intraspinal sprouting of unmyelinated pelvic afferents after complete spinal cord injury is...
TRANSCRIPT

IAA
S
SoU4
AenpcbtwpdoupAsn(awtdawuraI
Kp
Ap(1
2
si*EAauHragS
Neuroscience 159 (2009) 369–379
0d
NTRASPINAL SPROUTING OF UNMYELINATED PELVIC AFFERENTSFTER COMPLETE SPINAL CORD INJURY IS CORRELATED WITH
UTONOMIC DYSREFLEXIA INDUCED BY VISCERAL PAINcsbflsnbv1a(ieirrWotso(o
buafihtmppan1srtfitcwd(no
. HOU,1,2 H. DUALE1 AND A. G. RABCHEVSKY*
pinal Cord and Brain Injury Research Center, Department of Physi-logy, B471, Biomedical and Biological Sciences Research Building,niversity of Kentucky, 741 South Limestone Street, Lexington, KY0536-0509, USA
bstract—Autonomic dysreflexia is a potentially life-threat-ning hypertensive syndrome following high thoracic (T) spi-al cord injury (SCI). It is commonly triggered by noxiouselvic stimuli below the injury site that correlates with in-reased sprouting of primary afferent C-fibers into the lum-osacral (L/S) spinal cord. We have recently demonstratedhat injury-induced plasticity of (L/S) propriospinal neurons,hich relay pelvic visceral sensations to thoracolumbar sym-athetic preganglionic neurons, is also correlated with theevelopment of this syndrome. To determine the phenotypef pelvic afferent fiber sprouts after SCI, cholera toxin sub-nit beta (CTb) was injected into the distal colon 2 weeksost–T4 transection/sham to label colonic visceral afferents.fter 1 week of transport, the (L/S) spinal cords were cryo-ectioned and immunohistochemically stained for CTb, theociceptive-specific marker calcitonin gene-related peptideCGRP), and the myelinated fiber marker RT97. Quantitativenalysis showed that the density of CGRP� afferent fibersas significantly increased in the L/S dorsal horns of T4-
ransected versus sham rats, whereas RT97� afferent fiberensity showed no change. Importantly, CTb-labeled pelvicfferent fibers were co-localized with CGRP� fibers, but notith RT97� fibers. These results suggest that the sprouting ofnmyelinated nociceptive pelvic afferents following high tho-acic SCI, but not myelinated fibers, contributes to hypertensiveutonomic dysreflexia induced by pelvic visceral pain. © 2009BRO. Published by Elsevier Ltd. All rights reserved.
ey words: distal colon, cholera toxin subunit beta, pelvicrimary afferent, neuronal plasticity.
utonomic dysreflexia is a potentially life-threatening hy-ertensive syndrome that develops after spinal cord injurySCI) above the sixth thoracic (T) spinal segment. It is
Authors contributed equally.Present address: Center for Neural Repair, Department of Neuro-ciences, University of California, San Diego Biomedical Science Build-
ng, Rm. 4033, 9500 Gilman Dr. La Jolla, CA 92093-0626.Corresponding author. Tel: �1-859-323-0267; fax: �1-859-257-5737.-mail address: [email protected] (A. G. Rabchevsky).bbreviations: CGRP, calcitonin gene-related peptide; ChAT, cholinecetyltransferase; CRD, colorectal distension; CTb, cholera toxin sub-nit beta; DGC, dorsal gray commissure; DRG, dorsal root ganglia;R, heart rate; HRP, horseradish peroxidase; LCP, lateral collate-
al projection; L/S, lumbosacral; LT, Lissauer’s tract; MAP, meanrterial pressure; MCP, medial collateral projection; NGF, nerve
(rowth factor; PBS, phosphate buffered saline; SCI, spinal cord injury;PN, sacral parasympathetic nucleus; T, thoracic.
306-4522/09 © 2009 IBRO. Published by Elsevier Ltd. All rights reserved.oi:10.1016/j.neuroscience.2008.12.022
369
haracterized by severe hypertension due to sudden, mas-ive discharge of the sympathetic preganglionic neuronselow the injury site, which when accompanied by barore-ex-mediated bradycardia defines this syndrome (Fine-tone and Teasell, 1993; Zagon and Smith, 1993). Auto-omic dysreflexia is commonly triggered by noxious stimulielow the injury site, particularly by the distension of pelviciscera (bowel and bladder) (Lindan et al., 1980; Karlsson,999). It is believed that acute autonomic dysreflexiarises due to loss of bulbospinal sympathetic inhibitionKrassioukov and Weaver, 1995, 1996). This is followed bynjury-induced increases in growth factor expression (Brownt al., 2004) that elicit progressive structural and electrophys-
ological changes in both primary afferents and spinal neu-ons that coincide with increased severity of autonomic dys-eflexia (Maiorov et al., 1997; Weaver et al., 1997; Krenz and
eaver, 1998a,b; Chau et al., 2000). Moreover, studies fromur laboratory have shown that following complete T4 spinal
ransection, both primary afferent fiber sprouting into lumbo-acral (L/S) dorsal horns (Cameron et al., 2006) and plasticityf L/S propriospinal neurons in the dorsal gray commissureDGC) (Hou et al., 2008) correlate temporally with the devel-pment of autonomic dysreflexia.
Nevertheless, there is still uncertainty regarding whichranch of the pelvic primary afferents, the myelinated ornmyelinated fibers, contributes to the development ofutonomic dysreflexia. The unmyelinated pelvic afferentbers, which convey thermal and nociceptive information,ave been shown to contain calcitonin gene-related pep-ide (CGRP) (Keast and De Groat, 1992). Alternatively, theonoclonal antibody for 200 kDa neurofilament subunit inhosphorylated form, RT97, which is exclusively ex-ressed in A-fiber afferent neurons, is a marker for myelin-ted primary afferent fibers in both somatic and visceralerves (Perry et al., 1991; Sann et al., 1995; Wang et al.,998). Yoshimura et al. (1998) reported that capsaicin-ensitive neurons (unmyelinated fibers) in L6/S1 dorsaloot ganglia (DRG) were dramatically reduced in spinal-ransected rats compared with shams. Conversely, neuro-lament-rich DRG neurons (myelinated fibers) were de-ected at a significantly greater percentage after spinalord transection (Yoshimura et al., 1998). On the contrary,e and others have shown that experimental autonomicysreflexia induced by noxious colorectal distensionCRD) in spinal-transected rats correlates with profuseerve growth factor (NGF)–mediated intraspinal sproutingf CGRP� primary afferent fibers into L/S spinal segments
termination sites of pelvic visceral sensory axons) (Krenz
a2
sls2adullsfcms
A
AtKNmfkUilo
bpc(psM2UPriIaota
C
Tn
(cepHBfpswtcasic
A
TpadgcccC
D
TpwCpdmrt(Spsse2s(ss
I
TiwTl
T
A
CCRC
S. Hou et al. / Neuroscience 159 (2009) 369–379370
nd Weaver, 1998a; Weaver et al., 2001; Cameron et al.,006; Rabchevsky, 2006).
Primary sensory afferent fibers innervating the de-cending colon in rats run in the pelvic and hypogastric/umbar colonic nerves, distributing mainly to the L6/S1pinal level (Ness and Gebhart, 1987; Al-Chaer and Traub,002). To characterize the relative contribution of myelin-ted versus unmyelinated sensory fiber sprouting to theevelopment of autonomic dysreflexia, cholera toxin sub-nit beta (CTb) was injected into the distal colon of se-
ected T4-transected versus sham rats to label distal co-onic afferents and their terminal arbors within the L/Spinal cord. Our results demonstrate, for the first time, thatollowing high T SCI, the sprouting of unmyelinated noci-eptive pelvic afferents into the L/S spinal cord, but notyelinated fibers, is correlated with dysreflexic hyperten-
ion induced by visceral pain.
EXPERIMENTAL PROCEDURES
nimals and surgery
ll animal housing conditions, surgical procedures and post-opera-ive care techniques were conducted according to the University ofentucky Institutional Animal Care and Use Committee and theational Institutes of Health animal care guidelines to ensure mini-izing the number of animals used and any potential suffering. Adult
emale Wistar rats (�200–250 g) were anesthetized with a mixture ofetamine (80 mg/kg, i.p.; Fort Dodge Animal Health, Fort Dodge, IA,SA) and xylazine (10 mg/kg, i.p.; Butler, Columbus, OH, USA). The
njured group received complete T4 transection following T3 vertebralaminectomy (n�18), in contrast to the sham group which receivednly T3 laminectomy (n�15).
