invasive animals in marshes
TRANSCRIPT

41
Relative to other marine systems, salt marshes and estuaries are highly susceptible toinvasion, and impacts by exotic species in these systems seem particularly pronounced.These impacts range from purely trophic and competitive effects that can lead toreplacement of native species by exotics, to physical transformation by exotic speciesthat engineer habitat and alter large-scale abiotic and hydrographic properties of themarsh environment. I discuss several examples of each of these, as well as threemechanisms that promote high establishment rates and strong competitive effects ofnonnative species in marshes.
Although the problem of exotic species in marsh systems is substantial, marshes’tractable, discrete boundaries make intervention more successful than in other marinesystems, such as the open coast. However, protocols and policies (ideally standardizedat a national or international level) need to be in place for eradication or containmentof incipient invasions, which often require fast action to be effective. Protocols andmonitoring efforts should aim not only to detect newly introduced species but also toquantify the dynamics and impacts of established invaders to enable prioritization ofintervention efforts. Estuaries and associated marshes are a heavily invaded habitatthat must be well managed to mitigate the increasing ecological impacts of exoticspecies on native species and the valuable ecosystems services they provide.
3
Invasive Animals in Marshes
biological agents of change
James E. Byers
Salt marshes are one of the most anthropogeni-cally impacted marine ecosystems (Cairnes 1993;Kennish 2001; Nicholls 2004). Historically, phys-ical and chemical impacts to salt marshes havebeen the most conspicuous and influential.Physical changes include extensive habitat con-version (e.g., filling and dredging) and altered hy-drography that stem, especially in recent times,from marshes occupying valuable coastal real
estate. Kennish (2001) calculates that more than50 percent of original tidal salt marsh in theUnited States has been lost through such physicalalterations. Chemical impacts on marshes largelyresult from the close proximity of this marinehabitat to humans and the associated pollutantsthey produce. That is, salt marshes are typicallythe outlets for watersheds where large volumes ofanthropogenic pollutants are deposited (Fox et al.
Silliman_ch03.qxd 2/23/09 1:20 PM Page 41

1999; Sanger, Holland, and Scott 1999; Hollandet al. 2004). Furthermore, as the large number ofmarsh Environmental Protection Agency (EPA)Superfund sites attests, marshes historically havedirectly received large quantities of contaminantsbecause they were perceived as a convenient, val-ueless dumping ground.
While these physical and chemical impactshave affected marsh biota for decades, only rela-tively recently, with the surge in globalizationand consequent transport of nonnative species,have biological forces themselves been recog-nized as major agents of change (e.g., Ruiz et al.1999). By skewing optimal environmental con-ditions away from conditions to which nativespecies are adapted, many physical and chemi-cal impacts may have set the stage for morefrequent and successful biological invasions.Biological impacts of nonnative species rangefrom purely trophic and competitive effects thatcan lead to replacement of native species byexotics, to physical transformation by exoticspecies that engineer habitat and alter large-scale abiotic and hydrographic properties of the marsh environment. Perhaps the best-documented example of the severe biologicalimpacts possible from invasive species is SanFrancisco Bay, where more than 240 nonindige-nous species now reside, and 90 to 95 percent ofthe biomass is exotic in many areas of the bay(Cohen and Carlton 1998; Lee, Thimpson, andLowe 2003). Clearly, to fully understand modernimpacts on both the function and the taxonomiccomposition of estuaries and salt marshes, im-pacts of exotic species must be considered.
Relative to other marine systems, saltmarshes and estuaries are highly susceptible toinvasion, and impacts by exotic species in thesesystems seem particularly pronounced. I beginthis chapter by discussing underlying proper-ties of estuaries that may be important in ex-plaining these patterns. I then highlight someprominent impacts driven by invasive animalsin salt marshes, using examples of nonindige-nous species that exert large engineering effectsand ones whose impacts are limited to trophicor competitive effects. I briefly conclude with
recommendations for how to best monitor andmanage salt marshes against these threats.Although the focus of this chapter is animalinvasions, most of the messages here are ger-mane to invasions of all types.
ESTUARIES, INCLUDING THEIRASSOCIATED MARSHES, ARE THE MOSTINVADED MARINE HABITAT
Compared to open coasts, a much higher num-ber and proportion of exotic species are foundin embayments, including associated marshes(see, e.g., Ruiz et al. 1997, 2000; Reise, Gollash,and Wolff 2002; Nehring 2002). For example, inElkhorn Slough, California, Wasson, Fenn, andPearse (2005) documented 526 invertebratescomprised of 443 natives, 58 exotics, and 25cryptogens (species whose geographic origin isuncertain). The surrounding rocky intertidalopen coast contained 588 species, of which only8 were exotic and 13 cryptogenic. The numberand proportion of exotics were significantlyhigher in the estuary (11 percent) than on the ad-jacent coast (1 percent). Furthermore, exoticspecies in the estuary were not only more di-verse but also more abundant and conspicuousthan on the open coast (e.g., the mudsnailBatillaria attramentaria, the orange spongeHymeniacidon, the reef-building tube wormFicopomatus enigmaticus). Similarly, while morethan 240 nonnative species are known from SanFranscico Bay, fewer than 10 are found on theadjacent outer coast (Ruiz et al. 1997).
Perhaps even more enigmatic is the observa-tion that in many cases, even species that aretypically open coast residents in their nativehabitats often remain in embayments andmarshes in their introduced range (Griffiths2000; Robinson et al. 2005; Wasson et al.2005). For example, the snail Littorina saxatilis,which is almost exclusively coastal in easternNorth America and Europe where it is native,has not left the confines of San Francisco Baywhere it has been established for more than adecade (Carlton and Cohen 1998; W. Miller,personal communication).
42 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
Silliman_ch03.qxd 2/23/09 1:20 PM Page 42

