is automatic imitation based on goal coding or movement coding?
TRANSCRIPT

Research Article
Is Automatic Imitation Based onGoal Coding or Movement Coding?
A Comparison of Goal-Directed and Goal-Less Actions
Claudia Chiavarino, Stefano Bugiani, Elisa Grandi, and Livia Colle
Center for Cognitive Science, Department of Psychology, University of Turin, Italy
Abstract. A key issue for research on automatic imitation is whether it occurs primarily at the level of movements, that is, by automaticallyactivating a representation of the movement/effector involved in the execution of the observed action, or at the level of goals, that is, bytriggering a representation of the action goal, irrespective of how the motor act is physically instantiated. The present study presents twoexperiments aimed at investigating the contribution of movement coding and goal coding to automatic imitation, by assessing participants’performance in a spatial compatibility task where the observed stimuli were goal-directed and goal-less actions, which have been demonstratedto elicit, respectively, goal and movement coding. We found a significant automatic imitation effect both when the stimuli were goal-lessactions and when they were actions directed toward a goal. However, the effect was stronger for the goal-less actions, even after controlling forsaliency effects. These results suggest that goal coding contributes to automatic imitation, but to a lesser degree compared to movement coding.The implications of these results for theory and research on automatic imitation are discussed.
Keywords: automatic imitation, visuomotor priming, movement coding, goal coding, spatial compatibility
Automatic imitation, that is, the involuntary tendency toreproduce the actions we observe, has been receivingincreasing attention in psychological research, especiallyafter the discovery, in the human brain, of areas containingmirror neurons, which discharge both during execution ofan action and during observation of another agent perform-ing the same action (Chong, Cunnington, Williams,Kanwisher, & Mattingley, 2008; Kilner, Neal, Weiskopf,Friston, & Frith, 2009; Mukamel, Ekstrom, Kaplan,Iacoboni, & Fried, 2010). A number of studies have relatedautomatic imitation not only to our tendency to involun-tarily reproduce actions, postures, and expressions of thepeople we interact with, but also to our capacity tointentionally imitate movements and goals of other agents(Ferrari, Bonini, & Fogassi, 2009; Iacoboni, 2005).
A key issue for research on automatic imitation iswhether it occurs primarily at the level of movements orat the level of goals. The notion of ‘‘goal’’ has been usedto indicate: (i) generally, the accomplishment of an out-come, whether it is grasping an object, reaching for a loca-tion in space, or achieving the end position of a movement(Liepelt, Von Cramon, & Brass, 2008; Longo, Kosobud, &Bertenthal, 2008); (ii) more specifically, a physical, visibletarget, object, or goal toward which an action is directed(Bouquet, Shipley, Capa, & Marshall, 2011; Wild,Poliakoff, Jerrison, & Gowen, 2010; Wohlschl�ger &Bekkering, 2002). In the present paper, we will refer to thissecond, more definite, meaning of ‘‘goal.’’ In this sense,pressing a button or touching a dot drawn on a table aregoal-directed actions, because they are specifically directed
toward a physical object, while tapping on the surface of atable where no target for the movement is visible consti-tutes a goal-less action.
Studies on the mirror effects elicited by passive actionobservation suggest that our representation of actions ishierarchically organized. Specifically, a low-level modalityis concerned with describing the physical features of theobserved scene, such as the muscular pattern and the effec-tor involved in the execution of the observed action (move-ment coding), and is primarily activated when we observeactions devoid of a goal. A higher-level modality is insteadresponsible for transforming the perceptual features of theobserved scene into an abstract coding of the motor act,independent of physical instantiation (goal coding), and ismostly triggered by observation of actions directed towarda goal (Cattaneo, Caruana, Jezzini, & Rizzolatti, 2009;Cattaneo, Sandrini, & Schwarzbach, 2010; van Elk, vanSchie, & Bekkering, 2008).
A similar distinction between movement and goal cod-ing is found in the literature on intentional imitation to thegoal-directed theory of imitation (GOADI; Bekkering,Wohlschl�ger, & Gattis, 2000; Wohlschl�ger, Gattis, &Bekkering, 2003), when required to imitate observedactions, people deconstruct them into a hierarchy of compo-nent features (e.g., effector, movement path, end-state, etc.)and then use this representation as the basis for reproducingthe action. When an object is present on the scene, as in thecase of goal-directed actions, it generally occupies the high-est position in the hierarchy, and as such it is reproducedfaster and more accurately. Instead, when people imitate
� 2013 Hogrefe Publishing Experimental Psychology 2013DOI: 10.1027/1618-3169/a000190
Author’s personal copy (e-offprint)

actions not directed toward a goal, the action feature thatbecomes prioritized for processing resources is usuallythe effector used by the actor (Chiavarino, Apperly, &Humphreys, 2008; Gleissner, Meltzoff, & Bekkering,2000; Wild et al., 2010; Wohlschl�ger & Bekkering,2002; Wohlschl�ger et al., 2003). However, different typesof saliency effects can influence the hierarchical organiza-tion of the component features of an action during imita-tion. For example, when, within an object-directed action,the saliency of the effector is increased (e.g., with a colorcue), reproduction of the effector becomes more accuratethan reproduction of the object (Bird, Brindley, Leighton,& Heyes, 2007).
A valuable insight into the hierarchical organization ofaction features is also offered by the spatial compatibilityliterature. For instance, the model of Heister, Schroeder-Heister, and Ehrenstein (1990) distinguishes three S-Rcompatibility factors: (1) spatial coding of response keys,(2) spatial coding of effector position, and (3) spatio-ana-tomical mapping. These factors jointly contribute to perfor-mance, with the effect contributed by each factor weightedaccording to its rank order. When stimuli and responses areconstituted by biological movements and there is no S-Ranatomical correspondence (i.e., when the third factor doesnot occur), RTs tend to be longer. Also, because the ana-tomical distinction is the lowest contributor in the hierar-chy, other factors that influence the reference frame forcoding the spatial location of the response keys or of theeffector (such as saliency effects) may reduce the contribu-tion of the anatomical distinction.
In the research on automatic imitation, the contributionof movement coding has been thoroughly documented by awide number of studies focusing on actions devoid of agoal, such as lifting/tapping finger movements, oropening/closing hand movements (Bertenthal, Longo, &Kosobud, 2006; Brass, Bekkering, Wohlschl�ger, & Prinz,2000; Catmur & Heyes, 2011; Leighton & Heyes, 2010;Press, Bird, Walsh, & Heyes, 2008). However, to date,there is no conclusive evidence on the role of goal codingin automatic imitation, because the few studies that havedirectly compared goal-less and goal-directed actions eitherhave not used closely matched stimuli (Liepelt, Prinz, &Brass, 2010), or they have not differentiated the effect ofautomatic imitation from similar effects, such as spatialcompatibility (Bouquet et al., 2011), that is, the facilitationeffect for stimuli and responses that occur on thesame left-right or up-down spatial alignment (Hommel &Prinz, 1997).
The aim of the present work was to investigate the con-tribution of goal and movement coding to automatic imita-tion. To this end, we showed participants identical fingertapping actions, but in one case movements were goal-directed, that is, they ended on the keys of a responsebox, while in the other case movements were goal-less, thatis, they ended on a flat surface. Based on the previouslyreviewed literature, we assumed that goal-directed actionswould primarily elicit a coding based on the goal of theaction, whereas actions devoid of a goal would automati-cally trigger movement coding. We used a stimulus-response compatibility paradigm developed by Brass
et al. (2000) and adapted by Bertenthal et al. (2006), inwhich participants had to respond with a finger key pressto the spatial location (left or right side) of an eliciting stim-ulus, consisting of a finger movement, irrespective of theanatomical identity of the finger (index or middle finger)that was moved. In such situation, longer RTs for anatom-ically incompatible (i.e., responses performed by a differentfinger than the model) than for anatomically compatible(i.e., responses performed by the same finger as the model)movements are a measure of automatic imitation, in thatthey reflect the involuntary tendency to respond with thesame effector used in the observed action (even though itis task irrelevant). If movement coding plays the most cru-cial role in automatic imitation, we should find significantlygreater anatomical interference in the case of goal-lessactions. However, if goal and movement coding equallycontribute to automatic imitation, we should find similaranatomical interference for goal-directed and goal-lessactions.
Additionally, as a control condition, we verifiedwhether the presence of an action goal would modulatean effect similar to but distinct from automatic imitation,namely spatial compatibility. Spatial compatibility has beenshown to affect agents’ performance in automatic imitationtasks (Jansson, Wilson, Williams, & Mon-Williams, 2007),and therefore it is crucial to design experiments in a waythat allows researchers to differentiate spatial compatibilityfrom automatic imitation. With regard to the distinctionbetween goal-less and goal-directed actions, spatialcompatibility effects have been consistently observed instudies that used goal-less action stimuli (Bertenthalet al., 2006; Catmur & Heyes, 2011), and it has been ascer-tained that they are sensitive also to the spatial location ofresponse goals (e.g., the key being pressed, see Riggio,Gawryszewski, & Umilt�, 1986). In the present work wedirectly compared the effects of spatial compatibilityfollowing observation of closely matched goal-less andgoal-directed actions. Based on the available evidence,we expected to find a similar effect of spatial compatibilityfor goal-less as well as for goal-directed actions.