The spinal cord was completely transected with a #11 scalpellade at the T4 level following the laminectomy, as describedreviously (Cameron et al., 2006). After surgical operations wereomplete, the erector spinae muscles were sutured with 3-0 VycrilEthicon, Sommerfield, NJ, USA), the field was disinfected withovidone–iodine solution (Nova Plus, Irving, TX, USA), and thekin was closed with Michel wound clips (Roboz, Gaithersburg,D, USA). For post-operative care, animals were administered0 ml lactated Ringer’s solution (Baxter Healthcare, Deerfield, IL,SA) and 33 mg/kg cephazolin (Apothecon, Bristol-Myers Squibb,rinceton, NJ, USA) s.c. immediately after surgery, and for injured
ats twice daily for up to 10 days to maintain hydration and controlnfection. Buprenorphine (0.035 mg/kg; Hospira, Inc., Lakeforest,L, USA) was also administered s.c. once after recovery fromnesthesia and twice daily for the next 3 days to control post-perative pain. Bladders of injured rats were manually expressedwice daily until automatic bladder-emptying reflex developed atbout 10 days postinjury.
Tb injections and tracing
wo weeks after injury/sham, selected T4-transected (n�7) andon-transected (n�7) rats were reanesthetized with ketamine
able 1. Complete information and immunogen of primary antibodies
ntiserum Species Clonality Working dilution Source cata
Tb Goat Polyclonal 1:1500 List BiologiGRP Rabbit Polyclonal 1:2000 Sigma-AldrT97 Mouse Monoclonal 1:400 Chemicon,
hAT Goat Polyclonal 1:100 Chemicon, AB144P/80 mg/kg, i.p.) and xylazine (10 mg/kg, i.p.) to inject the distalolon with CTb, as previously described with modifications (Val-ntino et al., 2000). Briefly, a laparotomy was made to expose theelvic viscera. With a Hamilton microsyringe (33-gauge needle;amilton, Reno, NV, USA), 6 �l of CTb solution (1% in dH2O; Listiological Laboratories, Campbell, CA,USA) was injected circum-
erentially beneath the serosal layer in the distal colon, 1–2 cmroximal to the anus. The injections were made in six adjacentites surrounding the colon (1 �l/site). Injection sites were sealedith a drop of tissue adhesive (3 M Vetbond™, St. Paul, MN, USA)
o minimize leakage of tracer before the wound was sutured. Toontrol for CTb labeling specificity, CTb solution was topicallypplied on the ventral surface of the distal colon of an additionalham and an injured rat (n�1 per group). After 1 week post-CTbnjection to allow for transport, animals were perfused and spinalords were harvested for histology.
ssessing autonomic dysreflexia with CRD
wo weeks after T4 spinal transection, femoral cannulas were im-lanted in three randomly selected rats to verify the incidence ofutonomic dysreflexia during noxious CRD the following day, asetailed previously (Cameron et al., 2006). An injured rat was re-arded as dysreflexic if CRD, produced by inflation of a cardiacatheter balloon at a pressure that is known to stimulate the noci-eptive fibers, created a rise in mean arterial pressure (MAP) and aoncomitant decrease in heart rate (HR) for as long as the period ofRD.
issection and tissue processing
hree weeks after T4-transection/sham (n�33), including 1 weekost-CTb injections in the tracing cohort (n�7 per group), animalsere overdosed with sodium pentobarbital (150 mg/kg; Abbott,hicago, IL, USA) and perfused transcardially with 0.1 M phos-hate buffered saline (PBS), pH 7.4, followed by 4% paraformal-ehyde in PBS. A 6 cm long spinal cord extending from the conusedullaris to the transection site was removed, post-fixed for 4 h,
insed in 0.2 M phosphate buffer (PB) overnight, and cryopro-ected in 20% sucrose in 0.1 M PBS. The 3 cm caudal segments�T12–S3) were embedded in gum tragacanth (Sigma-Aldrich,t. Louis, MO, USA) in 20% sucrose/PBS for cryosectioning, asreviously detailed (Cameron et al., 2006). Embedded spinal cordegments were, snap-frozen in acetone chilled to �40 °C andtored at �80 °C until sectioning on a cryostat (Microm Laborg-rate, Walldorf, Germany). Approximately 30 consecutive rows of0 �m transverse cryosections separated by 100 �m from eachpinal cord were placed onto each of 10 adjacent glass slidesSuperfrost plus, Fisher Scientific, Pittsburgh, PA, USA) in twoeries, as previously detailed (Cameron et al., 2006). All mountedlides were stored at �20 °C until staining procedures.
mmunofluorescent histochemistry
he complete information of all primary antibodies used is detailedn Table 1. For CTb staining, slides with mounted coronal sectionsere thawed and pre-incubated in 0.1 M PBS containing 0.5%riton-X and 5% normal donkey serum (Vector Laboratories, Bur-
ingame, CA, USA) for 1 h, followed by incubation with goat
umber Immunogen
atories, 703/7032A5 CTb (choleragenoid)8/101K4846 CGRP-KLHTR1418670 200 kDa Neurofilament polypeptide
applied
log/lot n
cal Laborich, C819CBL212/
0608037072 ChAT purified from human placental enzyme

aodLt
s100spwrrICfiabs
ismrhds
as5w7c
C
ApisuOCtiAvts
mttiSnaeawRescrm
tt
S
UTIw
Ac
Ifpflw
SC
anrgtlwIe(esrg
ltd
Ffirb
S. Hou et al. / Neuroscience 159 (2009) 369–379 371
nti-CTb (List Biological Laboratories, 1:1500) in same buffervernight at 4 °C. The slides were then rinsed before applyingonkey anti-goat conjugated to FITC (Jackson ImmunoResearchaboratories, West Grove, PA, USA; 7.5 �g/ml) for 3 h at room
emperature.For CGRP� or RT97� afferent fibers, immunohistochemical
taining was conducted with rabbit anti-rat CGRP (Sigma-Aldrich,:2000) or mouse anti-rat RT97 (Chemicon, Temecula, CA, USA,.25 �g/ml) on coronal sections. Sections were pre-incubated in.1 M PBS containing 0.5% Triton-X and 5% normal donkeyerum for 1 h at room temperature, followed by incubation withrimary antibodies in the same buffer overnight at 4 °C. The slidesere then rinsed before applying secondary antibodies for 3 h at
oom temperature. The secondary antibodies were donkey anti-abbit or donkey anti-mouse conjugated to Texas-Red (JacksonmmunoResearch Laboratories; 7.5 �g/ml). To observe whetherTb-labeled afferents co-localized with either CGRP� or RT97�
ber, double immunostaining with both CTb and CGRP or RT97ntibodies was conducted. In addition, double immunostaining foroth CGRP and RT97 antibodies was performed to rule out pos-ible co-localization.
Using commercially available monoclonal or polyclonal antibod-es against rat choline acetyltransferase (ChAT), we were unable touccessfully double-label ChAT in putative autonomic or somaticotoneurons with the CTb antibody we employed. Therefore, the
emaining series of sections were incubated with reliable goat anti-uman ChAT (Chemicon, 1:100) overnight at 4 °C, followed byonkey anti-goat conjugated to Texas Red (Jackson ImmunoRe-earch Laboratories; 7.5 �g/ml) for 3 h at room temperature.
All control slides had the primary antibody omitted beforepplying the secondary. After final rinses, slides were cover-lipped using Vectashield mounting medium (Vector) containing
�M Hoechst 33342 nuclear dye (Sigma-Aldrich), and sealedith Cutex nail hardener (Jackson, WY, USA). Adobe Photoshop.0 was used to manipulate photomicrographs only for brightness/ontrast and to construct all figures.
Tb-labeling and densitometric quantification
n Olympus IX81 spinning confocal microscope (Olympus Cor-oration, Melville, NY, USA) was used for high magnification dual
mmunofluorescence microscopy of CTb-labeled cells. For all den-itometric analyses, including RT97� myelinated and CGRP�
nmyelinated fibers, digital images were captured using anptronics digital video camera (Optronics Corporation, Goleta,A, USA). Densitometry for both groups, with and without CTb
racing, was conducted in a blinded fashion using the Bioquant®
mage analysis program (Nova Prime, V6.70.10; Bioquant Imagenalysis Corporation, Nashville, TN, USA) on coronal sectionsiewed under an Olympus BX51 microscope (Olympus Corpora-ion) using established methods (Cameron et al., 2006; Rabchev-ky et al., 2007).