Clearly this pattern of a higher number ofinvaders in marshes and estuaries may be duein part to a sampling bias. Embayments andmarshes are located nearshore in close proxim-ity to humans, who as a result have examinedthese habitats more than most other marinehabitats (e.g., Ruiz et al. 1997, 2000; Cohen andCarlton 1998; Hewitt et al. 1999). In addition,compared to open marine systems, saltmarshes have discrete boundaries and a highproportion of benthic, tractable species.
However, three general, spatially variable fac-tors likely contribute to a true pattern of highernumbers of introduced species in estuaries andmarshes. First, bays and their associatedmarshes receive vast quantities of exotic propag-ules from ballast water. In U.S. ports alone, tensof millions of metric tons of ballast water fromports of origin all over the world are dischargedyearly, with each liter containing up to tenzooplankton organisms (Verling et al. 2005).Second, bays are retention zones where larvaeoften are not advected away. Byers and Pringle(2006) demonstrated that advection typical ofopen coastlines makes retention, and thus estab-lishment, difficult and may be largely responsi-ble for the dearth of invasive species in thoseenvirons. Third, marsh and estuarine habitatsbest match the habitat from which most non-indigenous species propagules are exported. Twoof the biggest vectors for nonindigenous marinespecies are ballast water and aquaculture, partic-ularly shellfish imports (Ruiz et al. 2000). Thesesources most often originate from estuaries andbays, and the similarity of source and recipienthabitats, especially in the case of intentionally in-troduced oysters and their associated communi-ties (Ruesink et al. 2005), results in a high rate ofsuccessful establishment.
EXOTIC SPECIES MAY HAVE GREATERIMPACTS IN ESTUARIES AND MARSHESTHAN OTHER MARINE HABITATS
Having more nonindigenous species in estuar-ies and marshes increases not only their cumu-lative impact but also the odds of having species
with particularly high per capita impact. Thatis, having more exotic species essentially cre-ates a sampling effect, whereby more speciessimply means species with greater impacts havea higher probability of being among thoseestablished.
However, even after we standardize for thenumber of introduced species, we may still ex-pect marshes to experience larger impacts fromexotic species for at least two additional reasons.First, as emphasized previously (and through-out this book in general), salt marshes are oneof the most anthropogenically disturbed habi-tats. Pollutants, eutrophication, habitat filling,drainage, dredge spoils dumping, and channel-ization are a few of the many, often severe abi-otic alterations humans have imposed on thishabitat type (e.g., Kennish 1992, 2001; Valiela,Rutecki, and Fox 2004; see also chaps. 8 and 9,this volume). The novel and sustained environ-mental changes that anthropogenic distur-bances impose may often be enough to move aspecies out of the parameter space that definedits evolutionary history and to which it wasadapted—a process dubbed selection regimemodification (SRM) (Byers 2002a). A nativemarsh species may therefore suddenly find it-self in an environment that in key ways is just asnovel as it is to a nonindigenous species (Byers2002a). SRM can thus accentuate competitiveimpacts of exotics on natives by eliminating anative species’ prior resident effect or “homecourt advantage.” That is, disturbances increaseinvader establishment and impact not onlyby creating new microhabitats, introducingpropagules, and decreasing populations ofnative species that can resist invasion, but alsoby potentially weakening the per capita ability ofthe native biota to resist invaders. Becausemarshes are usually heavily altered by humans,they are a prime environment for selectionregime modification and thus large resultantimpacts through competition with exoticspecies (Byers 2000b, 2002a).
Second, the retentive environment thatcontributes to increased exotic establishmentin marshes also likely enhances their
i n v a s i v e a n i m a l s i n m a r s h e s 43
Silliman_ch03.qxd 2/23/09 1:20 PM Page 43

population-level competitive effects (Byers andPringle 2006; Byers 2009). Within marshes,populations tend to be closed, and impacts ofexotic species on natives may therefore beintensified. Specifically, because there is tightcoupling of adults and successive generations,local, density-dependent impacts of exoticspecies may directly effect a lower populationgrowth rate of the resident population. Theseparation between disparate marshes along acoast helps ensure their insulation and provideslittle chance that a declining native populationwill receive a rescue effect from an immigrationpulse.
Although the closed nature of salt marshand estuarine habitats increases the propensityfor high competitive impacts, it also has positiveimplications. This tendency toward closed pop-ulations should make invasive species thereeasier to control, mitigate, or eradicate. In fact,nearly all marine eradications are done inembayments. For example, Anderson (2005)describes the successful response of variousgovernment and nonprofit agencies respondingto incipient invasions of Caulerpa taxifolia in asmall southern California lagoon. Similarly,Hutchings, Hilliard, and Coles (2002) discussthe discovery of a problematic invasive mussel,Mytilopsis sallei, in two harbors near Darwin,Australia. The harbors were quarantined, andlarge doses of sodium hypochlorite and coppersulfate were added to poison the mussel. Theprocess successfully eliminated Mytilopsis de-spite densities of the mussel that had alreadyreached ten thousand square meters (Kuris2003). In contrast, few if any eradication at-tempts on the open coast have been reported,except for one that was successful (Culver andKuris 2000). An established population ofthe African shell-boring sabellid polychaete,Terebrasabella heterouncinata, was eliminatedfrom a coastal site in California by removing1.6 million potential snail hosts in the infectedarea (Culver and Kuris 2000). Although eradi-cation was successful, it was only possible dueto an extremely anomalous, localized distribu-tion of the invader.
EXAMPLES OF PROBLEMATICESTUARINE/MARSH ANIMAL INVADERS
Before we delve into some examples, it is worthstating that the depiction and assessment of im-pacts by nonindigenous species in marshes (aswell as most every ecosystem) is likely conserv-ative. Although there is a growing body of re-search on impacts of marine invaders (e.g.,Grosholz 2002), many early invasions occurredwith little notice. Presumably many of theirimmediate and pronounced impacts could haveoccurred long ago (Cohen and Carlton 1998),setting a different baseline for benchmarkingmodern-day changes (Dayton et al. 1998). A fur-ther issue that compounds the difficulty of as-sessing impact is the problem of cryptogenicspecies, species whose definitive native geo-graphic distribution is unknown. For example,Ruiz et al. (2000) tallied 298 exotic invertebrateand algal species in coastal and estuarine watersof North America; however, this figure excludescryptogenic species, hundreds of which may infact be exotic species that just have not beenidentified as such. Even if impacts by suchcryptogenic species are observed, the impactscannot be definitively ascribed to an exoticspecies. Carlton (1996) estimates that 37 percentof the total number of known exotic and crypto-genic species in San Francisco Bay are crypto-genic; in Chesapeake Bay, this percentage is farhigher. Robinson et al. (2005) report that SouthAfrica has ten marine species that are confirmedas exotic and twenty-two as cryptogenic.
As in the ecological literature, the taxa onwhich my examples are focused are better stud-ied because they are larger and economically orculturally important. The invasion history ofmany fish, crustaceans, and mollusks, for ex-ample, is typically well known due to their im-portance as a human food source (and formollusks also their interest to early shell collec-tors). Ray (2005b) calculates that in the PacificNorthwest and Alaska, 47 of the 162 exotic ma-rine and estuarine animal species are mollusksand 39 are crustaceans, thus combining for overhalf of the total. Similarly, Ruiz et al. (2000)
44 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
Silliman_ch03.qxd 2/23/09 1:20 PM Page 44

