700m) consisted of slumpedmaterial. Benthic foraminiferal assem-blages in BGT086 were examined quantitatively, while those in BGT096 were examined qualitatively only forcomparison. Core BGT086 shows a transition from a low-diversity, Cibicidoides pachyderma-dominated commu-nity with subdominant Cassidulina curvata to a later high-diversity, low-dominance Bulimina alazanensis–Osangularielloides rugosa–Epistominella exigua community. The former is indicative of low-productivity, andthe latter of high-productivity, surface water. The restriction of E. exigua to the core's upper part may reflect achange from an aseasonal to a seasonal organic flux. The faunal change was detected by both SHE and clusteranalyses and an assemblage turnover index (ATI). It may be related to either (a) expansion of the Orinocoplume due to northward migration of the ITCZ approximately 600 yr after the end of the Younger Dryas,(b) eastward progradation of the Orinoco Delta, which would in turn push the front of the Orinoco plume east-ward, or (c) both. No other boundary detected was common to all three procedures. A boundary between 124and 130 cm below the seafloor detected by SHE analysis might equate to the 8000–7500 yr BP meltwaterpulse 1C. While some specialist species were restricted to the C. pachyderma or the B. alazanensis–O. rugosa–E. exigua communities, the percentage abundance of some others, concluded to be relative generalists(e.g., Bulimina aculeata and Sphaeroidina bulloides), varied little through the core. The undisturbed section ofcore BGT096 showed the same general faunal succession as core BGT086. The slumped material in this corewas rich in Eponides regularis, which was rare elsewhere. This species shows that the allochthonous sedimentwas derived from the outer shelf and upper slope (100–400 m water depth) during Late Pleistocene time.
© 2015 Elsevier B.V. All rights reserved.
1. Introduction
The benthic foraminiferal palaeoecology and ecostratigraphy of thebathyal Orinoco deep-sea fan, in the western tropical Atlantic Ocean,are little known, even though the adjacent island of Trinidad has beena source of oil and natural gas for over a century (Carr-Brown, 2007;Furlonge and Kaiser, 2010) during which foraminiferal biostratigraphyplayed a major role in hydrocarbon exploration (Saunders and Bolli,1979). Exploration around Trinidad and Tobago to date has accessedfields in onshore regions or beneath the neritic (typically b150 mwater depth) continental shelf, and the few studies of late Pleistoceneand Holocene benthic foraminifera associated with this hydrocarbonexploration have focussed on neritic depths proximal to the Orinoco
.
Delta (Drooger and Kaasschieter, 1958; Kruit, 1954; van der Zwaanand Jorissen, 1991; Wilson, 2006, 2010). Comparison of geographicmaps of the bathyal to abyssal Orinoco fan and surface waters in Callecet al. (2010), Agard and Gobin (2000) and Kempler (2012) shows thatthe proximal part of the fan lies beneath an offshore hyperpycnal plumewith high surface productivity. The impact of this plume on fan foraminif-eral assemblages is unknown.
Nearby,Gaby andSenGupta (1985) examined lateQuaternary abyssal(N2000 m water depth) foraminifera in eight piston cores from theVenezuela Basin of the eastern Caribbean Sea. They found distinct upperPleistocene and Holocene assemblages, the Holocene assemblage beingdominated by Planulina wuellerstorfi (Schwager) and the upper Pleisto-cene assemblage being co-dominated by Massilina sp., Globocassidulinasubglobosa (Brady), and Nummoloculina irregularis (d'Orbigny). Theyfound neither Bulimina aculeata d'Orbigny, nor Uvigerina peregrinaCushman. Galluzzo et al. (1990) documented the distribution of benthic

86 B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
foraminifera in the Holocene of the Grenada Basin, ~200 km NW ofTrinidad, where they found an assemblage dominated by B. aculeata,B. alazanensis Cushman, Osangularia culter (Parker & Jones), Uvigerinaauberiana, d'Orbigny, and U. peregrina between 1000 and 2100 m. Thisthey thought was associated with Sub Antarctic Intermediate Water andindicative of low dissolved oxygen levels. Sen Gupta et al. (1991) foundonly subtle changes in the benthic foraminiferal fauna at the base of theHolocene, as defined using an influx of the planktonic foraminiferGloborotalia menardii, within the Grenada Basin. Wilson and Costelloe(2011) examined benthic foraminifera in the Quaternary of DSDP Hole148 (13°25.12′ N, 63°43.25′ W, water depth 1232 m) near the easternedge of the Orinoco plume. Only Sigmoilopsis schlumbergeri (Silvestri),U. auberiana, U. peregrina, and B. aculeata each formed N5% of the total as-semblage from DSDP Hole 148. Species of Uvigerina and Bulimina, whichperhaps due to an enhanced organic matter loading are indicative oflow dissolved-oxygen content (Kaiho, 1994), respectively formed 22.6%and 14.3% of the total assemblage. Cibicidoides pachyderma (Rzehak)[recorded as C. floridanus (Cushman)], which is indicative of oxic bottomwaters (Kaiho, 1994), formed only 2.6% of the total assemblage. Hofker(1983) examined the benthic foraminifera offshore Guyana and Surinam,~700–1000 km SW of Trinidad, using samples primarily from the conti-nental shelf but including five samples from upper to middle bathyaldepths (207–940m).He concluded that the bathyal fauna,with abundantB. aculeata, B. alazanensis, B. striata mexicana Cushman, C. pachyderma[as C. pseudoungerianus (Cushman)], O. culter, Sphaeroidina bulloidesd'Orbigny, and U. peregrina, are indicative of the upwelling of coolwater. He noted that, due to the influence of Amazon water, Secchi discvisibility on the continental shelf off Guyana “is nowhere more than20 m, and often less”. Despite the work by Hofker (1983), Gaby and SenGupta (1985), Galluzzo et al. (1990) and Wilson and Costelloe (2011),those wishing to interpret the deeper water palaeoenvironments of thePleistocene and older Neogene succession of Trinidad have relied on ob-served foraminiferal distributions in the western Gulf of Mexico (Culver,1988; Pflum and Frerichs, 1976; Phleger and Parker, 1951; Poag, 1981),perhaps because that area is influenced byMississippi outflow and Secchidisc visibility there is comparable to that off Trinidad and Guyana(Manheim et al., 1972). Alternatively, they have used the subjectivebiofacies model developed for the Trinidad Neogene by Batjes (1968).These biofacies, named after the dominant species, are the shallow-water Miliammina, Ammonia and Trochammina biofacies, the more openwater Hanzawaia, Buliminella, Eggerella, Uvigerina and Bolivina floridanabiofacies, and the predominantly agglutinated, deeper waterGlomospira–Alveovalvulina–Cyclammina biofacies. Jones (1998) usedboth Batjes (1968) system of biofacies and data from the Gulf of Mexicowhen undertaking palaeoenvironmental interpretations of benthic fora-miniferal assemblages in the upper Miocene and Pliocene formations ofTrinidad.
Hydrocarbon exploration off Trinidad is now moving into deeperwater on the continental slope. This necessitates a study of the benthic fo-raminifera in the upper Quaternary of the Orinoco deep-sea fan. This is anarea of considerable geological and oceanographic complexity, however,consisting of amajor river delta and fan system(theOrinocoDelta, the ad-jacent, shallow submarine Orinoco Deltana Platform, and the deeperwater Orinoco deep-sea fan) building across the transpressional tectonicplate boundary between the Caribbean and South American plates(Aslan et al., 2003; Callec et al., 2010; Garciacaro et al., 2011), thelocus of which currently extends across central Trinidad (Prenticeet al., 2010).
Phleger (1976) noted that the benthic foraminiferal community atdepth varies according to the primary productivity of surface waters,the latter being high in shelf areas affected by outflow from a largeriver. He found that the benthic foraminiferal fauna in very high produc-tion areas, such as beneath the offshore hyperpycnal plume off the east-ernMississippi Delta (see Chin-Leo and Benner, 1992), has lowdiversityand is dominated by one to three species that collectively constituteN95% of the population. Off the Mississippi the surface productivity is
highest at intermediate salinity levels (chlorides 15–30 ppt), decreasinginto the normal-marine waters of the open Gulf of Mexico. The OrinocoRiver being the fourth largest in terms of outflow worldwide (Hu et al.,2004), of particular interest to hydrocarbon exploration aroundTrinidad is the question of how the bathyal benthic foraminiferal com-munity responded to sea-level change and the concomitant migrationof the Orinoco Delta and its associated offshore oceanographic features.Furthermore, water and mud from the River Amazon, the world's larg-est river in terms of outflow (Oki and Kanae, 2006), is carried NWfrom the Amazon river mouth by the Guyana Current towardsTrinidad and eastern Venezuela such that about half the mud formingthe Orinoco Delta is of Amazon origin (Aslan et al., 2003). Low salinityAmazon water contributes to the Orinoco plume. During the Pliocene,when glacially-driven sea-level change was muted in comparison tothe Pleistocene, sea level rise and coupled sedimentation-drivenprogradation caused the Orinoco Delta to alternately retreat and ad-vance on the order of 100 km across the continental shelf (Chen et al.,2014), and similar translations are to be expected during the Pleisto-cene. Knowledge of how the benthic community reacted to these trans-gressions and regressions (both normal and forced)would aid greatly ininterpreting the evolution of the Orinoco fan. This paper examines thebenthic foraminiferal community across the Pleistocene–Holoceneboundary (Termination I — see Cheng et al., 2009), when sea levelrose ~120 m (Poag and Valentine, 1976), on the modern-day middlebathyal (500–1000 m) continental slope east of Trinidad.
Some comments can be made from the results of this study regard-ing the relative niche widths of benthic foraminiferal species. The fun-damental niche is an n-dimensional hypervolume where each of the ndimensions comprises a physical or biological factor (Hutchinson,1957; Soberón and Peterson, 2005). The ecological niches of individualbenthic foraminiferal species are not yet well understood (Murray,2001). Factors investigated that determine their fundamental andrealised niches (the pre-interaction and post-interaction niches ofMorin, 1999) comprise primarily food (organic carbon) and dissolvedoxygen (Van der Zwaan et al., 1999), nitrate (Piña-Ochoa et al., 2010)and light (Hallock, 1984). These factors being consumptive resources(Jeffries and Lawton, 1984), these studies overlook the role of speciesinteractions. Nevertheless, it is apparent that the fundamental nichewidth varies between species, there being distinct generalists (withbroad fundamental niches) and comparative specialists. van derZwaan et al. (1999) suggested that foraminifera generally are not steno-topic to some environmental variables such as temperature and salinity,having adopted a beneficial generalist strategy in which low degree ofspecialisation prevents rapid extinction. However, not all benthic fora-minifera are generalists. Kaiho (1994) distinguished oxic and dysoxicbenthic foraminifera, while Sen Gupta and Machain-Castillo (1993)showed that some endobenthic foraminifera are conspicuously domi-nant in bathyal oxygen minimum zones. Comparing epiphytic andsediment-dwelling foraminiferal communities around Long Island,Matera and Lee (1972) concluded that Elphidium incertum (Williamson)and Ammotium salsum (Cushman and Brönnimann) are generalists,being common in both epiphytic and sediment communities. Steineckand Bergstein (1979) suggested that modern Ammobaculites exiguusCushman and Brönnimann and Ammonia beccarii (Linnaeus) are oppor-tunistic generalists, occupying a wider range of paralic environmentsthan do other species. This generalist behaviour for A. beccarii may bemore apparent than real, however, authors having applied this nameto many forms (Hayward et al., 2004). Here we posit the generalistand relative specialist nature of some species recovered from the Orinocofan.
