intraspecific variability in nestling growth and fledging behavior of cassin's auklets at...
TRANSCRIPT

The Condor 991361-371 0 The Cooper Ornithological Society 1997
INTRASPECIFIC VARIABILITY IN NESTLING GROWTH AND FLEDGING BEHAVIOR OF CASSIN’S AUKLETS AT
TRIANGLE ISLAND, BRITISH COLUMBIA’
YOLANDA E. MORBEY AND RONALD C. YDENBERG Behavioral Ecology Research Group, Department of Biological Sciences, Simon Fraser University, Burnaby,
B.C., V5A I.%, Canada, e-mail: [email protected]
Abstract. We compared nestling growth and fledging behavior of Cassin’s Auklets (Pry- choramphus aleuticus) at Triangle Island, British Columbia to the predictions of a model that considers the timing of fledging to be an adaptive strategy. In the model, fledging mass and age depend on nestling growth rate, time remaining in the season, and the contrasting mortality costs and growth benefits experienced before and after fledging. As predicted, fast growing nestlings fledged heavier and younger than slow growing nestlings. Growth rates declined over the season and fledging behavior varied accordingly. When the seasonal vari- ation in growth rates was statistically controlled, late nestlings did not fledge lighter and younger, in contrast to the model’s predictions. Late in the season, nestlings reached a greater peak mass than expected based on their slower growth rates. Also, nestlings that grew more slowly due to a higher frequency of handling reached a higher peak mass than less frequently measured nestlings. We consider the possibility that parents adaptively modified their nes- tling’s growth trajectory by altering provisioning behavior in response to nestling condition. The nesting habitat influenced fledging. Fast growing nestlings fledged at similar ages in both level and steep nest sites. In contrast, slow growing nestlings fledged at younger ages, but similar masses, in level sites compared to steep sites. We consider the possibility that the difference in parental predation risk between level and steep nest sites influenced pro- visioning decisions of parents, and consequently, fledging decisions of nestlings.
Key words: fledging, nestling growth, Cassin’s Auklet, Triangle Island.
INTRODUCTION
In the family Alcidae, when nestlings leave the nest and go to sea, they encounter a different set of ecological conditions. An ontogenetic niche shift such as this typically involves a change in growth rate and mortality risk (Werner 1986, 1988). A tradeoff exists if one niche offers fast growth but high mortality, whereas the other of- fers low mortality but slow growth. This tradeoff is considered to hold for alcids, because the nest is assumed to be relatively safer than the ocean; however, juvenile growth rates in the nest are lower, due to relatively long provisioning trips by parents (Ydenberg 1989 and references there- in). The optimal condition (usually mass) and time to shift niches can be calculated by an al- gorithm that balances the fitness benefits of growing to a larger size against the costs of mor- tality (Ludwig and Rowe 1990, Rowe and Lud- wig 1991). Predictions about how fledging mass and age should vary with habitat-specific growth and mortality rates and with time remaining in the season have been generated by models of
’ Received 17 June 1996. Accepted 5 December 1996.
this type (Ydenberg 1989, Ydenberg et al. 1995).
The basic fledging model calculates optimal fledging mass and age under the following as- sumptions. First, juvenile growth trajectories dif- fer between the nest and ocean. Juveniles grow slowly while in the nest, but grow more quickly after fledging. Second, juveniles risk greater mortality in the ocean than in the nest. Third, the season during which growth is possible is limited in duration. Finally, survival to breeding depends on the final mass attained at the end of the season. The model explains two frequently reported patterns of intraspecific variation in fledging behavior (reviewed in Ydenberg et al. 1995). First, later in the season, nestlings fledge lighter and at younger ages because they take advantage of the higher growth rate in the ocean at the risk of higher mortality. When time avail- able for growth is limited, individuals facing greater time pressure are expected to take great- er risks (Clark 1994). Second, fast growing nest- lings are expected to fledge heavier and at youn- ger ages. This prediction was generated by vary- ing the nestling growth rate parameter in the model.
[3611

362 YOLANDA E. MORBEY AND RONALD C. YDENBERG
Parental provisioning decisions based on the costs and benefits of provisioning nestlings also can affect the optimal fledging time (Ydenberg 1989, Clark and Ydenberg 1990a, Harfenist and Ydenberg 1995). Specifically, in areas of a col- ony that are more dangerous to parents provi- sioning nestlings, nestlings are expected to fledge sooner, and consequently at lower masses.
An alternative hypothesis is that the seasonal fledging mass decline is a direct consequence of slower growth rates, caused by a seasonal de- cline in food availability, egg size, temperature, the effort invested by parents (Gaston 1985), or by the delayed breeding of poorer quality par- ents (Hatchwell 1991). Parental quality and food availability definitely influence the growth rate and fledging of nestlings (Gaston et al. 1983, Wehle 1983, Barrett et al. 1987, Emslie et al. 1992) and it follows logically that slower grow- ing nestlings will be smaller at a given age. However, because fledging age can vary, a pos- itive correlation between nestling growth rate and fledging mass is not a necessary conse- quence of slow growth under this hypothesis. A significant contribution of the fledging model is that it provides an explicit explanation for the positive correlation between nestling growth rate and fledging mass, based on nestlings using a condition- and time-dependent fledging strategy.
Although the assumptions of the fledging model are still debated, comparative (Ydenberg et al. 1995) and experimental (Harfenist 1995, Harfenist and Ydenberg 1995) studies support its predictions. From natural observations and experimental manipulation of Rhinoceros Auklet (Cerorhirtcu monocerata) nestling growth rate, Harfenist (1995) demonstrated that faster grow- ing nestlings fledged heavier and younger. Also, despite an absence of a seasonal decline in growth rates, nestlings fledged lighter and youn- ger later in the season. In areas of the colony with relatively high predation risk to parents, nestlings fledged sooner (Harfenist and Yden- berg 1995).