determine that half of the identified exotic in-vertebrate and algae species in coastal and estu-arine waters North America are mollusks andcrustaceans. Nonnative fish species are also alarge contributor (Ruiz et al. 2000; Ray 2005a).For these species, it is not uncommon to havesome background data on their invasion historyand sometimes even their population dynam-ics. In contrast, many microorganisms, includ-ing diatoms, protozoans, and fungi, are easilytransported unintentionally by humans, yet lit-tle historic record exists of these organismsbecause early natural historians seldom had theinterest, let alone the expertise to identify them.Even today, many of these taxa are overlooked.Thus, the ease of incidental introduction andthe likelihood of invasion detection are in-versely correlated (Ruiz et al. 2000). Therefore,as a preface to the following examples, it is im-portant to keep in mind that we know littleabout the impacts from what are likely to be themost common invaders.
The examples that follow are organized bytype of impact: invasive ecosystem engineersthat change the structural character of thesystem and other invasive species that changecompetitive or trophic relationships betweenspecies. While there is a growing list ofproblematic, conspicuous invaders of NorthAmerican estuaries and marshes (e.g., Chinesemitten crab Eriocheir sinensis, in San FranciscoBay [Rudnick et al. 2003]; Atlantic ribbed mus-sel Guekensia demissa, in California and BajaMexico [Torchin, Hechinger, et al. 2005]), I haveselected examples to illustrate some of the best-documented, quantified impacts.
PROMINENT EXAMPLES OF EXOTIC
ECOSYSTEM ENGINEERS
Organisms that create, modify, or destroy struc-ture often disproportionately affect the commu-nities they invade. These so-called ecosystemengineers (Jones, Lawton, and Shachak 1994)essentially alter the entire playing field onwhich ecological interactions take place bychanging habitat structure, refuge availability,and even abiotic processes (e.g., hydrography)
(Wright and Jones 2006; Crain and Bertness2006). Especially in urban areas, wheremarshes are often already small fractions oftheir original extent, any habitat conversion byecosystem engineers can alter a substantialproportion of remaining marsh habitat. In ter-restrial realms, ecosystem engineers are pre-dominantly plant species, whereas in marineenvironments, it is largely animals that fill thisrole (e.g., oysters, coral, tube worms) with occa-sional contributions from vegetation (e.g.,macroalgae) (Crooks 2002). As interfaces be-tween terrestrial and marine environments, saltmarshes have a mix of plant and animal engi-neers. Traditionally, the focus in marshes hasbeen on the structural changes caused by intro-duced angiosperms (see chapters on Spartinaand Phragmites in this volume), but plenty of in-vasive animal species are causing structuralchange as well. Examples of organisms thathave been shown to be important habitatengineers include oysters, the tube wormFicopomatus enigmaticus, the mussel Musculistasenhousia, the burrowing isopod Sphaeromaquoyanum, and nutria (Myocastor coypus). Thesespecies engineer habitat in their native range aswell; however, in a novel environment wheretheir habitat structure arises de novo or wherethere are few checks on their abundance andthus the scale or rate at which habitat is altered,they can strongly affect native biota that do notshare a common evolutionary history with theinvasive engineer (Crooks 2002).
FICOPOMATUS ENIGMATICUS
Ficopomatus enigmaticus is a cosmopolitan,reef-building serpulid polychaete introduced tomany estuaries worldwide, including marshesof California and coastal lagoons of Argentina(fig. 3.1). In Mar Chiquita Lagoon, Argentina,reefs composed of thousands of calcareoustubes of this species cover roughly 80 percent ofthe lagoon. Reefs, which can be up to 4 metersin diameter and 0.5 meter high, increase habitatstructure, modify the abundance of species thatuse it for shelter, and change the pattern of dis-tribution of soft-bottom species (Schwindt and
i n v a s i v e a n i m a l s i n m a r s h e s 45
Silliman_ch03.qxd 2/23/09 1:20 PM Page 45

Iribarne 2000). The reefs built by Ficopomatusin Mar Chiquita also greatly affect the physicalenvironment by altering the bedload sedimenttransport and water flow (Schwindt, Iribarne,and Isla 2004). In general, ecosystem engineer-ing species, such as F. enigmaticus, that invadesoft sediment environments and create hardsubstrate tend to have large impacts on commu-nity composition. This is because hard sub-strata are novel to most marshes, and thus theyoften provide exotic species equal opportunityto compete for space because native marshspecies are not specifically adapted to this habi-tat type. Thus, transformation of soft bottom es-tuarine habitat into hard substratum is a primeexample of selection regime modification, andit has been found to be associated with in-creases in invasive species (Wasson et al. 2005;Tyrrell and Byers 2007).
MUSCULISTA SENHOUSIA
Several decades ago, this Asian mytilid musselinvaded marshes and associated mudflats ofMission Bay (San Diego), San Francisco Bay,and Puget Sound among others along thePacific coast of North America (Crooks 1998).
The mussel anchors itself through byssal matsit produces that dramatically alter soft sedimenthabitats. Crooks (1998) noted marked changesto sedimentary properties and to the residentbiota. Densities of macrofauna as well asspecies richness were typically higher insidethan outside mussel mats. A tanaid amphipod,Leptochelia dubia, and the gastropod Barleeiasubtenuis were particularly enhanced withinmussel mats. In contrast, the abundance ofnative, filter-feeding clams declined, possiblybecause of competition for food (Crooks 2001).Through a series of experiments that comparedcommunity effects of live mussel mats to struc-tural mimics, Crooks and Khim (1999), demon-strated that the physical structure of themussels far outweighed the effects of livingmussels. In fact, they determined specificallythat the structure provided by the mussel matswas more influential than the mussel shells.In general, the effects of Musculista andFicopomatus agree with observed effects of otherhabitat-modifying exotic ecosystem engineers,illustrating dramatic effects on biota by non-native species capable of creating and alteringphysical structure.
46 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
FIGURE 3.1 Ficopomatus enigmaticus reefs in Argentina. Photo courtesy of MartinBruschetti.
Silliman_ch03.qxd 2/23/09 1:20 PM Page 46
SPHAEROMA QUOYANUM
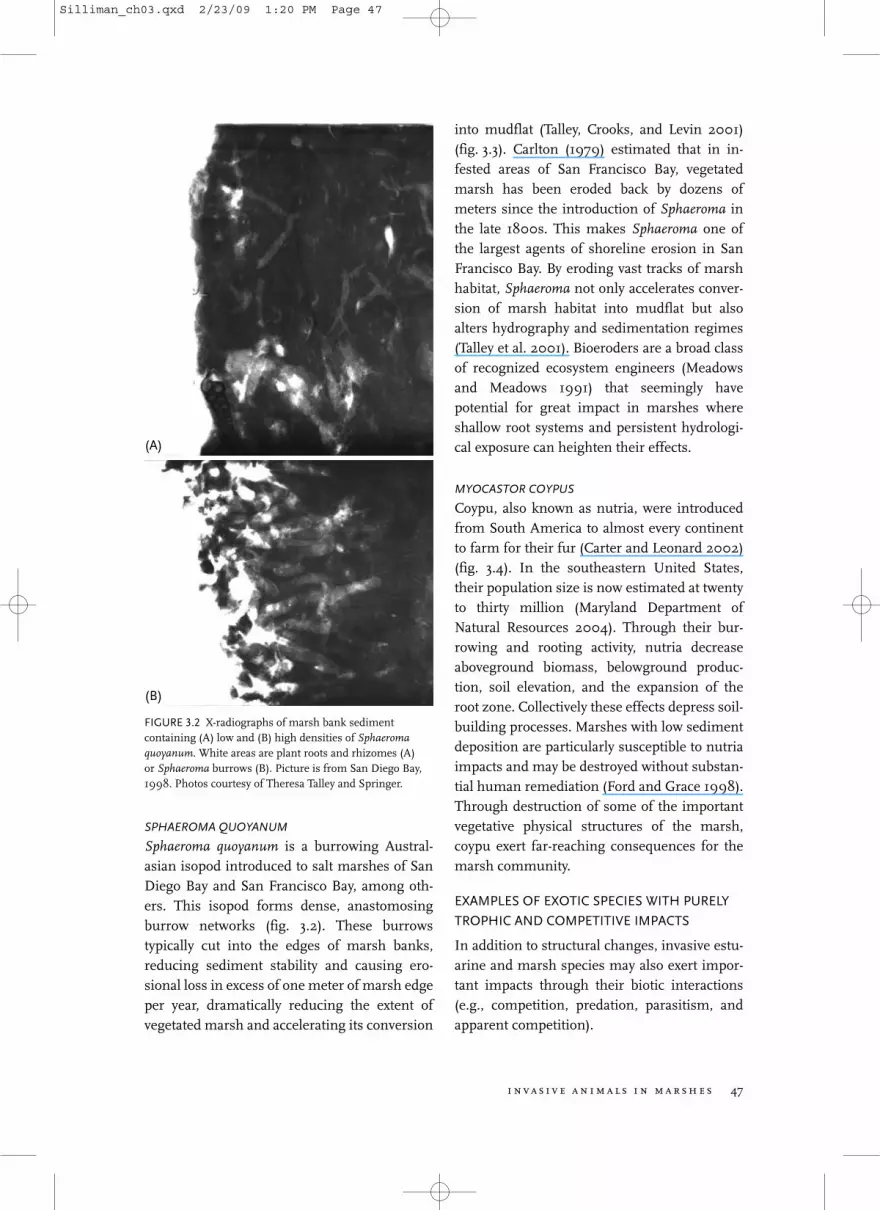
Sphaeroma quoyanum is a burrowing Austral-asian isopod introduced to salt marshes of SanDiego Bay and San Francisco Bay, among oth-ers. This isopod forms dense, anastomosingburrow networks (fig. 3.2). These burrowstypically cut into the edges of marsh banks,reducing sediment stability and causing ero-sional loss in excess of one meter of marsh edgeper year, dramatically reducing the extent ofvegetated marsh and accelerating its conversion
into mudflat (Talley, Crooks, and Levin 2001)(fig. 3.3). Carlton (1979) estimated that in in-fested areas of San Francisco Bay, vegetatedmarsh has been eroded back by dozens ofmeters since the introduction of Sphaeroma inthe late 1800s. This makes Sphaeroma one ofthe largest agents of shoreline erosion in SanFrancisco Bay. By eroding vast tracks of marshhabitat, Sphaeroma not only accelerates conver-sion of marsh habitat into mudflat but alsoalters hydrography and sedimentation regimes(Talley et al. 2001). Bioeroders are a broad classof recognized ecosystem engineers (Meadowsand Meadows 1991) that seemingly havepotential for great impact in marshes whereshallow root systems and persistent hydrologi-cal exposure can heighten their effects.
MYOCASTOR COYPUS
Coypu, also known as nutria, were introducedfrom South America to almost every continentto farm for their fur (Carter and Leonard 2002)(fig. 3.4). In the southeastern United States,their population size is now estimated at twentyto thirty million (Maryland Department ofNatural Resources 2004). Through their bur-rowing and rooting activity, nutria decreaseaboveground biomass, belowground produc-tion, soil elevation, and the expansion of theroot zone. Collectively these effects depress soil-building processes. Marshes with low sedimentdeposition are particularly susceptible to nutriaimpacts and may be destroyed without substan-tial human remediation (Ford and Grace 1998).Through destruction of some of the importantvegetative physical structures of the marsh,coypu exert far-reaching consequences for themarsh community.
EXAMPLES OF EXOTIC SPECIES WITH PURELY
TROPHIC AND COMPETITIVE IMPACTS
In addition to structural changes, invasive estu-arine and marsh species may also exert impor-tant impacts through their biotic interactions(e.g., competition, predation, parasitism, andapparent competition).
i n v a s i v e a n i m a l s i n m a r s h e s 47
FIGURE 3.2 X-radiographs of marsh bank sedimentcontaining (A) low and (B) high densities of Sphaeromaquoyanum. White areas are plant roots and rhizomes (A)or Sphaeroma burrows (B). Picture is from San Diego Bay,1998. Photos courtesy of Theresa Talley and Springer.
(A)
(B)
Silliman_ch03.qxd 2/23/09 1:20 PM Page 47