2. Regional setting
The complex slope east of Trinidad is characterised by a sand-richdeep-sea fan building across the active Barbados accretionary prism.The fan is traversed by at least four NE–SW anticlines and more than

87B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
ten NW–SE trending growth faults (Wood, 2000, figure 2). There arethree types of mass transport complexes in the region: (1) shelf-attached systems fed by shelf-edge deltas with sediment input con-trolled by sea level change; (2) slope-attached systems where upperslope sediments failed catastrophically due to earthquakes or gas-hydrate disruptions; and (3) locally detached systems that formedwhen local seafloor instabilities triggered small collapses (Moscardelliand Wood, 2008).
Environmental and palaeoevironmental factors are known tochange rapidly eastwards from Trinidad and northern South America,but, with the exception of Wilson and Costelloe (2011), regional inter-pretations of foraminiferal ecostratigraphy have not yet taken accountof this marked west–east gradient. Schmoker et al. (2013) noted thatthe modern waters of the tropical central Atlantic Ocean vary littleover the annual cycle, with high temperatures and low nutrient avail-ability. These waters have the lowest primary productivity worldwide.Palaeoenvironments on the Ceará Rise (~1000 km NE of the AmazonRiver mouth) were relatively stable across the late Pleistocene and Ho-loceneMarine Isotope Stages (MIS) 1–3, and hence across Termination I(which separates MIS 1 fromMIS 2; Foster and Sexton, 2014). This wasdespite an apparent fivefold increase in the upwelling rate of deeperwaters in the western tropical Atlantic during the last glacial (MIS2) (Foster and Sexton, 2014) and a major cosmic-impact event at theonset of the Younger Dryas cooling episode at ≈12,800 ± 150 yearsBP (Kinzie et al., 2014) that effected North America, Europe and theNorth Atlantic Ocean and Caribbean Sea. Glacial-interglacial sea surfacepalaeotemperatures in the open western tropical Atlantic differed byonly about 2 °C (cf. 4–6 °C in the eastern tropical Atlantic) (Foster andSexton, 2014, figure 1A) and western tropical Atlantic primary produc-tivity at the ocean surface changed little, averaging ~30 g C m−2 yr−1
(Rühlemann et al., 1996). This primary productivity suggests that theCeará Rise occupied a low productivity region throughout the later Qua-ternary. Nearer shore, the western tropical Atlantic Ocean along the NESouth Americanmargin was, in contrast, more variable during late Qua-ternary times. It is currently being affected by outflow from the Amazonand Orinoco rivers (Hu et al., 2004) and localised upwelling of nutrient-rich water (Agard and Gobin, 2000; Pascual et al., 2009; Wilson andHayek, 2014b). This engenders a mean primary productivity at thewater surface of 1.1–1.2 g Cm−2 day−1 along the present-day coastlineand around the island of Trinidad (cf. 0.2–0.4 g C m−2 day−1 on theCeará Rise; Foster and Sexton, 2014,figure 1B). The nutrient-richwatersof the Amazon and Orinoco Rivers conjoin to form an offshorehyperpycnal lens (the Orinoco plume) that meets the open Atlanticwater along an offshore front across which there is little mixing (DelCastillo et al., 1999; López et al., 2013). This is apparently reflected inthe offshore distribution of terrestrial pollen derived from the OrinocoRiver, which is concentrated in a narrow band that parallels the shoreline(Muller, 1959). Currently the eastern edge of the Orinoco plume,which ismarked by a sharp boundary between high- and low-productivity surfacewater, at the October height of the northern South American rainy seasonlies at ~60°W (Kempler, 2012). It extends approximately N–S, and is thusonly subparallel to the SE/NW-trending bathymetric contours in thestudy area.Wilson (2008a) suggested on the basis of recent benthic ostra-cod distributions that there occurs within the Orinoco plume off SETrinidad a secondary front that separates Orinoco water from thatderived from the Amazon. This secondary front extended acrossthe shelf perpendicular to the bathymetric contours, compromisingthe use of ostracod and foraminiferal species as palaeodepth indica-tors on the continental shelf within the hydrocarbon-rich ColumbusBasin (for details of which, see Wood, 2000). Within the Orinocoplume, the Amazon and Orinoco waters around Trinidad inducethe development of a rich microphytoplankton and mesozooplanktoncommunity indicative of a herbivorous-metazoan food chain andof conditions favourable for an enhanced net carbon flux to the sea-floor (Figueroa, 2007) compared to the open tropical Atlantic Ocean(Schmoker et al., 2013).
The area covered by the Orinoco plume changed throughout at leastthe late Pleistocene and Holocene, when the hydrological cycle of thenorthern South American continent was impacted greatly by latitudinalmigrations of the insolation-driven inter-tropical convergence zone(ITCZ) and associated monsoon activity (Arbuszewski et al., 2013).The ITCZ had amean annual position of 2°S during the Last Glacial Max-imum and the Younger Dryas, but shifted to northern South America(10°–12°N) during the early Holocene (Hoffmann et al., 2014). Thisled to dry Late Pleistocene and wet Holocene palaeoclimates in thenorthern Venezuela–Trinidad region. These hydrological changes arereflected in the tests of the nearshore planktonic foraminiferalpalaeoecommunity. Hoffmann et al. (2014) examined Ba/Ca ratios andδ18Oseawater in the surface-dwelling planktonic foraminiferaGlobigerinoides ruber (pink) in a core taken off eastern Trinidad(11°36.53′N, 60°57.86′W; 852 m). They found an abrupt increasein the Ba/Ca ratio in the early Holocene, ~600 yr. after the end ofthe Younger Dryas (YD) cold interval at ca. 10.8 ka. This indicatesa major reorganisation of moisture sources in northern SouthAmerica at the time. In contrast, however, the salinity dependent δ-18Oseawater from the same samples decreased gradually starting atthe end of the YD. They suggested that the Ba/Ca ratio documentsan abrupt increase in Ba-rich waters of a northern Andean sourcecaused by an insolation-driven shift of the ITCZ.
It is not yet known in detail how the plume/open Atlantic front illus-trated by Odriozola et al. (2007), figure 3) migrated during the glacial–interglacial cycles of the Pleistocene. This paper addresses this ques-tion by examining the benthic foraminiferal palaeoecology andecostratigraphy in two piston cores taken in the vicinity of the east-ern rainy-season boundary of the present day Orinoco plume.
3. Materials and methods
A proprietary suite of piston cores was collected from the slope eastof Trinidad between 09° 57.8379′–10° 19.7871′N and 059° 56.0245′–060° 19.2332′W, water depths 168–700 m (Ron Daniel, writtencommunication). Samples from two of these (BGT086, 10.258°N,59.956°W, water depth 626 m; BGT096, 10.330°N, 59.954°W, waterdepth 700 m; Fig. 1) were released to the authors for study. Accordingto the timeof year, the sites of piston cores BGT086andBGT096 at presentlay either east (dry season) or west (rainy season) of the offshore front.The chronology of these cores is extrapolated from the nearby coreM78/1-235-1 (11.607°N, 60.96°W, water depth 852 m) of Hoffmannet al. (2014).
Care was taken to avoid the anticlines and faults recorded by Wood(2000) during core selection. Furthermore, the slope-forming clinoformscan show considerable deformation due to themass transport complexesrecorded by Moscardelli and Wood (2008). Although an attempt wasmade to avoid transported sediment during core selection, someslumping is evident at the base of core BGT096. While high-resolutionCHIRP sonar profiles showed that the site of core BGT086 was unde-formed, with reflectors paralleling the seafloor throughout and the firstmarked reflector ~200 cm below the seafloor (cmbsf), Site BGT096showed deformation from ~280 cmbsf but undeformed sedimentabove. Core BGT086 comprised silty clays (colour 5Y 4/1) while BGT096comprised clay (colour 5Y 3/1). The gross appearance of the slumpedma-terial did not differ from that of the overlying in situ sediment. CoreBGT086was analysed quantitatively, but, in view of the soft sediment de-formation, BGT096 was primarily analysed qualitatively.
Samples had been taken where possible from both cores from4 cm-thick slices spaced 10 cm apart, although selected samples wereretained by the donor for other analyses. The donated samples werewashed over a 63 μm mesh to remove silt and clay and the residuedried over a gentle heat before being spread evenly over a pickingtray. Specimens were picked from the first three rows in the tray to ob-tain N250 benthic foraminifera. For core BGT086, sample BGT086:380–384 cmbsf yielded 249 benthic foraminifera only in its entirety;
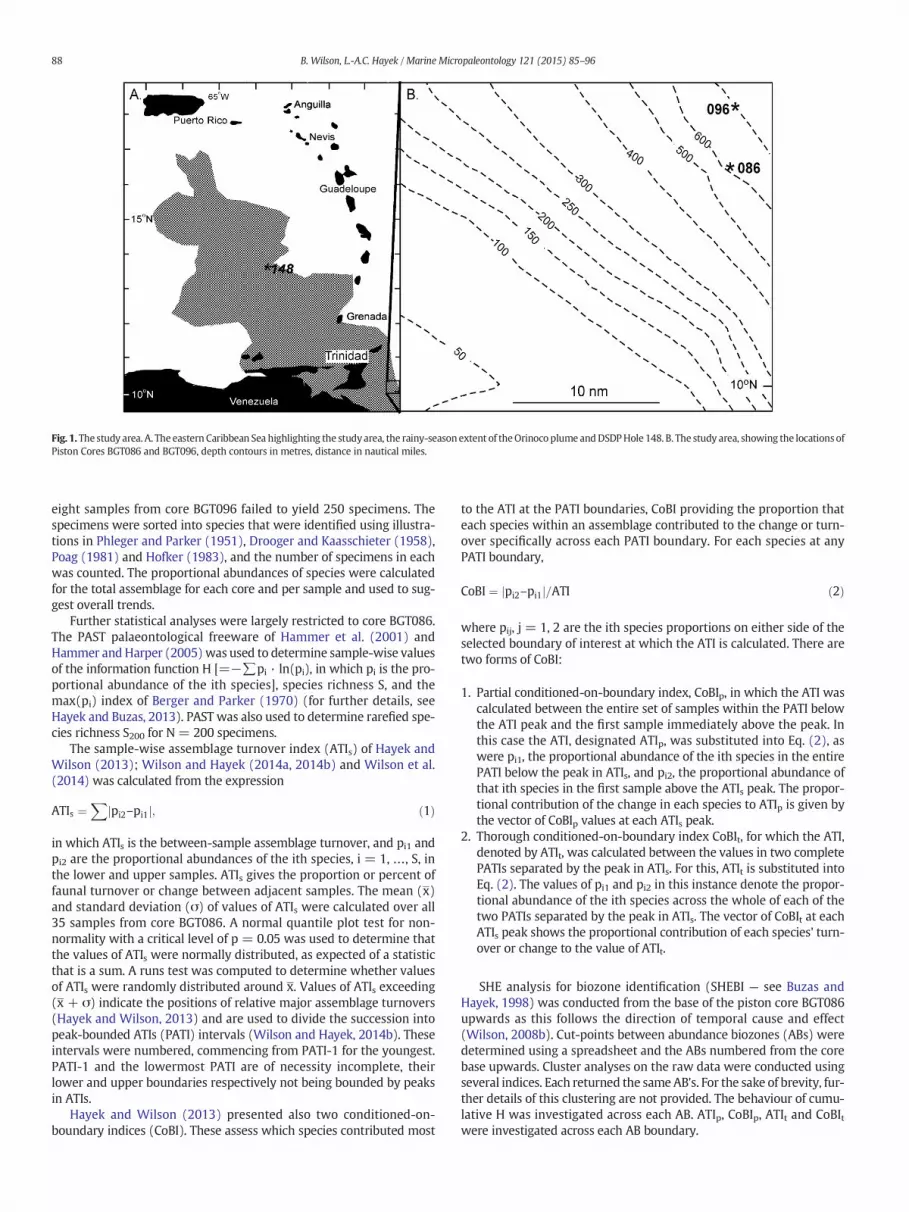
Fig. 1.The study area. A. The eastern Caribbean Sea highlighting the studyarea, the rainy-seasonextent of theOrinoco plume andDSDPHole 148. B. The study area, showing the locations ofPiston Cores BGT086 and BGT096, depth contours in metres, distance in nautical miles.
