isotype profiles induced in balb/c mice during foot and mouth disease (fmd) virus infection or...
TRANSCRIPT

0264-410X(95)00078-6
Vaccine. Vol. 13, No. 10, pp. 953-960, 1995 Copyright 0 1995 Elsevier Science Ltd
Printed in Great Britain. All rights reserved 0264-410X/95 $lO+O.OO
Isotype profiles induced in Balbk mice during foot and mouth disease (FMD) virus infection or immunization with different FMD vaccine formulations
D. M. PCrez Filgueira *, A. Berinsteinj-, E. Smitsaart*, M. V. Borca? and A. M. Sadir*$
The IgG iso type response in Balblc mice infected with FMD V or immunized with dtflerent vaccine formulations using inactivated virus particles as antigen was analyzed at various times post-inoculation. For this purpose an ELISA based on polyclonal antibodies for detection and quanttfication of mouse IgG isotypes with FMD virus (FMDV) specificity was developed. Three immunomodulators, which have been shown to be very eflective in inducing strong and long-lasting antibody responses (Bahnemann, Arch. Virol. 1975, 47, 47-56; Polatnik and Bachrach, Appl. Microbial. 1964, 12, 368-376), were employed to formulate dtyerent vaccines using aqueous and oil vehicles: a water-soluble fraction of the cell wall of Mycobacterium sp., a purtjied extract of lipopolysacharide from Brucella ovis and a synthetic lipoamide, Avridine. Infected animals between 14 and 60 days post- inoculation (d.p.i.) showed responses dominated by IgG2b, followed by IgG1, IgG2a and IgG3, respectively. The IgG3 isotype was thefirst, together with IgGl, to be elicited during the first 7 days after infection, whereas no IgG3 activity was detected in vaccinated animals at any time. With formulations including immunomodulators, persisting high levels of IgG2b (similar to those of infected animals) were detected until 180 d.p.i., while with conventional vaccines IgG2b responses were detected up to 60 d.p.i. Animals vaccinated with formulations including these immunomodulators presented an augmented resistance to viral challenge at 210 d.p.i. in relation with those immunized with conventional vaccines. The possible relationship sf these d@erences in the isotype response and protection is discussed.
Keywords: Foot and mouth disease; mouse isotypes; immunomodulators
Foot and mouth disease (FMD) is a widespread infec- tious disease affecting cloven-hoofed animals with severe economic consequences’. Animals infected with FMD virus (FMDV) develop a state of immunity characterized by high titers of virus serotype-specific neutralizing anti- bodies (NAb) which persist for at least 18 months’. In contrast to infection, current inactivated virus vaccines elicit lower antibody responses for shorter periodss. Pro- tection against FMDV infection has been commonly related to the level of NAb in serum+‘. However, in some cases this correlation is not clearly demonstrable, perhaps indicating the existence of other antibody-
*Institute de Virologia, Centro de lnvestigaciones en Ciencias Veterinarias, INTA-Castelar, CC77, Moron, Pcia, Buenos Aires, Argentina. tPresent address: Plum Island Animal Disease Center, USDA, Greeport, NY 11944, USA. fAuthor to whom correspondence should be addressed. (Received 20 April 1994; revised 8 November 1994; accepted 18 November 1994)
mediated mechanisms besides antibody neutralization. A recent review’ suggests that phagocytic processes are involved in virus clearance and that phagocytosis is enhanced when the virus is complexed with antibodies of the appropriate specificity and affinity.
Adult mice are not susceptible to natural infection, but may be experimentally infected’. As in the natural host, infection elicits high levels of circulating NAb which persist life-long . lo In this model FMD vaccines prepared with inactivated virus and adjuvants also induce poorer titers of antibodies which persist for less time than those induced by live virus1’*12. These differences may reflect not only quantitative but also qualitative aspects related to the antibody response, such as affinity maturation and immunoglobulin isotype profile.
Mouse serum IgG comprises four isotypes IgGl, IgG2a, IgG2b and IgG3 13,14 Murine IgG subclasses _ differ in their physical and biochemical characteristics, as well as in their biological properties which are com- monly related to the Fc region of the molecules. Some of
Vaccine 1995 Volume 13 Number 10 953

lsotype profiles induced in mice: D. M. Pkrez Filgueira et al.
these subclasses are able to activate the complement system through the Fc portion when bound to antigen15, and receptors for Fc of some IgG isotypes are present on different cells including lymphocytes, macrophages, platelets and neutrophils mediating different pathways of the cellular immune response16. It is also known that patterns of isotype in an immune response are usually restricted by the nature of the antigen for which they are specific”.18 and by the form in which the antigens are presented to the immune system”. Study of the IgG isotypes elicited in response to viral infection or immu- nization is therefore important when considering the mechanisms of in vivo protection.