BATILLARIA ATTRAMENTARIA
An exotic snail, Batillaria attramentaria, hassuccessfully invaded several salt marshes andmud flats in northern California, Washington,and British Columbia (Byers 1999) (fig. 3.5). InCalifornia, populations of the native mud snail,Cerithidea californica, have declined precipi-tously. Experimental manipulations demon-strated that the snail species competedexploitatively for epipelic diatoms colonizingthe surface of the marsh mud. Although the twospecies did not differ in their effect on resourcelevels at any experimental snail density, theintroduced snail was always more efficient at
48 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
FIGURE 3.3 (A) Extensiveburrowing by Sphaeromaquoyanum in vertical marshbanks, San Diego Bay. (B) Such burrowing loosenssediment, increasing erosion,undercutting banks, andreleasing chunks of marshsurface and reducing marshhabitat. Corte Madera Marsh,San Francisco Bay. Photoscourtesy of Theresa Talley andSpringer.
converting limited resources to tissue growth.Batillaria’s enhanced resource conversion effi-ciency provides a sufficient explanation for itssuccessful invasion and subsequent exclusionof Cerithidea (Byers 2000a). Byers andGoldwasser (2001) subsequently combinedthese detailed, quantitative field data onBatillaria and its interactions with Cerithidea inan individual-based model. In empirically para-meterized simulations, the native snail was dri-ven extinct within fifty-five to seventy years afterthe introduction of Batillaria, closely matchingdirect field estimates of Cerithidea’s time to dis-placement (Byers 1999).
(A)
(B)
Silliman_ch03.qxd 2/23/09 1:20 PM Page 48

Replacement of one mud snail for anothermight seem like a superficial biotic change.However, because of its superior resource con-version efficiency, Batillaria achieves higher den-sities than Cerithidea, thus suppressing thediatom standing stock to very low levels. Such re-source depression is likely to dramatically affectother benthic grazers within the marsh, such asghost shrimp and other snails. But the largestimpact stemming from the replacement ofnative Cerithidea was uncovered only recently.
Both Batillaria and Cerithidea are first inter-mediate hosts for trematode parasites. All butone of these trematode species must obligately
pass through a second intermediate host andthen on to a final host during their life cycles.Depending on the trematode species, mollusks,crustaceans, or fishes may serve as second in-termediate hosts for the metacercarial cysts.The trematode life cycle is completed when asecond intermediate host is eaten by the finalhost, typically a shorebird. Cerithidea californicahosts at least eighteen native trematode speciesthroughout its range in California (Martin1972). Batillaria, however, hosts just a singletrematode species—itself a nonnative species(Cercaria batillariae) (Torchin, Byers, andHuspeni 2005; fig. 3.6). In Elkhorn Slough,California, hundreds of cysts of C. batillariaewere found in all individuals of the three fishspecies examined, including within physiologi-cally sensitive regions like the pericardium.
Because trematode parasites are highlyspecific for the species of snail they infect, inmarshes where C. californica becomes extir-pated by Batillaria, Cerithidea’s parasites willalso become locally extinct. The removal of
i n v a s i v e a n i m a l s i n m a r s h e s 49
FIGURE 3.4 Nutria (Myocastor coypus) in marsh. Photocourtesy of Guerry O’Holm.
FIGURE 3.5 The exotic mudsnail, Batillaria attramentaria(the two snails pictured on the right), has successfullyinvaded several salt marshes and mud flats in northernCalifornia, where it contributes to the exclusion of thenative confamilial mudsnail, Cerithidea californica (the two snails pictured on the left). Photo by Jeb Byers andJohn Meyer.
FIGURE 3.6 Cercaria of the multihost trematode parasite,Cercaria batillariae. This nonnative trematode is found inmarshes along the Pacific coast of North America where itinfects only a single species of first intermediate host—thenonnative mudsnail, Batillaria attramentaria. The cercariaeare released from infected snails and penetrate into nativeestuarine fish as a second intermediate host. Length ofcercariae is about 0.5 millimeter. Photo courtesy of ToddHuspeni.
Silliman_ch03.qxd 2/23/09 1:20 PM Page 49