To determine whether the model could be generalized to another semi-precocial species, we studied nestling growth and fledging param- eters of Cassin’s Auklets (Pfychorumphus aleu- ticus). Information from previous accounts of Cassin’s Auklets’ growth and fledging behavior is insufficient to support or reject the fledging model, primarily because the relationships be- tween nestling growth rates, fledging parame-
ters, and time of year were not considered mul- tivariately. In addition to documenting the nat- ural variation in growth and fledging parameters, we also tested for an effect of growth rate on fledging parameters by exposing nestlings to one of two disturbance regimes. Increased distur- bance was expected to adversely affect nestling growth (Evans 1981, Harris and Wanless 1984 in Nettleship and Birkhead 1985, Harfenist 1991). We expected a higher mortality risk to adults on level nest sites, because escape from predators could be impaired (Nettleship 1972) and therefore we compared growth and fledging behavior between level and steep sites. Finally, we consider the consequences adaptive modifi- cation of growth trajectories has on the construc- tion and interpretation of the fledging model.
METHODS STUDY SITE AND STUDY SPECIES
Studies were conducted in 1994 on Triangle Is- land, located 45 km northwest of Cape Scott, Vancouver Island, British Columbia (50” 52’ N, 129” 05’ W, area = 44 ha, elevation = 194 m, figure in Vermeer et al. 1979). Triangle Island supports the world’s largest breeding colony of Cassin’s Auklets, with 547,000 breeding pairs (Rodway et al. 1990). Campbell et al. (1990) and Manuwal and Thoresen (1993) have re- viewed the geographical distribution, feeding behavior, breeding biology, phenology, and oth- er aspects of Cassin’s Auklet natural history. Cassin’s Auklets have a monogamous mating system, nest in burrows, and have a one-egg clutch. During the breeding season, adults feed offshore diurnally and visit the colony noctur- nally, delivering a load of zooplankton to nest- lings in a specialized throat pouch. Food loads are regurgitated directly to the nestling. Nestling growth approximates a logistic growth function, except prior to fledging when nestlings typically lose mass (Sealy 1973, Vermeer and Cullen 1982, Morbey 1995). Nestlings fledge with com- pleted juvenal plumage at 39-57 days old and 65-100% of mean adult body mass (Manuwal 1974, Vermeer 1981, 1987, Ainley et al. 1990). Glaucous-winged Gulls (Lams glaucescens) prey on Cassin’s Auklet nestlings (Rodway et al. 1990), and circumstantial evidence suggests that Bald Eagles (Haliaeetus leucocephalus) and Peregrine Falcons (Falco peregrinus) are impor- tant predators on adults (Rodway et al. 1990, Morbey, pers. observ.).

GROWTH AND FLEDGING BEHAVIOR OF CASSIN’S AUKLETS 363
SAMPLING PROTOCOL
Five study sites were delineated within areas of the Cassin’s Auklet colony that had high densi- ties of burrows, were distinct from the Rhinoc- eros Auklet and Tufted Puffin (Fratercula cir- rhata) colonies, and were easily accessible from the shore. In the southwest slope region of the colony, one site was located on level ground, whereas three sites were located on the lower steep slopes. An additional level site was located in the west slope region of the colony. During the incubation period, we selected 238 burrows for excavation based on signs of current occu- pancy such as worn entrances or fecal matter. Excavation consisted of digging vertical holes to allow access to all areas of the burrow. Access holes were patched with square-cut shingles and covered with soil and vegetation to reduce ero- sion. A total of 73 burrows were excavated on the two ‘level’ sites and 165 on the three ‘steep’ sites.
Egg length and egg width were measured to the nearest 0.1 mm with vernier calipers. Start- ing on lo-12 May, we checked each burrow ev- ery three days until the egg hatched. We also recorded whether the egg was missing, cold, or depredated by a rodent, identifiable by the egg- shell remains. We continued to visit burrows with cold eggs until they disappeared or were depredated.
We estimated hatchling age (O-3 days) based on each nestling’s size at first encounter. The range of hatchling wing chord lengths (16.0- 22.2 mm) was categorized into hatchling age classes. For wing chord lengths of 16.0-17.8 mm, hatchlings were considered 0 days old; 17.9-18.9 mm, 1 day old; 19.0-19.9 mm, 2 days old; and 20.0-22.2 mm, 3 days old. Six wet, downy nestlings considered newly hatched upon discovery had a mean wing chord length that fell within the O-days-old class (17.6 ? 0.6 mm, iz = 6). Although some variation in size at first measurement is due to hatchling size, we believe most variation is due to age. Hatching date is the date at which nestlings were first measured minus hatchling age.
Nestlings were measured on one of two dis- turbance schedules to assess whether increased handling affected nestling growth and fledging parameters. Half the nestlings were measured frequently (at hatching, 5 days of age, then ev- ery fifth day until fully feathered, and then every
second day until fledged) and half the nestlings were measured less often (at hatching, 5 days of age, 25 days of age, then every fifth day until fully feathered, and then every second day until fledged). The latter nestlings were not handled during the period of linear growth between 5 and 25 days. The former treatment will be referred to as the high disturbance (HD) group, the latter as the low disturbance (LD) group. All nestlings were banded with a U.S. Fish and Wildlife Ser- vice stainless steel band at 25 days of age. Nest- lings were measured between 09:OO and 15:00 PST, weather permitting. While handling nest- lings, none regurgitated their meals, indicating that no meals were lost due to handling.
We considered a nestling ‘fledged’ if it was absent from its burrow and was older than 30 days. There was no direct evidence of predation on nestlings of this age. We confirmed most de- partures by a subsequent nest check, usually on the next day but occasionally two days later. Only two out of 139 burrows still had a nestling at the confirmation check. During handling, we also checked whether nestlings were banded to be sure they were not visiting from a nearby burrow.
At each burrow visit, nestling mass was mea- sured to the nearest 0.5 g (for masses < 50 g) or 1 g (for masses > 50 g) using a Pesola spring scale. Flattened wing chord length was mea- sured to the nearest 0.1 mm (for wing lengths < 25 mm) with vernier calipers or 1 mm (for wing lengths > 25 mm) with a ruler. The number of ticks on the plantar surface of both webs was recorded (Morbey 1996).
