juliana ribeiro mariotto estÁgio de docÊncia enzimas
TRANSCRIPT

JULIANA RIBEIRO MARIOTTO
ESTÁGIO DE DOCÊNCIA
Enzimas
JULHO 2006

ENZIMAS
1 Conceito
Enzimas são catalisadores biológicos, formados por longas cadeias de moléculas
pequenas, chamadas de aminoácidos. São, portanto, um tipo de proteína com atividade
catalítica, sendo encontradas na natureza em todos os seres vivos. Sua função é viabilizar a
atividade das células, quebrando moléculas ou juntando-as para formar novos compostos. A
singularidade desses compostos decorre do elevado grau de especificidade ao substrato em
condições moderadas, sob as quais atuam. Algumas enzimas são específicas para determinado
tipo de ligação química, como por exemplo, a capacidade da α-amilase de romper unicamente
as ligações α-1,4 das moléculas de amido. Outras são específicas para um tipo particular de
isômeros ópticos, como a oxidação da β-D-glicose pela glicose oxidase.
“Toda a enzima é uma proteína, mas nem toda proteína é uma enzima”.
2 Características das Enzimas
• São produtos naturais biológicos;
• Apresentam um alto grau de especificidade;
• Reações baratas e seguras;
• Possuem mecanismo de “turnover”, desempenhando a mesma função consecutivamente,
sem serem consumidas no processo;
• São altamente eficientes, acelerando a velocidade das reações de 108 a 1011 vezes;
• São econômicas, reduzindo a energia de ativação necessária para a reação catalisada;
• Não são tóxicas.
3 Proteínas
Exceto um grupo de moléculas de RNA com propriedades catalíticas (ribozimas),
todas as enzimas são proteínas. A atividade catalítica depende da sua conformação protéica
nativa, sendo esta perdida quando a enzima é desnaturada ou dissociada em subunidades.

Os componentes das enzimas, os aminoácidos, apresentam um átomo de carbono
ligado a uma carboxila, a um grupo amino e a um átomo de hidrogênio. O quarto substituinte
é uma cadeia (grupo R) específica para cada aminoácido (Figura 1).
Figura 1. Representação esquemática do aminoácido.
Para serem ativas, algumas enzimas requerem apenas seus resíduos de aminoácidos.
Outras requerem componentes químicos adicionais chamados cofatores. Um cofator pode ser
um ou mais íons inorgânicos, tais como Fe2+, Mg2+, Mn2+ ou Zn2+, ou uma molécula orgânica
complexa, chamada coenzimas. Algumas enzimas requerem ambos, coenzima e íon metálico
para exibirem sua atividade. Uma coenzima, ou íon metálico, que está covalentemente ligada
à parte protéica da enzima é chamada grupo prostético. Uma enzima completa,
cataliticamente ativa, unida à sua coenzima e/ou íons metálicos, é chamada de holoenzima. A
parte exclusivamente protéica desta enzima é chamada de apoenzima ou apoproteína.
3.1 Coenzimas
São aceptores de átomos ou grupos funcionais retirados do substrato em uma dada
reação e como doadores destes mesmos grupos ao participarem de uma outra reação e, por
isto, diz-se que as coenzimas são transportadoras de determinados grupos (Tabela 1). O fato
de as coenzimas estarem sendo constantemente recicladas permite que suas concentrações
celulares possam ser bastante reduzidas, muito menores do que as concentrações do substrato.
Tabela 1. Coenzimas e grupos aos quais se ligam/desligam em diferentes reações.
Coenzima Grupo transportado Adenosina trifosfato (ATP) Fosfato
Biotina CO2 Coenzima A Acila
Flavina adenina dinucleotídeo (FAD) Hidrogênio Tetraidrofolato Carbono
Nicotinamida adenina dinucleotídeo (NAD+) Hidreto Tiamina pirofosfato (TPP) Aldeído