88 B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
eight samples from core BGT096 failed to yield 250 specimens. Thespecimens were sorted into species that were identified using illustra-tions in Phleger and Parker (1951), Drooger and Kaasschieter (1958),Poag (1981) and Hofker (1983), and the number of specimens in eachwas counted. The proportional abundances of species were calculatedfor the total assemblage for each core and per sample and used to sug-gest overall trends.
Further statistical analyses were largely restricted to core BGT086.The PAST palaeontological freeware of Hammer et al. (2001) andHammer andHarper (2005) was used to determine sample-wise valuesof the information function H [=−∑pi · ln(pi), in which pi is the pro-portional abundance of the ith species], species richness S, and themax(pi) index of Berger and Parker (1970) (for further details, seeHayek and Buzas, 2013). PAST was also used to determine rarefied spe-cies richness S200 for N = 200 specimens.
The sample-wise assemblage turnover index (ATIs) of Hayek andWilson (2013); Wilson and Hayek (2014a, 2014b) and Wilson et al.(2014) was calculated from the expression
ATIs ¼X
pi2–pi1j j; ð1Þ
in which ATIs is the between-sample assemblage turnover, and pi1 andpi2 are the proportional abundances of the ith species, i = 1, …, S, inthe lower and upper samples. ATIs gives the proportion or percent offaunal turnover or change between adjacent samples. The mean (x)and standard deviation (σ) of values of ATIs were calculated over all35 samples from core BGT086. A normal quantile plot test for non-normality with a critical level of p = 0.05 was used to determine thatthe values of ATIs were normally distributed, as expected of a statisticthat is a sum. A runs test was computed to determine whether valuesof ATIs were randomly distributed around x. Values of ATIs exceeding(x + σ) indicate the positions of relative major assemblage turnovers(Hayek and Wilson, 2013) and are used to divide the succession intopeak-bounded ATIs (PATI) intervals (Wilson and Hayek, 2014b). Theseintervals were numbered, commencing from PATI-1 for the youngest.PATI-1 and the lowermost PATI are of necessity incomplete, theirlower and upper boundaries respectively not being bounded by peaksin ATIs.
Hayek and Wilson (2013) presented also two conditioned-on-boundary indices (CoBI). These assess which species contributed most
to the ATI at the PATI boundaries, CoBI providing the proportion thateach species within an assemblage contributed to the change or turn-over specifically across each PATI boundary. For each species at anyPATI boundary,
CoBI ¼ pi2–pi1j j=ATI ð2Þ
where pij, j = 1, 2 are the ith species proportions on either side of theselected boundary of interest at which the ATI is calculated. There aretwo forms of CoBI:
1. Partial conditioned-on-boundary index, CoBIp, in which the ATI wascalculated between the entire set of samples within the PATI belowthe ATI peak and the first sample immediately above the peak. Inthis case the ATI, designated ATIp, was substituted into Eq. (2), aswere pi1, the proportional abundance of the ith species in the entirePATI below the peak in ATIs, and pi2, the proportional abundance ofthat ith species in the first sample above the ATIs peak. The propor-tional contribution of the change in each species to ATIp is given bythe vector of CoBIp values at each ATIs peak.
2. Thorough conditioned-on-boundary index CoBIt, for which the ATI,denoted by ATIt, was calculated between the values in two completePATIs separated by the peak in ATIs. For this, ATIt is substituted intoEq. (2). The values of pi1 and pi2 in this instance denote the propor-tional abundance of the ith species across the whole of each of thetwo PATIs separated by the peak in ATIs. The vector of CoBIt at eachATIs peak shows the proportional contribution of each species' turn-over or change to the value of ATIt.
SHE analysis for biozone identification (SHEBI — see Buzas andHayek, 1998) was conducted from the base of the piston core BGT086upwards as this follows the direction of temporal cause and effect(Wilson, 2008b). Cut-points between abundance biozones (ABs) weredetermined using a spreadsheet and the ABs numbered from the corebase upwards. Cluster analyses on the raw data were conducted usingseveral indices. Each returned the same AB's. For the sake of brevity, fur-ther details of this clustering are not provided. The behaviour of cumu-lative H was investigated across each AB. ATIp, CoBIp, ATIt and CoBItwere investigated across each AB boundary.
89B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
4. Results
4.1. Piston core BGT086
A total of 10,415 benthic foraminifera were obtained from thiscore (x = 297 per sample, σ = 20.2 specimens; online supplemen-tary data file 1). These were placed in 152 species, of which 31 wereeither left in open nomenclature or merely compared with previ-ously named species. Thirty species (19.7%) were singletons, while 61
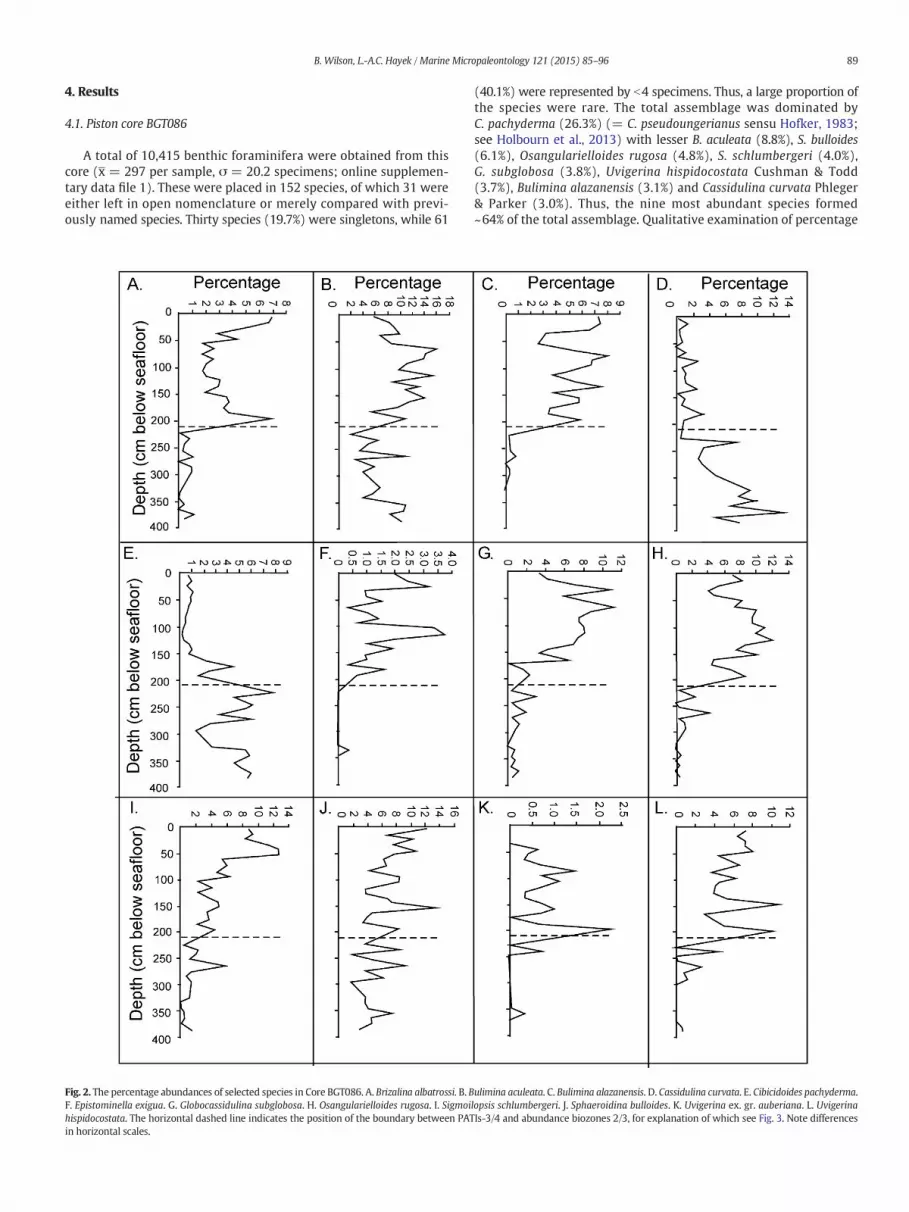
Fig. 2. The percentage abundances of selected species in Core BGT086. A. Brizalina albatrossi. B. BF. Epistominella exigua. G. Globocassidulina subglobosa. H. Osangularielloides rugosa. I. Sigmoihispidocostata. The horizontal dashed line indicates the position of the boundary between PATin horizontal scales.
(40.1%) were represented by b4 specimens. Thus, a large proportion ofthe species were rare. The total assemblage was dominated byC. pachyderma (26.3%) (= C. pseudoungerianus sensu Hofker, 1983;see Holbourn et al., 2013) with lesser B. aculeata (8.8%), S. bulloides(6.1%), Osangularielloides rugosa (4.8%), S. schlumbergeri (4.0%),G. subglobosa (3.8%), Uvigerina hispidocostata Cushman & Todd(3.7%), Bulimina alazanensis (3.1%) and Cassidulina curvata Phleger& Parker (3.0%). Thus, the nine most abundant species formed~64% of the total assemblage. Qualitative examination of percentage
ulimina aculeata. C. Bulimina alazanensis. D. Cassidulina curvata. E. Cibicidoides pachyderma.lopsis schlumbergeri. J. Sphaeroidina bulloides. K. Uvigerina ex. gr. auberiana. L. UvigerinaIs-3/4 and abundance biozones 2/3, for explanation of which see Fig. 3. Note differences

90 B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
abundances of these species (Fig. 2) suggests that they fall into threegroups:
1. Species most abundant towards the base of the core and decreasingin abundance abruptly (C. pachyderma, C. curvata; Fig. 2)
2. Species most abundant towards the top of the core and increasing inabundance abruptly (B. alazanesis, O. rugosa, U. hispidocostata)
3. Species increasing gradually in abundance through the core(B. aculeata, S. bulloides).
Epistominella exigua (Brady), although never forming N4% of anysample, is abundant only at 194 cmbsf and above and so belongs inGroup 2 (Fig. 2F). G. subglobosa (Fig. 2G) also belongs in this group.