Finally, a further aim of the present work was to lookinto the influence of the order of presentation of the ana-tomically compatible and incompatible stimuli on the sizeof the automatic imitation effect, and of the spatially com-patible and incompatible stimuli on the size of the spatialcompatibility effect. Regarding automatic imitation,Bertenthal et al. (2006) presented anatomically compatibleand incompatible trials in separate mini-blocks and found arather small automatic imitation effect (9 ms), whereasCatmur and Heyes (2011) presented compatible and incom-patible trials randomly intermixed and found a more robustautomatic imitation effect (19 ms). We thus compared acompletely blocked presentation of the anatomicallycompatible and incompatible stimuli (with two differentcompatible stimuli, e.g., the index and the middle fingerof the left hand, and two incompatible stimuli, e.g., theindex and the middle finger of the right hand) and a com-pletely random presentation of the same compatible andincompatible stimuli, in order to directly assess whetherblocked presentation, by allowing the development of
2 C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding?
Author’s personal copy (e-offprint)
Experimental Psychology 2013 � 2013 Hogrefe Publishing

response strategies, indeed produces smaller automatic imi-tation effects than random presentation. As for the spatialcompatibility effect, whereas Bertenthal et al. (2006) andCatmur and Heyes (2011) found very similar results(41 ms and 42 ms, respectively) irrespective of the orderof presentation of the stimuli (blocked vs. random), the spa-tial compatibility literature shows that when spatially com-patible and incompatible mappings are intermixed within ablock, the spatial compatibility effect tends to be reduced oreliminated (Proctor & Vu, 2006). We therefore expectedthat the completely blocked presentation of the spatiallycompatible and incompatible stimuli might result in agreater spatial compatibility effect than the randompresentation.
Experiment 1
Method
Participants
Forty-eight students (24 females; mean age 24.9 €2.6 years) of the University of Turin took part in thisexperiment. All were right-handed, had normal or cor-rected-to-normal vision, and were na�ve to the purpose ofthe research. All the participants gave their informed con-sent prior to the inclusion in this study, which was per-formed in accordance with the ethical standards of the1964 Declaration of Helsinki.
Apparatus and Material
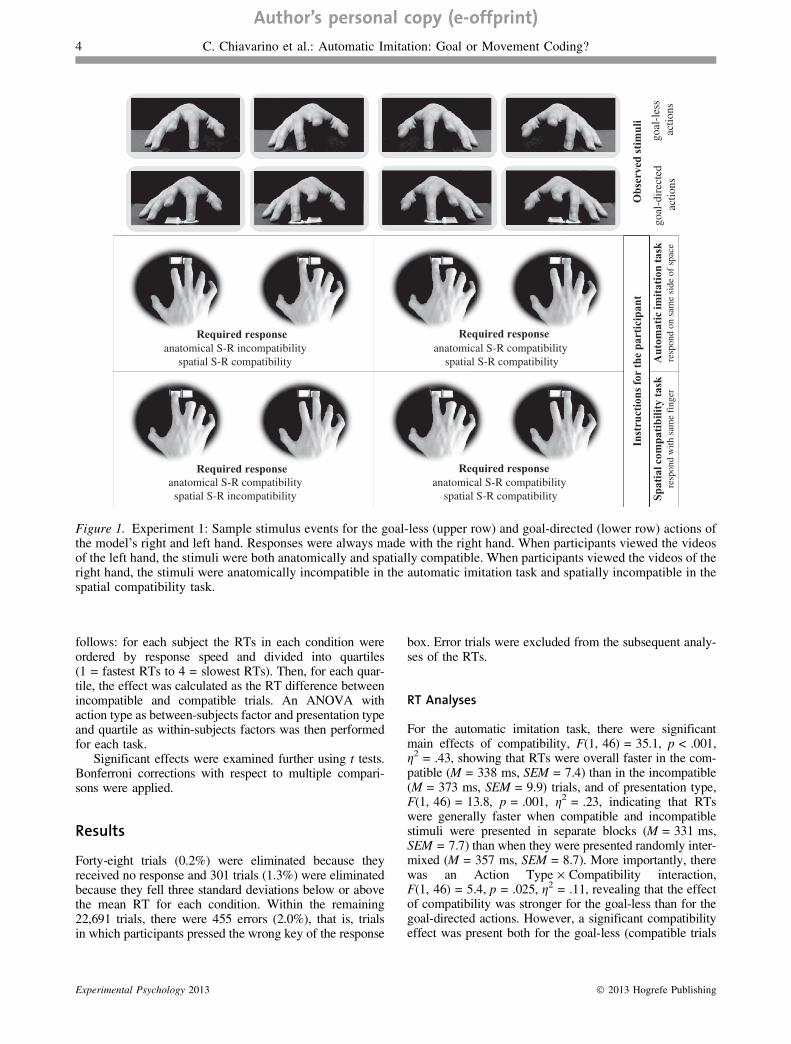
Videos of finger tapping goal-less and goal-directed actionsexecuted by a model’s right hand were filmed on a blackbackground in a third-person perspective, that is, with themodel sitting in front of the camera. The model wasinstructed to perform the very same movement (providedthe action still looked natural) in both types of videos,but in the goal-less actions the finger movements endedon a flat surface, while in the goal-directed actions thefinger movements ended on the keys of a response box(Figure 1). Sound effects were not included in the videos.
Four videos were filmed: goal-less and goal-directedmovements of the right index finger, and goal-less andgoal-directed movements of the right middle finger. Fiveframes were extracted from each video and, by reversingeach frame, identical sequences were obtained representingmovements of the index and middle finger of the left hand.
New video sequences were then produced using thesoftware Presentation (Neurobehavioral Systems, Albany,NY): in each video the first frame, portraying the initialposition of the hand, remained visible for 1,533 ms; the fol-lowing three frames, representing the movement sequence,lasted 38 ms each; the last frame, depicting the final posi-tion of the hand, remained visible for 886 ms. Eight typesof trials resulted from the combination of three variables:
finger (index, middle), hand (right, left), and type of action(goal-less, goal-directed). Each trial was preceded by a fix-ation cross for 1,000 ms and followed by a black screen for2,467 ms, therefore lasting 6,000 ms in total.
Procedure
Participants were tested individually in a dimly lit room.They sat approximately 60 cm from the monitor of a com-puter (17 inch, refresh rate 60 Hz), with their right indexand middle finger resting, respectively, on the left and rightkeys of a response box.
Each participant was asked to perform two differenttasks. The automatic imitation task required to respond witha key press of the finger from the same side of space as thefinger used by the model. Each participant was also askedto perform a spatial compatibility task, which required torespond with a key press of the same finger used by themodel. Participants’ response times were measured fromthe second frame of each video sequence, correspondingto the onset of the presented finger movement.
When participants observed the videos of the left hand,the instructions of the two tasks produced identicalresponses, because the index finger always appeared onthe left of the screen (requiring the pressing of the leftkey by the participant’s right index finger) and the middlefinger always appeared on the right of the screen (requiringthe pressing of the right key by the participant’s right mid-dle finger). In other words, when participants viewed thevideos of the left hand, the stimuli were both anatomicallyand spatially compatible. In contrast, when participantsobserved the videos of the right hand, the stimuli were ana-tomically incompatible in the automatic imitation task andspatially incompatible in the spatial compatibility task.A RT difference between responses to compatible andincompatible trials in the automatic imitation task wouldreflect the occurrence of automatic imitation, while a RTdifference between responses to compatible and incompat-ible trials in the spatial compatibility task would indicatethe presence of spatial compatibility.
Participants executed each of the two tasks twice, oncewith a blocked and once with a random stimulus presenta-tion. Every condition comprised two blocks of 60 trialseach, for a total of 480 trials, and was preceded by 8 prac-tice trials, which were excluded from further analysis.
Design and Statistical Analyses
Separate ANOVAs with action type (goal-less vs. goal-directed) as between-subjects factor and compatibility(automatic imitation vs. spatial compatibility) and presenta-tion type (blocked vs. random presentation of the left- andright-hand stimuli) as within-subjects factors were con-ducted on the RTs and on the error data of the automaticimitation and of the spatial compatibility task.
The time course of the automatic imitation and of thespatial compatibility effect across trials was measured as
C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding? 3
Author’s personal copy (e-offprint)
� 2013 Hogrefe Publishing Experimental Psychology 2013
follows: for each subject the RTs in each condition wereordered by response speed and divided into quartiles(1 = fastest RTs to 4 = slowest RTs). Then, for each quar-tile, the effect was calculated as the RT difference betweenincompatible and compatible trials. An ANOVA withaction type as between-subjects factor and presentation typeand quartile as within-subjects factors was then performedfor each task.
Significant effects were examined further using t tests.Bonferroni corrections with respect to multiple compari-sons were applied.
Results
Forty-eight trials (0.2%) were eliminated because theyreceived no response and 301 trials (1.3%) were eliminatedbecause they fell three standard deviations below or abovethe mean RT for each condition. Within the remaining22,691 trials, there were 455 errors (2.0%), that is, trialsin which participants pressed the wrong key of the response
box. Error trials were excluded from the subsequent analy-ses of the RTs.