Em alguns casos, a coenzima encontra-se covalentemente ligada à molécula
enzimática, constituindo um grupo prostético da proteína; em outros casos, a coenzima é uma
molécula “livre”, reunindo-se à enzima apenas no momento da catálise.
Algumas coenzimas são integralmente sintetizadas pelas células; outras apresentam
em sua molécula um componente orgânico que não pode ser sintetizado pelos animais
superiores. Esse componente, ou um precursor imediato, deve então ser obtido através da
dieta, constituindo uma vitamina. As vitaminas são compostos orgânicos indispensáveis ao
crescimento e funções normais dos animais superiores. Estas são classicamente divididas em
hidrossolúveis e lipossolúveis, sendo as vitaminas hidrossolúveis aquelas que possuem função
de coenzima ou fazem parte de moléculas de coenzimas.
4 Estruturas das enzimas
4.1 Estrutura primária
Na molécula protéica, os aminoácidos estão ligados uns aos outros através da ligação
peptídica (Figura 2), estabelecida entre o grupo α-carboxila de um aminoácido e o grupo α-
amino do aminoácido subseqüente, formando uma longa cadeia. Grupos carboxila e grupos
amino do radical R jamais participam da ligação peptídica.
O que caracteriza cada enzima é o número de aminoácidos componentes de sua cadeia
e a ordem em que eles se encontram, ou seja, a sua estrutura primária. Assim, apesar de
constituídas por apenas 20 aminoácidos diferentes, as possibilidades de estruturas diversas
para as proteínas são muito grandes. A estrutura primária é responsável pelas estruturas de
ordem superior que a proteína exibe em sua forma celular, ou nativa.
Figura 2. Esquema da formação da ligação peptídica.

4.2 Estrutura secundária
Dois tipos diferentes de organização regular, chamadas estruturas secundárias, são
encontradas nas enzimas. A cadeia peptídica pode ter segmentos organizados em α-hélice,
sendo formada e estabilizada por pontes de hidrogênio estabelecidas entre o átomo de
nitrogênio e o átomo de oxigênio (Figura 3).
Cada ligação peptídica oferece os elementos para a formação da ponte de hidrogênio:
um átomo de hidrogênio covalentemente ligado ao nitrogênio e o oxigênio, preso ao carbono
por uma dupla ligação. A ligação por ponte de hidrogênio é estabelecida entre os átomos
constituintes de uma ligação peptídica qualquer e os átomos da quarta ligação peptídica
subseqüente, que a volta da hélice aproximou da primeira. Cada ponte de hidrogênio é uma
ligação fraca, mas o grande número dessas interações confere muita estabilidade à estrutura
que mantêm. (Figura 4).
Figura 3. Ponte de hidrogênio, formada com os elementos da ligação peptídica.
Figura 4. Esquema da α-hélice, estabilizada por pontes de hidrogênio.

A folha β-pregueada é formada por um arranjo paralelo de dois ou mais segmentos de
cadeias peptídicas, mantidas por pontes de hidrogênio formadas pelos mesmos elementos que
constituem as pontes de hidrogênio da α-hélice (Figura 5). Nesse caso a ponte de hidrogênio
une dois segmentos distintos da cadeia protéica.
Figura 5. Esquema da folha β-pregueada, estabilizada por pontes de hidrogênio.
As enzimas apresentam, em sua conformação espacial, as duas estruturas secundárias;
parte da cadeia está organizada em α-hélice e parte em folha β-pregueada. Aparecem também
regiões de conformação irregular, conectando os segmentos com arranjo definido (Figura 6).
Figura 6. Estrutura da lisozima, indicando segmentos em α-hélice, segmentos em folha β-pregueada e
regiões sem estrutura regular.