In this report we evaluated the immune response elicited by different immunizations against FMDV in mice. Using an ELISA specifically developed for this purpose, we determined the isotype profiles induced in each case and we discussed the immunological pathway that operates depending on the immunization protocol and the level of protection induced. It is possible that similar mechanisms were required to obtain an effective immune response in the natural host. Importantly, re- sults obtained in our laboratory indicate that sero- neutralizing B-cell epitopes located in discrete regions of VP1 of FMDV type 0, are recognized by both mice and cattle immune sera2@22. This study will help in the design of new vaccines for the natural host that would elicit the production of antibodies of the desired isotype and biological function.
MATERIALS AND METHODS
Mice
Sixty to 90 day-old male and newborn Balb/c mice from our colony were used thoroughly in the exper- iments described below.
virus FMDV serotype 01 Campos (OlC) was used to infect
adult mice by the intraperitoneal (i.p.) route with lo4 suckling mouse 50% lethal doses (SMLD 50%) per mouse, diluted in 0.5 ml of phosphate-buffered saline (PBS) pH 7.49.
Purified whole virus particles (146s) used as antigen in vaccines were prepared from tissue culture supernatants obtained by one passage of our virus stock in BHK 21 monolayers and then inactivated with binary ethyl- enimine (BEI) according to Bahnemann23. Inactivated virus was purified in a 15-45% sucrose gradient’“, and concentration was measured at 259 nm.
Vaccine formulations
Vaccines were prepared, all containing 0.1 pg of inactivated virus per dose diluted in PBS (pH 7.4). Five different formulations were employed to immu- nize mice: (1) a water-in-oil emulsion according to the INTA formulation25 > ‘2] a conventional aluminum hy- droxide (2.5%) vaccine 6, (3) a water-in-oil emulsion containing Avridine ((N,N dioctadecyl-JV’,N’-bis) (2- hydroxyethyl) propanediamine) (Pfizer, Inc., USA$, added to the oil phase at a dose of 1 mg per mouse2 , and two aluminum hydroxide (2.5%) formulations in- cluding (4) 20 pg per dose of lipopolysacharide of
Bpucellu ovis (LPS)“, or (5) a water-soluble fraction (WSF) (10 pug per dose) of the cell wall of Myco- bacterium sp. added as described previouslyZ6.
Serology
Infected animals were bled at 3, 7, 14, 21, 45 and 60 days post-immunization (d.p.i.). At each time groups of three mice were used and individual serum samples were pooled and stored at - 20°C until processed. In a similar way, groups of mice inoculated with conven- tional water-in-oil and aluminum hydroxide vaccines were bled at the same intervals plus an additional time at 180 d.p.i, while sera from animals immunized with vaccines containing Avridine, WSF or LPS were taken only at 60 and 180 d.p.i.
Sera from 20 normal male mice were pooled and used as a negative control to measure the background activity in all the experiments.
Pooled sera were titrated for FMDV Nab using suckling mice as described by Cunha et al. (1957)2s and their titres expressed as seroneutralizing indexes (SNI).
Quantification of mouse Ig isotypes by ELISA
ELISA plates (Nunc Immunoplate 1 Maxisorp, Denmark) were sensitized overnight at 4°C with rabbit anti-FMDV 01 C antiserum (capture serum) diluted 1:300 in carbonate-bicarbonate buffer pH 9.629. The following steps were performed using PBST/S% horse normal serum (HNS) as dilution buffer with incubations at 37°C for 1 h in constant agitation, except for the substrate step. The HNS was selected among diverse blocking substances, which were assayed to prevent non-specific reactions. Coated plates were then consecu- tively incubated with virus (1:5 dilution of a lo*.’ SMLD 5O%/ml tissue culture supernatant), sample mouse serum (in threefold dilution series), goat anti-mouse IgG sub- classes (IgGl, IgG2a, IgG2b or IgG3) serum (Sigma, USA) and rabbit anti-goat peroxidase conjugate 1:400 (Cappel, Swiss). O-phenylenediamine (0PD)/H20, was used as peroxidase substrate diluted in citrate- phosphate buffer (pH 4.5). Substrate reaction was stopped after 10 min by addition of 2 M H,SO, and the absorbance read at 490 nm in a Dynatech MR5000 Microplate Reader (Dynatech Laboratories, UK).
Isotypes in normal Balb/c mouse serum are not rep- resented in equal concentrations’4.30. This fact could modify final titres by competition between isotypes, underestimatin those subclasses presented at lower concentrations J? . Capture serum and virus dilutions were selected in order to obtain an excess of captured antigen to reduce the effect of any competition (data not shown). If competition occurs it should be reduced at high test serum dilutions, so that different isotype pro- files should be obtained when isotype titres are calcu- lated at different end-point sample serum dilutions. We found no significant differences when comparing profiles obtained using different end-point dilutions, indicating that competition under our conditions was negligible.
The specificity of goat anti-mouse IgG subclass reagents was assessed by substituting sample mouse serum, by one of four monoclonal antibodies (mAb) against FMDV serotype 01, each one with a different
954 Vaccine 1995 Volume 13 Number 10

Table 1 Description of the mAbs used to establish specificity of anti-isotype reagents
mAb Virus specificity lsotype
12FE9.2.1 FMDV 01 Brugge IgGl 1 OGA4.2.2 FMDV 01 Brugge lgG3 4c9 FMDV 01 Suisse IgG2a lC6 FMDV 01 Suisse IgG2b
isotype. Such mAbs were used throughout as controls for specificity.