The between-sample assemblage turnover index ranged fromATIs = 0.27–1.48 (x = 0.61, σ = 0.21) (Fig. 3A). In only three placeswas ATIs N x + σ. This was used to delimit four PATIs between:
1. 0–194 cmbsf, 20 samples (PATI-1),2. 194–234 cmbsf, 1 sample (PATI-2),3. 234–280 cmbsf, 5 samples (PATI-3),4. 280–384 cmbsf, 9 samples (PATI-4).
The boundary between PATI-1 and PATI-2 encompasses a 20 cm gapfromwhich samples were retained for use elsewhere. The boundary be-tween these PATIs is here drawn at 214 cmbsf, at the midpoint of thisgap. A second gap, between 294 and 320 cmbsf, lay within PATI-4.ANOVA and post hoc Tukey–Cramer's Q were used to compare meanATIs for the three PATIs containing ≥2 samples. This showed thatmean ATIs did not differ between these PATIs (F = 0.699, p = 0.51,df = 2).
Species richness per sample ranged from S = 22–61. Species rich-ness rarefied to 200 specimens per sample (S200) ranged from 17.9–49.2 (at 220–224 cmbsf and 10–14 cmbsf respectively). S200 is highertowards the top of the piston core (Fig. 3B). This pattern was investigat-ed further across those PATIs with ≥2 samples using ANOVA, post hocTukey–Cramer's Q and ln(S200 + 1). ANOVA showed that ln(S200 + 1)differed between at least two PATIs (F = 22.97, p b 0.0001). Tukey–Cramer's Q was not necessary to show that mean S200 did not differ be-tween PATI-3 and PATI-4, which both had a mean of 28.2 species, butthis test showed that that mean S200 in PATI-1 was significantly differ-ent (mean S200=39.3). Since S200 in PATI-2was less than theminimumin either PATI-3 or 4, S200 did not increase linearly upwards through thecore, but changed in a step across PATI-2. The information functionshowed the same pattern as S200, varying little above 194 cmbsf. This in-crease in diversity is accompanied by a decrease in dominance, asshown by low levels of max(pi) above 154 cmbsf.
Fig. 3. Statistical measures in Core BGT086. A. Sample-wise assemblage turnover index ATIs. Thedashed lines mark PATI boundaries. Data points are placed at the midpoint between adjacentposition of abundance biozone boundaries from SHE analysis. C. The information function H. D
The partial assemblage turnover index across the three PATI bound-aries ranged from ATIp = 0.564–1.481 (i.e., the assemblage turnoverwas 56.4%–148.1%) across the PATI-4/3 and PATI-2/1 boundaries re-spectively (Table 1). The number of species contributing to ATIp(i.e., for which CoBIp N 0) differed between the three peaks at the PATIboundaries (PATI-4/3 = 77 species, PATI-3/2 = 70 species, PATI-2/1 = 53 species). However, only 29 species had a CoBIp N 0.01(i.e., contributed N1.0% to ATIp) across any one boundary, while onlyseven species had a COBIp N 0.05 across any PATI boundary(Brizalina subaenariensis mexicana, B. aculeata, B. marginata, C. curvata,C. pachyderma, O. rugosa, U. hispidocostata). Of these, B. subaenariensismexicana and B marginata had a CoBIp N 0.05 across one boundary only,reflecting their more common occurrence towards the bottom of thecore. Single values of CoBIp N 0.05 across one boundary only forO. rugosa and U. hispidocostata reflected their more common occurrencetowards the core top. Only B. aculeata and C. pachyderma presentedCOBIp N 0.05 across all the PATI boundaries, reflecting a significant nega-tive correlation between the proportional abundances of these species(r=−0.58, p b 0.0001) as B. aculeata replaces C. pachyderma as the dom-inant species towards the top of the core.
The thorough assemblage turnover index across the three PATIboundaries ranged from ATIt = 0.418–1.560 (i.e., the assemblage turn-over was 41.8%–156.0%) across the PATI-4/3 and PATI-2/1 boundariesrespectively (Table 2). The number of species contributing to ATIt dif-fered between the three PATI boundaries (PATI-4/3 = 87 species,PATI-3/2 = 70 species, PATI-2/1 = 145 species). However, only 24species had a COBIt N 0.01 across any one boundary, while only five spe-cies had a COBIt N 0.05 across any PATI boundary (B. subaenariensismexicana, B. aculeata, C. curvata, C. pachyderma, U. dirupta). OnlyC. pachyderma had a COBIt N 0.05 across all three boundaries, whileB. aculeata had a COBIt N 0.05 across the uppermost two boundaries.
SHE analysis for biozone identification (SHEBI) indicated the pres-ence of four abundance biozones (ABs):
1. AB1, 330–384 cmbsf, six samples2. AB2, 220–324 cmbsf, nine samples3. AB3, 130–194 cmbsf, seven samples4. AB4, 0–124 cmbsf, thirteen samples.
Of these, the AB2/3 boundary coincided with a 20 cm gap fromwhich sampleswere retained for use elsewhere. The boundary betweenthese two ABs is drawn at 214 cmbsf, at the midpoint of the gap. A sec-ond gap, between 294 and 320 cmbsf, laywithin AB2. Although four ABsseparated by three AB boundaries were recognised, only one of theseboundaries was common to both ATI and SHE analysis (between PATI-
vertical dashed line indicates a value of mean ATIs plus one standard deviation; horizontalsamples. B. Rarefied species richness for 200 specimens, S200. Horizontal lines mark the. Dominance, measured using the max(pi) index.

Table 1Partial assemblage turnover index (ATIp) and partial conditioned-on-boundary indices(CoBIp) for PATI boundaries detected in Core BGT086. Only values of CoBIp exceeding0.01 are shown. Values of CoBIp in bold exceed 0.05.
PATI boundary 3/4 2/3 1/2
ATIp 0.564 0.634 1.481CoBIpBrizalina albatrossi – – 0.046Brizalina barbata 0.014 – –Brizalina subaenariensis mexicana 0.074 – –Brizalina translucens 0.010 – –Bulimina aculeata 0.079 0.058 0.061Bulimina alazanensis – – 0.037Bulimina marginata 0.080 0.048 0.011Bulimina striata mexicana – 0.046 –Cassidulina curvata 0.072 0.057 –Cassidulina laevigata – – 0.013Cassidulina norcrossi australis 0.011 – –Cibicidoides bradyi – – 0.013Cibicidoides pachyderma 0.332 0.414 0.420Fissurina marginata 0.014 – –Globocassidulina subglobosa – 0.015 0.013Globocassidulina subglobosa subcalifornica – – 0.011Lenticulina norvangilae 0.011 – –Martinottiella communis 0.018 – –Nummoloculina irregularis 0.011 0.010 –Oridorsalis umbonatus 0.015 0.020 –Osangularielloides rugosa – 0.016 0.057Pullenia bulloides – – 0.015Quinqueloculina cruziana – 0.038 0.018Sigmoilopsis schlumbergeri 0.013 0.032 0.028Sphaeroidina bulloides 0.018 0.030 0.032Uvigerina dirupta 0.023 – 0.033Uvigerina ex gr. auberiana – – 0.015Uvigerina hispidocostata – 0.023 0.068Uvigerina mediterranea – 0.020 –
Table 3Partial assemblage turnover index (ATIp) and partial conditioned-on-boundary indices(CoBIp) for abundance biozone boundaries from SHE analysis for biozone identificationdetected in Core BGT086. Only values of CoBIp exceeding 0.01 are shown. Values of CoBIpin bold exceed 0.05.
Abundance biozone boundary 1/2 2/3 3/4
ATIp 0.787 0.934 0.901CoBIpBrizalina albatrossi 0.011 – 0.011Brizalina barbata – – 0.052Brizalina subaenariensis mexicana – – 0.120Brizalina translucens – – 0.034Bulimina aculeata 0.080 0.048 0.034Bulimina marginata – 0.010 0.130Bulimina striata mexicana 0.070 0.064 0.090Cassidulina curvata 0.041 0.095 0.038Cibicidoides pachyderma 0.569 0.550 0.122Dentalina communis 0.015 – –Fissurina marginata 0.012 0.014 –Globocassidulina subglobosa 0.011 – –Lenticulina calcar – 0.014 –Lenticulina gibba – – 0.011Lenticulina norvangilae 0.019 – –Lenticulina rotulata – 0.021 –Nonionella labradorica – – 0.030Nummoloculina irregularis 0.014 0.011 –Oridorsalis umbonatus – 0.023 –Osangularielloides rugosa – – 0.011Sigmoilopsis schlumbergeri – – 0.013Sphaeroidina bulloides 0.046 0.044 0.013Uvigerina dirupta – 0.034 0.177
91B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
3/4 and AB2/3). Cumulative H increased across AB1, 3 and 4, whereasfor AB2 the slope on the equation for H vs. lnN is negative but notsignificant.
The partial assemblage turnover index for AB boundaries fromSHEBI ranged from ATIp = 0.79–0.93 (AB1/2 and AB2/3 boundaries
Table 2Thorough assemblage turnover index (ATIt) and thorough conditioned-on-boundary indi-ces (CoBIt) for PATI boundaries detected in Core BGT086. Only values of CoBIt exceeding0.01 are shown. Values of CoBIt in bold exceed 0.05.
PATI boundary 3/4 2/3 1/2
ATIt 0.418035 0.633964 1.55967COBItBrizalina albatrossi – – 0.021Brizalina barbata 0.016 – –Brizalina simplex 0.012 – –Brizalina subaenariensis mexicana 0.099 – –Brizalina translucens 0.014 – –Bulimina aculeata 0.041 0.058 0.057Bulimina alazanensis – – 0.031Bulimina marginata 0.033 0.048 0.010Bulimina striata mexicana 0.013 0.046 –Cassidulina curvata 0.081 0.057 –Cassidulina laevigata – – 0.020Cassidulina norcrossi australis 0.014 – –Cibicidoides pachyderma 0.226 0.414 0.427Globocassidulina subglobosa subcalifornica – – 0.030Globocassidulina subglobosa 0.016 0.015 0.037Nummoloculina irregularis 0.014 0.010 –Oridorsalis umbonatus 0.013 0.020 –Osangularielloides rugosa 0.024 0.016 0.048Quinqueloculina cruziana – 0.038 0.017Sigmoilopsis schlumbergeri 0.043 0.032 0.036Sphaeroidina bulloides 0.029 0.030 0.021Uvigerina dirupta 0.058 – 0.030Uvigerina hispidocostata 0.030 0.023 0.038Uvigerina mediterranea 0.026 0.020 –
respectively; Table 3). Forty-six, 43 and 51 species contributed to ATIpacross the AB boundaries 1/2, 2/3 and 3/4 respectively. The meannumber of species contributing to ATIp across the AB and PATI bound-aries was compared using Student's t-test and the transformationln(S + 1). The result was statistically significant (mean across ABboundaries = 46.7 species, mean across PATI boundaries = 66.7 spe-cies; t = 2.83, p = 0.047, df = 4). The non-parametric test of meanslikewisewas significant. Twenty three species had a COBIp N 0.01 acrossany oneAB boundary, while 13 had a COBIp N 0.01 across oneAB bound-ary only. Six species had a COBIp N 0.05 across any AB boundary(Brizalina barbata, B. aculeata, B. marginata, B. striata mexicana,C. curvata, C. pachyderma). Only B. aculeata presented COBIp N 0.05across all three AB boundaries, while C. pachyderma had COBIp N 0.05across the lower two AB boundaries only. This reflects the significantnegative correlation between the proportional abundances ofB. aculeata and C. pachyderma.