RT Analyses
For the automatic imitation task, there were significantmain effects of compatibility, F(1, 46) = 35.1, p < .001,g2 = .43, showing that RTs were overall faster in the com-patible (M = 338 ms, SEM = 7.4) than in the incompatible(M = 373 ms, SEM = 9.9) trials, and of presentation type,F(1, 46) = 13.8, p = .001, g2 = .23, indicating that RTswere generally faster when compatible and incompatiblestimuli were presented in separate blocks (M = 331 ms,SEM = 7.7) than when they were presented randomly inter-mixed (M = 357 ms, SEM = 8.7). More importantly, therewas an Action Type · Compatibility interaction,F(1, 46) = 5.4, p = .025, g2 = .11, revealing that the effectof compatibility was stronger for the goal-less than for thegoal-directed actions. However, a significant compatibilityeffect was present both for the goal-less (compatible trials
Obs
erve
dst
imul
i
Aut
omat
icim
itatio
nta
sk
resp
ond
onsa
me
side
of
spac
e
anatomical S-R incompatibilityspatial S-R compatibility
anatomical S-R compatibility spatial S-R compatibility
anatomical S-R compatibility spatial S-R compatibility
anatomical S-R compatibilityspatial S-R incompatibility Sp
atia
lcom
patib
ility
task
re
spon
dw
ithsa
me
fing
ergo
al-d
irec
ted
actio
nsgo
al-l
ess
actio
ns
Inst
ruct
ions
for
the
part
icip
ant
Required response Required response
Required response Required response
Figure 1. Experiment 1: Sample stimulus events for the goal-less (upper row) and goal-directed (lower row) actions ofthe model’s right and left hand. Responses were always made with the right hand. When participants viewed the videosof the left hand, the stimuli were both anatomically and spatially compatible. When participants viewed the videos of theright hand, the stimuli were anatomically incompatible in the automatic imitation task and spatially incompatible in thespatial compatibility task.
4 C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding?
Author’s personal copy (e-offprint)
Experimental Psychology 2013 � 2013 Hogrefe Publishing
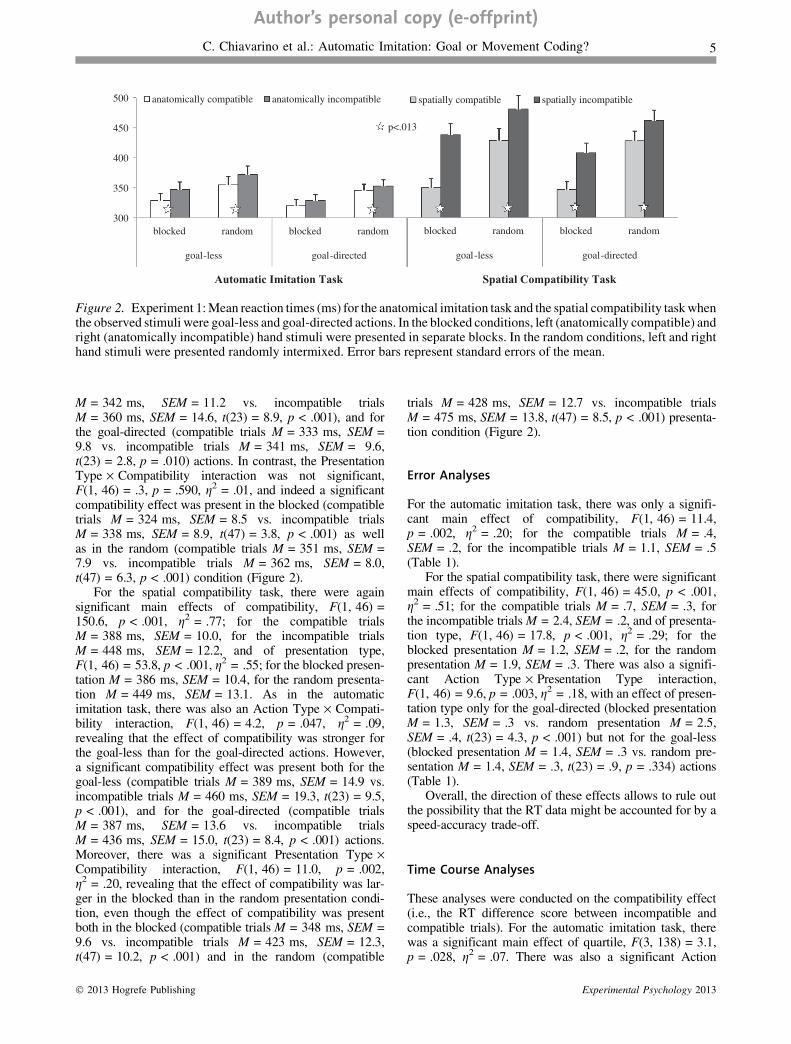
M = 342 ms, SEM = 11.2 vs. incompatible trialsM = 360 ms, SEM = 14.6, t(23) = 8.9, p < .001), and forthe goal-directed (compatible trials M = 333 ms, SEM =9.8 vs. incompatible trials M = 341 ms, SEM = 9.6,t(23) = 2.8, p = .010) actions. In contrast, the PresentationType · Compatibility interaction was not significant,F(1, 46) = .3, p = .590, g2 = .01, and indeed a significantcompatibility effect was present in the blocked (compatibletrials M = 324 ms, SEM = 8.5 vs. incompatible trialsM = 338 ms, SEM = 8.9, t(47) = 3.8, p < .001) as wellas in the random (compatible trials M = 351 ms, SEM =7.9 vs. incompatible trials M = 362 ms, SEM = 8.0,t(47) = 6.3, p < .001) condition (Figure 2).
For the spatial compatibility task, there were againsignificant main effects of compatibility, F(1, 46) =150.6, p < .001, g2 = .77; for the compatible trialsM = 388 ms, SEM = 10.0, for the incompatible trialsM = 448 ms, SEM = 12.2, and of presentation type,F(1, 46) = 53.8, p < .001, g2 = .55; for the blocked presen-tation M = 386 ms, SEM = 10.4, for the random presenta-tion M = 449 ms, SEM = 13.1. As in the automaticimitation task, there was also an Action Type · Compati-bility interaction, F(1, 46) = 4.2, p = .047, g2 = .09,revealing that the effect of compatibility was stronger forthe goal-less than for the goal-directed actions. However,a significant compatibility effect was present both for thegoal-less (compatible trials M = 389 ms, SEM = 14.9 vs.incompatible trials M = 460 ms, SEM = 19.3, t(23) = 9.5,p < .001), and for the goal-directed (compatible trialsM = 387 ms, SEM = 13.6 vs. incompatible trialsM = 436 ms, SEM = 15.0, t(23) = 8.4, p < .001) actions.Moreover, there was a significant Presentation Type ·Compatibility interaction, F(1, 46) = 11.0, p = .002,g2 = .20, revealing that the effect of compatibility was lar-ger in the blocked than in the random presentation condi-tion, even though the effect of compatibility was presentboth in the blocked (compatible trials M = 348 ms, SEM =9.6 vs. incompatible trials M = 423 ms, SEM = 12.3,t(47) = 10.2, p < .001) and in the random (compatible
trials M = 428 ms, SEM = 12.7 vs. incompatible trialsM = 475 ms, SEM = 13.8, t(47) = 8.5, p < .001) presenta-tion condition (Figure 2).
Error Analyses
For the automatic imitation task, there was only a signifi-cant main effect of compatibility, F(1, 46) = 11.4,p = .002, g2 = .20; for the compatible trials M = .4,SEM = .2, for the incompatible trials M = 1.1, SEM = .5(Table 1).
For the spatial compatibility task, there were significantmain effects of compatibility, F(1, 46) = 45.0, p < .001,g2 = .51; for the compatible trials M = .7, SEM = .3, forthe incompatible trials M = 2.4, SEM = .2, and of presenta-tion type, F(1, 46) = 17.8, p < .001, g2 = .29; for theblocked presentation M = 1.2, SEM = .2, for the randompresentation M = 1.9, SEM = .3. There was also a signifi-cant Action Type · Presentation Type interaction,F(1, 46) = 9.6, p = .003, g2 = .18, with an effect of presen-tation type only for the goal-directed (blocked presentationM = 1.3, SEM = .3 vs. random presentation M = 2.5,SEM = .4, t(23) = 4.3, p < .001) but not for the goal-less(blocked presentation M = 1.4, SEM = .3 vs. random pre-sentation M = 1.4, SEM = .3, t(23) = .9, p = .334) actions(Table 1).
Overall, the direction of these effects allows to rule outthe possibility that the RT data might be accounted for by aspeed-accuracy trade-off.
Time Course Analyses
These analyses were conducted on the compatibility effect(i.e., the RT difference score between incompatible andcompatible trials). For the automatic imitation task, therewas a significant main effect of quartile, F(3, 138) = 3.1,p = .028, g2 = .07. There was also a significant Action
300
350
400
450
500
blocked random blocked random
goal-less goal-directed
anatomically compatible anatomically incompatible
Automatic Imitation Task
blocked random blocked random
goal-less goal-directed
spatially compatible spatially incompatible
Spatial Compatibility Task
p<.013
Figure 2. Experiment 1: Mean reaction times (ms) for the anatomical imitation task and the spatial compatibility task whenthe observed stimuli were goal-less and goal-directed actions. In the blocked conditions, left (anatomically compatible) andright (anatomically incompatible) hand stimuli were presented in separate blocks. In the random conditions, left and righthand stimuli were presented randomly intermixed. Error bars represent standard errors of the mean.