Of the initial 238 burrows, 157 nestlings fledged (73 in the HD group, 79 in the LD group; 5 with a condition termed ‘shut-eye’). Nestlings with ‘shut-eye’ experienced weight loss and a general weakening accompanied by permanently shut eyes. The cause of their symp- toms is unknown. Seventy burrows failed be- cause of egg predation or abandonment (48) or nestling death (22). Eleven nestlings were not followed to fledging because of the threat of bur- row collapse or because it was too late in the season (‘stopped’).
DATA ANALYSES
For the analyses of breeding success and phe- nology, the ‘stopped’ burrows are excluded. For the analysis of egg size variation, all burrows are included. The growth and fledging analyses

364 YOLANDA E. MORBEY AND RONALD C. YDENBERG
include HD and LD nestlings that fledged, but excludes the five nestlings with ‘shut-eye.’
Hatching success is (n eggs hatched/n eggs laid).lOO%; fledging success is (n nestlings fledged/n eggs hatched). 100%. Egg mortality in- cludes cold, abandoned eggs, missing eggs, and egg-shell fragments. Nestling mortality includes dead and missing nestlings. Seasonal variation in nestling mortality was analyzed with chi- square tests: for each live day period over the entire hatching interval, the numbers of success- ful and unsuccessful burrows were compared. Egg mortality and nestling mortality were com- pared between habitats (level vs. steep) using chi-square analysis.
We compared egg size (egg length and egg width) between habitats using multivariate anal- ysis of variance (MANOVA). A significant MANOVA indicates that egg size differs be- tween habitats, but in order to describe how egg length and egg width are affected differentially by habitat, univariate analyses of variance (AN- OVAs) are required. If a MANOVA is signifi- cant, ANOVAs on the same data may not pro- vide as convincing evidence of significance and therefore, the significance values generated by the latter tests can be treated more liberally than usual.
We compared egg size between ‘fates’ by the same procedure. ‘Fate’ has three levels: eggs that did not hatch, successfully hatched eggs with nestlings that fledged, and successfully hatched eggs with nestlings that died. Since ‘fate’ had more than two levels, we used a mul- tiple comparison test (an option in the ANOVA) to compare egg length and egg width between the different fates (SAS Institute 1989).
The variables fledging age, fledging mass, and fledging wing length are the last recorded age, mass, and wing chord length prior to fledging. Nestlings lose mass prior to fledging, thus peak (maximum) mass and age also were considered. Daily growth rates were estimated for the linear phase of growth (Harfenist 1995) between the ages of 5 and 25 days. Mass growth rate equals (mass at day 25 - mass at day S)/(age at day 25 - age at day 5). Similarly, wing growth rate equals (wing length at day 25 - wing length at day S)/(age at day 25 - age at day 5).
We analyzed growth and fledging behavior by focusing on the following three questions: (1) how does mass growth rate influence fledging behavior, (2) how does disturbance regime affect
fledging behavior, and (3) how does fledging be- havior vary over the season? In this framework, mass growth rate, treatment, habitat, and hatch- ing date are the independent variables; fledging age, fledging mass, fledging wing length, peak age, and peak mass are the dependent variables. We first tested the full model using MANOVA (i.e., how do the independent variables influence the dependent variables?). All the interaction ef- fects between the independent variables were in- cluded. Nonsignificant interaction effects were sequentially dropped from the full model, fol- lowed each time by reanalysis. A significant in- teraction effect means that the relationship be- tween a continuous variable (mass growth rate or hatching date) differs between the levels of a class variable (treatment and/or habitat).
To explain the variation in fledging behavior when a significant interaction effect is retained in the selected model, reanalysis by MANOVA was required for each level of the class variable that is responsible for the interaction. When a MANOVA was significant, a series of linear re- gression tests provided information about how each fledging parameter varied with growth rate or hatching date. Analysis of covariance (AN- COVA) was required if a class variable in the selected MANOVA was significant.
We fitted a linear equation to describe the re- lationship between mass growth rate and hatch- ing date, and partitioned the seasonal variation in fledging behavior into seasonal (hatching date) effects and growth rate effects. We then used multiple regression to assess the impact of each of these components on fledging behavior.
We used SAS statistical software for all anal- yses (SAS Institute 1989). The a-level was set at 0.05, t-tests are two-tailed, and F-statistics are based on partial (type III) sums of squares. We present means as t 2 SD (n), the Wilks’ A F- statistics for MANOVAs, and the slope (m) and coefficient of determination (R2) for significant regressions. For hatching date values used in analyses, 1 = 1 March.
RESULTS
TIMING OF BREEDING AND BREEDING SUCCESS
Hatching dates (n = 169) ranged over 32 days (6 May-7 June) with a median hatching date of 17 May; fledging dates (n = 157) ranged over 40 days (13 June-23 July) with a median fledg- ing date of 1 July (Fig. 1). Neither hatching

GROWTH AND FLEDGING BEHAVIOR OF CASSIN’S AUKLETS 365
29-April 14-May 29-May 13-June 28-June 13-July 28-July
DATE
FIGURE 1. Distribution of hatching dates (before 10 June: level sites: n = 44, solid bars; steep sites: n = 125, shaded bars) and fledging dates (after 10 June: level sites: n = 45, solid bars; steep sites: n = 112, shaded bars). We could not estimate hatching date for ten burrows.
dates nor fledging dates differed between level and steep habitats (F,, ,67 = 0.08, P > 0.5 and F 1, 155 = 2.27, P = 0.1, respectively).