Monoclonal antibodies MAbs used to test specificity are described in Table 1
and were kindly provided by Dr D. 0. Morgan from Plum Island Animal Disease Center, USDA, USA (12FE9.2.1 and lOGA4.2.2; ref. 32) and by Dr J. R. Crowther from Pirbright Laboratories, AFRC, UK (4C9 and lC6; ref. 33). All these mAbs showed high neutralizing activity against FMDV 01 Campos in mice (data not shown)
RESULTS
Specificity of anti-mouse IgG isotypes reagents Different dilutions of each anti-mouse isotype serum
were tested in order to obtain the maximum specificity for the homologous isotype and the lower cross- reactivity with the others (Figure 1). Optimal dilutions were l/2000 for goat anti-IgGl, -IgG2a, -1gG3 and 113000 for anti-IgG2b. In all cases anti-isotype sera were 3-7 times more specific for the corresponding isotype than for the heterologous isotypes.
lsotype profiles induced in mice: D. M. Phrez Filgueira et al.
Antibody concentration of each anti-IgG isotype re- agent as well as its affinity for the corresponding sub- class differed, so it was necessary to correct for these different sensitivities before comparing each IgG isotype titre in the immune sera. As is shown in Figure 2, in order to establish the dilution that optimized the speci- ficity of each anti-isotype reagent, mAbs were used in excess in all cases (diluted 1:5). It was assumed that initial amounts of mAbs bound to the plates were similar for all the isotypes. Thus, differences in maximal optical density (O.D.) obtained for each anti-isotype serum with the mAb of the homologous isotype, re- flected potency differences among the anti-IgG isotype reagents (Figure 2). Correction factors were used (Table 2) to standardize the O.D. so that they could be con- verted to an isotype titre in the four systems, then obtaining a more accurate relative titre, as described by McKendall and WOOED.
IgG subclass responses in infected animals Isotypic profiles and SNI obtained at 3, 7, 14, 21, 45
and 60 d.p.i. for infected mice are presented in Figure 3. No anti-FMDV IgG subclasses were detected until 7 d.p.i., when IgGl and IgG3 activities were clearly de- tected. Two weeks post-infection the other isotypes made their appearance, while IgGl and IgG3 showed elevated levels. This pattern registered at this time was maintained up to 60 d.p.i., with predominant levels of IgG2b, followed by IgGl, and then IgG2a and IgG3.
IgG subclass responses in animals vaccinated with conventional vaccines
In the animals immunized with the aluminum hydrox- ide vaccine, IgGl was the predominant isotype showing
0.01 I I I I I I 0.0 I I I I I I 2.0 2.5 3.0 3.5 4.0 4.5 5.0 2.0 2.5 3.0 3.5 4.0 4.5 5.0
Anti-IgGI log reciprocal dilution Anti-IgG2a log reciprocal dilution
P-J 0.8r
t
l -�-,_, E 0.6 E \
2.5 3.0 3.5 4.0 4.5 5.0
I.O-
0.8 -
1 2.0 2.5 3.0 3.5 4.0 4.5 5.0
Anti-IgG2b log reciprocal dilution Anti-IgG3 log reciprocal dilution
Figure 1 ELISA to determine the specificity of goat anti-mouse IgG isotypes reagents. Captured virus was incubated with normal mouse serum (-) diluted l:lO, or with sets of mAbs (diluted 1:5) of IgGl (O), lgG2a (W), lgG2b (a), or lgG3 (A) isotypes. Various dilutions of goat anti-IgGl @&g-llF2a (b), -IgG2b (c), or -lgG3 (d) were then added to each set of mAbs. Arrows indicate optimal working dilution for each anti-isotype
Vaccine 1995 Volume 13 Number 10 955

lsolype profiles induced in mice: D. M. Pkrez Filgueira et al.
(4
0.8$- Anti-IgGl
0.6 -
mAb log reciprocal dilution
Anti-IgG2b
I I I 1.5 2.5 3.5
mAb log reciprocal dilution
(b)
0.8r Anti-IgG2a
0.6
0.6
0.4
I I I
I .5 2.5 3.5
mAb log reciprocal dilution
(4
:i,_“\* 0.0 I I I I
0.5 I.5 2.5 3.5 4.5
mAb log reciprocal dilution
Figure 2 Activity of anti-isotypes reagents against different dilutions of the set of mAb. Optimal dilution of each anti-IgG subclass sera (1:2000 for anti-IgGl (a), anti-lgG2a (b) and anti-lgG3 (d); and 1:3000 for anti-lgG2b (c)); were added to mAbs with IgGl ( q ), IgG2a ( n ), IgG2b (+) and lgG3 (A) isotype in the same system as described for Figure 7
Table 2 Correction factors were calculated as the ratio between absorbance obtained for anti-lgG3 (at optimal dilution) with its homologous mAb (lOGA4.2.2) diluted 15; and the ones obtained for anti-IgGl, -IgG2a or -IgG2b with their homologous mAbs (12FE9.2.1, 4C9 and lC6, respectively) in the same conditions. Apparent final dilutions for IgGl, IgG2a and IgG2b were adjusted by the respective factors to compensate their lower potency respect to anti-lgG3 serum
IgG subclass
IgGl IgG2a IgG2b lgG3
A492 nm
0.724 0.403 0.676 1.183
Correction factor
1.64 2.93 1.75 1 .oo
its highest titre by 60 d.p.i. IgG2a activity was tran- siently detected at 45 and 60 d.p.i. at lower concen- trations than for IgGl. The IgG2b isotype was the less abundant of all isotypes, and was only detected at 60 d.p.i. No IgG3 was detected during all the experimental periods (Figure 40).