C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding? 5
Author’s personal copy (e-offprint)
� 2013 Hogrefe Publishing Experimental Psychology 2013
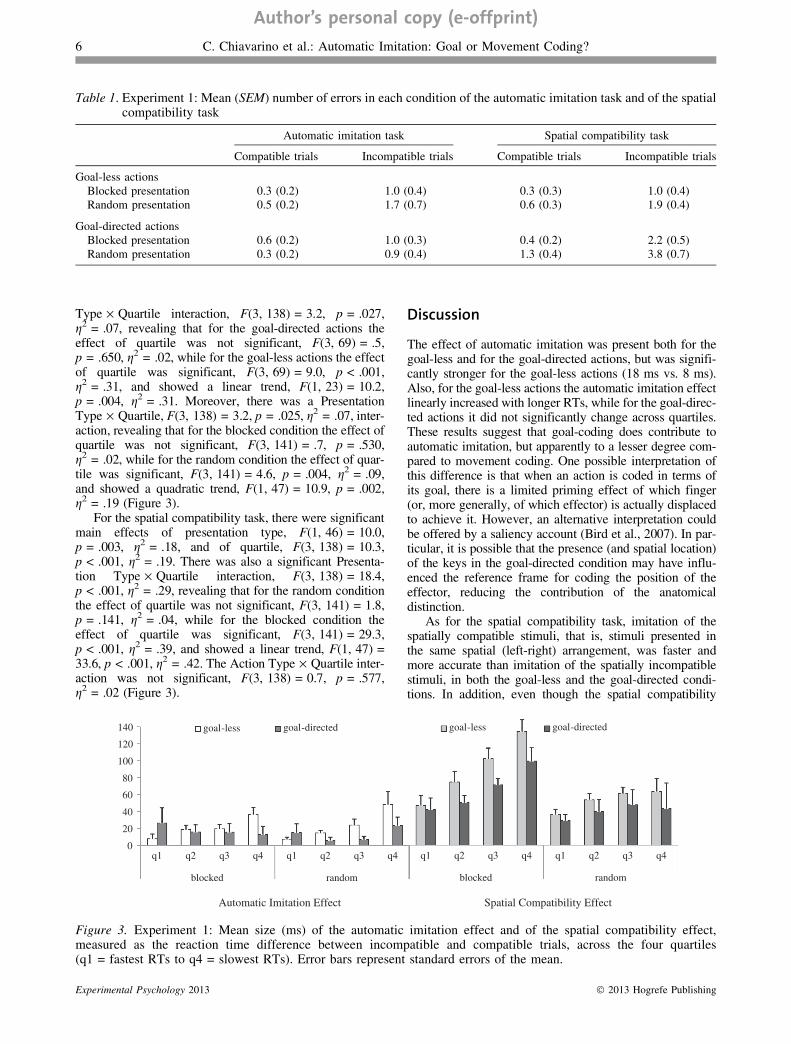
Type · Quartile interaction, F(3, 138) = 3.2, p = .027,g2 = .07, revealing that for the goal-directed actions theeffect of quartile was not significant, F(3, 69) = .5,p = .650, g2 = .02, while for the goal-less actions the effectof quartile was significant, F(3, 69) = 9.0, p < .001,g2 = .31, and showed a linear trend, F(1, 23) = 10.2,p = .004, g2 = .31. Moreover, there was a PresentationType · Quartile, F(3, 138) = 3.2, p = .025, g2 = .07, inter-action, revealing that for the blocked condition the effect ofquartile was not significant, F(3, 141) = .7, p = .530,g2 = .02, while for the random condition the effect of quar-tile was significant, F(3, 141) = 4.6, p = .004, g2 = .09,and showed a quadratic trend, F(1, 47) = 10.9, p = .002,g2 = .19 (Figure 3).
For the spatial compatibility task, there were significantmain effects of presentation type, F(1, 46) = 10.0,p = .003, g2 = .18, and of quartile, F(3, 138) = 10.3,p < .001, g2 = .19. There was also a significant Presenta-tion Type · Quartile interaction, F(3, 138) = 18.4,p < .001, g2 = .29, revealing that for the random conditionthe effect of quartile was not significant, F(3, 141) = 1.8,p = .141, g2 = .04, while for the blocked condition theeffect of quartile was significant, F(3, 141) = 29.3,p < .001, g2 = .39, and showed a linear trend, F(1, 47) =33.6, p < .001, g2 = .42. The Action Type · Quartile inter-action was not significant, F(3, 138) = 0.7, p = .577,g2 = .02 (Figure 3).
Discussion
The effect of automatic imitation was present both for thegoal-less and for the goal-directed actions, but was signifi-cantly stronger for the goal-less actions (18 ms vs. 8 ms).Also, for the goal-less actions the automatic imitation effectlinearly increased with longer RTs, while for the goal-direc-ted actions it did not significantly change across quartiles.These results suggest that goal-coding does contribute toautomatic imitation, but apparently to a lesser degree com-pared to movement coding. One possible interpretation ofthis difference is that when an action is coded in terms ofits goal, there is a limited priming effect of which finger(or, more generally, of which effector) is actually displacedto achieve it. However, an alternative interpretation couldbe offered by a saliency account (Bird et al., 2007). In par-ticular, it is possible that the presence (and spatial location)of the keys in the goal-directed condition may have influ-enced the reference frame for coding the position of theeffector, reducing the contribution of the anatomicaldistinction.
As for the spatial compatibility task, imitation of thespatially compatible stimuli, that is, stimuli presented inthe same spatial (left-right) arrangement, was faster andmore accurate than imitation of the spatially incompatiblestimuli, in both the goal-less and the goal-directed condi-tions. In addition, even though the spatial compatibility
Table 1. Experiment 1: Mean (SEM) number of errors in each condition of the automatic imitation task and of the spatialcompatibility task
Automatic imitation task Spatial compatibility task
Compatible trials Incompatible trials Compatible trials Incompatible trials
Goal-less actionsBlocked presentation 0.3 (0.2) 1.0 (0.4) 0.3 (0.3) 1.0 (0.4)Random presentation 0.5 (0.2) 1.7 (0.7) 0.6 (0.3) 1.9 (0.4)
Goal-directed actionsBlocked presentation 0.6 (0.2) 1.0 (0.3) 0.4 (0.2) 2.2 (0.5)Random presentation 0.3 (0.2) 0.9 (0.4) 1.3 (0.4) 3.8 (0.7)
0
20
40
60
80
100
120
140
q1 q2 q3 q4 q1 q2 q3 q4
blocked random
goal-less goal-directed
Automatic Imitation Effect
q1 q2 q3 q4 q1 q2 q3 q4
blocked random
goal-less goal-directed
Spatial Compatibility Effect
Figure 3. Experiment 1: Mean size (ms) of the automatic imitation effect and of the spatial compatibility effect,measured as the reaction time difference between incompatible and compatible trials, across the four quartiles(q1 = fastest RTs to q4 = slowest RTs). Error bars represent standard errors of the mean.
6 C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding?
Author’s personal copy (e-offprint)
Experimental Psychology 2013 � 2013 Hogrefe Publishing

effect was stronger for the goal-less (71 ms) than for thegoal-directed (49 ms) actions, the analysis of the timecourse of the spatial compatibility effect did not signifi-cantly differ between the two conditions. These findingsare consistent with the notion that spatial compatibilityrobustly depends both on goal coding and on movementcoding (Riggio et al., 1986).
Finally, relative to the effect of the order of presentationof the stimuli, in the automatic imitation task we found thatthe blocked presentation (14 ms) produced an effect of sim-ilar size to the random presentation (11 ms); however, inthe random condition the effect of automatic imitationincreased with longer RTs, while in the blocked conditionit did not significantly change across quartiles. This sug-gests that: (i) when anatomically compatible and incompat-ible stimuli are presented in separate blocks, it may bepossible to develop response strategies, whereas this doesnot happen when stimuli are presented randomly inter-mixed; (ii) in our task these strategies were not effectiveenough to result in a significant difference in the size ofthe automatic imitation effect between the blocked andthe random conditions. As for the spatial compatibility task,consistent with the spatial compatibility literature, ourresults showed a stronger effect in the blocked (75 ms) thanin the random (47 ms) presentation condition, as well as anincrease of the effect as a function of quartile only in theblocked condition. In order to better control the contribu-tion of goal and movement coding to automatic imitationand spatial compatibility, we run an additional experiment.
Experiment 2
Experiment 1 was not designed to directly compare theeffects of spatial compatibility and automatic imitation.Differences in task instruction may result in differences intask difficulty and therefore in response speed, as well asin potentially different coding strategies.
The experimental procedure used in this second exper-iment allowed a direct comparison between the automaticimitation and the spatial compatibility effects. Importantly,it also allowed to control the saliency of the frame of refer-ence provided by the response keys in the goal-directedcondition. We expected to find that automatic imitationand spatial compatibility are indeed two distinct phenom-ena, characterized by a different time course. We alsoexpected that, even though saliency effects may modulategoal and movement coding processes, automatic imitationwould rely more on movement than on goal coding, whilespatial compatibility would similarly depend on bothprocesses.
Method
Participants
Forty students (20 females; mean age 24.0 € 5.5 years) ofthe University of Turin took part in this experiment. All
were right-handed, had normal or corrected-to-normalvision, were na�ve to the purpose of the research, and gavetheir informed consent prior to the inclusion in this study.