Overall hatching success was 79% and fledg- ing success was 88%. Fifteen of the initial 227 eggs experienced mortality before hatching checks began on 9 May. Egg mortality rate in- creased as the season progressed (Fig. 2). Most nestling mortality (79%) occurred before 10 days of age. Dead nestlings found in burrows accounted for 64% of total nestling mortality and none of these exhibited signs of predation. Nestling mortality did not vary over the season (x24 = 2.9, P > 0.5). Level and steep sites did not differ in egg mortality (26% vs. 19%, x21 = 1.7, P = 0.2) or nestling mortality (10% vs. 13%, x2, = 0.3, P > 0.5). Reproductive success did not differ between level and steep sites (66% vs. 70%, respectively; x2, = 0.4, P > 0.5).
EGG SIZE VARIATION
Mean egg length was 47.4 -C 1.8 mm (209); mean egg width was 34.1 + 1.1 mm (209). Egg size did not differ between the level and steep sites (F2, 206 = 0.92, P = 0.4), but did differ among eggs with different fates (F2. ,95 = 4.53, P = 0.01). The multiple comparison test indi- cated no difference in egg width between fates, but eggs that did not hatch were shorter [46.7 +- 1.6 mm (37)] than successfully hatched eggs [47.6 + 1.8 mm (161)], regardless of whether or not young fledged.
GROWTH AND FLEDGING PARAMETERS
Table 1 shows the summary statistics describing growth rates, the peak variables, and the fledg- ing variables. Table 2 summarizes the MANO- VA model selection procedure to analyze vari- ation in the peak and fledging variables.
0 1 O-May PO-May 30-May 09-June
DATE
1
0.8
0’ 0.6 F
8 0.4 8
:: 0.2
FIGURE 2. Egg mortality over the season. The solid bars indicate the number of incubated eggs during each five day period. The shaded bars indicate the proportion of egg mortality during each five day period.
366 YOLANDA E. MORBEY AND RONALD C. YDENBERG
TABLE 1. Summary statistics for growth rates and the peak and fledging variables for all nestlings.
Variable 5 2 SD(n)
Growth rate (g.day-‘) 4.49 f 0.69 (140) Wing growth rate (mm.day-‘) 2.82 % 0.41 (141) Peak age (days) 42 5 4 (147) Peak mass (g) 171 -+ 12 (152) Fledging age (days) 462 3 (147) Fledging mass (g) 162 ? 12 (151) Fledging wing length (mm) 125 ? 4 (151)
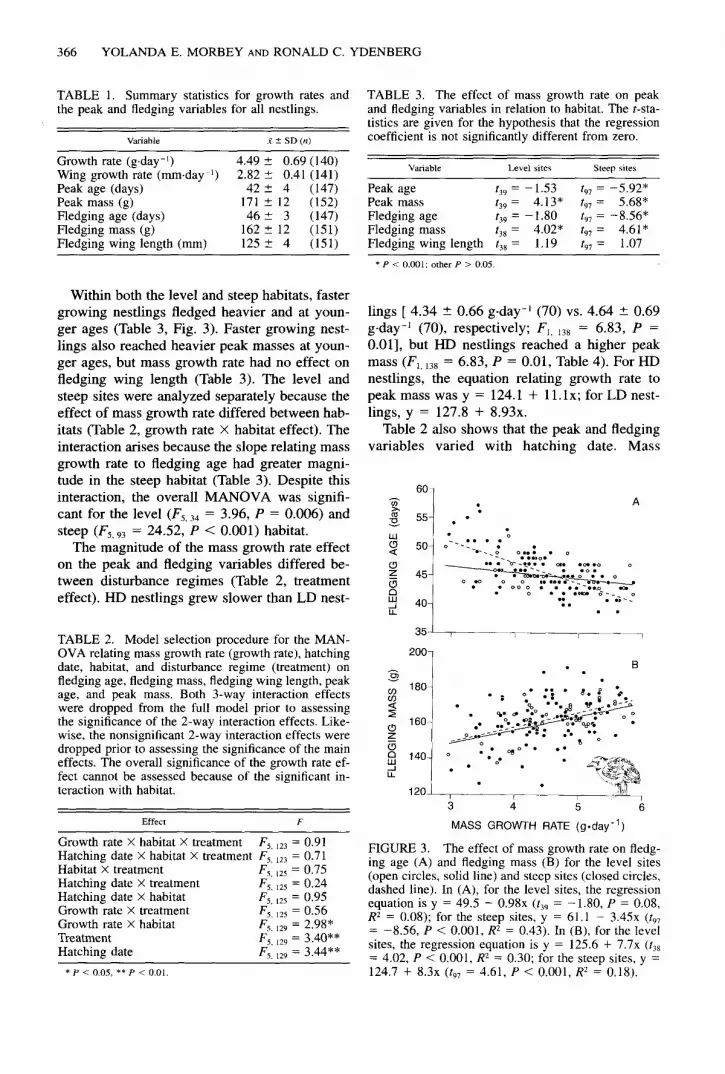
Within both the level and steep habitats, faster growing nestlings fledged heavier and at youn- ger ages (Table 3, Fig. 3). Faster growing nest- lings also reached heavier peak masses at youn- ger ages, but mass growth rate had no effect on fledging wing length (Table 3). The level and steep sites were analyzed separately because the effect of mass growth rate differed between hab- itats (Table 2, growth rate X habitat effect). The interaction arises because the slope relating mass growth rate to fledging age had greater magni- tude in the steep habitat (Table 3). Despite this interaction, the overall MANOVA was signifi- cant for the level (F5,34 = 3.96, P = 0.006) and
steep (Fs, 93 = 24.52, P < 0.001) habitat. The magnitude of the mass growth rate effect
on the peak and fledging variables differed be- tween disturbance regimes (Table 2, treatment effect). HD nestlings grew slower than LD nest-
TABLE 2. Model selection procedure for the MAN- OVA relating mass growth rate (growth rate), hatching date, habitat, and disturbance regime (treatment) on fledging age, fledging mass, fledging wing length, peak age, and peak mass. Both 3-way interaction effects were dropped from the full model prior to assessing the significance of the 2-way interaction effects. Like- wise, the nonsignificant 2-way interaction effects were dropped prior to assessing the significance of the main effects. The overall significance of the growth rate ef- fect cannot be assessed because of the significant in- teraction with habitat.