All sections were photographed with the same 10� objectiveagnification (10� eyepiece) and with the same exposure set-
ings. In all sections analyzed, our region of interest encompassedhe unilateral (right) dorsal gray matter above the central canal,ncluding Lissauer’s tract (LT) on the surface of dorsal horn (Fig. 1).ince CTb-labeled cells were seen in the sacral parasympatheticucleus (SPN), this region was excluded from all densitometricnalyses (see Discussion, Methodological considerations). Forach animal, three serial spinal cord sections separated by 1 mmnd centering on the L6/S1 level, as identified by cytoarchitecture,ere selected for quantification. To measure CTb�/CGRP�/T97� immunoreactivity, labeled fibers were “thresholded” onach digital photograph, using the Bioquant program. For eachection, the percent area occupied by immunolabeled fibers wasalculated as the ratio of immunoreactive area divided by theegion of interest area in unilateral dorsal gray matter �100. The
ean percentage of immunoreactive area occupied in three sec- gions was calculated, followed by the mean for all animals in areatment group.
tatistical analysis
npaired Student’s t-test were used to compare data between4-transected and non-transected groups, using StatView (SAS
nstitute, Cary, NC, USA). Significance throughout all experimentsas set at P�0.05. Data are represented as mean�S.D.
RESULTS
utonomic dysreflexia: noxious CRD-inducedhanges in MAP and HR
n all three selected rats with T4-transection, CRD per-ormed 2 weeks post-SCI elicited MAP increases accom-anied by reduced HR characteristic of autonomic dysre-exia (data not shown). These results are in accordanceith what we have published (Cameron et al., 2006).
prouting of pelvic primary afferents withoutTb tracing
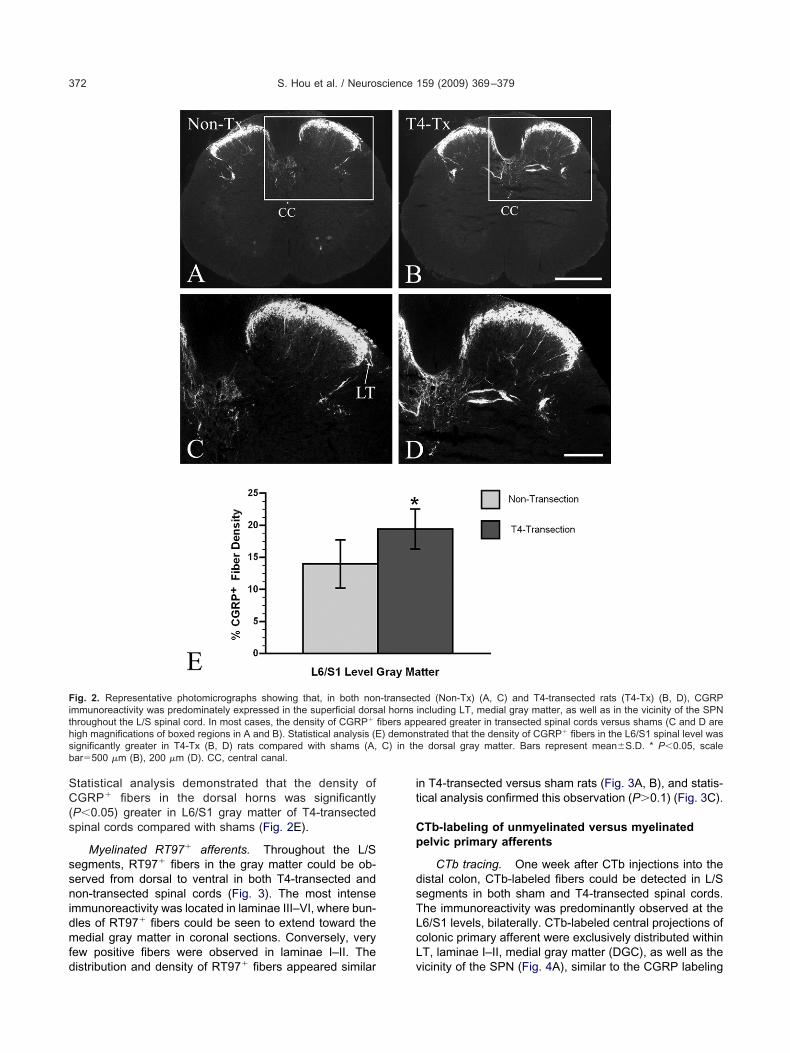
Unmyelinated CGRP� afferents. In both T4-transectednd sham rats, CGRP immunoreactivity was predomi-ately expressed in fibers and punctate terminals in threeegions: the superficial dorsal horn (laminae I–II), medialray matter (above the central canal), as well as peri-SPNhroughout the L/S spinal cord (Fig. 2). LT located aboveamina I and along the lateral edge of the dorsal hornere also immunolabeled (Nadelhaft and Booth, 1984).
n addition, CGRP� branching fibers in laminae III–Vxtended individually and in bundles toward the DGClamina X). Notably, CGRP� fibers could not be seenxtending into the ventral horn in either T4-transected orham spinal cords (Fig. 2A–D). However, CGRP� neu-ons were frequently observed in the ventral horn of bothroups (Fig. 2A).
Qualitatively, CGRP labeling density and distribution inaminae I–II at the L6/S1 level appeared similar in T4-ransected and sham rats. However, following injury theensity of CGRP� fibers appeared increased in other re-
ig. 1. Photomicrographs illustrating our region of interest for quanti-cation (shaded inset) of CGRP�, RT97�, or CTb� fiber density in theight upper dorsal gray matter at the L6/S1 spinal level. Scalear�500 �m. CC, central canal.
ions, particularly in laminae III–V and DGC (Fig. 2A–D).
SC(s
ssnidmfd
it
Cp
dsTLcL
Fithsb
S. Hou et al. / Neuroscience 159 (2009) 369–379372
tatistical analysis demonstrated that the density ofGRP� fibers in the dorsal horns was significantly
P�0.05) greater in L6/S1 gray matter of T4-transectedpinal cords compared with shams (Fig. 2E).
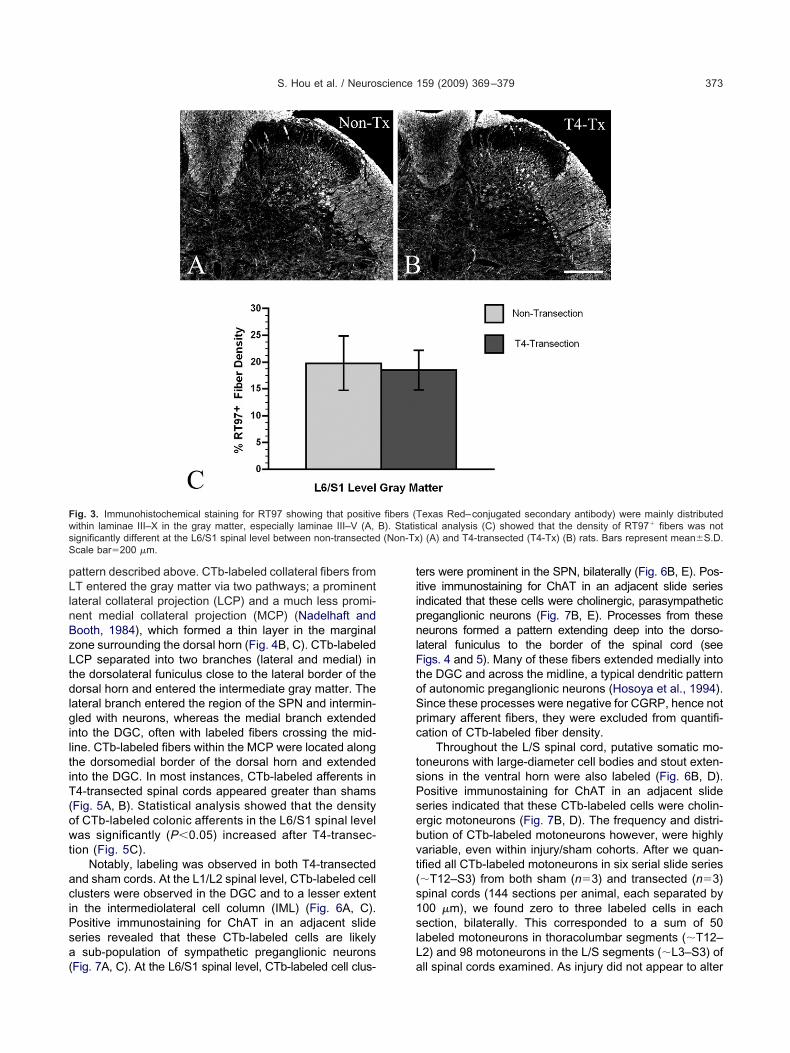
Myelinated RT97� afferents. Throughout the L/Segments, RT97� fibers in the gray matter could be ob-erved from dorsal to ventral in both T4-transected andon-transected spinal cords (Fig. 3). The most intense
mmunoreactivity was located in laminae III–VI, where bun-les of RT97� fibers could be seen to extend toward theedial gray matter in coronal sections. Conversely, very
ew positive fibers were observed in laminae I–II. The
ig. 2. Representative photomicrographs showing that, in both nonmmunoreactivity was predominately expressed in the superficial dorshroughout the L/S spinal cord. In most cases, the density of CGRP�
igh magnifications of boxed regions in A and B). Statistical analysis (Eignificantly greater in T4-Tx (B, D) rats compared with shams (A,ar�500 �m (B), 200 �m (D). CC, central canal.
istribution and density of RT97� fibers appeared similar v
n T4-transected versus sham rats (Fig. 3A, B), and statis-ical analysis confirmed this observation (P�0.1) (Fig. 3C).