Apparatus and Material
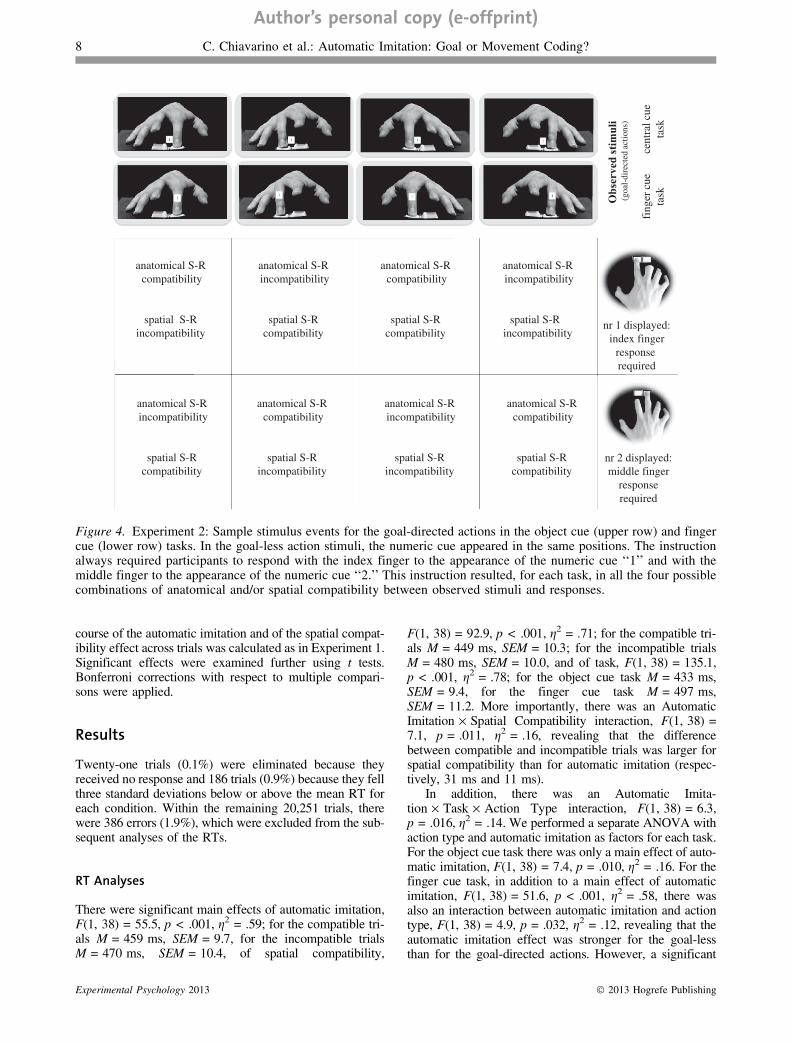
The same video sequences of goal-directed and goal-lessfinger movements employed in Experiment 1 were used.However, they were modified in order to create two newtasks. In the first task, starting from the second frame ofeach sequence (i.e., when the finger began moving), anumeric cue (‘‘1’’ or ‘‘2’’) appeared on the moving finger(finger cue task). Because participants were instructed torespond to the numeric cue, this manipulation increasedthe saliency of the effector, both in the goal-directed andin the goal-less conditions, compared to Experiment 1. Inthe second task, the numeric cue appeared between theresponse keys (in the goal-directed condition) or in the cor-responding position on the flat surface (in the goal-less con-dition) (object cue task). This manipulation resulted in anincrease of the saliency of the goal or of the end positionof the action, and at the same time in a decreased saliencyof the spatial reference frame compared to Experiment 1(Figure 4).
Procedure
Each participant was asked to perform both the finger cuetask and the object cue task. In both tasks, participants wereinstructed to execute, with their right hand, an index fingermovement in response to the presentation of the number‘‘1’’ and a middle finger movement in response to the pre-sentation of the number ‘‘2.’’ Simultaneous with the appear-ance of the numeric cue, the same video sequences used inExperiment 1 were presented. The combination of theobserved movements with the numeric cues resulted in a2 · 2 manipulation of automatic imitation and spatial com-patibility for each task (Figure 4).
Participants’ response times were measured from thesecond frame of each video sequence, corresponding tothe onset of the numeric cue (and of the finger movement).Every condition comprised two blocks of 64 trials each, fora total of 512 trials, and was preceded by 8 practice trials,which were excluded from further analysis.
Design and Statistical Analyses
An ANOVA with action type (goal-less vs. goal-directed)as between-subjects factor and automatic imitation (com-patible vs. incompatible trials), spatial compatibility (com-patible vs. incompatible trials), and task (object cue vs.finger cue) as within-subjects factors was conducted onthe RTs and on the error data of the automatic imitationtask and of the spatial compatibility task. Because, withthe present set-up, observation of each hand could lead toboth compatible and incompatible mappings, the presenta-tion type factor was not included in the analyses. The time
C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding? 7
Author’s personal copy (e-offprint)
� 2013 Hogrefe Publishing Experimental Psychology 2013
course of the automatic imitation and of the spatial compat-ibility effect across trials was calculated as in Experiment 1.Significant effects were examined further using t tests.Bonferroni corrections with respect to multiple compari-sons were applied.
Results
Twenty-one trials (0.1%) were eliminated because theyreceived no response and 186 trials (0.9%) because they fellthree standard deviations below or above the mean RT foreach condition. Within the remaining 20,251 trials, therewere 386 errors (1.9%), which were excluded from the sub-sequent analyses of the RTs.
RT Analyses
There were significant main effects of automatic imitation,F(1, 38) = 55.5, p < .001, g2 = .59; for the compatible tri-als M = 459 ms, SEM = 9.7, for the incompatible trialsM = 470 ms, SEM = 10.4, of spatial compatibility,
F(1, 38) = 92.9, p < .001, g2 = .71; for the compatible tri-als M = 449 ms, SEM = 10.3; for the incompatible trialsM = 480 ms, SEM = 10.0, and of task, F(1, 38) = 135.1,p < .001, g2 = .78; for the object cue task M = 433 ms,SEM = 9.4, for the finger cue task M = 497 ms,SEM = 11.2. More importantly, there was an AutomaticImitation · Spatial Compatibility interaction, F(1, 38) =7.1, p = .011, g2 = .16, revealing that the differencebetween compatible and incompatible trials was larger forspatial compatibility than for automatic imitation (respec-tively, 31 ms and 11 ms).
In addition, there was an Automatic Imita-tion · Task · Action Type interaction, F(1, 38) = 6.3,p = .016, g2 = .14. We performed a separate ANOVA withaction type and automatic imitation as factors for each task.For the object cue task there was only a main effect of auto-matic imitation, F(1, 38) = 7.4, p = .010, g2 = .16. For thefinger cue task, in addition to a main effect of automaticimitation, F(1, 38) = 51.6, p < .001, g2 = .58, there wasalso an interaction between automatic imitation and actiontype, F(1, 38) = 4.9, p = .032, g2 = .12, revealing that theautomatic imitation effect was stronger for the goal-lessthan for the goal-directed actions. However, a significant
anatomical S-Rincompatibility
spatial S-Rcompatibility
anatomical S-Rincompatibility
spatial S-Rcompatibility
anatomical S-Rcompatibility
spatial S-Rincompatibility
anatomical S-Rcompatibility
spatial S-Rincompatibility
anatomical S-Rcompatibility
spatial S-Rcompatibility
anatomical S-Rcompatibility
spatial S-Rcompatibility
anatomical S-Rincompatibility
spatial S-Rincompatibility
anatomical S-Rincompatibility
spatial S-Rincompatibility
Obs
erve
dst
imul
i(g
oal-
dire
cted
actio
ns)
cent
ralc
ueta
skfi
nger
cue
task
nr 1 displayed:index finger
response required
nr 2 displayed:middle finger
response required
Figure 4. Experiment 2: Sample stimulus events for the goal-directed actions in the object cue (upper row) and fingercue (lower row) tasks. In the goal-less action stimuli, the numeric cue appeared in the same positions. The instructionalways required participants to respond with the index finger to the appearance of the numeric cue ‘‘1’’ and with themiddle finger to the appearance of the numeric cue ‘‘2.’’ This instruction resulted, for each task, in all the four possiblecombinations of anatomical and/or spatial compatibility between observed stimuli and responses.