multiple native trematode species will ulti-mately result in local eliminations of trematodeinfections in mollusks, crustaceans, and poten-tially several fishes that serve as second inter-mediate hosts for C. californica’s parasites(Martin 1972). Although the exact manifesta-tions are still unclear, the reduction in infec-tions will likely alter host population dynamics(Lafferty 1992) and potentially the metabolism,foraging, and abundance of shorebirds as well(Lafferty and Morris 1996). Thus, the most far-reaching effects of Cerithidea’s loss on the com-munity structure of the native marsh systemseems very likely to stem from the concomitantloss of its dependent, trophically transmittedtrematode species.
VENERUPIS PHILIPPINARUM
Even in typically closed marsh populations,not every species exerts competitive effects(Byers 2009). Filter (suspension) feedingspecies in particular have been suggested toexperience reduced competition because theirplanktonic food source is often not a limitingfactor (Levinton 1972; Peterson 1979; Byers2009). A nonindigenous suspension-feedingspecies, Venerupis philippinarum (� Ruditapesphilippinarum, � Tapes japonica), the Japaneselittleneck clam, was accidentally introducedinto the eastern Pacific in the 1930s withimported oyster seed from Japan and is nowfound in low-energy embayments and marsh-associated mudflats from British Columbia tosouthern California (Quayle 1941; Haderlieand Abbott 1980). It is the most prolific intro-duced clam species in the San Juan Islands,Washington, and accounts for 50 percent ofthe annual commercial landings of hard-shellclams in the state (Washington Department ofFish and Wildlife 2000).
Byers (2005) experimentally examined theeffects of high Venerupis densities on mortality,growth, and fecundity of the confamilial clam,Protothaca staminea, and whether differences inpredator abundance mitigate density dependenteffects. Even at densities 50 percent higher thanthose found naturally in the field, Venerupis had
no direct effect on itself or Protothaca. Rather,the variable of overwhelming influence on theclams was crab biomass, which decreasedgrowth of both species and increased mortalityof Venerupis. The annualized loss rate ofVenerupis was 50 percent when exposed to exca-vating crab predators due to a very shallowburial depth, nearly three centimeters shallowerthan Protothaca (Byers 2005). In fact, when ex-posed to predators, Venerupis was up to seventimes more likely to be taken than Protothaca,whose mortality did not differ significantly withcrab abundance.
By taking the brunt of predator pressure,Venerupis seems to play a sacrificial role that atleast partially protects Protothaca from predatormortality. However, high consumption ofVenerupis likely has negative effects on otherinvertebrate species. When an exotic species isconsumed by a native predator, losses to theexotic population are converted to additionalpredator biomass. Hence, predation on an ex-otic species can indirectly harm the predator’snative prey via apparent competition (Rand andLouda 2004). Noonburg and Byers (2005)demonstrate conditions under which resourcecompetition from an invasive species is lessdetrimental to a native consumer than in-creased losses to a native predator populationthat is boosted by the invader.
Thus, although no direct competitive effectof Venerupis on Protothaca was detected, byserving as an easy prey source for crabs, thisnonindigenous prey species may be boostingregional crab abundance and productivity.Given the ubiquity of Venerupis throughout theWest Coast of North America, this crab foodsubsidy could be substantial. In addition, thethin-shelled, nonindigenous Nuttallia obscurata,present in high abundance throughout the U.S.Northwest and British Columbia coast, is alsoan easy, novel prey item for crabs when it occursin areas without appropriate physical refuges(Byers 2002b). Because Cancer crabs are omniv-orous predators, their increase (particularly theless harvested species, C. gracilis and C. produc-tus) potentially affects many other native prey
50 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
Silliman_ch03.qxd 2/23/09 1:20 PM Page 50

species, including worms, fish, crustaceans,and other bivalves. Subsidies of native predatorsmay be a largely underappreciated means bywhich nonindigenous species that are con-sumed heavily by native predators enhance ap-parent competition and thus escalate the impactof predators on native species (Courchamp,Langlais, and Sugihara 2000; Byers 2002b,2005; Noonburg and Byers 2005).
CARCINUS MAENAS
Due to inadvertent human transport, theEuropean green crab, Carcinus maenas, is now acosmopolitan species (Geller et al. 1997;Hidalgo. Baron, and Orensanz 2005). Its im-pacts have been well studied in marshes andembayments of the western coast of NorthAmerica. Due to its omnivorous diet, Carcinuswas found to significantly decrease the abun-dance of several invertebrate prey species inBodega Bay, California (Grosholz et al. 2000).Most notably, two native clams, Nutricola tan-tilla and N. confusa, and a native shorecrab,Hemigrapsus oregonensis, decreased five- to ten-fold within three years of the introduction ofCarcinus. Carcinus indirectly promoted severalpolychaete and crustacean species, which ap-parently were not major prey items but ratherbenefited from Carcinus’s removal of compet-ing species (Grosholz et al. 2000). In addition,Carcinus predation on Nutricola clam speciesfacilitated spread of an introduced competitorclam, Gemma gemma throughout BodegaHarbor (Grosholz 2005). Carcinus thus has bothdirect and indirect predatory effects.
Similarly, on the East Coast of NorthAmerica, where it has been established for overone hundred years, Carcinus has been shown tohave equally profound predatory impacts, par-ticularly on clams. In Nova Scotia, Floyd andWilliams (2004) measured 80 percent reduc-tions of small Mya arenaria within less thanfour months, with consumption rates of threeto twenty-two clams per crab per day. Whitlow,Rice, and Sweeney (2003) also measured highpredation rates of Carcinus on M. arenaria.Although these authors demonstrated that Mya
was able to partially mitigate the high predationrate from the excavating crabs by burrowingmore deeply in the sediment, deeper burialtypically decreases feeding efficiency and thusgrowth rates of clams (Zaklan and Ydenberg1997). Thus, predatory effects of Carcinus canbe both density and trait mediated.
RECOMMENDATIONS ANDCONCLUSIONS
With increasing globalization, the influx of non-indigenous species to salt marshes is not likelyto abate soon. The relatively retentive nature ofmost embayments, estuaries, and associatedmarshes seems to contribute both to higher in-vasive establishment as well as to strong com-petitive effects. Because nonindigenous speciesoften originate from estuaries and bays, thesimilarity of source and recipient habitats alsolikely contributes to a high rate of successfulestablishment. Finally, the high rate of distur-bance to marsh environments suggests thatselection regimes may be sufficiently altered topromote high establishment rates and subse-quent impact of introduced species.
Policies to reduce the supply rate of exoticpropagules, such as new mandatory ballastwater exchange during transoceanic crossings(Federal Register 2004) are extremely com-mendable, since preventing invasions in thefirst place is the most proactive policy. Butbecause this policy alone will not eliminatefuture invasions or mitigate effects of invadersalready present, it is important to maintainvigilance in marsh habitats. In the UnitedStates, the Environmental Protection Agency’sEnvironmental Monitoring and AssessmentProgram and several National Oceanographicand Atmospheric Administration subagenciessuch as Sea Grant have conducted periodicregional inventories of marshes and bays forexotic species (Cohen et al. 2001; Lee et al.2004; Pederson et al. 2005). Taking these effortsone step further to implement a standardized,national or international protocol to detect in-cipient invasions would strengthen our approach
i n v a s i v e a n i m a l s i n m a r s h e s 51
Silliman_ch03.qxd 2/23/09 1:20 PM Page 51