Sixty days p.i. the WSF-containing vaccine induced predominant levels of IgGl followed by IgG2b and IgG2a in concentrations three- and eightfold lower than the ones for IgG1. respectively (Figure 5b). On the other hand, LPS generated an IgG2a level two- and sixfold higher than the IgGl and IgG2b ones, respectively (Figure 5~). Vaccines including WSF or LPS induced at 180 d.p.i. almost the same isotypic profile with produc- tion of IgG2b and IgG2a, isotypes which were not present in the conventional aqueous formulation at that time (Figure 5~).
On the other hand, animals immunized with the The water-in-oil vaccine including Avridine clearly water-in-oil formulation presented IgG2a as the pre- produced dominant titres of IgGl, followed by IgG2a dominant isotype from 21 d.p.i. IgGl was detected 45 and IgG2b at 60 d.p.i. (Figure 6b). On the other hand d.p.i., but at twofold lower concentration than those of conventional water-in-oil vaccine generated higher levels IgG2a. Sixty d.p.i. IgGl titres presented an increment of IgG2a followed by IgGl and IgG2b by that time as described above for the aluminum hydroxide vac- (Figure 6~). Six months p.i. IgG2a was still the predomi- cine, and almost equaled IgG2a titres. At 180 d.p.i. nant isotype in the sera of the animals immunized with IgG2a and IgGl titres were still similar but lower than the conventional formulation followed by IgGl, but at 60 d.p.i. As in the case of the animals immunized IgG2b could not be detected. By the same time, the with the aqueous vaccine, IgG2b was only detected at experimental formulation generated dominant titres of 60 d.p.i. and there was no IgG3 activity at any time IgG2b and IgGl, with production of IgG2a but to a (Figure 4b). lesser extent. Like with the conventional formulations
For both formulations those isotypes presented at higher concentrations were also the first to be elicited.
IgG isotype responses to vaccines containing immunomodulators
The pattern of activity of IgG isotypes in animals vaccinated with formulations including Avridine, WSF or LPS was established at 60 and 180 d.p.i. and com- pared with the one obtained with the same vehicles without these immunomodulators.
956 Vaccine 1995 Volume 13 Number 10

lsotype profiles induced in mice: D. M. P&ez Filgueira et al.
80-
70-
13 M 60-
z 2 50-
: 40-
30 -
0 3 7 14 21 45 60
d.p.i.
Figure 3 SNI and isotype profiles determined in infected animals. Results are expressed as percentage of the addition of the titres obtained for each isotype. Titres for each subclass were defined as the reciprocal of the corrected final dilution. The final dilution for each isotype was defined as the highest serum dilution that produced at least an absorbance twice as high as the normal mice serum diluted 1 :lO. Final titres in all cases were the mean of two independent assays performed in duplicate
(4 Aluminium hydroxide formation
90
80
0, 70
; 60
g 50
EF 40
21 45 60 180
(b)
100
90
80
‘3, 70
; 60
s k
50
* 40
30
20
I 0
0
d.p.i.
Water in oil formation
21 45 60 d.p.i.
Figure 4 SNI and isotype profiles in animals immunized with conventional vaccines. lsotypes in mice vaccinated with aluminum (a) or water in oil (b) formulations were detected since day 21 p.i. Results are expressed as in Figure 3
no IgG3 activity could be detected in the animals immunized with any experimental formulation during all the experimental period.
DISCUSSION
Many studies have been conducted to determine the IgG isotype response induced in mice immunized with viral
antigens or infected with active virus3438 but to our knowledge none of them were done in the FMDV system.
Coutelier et al. 36,39.40 reported that viral infections generally induced antibodies of the IgG2a, and to a lesser extent of the IgG2b isotypes. Such restriction was demonstrated for a wide range of RNA and DNA viruses, and was independent of the route of inoculation or mouse strain. However, patterns others than this were
Vaccine 1995 Volume 13 Number 10 957
lsotype profiles induced in mice: D. M. Pbrez Filgueira et al.
“‘1 (a)
30
20
IO-
O- 60 60 180 60 180
~ (b) WSNI
I EiIgGl
I
I 0 IgC2a
I
d.p.i.