Tb-labeling of unmyelinated versus myelinatedelvic primary afferents
CTb tracing. One week after CTb injections into theistal colon, CTb-labeled fibers could be detected in L/Segments in both sham and T4-transected spinal cords.he immunoreactivity was predominantly observed at the6/S1 levels, bilaterally. CTb-labeled central projections ofolonic primary afferent were exclusively distributed withinT, laminae I–II, medial gray matter (DGC), as well as the
ted (Non-Tx) (A, C) and T4-transected rats (T4-Tx) (B, D), CGRPincluding LT, medial gray matter, as well as in the vicinity of the SPNeared greater in transected spinal cords versus shams (C and D arestrated that the density of CGRP� fibers in the L6/S1 spinal level wase dorsal gray matter. Bars represent mean�S.D. * P�0.05, scale
-transecal hornsfibers app) demonC) in th
icinity of the SPN (Fig. 4A), similar to the CGRP labeling
pLlnBzLtdlgiltiT(owt
aciPsa(
tiipnlFtoSpc
tsPsebvt(s1slL
Fws (Non-TxS
S. Hou et al. / Neuroscience 159 (2009) 369–379 373
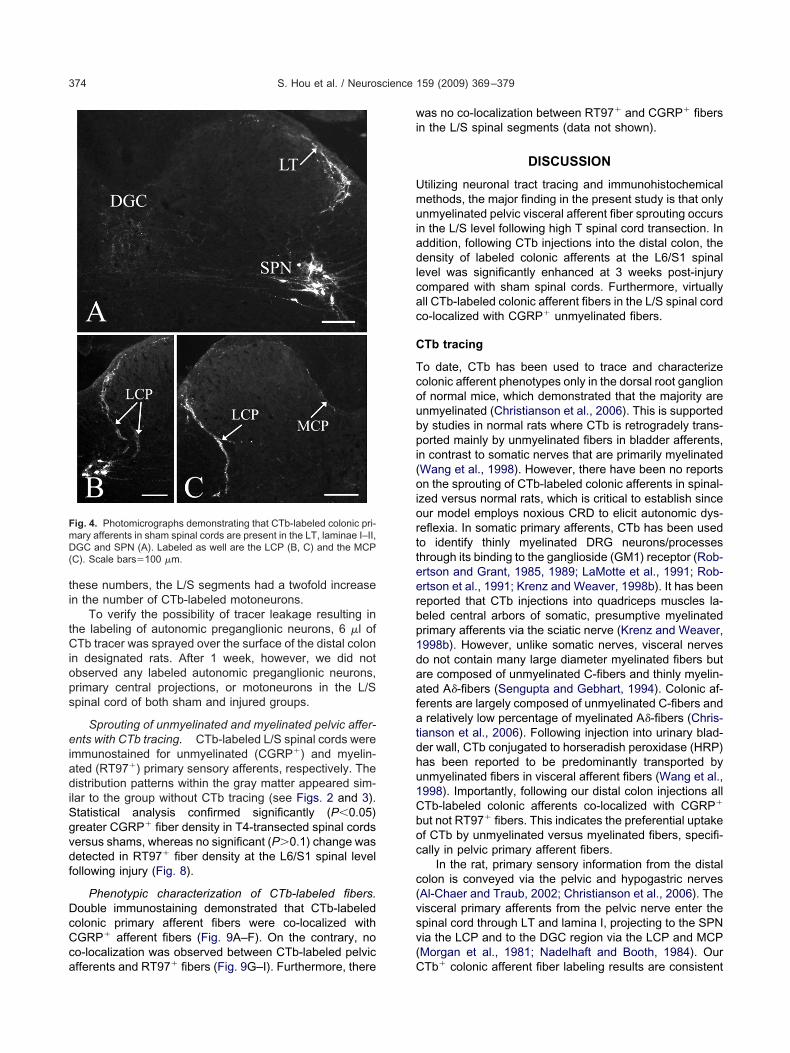
attern described above. CTb-labeled collateral fibers fromT entered the gray matter via two pathways; a prominent
ateral collateral projection (LCP) and a much less promi-ent medial collateral projection (MCP) (Nadelhaft andooth, 1984), which formed a thin layer in the marginalone surrounding the dorsal horn (Fig. 4B, C). CTb-labeledCP separated into two branches (lateral and medial) inhe dorsolateral funiculus close to the lateral border of theorsal horn and entered the intermediate gray matter. The
ateral branch entered the region of the SPN and intermin-led with neurons, whereas the medial branch extended
nto the DGC, often with labeled fibers crossing the mid-ine. CTb-labeled fibers within the MCP were located alonghe dorsomedial border of the dorsal horn and extendednto the DGC. In most instances, CTb-labeled afferents in4-transected spinal cords appeared greater than shamsFig. 5A, B). Statistical analysis showed that the densityf CTb-labeled colonic afferents in the L6/S1 spinal levelas significantly (P�0.05) increased after T4-transec-
ion (Fig. 5C).Notably, labeling was observed in both T4-transected
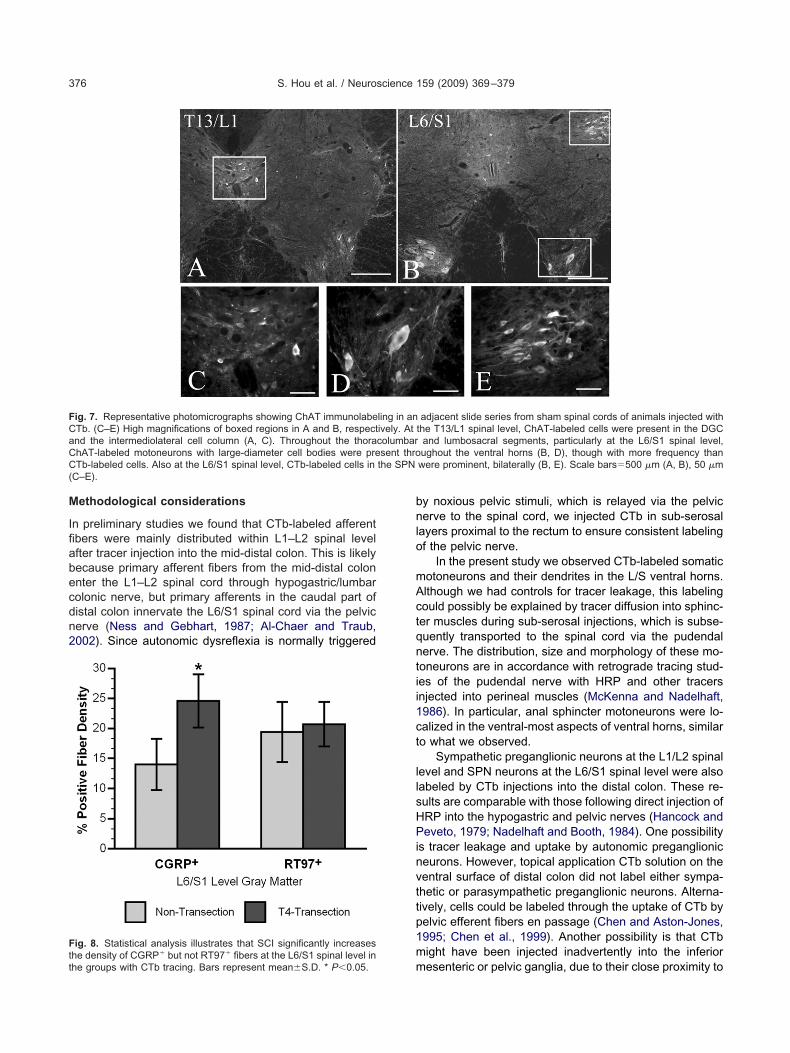
nd sham cords. At the L1/L2 spinal level, CTb-labeled celllusters were observed in the DGC and to a lesser extentn the intermediolateral cell column (IML) (Fig. 6A, C).ositive immunostaining for ChAT in an adjacent slideeries revealed that these CTb-labeled cells are likely
sub-population of sympathetic preganglionic neurons
ig. 3. Immunohistochemical staining for RT97 showing that positiveithin laminae III–X in the gray matter, especially laminae III–V (A,ignificantly different at the L6/S1 spinal level between non-transectedcale bar�200 �m.