Effect F
Growth rate X habitat X treatment F5, ,23 = 0.91 Hatching date X habitat X treatment F5, ,23 = 0.71 Habitat X treatment F 5.125 = 0.75 Hatching date X treatment F 5. 125 = 0.24 Hatching date X habitat F 5, 125 = 0.95 Growth rate X treatment F 5. 125 = 0.56 Growth rate X habitat F 5, ,29 = 2.98* Treatment F 5, ,Z9 = 3.40** Hatching date F 5, ,pJ = 3.44**
* P < 0.05, ** P < 0.01.
TABLE 3. The effect of mass growth rate on peak and fledging variables in relation to habitat. The t-sta- tistics are given for the hypothesis that the regression coefficient is not significantly different from zero.
Variable Level sites Steep sites
Peak age t 39 = -1.53 t9, = -5.92* Peak mass t,, = 4.13* t9, = 5.68* Fledging age 139 = -1.80 t9, = -8.56* Fledging mass t,8 = 4.02* t9, = 4.61* Fledging wing length t,, = 1.19 t9, = 1.07
* P < 0.001: other P > 0.05
lings [ 4.34 ? 0.66 g.day-r (70) vs. 4.64 2 0.69 g.day-’ (70), respectively; F,, ,38 = 6.83, P = 0.011, but HD nestlings reached a higher peak mass (F,, ,38 = 6.83, P = 0.01, Table 4). For HD nestlings, the equation relating growth rate to peak mass was y = 124.1 + 11.1x; for LD nest- lings, y = 127.8 + 8.93x.
Table 2 also shows that the peak and fledging variables varied with hatching date. Mass
60 3 1 . A 2 .
p_ 55 :
LL 35L, l * ? 2001
. 0. B
.
3 4 5
MASS GROWTH RATE (g-clay-‘)
FIGURE 3. The effect of mass growth rate on fledg- ing age (A) and fledging mass (B) for the level sites (open circles, solid line) and steep sites (closed circles, dashed line). In (A), for the level sites, the regression equation is y = 49.5 - 0.98x (t39 = - 1.80, P = 0.08, RZ = 0.08); for the steep sites, y = 61.1 - 3.45x (t9, = -8.56, P < 0.001, R* = 0.43). In (B), for the level sites, the regression equation is y = 125.6 + 7.7x (t,* = 4.02, P < 0.001, R2 = 0.30; for the steep sites, y = 124.7 + 8.3x (t9, = 4.61, P < 0.001, RZ = 0.18).

GROWTH AND FLEDGING BEHAVIOR OF CASSIN’S AUIUETS 361
TABLE 4. ANCOVA to determine how the magni- tude of the growth rate effect on the peak and fledging variables differed between disturbance regimes (HD vs. LD). For each independent variable, the F-statistic and significance of the treatment effect are presented, based on the statistical model including mass growth rate and treatment as dependent variables. The MAN- OVA in Table 2 indicates no interaction between growth rate and treatment.
Vanable
Peak age Peak mass Fledging age Fledging mass Fledging wing length
* P < O.(H)l: other P > 0.05.
F
F 2. L37 = 1.33 F 2. 137 = 12.21* F 2. 137 = 1.03 F 2, 136 = 3.22 F 2,136 = I.96
growth rate influenced the peak and fledging variables (Table 2), and because mass growth rate declined with hatching date (Fig. 4), some seasonal variation in the peak and fledging vari- ables was expected. In particular, with slower mass growth rates later in the season, fledging age and peak age should increase and fledging mass and peak mass should decrease. The ef- fects were in the expected direction, but were significant only for fledging age (Table 5, Model 1). A hatching date effect was found only by statistically controlling for the effect of mass growth rate. This analysis (Table 5, Model 2) indicates that later in the season, nestlings reached higher peak masses than expected given their mass growth rates. In contrast, the seasonal increase in fledging age could be attributed to the seasonal decline in mass growth rates.
There also was seasonal variation in fledging wing length. With faster wing growth rate, nest- lings had longer wings at fledging (t,,, = 5.85, P < 0.001, m = 4.5, R* = 0.20), and both wing growth rate and fledging wing length declined over the season (f,39 = -7.12, P < 0.001, m = -0.04, R* = 0.27, and Table 5). However, the seasonal variation in fledging wing length did not differ from that expected based on the sea- sonal decline in wing growth rates (t,37 = 0.12, P > 0.5).
DISCUSSION
The observed natural variation in nestling growth rates and fledging mass and age matched the predictions of the fledging model: fast grow- ing nestlings fledged heavier and younger than slow growing nestlings. Fast growing nestlings
HATCHING DATE
FIGURE 4. Mass growth rate vs. hatching date for all nestlings in both treatments (y = 8.15 - 0.05x, tliB = -4.55, P < 0.001, RZ = 0.13).
also reached a higher peak mass at a younger age.
On the basis of the fledging model, we ex- pected late nestlings to fledge lighter and youn- ger because of the impending onset of winter. However, we observed a seasonal increase in fledging age, which was attributable to the sea- sonal decline in mass growth rates. We believe that the seasonal decline in growth rates had in- sufficient magnitude to cause detectable season- al variation in fledging mass. In contrast to our expectations, neither fledging age nor fledging mass varied with hatching date independently of the growth rate effect. The cause of the seasonal decline in growth rates is unresolved (Morbey 1995). Other studies indicate that low growth rates are associated not only with poor feeding conditions (Wehle 1983, Barrett et al. 1987), but also with young or inexperienced parents (Gas- ton et al. 1983); the seasonal decline observed
TABLE 5. Statistics for two regression models to sep- arate the effects of hatching date and mass growth rate on the peak and fledging variables. Model 1 is the effect of hatching date and model 2 is the effect of hatching date and growth rate. The t-statistics are giv- en for the hypothesis that the regression coefficient of the hatching date effect is not significantly different from zero. Mass growth rate did not affect fledging wing length; therefore, we did not partial out an effect of mass growth rate.