Aluminum WSF LPS
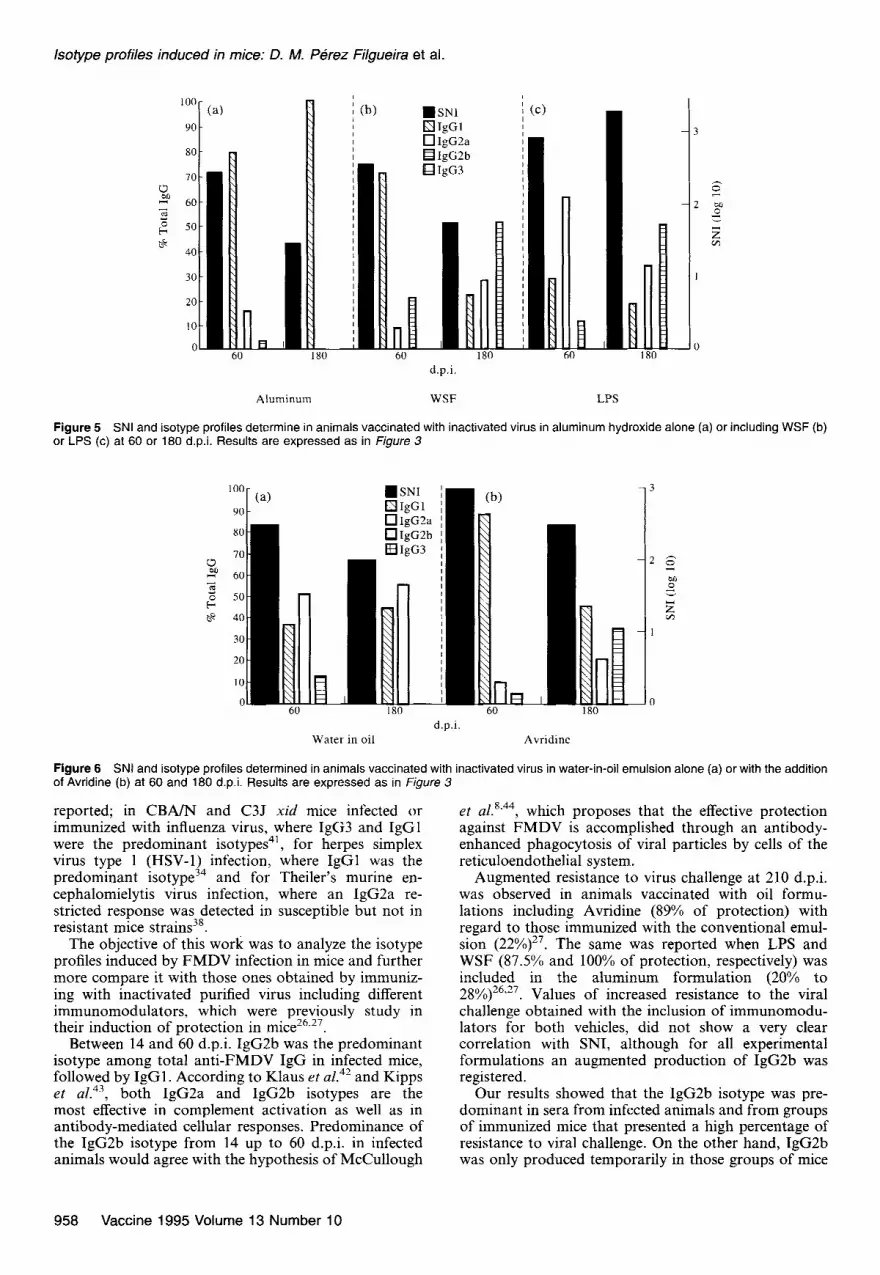
Figure 5 SNI and isotype profiles determine in animals vaccinated with inactivated virus in aluminum hydroxide alone (a) or including WSF (b) or LPS (c) at 60 or 180 d.p.i. Results are expressed as in figure 3
80
70 0 on 3 60
z?
g 50
EF 40
0 60 180
Water in oil
(b)
d.p.i.
60 180
Avridine
Figure 6 SNI and isotype profiles determined in animals vaccinated with inactivated virus in water-in-oil emulsion alone (a) or with the addition of Avridine (b) at 60 and 180 d.p.i. Results are expressed as in Figure 3
reported; in CBA/N and C3J xid mice infected or immunized with influenza virus, where IgG3 and IgGl were the predominant isotypes4’, for herpes simplex virus type 1 (HSV-1) infection, where IgGl was the predominant isotype34 and for Theiler’s murine en- cephalomielytis virus infection, where an IgG2a re- stricted response was detected in susceptible but not in resistant mice strains3*.
The objective of this work was to analyze the isotype profiles induced by FMDV infection in mice and further more compare it with those ones obtained by immuniz- ing with inactivated purified virus including different immunomodulators, which were previously study in their induction of protection in mice’6,27.