Fig. 7A, C). At the L6/S1 spinal level, CTb-labeled cell clus- a
ers were prominent in the SPN, bilaterally (Fig. 6B, E). Pos-tive immunostaining for ChAT in an adjacent slide seriesndicated that these cells were cholinergic, parasympatheticreganglionic neurons (Fig. 7B, E). Processes from theseeurons formed a pattern extending deep into the dorso-
ateral funiculus to the border of the spinal cord (seeigs. 4 and 5). Many of these fibers extended medially into
he DGC and across the midline, a typical dendritic patternf autonomic preganglionic neurons (Hosoya et al., 1994).ince these processes were negative for CGRP, hence notrimary afferent fibers, they were excluded from quantifi-ation of CTb-labeled fiber density.
Throughout the L/S spinal cord, putative somatic mo-oneurons with large-diameter cell bodies and stout exten-ions in the ventral horn were also labeled (Fig. 6B, D).ositive immunostaining for ChAT in an adjacent slideeries indicated that these CTb-labeled cells were cholin-rgic motoneurons (Fig. 7B, D). The frequency and distri-ution of CTb-labeled motoneurons however, were highlyariable, even within injury/sham cohorts. After we quan-ified all CTb-labeled motoneurons in six serial slide series�T12–S3) from both sham (n�3) and transected (n�3)pinal cords (144 sections per animal, each separated by00 �m), we found zero to three labeled cells in eachection, bilaterally. This corresponded to a sum of 50
abeled motoneurons in thoracolumbar segments (�T12–2) and 98 motoneurons in the L/S segments (�L3–S3) of
exas Red–conjugated secondary antibody) were mainly distributedtical analysis (C) showed that the density of RT97� fibers was not) (A) and T4-transected (T4-Tx) (B) rats. Bars represent mean�S.D.
fibers (TB). Statis
ll spinal cords examined. As injury did not appear to alter
ti
tCiops
eiadiSgvdf
DcCca
wi
Umuiadlcac
C
Tcoubpi(oiortteerbp1daafatdhu1Cboc
c(vsv(
FmD(
S. Hou et al. / Neuroscience 159 (2009) 369–379374
hese numbers, the L/S segments had a twofold increasen the number of CTb-labeled motoneurons.
To verify the possibility of tracer leakage resulting inhe labeling of autonomic preganglionic neurons, 6 �l ofTb tracer was sprayed over the surface of the distal colon
n designated rats. After 1 week, however, we did notbserved any labeled autonomic preganglionic neurons,rimary central projections, or motoneurons in the L/Spinal cord of both sham and injured groups.
Sprouting of unmyelinated and myelinated pelvic affer-nts with CTb tracing. CTb-labeled L/S spinal cords were
mmunostained for unmyelinated (CGRP�) and myelin-ted (RT97�) primary sensory afferents, respectively. Theistribution patterns within the gray matter appeared sim-
lar to the group without CTb tracing (see Figs. 2 and 3).tatistical analysis confirmed significantly (P�0.05)reater CGRP� fiber density in T4-transected spinal cordsersus shams, whereas no significant (P�0.1) change wasetected in RT97� fiber density at the L6/S1 spinal levelollowing injury (Fig. 8).
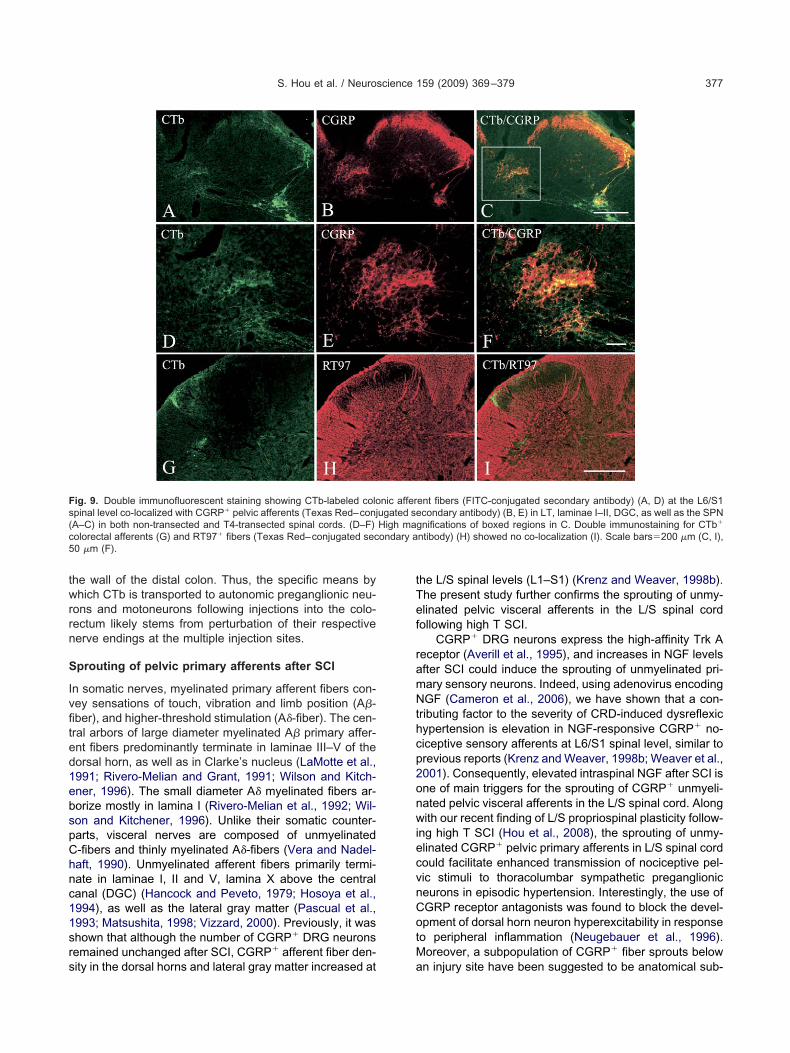
Phenotypic characterization of CTb-labeled fibers.ouble immunostaining demonstrated that CTb-labeledolonic primary afferent fibers were co-localized withGRP� afferent fibers (Fig. 9A–F). On the contrary, noo-localization was observed between CTb-labeled pelvic
ig. 4. Photomicrographs demonstrating that CTb-labeled colonic pri-ary afferents in sham spinal cords are present in the LT, laminae I–II,GC and SPN (A). Labeled as well are the LCP (B, C) and the MCP
C). Scale bars�100 �m.
fferents and RT97� fibers (Fig. 9G–I). Furthermore, there C
as no co-localization between RT97� and CGRP� fibersn the L/S spinal segments (data not shown).
DISCUSSION
tilizing neuronal tract tracing and immunohistochemicalethods, the major finding in the present study is that onlynmyelinated pelvic visceral afferent fiber sprouting occurs
n the L/S level following high T spinal cord transection. Inddition, following CTb injections into the distal colon, theensity of labeled colonic afferents at the L6/S1 spinal
evel was significantly enhanced at 3 weeks post-injuryompared with sham spinal cords. Furthermore, virtuallyll CTb-labeled colonic afferent fibers in the L/S spinal cordo-localized with CGRP� unmyelinated fibers.