8 C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding?
Author’s personal copy (e-offprint)
Experimental Psychology 2013 � 2013 Hogrefe Publishing
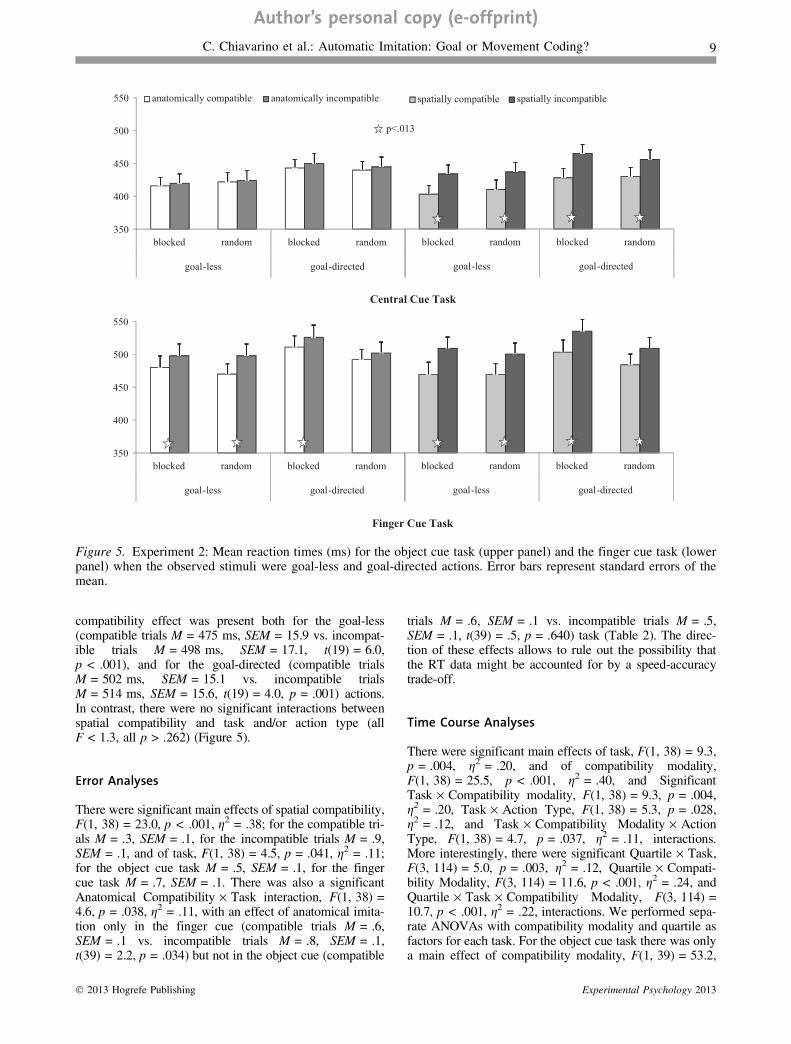
compatibility effect was present both for the goal-less(compatible trials M = 475 ms, SEM = 15.9 vs. incompat-ible trials M = 498 ms, SEM = 17.1, t(19) = 6.0,p < .001), and for the goal-directed (compatible trialsM = 502 ms, SEM = 15.1 vs. incompatible trialsM = 514 ms, SEM = 15.6, t(19) = 4.0, p = .001) actions.In contrast, there were no significant interactions betweenspatial compatibility and task and/or action type (allF < 1.3, all p > .262) (Figure 5).
Error Analyses
There were significant main effects of spatial compatibility,F(1, 38) = 23.0, p < .001, g2 = .38; for the compatible tri-als M = .3, SEM = .1, for the incompatible trials M = .9,SEM = .1, and of task, F(1, 38) = 4.5, p = .041, g2 = .11;for the object cue task M = .5, SEM = .1, for the fingercue task M = .7, SEM = .1. There was also a significantAnatomical Compatibility · Task interaction, F(1, 38) =4.6, p = .038, g2 = .11, with an effect of anatomical imita-tion only in the finger cue (compatible trials M = .6,SEM = .1 vs. incompatible trials M = .8, SEM = .1,t(39) = 2.2, p = .034) but not in the object cue (compatible
trials M = .6, SEM = .1 vs. incompatible trials M = .5,SEM = .1, t(39) = .5, p = .640) task (Table 2). The direc-tion of these effects allows to rule out the possibility thatthe RT data might be accounted for by a speed-accuracytrade-off.
Time Course Analyses
There were significant main effects of task, F(1, 38) = 9.3,p = .004, g2 = .20, and of compatibility modality,F(1, 38) = 25.5, p < .001, g2 = .40, and SignificantTask · Compatibility modality, F(1, 38) = 9.3, p = .004,g2 = .20, Task · Action Type, F(1, 38) = 5.3, p = .028,g2 = .12, and Task · Compatibility Modality · ActionType, F(1, 38) = 4.7, p = .037, g2 = .11, interactions.More interestingly, there were significant Quartile · Task,F(3, 114) = 5.0, p = .003, g2 = .12, Quartile · Compati-bility Modality, F(3, 114) = 11.6, p < .001, g2 = .24, andQuartile · Task · Compatibility Modality, F(3, 114) =10.7, p < .001, g2 = .22, interactions. We performed sepa-rate ANOVAs with compatibility modality and quartile asfactors for each task. For the object cue task there was onlya main effect of compatibility modality, F(1, 39) = 53.2,
350
400
450
500
550
blocked random blocked random
goal-less goal-directed
anatomically compatible anatomically incompatible
blocked random blocked random
goal-less goal-directed
spatially compatible spatially incompatible
p<.013
Central Cue Task
350
400
450
500
550
blocked random blocked random
goal-less goal-directed
blocked random blocked random
goal-less goal-directed
Finger Cue Task
Figure 5. Experiment 2: Mean reaction times (ms) for the object cue task (upper panel) and the finger cue task (lowerpanel) when the observed stimuli were goal-less and goal-directed actions. Error bars represent standard errors of themean.
C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding? 9
Author’s personal copy (e-offprint)
� 2013 Hogrefe Publishing Experimental Psychology 2013
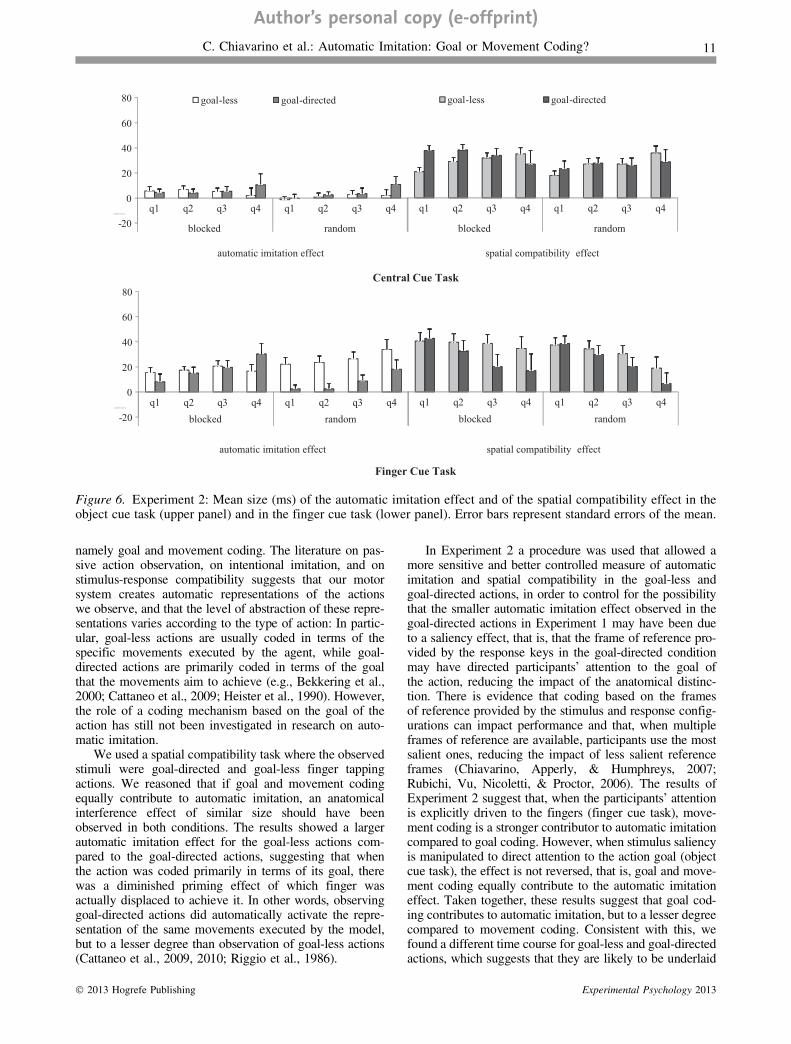
p < .001, g2 = .58, and of quartile, F(3, 117) = 3.3,p = .022, g2 = .08, with the quartile function showing acubic trend, F(1, 39) = 4.4, p = .043, g2 = .10 andQuartile · Compatibility interaction, F(3, 117) = 0.3,p = .837, g2 = .01. For the finger cue task, instead, besidea main effect of compatibility modality, F(1, 39) = 6.6,p = .014, g2 = .15, there was a significant Quartile · Com-patibility Modality interaction, F(3, 117) = 20.3, p < .001,g2 = .34, revealing that the effect of automatic imitationincreased with longer RTs, F(3, 117) = 7.5, p < .001,g2 = .16, with a linear trend, F(1, 39) = 9.7, p = .003,g2 = .20, while the effect of spatial compatibility decreasedover quartiles, F(3, 117) = 13.0, p < .001, g2 = .25, againwith a linear trend, F(1, 39) = 17.3, p < .001, g2 = .31.