4.3 Estrutura terciária
Esta estrutura descreve a conformação tridimensional que a molécula protéica assume
em solução, explicando o dobramento da cadeia peptídica com os enrolamentos, dobras e
voltas que a compõem e que a levam a uma forma geral globular. As ligações químicas que
estabelecem e mantêm a estrutura terciária são formadas sempre entre os grupos R dos
aminoácidos. Estes grupos R podem ser divididos em dois tipos: apolares (hidrofóbicos) e
polares. Como a água é um solvente polar, os grupos R apolares tendem a aproximar-se uns
dos outros, excluindo a água, situando-se no interior da molécula, enquanto os grupos polares
voltam-se para a superfície. Essa localização dos grupos R constitui uma força poderosa de
dobramento da cadeia polipeptídica.
Outras forças devem ser consideradas. Grupos R com carga elétrica positiva fazem
ligações eletrostáticas com grupos R com carga negativa. Outras forças importantes são as
pontes de hidrogênio, formadas entre grupos R. Ao contrário das pontes de hidrogênio da
estrutura secundária, estas não apresentam qualquer padrão regular, pois a localização dos
grupos R, capazes de oferecer os elementos para a formação da ponte de hidrogênio, estão
distribuídos irregularmente ao longo da cadeia peptídica segundo a estrutura primária da
enzima.
As interações responsáveis pela estrutura tridimensional das enzimas são todas forças
fracas. Mas pode ser encontrada também uma ligação covalente fazendo parte das ligações
que mantêm a estrutura terciária: são as pontes dissulfeto, ou pontes S-S, formadas pela
oxidação de dois grupos –SH, cada um dos quais presente na cadeia lateral de um resíduo de
cisteína, o único aminoácido a apresentar –SH no grupo R. Designa-se resíduo de aminoácido
à fração da molécula de aminoácido efetivamente inserida na cadeia protéica. A Figura 7
mostra as principais ligações da estrutura terciária.
Obs: A forma espacial da enzima, responsável pela sua função, é resultado indireto de sua
estrutura primária.

Figura 7. Esquema das principais ligações responsáveis pela estrutura terciária das enzimas. (a) interação hidrofóbica; (b) ponte dissulfeto; (c) ligação eletrostática; (d) ponte de hidrogênio; (e) região
sem estrutura definida.
4.4 Estrutura quaternária
Estrutura quaternária é a organização presente nas proteínas compostas de uma cadeia
polipeptídica e descreve quantos e quais monômeros compõem a molécula e como estes
monômeros estão associados. As forças que mantêm unidos os monômeros são as mesmas
que mantêm a estrutura terciária: interações hidrofóbicas, pontes de hidrogênio e ligações
salinas, formadas, entre grupos R de aminoácidos pertencentes a cadeias polipeptídicas
diferentes (Figura 8). A exceção são as pontes dissulfeto, ausentes na estrutura quaternária.
Figura 8. Estrutura quaternária da hemoglobina (proteína não-enzimática), composta por quatro
cadeias polipeptídicas, iguais duas a duas.

5 Classificação e nomenclatura das enzimas
A Comissão de Enzimas (CE) da União Internacional de Bioquímica (IUB) coloca
todas as enzimas em seis grandes classes (Tabela 2), cada uma delas comportando subclasses
conforme o tipo de reação catalisada. A cada enzima é assinalado um número classificatório
de quatro dígitos de um nome sistemático:
• Primeiro número designa a qual das seis classes a enzima pertence;
• Segundo número indica o tipo de ligação que a enzima atua;
• Terceiro número é uma subclassificação do tipo de ligação;
• Quarto número é apenas um número de série.
Tabela 2. Principais classes das enzimas.
Nº Classe da enzima Tipo de reação catalisada Atuação
1 Oxidorredutases Reações de oxidação-redução ou transferência de elétrons
CH-OH C=O C=O-
CH-NH2 CH-NH-
NADH, NADPH
2 Transferases Transferem grupos funcionais entre moléculas
Grupos com um carbono Grupos aldeído ou cetona
Grupos acil Grupos glicosil Grupos fosfatos
Grupos contendo enxofre
3 Hidrolases Reações de Hidrólise
Ésteres Ligações glicosídicas Ligaçõespeptídicas Outras ligações C-N
Anidridos ácidos
4 Liases
Catalisam a quebra de ligações covalentes e a remoção de
moléculas de água, amônia e gás carbônico
=C=C= =C=O =C=N-
5 Isomerases Transferência de grupos dentro da
mesma molécula para formar isômeros
racemases
6 Ligases
Catalisam reações de formação de novas moléculas a partir da ligação entre duas pré-existentes, sempre
às custas de energia
C-O C-S C-N C-C