Tb tracing
o date, CTb has been used to trace and characterizeolonic afferent phenotypes only in the dorsal root ganglionf normal mice, which demonstrated that the majority arenmyelinated (Christianson et al., 2006). This is supportedy studies in normal rats where CTb is retrogradely trans-orted mainly by unmyelinated fibers in bladder afferents,
n contrast to somatic nerves that are primarily myelinatedWang et al., 1998). However, there have been no reportsn the sprouting of CTb-labeled colonic afferents in spinal-
zed versus normal rats, which is critical to establish sinceur model employs noxious CRD to elicit autonomic dys-eflexia. In somatic primary afferents, CTb has been usedo identify thinly myelinated DRG neurons/processeshrough its binding to the ganglioside (GM1) receptor (Rob-rtson and Grant, 1985, 1989; LaMotte et al., 1991; Rob-rtson et al., 1991; Krenz and Weaver, 1998b). It has beeneported that CTb injections into quadriceps muscles la-eled central arbors of somatic, presumptive myelinatedrimary afferents via the sciatic nerve (Krenz and Weaver,998b). However, unlike somatic nerves, visceral nerveso not contain many large diameter myelinated fibers butre composed of unmyelinated C-fibers and thinly myelin-ted A�-fibers (Sengupta and Gebhart, 1994). Colonic af-erents are largely composed of unmyelinated C-fibers and
relatively low percentage of myelinated A�-fibers (Chris-ianson et al., 2006). Following injection into urinary blad-er wall, CTb conjugated to horseradish peroxidase (HRP)as been reported to be predominantly transported bynmyelinated fibers in visceral afferent fibers (Wang et al.,998). Importantly, following our distal colon injections allTb-labeled colonic afferents co-localized with CGRP�
ut not RT97� fibers. This indicates the preferential uptakef CTb by unmyelinated versus myelinated fibers, specifi-ally in pelvic primary afferent fibers.
In the rat, primary sensory information from the distalolon is conveyed via the pelvic and hypogastric nervesAl-Chaer and Traub, 2002; Christianson et al., 2006). Theisceral primary afferents from the pelvic nerve enter thepinal cord through LT and lamina I, projecting to the SPNia the LCP and to the DGC region via the LCP and MCPMorgan et al., 1981; Nadelhaft and Booth, 1984). Our
Tb� colonic afferent fiber labeling results are consistent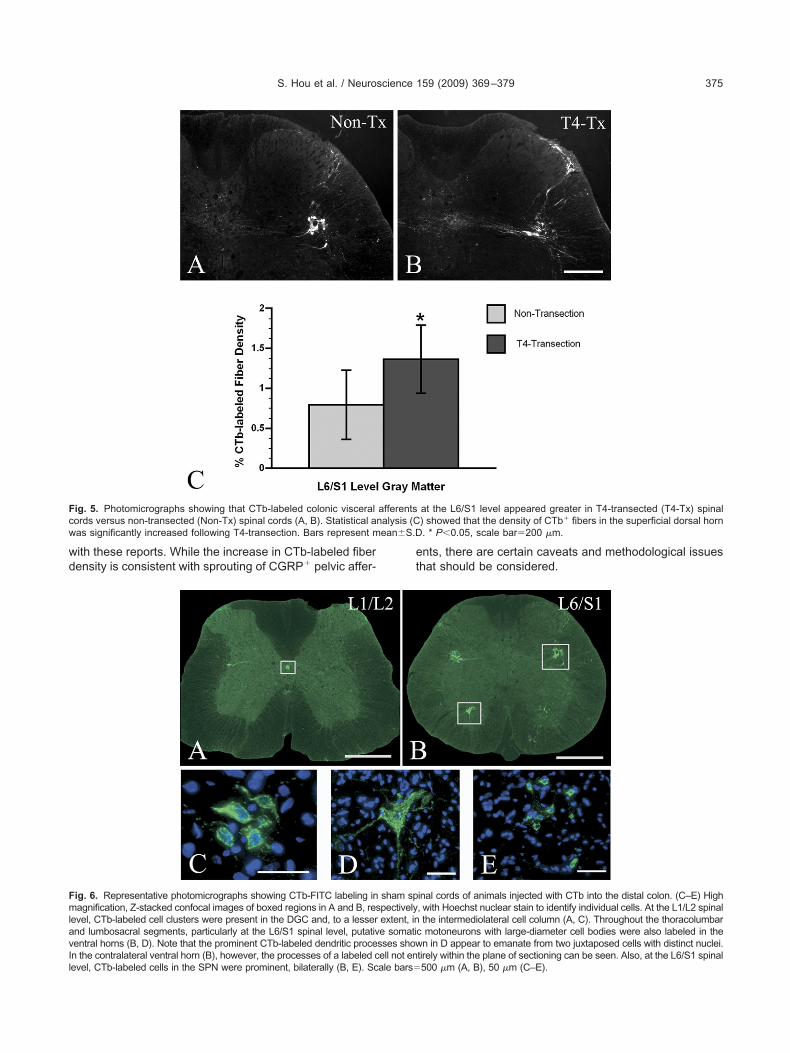
wd
et
Fc nalysis (Cw ean�S.D
FmlavIl
S. Hou et al. / Neuroscience 159 (2009) 369–379 375
ith these reports. While the increase in CTb-labeled fiberensity is consistent with sprouting of CGRP� pelvic affer-
ig. 5. Photomicrographs showing that CTb-labeled colonic visceralords versus non-transected (Non-Tx) spinal cords (A, B). Statistical aas significantly increased following T4-transection. Bars represent m
ig. 6. Representative photomicrographs showing CTb-FITC labeling inagnification, Z-stacked confocal images of boxed regions in A and B, res
evel, CTb-labeled cell clusters were present in the DGC and, to a lessernd lumbosacral segments, particularly at the L6/S1 spinal level, putativentral horns (B, D). Note that the prominent CTb-labeled dendritic proce
n the contralateral ventral horn (B), however, the processes of a labeled cell not enevel, CTb-labeled cells in the SPN were prominent, bilaterally (B, E). Scale bars�nts, there are certain caveats and methodological issueshat should be considered.
at the L6/S1 level appeared greater in T4-transected (T4-Tx) spinal) showed that the density of CTb� fibers in the superficial dorsal horn. * P�0.05, scale bar�200 �m.
inal cords of animals injected with CTb into the distal colon. (C–E) High, with Hoechst nuclear stain to identify individual cells. At the L1/L2 spinalthe intermediolateral cell column (A, C). Throughout the thoracolumbar
c motoneurons with large-diameter cell bodies were also labeled in then in D appear to emanate from two juxtaposed cells with distinct nuclei.
afferents
sham sppectivelyextent, ine somatisses show
tirely within the plane of sectioning can be seen. Also, at the L6/S1 spinal500 �m (A, B), 50 �m (C–E).
M
Ifiabecdn2
bnlo
mActqntii1ct
llsHPinvttp1m
FCaCC the SPN(
Ftt
S. Hou et al. / Neuroscience 159 (2009) 369–379376
ethodological considerations
n preliminary studies we found that CTb-labeled afferentbers were mainly distributed within L1–L2 spinal levelfter tracer injection into the mid-distal colon. This is likelyecause primary afferent fibers from the mid-distal colonnter the L1–L2 spinal cord through hypogastric/lumbarolonic nerve, but primary afferents in the caudal part ofistal colon innervate the L6/S1 spinal cord via the pelvicerve (Ness and Gebhart, 1987; Al-Chaer and Traub,002). Since autonomic dysreflexia is normally triggered
ig. 7. Representative photomicrographs showing ChAT immunolabeTb. (C–E) High magnifications of boxed regions in A and B, respectnd the intermediolateral cell column (A, C). Throughout the thorahAT-labeled motoneurons with large-diameter cell bodies were preTb-labeled cells. Also at the L6/S1 spinal level, CTb-labeled cells in
C–E).
ig. 8. Statistical analysis illustrates that SCI significantly increases� �
mhe density of CGRP but not RT97 fibers at the L6/S1 spinal level inhe groups with CTb tracing. Bars represent mean�S.D. * P�0.05.
y noxious pelvic stimuli, which is relayed via the pelvicerve to the spinal cord, we injected CTb in sub-serosal
ayers proximal to the rectum to ensure consistent labelingf the pelvic nerve.
In the present study we observed CTb-labeled somaticotoneurons and their dendrites in the L/S ventral horns.lthough we had controls for tracer leakage, this labelingould possibly be explained by tracer diffusion into sphinc-er muscles during sub-serosal injections, which is subse-uently transported to the spinal cord via the pudendalerve. The distribution, size and morphology of these mo-oneurons are in accordance with retrograde tracing stud-es of the pudendal nerve with HRP and other tracersnjected into perineal muscles (McKenna and Nadelhaft,986). In particular, anal sphincter motoneurons were lo-alized in the ventral-most aspects of ventral horns, similaro what we observed.