even more (e.g., Wasson et al. 2002). Ad hocassessments have certainly led to some invadersbeing overlooked, presenting enormous prob-lems in establishing appropriate baselines ofimpact assessment (Dayton et al. 1998) andslowing potential mitigation by resource man-agers. Protocols and policies need to be in placefor eradication or containment of incipientinvasions, which often require fast action to beeffective (e.g., Anderson 2005).
In addition to detecting introduced speciesand following the dynamics of established ones,quantitative monitoring is important because itaids restoration. Specifically, it complementsadaptive management, which through frequentdata-driven assessments can refine interventiontechniques to restore native marsh populations,habitat, and ecosystem function. Clearlyrestoration efforts should be prioritized basedon an understanding of invaders’ impact, bethey engineering or trophic effects. Because im-pact assessment can be slow, if forthcoming atall, on first approximation such prioritizationmay do best to focus on invasive habitat engi-neers (e.g., oysters, reef-building polychaetes)because of their often far-reaching and lastinglegacy effects and their ability to transform abi-otic properties of a marsh and thus alter ecosys-tem services (Byers et al. 2006).
Because not all invasive species are detri-mental to native species or systems (e.g., Brunoet al. 2005; Byers 2009), monitoring marshesfor early impact detection may be crucial forproper prioritization of intervention efforts.However, it is very important to avoid false con-fidence in the promptness of impact detection insuch monitoring programs, because exotic im-pact is sometimes virtually undetectable untilafter the exotic species is extremely abundant(Byers and Goldwasser 2001). This lag in impactmay be especially pronounced if the mechanismof impact by the exotic species is to decreasebirths of a long-lived native species—a less con-spicuous impact than increasing death of adults.In such cases, demographic lag times necessitatealternative, faster-responding metrics for impactdetection than adult population densities.
As an illustration, Byers and Goldwasser(2001) sought to identify empirically measur-able quantities that provide the earliest warningof impact by the invasive snail Batillaria on thenative Cerithidea. Through an individual-basedmodel parameterized with empirical data, theytracked many population- and individual-levelresponses of Cerithidea to Batillaria’s invasion,including population density, biomass, eggproduction, mean size, proportion of parasitizedindividuals, and individual growth rate, as wellas availability of shared food resources. In modelsimulations, the initial number of invadingBatillaria was set to guarantee extinction ofCerithidea within ninety years. Despite a rapidinitial increase in the invader population, allmetrics for Cerithidea were slow to exhibit signsof impact. Most took at least twenty-five yearsfrom invasion to exhibit detectable changes, bywhich time the exotic snail was established at ex-tremely high densities. Cerithidea egg produc-tion was the fastest, most consistent responsemetric, exhibiting declines within twenty totwenty-five years after invasion in about 90 per-cent of simulations. Monitoring programs andrisk assessment analyses must identify and con-centrate on reliable, early-warning metrics.
Habitat alteration and biotic effects by exoticspecies now join anthropogenic physical alter-ations and chemical inputs as dominantimpacts on estuarine and marsh environments.Dramatic modification of selection regimes inmarshes suggests that synergism between thesedisturbance agents enhances invasion rates andimpacts. Therefore, policies to protect marshesand estuaries from physical and chemical an-thropogenic disturbance should simultaneouslyimprove their resistance and resilience to bio-logical invasions. Although the closed nature ofmost estuarine and marsh habitats and theirpopulations increases the propensity forimpacts by nonindigenous species, it alsomakes the species easier to document, track,control, and potentially eradicate. For example,the tractable, discrete boundaries of marsh sys-tems certainly make intervention more success-ful than in other marine systems such as the
52 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
Silliman_ch03.qxd 2/23/09 1:20 PM Page 52

open coast with its porous boundaries, compli-cating currents, and an influx of propagulesfrom elsewhere. Marshes are a jeopardized,increasingly compromised habitat to which wemust pay strong attention; otherwise, we risklosing many of the native species and valuableecosystems services they provide.
Acknowledgments. I thank Irit Altman, TyraByers, John Meyer, Whitman Miller, and TedGrosholz for helpful edits and discussions onthis chapter.
REFERENCES
Anderson, L. W. J. 2005. California’s reaction toCaulerpa taxifolia: A model for invasive speciesrapid response. Biological Invasions 7: 1003–1016.
Bruno, J. E., J. D. Fridley, K. D. Bromberg, and M. D.Bertness. 2005. Insights into biotic interactionsfrom studies of species invasions. Pages 13–40 inD. F. Sax, J. J. Stachowicz, and S. D. Gaines (eds.),Species Invasions: Insights into Ecology, Evolution,and Biogeography. Sunderland, MA: Sinauer.
Byers, J. E. 1999. The distribution of an introducedmollusc and its role in the long-term demise of anative confamilial species. Biological Invasions 1:339–353.
———. 2000a. Competition between two estuarinesnails: Implications for invasions of exoticspecies. Ecology 81: 1225–1239.
———. 2000b. Differential susceptibility to hypoxiaaids estuarine invasion. Marine Ecology ProgressSeries 203: 123–132.
———. 2002a. Impact of nonindigenous species onnatives enhanced by anthropogenic alteration ofselection regimes. Oikos 97: 449–458.
———. 2002b. Physical habitat attribute mediatesbiotic resistance to nonindigenous species inva-sion. Oecologia 130: 146–156.
———. 2005. Marine reserves enhance abundancebut not competitive impacts of a harvested non-indigenous species. Ecology 86: 487–500.
———. 2009. Competition in Marine Invasions.In: Biological invasions in marine ecosystems:Ecological, management, and geographic perspec-tives. Eds.: Gil Rilov & Jeff Crooks. Springer-Verlag. pp. 245–260.
Byers, J. E., K. Cuddington, C. G. Jones, T. S. Talley, A.Hastings, J. Lambrinos, J. A. Crooks, and W. G.Wilson. 2006. Using ecosystem engineers to re-store ecological systems. Trends in Ecology andEvolution 21, no. 9: 493–500.
Byers, J. E., and L. Goldwasser. 2001. Exposing themechanism and timing of impact of nonindige-nous species on native species. Ecology 82:1330–1343.
Byers, J. E., and J. M. Pringle. 2006. Going againstthe flow: Retention, range limits and invasions inadvective environments. Marine Ecology ProgressSeries 313: 27–41.
Cairnes, J. 1993. Is restoration ecology practical?Restoration Ecology March: 3–7.
Carlton, J. T. 1979. History, biogeography, and ecol-ogy of the introduced marine and estuarine inver-tebrates of the Pacific coast of North America.Unpublished PhD diss., University of California,Davis.
———. 1996. Biological invasions and cryptogenicspecies. Ecology 77: 1653–1655.
Carlton, J. T., and A. N. Cohen. 1998. Periwinkle’sprogress: The Atlantic snail Littorina saxatilis(Mollusca: Gastropoda) establishes a colony on aPacific shore. Veliger 41: 333–338.
Carter, J., and B. P. Leonard. 2002. A review of theliterature on the worldwide distribution, spreadof, and efforts to eradicate the coypu (Myocastorcoypus). Wildlife Society Bulletin 30: 162–175.
Cohen, A., H. Berry, C. Mills, D. Milne, K. Britton-Simmons, M. Wonham, D. Secord, J. Barkas, B.Bingham, B. Bookheim, J. E. Byers, J. Chapman,J. Cordell, B. Dumbauld, A. Fukuyama, L. Harris,A. Kohn, K. Li, T. Mumford, V. Radashevsky, A.Sewell, and K. Welch. 2001. Washington state ex-otics expedition 2000: A rapid assessment surveyof exotic species in the shallow waters of ElliottBay, Totten and Eld Inlets, and Willapa Bay.Unpublished manuscript, Nearshore HabitatProgram, Washington State Department ofNatural Resources, Olympia.
Cohen, A. N., and J. T. Carlton. 1998. Acceleratinginvasion rate in a highly invaded estuary. Science279: 555–558.
Courchamp, F., M. Langlais, and G. Sugihara. 2000.Rabbits killing birds: Modelling the hyperpreda-tion process. Journal of Animal Ecology 69:154–164.
Crain, C. M., and M. D. Bertness. 2006. Ecosystemengineering across environmental gradients:Implications for conservation and management.BioScience 56: 211–218.
Crooks, J. A. 1998. Habitat alteration and community-level effects of an exotic mussel, Musculista sen-housia. Marine Ecology Progress Series 162: 137–152.
———. 2001. Assessing invader roles within chang-ing ecosystems: Historical and experimental per-spectives on an exotic mussel in an urbanizedlagoon. Biological Invasions 3: 23–36.
i n v a s i v e a n i m a l s i n m a r s h e s 53
Silliman_ch03.qxd 2/23/09 1:20 PM Page 53