The ATIt across the three AB boundaries ranged from 0.48 to 1.01across the AB1/2 and AB2/3 boundaries respectively (Table 4). Thenumber of species contributing to ATIt differed over the three AB bound-aries (AB1/2= 88 species, AB2/3= 110 species, AB3/4= 143 species).Themeannumber of species contributing to ATIp across the AB and PATIboundaries was compared in the same manner as for ATIt. The resultwas not statistically different (mean across AB boundaries= 113.7 spe-cies, mean across PATI boundaries= 100.7 species; t= 0.58, p=0.593,df = 4). Thirty species had a COBIt N 0.01 across any one boundary, in-cluding E. exigua, which presented COBIt N 0.01 across both the AB2/3and AB3/4 boundaries. This reflects the occurrence of this species in theupper part of the core. Ten species had a COBIt N 0.05 across any onePATI boundary (B. subaenariensis mexicana, B. aculeata, B. marginata,C. curvata, C. pachyderma, G. subglobosa, O. rugosa, S. schlumbergeri,Uvigerina dirupta and U. hispidocostata). Only C. pachyderma had aCOBIt N 0.05 across all three AB boundaries, while B. aculeata had aCOBIt N 0.05 across the lowermost two boundaries.
4.2. Piston core BGT096
This piston core yielded 10,644 benthic foraminifera (x = 273 persample, σ = 61.7 specimens). These were placed in 147 species, of

Table 4Thorough assemblage turnover index (ATIt) and thorough conditioned-on-boundaryindices (CoBIt) for abundance biozone boundaries from SHE analysis for biozone identifica-tion detected in Core BGT086. Only values of CoBIt exceeding 0.01 are shown. Values of CoBItin bold exceed 0.05.
Brizalina albatrossi – 0.031 0.015Brizalina barbata 0.016 – –Brizalina subaenariensis mexicana 0.068 0.027 0.012Bulimina aculeata 0.067 0.055 –Bulimina alazanensis – 0.047 –Bulimina marginata 0.075 0.019 –Bulimina striata mexicana 0.024 0.026 0.016Cassidulina curvata 0.084 0.027 0.016Cassidulina laevigata – 0.025 –Cibicides sp. – – 0.014Cibicidoides bradyi – – 0.012Cibicidoides mollis – 0.012 0.013Cibicidoides pachyderma 0.191 0.251 0.295Epistominella exigua – 0.011 0.016Eponides regularis 0.011 – –Gavelinopsis praegeri – – 0.021Globocassidulina subglobosa subcalifornica – 0.029 0.043Globocassidulina subglobosa – 0.025 0.103Lenticulina calcar 0.014 – –Martinottiella pallida – – 0.019Nummoloculina irregularis 0.021 – –Oridorsalis umbonatus 0.014 – –Osangularielloides rugosa 0.016 0.063 0.021Pseudononion atlanticum 0.010 – –Pullenia bulloides – 0.011 –Sigmoilopsis schlumbergeri 0.028 0.021 0.078Sphaeroidina bulloides – 0.020 –Uvigerina dirupta 0.120 0.084 –Uvigerina hispidocostata 0.016 0.055 0.018Uvigerina mediterranea 0.017 – –
92 B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
which 26 were either left in open nomenclature or merely comparedwith previously named species. Thirty five species (23.8%) were single-tons, while 58 (39.5%) were represented by b4 specimens. Thus, a largeproportion of the species were rare. The total assemblage was dominat-ed by C. pachyderma (25.2%) with lesser B. aculeata (13.0%), Eponidesregularis (6.7%), U. hispidocostata (5.9%), C. curvata (4.5%), S. bulloides(4.2%), S. schlumbergeri (3.6%) and Bulimina striata mexicana (3.2%).For distributions, see Fig. 4. These eightmost abundant species togetherformed 66.2% of the total assemblage. Of these, C. pachyderma,S. bulloides, C, curvata, U. hispidocostata and S. schlumbergeri formedN3% of the total assemblage from core BGT086. E. regularis Phleger andParker formed up to 67% of samples below 350 cmbsf in CoreBGT096 but only 0.7% of the total assemblage from Core BGT086. Itwas closely associated with Cassidulina laevigata d'Orbigny (1.9% ofrecovery from Core BGT086), which comprised 2.4% of recoveryfrom Core BGT196, but formed up to 12% of recovery from samplesbelow 280 cmbsf.
Some species distributions in core BGT096 are comparable to thosein core BGT086. B. aculeata ranged though the biofacies from whichthe slumping belowwas derived, forming amean of 7.5% of each samplebelow 280 cmbsf. In the in situ material above 274 cmbsf this spe-cies formed a mean of 15.6% of each sample (Fig. 4). In contrast,B. alazanensis was found primarily above 244 cmbsf, which suggeststhat this might correlate with 190 cmbsf in core BGT086. C. pachydermawas rare in the slumped interval below 290 cmbsf, increased abruptlyin abundance between 290 and 280 cmbsf and thereafter decreased inabundance to the top of the core. It lacked the marked, step-like de-crease in abundance found between 150 and 164 cmbsf in coreBGT086. S. schlumbergeri was rare in the slumped section but in-creased gradually in abundance through the in situ material, andU. hispidocostata showed a comparable distribution. E. exigua neverformed N1.4% of any sample, but was mostly concentrated towardsthe core top, between 20 and 140 cmbsf. COBIt N 0.01 across theAB2/3 and AB3/4 boundaries showed this species in core BGT086to be abundant in the upper part of that core.
5. Discussion
CHIRP profiles showed that core BGT086 contained in situ sedimentthroughout while core BGT096 contained some slumped material nearthe core base. Some conjecture can be made regarding the age and ori-gin of the slumped material. The difference in the abundant (N3%) spe-cies between the two cores is at least in part due to the slumping below~3 mbsf in Core BGT096, which brought in material from a differentbiofacies. A zone of carbonate-rich, iron-stained sand of early Holoceneage along the shelf edge east of Trinidad (60–70 mwater depth) marksthe low sea level of the late Quaternary (Carr-Brown, 1972). The Holo-cene on the shelf is, however, typically only ~0.05–1.6m thick,most Ho-locene sediment having been sequestered in the delta itself. Thiscarbonate-rich zone can be followed SE into northern Brazil, where sim-ilar sediment is found off the River Amazon at depths of N70 m (Vital,2014). Off Trinidad it is rich in Amphistegina gibbosa d'Orbigny(Drooger and Kaasschieter, 1958; Wilson, 2010), and it might be ex-pected that slumped material derived from it would contain specimensof this species. No such specimenswere found in the present study. Thismay be because the zone had not developed at the time of deposition ofthe slumpedmaterial in BGT096. The allochthonousmaterial in BGT096contained abundant E. regularis, whichwas rare elsewhere in both coresand had evidently brought sediment downslope from another biofaciesto the BGT096 site (water depth 700 m). Drooger and Kaasschieter(1958) recorded abundant E. regularis down to 150 m off Trinidad,while Hofker (1983) did not find it in samples from water deeperthan 400 m off Surinam and Guyana. Poag (1981) reported thatE. regularis is in the Gulf of Mexico “most frequently recorded inmoder-ate to low numbers on [the] outer shelf and upper to middle slope.”Thus the displaced material in BGT096 was derived from a mass trans-port complex of pre-Holocene age of outer shelf to upper slope origin.It is not clear, however, if it is of type 2 (upper slope-attached systemsdue to earthquakes or gas-hydrate disruptions) or 3 (locally detachedsystems formed by local seafloor instabilities) of the classification ofmass transport complexes presented by Moscardelli and Wood (2008).
When this displaced sediment is excluded, both cores show a transi-tion from an early low-diversity, C. pachyderma-dominated communitywith subdominant C. curvata to a later high-diversity, low-dominanceB. alazanensis–O. rugosa–E. exigua community. Galluzzo et al. (1990)suggested a B. aculeata–B. alazanensis association in the Grenada Basinto be indicative of low dissolved oxygen levels. Wilson and Costelloe(2011) found species of Uvigerina and Bulimina, indicative of a de-pressed dissolved-oxygen content (Kaiho, 1994), to be abundantbelow the Orinoco Plume on the Aves Ridge of the central EasternCaribbean Sea, where C. pachyderma was rare. Large Cibicidoides areindicative of oxic bottom waters (Kaiho, 1994). Wilson (2013) foundabundant C. pachyderma in the bathyal upper Quaternary of theSantaren Channel, off the western Bahama Platform, in associationwith abundant G. subglobosa and Planulina ariminensis, which he con-cluded to reflect a low flux of organic carbon. This suggests that thechange in fauna in both core BGT086 (whichwas detected by SHE anal-ysis, cluster analysis and ATI) and core BGT096 marked a change from alower to a higher organic carbon flux, which in turn depressed dissolvedoxygen levels.
There are two possible causes for the change in the organic flux. Inthe SE Caribbean region and adjacent tropical Atlantic Ocean, low-productivity waters occur in the open ocean while the nearshore,hyperpycnal Orinoco plume consists of high-productivitywater. The in-crease in flux at the sampled sites may reflect eastward progradation ofthe Orinoco Delta and Plataforma Deltana following the end Pleistocenetransgression. The transgression would have initially moved the locusof deposition westwards, but subsequent progradation might havebrought organic-rich prodelta clays to the site of deposition. Alterna-tively, the northward shift of the summertime ITCZ starting ~600 yrafter the end of the Younger Dryas (YD) cold interval at ca. 10.8 ka(Hoffmann et al., 2014) would have increased the volume of rainy

Fig. 4. The percentage abundances of selected species in Core BGT096. A. Brizalina albatrossi. B. Bulimina aculeata. C. Bulimina alazanensis. D. Cassidulina curvata. E. Cibicidoides pachyderma.F. Epistominella exigua. G. Eponides regularis.H.Globocassidulina subglobosa. I.Osangularielloides rugosa. J. Sigmoilopsis schlumbergeri. K. Sphaeroidina bulloides. L.Uvigerina hispidocostata. Thehorizontal dashed line indicates the top of the slumped interval interpreted from the CHIRP profile.