In addition, there was a significant Quartile · Compati-bility Modality · Action Type interaction, F(3, 114) = 8.8,p < .001, g2 = .19, and separate ANOVAs with compatibil-ity modality and quartile as factors were performed for eachaction type. For the goal-directed actions there were a signif-icant main effect of compatibility modality, F(1, 19) = 9.7,p = .006, g2 = .34, and a significant Quartile · Compatibil-ity Modality interaction, F(3, 57) = 15.1, p < .001,g2 = .44, revealing that the effect of automatic imitationincreased with longer RTs, F(3, 57) = 10.2, p < .001,g2 = .35, with a linear trend, F(1, 19) = 12.3, p = .002,g2 = .39, while the effect of spatial compatibility decreasedover quartiles, F(3, 57) = 5.4, p = .002, g2 = .25, againwith a linear trend, F(1, 19) = 6.3, p = .022, g2 = .25. Incontrast, for the goal-less actions there was only a maineffect of compatibility modality, F(1, 19) = 18.7,p < .001, g2 = .50, and neither the automatic imitationeffect, F(3, 57) = 1.0, p = .414, g2 = .05, nor the spatialcompatibility effect, F(3, 57) = 0.6, p = .592, g2 = .03, sig-nificantly changed over quartiles (Figure 6).
Discussion
In Experiment 2 we developed a new task to directly com-pare the effects of automatic imitation and spatial compat-
ibility in the goal-directed and goal-less actions. In line withExperiment 1, the results showed that the differencebetween the spatially compatible and incompatible trialswas larger than the difference between the anatomicallycompatible and incompatible ones. Also, in the finger cuetask the time courses of the two effects differed: Automaticimitation increased, whereas spatial compatibilitydecreased, with longer RTs. These are additional findingsconsistent with the recent literature which considers spatialcompatibility and automatic imitation as two distinct phe-nomena (Catmur & Heyes, 2011; however, note that in thatstudy the effect of spatial compatibility did not decrease,but remained constant over quartiles).
In Experiment 2 we also controlled for stimulus saliencyby adding to the stimuli used in Experiment 1, for both thegoal-directed and the goal-less actions, a numeric cuewhich directed participants’ attention, in one case, to themovement dimension (finger cue task) and, in the othercase, to the goal dimension (object cue task). In the fingercue task, the goal-less actions were more affected by thecompatibility effect than the goal-directed ones. This find-ing replicates the results of Experiment 1 and suggests that,even when the difference in the saliency of the anatomicaldimension between the conditions is eliminated, the contri-bution of movement coding to automatic imitation is stron-ger than that of goal coding.
In the object cue task, however, the automatic imitationeffect did not differ between the goal-less and the goal-directed actions. This finding suggests that increasing thesaliency of the goal (or end position) of the action is notenough to increase the contribution of goal coding over thatof movement coding. Taken together, these results confirmthat goal coding is a weaker contributor than movementcoding to automatic imitation.
General Discussion
The main goal of the present study was to analyze the con-tribution of two distinct processes to automatic imitation,
Table 2. Experiment 2: Mean (SEM) number of errors in each condition of the object cue task and of the finger cue task
Anatomically compatible Anatomically incompatible
Spatiallycompatible
Spatiallyincompatible
Spatiallycompatible
Spatiallyincompatible
Object cue task Goal-less actionsBlocked presentation 0.5 (0.2) 0.9 (0.3) 0.5 (0.2) 0.8 (0.2)Random presentation 0.4 (0.1) 0.4 (0.1) 0.3 (0.1) 0.5 (0.2)
Goal-directed actionsBlocked presentation 0.2 (0.2) 0.8 (0.2) 0.2 (0.1) 1.0 (0.3)Random presentation 0.4 (0.2) 0.9 (0.3) 0.2 (0.1) 0.8 (0.3)
Finger cue task Goal-less actionsBlocked presentation 0.2 (0.1) 1.0 (0.3) 0.3 (0.2) 1.5 (0.4)Random presentation 0.2 (0.1) 1.0 (0.3) 0.4 (0.2) 1.0 (0.3)
Goal-directed actionsBlocked presentation 0.4 (0.1) 1.1 (0.3) 0.6 (0.2) 1.3 (0.3)Random presentation 0.3 (0.1) 0.7 (0.2) 0.4 (0.1) 1.0 (0.4)
10 C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding?
Author’s personal copy (e-offprint)
Experimental Psychology 2013 � 2013 Hogrefe Publishing
namely goal and movement coding. The literature on pas-sive action observation, on intentional imitation, and onstimulus-response compatibility suggests that our motorsystem creates automatic representations of the actionswe observe, and that the level of abstraction of these repre-sentations varies according to the type of action: In partic-ular, goal-less actions are usually coded in terms of thespecific movements executed by the agent, while goal-directed actions are primarily coded in terms of the goalthat the movements aim to achieve (e.g., Bekkering et al.,2000; Cattaneo et al., 2009; Heister et al., 1990). However,the role of a coding mechanism based on the goal of theaction has still not been investigated in research on auto-matic imitation.
We used a spatial compatibility task where the observedstimuli were goal-directed and goal-less finger tappingactions. We reasoned that if goal and movement codingequally contribute to automatic imitation, an anatomicalinterference effect of similar size should have beenobserved in both conditions. The results showed a largerautomatic imitation effect for the goal-less actions com-pared to the goal-directed actions, suggesting that whenthe action was coded primarily in terms of its goal, therewas a diminished priming effect of which finger wasactually displaced to achieve it. In other words, observinggoal-directed actions did automatically activate the repre-sentation of the same movements executed by the model,but to a lesser degree than observation of goal-less actions(Cattaneo et al., 2009, 2010; Riggio et al., 1986).
In Experiment 2 a procedure was used that allowed amore sensitive and better controlled measure of automaticimitation and spatial compatibility in the goal-less andgoal-directed actions, in order to control for the possibilitythat the smaller automatic imitation effect observed in thegoal-directed actions in Experiment 1 may have been dueto a saliency effect, that is, that the frame of reference pro-vided by the response keys in the goal-directed conditionmay have directed participants’ attention to the goal ofthe action, reducing the impact of the anatomical distinc-tion. There is evidence that coding based on the framesof reference provided by the stimulus and response config-urations can impact performance and that, when multipleframes of reference are available, participants use the mostsalient ones, reducing the impact of less salient referenceframes (Chiavarino, Apperly, & Humphreys, 2007;Rubichi, Vu, Nicoletti, & Proctor, 2006). The results ofExperiment 2 suggest that, when the participants’ attentionis explicitly driven to the fingers (finger cue task), move-ment coding is a stronger contributor to automatic imitationcompared to goal coding. However, when stimulus saliencyis manipulated to direct attention to the action goal (objectcue task), the effect is not reversed, that is, goal and move-ment coding equally contribute to the automatic imitationeffect. Taken together, these results suggest that goal cod-ing contributes to automatic imitation, but to a lesser degreecompared to movement coding. Consistent with this, wefound a different time course for goal-less and goal-directedactions, which suggests that they are likely to be underlaid
-20
0
20
40
60
80
q1 q2 q3 q4 q1 q2 q3 q4blocked random
compatibility effect (ms)
automatic imitation effect
q1 q2 q3 q4 q1 q2 q3 q4blocked random
spatial compatibility effect
Finger Cue Task
-20
0
20
40
60
80
q1 q2 q3 q4 q1 q2 q3 q4
blocked random
compatibility effect (ms)
goal-less goal-directed
automatic imitation effect
q1 q2 q3 q4 q1 q2 q3 q4
blocked random
goal-less goal-directed
spatial compatibility effect
Central Cue Task
Figure 6. Experiment 2: Mean size (ms) of the automatic imitation effect and of the spatial compatibility effect in theobject cue task (upper panel) and in the finger cue task (lower panel). Error bars represent standard errors of the mean.
C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding? 11
Author’s personal copy (e-offprint)
� 2013 Hogrefe Publishing Experimental Psychology 2013

by distinct processes. Note, however, that in the object cuetask the spatial saliency (i.e., left-right coding) of the cuewas reduced; thus, it is possible that a specific type of sal-iency, namely spatial saliency, may have contributed to theobserved difference between goal-less and goal-directedactions. It will be interesting for future studies to furtherinvestigate the effects of spatial saliency on automaticimitation.
Preliminary evidence that goal coding contributes toautomatic imitation was provided by Longo et al. (2008),who showed that top-down instructions can modulatewhether an action is coded in terms of its final goal (e.g.,‘‘tapping’’) or in terms of the movements performed. Ourresults further indicate that automatic imitation can be mod-ulated also by bottom-up features of the observed action,such as the position of the response cue. Consistent withthis, it has been observed that automatic imitation can beinfluenced by emotional primes, and more specifically thatthe anatomical compatibility effect increases when thestimulus movement is preceded by an affectively negativepicture (Grecucci, Koch, & Rumiati, 2011).
Our results further show that automatic imitation is sen-sitive to a number of features of the experimental set-upthat have often not been controlled in other studies. In par-ticular, in many experiments on automatic imitation stimulihave been presented in the same (left-right or up-down)spatial arrangement as that in which the participant’sresponses were executed, making it impossible to decidewhether the facilitation/interference between observed andexecuted actions had to be attributed to the tendency toreproduce a topographically similar response as the stimu-lus (automatic imitation), or to the tendency to respond inthe same relative position as the stimulus (spatial compati-bility), or to both factors together (Heyes, 2011). Recentfindings suggest that automatic imitation and spatial com-patibility are indeed two separate phenomena (Bertenthalet al., 2006; Catmur & Heyes, 2011), and our results pro-vide additional support to this hypothesis. In addition, theyare consistent with previous work showing that spatial com-patibility depends as much on goal coding and on move-ment coding (Riggio et al., 1986), and extend suchfindings also to the case of biological stimuli (finger’smovement).