Variable Model I Model 2
Peak age t,‘ls = 1.56 t,,, = -0.18 Peak mass 1145 = -0.63 t,,, = 2.36* Fledging age t&45 = 2.16* t,,, = -0.29 Fledging mass r144 = -1.28 t,,, = 1.15 Fledging wing length t,, = -2.93**
*P 4 0.05. ** P < 0.0,

368 YOLANDA E. MORBEY AND RONALD C. YDENBERG
here could be due to a combination of these fac- tors.
The mismatch between some of the predic- tions of the fledging model and the observations of Cassin’s Auklets could mean that the assump- tions of the model were not met. Our observa- tions support the assumption that nestling mor- tality is negligible near fledging, because most nestling mortality occurred much earlier. In the fledging model, optimal fledging mass and age are calculated in relation to a defined logistic growth trajectory. This function is perhaps too simplistic when the growth trajectory is flexible (Clark and Ydenberg 1990a) or when mass re- cession occurs. In our study, the assumption of a deterministic logistic growth trajectory was vi- olated, and we believe this has implications for supporting or rejecting the model.
Facultative adjustment of parental provision- ing behavior is thought to be especially impor- tant near the end of the nestling’s growth period. Recent models of fledging behavior in alcids suggest a conflict over the duration of parental care between parents and offspring late in the nestling period (Ydenberg 1989, Clark and Ydenberg 1990a, 1990b). From the parents’ per- spective, when visiting the colony is dangerous and the marginal return in terms of nestling growth from provisioning beyond a minimum requirement is sufficiently low, then termination or reduction of provisioning may result (Bertram 1988, Ydenberg 1989, Harfenist and Ydenberg 1995). From the nestling’s perspective, near the end of the nestling period, the benefits of staying and remaining safe in the nest must be consid- ered against the risk of losing too much weight.
A fascinating feature of our data is the sug- gestion that parents adaptively modified nest- lings’ growth trajectories. If intraspecific varia- tion in the shape of nestlings’ growth trajectories results from adult decisions about when and how much to provision, the optimal fledging mass and age combination could be reached by mod- ifying the growth trajectory. Three separate ob- servations support this view. First, nestlings that grew slower as a result of increased handling reached a higher peak mass than nestlings that were not handled during the period of maximum growth. The significant treatment effect in the MANOVA and the highly significant treatment effect on peak mass in the ANCOVA argue against this being a spurious result. Parents may have compensated for the poorer growth of han-
dled nestlings by feeding them to a higher peak mass, which in effect would accelerate fledging.
Second, late nestlings reached a larger peak mass than expected given their growth rates. Late season parents may have compensated by feeding slow growing nestlings to a higher peak mass. The negative effect of hatching date on fledging age when growth rate variation was controlled, although not significant, is consistent with this idea. However, other theories also might account for the observed seasonal varia- tion in fledging mass and age. If poorer quality parents breed later, modification of the growth trajectory may be a parental strategy to increase fitness regardless of a time limitation. A differ- ent explanation is that selection on synchrony is operating (Birkhead 1977, Gaston and Nettle- ship 1981). For example, contracting the range of fledging dates could function to swamp pred- ators, thereby reducing each fledgling’s risk of predation (Gaston and Nettleship 1981).
Third, fledging mass and age differed between level and steep habitats. Fast growing nestlings fledged at similar ages in both level and steep sites. In contrast, slow growing nestlings fledged at younger ages, but at similar masses, in level sites compared to steep sites. These patterns do not appear to be an artifact of sampling because the range of mass growth rates was similar be- tween habitats. We suggest this pattern resulted from parental adjustment of the growth trajec- tory under different predation risks: in the level (high predation) sites, parents may have invested less in nestlings near fledging, through reduction or termination of provisioning. To account for the similarity in fledging age for fast growers in both habitats, we suggest that parents invested the minimum time requirement in a fast growing nestling because the expected gain from invest- ment in a good condition nestling near fledging would be less than in a poor condition nestling.
Other studies have observed an effect of pre- dation on growth rates and/or fledging behavior (Nettleship 1972, Harris 1980, Wilson 1993, Harfenist and Ydenberg 1995); but only the Har- fenist and Ydenberg study attributes this to a facultative strategy. In areas of the study colony with high adult predation, Rhinoceros Auklet nestlings fledged sooner (Harfenist and Yden- berg 1995). Although not evident from their pre- sentation of the data, this pattern also was more pronounced for slow growing nestlings (A. Har- fenist, pers. comm.). Nettleship (1972) found

GROWTH AND FLEDGING BEHAVIOR OF CASSIN’S AUKLETS 369
that in regions of an Atlantic Puffin (Fratercula arctica) colony with less predation and less kleptoparasitism by gulls, parents provisioned nestlings more frequently and nestlings fledged heavier and younger.
Circumstantial evidence from the present study and corroborative evidence from others support our claim that the level and steep sites differed in adult predation risk. Cassin’s Auklets are clumsy on land and may be particularly sus- ceptible to Peregrine Falcons and Bald Eagles on level ground. In the Nettleship (1972) study on Great Island, Atlantic Puffins were particu- larly susceptible to attack by gulls on level ground because escape was slower. Bald Eagles tend to eat prey where they catch them and leave a characteristic pile of plucked feathers. On Tri- angle Island, many feather piles were found in level habitat but none in the steep habitat; how- ever, feather piles were observed only at the be- ginning of the season (Morbey, pers. observ.). The greater force of wind on the steep habitat may have caused feathers to blow away, but we suspect at least some feathers would be observed if predation occurred there.