Between 14 and 60 d.p.i. IgG2b was the predominant isotype among total anti-FMDV IgG in infected mice, followed by IgGl . According to Klaus et aL4’ and Kipps et a1.43, both IgG2a and IgG2b isotypes are the most effective in complement activation as well as in antibody-mediated cellular responses. Predominance of the IgG2b isotype from 14 up to 60 d.p.i. in infected animals would agree with the hypothesis of McCullough
et a1.8.44, which proposes that the effective protection against FMDV is accomplished through an antibody- enhanced phagocytosis of viral particles by cells of the reticuloendothelial system.
Augmented resistance to virus challenge at 210 d.p.i. was observed in animals vaccinated with oil formu- lations including Avridine (89% of protection) with regard to those immunized with the conventional emul- sion (22%) 27 The same was reported when LPS and . WSF (87.5% and 100% of protection, respectively) was included in the aluminum formulation (20% to 28%)26.17. Values of increased resistance to the viral challenge obtained with the inclusion of immunomodu- lators for both vehicles, did not show a very clear correlation with SNI, although for all experimental formulations an augmented production of IgG2b was registered.
Our results showed that the IgG2b isotype was pre- dominant in sera from infected animals and from groups of immunized mice that presented a high percentage of resistance to viral challenge. On the other hand, IgG2b was only produced temporarily in those groups of mice
958 Vaccine 1995 Volume 13 Number 10

lsoiype profiles induced in mice: D. M. Pbrez Filgueira et al.
immunized with conventional formulations that showed poor protective response to the viral challenge. Al- though further experiments are required to confirm the efficiency of this particular isotype in protection against FMDV in mice, these results indicate that the isotype profile elicited, besides the level of NAb, could be important when considering the efficacy of a vaccine formulation.
Infected animals also differ in relation to the vacci- nated ones in the presence of IgG3 activity. The IgG3 subclass presents the tendency to form aggregates by non-covalent bonds between the Fc region of other adjacent IgG3 molecules and fixes complement poorly . 45.14 Baxt er ~1.~~ found that all mAbs that caused FMDV aggregation were of the IgG3 subclass. These authors propose viral aggregation as a possible mech- anism of viral neutralization and suggest that it might be isotype-related. Mouse IgG3 production seems to be supported by an absence of T-cell-derived limphokines4’ that direct immunoglobulin expression to other sub- classes; then, it could be reflecting a failure to stimulate T cells capable of secreting limphokines. In viva exper- iments showed that transferred B cells from mice in- fected 8 days before were capable of abortin viraemia in irradiated animals, in the absence of T cells 7. Despite the low levels found for this isotype (that only represents 5% of the total IgG isotypes in normal mice sera) in relation with the others, it is noteworthy that it was the first isotype to be detected together with IgGl at 7 d.p.i. Taking together, these results might reflect a possible role of the IgG3 during the first steps of the immune response in infected animals or at least indicate that within these first days it is carried out with a low participation of the T cell system. In any case this restriction of IgG3 production obtained during FMDV infection again would indicate that the immune response is carried out in a different way in infected and vaccinated animals.
We conclude that differences in isotype profiles pre- sented here between virus-infected and vaccinated mice might indicate dissimilar conditions of T-cell priming. Further studies are required to clarify the relation be- tween the T-cell system stimulation, isotype of antibody induced and protection against FMDV.
ACKNOWLEDGEMENTS
We gratefully ackonwledge Dr Anthony J. Forman for critical review of the manuscript. These authors would also like to thank to Mr Antonio Varone and Mr Salvador Herrera for their excellent technical help.