93B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
season outflow from the Orinoco, which in turn would have enlargedthe area covered by the hyperpycnal Orinoco plume. It is probable, how-ever, that these effects operated in concert, the leading edge of theplume being driven eastwards by both the increase in precipitationand delta progradation.
The change in community was abrupt at the more southerly SiteBGT086, but gradual at Site BGT096. This might reflect the position ofthe two sites relative to the plume front. One hypothesis can be pro-posed, although the precise mechanics connected with it have yet to
be investigated. It is possible that the more southerly Site BGT086 wasinundated every year by the Orinoco plume following the northwardmigration of the ITCZ, but that Site BGT096was initially only infrequent-ly inundated. As the delta prograded and inundation at Site BGT096becamemore regular during the later Holocene, it encouraged a gradualtransition to a B. alazanensis–O. rugosa–E. exigua community.
The Miocene–Pliocene Uvigerina biofacies of Batjes (1968) showsslight similarity to the B. alazanensis–O. rugosa–E. exigua community ofthis study, containing B. subaenariensis, Bulimina inflata, Cassidulina cf.

94 B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
laevigata and some Lenticulina spp. However, Batjes (1968) did not re-cord O. rugosa or E. exigua, nor did he record any biofacies comparableto the C. pachyderma community recorded from the present study.
Wilson (2008a) using ostracods found a front between Orinoco andAmazonwater within the Orinoco plume thatwas not parallel to bathy-metric contours. It seems probable that the front at the plume's leadingedge similarly does not parallel bathymetric contours. If this is so, thenthe change in fauna at Sites BGT086 and BGT096 cannot be used as apalaeobathymetric marker. This would require some revision of themanner in which micropalaentologists in the SE Caribbean region usebenthic foraminifera to suggest palaeodepths, the change in the faunanot reflecting changes in palaeodepth alone, but incorporating also themigration of the Orinoco plume.
The change from the C. pachyderma community to the B. alazanensis–O. rugosa–E. exigua community is at Site BGT086 detected by both thesample-wise assemblage turnover index (ATIs), SHE analysis for biozoneidentification (SHEBI) and various cluster analyses. However, these tech-niques detected other boundaries that require some explanation.The arrival of the ITCZ post-dated the end of the Younger Dryas (YD,11,500 yr BP) by 600 years. Hoffmann et al. (2014) found that deposition-al rates on the Orinoco fan range from 13 to 42 cm yr−1. This wouldequate to 7.8–25.2 cm of deposition at Site BGT086 between the end ofthe YD and the later arrival of the ITCZ, here postulated to be between194 and 220 cmbsf. SHEBI did not detect a further AB boundary until324–330 cmbsf, over 1mbelow the change in community. ATIs, however,detected a turnover boundary between 224 and 230 cmbsf, and it is pos-sible that this equates to the end of the YD. The position of the beginningof the Younger Dryas might be determined using nanodiamonds pro-duced by the cosmic-impact event at the onset of the Younger Dryas(≈12,800± 150 years BP; Kinzie et al., 2014). However, this was outsidethe scope of the present project.
The causes of the other PATI and AB boundaries in core BGT086,which indicate points of marked faunal turnover, have to be deter-mined, but it is possible that these relate to sea level change.Fairbanks (1989) showed on the basis of downed coral reefs offBarbados, ~200 km N of Trinidad, that sea level was 121 ± 5 m belowpresent during the last glacial maximum. Sea level rise probably beganat 23.0 ± 0.6 ka, coincident with melting of the southern Laurentideice-sheet, which is marked by retreat from last glacial maximum mo-raines in Wisconsin, USA (Ullman et al., 2015). The sunken coral reefsaround Barbados show that deglacial sea level rise was marked bythree intervals of especially rapid rise called meltwater pulses (MWP —Blanchon, 2011). Between ~17,100 and 14,000 yr BP, sea level increasedby 20 m. This first phase of deglaciation was followed by a rise of 24 min b1000 years (MWP-1A — see fig. 7.1 in Murray-Wallace andWoodroffe, 2014) that corresponds to the latter part of Termination 1aof deep sea records. The rate of sea level rise subsequently slowed,being low at the beginning of the Younger Dryas. The later part of theYounger Dryas is marked by increasing rates of sea level rise, however,that culminated inMWP-1B between 11,500–11,000 yr BP. This is closelytied to Termination 1B in the deep-sea, during which sea level rose by~28 m. This event has already been suggested to equate to the ATI peakat 224–230 cmbsf. Following a period of comparatively low sea levelrise, the rate again increased between 8000 and 7500 yr BP (MWP-1C).This might equate to the AB boundary at 124–130 cm detected by SHEBI.
Seasonality of organicmatter fluxmay have played a role in the devel-opment of the B. alazanensis–O. rugosa–E. exigua community. E. exigua ischaracteristic of cold (b3 °C) and well oxygenated (N3.5 ml/l) waterwith a low carbon flux to the seafloor (b3 g C m−2 yr−1) (Murgese andDe Deckker, 2005). It is also indicative of periodic (seasonal) delivery ofphytodetritus to the seafloor (Smart et al., 1994). Being an opportunistthat lives within the phytodetrital layer, it is rarely found in the underly-ing sediment (Murray, 2006). The appearance of E. exigua in the upperpart of cores BGT086 and BGT096may therefore reflect the developmentof a seasonal phytodetrital rain following the arrival of theOrinoco plume.The nutrient-rich plume at present expands and contracts between the
northern South American rainy and dry seasons (Odriozola et al., 2007,figure 3). Thus, the plume's eastern boundary at times lies east of thecore sites, and at other times west of it. This migration of water with ahigher primary productivity would induce the seasonality in thephytodetrital flux. The onset of this seasonality is in core BGT086markedby the first consistent occurrence of E. exigua at 190–194 cmbsf, at thebase of PATI-1 and the base of AB3. This is the only boundary commonto both ATI and SHEBI. In core BGT096 E. exigua occurs mostly between20 and 140 cmbsf. That Batjes (1968) did not record E. exigua in his Mio-cene–Pliocene Uvigerina biofacies assemblage might reflect a lack of sea-sonality in the outflow from the Orinoco then, the mean position of theITCZ being considerably north of Trinidad at that time (Van Vliet-Lanoe,2007). Alternatively, E. exiguamight have formed such a small proportionof the Uvigerina biofacial assemblage that Batjes (1968) did not think itworthwhile mentioning in his qualitative study. This seems possible,Wilson and Hayek (2015) having found E. exigua in the Middle MioceneGloborotalia fohsi robusta planktonic foraminiferal Zone (= Zone N12)of the Cipero Formation of Central Trinidad, where it formed up to 2.5%of each sample.
The mean ATIs (0.61) in the bathyal core BGT086 did not differ sig-nificantly from that (0.58) in the upper Quaternary of the abyssal ODP1261A of the Demerara Rise (tobserved = 0.544, tcritical = 1.991, df =77, p = 0.588) (see Wilson and Hayek, 2014b), but was significantlyless than that (0.71) for the upper Quaternary in the abyssal ODP Hole994C of Blake Ridge (tobserved = 2.136, tcritical = 1.985, df = 97, p =0.035) recorded by Hayek and Wilson (2013). It has yet to be deter-mined, however, if this indicates there is little variation in the meanvalues of ATIs between bathyal and abyssal sites off northern SouthAmerica.
Graphs of percentage abundances in core BGT086 suggest that thefundamental niche width varies between species, as noted also byWilson and Hayek (2015). Species that only gradually increase in abun-dance through the core (B. aculeata, S. bulloides) will have widerfundamental niches than those more abundant towards the corebase (C. pachyderma, C. curvata) or top (B. alazanesis, O. rugosa,U. hispidocostata). Galluzzo et al. (1990) suggested that bothB. aculeata and B. alazanensis in the Holocene of the Grenada Basinwere part of a low-oxygen assemblage. However, B. alazanensisshowed more specialist behaviour in the cores BGT086 andBGT096 off eastern Trinidad than did B. aculeata. This is apparentlyat odds with the suggestion by Ujiié (1990) that B. aculeata was offJapan “characteristically dominant during glacial intervals”. Howev-er, Ujiié's (1990) frequency polygon shows that B. aculeata wasabundant across Termination II in both glacial and interglacialtimes. Reiss and Hottinger's (1984) observations on the depth dis-tributions of species of Amphistegina in the neritic Red Sea showthat congeneric species occupy different niches. The present studyshows that congeneric bathyal species such as B. aculeata andB. alazanensis may also occupy fundamental niches of markedly dif-ferent dimensions.
Some evidence suggests that the amount of flux alone cannot be in-voked as the sole factor determining the change in fauna through coreBGT086. S. bulloides occupies primarily areas with an organic carbonflux of 4–9 g m−2 yr−1, while B. striata mexicana and C. pachyderma(as C. pseudoungerianus), both shown by CoBIt for abundance biozoneboundaries to be more abundant towards the core base, occupy pri-marily areas with a flux of 3–8 g m−2 yr−1 and 5–12 g m−2 yr−1
(Altenbach et al., 1999). C. laevigata, concentrated towards thecore top, primarily occupies areas with an organic carbon flux of6–20 g m−2 yr−1, while Uvigerina ex gr. auberiana, showing asimilar distribution in the core, prefers areas with a flux of4–9 g m−2 yr−1. These flux rates show considerable overlap. Itthus is possible that seasonality of input plays a role in the funda-mental niche width of species other than E. exigua, although otherfactors such as the degree of degradation of the organic matter or ni-trate availability might also play a part.

95B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
Deshmukh (1986, p. 181) ponderedwhether communities, as objec-tive realities formed fromdiscrete collections of species populations, oc-curred such that certain groups of species populations, which differfrom adjacent communities, would recur in similar environments.Whittaker (1975) suggested that there are two end-member distri-butions of populations of different species along an environmentalgradient. In the first, communities are clearly defined by groups of pop-ulations having more or less coincident distributions. In the second,each population is independently distributed along the gradient suchthat no clearly defined communities are formed. In core BGT086, thereare three groups of species: (1) those common towards the core base(C. pachyderma, C. curvata); (2) those abundant towards the core top(B. alazanesis, O. rugosa, U. hispidocostata); and (3) those presentthroughout (B. aculeata, S. bulloides). Groups (1) and (2) conform toWhittaker's (1975) suggested discrete communities, while Group(3) is reminiscent of his suggested independent populations. Such a dis-tribution is not unprecedented. Randall (1970) examined the distribu-tion of plants along a beach transect in Barbados. He found somegroups of species to have similar distributions while others were inde-pendent. Those species showing greater independence, which are com-parable to the third group of species recorded here, are those deemed tobe generalists.