In the absence of any accurate measure of adult predation risk, we considered whether hab- itat differences in breeding success could sub- stantiate the claim that level habitat posed great- er risk. However, we believe factors other than the mortality of parents caused most of the egg and nestling mortality because egg and nestling mortality was strongly time- and age-dependent. Egg mortality was highest late in the season. We assume predation of eggs by rodents followed neglect or desertion of eggs (it is doubtful that a mouse or vole could chase an adult from its egg). Other causes of abandonment, such as em- bryo mortality, are not important in Cassin’s Auklets (Ainley et al. 1990). We suspect rodents specialized on eggs when eggs were abundant, taking advantage of increased egg neglect and desertion late in the year. Egg neglect occurs more frequently early and late during incubation (Astheimer 1991), and Emslie et al. (1992) found breeding experience increases the hatch- ing success of Cassin’s Auklets. If inexperienced parents delayed breeding, desertion would occur more frequently late in the year. Most nestling mortality occurred in young nestlings; we sug- gest most nestling mortality resulted from star- vation, failure to thermoregulate, or disease. Adult predation may have a slight contribution
to abandonment of eggs and starvation of nest- lings, but adult predation is unlikely to be cor- related with the developmental stage of the egg or nestling.
We cannot discount the possibility that the habitat differences in the fledging parameters re- sulted from another factor, such as parental qual- ity. The only measure we have with potential relevance to parental quality is egg size, but we are uncertain of its efficacy as an index. In Ra- zorbills (Alca to&z) and Gannets (S&u bussun- US), older parents lay larger eggs (Nelson 1966, Lloyd 1979), but this has not been documented in Cassin’s Auklets. Furthermore, in this study, egg volume had a minute influence on growth rates and did not vary over the season (Morbey 1995). Depredated or abandoned eggs were shorter than successfully hatched eggs, but this does not prove poorer quality parents laid short- er eggs, because rodents may depredate shorter eggs with greater success. Finally, neither egg length nor egg width differed between habitats.
Growth rate affected the fledging parameters of Cassin’s Auklets, supporting the predictions of the fledging model. The prediction that faster growing nestlings should fledge heavier and younger is the least controversial and most ro- bust component of the fledging model. The pre- diction that the fledging parameters should vary seasonally cannot yet be rejected. To explain the seasonal variation in the fledging parameters, it is necessary to determine if poorer quality par- ents are breeding later, how important synchrony of fledging is, and how and why individuals vary in their growth rate trajectories. We suggest that facultative adjustment of nestling growth trajectories in response to nestling condition and predation risk is an important and understudied area. Another area for future study is to assess whether the mass recession phenomenon results from parental provisioning decisions and nest- ling fledging decisions.
ACKNOWLEDGMENTS
We thank Tasha Smith and Jasper Lament who helped with the field work and data entry. Logistical support while in the field was provided by the Canadian Wild- life Service/Natural Science and Engineering Research Council of Canada (NSERC)/Simon Fraser University (SFU) Research Chair in Wildlife Ecology. We thank the crew of the Canadian Coast Guard vessel Narwhal for transportation from Victoria and Ecological Re- serves (B.C. Parks) for allowing access to Triangle Is- land. Brett Sandercock offered helpful comments on

370 YOLANDA E. MORBEY AND RONALD C. YDENBERG
an earlier draft. We also thank Steve Emslie and an anonymous reviewer for constructive suggestions to improve the manuscript. The research was supported by an Anne Vallte Ecological Scholarship and an SFU Graduate Fellowship to YEM and by an NSERC op- erating grant to RCY.
LITERATURE CITED
AINLEY, D. G., R. J. BOEKELHEIDE, S. H. MORRELL, AND C. S. STRONG. 1990. Cassin’s Auklet, p. 306-338. In D. G. Ainley and R. J. Boekelheide [eds.], Sea- birds of the Farallon Islands. Ecology, dynamics and structure of an upwelling-system community. Stanford Univ. Press, Stanford, CA.
ASTHEIMER, L. B. 1991. Embryo metabolism and egg neglect in Cassin’s Auklets. Condor 93:486-493:
BARRETT. R. T.. T ANKER-NILSSEN. I? RIKARDSEN. K. VALDE, N. Rov, AND W. VADER. 1987. The food, growth, and fledging success of Norwegian Puffin chicks Fratercula arcticu in 1980-1983. Ornis Stand. 18:73-83.
BERTRAM, D. E 1988. The provisioning of nestlings by parent Rhinoceros Auklets (Cerorhincu monocer- afu). M.Sc. thesis, Simon Fraser Univ., Burnaby, B.C., Canada.
BIRKHEAD, T R. 1977. Adaptive significance of the nestling period of Guillemots Uris aalge. Ibis 119544-549.
CAMPBELL, R. W., N. K. DAWE, I. MCTAGGART-COWAN, J. M. COOPER, G. W. KAISER, AND M. C. E. MCNALL. 1990. The birds of British Columbia. Vol. II. Nonpasserines. Roy. Br. Columbia Mus., Victoria, B.C., Canada.
CLARK, C. W. 1994. Antipredator behavior and the as- set-protection principle. Behav. Ecol. 5:159-170.
CLARK, C. W., AND R. C. YDENBERG. 1990a. The risks of parenthood. I. General theory and applications. Evol. Ecol. 4:21-34.
CLARK, C. W., AND R. C. YDENBERG. 1990b. The risks of parenthood. II. Parent-offspring conflict. Evol. Ecol. 4:312-325.
EMSLIE, S. D., W. J. SYDEMAN, AND I? PYLE. 1992. The importance of mate retention and experience on breeding success in Cassin’s Auklet (Ptychorum- phus aleuticus). Behav. Ecol. 3:189-195.
EVA& F! G. H. 1981. Ecology and behaviour of the Little Auk Alle alle in west Greenland. Ibis 123: l-18.
GASTON, A. J. 1985. Development of the young in the Atlantic Alcidae, p. 319-354. In D. N. Nettleship and T R. Birkhead [eds.], The Atlantic Alcidae. Academic Press, Toronto.