Exemplo: ATP + D-glicose ADP + D-glicose-6-fosfato
O nome sistemático formal da enzima que catalisa a reação é ATP:glicose
fosfotransferase, indicando que ela catalisa a transferência de um grupo fosfato do ATP para a
glicose. O seu número na classificação enzimática é 2.7.1.1; o primeiro dígito (2) denota o
nome da classe (transferase); o segundo dígito (7) a subclasse (fosfotransferase); o terceiro
dígito (1), fosfotransferases que trabalham com um grupo hidroxila como receptor; e o quarto
dígito (1), indica ser a D-glicose o receptor do grupo fosfato. Quando o nome sistemático de
uma enzima é muito longo ou de uso difícil, um nome trivial pode ser usado, no exemplo este
nome é hexoquinase.
6 Ação catalítica
O primeiro ponto a considerar é a grande diferença de tamanho entre a enzima e seus
substratos. Um exemplo numérico: a enzima uréase, que catalisa a reação de hidrólise da uréia
tem peso molecular aproximado de 500.000 dáltons, enquanto a uréia tem peso molecular de
60 e a água de 18. O investimento energético para a síntese de uma molécula protéica tão
grande justifica-se pela obtenção de uma estrutura muito precisa, com reentrâncias de forma
apropriada e com grupos químicos localizados em posições exatas para servir à catalise. Para
que esta seja exercida, os reagentes, chamados de substratos, devem ligar-se à molécula da
enzima em uma região específica, chamada sítio ativo. O sítio ativo é uma cavidade com
forma definida, aberta na superfície da molécula globular da enzima, constituída por grupos R
de aminoácidos. Essa forma do sítio ativo confere especificidade à catalise enzimática: para
ser reconhecida como substrato uma molécula deve ter a forma adequada para acomodar-se
no sítio ativo e os grupos químicos capazes de estabelecer ligações com os grupos R ali
presentes (Figura 9).
Figura 9. Esquema da ligação do substrato no sítio ativo.