6. Conclusions
Outflow from the Orinoco River currently forms a hyperpycnal, low-salinity plumewith high primary productivity that abuts sharply againstlow-productivity water of the western tropical Atlantic Ocean. Thisforms an extensive offshore front that currently lies east of Trinidadand crosses the Columbus Basin. However, the precise position of thefront varies between the northern South American rainy and dry sea-sons, which change the volume of Orinoco outflow. Bathyal benthicforaminifera suggest that the position of the front varied through atleast the late Quaternary, when the Orinoco delta was displaced west-ward during the transgressions accompanying glacial terminations,and then prograded eastward during highstands and ensuing regres-sions. The progradation after the end-Pleistocene was accompanied bya northward shift in the mean position of the intercontinental conver-gence zone (ITCZ), which would have increased Orinoco rainy-seasonoutflow and the areal extent of the Orinoco Plume. This northwardshift occurred about 600 yr after the end of the Younger Dryas.
Transgression induced the development of a low-productivity faunadominated by C. pachydermawith lesser C. curvata. In core BGT086 thisis abruptly replaced by a high-productivity B. alazanensis–O. rugosa–E. exigua assemblage, the change in fauna being detected by bothsample-wise assemblage turnover index (ATIs), SHE analysis and clusteranalysis. The presence of E. exigua in the latter assemblage but not theformer one indicates a change from a constant to a pulsed organic mat-ter flux. The transition between the two assemblages is here ascribed tothe arrival of the Orinoco plume following the abrupt migration of theITCZ, coupled with eastward progradation of the Orinoco delta. ATIsdetected a boundary in core BGT086 between PATI-3/4 that may equateto the end of the Younger Dryas and reflect the change in sea level atthat time. The causes of other peaks in ATIs and of abundance biozoneboundaries from SHE Analysis have yet to be determined and it is notyet clear if these other eventswere regional ones suitable for correlationbetween cores. While some species changed in proportional abundancemarkedly between the C. pachyderma–C. curvata and B. alazanensis–O. rugosa–E. exigua assemblages, others (B. aculeata, S. bulloides) didnot. This is concluded to reflect differences in the species' fundamentalniche widths. This indicates that the cogeneric B. alazanensis andB. aculeata have different niche widths, the former being a specialistspecies and the latter a generalist. Those using foraminifera to recon-struct the evolution of the Orinoco deep-sea fan will need to take intoaccount not only the impact of the offshore estuarine front on faunal as-semblages, but also the discordant relationships between the front at
the sea surface and palaeobathymetric contours. Those working offother major rivers might find similar phenomena.
Acknowledgements
Thanks are due to BG (Trinidad and Tobago) for sample donationand to Ron Daniel for aid in interpreting the CHIRP profiles. BW wouldlike to thank the Ministry of Energy and Energy Affairs of Trinidad andTobago for financial support for this project, and Andrew Jupiter inparticular for helping arrange this support. Constructive reviews byIan Boomer and Flavia Fiorini are gratefully acknowledged.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.marmicro.2015.11.004.
References
Agard, J.B.R., Gobin, J.F., 2000. The Lesser Antilles, Trinidad and Tobago. In: Sheppard, C.(Ed.)Seas at the Millennium: An Environmental Evaluation. Elsevier Science Limited,pp. 627–641.
Altenbach, A.V., Pflaumann, U., Schiebel, R., Thies, A., Timm, S., Trauth, M., 1999. Scalingpercentages and distributional patterns of benthic foraminifera with flux rates oforganic carbon. J. Foraminifer. Res. 29, 173–185.
Arbuszewski, J.A., Cléroux, C., Bradtmiller, L., Mix, A., 2013. Meridional shifts of the Atlanticintertropical convergence zone since the Last Glacial Maximum. Nat. Geosci. 6, 959–962.
Aslan, A., White, W.A., Warne, A.G., Guevara, E.H., 2003. Holocene evolution of the west-ern Orinoco Delta, Venezuela. Geol. Soc. Am. Bull. 115, 479–498.
Batjes, D.A.J., 1968. Palaeoecology of foraminiferal assemblages in the late Miocene cruseand forest formations of Trinidad. Antilles, Fourth Caribbean Geological Conference,1965, Trinidad, pp. 141–156.
Berger, W.H., Parker, F.L., 1970. Diversity of planktonic foraminifera in deep-sea sedi-ments. Science 168, 1345–1347.
Blanchon, P., 2011. Meltwater pulses. In: Hopley, D. (Ed.)Encyclopaedia of Modern CoralReefs. Springer, pp. 683–690.
Buzas, M.A., Hayek, L.-A.C., 1998. SHE analysis for biofacies identification. J. Foraminifer.Res. 28, 233–239.
Callec, Y., Deville, E., Desaubliaux, G., Griboulard, R., Huyghe, P., Mascle, A., Mascle, G.,Noble, M., Padron de Carillo, C., Schmitz, J., 2010. The Orinoco turbidite system: tec-tonic controls on sea-floor morphology and sedimentation. AAPG Bull. 94, 869–887.
Carr-Brown, B., 1972. The Holocene/Pleistocene contact in the offshore area east ofGaleota Point, Trinidad, West Indies. VI Conferencia Geologica Del Caribe, Margarita,Venezuela, pp. 381–397.
Carr-Brown, B., 2007. The contribution of Trinidad micropaleontology to global E&P.100 Years of Petroleum in Trinidad and Tobago: Celebrating a Century of CommercialOil Production. Government of Trinidad and Tobago, Port of Spain, Trinidad,pp. 158–167.
Chen, S., Steel, R.J., Dixon, J.F., Osman, A., 2014. Facies and architecture of a tide-dominated segment of the Late Pliocene Orinoco Delta (Morne L'Enfer Formation)SW Trinidad. Mar. Pet. Geol. 57, 208–232.
Cheng, H., Edwards, R.L., Broecker, W.S., Denton, G.H., Kong, X., Wang, Y., Zhang, R., Wang,X., 2009. Ice age terminations. Science 326, 248–252.
Chin-Leo, G., Benner, R., 1992. Enhanced bacterioplankton production and respiration atintermediate salinities in the Mississippi River plume. Mar. Ecol. Prog. Ser. 87,87–103.
Culver, S.J., 1988. New foraminiferal depth zonation of the northwestern Gulf of Mexico.PALAIOS 3, 69–85.
Del Castillo, C.E., Coble, P.G., Morell, J.M., López, J.M., Corredor, J.E., 1999. Analysis of theoptical properties of the Orinoco River plume by absorption and fluorescence spec-troscopy. Mar. Chem. 66, 35–51.
Deshmukh, I., 1986. Ecology and Tropical Biology. Blackwell Scientific Publications, PaloAlto, California (387 pp.).
Drooger, C.W., Kaasschieter, J.P., 1958. Foraminifera of the Orinoco–Trinidad–Paria Shelf.Report of the Orinoco Shelf Expedition, Verhandlungen Koninklijk NederlandAkademie Wetenschappelijke 4, pp. 1–108.
Fairbanks, R.G., 1989. A 17,000 year glacio-eustatic sea level record: influence of glacialmelting rates on the Younger Dryas event and deep-ocean circulation. Nature 342,637–642.
Figueroa, D.F., 2007. Variation of Planktonic Community Structure Along the OrinocoRiver Plume MSc Thesis Department of Marine Sciences. University of Puerto Rico,Mayaguez, p. 29.
Foster, G.L., Sexton, P.F., 2014. Enhanced carbon dioxide outgassing from the easternequatorial Atlantic during the last glacial. Geology 42, 1003–1006.
Furlonge, H., Kaiser, M., 2010. Overview of natural gas sector developments in Trinidadand Tobago. Int. J. Energy Sect. Manage. 4, 535–554.
Gaby, M.L., Sen Gupta, B.K., 1985. Late Quaternary benthic foraminifera of the VenezuelaBasin. Mar. Geol. 68, 125–144.
Galluzzo, J.J., Sen Gupta, B.K., Pujos, M., 1990. Holocene deep-sea foraminifera of theGrenada Basin. J. Foraminifer. Res. 20, 195–211.

96 B. Wilson, L.-A.C. Hayek / Marine Micropaleontology 121 (2015) 85–96
Garciacaro, E., Escalona, A., Mann, P., Wood, L., Moscardelli, L., Sullivan, S., 2011. Structuralcontrols on Quaternary deepwater sedimentation, mud diapirism, and hydrocarbondistribution within the actively evolving Columbus foreland basin, eastern offshoreTrinidad. Mar. Pet. Geol. 28, 149–176.
Hallock, P., 1984. Distribution of selected species of living algal symbiont-bearing forami-nifera on two Pacific coral reefs. J. Foraminifer. Res. 14, 250–261.
Hammer, Ø., Harper, D.A.T., 2005. Paleontological Data Analysis. Wiley-Blackwell, Oxford,UK.
Hammer, Ø., Harper, D.A.T., Ryan, P.D., 2001. PAST: paleontological statistics softwarepackage for education and data analysis. Palaeontol. Electron. (http://palaeo-electronica.org/2001_2001/past/issue2001_2001.htm).
Hayek, L.C., Buzas, M.A., 2013. On the proper and efficient use of diversity measures forindividual field samples. J. Foraminifer. Res. 43, 305–313.
Hayek, L.C., Wilson, B., 2013. Quantifying assemblage turnover and species contributionsat ecologic boundaries. PLoS One 8, e74999.
Hayward, B.W., Holzmann,M., Grenfell, H.R., Pawlowski, J., Triggs, C.M., 2004. Morpholog-ical distinction of molecular types in Ammonia—towards a taxonomic revision of theworld's most commonly misidentified foraminifera. Mar. Micropaleontol. 50,237–271.
Hoffmann, J., Bahr, A., Voigt, S., Schönfeld, J., Nürnberg, D., Rethemeyer, J., 2014.Disentangling abrupt deglacial hydrological changes in northern South America:insolation versus oceanic forcing. Geology 42, 579–582.
Hofker, J., 1983. Zoological exploration of the continental shelf of Surinam: the foraminif-era of the shelf of Surinam and the Guyanas. Zoologische Verhandelingen Uitgegevendoor het Rijksmuseum van Natuurlijke Histoire te Leiden 201, pp. 1–75.
Holbourn, A., Henderson, A.S., MacLeod, N., 2013. Atlas of Benthic Foraminifera. JohnWiley and Sons, Chichester, UK.
Hu, C., Montgomery, E.T., Schmitt, R.W., Muller-Karger, F.E., 2004. The dispersal of the am-azon and Orinoco River water in the tropical Atlantic and Caribbean Sea: observationfrom space and S-PALACE floats. Deep-Sea Res. II 51, 1151–1171.
Hutchinson, G.E., 1957. Concluding remarks. Cold Spring Harb. Symp. Quant. Biol. 22,415–427.
Jeffries, M.J., Lawton, J.H., 1984. Enemy free space and the structure of ecological commu-nities. Biol. J. Linn. Soc. 23, 269–286.
Jones, R.W., 1998. Palaeonenvironmental interpretation of the Late Miocene and Plioceneof Trinidad based on micropalaeontological data. In: Ali, W., Paul, A., Young On, V.(Eds.), Transactions of the 3rd Geological Conference of the Geological Society ofTrinidad and Tobago and 14th Caribbean Geological Conference, Port-of-Spain,Trinidad, pp. 88–101.
Kaiho, K., 1994. Benthic foraminiferal dissolved-oxygen index and dissolved-oxygenlevels in the modern ocean. Geology 22, 719–722.