REFERENCES
Cottral, G. Foot and mouth disease virus. In: Foreign Animal Disease, 3rd ed. Committee on Foreign Animal Diseases, United States Animal Health Association, Richmond, VA, 1975
Cunliffe, HR. Observations on the duration of immunity in cattle after experimental infection with foot and mouth disease virus. Cornell Vet. 1964, 54, 501-510 Morgan, D.O., Moore, D. and McKercher, P. Vaccination against foot and mouth disease. In: New Developments with Human and Veterinary Vaccines (eds Mizsaht, A., Hertman, I., Klingberg, M.A. and Kohn, A.). Alan A. Liss, New York, 1980, pp. 169-178
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
Van Bekkum, J.G. Correlation between serum antibody levels and protection against challenge with FMDV. Session of the Research Group of the Standing Technical Committee of the European Commission for the Control of FMD, Brescia, Italy. Food and Agriculture Organization, Rome, 1969, pp. 38-41 Knudsen, R., Groocock, C. and Andersen, A. Immunity to foot and mouth disease virus in guinea pigs: clinical and immune response. Infect. Immun. 1979, 24, 787-792 Sutmoller, P. and Vieira, A. The relationship of neutralizing antibody titres for FMDV and the protection of cattle. Bol. Cent. Panam. Fiebre Aftosa 1980, 39, 57-62 Pay, T.W.F., Hingley, P.H., Radlett, P.J., Black, L. and 0’ Reilly, K.J. The correlation of the 140s antigen dose with the serum neutralizing antibody response and with protection from challenge induced by FMD vaccine. Session of the Research Group of the Standing Technical Committee of the European Commission for the Control of FMD, Leystad, The Netherlands. Food and Agriculture Organization, Rome, 1983, pp. 52-55 McCullough, KC., De Simone, F., Brocchi, E., Capucci, L., Crowther, J.R. and Kihm, U. Protective immune response against foot-and-mouth disease. J. Viral. 1992, 66, 1835-l 840 Fernandez, F.F., Borca, M.V., Sadir, A.M., Fondevila, N., Mayo, J. and Schudel, A.A. Foot and mouth disease virus (FMDV) experimental infection: susceptibility and immune response of adult mice. Vet. Microbial. 1986, 12, 15-24 Borca, M.V., FernBndez, F.F., Sadir, A.M. and Schudel, A.A. Reconstitution of immunosuppressed mice with mononuclear cells from donors sensitized to foot and mouth disease virus (FMDV). Vet. Microbial. 1984, 10, l-11 Lopez, O.J., Borca, M.V., Fernandez, F.F., Braun, M. and Schudel, A.A. Immune response to foot and mouth disease virus in an experimental murine model. II. Basis of persistent antibody reaction. Vet Immunol. Immunopathol. 1990, 24, 313-321 Piatti, P., Berinstein, A., Lopez, 0. J, Borca, M.V., Fernandez, F.F., Schudel, A.A., and Sadir, A.M. Comparison of the immune response elicited by infectious and inactivated foot and mouth disease virus in mice. J. Gen. Viral. 1991, 72, 1691-1694 Fahey, J., Wunderlich, J. and Michell, R. The immunoglobulins of mice: 7S gamma 2,7S gamma 1, gamma 1 A (&!A), and 18s gamma 1 M-globulins. J. Exp. Med. 1964, 120, 223-242 Grey, H.M., Hirst, J.W. and Cohn, M. A new mouse immuno- globulin: lgG3. J. fxp. Med. 1971, 133, 289-304 Dorrington, K. The structural basis for the functional versa- tility of the immunoglobulin G. Can. J. Biochem. 1978, 56, 1087-1101 Burton, 0. lmmunoglobulin G, functional sites. Molec. lmmunol. 1985, 22, 161-206 Perlmutter, R. M., Hansburg, D., Briles, D.E., Nicolotti, D.A. and Davie, J. Subclass restriction of murine antibodies. J. Immun. 1978, 72, 566-572 Almond, N.M. and Parkhouse, R.M. lmmunoglobulin class specific responses to biochemically defined antigens of Tri- chine/la spiralis. Parasite Immunol. 1986, 6, 391-406 Balcovic, E.S., Florack, J.A. and Six, H.R. lmmunoglobulin G subclass responses of mice to influenza virus antigens given in different forms. Antiviral Res. 1987, 6: 151-160 Garmendia, A.E., Borca, M.V., Morgan, D.O. and Baxt, B. Analysis of the foot-and-mouth disease virus neutralizing idio- types from immune bovine and swine with anti-murine idiotype antibody probes. J. Immun. 1989, 143, 3015-3019 Borca, M.V., Garmendia, A.E., Baxt, B., Moore, D., Srikumaran, S. and Morgan, D.O. Cross-reactive idiotopes among anti foot-and-mouth disease virus neutralizing antibodies. Immunol- ogy 1993,79,368-374 Zamorano, P, Wigdorovitz, A., Perez Filgueira, D.M., Carrillo, C.. Sadir. A.M. and Borca. M.V. bkiDDinCl of B and T epitopes in a 26mer’synthetic peptide containing the major immunogenic site of VP1 FMDV 01 Campos. Manuscript in preparation Bahnemann, H. Binary ethylenimine as an inactivant for foot-and-mouth disease virus and its application for vaccine production. Arch. Virol. 1975, 47, 47-56 Polatnick, J. and Bachrach, H. Production and purification of milligram amounts of foot and mouth disease virus from baby hamster kidney cell cultures. Appl. Microbial. 1964, 12, 368-376 Rivenson, S., Sadir, A.M., Gaggino, O.P., Marcovecchio, F.E., Zabal, 0. and Laporte, 0. Estudio comparative en bovinos de
Vaccine 1995 Volume 13 Number 10 959

lsoiype profiles induced in mice: D. M. Perez Filgueira et al.