———. 2002. Characterizing ecosystem-level conse-quences of biological invasions: The role ofecosystem engineers. Oikos 97: 153–166.
Crooks, J. A., and H. S. Khim. 1999. Architectural vs.biological effects of a habitat-altering, exotic mus-sel, Musculista senhousia. Journal of ExperimentalMarine Biology and Ecology 240: 53–75.
Culver, C. S., and A. M. Kuris. 2000. The apparenteradication of a locally established introducedmarine pest. Biological Invasions 2: 245–253.
Dayton, P. K., M. J. Tegner, P. B. Edwards, and K. L.Riser. 1998. Sliding baselines, ghosts, and re-duced expectations in kelp forest communities.Ecological Applications 8: 309–322.
Federal Register. 2004. Mandatory ballast water man-agement program for U.S. waters. Federal Register69: 44952–44961.
Floyd, T., and J. Williams. 2004. Impact of green crab(Carcinus maenas L.) predation on a population ofsoft-shell clams (Mya arenaria L.) in the southernGulf of St. Lawrence. Journal of Shellfish Research23: 457–462.
Ford, M. A., and J. B. Grace. 1998. Effects of verte-brate herbivores on soil processes, plant biomass,litter accumulation and soil elevation changes in acoastal marsh. Journal of Ecology 86: 974–982.
Fox, W. M., M. S. Johnson, S. R. Jones, R. T. Leah, andD. Copplestone. 1999. The use of sediment coresfrom stable and developing salt marshes to recon-struct historical contamination profiles in theMersey Estuary, UK. Marine EnvironmentalResearch 47: 311–329.
Geller, J. B., E. D. Walton, E. D. Grosholz, and G. M.Ruiz. 1997. Cryptic invasions of the crab Carcinusdetected by molecular phylogeography. MolecularEcology 6: 901–906.
Griffiths, C. L. 2000. Overview on current problemsand future risks. Pages 235–241 in G. Preston, G.Brown, and E. van Wyk (eds.), Best ManagementPractices for Preventing and Controlling InvasiveAlien Species. Capetown: Working for WaterProgramme.
Grosholz, E. D. 2002. Ecological and evolutionaryconsequences of coastal invasions. Trends inEcology and Evolution 17: 22–27.
———. 2005. Recent biological invasion may hasteninvasional meltdown by accelerating historical in-troductions. Proceedings of the National Academy ofSciences of the USA 102: 1088–1091.
Grosholz, E. D., G. M. Ruiz, C. A. Dean, K. A. Shirley,J. L. Maron, and P. G. Connors. 2000. The im-pacts of a nonindigenous marine predator in aCalifornia bay. Ecology 81: 1206–1224.
Haderlie, E. C., and D. P. Abbott. 1980. Bivalvia: theclams and allies. Pages 355–411 in R. H. Morris,
D. P. Abbott, and E. C. Haderlie (eds.), IntertidalInvertebrates of California. Stanford, CA: StanfordUniversity Press.
Hewitt, C. L., M. L. Campbell, R. E. Thresher, andR. B. Martin. 1999. Marine biological invasion ofPort Phillip Bay, Victoria. Hobart, Australia:Centre for Research on Introduced Marine Pests.
Hidalgo, F. J., P. J. Baron, and J. M. Orensanz. 2005.A prediction come true: the green crab invades thePatagonian coast. Biological Invasions 7: 547–552.
Holland, A. F., D. M. Sanger, C. P. Gawle, S. B.Lerberg, M. S. Santiago, G. H. M. Riekerk, L. E.Zimmerman, and G. I. Scott. 2004. Linkages be-tween tidal creek ecosystems and the landscapeand demographic attributes of their watersheds.Journal of Experimental Marine Biology and Ecology298: 151–178.
Hutchings, P. A., R. W. Hilliard, and S. L. Coles.2002. Species introductions and potential for ma-rine pest invasions into tropical marine commu-nities, with special reference to the Indo-Pacific.Pacific Science 56: 223–233.
Jones, C. G., J. H. Lawton, and M. Shachak. 1994.Organisms as ecosystem engineers. Oikos 69:373–386.
Kennish, M. J. 1992. Ecology of estuaries:Anthropogenic effects. Boca Raton, FL: CRC.
———. 2001. Coastal salt marsh systems in the US:A review of anthropogenic impacts. Journal ofCoastal Research 17: 731–748.
Kuris, A. M. 2003. Eradication of introduced marinepests. Pages 549–556 in D. J. Rapport, W. L.Lasley, D. E. Rolston, N. O. Nielsen, C. O. Qualset,and A. B. Damania (eds.), Managing for HealthyEcosystems. Boca Raton, FL: CRC.
Lafferty, K. D. 1992. Foraging on prey that aremodified by parasites. American Naturalist 140:854–867.
Lafferty, K. D., and A. K. Morris. 1996. Altered behav-ior of parasitized killifish increases susceptibilityto predation by bird final hosts. Ecology 77:1390–1397.
Lee, H., W. Nelson, J. Lamberson, and D. A. Reusser.2004. Regional Assessment of the InvasiveMacrobenthos in the Small West Coast Estuaries.Washington, DC: Environmental ProtectionAgency; Environmental Monitoring andAssessment Program.
Lee, H., B. Thompson, and S. Lowe. 2003. Estuarineand scalar patterns of invasion in the soft-bottombenthic communities of the San FranciscoEstuary. Biological Invasions 5: 85–102.
Levinton, J. 1972. Stability and trophic structure indeposit-feeding and suspension-feeding commu-nities. American Naturalist 106: 472–486.
54 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
Silliman_ch03.qxd 2/23/09 1:20 PM Page 54