Kempler, S., 2012. Classic CZCS Scenes Chapter 5: Orinoco River Plume in the Caribbean.(http://disc.sci.gsfc.nasa.gov/education-and-outreach/additional/science-focus/classic_scenes/05_classics_orinoco.shtml).
Kinzie, C.R., Hee, S.S.Q., Stich, A., Tague, K.A., Mercer, C., Razink, J.J., Kennett, D.J., DeCarli,P.S., Bunch, T.E., Wittke, J.H., Israde-Alcántara, I., Bischoff, J.L., Goodyear, A.C.,Tankersley, K.B., Kimbel, D.R., Culleton, B.J., Jon, M.E., Stafford, T.W., Kloosterman,J.B., Moore, A.M.T., Firestone, R.B., Tortosa, J.E.A., Pardo, J.F.J., West, A., Kennett, J.P.,Wolbach, W.S., 2014. Nanodiamond-rich layer across three continents consistentwith major cosmic impact at 12,800 Cal BP. J. Geol. 122, 475–506.
Kruit, C., 1954. Distribution of faunal remains, 2: microfauna. In: Van Andel, T.H., Postma,H. (Eds.), Recent Sediments of the Gulf of Paria: Reports of the Orinoco Shelf Expedi-tion vol. 1. North-Holland Publishing Company, Amsterdam, Holland, pp. 117–134.
López, R., López, J.M., Morell, J., Corredor, J.E., Del Castillo, C.E., 2013. Influence of theOrinoco River on the primary production of eastern Caribbean surface waters.J. Geophys. Res.: Oceans 118, 4617–4632.
Manheim, F.T., Hathaway, J.C., Uchupi, E., 1972. Suspendedmatter in surface waters of thenorthern Gulf of Mexico. Limnol. Oceanogr. 17, 17–27.
Matera, N.J., Lee, J.J., 1972. Environmental factors affecting the standing crop of foraminif-era in sublittoral and psammolittoral communities of a Long Island salt marsh. Mar.Biol. 14, 89–103.
Morin, P.J., 1999. Community Ecology. Blackwell Science, Malden, Massachusetts.Moscardelli, L., Wood, L., 2008. New classification system for mass transport complexes in
offshore Trinidad. Basin Res. 20, 73–98.Muller, J., 1959. Palynology of Recent Orinoco delta and shelf sediments; reports of the
Orinoco shelf expedition, volume 5. Micropaleontology 5, 1–32.Murgese, D.S., De Deckker, P., 2005. The distribution of deep-sea benthic foraminifera in
core tops from the eastern Indian Ocean. Mar. Micropaleontol. 56, 25–49.Murray, J.W., 2001. The niche of benthic foraminifera, critical thresholds and proxies. Mar.
Micropaleontol. 41, 1–7.Murray, J.W., 2006. Ecology and Applications of Benthic Foraminifera. Cambridge University
Press, Cambridge, UK.Murray-Wallace, C.V., Woodroffe, C.D., 2014. Quaternary Sea-level Changes: A Global
Perspective. Cambridge University Press, Cambridge, UK.Odriozola, A.L., Varela, R., Hu, C., Astor, Y., Lorenzoni, L., Müller-Karger, F.E., 2007. On the
absorption of light in the Orinoco River plume. Cont. Shelf Res. 27, 1447–1464.Oki, T., Kanae, S., 2006. Global hydrological cycles and world water resources. Science 313,
1068–1072.Pascual, A., García, B.M., Lázaro, J.R., Pujos, M., 2009. Asociaciones de foraminíferos
bentónicos recientes en la plataforma marina de las Guayanas. Geogaceta 46, 75–78.Pflum, C.E., Frerichs, W.E., 1976. Gulf of Mexico deep-water foraminifers. Cushman Foun-
dation for Foraminiferal Research, Special Publication 14, pp. 1–125.Phleger, F.B., 1976. Benthic foraminiferids as indicators of organic production in marginal
marine areas, first international symposium on benthonic foraminifera of continentalmargins. Part A: ecology and biology. Mar. Sed. Spec. Publ. 1, 107–117.
Phleger, F.B., Parker, F.L., 1951. Foraminifera species, ecology of foraminifera, northwestGulf of Mexico. Geological Society of America Memoir 1–64.
Piña-Ochoa, E., Høgslund, S., Geslin, E., Cedhagen, T., Revsbech, N.P., Nielsen, L.P.,Schweizer, M., Jorissen, F., Rysgaard, S., Risgaard-Petersen, N., 2010. Widespreadoccurrence of nitrate storage and denitrification among Foraminifera and Gromiida.Proc. Natl. Acad. Sci. 107, 1148–1153.
Poag,W.C., 1981. Ecologic Atlas of Benthic Foraminifera of the Gulf of Mexico. HutchinsonRoss Publishing Company, Woods Hole.
Poag, C.W., Valentine, P.C., 1976. Biostratigraphy and ecostratigraphy of the Pleistocenebasin Texas–Louisiana continental shelf. Gulf Coast Assoc. Geol. Soc. Trans. 26,185–256.
Prentice, C.S., Weber, J.C., Crosby, C.J., Ragona, D., 2010. Prehistoric earthquakes on theCaribbean–South American plate boundary, Central Range fault, Trinidad. Geology38, 675–678.
Randall, R.E., 1970. Vegetation and environment on the Barbados coast. J. Ecol. 58,155–172.
Reiss, Z., Hottinger, L., 1984. The Gulf of Aqaba: Ecological Micropaleontology. Springer-Verlag, Berlin.
Rühlemann, C., Frank, M., Hale, W., Mangini, A., Mulitza, S., Müller, P.J., Wefer, G., 1996.Late Quaternary productivity changes in the western equatorial Atlantic: evidencefrom 230Th-normalized carbonate and organic carbon accumulation rates. Mar.Geol. 135, 127–152.
Saunders, J.B., Bolli, H.M., 1979. Trinidad's contribution to world biostratigraphy. FourthLatin American Geological Congress, Trinidad and Tobago, pp. 781–795.
Schmoker, C., Hernández-León, S., Calbet, A., 2013. Microzooplankton grazing in theoceans: impacts, data variability, knowledge gaps and future directions. J. PlanktonRes. 35, 691–706.
Sen Gupta, B.K., Machain-Castillo, M.L., 1993. Benthic foraminifera in oxygen-poorhabitats. Mar. Micropaleontol. 20, 183–201.
Sen Gupta, B.K., Pujos, M., Pons, J.-C., Galluzzo, J.J., Aharon, P., Parra, M., 1991. Micropale-ontological, mineralogical, and geochemical record in Late Quaternary cores fromabyssal Grenada Basin. Geo-Mar. Lett. 11, 2–8.
Smart, C.W., King, S.C., Gooday, A.J., Murray, J.W., Thomas, E., 1994. A benthic foraminiferalproxy of pulsed organic matter paleofluxes. Mar. Micropaleontol. 23, 89–99.
Soberón, J., Peterson, A.T., 2005. Interpretation ofmodels of fundamental ecological nichesand species' distributional areas. Biodivers. Inform. 2, 1–10.
Steineck, P.L., Bergstein, J., 1979. Foraminifera from Hommocks salt-marsh, LarchmontHarbor, New York. J. Foraminifer. Res. 9, 147–158.
Ujiié, H., 1990. Bathyal benthic foraminifera in a piston core from east off the Miyakoislands, Ryukyu Island Arc. Bull. Coll. Sci. Univ. Ryukyus 49, 1–60.
Ullman, D.J., Carlson, A.E., LeGrande, A.N., Anslow, F.S., Moore, A.K., Caffee, M., Syverson,K.M., Licciardi, J.M., 2015. Southern Laurentide ice-sheet retreat synchronous withrising boreal summer insolation. Geology 43, 23–26.
Van Vliet-Lanoe, B., 2007. The autocyclic nature of glaciations. Bull. Soc. Géol. Franc. 178,247–262.
Vital, H., 2014. The north and northeast Brazilian tropical shelves. In: Chiocci, F.L., Chivas,A.R. (Eds.), Continental Shelves of the World: Their Evolution During the Last Glacio-eustatic Cycle. Geological Society Memoir, London, pp. 35–46.
Whittaker, R.H., 1975. Communities and Ecosystems. MacMillan, New York.Wilson, B., 2006. Trouble in paradise? A comparison of 1953 and 2005 benthonic forami-
niferal seafloor assemblages at the Ibis Field, offshore eastern Trinidad, West Indies.J. Micropalaeontol. 25, 157–164.
Wilson, B., 2008a. Distributions of ostracod (Crustacea) biofacies on the continental shelfoff south-east Trinidad, western central Atlantic Ocean, suggest the location of an off-shore river-induced front within the Orinoco Plume. Senckenb. Lethaea 88, 199–211.
Wilson, B., 2008b. Using SHEBI (SHE analysis for Biozone Identification): to proceed fromthe top down or the bottom up? A discussion using two Miocene foraminiferal suc-cessions from Trinidad, West Indies. PALAIOS 23, 636–644.
Wilson, B., 2010. The significance of iron-stained foraminifera off SE Trinidad,West Indies,western central Atlantic Ocean. Geol. Mag. 147, 728–736.
Wilson, B., 2013. Ecostratigraphic regime shift during late Quaternary Marine IsotopeStages 8–9 in Santaren Channel, western tropical Atlantic Ocean: benthonic forami-niferal evidence from ODP Hole 1006A. J. Foraminifer. Res. 43, 143–153.
Wilson, B., Costelloe, A., 2011. Benthonic foraminiferal paleoecology of the Pleistocene inDSDP Hole 148, Aves Ridge, Eastern Caribbean Sea. J. Foraminifer. Res. 41, 363–370.
Wilson, B., Hayek, L.C., 2014a. Ontology confounds reproducibility in ecology and climatescience. Life 2, 13–30.
Wilson, B., Hayek, L.C., 2014b. Foraminifera on the Demerara Rise offshore Surinam:crustal subsidence or shallowing of an oxygen minimum zone? Geol. Mag. 152,788–801.
Wilson, B., Hayek, L. C., 2015. Distinguishing relative specialist and generalist species inthe fossil record. Mar. Micropaleontol., v. 119, p. 7–16.
Wilson, B., Coimbra, J.C., Hayek, L.C., 2014. Ostracoda (Arthropoda, Crustacea) in aMiocene oxygen minimum zone, Trinidad,West Indies: a test of the platycopid signalhypothesis. J. S. Am. Earth Sci. 54, 210–216.
Wood, L.J., 2000. Chronostratigraphy and tectonostratigraphy of the Columbus Basin,eastern offshore Trinidad. AAPG Bull. 84, 1905–1928.
van der Zwaan, G.J., Jorissen, F.J., 1991. Biofacial patterns in river-induced shelf anoxia. In:Tyson, R.V., Pearson, T.H. (Eds.), Modern and Ancient Continental Shelf Anoxia.Geological Society of London, pp. 65–82.
van der Zwaan, G.J., Duijnstee, I.A.P., den Dulk, M., Ernst, S.R., Jannink, N.T., Kouwenhoven,T.J., 1999. Benthic foraminifers: proxies or problems? A review of paleoecologicalconcepts. Earth Sci. Rev. 46, 213–236.