6.1 Modelos enzima e substrato
Emil Fischer propôs, em 1894, que as enzimas se ajustavam como o modelo “chave e
fechadura”, isto é, eram estruturalmente complementares em tamanho, forma e natureza
química a seus substratos (Figura 10).
Figura 10. Modelo chave-fechadura.
Uma enzima completamente complementar a seu substrato seria pouco eficiente. A
aproximação e a ligação do substrato à enzima altera o balanço de forças responsáveis pela
manutenção da estrutura tridimensional da enzima, amoldando sua forma à forma do substrato
e fazendo-a adquirir uma nova conformação, ideal para a catálise (Koshland, 1958, modelo do
encaixe induzido). A conformação dos substratos também é tensionada e distorcida,
aproximando-se da conformação do estado de transição (Figura 11). É esse conjunto de
mecanismos que torna a catálise enzimática tão eficiente.
Figura 11. Esquema do modelo de encaixe induzido.
7 Velocidade das reações
Uma reação enzimática simples pode ser escrita assim:
E + S ES EP E + P
E, S e P representam a enzima, o substrato e o produto. ES e EP são complexos da
molécula da enzima com o substrato e com o produto, respectivamente.
A função de um catalisador é aumentar a velocidade de uma reação, não afetando o
equilíbrio da reação. Qualquer reação pode ser descrita por um diagrama de coordenadas da
reação (Figura 12), sendo esta uma representação gráfica do curso energético da reação. A
energia nos sistemas biológicos é descrita em termos da energia livre G. Em sua forma normal
estável ou nível basal, qualquer molécula (S ou P) contém uma quantidade característica de
energia livre.
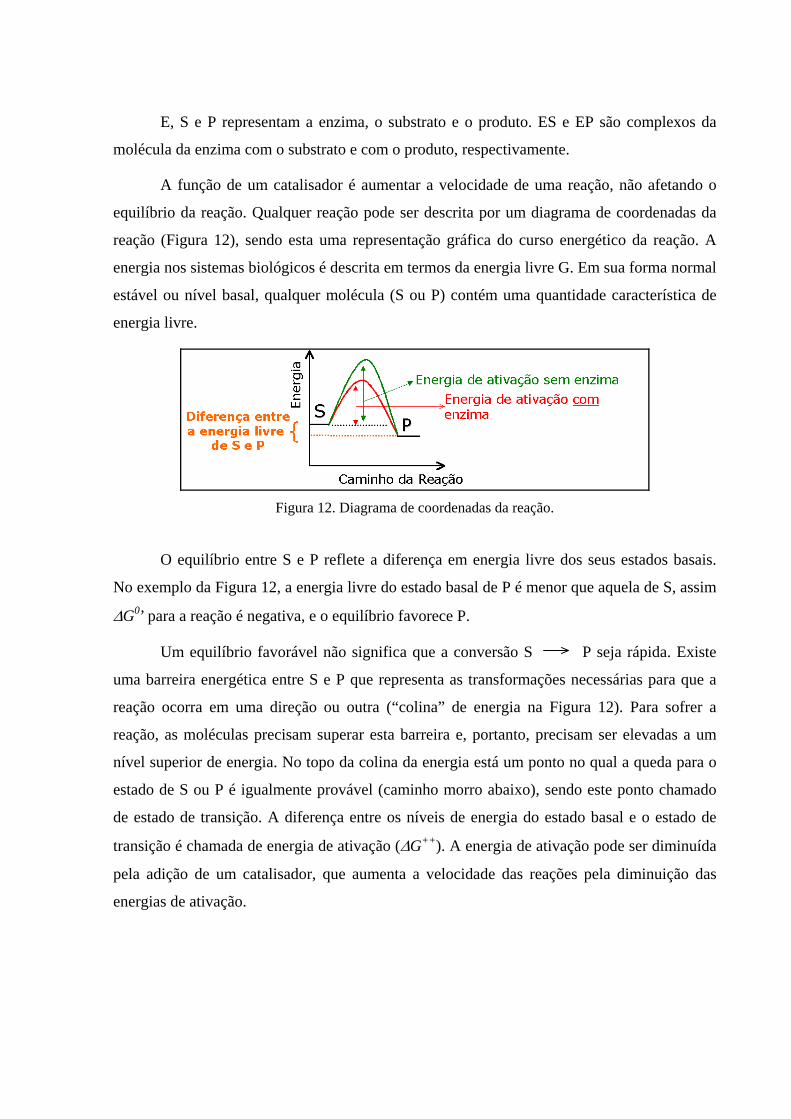
Figura 12. Diagrama de coordenadas da reação.
O equilíbrio entre S e P reflete a diferença em energia livre dos seus estados basais.
No exemplo da Figura 12, a energia livre do estado basal de P é menor que aquela de S, assim
∆G0’ para a reação é negativa, e o equilíbrio favorece P.
Um equilíbrio favorável não significa que a conversão S P seja rápida. Existe
uma barreira energética entre S e P que representa as transformações necessárias para que a
reação ocorra em uma direção ou outra (“colina” de energia na Figura 12). Para sofrer a
reação, as moléculas precisam superar esta barreira e, portanto, precisam ser elevadas a um
nível superior de energia. No topo da colina da energia está um ponto no qual a queda para o
estado de S ou P é igualmente provável (caminho morro abaixo), sendo este ponto chamado
de estado de transição. A diferença entre os níveis de energia do estado basal e o estado de
transição é chamada de energia de ativação (∆G++). A energia de ativação pode ser diminuída
pela adição de um catalisador, que aumenta a velocidade das reações pela diminuição das
energias de ativação.

8 Medida da atividade enzimática
A dosagem de enzimas é sempre feita através da medida de sua atividade, que é
avaliada pela velocidade da reação que a enzima catalisa. Para efetuar essas dosagens, uma
amostra da solução contendo a enzima é incubada com concentrações altas de substrato (para
garantir a velocidade máxima e impedir que pequenas variações na concentração do substrato
possam afetar as medidas). A velocidade da reação é medida e expressa em Unidades
Internacionais. Uma Unidade Internacional (U) é a quantidade de enzima capaz de formar 1
µmol de produto por minuto em condições ótimas de medida (pH, temperatura, etc.),
especificadas para cada caso.