26
27
28
29
30
31
32
33
34
35
36
dos vacunas antiaftosa: oleosa e hidroxisaponinada. Rev. Med. (Buenos Aires) 1982, 63, 364-370 Berinstein, A., Piatti, P., Gaggino, O.P., Schudel, A.A. and Sadir, A.M. Enhancement of the immune response elicited with foot- and-mouth disease virus vaccines by an extract of the Myco- bacterium sp. wall. Vaccine 1991, 9, 883-888 Berinstein, A., Perez Filgueira, M., Schudel, A.A., Zamorano, P., Borca, M. and Sadir, A.M. Avridine and LPS from B. ovis: effect on the memory induced by FMDV vaccination. Vaccine 1993, 11,1295-1301 Cunha IX, Junior, J., Serraro, W. and Torruela, I. El uso de ratones lactantes en la evaluation de 10s anticuerpos contra el virus de la fiebre aftosa y su significancia inmunologica. Gac. Vet. 1957, 110, 243-267 Mulcahy, G., Gale, C., Robertson, P., lyisan, S., DiMarchi, R.D. and Doel, T.R. lsotype responses of infected, virus-vaccinated and peptide-vaccinated cattle to Foot-and-Mouth disease virus. Vaccine 1990, 8, 249-257 Natsuume-Sakai, S., Motonishi, K. and Migita, S. Quantitative estimation of five classes of immunoglobulin in inbred mouse strains. Immunology 1977, 32, 861-866 Djurup, R. and Weeke, B. Methods of detecting IgG subclass proteins and antibodies. In: Basic and Clinical Aspects of IgG Subclasses (Ed. Shakib, F.). S. Karger, Basel, 1986, pp. 91-110 Stave, J.W., Card, J.L., Morgan, D.O. and Vakharia, V.N. Neutralization sites of type 01 foot-and-mouth disease virus defined by monoclonal antibodies and neutralization-escape virus variants. Virology 1987, 162, 21-29 McCullough, K.C., Crowther, J.R., Butcher, R.N., Carpenter, W.C., Brocchi, E., Capucci, L. and De Simone, F. Immune protection against foot-and-mouth disease virus studied using virus neutralizing and non-neutralizing concentrations of mono- clonal antibodies. Immunology 1986, 58, 421-428 McKendall, R.R. and Woo, W. Murine IgG subclasses re- sponses to herpes simplex virus type 1 and polypeptides. J. Gen. Virol. 1988, 69, 847-857 Thomsen, A.R., Volkert, M. and Rajewsky, K. Different isotype profiles in virus-specific antibodies in acute and persistent lymphocytic choriomeningitis virus infection in mice. J. Immun. 1986, 136,2892-2895 Coutelier, J.-P., van der Logt, F., Heessen, W.A., Warnier, G. and Van Snick, J. IgGPa restriction of murine antibodies elicited by viral infections. J. Exp. Med. 1987, 165, 64-69
37
38
39
40
41
42
43
44
45
46
47
Hockart, M., Mackenzie, J. and Steward, G. The IgG subclass response induced by wild-type, cold-adapted and purified haemogglutinin from influenza virus A/Queensland/G/72 in CBAfCaH mice. J. Gen. Virol. 1988, 69, 1872-l 882 Peterson J.D., Waltenbaugh, C. and Miller, SD. IgG subclass responses to Theiler’s murine encephalomyelitis virus infection and immunization suggest a dominant role for Thl cells in susceptible mouse strains. Immunology 1992, 75, 652-658 Coutelier, J.-P., van der Logt, F., Heessen, W.A., Vink, A. and Van Snick, J. Virally induced modulation of murine IgG antibody subclasses. J. fxp. Med. 1988, 165, 2373-2380
Coutelier, J.-P., van der Logt, F. and Heessen, F.W.A. IgG subclass distribution of primary and secondary immune responses concomitant with viral infections. J. Exp. Med. 1991, 165,64-69
Reale, M.A., Bona, C.A. and Schulman, J.L. lsotype profiles of anti-influenza antibodies in mice bearing the xiddefect. J. Viral. 1985, 53, 4255429 Klaus, G.G.B., Pepys, M.B., Kitajina, K. and Asconas, B.A. Activation of mouse complement by different classes of mouse antibody. J. Immun. 1979, 38, 687-695
Kipps, T.J., Parham, P., Punt, J and Herzemberg, L.A. Impor- tance of immunoglobulin isotype in human antibody-dependent, cell-mediated cytotoxicity directed by murine monoclonal anti- bodies J. Exp. Med. 1985, 161, 1-17 McCullough, KC., Parkinson, D. and Crowther, J.R. Opsoniz- ation enhanced phagocytosis of foot-and-mouth disease virus. Immunology 1988,65, 187-191
Greenspan, N.S. and Cooper, L.J.N. Intermolecular cooper- ativity: a clue to why mice have lgG3? Immunol. Today 1992, 13, 164-168
Baxt, B., Morgan, D.O., Robertson, B.H. and Tlmpone, CA. Epitopes of foot and mouth disease virus outer capsid protein VP1 involved in neutralization and cell attachment. J. Viral. 1984, 15,289-309 Borca, M.V., Fernandez, F.F., Sadir, A.M., Braun, M. and Schudel, A.A. The immune response to foot-and-mouth dis- ease virus in a murine model: effective thymus-independent primary and secondary reaction. Immunology 1988, 59, 261-267
960 Vaccine 1995 Volume 13 Number 10
Effect of severe aortic banding above the renal arteries on nitric oxide synthase isotype expression
Isotype-specific inhibition of the phosphatidylinositol-3-kinase pathway in hematologic malignancies