Another factor that has often not been controlled inother studies is the order of presentation of the stimuli,which may potentiate or weaken the automatic imitationeffect. The difference between blocked and random stimu-lus presentation that was found in Experiment 1 is consis-tent with the notion that blocks of spatially compatiblemappings recruit a direct response-selection route basedon overlearned associations, while blocks of incompatiblemappings and randomly intermixed compatible and incom-patible mappings require the suppression of this route (as itwould lead to the wrong response in incompatible trials)and recruit instead an indirect route based on intentionalresponse identification (De Jong, 1995; Vu & Proctor,2004). However, it is worth noting that in the compatibleblocks of the blocked presentation condition the task canbe (perhaps more easily) performed as a spatial responsetask, that is, participants could realize that the correct
response is always spatially compatible, and thereforethey would not need to process which finger was moving.Therefore, it is possible that the significant reduction inRTs in these blocks may be accounted for by such a strat-egy. To conclude, it seems important for future studies onautomatic imitation to take into account a number of differ-ent factors and to measure their specific contribution to theobserved effects.
Acknowledgments
This research was supported by funding from San PaoloFoundation (Neuroscience Programme: Action representa-tions and their impairment, 2009–2012). We wish to thankFilippo Migliorati for his valuable assistance with Presenta-tion software, Valentina Laudati for data collection onExperiment 2, and three anonymous reviewers for theirgenerous feedback.
References
Bekkering, H., Wohlschl�ger, A., & Gattis, M. (2000). Imitationof gestures in children is goal-directed. The QuarterlyJournal of Experimental Psychology, 53, 153–164.
Bertenthal, B. I., Longo, M. R., & Kosobud, A. (2006).Imitative response tendencies following observation of no-goal directed actions. Journal of Experimental Psychology:Human Perception and Performance, 32, 210–225.
Bird, G., Brindley, R., Leighton, J., & Heyes, C. (2007). Generalprocesses, rather than ‘‘goals,’’ explain imitation errors.Journal of Experimental Psychology: Human Perceptionand Performance, 33, 1158–1169.
Bouquet, C. A., Shipley, T. F., Capa, R. L., & Marshall, P. J.(2011). Goal-directed actions are more contagious than non-goal-directed actions. Experimental Psychology, 58, 71–78.
Brass, M., Bekkering, H., Wohlschl�ger, A., & Prinz, W. (2000).Compatibility between observed and executed finger move-ments: Comparing symbolic, spatial and imitative cues.Brain and Cognition, 44, 124–143.
Catmur, C., & Heyes, C. (2011). Time course analyses confirmindependence of automatic imitation and spatial compati-bility effects. Journal of Experimental Psychology: HumanPerception and Performance, 37, 409–421.
Cattaneo, L., Caruana, F., Jezzini, A., & Rizzolatti, G. (2009).Representation of goal and movements without overt motorbehavior in the human motor cortex: A transcranialmagnetic stimulation study. The Journal of Neuroscience,29, 11134–11138.
Cattaneo, L., Sandrini, M., & Schwarzbach, J. (2010). Repre-sentation of goal state-dependent TMS reveals a hierarchicalrepresentation of observed acts in the temporal, parietal, andpremotor cortices. Cerebral Cortex, 20, 2252–2258.
Chiavarino, C., Apperly, I. A., & Humphreys, G. W. (2007).Exploring the functional and anatomical bases of mirror-image and anatomical imitation: The role of the frontallobes. Neuropsychologia, 47, 784–795.
Chiavarino, C., Apperly, I. A., & Humphreys, G. W. (2008).The effect of action goal hierarchy on the coding of objectorientation in imitation tasks: Evidence from patients withparietal lobe damage. Cognitive Neuropsychology, 25,1011–1026.
Chong, T. T.-J., Cunnington, R., Williams, M. A., Kanwisher,N., & Mattingley, J. B. (2008). fMRI adaptation revealsmirror neurons in human inferior parietal cortex. CurrentBiology, 18, 1576–1580.
12 C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding?
Author’s personal copy (e-offprint)
Experimental Psychology 2013 � 2013 Hogrefe Publishing

De Jong, R. (1995). Strategical determinants of compatibilityeffects with task uncertainty. Acta Psychologica, 88, 187–207.
Ferrari, P. F., Bonini, L., & Fogassi, L. (2009). From monkeymirror neurons to primate behaviours: Possible ‘‘direct’’ and‘‘indirect’’ pathways. Philosophical Transactions of theRoyal Society of London, Series B: Biological Sciences,364, 2311–2323.
Gleissner, B., Meltzoff, A. N., & Bekkering, H. (2000).Children’s coding of human action: Cognitive factorsinfluencing imitation in 3-year-olds. Developmental Science,3, 405–414.
Grecucci, A., Koch, I., & Rumiati, R. I. (2011). The role ofemotional context in facilitating imitative actions. ActaPsychologica, 138, 311–315.
Heister, G., Schroeder-Heister, P., & Ehrenstein, W. H. (1990).Spatial coding and spatio-anatomical mapping: Evidence fora hierarchical model of spatial stimulus-response compati-bility. In R. W. Proctor & T. G. Reeve (Eds.), Stimulus-response compatibility: An integrated perspective (pp. 117–143). Amsterdam, The Netherlands: North-Holland.
Heyes, C. (2011). Automatic imitation. Psychological Bulletin,137, 463–483.
Hommel, B. & Prinz, W. (Eds.). (1997). Theoretical issuesin stimulus-response compatibility. Amsterdam, TheNetherlands: Elsevier.
Iacoboni, M. (2005). Neural mechanisms of imitation. CurrentOpinion in Neurobiology, 15, 632–637.
Jansson, E., Wilson, A. D., Williams, J. H., & Mon-Williams,M. (2007). Methodological problems undermine tests of theideo-motor conjecture. Experimental Brain Research, 182,549–558.
Kilner, J. M., Neal, A., Weiskopf, N., Friston, K. J., & Frith,C. D. (2009). Evidence of mirror neurons in human inferiorfrontal gyrus. The Journal of Neuroscience, 29, 10153–10159.
Leighton, J., & Heyes, C. (2010). Hand to mouth: Automaticimitation across effector systems. Journal of ExperimentalPsychology: Human Perception and Performance, 36,1174–1183.
Liepelt, R., Prinz, W., & Brass, M. (2010). When do we simulatenon-human agents? Dissociating communicative and non-communicative actions. Cognition, 115, 426–434.
Liepelt, R., Von Cramon, D. Y., & Brass, M. (2008). What ismatched in direct matching? Intention attribution modulatesmotor priming. Journal of Experimental Psychology:Human Perception and Performance, 34, 578–591.
Longo, M. R., Kosobud, A., & Bertenthal, B. I. (2008).Automatic imitation of biomechanically possible andimpossible actions: Effects of priming movements versusgoals. Journal of Experimental Psychology: HumanPerception and Performance, 34, 489–501.
Mukamel, R., Ekstrom, A. D., Kaplan, J., Iacoboni, M., & Fried,I. (2010). Single-neuron responses in humans during execu-tion and observation of actions. Current Biology, 20, 750–756.
Press, C., Bird, G., Walsh, E., & Heyes, C. (2008). Automaticimitation of intransitive actions. Brain and Cognition, 67,44–50.
Proctor, R. W., & Vu, K.-P. L. (2006). Stimulus-responsecompatibility principles: Data, theory, and application.Boca Raton, FL: CRC Press.
Riggio, L., Gawryszewski, L. G., & Umilt�, C. (1986). What iscrossed in crossed-hand effects? Acta Psychologica, 62, 89–100.
Rubichi, S., Vu, K.-P. L., Nicoletti, R., & Proctor, R. W. (2006).Spatial coding in two dimensions. Psychonomic Bulletin &Review, 13, 201–216.
Van Elk, M., van Schie, H. T., & Bekkering, H. (2008).Conceptual knowledge for understanding other’s actions isorganized primarily around action goals. ExperimentalBrain Research, 189, 99–107.
Vu, K.-P. L., & Proctor, R. W. (2004). Mixing compatible andincompatible mappings: Elimination, reduction, andenhancement of spatial compatibility effects. The QuarterlyJournal of Experimental Psychology Section A, 57, 539–556.
Wild, K. S., Poliakoff, E., Jerrison, A., & Gowen, E. (2010).The influence of goals on movement kinematics duringimitation. Experimental Brain Research, 204, 353–360.
Wohlschl�ger, A., & Bekkering, H. (2002). Is human imitationbased on a mirror-neuron system? Experimental BrainResearch, 143, 335–341.
Wohlschl�ger, A., Gattis, M., & Bekkering, H. (2003). Actiongeneration and action perception in imitation: An instance ofthe ideomotor principle. Philosophical Transactions of theRoyal Society of London, Series B, Biological Sciences, 358,501–515.
Received November 23, 2011Revision received October 15, 2012Accepted October 18, 2012Published online February 21, 2013
Claudia Chiavarino
Department of PsychologyCenter for Cognitive ScienceUniversity of TurinVia Po 1410123 TurinItalyTel. +39 011 6703068Fax +39 011 8146231E-mail [email protected]
C. Chiavarino et al.: Automatic Imitation: Goal or Movement Coding? 13
Author’s personal copy (e-offprint)
� 2013 Hogrefe Publishing Experimental Psychology 2013