Martin, W. E. 1972. An annotated key to the Cercariaethat develop in the snail Cerithidea californica.Bulletin of the Southern California Academy ofSciences 71: 39–43.
Maryland Department of Natural Resources. 2004.Invasive and Exotic Species: Nutria. Baltimore:Wildlife and Heritage Service.
Meadows, P. S., and A. Meadows, eds. 1991. TheEnvironmental Impact of Burrowing Animals andAnimal Burrows. Oxford: Clarendon.
Nehring, S. 2002. Biological invasions into Germanwaters: An evaluation of the importance of differ-ent human-mediated vectors for nonindigenousmacrozoobenthic species. Pages 373–383 in E.Leppakoski, S. Gollasch, and S. Olenin (eds.),Invasive Aquatic Species of Europe. Distribution,Impacts, and Management. Dordrecht: KluwerAcademic.
Nicholls, R. J. 2004. Coastal flooding and wetlandloss in the 21st century: Changes under the SRESclimate and socio-economic scenarios. GlobalEnvironmental Change—Human and PolicyDimensions 14: 69–86.
Noonburg, E. G., and J. E. Byers. 2005. More harmthan good: When invader vulnerability to preda-tors enhances impact on native species. Ecology86: 2555–2560.
Pederson, J., R. Bullock, J. T. Carlton, J. Dijkstra,N. Dobroski, P. Dyrynda, R. Fisher, L. Harris,N. Hobbs, G. Lambert, E. Lazo-Wasem,A. Mathieson, M. Miglietta, J. Smith, J. Smith III,and M. Tyrrell. 2005. Marine Invaders in theNortheast: Rapid Assessment of Nonnative MarineSpecies of Floating Dock Communities. Report ofthe August 3–9, 2003 survey. Cambridge, MA:MIT Sea Grant College Program.
Peterson, C. H. 1979. Predation, competitive exclusion,and diversity in the soft-sediment benthic commu-nities of estuaries and lagoons. Pages 233–263 inR. J. Livingston (ed.), Ecological Processes in Coastaland Marine Systems. New York: Plenum.
Quayle, D. B. 1941. The Japanese “little neck” clam ac-cidentally introduced into British Columbia wa-ters. Progress Report Fisheries Research Board ofCanada Pacific Coast Station 48: 17–18.
Rand, T. A., and S. M. Louda. 2004. Exotic weed inva-sion increases the susceptibility of native plantsattack by a biocontrol herbivore. Ecology 85:1548–1554.
Ray, G. L. 2005a. Invasive marine and estuarineanimals of the North Atlantic. 19. ERDC/TN ANSRP-05-1. http://el.erdc.usace.army.mil/ansrp/pdfs/ansrp05-6.pdf.
———. 2005b. Invasive marine and estuarineanimals of the Pacific Northwest and Alaska.
18. ERDC/TN ANSRP-05-6. http://el.erdc.usace.army.mil/ansrp/pdfs/ansrp05-6.pdf.
Reise, K., S. Gollasch, and W. J. Wolff. 2002.Introduced marine species of the North Seacoasts. Pages 260–266 in E. Leppakoski, S.Gollasch, and S. Olenin (eds.), Invasive AquaticSpecies of Europe: Distribution, Impacts, andManagement. Dordrecht: Kluwer Academic.
Robinson, T. B., C. L. Griffiths, C. McQuaid, and M.Rius. 2005. Marine alien species of South Africa:Status and impacts. African Journal of MarineScience 27: 297–306.
Rudnick, D. A., K. Hieb, K. F. Grimmer, and V. H.Resh. 2003. Patterns and processes of biological in-vasion: The Chinese mitten crab in San FranciscoBay. Basic and Applied Ecology 4: 249–262.
Ruesink, J. L., H. S. Lenihan, A. C. Trimble, K. W.Heiman, F. Micheli, J. E. Byers, and M. C. Kay.2005. Introduction of nonnative oysters:Ecosystem effects and restoration implications.Annual Review of Ecology Evolution and Systematics36: 643–689.
Ruiz, G. M., J. T. Carlton, E. D. Grosholz, and A. H.Hines. 1997. Global invasions of marine andestuarine habitats by non indigenous species:Mechanisms, extent, and consequences.American Zoologist 37: 621–632.
Ruiz, G. M., P. W. Fofonoff, J. T. Carlton, M. J.Wonham, and A. H. Hines. 2000. Invasion ofcoastal marine communities in North America:Apparent patterns, processes, and biases. AnnualReview of Ecology and Systematics 31: 481–531.
Ruiz, G. M., P. W. Fofonoff, A. H. Hines, and E. D.Grosholz. 1999. Nonindigenous species asstressors in estuarine and marine communities:Assessing invasion impacts and interactions.Limnology and Oceanography 44: 950–972.
Sanger, D. M., A. F. Holland, and G. I. Scott. 1999.Tidal creek and salt marsh sediments in SouthCarolina coastal estuaries: II. Distribution oforganic contaminants. Archives of EnvironmentalContamination and Toxicology 37: 458–471.
Schwindt, E., and O. O. Iribarne. 2000. Settlementsites, survival and effects on benthos of an intro-duced reef-building polychaete in a SW Atlanticcoastal lagoon. Bulletin of Marine Science 67: 73–82.
Schwindt, E., O. O. Iribarne, and F. I. Isla. 2004.Physical effects of an invading reef-buildingpolychaete on an Argentinean estuarine environ-ment. Estuarine Coastal and Shelf Science 59:109–120.
Talley, T. S., J. A. Crooks, and L. A. Levin. 2001. Habitatutilization and alteration by the invasive burrowingisopod, Sphaeroma quoyanum, in California saltmarshes. Marine Biology 138: 561–573.
i n v a s i v e a n i m a l s i n m a r s h e s 55
Silliman_ch03.qxd 2/23/09 1:20 PM Page 55

Torchin, M. E., J. E. Byers, and T. C. Huspeni. 2005.Differential parasitism of native and introducedsnails: Replacement of a parasite fauna. BiologicalInvasions 7: 885–894.
Torchin, M. E., R. F. Hechinger, T. C. Huspeni, K. L.Whitney, and K. D. Lafferty. 2005. The introducedribbed mussel (Geukensia demissa) in Estero dePunta Banda, Mexico: Interactions with the nativecord grass, Spartina foliosa. Biological Invasions 7:607–614.
Tyrrell, M. C., and J. E. Byers. 2007. Do artificial sub-strates favor nonindigenous fouling species overnatives? Journal of Experimental Marine Biologyand Ecology 342: 54–60.
Valiela, I., D. Rutecki, and S. Fox. 2004. Salt marshes:Biological controls of food webs in a diminishingenvironment. Journal of Experimental MarineBiology and Ecology 300: 131–159.
Verling, E., G. M. Ruiz, L. D. Smith, B. Galil, A. W.Miller, and K. R. Murphy. 2005. Supply-side inva-sion ecology: Characterizing propagule pressure incoastal ecosystems. Proceedings of the Royal Society ofLondon B, Biological Sciences 272: 1249–1256.
Washington Department of Fish and Wildlife. 2000.Untitled document available at http://www.wa.gov/wdfw/fish/shelfish/beachreg/1clam.htm#manila.
Wasson, K., K. Fenn, and J. S. Pearse. 2005. Habitatdifferences in marine invasions of centralCalifornia. Biological Invasions 7: 935–948.
Wasson, K., D. Lohrer, M. Crawford, and S. Rumrill.2002. Nonnative species in our nation’s estuaries:a framework for an invasion monitoring program.National Estuarine Research Reserve Technical ReportSeries 2002: 1.
Whitlow, W. L., N. A. Rice, and C. Sweeney. 2003.Native species vulnerability to introduced preda-tors: Testing an inducible defense and a refugefrom predation. Biological Invasions 5: 23–31.
Wright, J. P., and C. G. Jones. 2006. The concept oforganisms as ecosystem engineers ten years on:Progress, limitations, and challenges. BioScience56: 203–209.
Zaklan, S. D., and R. Ydenberg. 1997. The body size-burial depth relationship in the infaunal clamMya arenaria. Journal of Experimental MarineBiology and Ecology 215: 1–17.
56 i n v a s i o n s i n n o r t h a m e r i c a n s a l t m a r s h e s
Silliman_ch03.qxd 2/23/09 1:20 PM Page 56