Sympathetic preganglionic neurons at the L1/L2 spinalevel and SPN neurons at the L6/S1 spinal level were alsoabeled by CTb injections into the distal colon. These re-ults are comparable with those following direct injection ofRP into the hypogastric and pelvic nerves (Hancock andeveto, 1979; Nadelhaft and Booth, 1984). One possibility
s tracer leakage and uptake by autonomic preganglioniceurons. However, topical application CTb solution on theentral surface of distal colon did not label either sympa-hetic or parasympathetic preganglionic neurons. Alterna-ively, cells could be labeled through the uptake of CTb byelvic efferent fibers en passage (Chen and Aston-Jones,995; Chen et al., 1999). Another possibility is that CTbight have been injected inadvertently into the inferior
adjacent slide series from sham spinal cords of animals injected withthe T13/L1 spinal level, ChAT-labeled cells were present in the DGC
and lumbosacral segments, particularly at the L6/S1 spinal level,ughout the ventral horns (B, D), though with more frequency thanwere prominent, bilaterally (B, E). Scale bars�500 �m (A, B), 50 �m
ling in anively. Atcolumbarsent thro
esenteric or pelvic ganglia, due to their close proximity to
twrrn
S
Ivfited1ebspChnc11srs
tTef
ramNthcp2onwiecvnCotM
Fs(c ondary a5
S. Hou et al. / Neuroscience 159 (2009) 369–379 377
he wall of the distal colon. Thus, the specific means byhich CTb is transported to autonomic preganglionic neu-
ons and motoneurons following injections into the colo-ectum likely stems from perturbation of their respectiveerve endings at the multiple injection sites.
prouting of pelvic primary afferents after SCI
n somatic nerves, myelinated primary afferent fibers con-ey sensations of touch, vibration and limb position (A�-ber), and higher-threshold stimulation (A�-fiber). The cen-ral arbors of large diameter myelinated A� primary affer-nt fibers predominantly terminate in laminae III–V of theorsal horn, as well as in Clarke’s nucleus (LaMotte et al.,991; Rivero-Melian and Grant, 1991; Wilson and Kitch-ner, 1996). The small diameter A� myelinated fibers ar-orize mostly in lamina I (Rivero-Melian et al., 1992; Wil-on and Kitchener, 1996). Unlike their somatic counter-arts, visceral nerves are composed of unmyelinated-fibers and thinly myelinated A�-fibers (Vera and Nadel-aft, 1990). Unmyelinated afferent fibers primarily termi-ate in laminae I, II and V, lamina X above the centralanal (DGC) (Hancock and Peveto, 1979; Hosoya et al.,994), as well as the lateral gray matter (Pascual et al.,993; Matsushita, 1998; Vizzard, 2000). Previously, it washown that although the number of CGRP� DRG neuronsemained unchanged after SCI, CGRP� afferent fiber den-
ig. 9. Double immunofluorescent staining showing CTb-labeled colopinal level co-localized with CGRP� pelvic afferents (Texas Red–conjA–C) in both non-transected and T4-transected spinal cords. (D–F)olorectal afferents (G) and RT97� fibers (Texas Red–conjugated sec0 �m (F).
ity in the dorsal horns and lateral gray matter increased at a
he L/S spinal levels (L1–S1) (Krenz and Weaver, 1998b).he present study further confirms the sprouting of unmy-linated pelvic visceral afferents in the L/S spinal cordollowing high T SCI.
CGRP� DRG neurons express the high-affinity Trk Aeceptor (Averill et al., 1995), and increases in NGF levelsfter SCI could induce the sprouting of unmyelinated pri-ary sensory neurons. Indeed, using adenovirus encodingGF (Cameron et al., 2006), we have shown that a con-
ributing factor to the severity of CRD-induced dysreflexicypertension is elevation in NGF-responsive CGRP� no-iceptive sensory afferents at L6/S1 spinal level, similar torevious reports (Krenz and Weaver, 1998b; Weaver et al.,001). Consequently, elevated intraspinal NGF after SCI isne of main triggers for the sprouting of CGRP� unmyeli-ated pelvic visceral afferents in the L/S spinal cord. Alongith our recent finding of L/S propriospinal plasticity follow-
ng high T SCI (Hou et al., 2008), the sprouting of unmy-linated CGRP� pelvic primary afferents in L/S spinal cordould facilitate enhanced transmission of nociceptive pel-ic stimuli to thoracolumbar sympathetic preganglioniceurons in episodic hypertension. Interestingly, the use ofGRP receptor antagonists was found to block the devel-pment of dorsal horn neuron hyperexcitability in responseo peripheral inflammation (Neugebauer et al., 1996).oreover, a subpopulation of CGRP� fiber sprouts below
ent fibers (FITC-conjugated secondary antibody) (A, D) at the L6/S1condary antibody) (B, E) in LT, laminae I–II, DGC, as well as the SPN
gnifications of boxed regions in C. Double immunostaining for CTb�
ntibody) (H) showed no co-localization (I). Scale bars�200 �m (C, I),
nic afferugated seHigh ma
n injury site have been suggested to be anatomical sub-

sH
Ihnltniasaisospon
AK(e
A
A
B
C
C
C
C
C
C
F
H
H
H
KK
K
K
K
K
L
L
M
M
M
M
N
N
N
O
P
P
S. Hou et al. / Neuroscience 159 (2009) 369–379378
trates for chronic pain syndromes (Christensen andulsebosch, 1997; Ondarza et al., 2003).
CONCLUSION
n summary, the present study demonstrates that followingigh T SCI the selective sprouting of CGRP� unmyelinatedociceptive pelvic afferents into the L/S spinal cord corre-
ates with the development of autonomic dysreflexia. Fur-hermore, CTb injections into the distal colon predomi-antly labeled unmyelinated pelvic visceral afferent fibers,
n contrast to previous studies in which myelinated somaticfferent fibers were labeled with CTb. Our results demon-trate that the sprouting of unmyelinated nociceptive pelvicfferents, but not myelinated fibers, likely mediates the
nitiation of CRD-evoked autonomic dysreflexia. Futuretudies will focus on employing viral-mediated site-specificver-expression of neurotrophin-3 (NT-3) to enhance theurvival and growth of myelinated mechanoceptive and/orroprioceptive sensory neurons to rule out the contributionf myelinated fiber sprouting to the development of auto-omic dysreflexia.
cknowledgments—This work was supported by grants fromSCHIRT #3-11 (A.G.R.), NIH/NINDS R01 NS049901-01
A.G.R.) and P30 NS051220. Authors are grateful for the technicalxpertise of Travis Lyttle, MS and Joseph Whelan, MS.
REFERENCES
l-Chaer ED, Traub RJ (2002) Biological basis of visceral pain: recentdevelopments. Pain 96:221–225.
verill S, McMahon SB, Clary DO, Reichardt LF, Priestley JV (1995)Immunocytochemical localization of trkA receptors in chemicallyidentified subgroups of adult rat sensory neurons. Eur J Neurosci7:1484–1494.
rown A, Ricci MJ, Weaver LC (2004) NGF message and proteindistribution in the injured rat spinal cord. Exp Neurol 188:115–127.
ameron AA, Smith GM, Randall DC, Brown DR, Rabchevsky AG(2006) Genetic manipulation of intraspinal plasticity after spinalcord injury alters the severity of autonomic dysreflexia. J Neurosci26:2923–2932.
hau D, Johns DG, Schramm LP (2000) Ongoing and stimulus-evoked activity of sympathetically correlated neurons in the inter-mediate zone and dorsal horn of acutely spinalized rats. J Neuro-physiol 83:2699–2707.
hen S, Aston-Jones G (1995) Evidence that cholera toxin B subunit(CTb) can be avidly taken up and transported by fibers of passage.Brain Res 674:107–111.
hen S, Yang M, Miselis RR, Aston-Jones G (1999) Characterizationof transsynaptic tracing with central application of pseudorabiesvirus. Brain Res 838:171–183.
hristensen MD, Hulsebosch CE (1997) Chronic central pain afterspinal cord injury. J Neurotrauma 14:517–537.
hristianson JA, Traub RJ, Davis BM (2006) Differences in spinaldistribution and neurochemical phenotype of colonic afferents inmouse and rat. J Comp Neurol 494:246–259.
inestone HM, Teasell RW (1993) Autonomic dysreflexia after brain-stem tumor resection. A case report. Am J Phys Med Rehabil72:395–397.
ancock MB, Peveto CA (1979) Preganglionic neurons in the sacralspinal cord of the rat: an HRP study. Neurosci Lett 11:1–5.