GASTON, A. J., G. CHAPDELAINE, AND D. G. NOBLE. 1983. The growth of Thick-billed Murre chicks at colonies in Hudson Strait: inter- and intra-colony variation. Can. J. Zool. 61:2465-2475.
GASTON, A. J., AND D. N. NE~LESHIP. 1981. The Thick-billed Murres of Prince Leopold Island-a study of the breeding ecology of a colonial high arctic seabird. Can. Wildl. Serv. Monogr. Series 6.
HARFENIST, A. 1991. Effects of growth rate and paren- tal predation risk on fledging of Rhinoceros Auk-
lets (Cerorhinca monocerata). M.Sc. thesis, Simon Fraser Univ., Burnaby, B.C., Canada.
HARFENIST, A. 1995. Effects of growth-rate variation on fledging of Rhinoceros Auklets (Cerorhinca monocerutu). Auk 112:60-66.
HARFENIST, A., AND R. C. YDENBERG. 1995. Parental provisioning and predation risk in Rhinoceros Auklets (Cerorhinca monoceratu): effects on nest- ling growth and fledging. Behav. Ecol. 6:82-86.
HARRIS, M. P 1980. Breeding performance of puffins Fruterculu urcticu in relation to nest density, lay- ing date and year. Ibis 122:193-209.
HATCHWELL, B. J. 1991. An experimental study of the effects of timing of breeding on the reproductive success of Common Guillemots (Uriu an&e). J. Anim. Ecol. 60:721-736.
LLOYD, C. S. 1979. Factors affecting breeding of Ra- zorbills Alcu tordu on Stockholm. Ibis 121: 165- 176.
LUDWIG, D., AND L. ROWE. 1990. Life-history strate- gies for energy gain and predator avoidance under time constraints. Am. Nat. 135:686-707.
MANUWAL, D. A. 1974. The natural history of Cassin’s Auklet (Ptychoramphus aleuticus). Condor 76: 421-431.
MANUWAL, D. A., AND A. C. THORESEN. 1993. Cassin’s Auklet (Ptychorumphus uleuticus). In A. Poole and E Gill [eds.], The birds of North America, No. 50. The Academy of Natural Sciences, Philadel- phia and The American Ornithologists’ Union, Washington, DC.
MORBEY, Y. E. 1995. Fledging variability and the ap- plication of fledging models to the behaviour of Cassin’s Auklets (Ptychorumphus uleuticus) at Triangle Island, British Columbia. M.Sc. thesis, Simon Fraser Univ., Burnaby, B.C., Canada.
MORBEY, Y. E. 1996. The abundance and effects of ticks (Ixodes uriue) on nestling Cassin’s Auklets (Ptychoramphus aleuticus) at Triangle Island, British Columbia. Can. J. Zool. 74: 1585-1589.
NELSON, J. B. 1966. The breeding biology of the Gan- net Sulu f~ussuna on the Bass Rock, Scotland. Ibis 108:584-626.
NE~LESHIP, D. N. 1972. Breeding success of the Com- mon Puffin (Fraterculu arctica L.) on different habitats at Great Island, Newfoundland. Ecol. Monogr. 42:239-268.
NETTLESHIP, D. N. AND 'I R. BIRKHEAD [EDS.]. 1985. The Atlantic Alcidae. Academic Press, Toronto.
RODWAY, M. S., M. J. E LEMON, AND K. R. SUMMERS. 1990. British Columbia seabird colony inventory: report #4-Scott Islands. Census results from 1982 to 1989 with reference to the Nestucca oil spill. Tech. Rep. Series No. 86. Can. Wildl. Serv., Pacific and Yukon Region, B.C.
ROWE, L., AND D. LUDWIG. 1991. Size and timing of metamorphosis in complex life cycles: time con- straints and variation. Ecology 72:413-427.
SAS INSTITUTE INC. 1989. SAS/STAT user’s guide. Version 6, 4th ed., Vol. 2. SAS Institute, Inc., Cary, NC.
SEALY, S. G. 1973. Adaptive significance of post- hatching developmental patterns and growth rates in the Alcidae. Ornis Stand. 4:113-121.

GROWTH AND FLEDGING BEHAVIOR OF CASSIN’S AUKLETS 371
VERMEER, K. 1981. The importance of plankton to Cassin’s Auklets during breeding. J. Plank. Res. 3:315-329.
VERMEER, K. 1987. Growth and nestling periods of Cassin’s Auklets: adaptations of planktivorous auklets to breeding at northern latitudes. Can. Tech. Rep. Hydrog. Ocean Sci. No. 93.
VERMEER, K., AND L. CULLEN. 1982. Growth compar- ison of a plankton- and a fish-feeding alcid. Murrelet 63:34-39.
VERMEER, K., R. A. VERMEER, K. R. SUMMERS, AND R. R. BILLINGS. 1979. Numbers and habitat selection of Cassin’s Auklet breeding on Triangle Island, British Columbia. Auk 96: 143-15 1.
WEHLE, D. H. S. 1983. The food, feeding and devel- opment of young Tufted and Homed Puffins in Alaska. Condor 85:427-442.
WERNER, E. E. 1986. Amphibian metamorphosis: growth rate. predation risk and the optimal size at transformat&. Am. Nat. 128:319-3‘41.
WERNER. E. E. 1988. Size. scaling and the evolution of complex life cycles, p. 60-81. In B. Ebenman and L. Persson [eds.], Size-structured populations. Springer-Verlag, Berlin.
WILSON, U. W. 1993. Rhinoceros Auklet burrow use, breeding success, and chick growth: gull-free vs. gull-occupied habitat. J. Field Ornithol. 64:256- 261.
YDENBERG, R. C. 1989. Growth mortality tradeoffs and the evolution of juvenile life histories in the Al- cidae. Ecology 76: 1494-1506.
YDENBERG. R. C.. C. W. CLARK. AND A. HARFENIST. 1995. ‘Intraspecific fledging &ass variation in the Alcidae, with special reference to the seasonal fledging mass decline. Am. Nat. 145:413-433.