osoya Y, Nadelhaft I, Wang D, Kohno K (1994) Thoracolumbar
sympathetic preganglionic neurons in the dorsal commissural nu-cleus of the male rat: an immunohistochemical study usingretrograde labeling of cholera toxin subunit B. Exp Brain Res98:21–30.
ou S, Duale H, Cameron AA, Abshire SM, Lyttle TS, Rabchevsky AG(2008) Plasticity of lumbosacral propriospinal neurons is associ-ated with the development of autonomic dysreflexia after thoracicspinal cord transection. J Comp Neurol 509:382–399.
arlsson AK (1999) Autonomic dysreflexia. Spinal Cord 37:383–391.east JR, De Groat WC (1992) Segmental distribution and peptide
content of primary afferent neurons innervating the urogenital or-gans and colon of male rats. J Comp Neurol 319:615–623.
rassioukov AV, Weaver LC (1995) Episodic hypertension due toautonomic dysreflexia in acute and chronic spinal cord-injured rats.Am J Physiol 268:H2077–H2083.
rassioukov AV, Weaver LC (1996) Morphological changes in sym-pathetic preganglionic neurons after spinal cord injury in rats.Neuroscience 70:211–225.
renz NR, Weaver LC (1998a) Changes in the morphology of sympa-thetic preganglionic neurons parallel the development of auto-nomic dysreflexia after spinal cord injury in rats. Neurosci Lett243:61–64.
renz NR, Weaver LC (1998b) Sprouting of primary afferent fibersafter spinal cord transection in the rat. Neuroscience 85:443–458.
aMotte CC, Kapadia SE, Shapiro CM (1991) Central projections ofthe sciatic, saphenous, median, and ulnar nerves of the rat dem-onstrated by transganglionic transport of choleragenoid-HRP (B-HRP) and wheat germ agglutinin-HRP (WGA-HRP). J Comp Neu-rol 311:546–562.
indan R, Joiner E, Freehafer AA, Hazel C (1980) Incidence andclinical features of autonomic dysreflexia in patients with spinalcord injury. Paraplegia 18:285–292.
aiorov DN, Krenz NR, Krassioukov AV, Weaver LC (1997) Role ofspinal NMDA and AMPA receptors in episodic hypertension inconscious spinal rats. Am J Physiol 273:H1266–H1274.
atsushita M (1998) Ascending propriospinal afferents to area X(substantia grisea centralis) of the spinal cord in the rat. Exp BrainRes 119:356–366.
cKenna KE, Nadelhaft I (1986) The organization of the pudendalnerve in the male and female rat. J Comp Neurol 248:532–549.
organ C, Nadelhaft I, de Groat WC (1981) The distribution of visceralprimary afferents from the pelvic nerve to Lissauer’s tract and thespinal gray matter and its relationship to the sacral parasympa-thetic nucleus. J Comp Neurol 201:415–440.
adelhaft I, Booth AM (1984) The location and morphology of pregan-glionic neurons and the distribution of visceral afferents from the ratpelvic nerve: a horseradish peroxidase study. J Comp Neurol226:238–245.
ess TJ, Gebhart GF (1987) Characterization of neuronal responsesto noxious visceral and somatic stimuli in the medial lumbosacralspinal cord of the rat. J Neurophysiol 57:1867–1892.
eugebauer V, Rumenapp P, Schaible HG (1996) Calcitonin gene-related peptide is involved in the spinal processing of mechanosen-sory input from the rat’s knee joint and in the generation andmaintenance of hyperexcitability of dorsal horn-neurons duringdevelopment of acute inflammation. Neuroscience 71:1095–1109.
ndarza AB, Ye Z, Hulsebosch CE (2003) Direct evidence of primaryafferent sprouting in distant segments following spinal cord injury inthe rat: colocalization of GAP-43 and CGRP. Exp Neurol184:373–380.
ascual JI, Insausti R, Gonzalo LM (1993) Urinary bladder innervationin male rat: termination of primary afferents in the spinal cord asdetermined by transganglionic transport of WGA-HRP. J Urol150:500–504.
erry MJ, Lawson SN, Robertson J (1991) Neurofilament immunore-activity in populations of rat primary afferent neurons: a quantitativestudy of phosphorylated and non-phosphorylated subunits. J Neu-
rocytol 20:746–758.
R
R
R
R
R
R
R
S
S
V
V
V
W
W
W
W
Y
Z
S. Hou et al. / Neuroscience 159 (2009) 369–379 379
abchevsky AG (2006) Segmental organization of spinal reflexesmediating autonomic dysreflexia after spinal cord injury. Prog BrainRes 152:265–274.
abchevsky AG, Sullivan PG, Scheff SW (2007) Temporal-spatialdynamics in oligodendrocyte and glial progenitor cell numbersthroughout ventrolateral white matter following contusion spinalcord injury. Glia 55:831–843.
ivero-Melian C, Grant G (1991) Choleragenoid horseradish peroxi-dase used for studying projections of some hindlimb cutaneousnerves and plantar foot afferents to the dorsal horn and Clarke’scolumn in the rat. Exp Brain Res 84:125–132.
ivero-Melian C, Rosario C, Grant G (1992) Demonstration of trans-ganglionically transported choleragenoid in rat spinal cord by im-munofluorescence cytochemistry. Neurosci Lett 145:114–117.
obertson B, Grant G (1985) A comparison between wheat germagglutinin-and choleragenoid-horseradish peroxidase as antero-gradely transported markers in central branches of primary sen-sory neurones in the rat with some observations in the cat. Neu-roscience 14:895–905.
obertson B, Grant G (1989) Immunocytochemical evidence for thelocalization of the GM1 ganglioside in carbonic anhydrase-contain-ing and RT 97-immunoreactive rat primary sensory neurons.J Neurocytol 18:77–86.
obertson B, Perry MJ, Lawson SN (1991) Populations of rat spinalprimary afferent neurons with choleragenoid binding compared withthose labelled by markers for neurofilament and carbohydrate groups:a quantitative immunocytochemical study. J Neurocytol 20:387–395.
ann H, McCarthy PW, Jancso G, Pierau FK (1995) RT97: a markerfor capsaicin-insensitive sensory endings in the rat skin. Cell Tis-sue Res 282:155–161.
engupta JN, Gebhart GF (1994) Characterization of mechanosensi-tive pelvic nerve afferent fibers innervating the colon of the rat.
J Neurophysiol 71:2046–2060.alentino RJ, Kosboth M, Colflesh M, Miselis RR (2000) Transneuro-nal labeling from the rat distal colon: anatomic evidence for regu-lation of distal colon function by a pontine corticotropin-releasingfactor system. J Comp Neurol 417:399–414.
era PL, Nadelhaft I (1990) Conduction velocity distribution of afferentfibers innervating the rat urinary bladder. Brain Res 520:83–89.
izzard MA (2000) Increased expression of spinal cord Fos proteininduced by bladder stimulation after spinal cord injury. Am JPhysiol Regul Integr Comp Physiol 279:R295–R305.
ang HF, Shortland P, Park MJ, Grant G (1998) Retrograde andtransganglionic transport of horseradish peroxidase-conjugatedcholera toxin B subunit, wheatgerm agglutinin and isolectin B4from Griffonia simplicifolia I in primary afferent neurons innervatingthe rat urinary bladder. Neuroscience 87:275–288.
eaver LC, Cassam AK, Krassioukov AV, Llewellyn-Smith IJ (1997)Changes in immunoreactivity for growth associated protein-43suggest reorganization of synapses on spinal sympathetic neuronsafter cord transection. Neuroscience 81:535–551.
eaver LC, Verghese P, Bruce JC, Fehlings MG, Krenz NR, MarshDR (2001) Autonomic dysreflexia and primary afferent sproutingafter clip-compression injury of the rat spinal cord. J Neurotrauma18:1107–1119.
ilson P, Kitchener PD (1996) Plasticity of cutaneous primaryafferent projections to the spinal dorsal horn. Prog Neurobiol 48:105–129.
oshimura N, Erdman SL, Snider MW, de Groat WC (1998) Effects ofspinal cord injury on neurofilament immunoreactivity and capsaicinsensitivity in rat dorsal root ganglion neurons innervating the uri-nary bladder. Neuroscience 83:633–643.
agon A, Smith AD (1993) Monosynaptic projections from the rostralventrolateral medulla oblongata to identified sympathetic pregan-
glionic neurons. Neuroscience 54:729–743.(Accepted 5 December 2008)(Available online 24 December 2008)