interleukin-1 (il-1) directly and indirectly promotes hematopoietic cell growth through type i il-1...
TRANSCRIPT

Interleukin-l (IL-1) Directly and Indirectly Promotes Hematopoietic Cell Growth Through Type I IL-1 Receptor
By Kjetil Hestdal, Francis W. Ruscetti, Richard Chizzonite, Mariaestela Ortiz, John M. Gooya, Dan L. Longo, and Jonathan R. Keller
Interleukin-l (IL-l) has been shown to stimulate hematopoi- etic progenitor cell growth both in vitro and in vivo. Al- though IL-l alone lacks the ability to promote hematopoietic progenitor growth in vitro, it is a potent synergistic factor in combination with other colony-stimulating factors (CSFs). Because it was unknown whether type I (~80). type II (~68). or other IL-l-binding proteins mediated the synergistic ef- fects of IL-l on purified progenitor cells, we used the differ- ence in immunoreactivity between type I and type II IL-l receptor (IL-1R) to better assess the role of these receptors in hematopoietic progenitor growth. Therefore, the synergistic effects of IL-la on IL-3-, CSF-1-, and granulocyte macro- phage (GM)-CSF-induced progenitor growth, both in CFU- c and single-cell assays, were determined in the presence of monoclonal antibodies (MoAbs) 35F5 and 4E2 that block the binding of IL-la to type I and type II IL-lR, respectively. The synergistic effect of IL-la on IL-3 responsive Lin- and Lin”
NTERLEUKIN-1 (IL-1) has been shown to have pro- found stimulatory effects on hematopoietic cell growth
both in vitro and in vivo.”” Although IL-1 alone does not induce the proliferation and differentiation of hematopoietic progenitor cells in vitro, IL-1 synergizes with the colony- stimulating factors (CSFs) to promote the growth of bone marrow cells (BMCs) in soft-agar colony assays.l2-I6 For example, IL-1 is a necessary growth factor for the prolifera- tion of high-proliferative potential colony-forming cells, one of the most primitive progenitors detected in ~ i t r o . ’ ~ , ’ ~ Be- cause K- l has been shown to induce production of hemato- poietic growth factors (HGFs) in vitro and in it has been proposed that IL-l might indirectly promote the growth of hematopoietic progenitor cells in vitro. However, in sin- gle-cell assays using purified progenitors, we have recently shown that IL-1 can directly synergize with some HGFs to promote cell In this regard, it has been shown that IL-l can upmodulate the expression of CSF receptors on purified progenitor cells and thus could mediate its direct synergistic effects through this mechanism.”.2’
The biologic response of IL-1 is mediated through specific high-affinity cell-surface receptors, and two distinct gene products have been cloned.24325 The type I IL- 1 receptor (IL- 1R I; p80) is expressed on T cells, fibroblasts, and endothelial cells, whereas the type I1 IL-1R (IL-1R 11; p68) is expressed on B cells, granulocytes, and macrophage^.^^-^* As indicated above, although IL-1 has direct effects on the growth of immature hematopoietic progenitors cells, we have been un- able to detect the expression of IL- 1 Rs using radioiodinated IL- l a on these population^.^^ However, we and others have shown that mature myeloid cells express type I IL-lR,25.30 and that HGFs can upmodulate the expression of p68, IL- 1R I1 on purified progenitor cells.29
Type I and type I1 IL-1R show different immunoreactiv- it^,^' and can be distinguished using monoclonal antibodies (MoAbs) 35F5 and 4E2 that specifically block the binding of radioiodinated IL-1 to IL-1R I and 11, respectively. Al- though no biologic effects have been assigned to the type I1 IL-IR, it has been shown that MoAb 35F5 can inhibit IL-
I
Blood, Vol 84, No 1 (July l), 1994: pp 125-132
Thy-l+ progenitors was indirectly mediated and could be inhibited by MoAb 35F5. In contrast, IL-la directly syner- gized with CSF-1 and GM-CSF to promote progenitor cell growth. The direct synergistic effect of IL-la on CSF-l-in- duced progenitor growth was observed in all progenitor populations examined (Lin-, Lin-Thy-l+, and Lin-Thy-1-1 and was inhibited by MoAb 35F5. However, the direct syner- gistic effect of IL-la on GM-CSF-responsive progenitors, Lin- and Lin-Thy-l+, was partially inhibited by MoAb 35F5. In contrast, the MoAb antitype II IL-1R (MoAb G21 could not inhibit the direct synergistic effects of IL-la on CSF-1- or GM-CSF-induced progenitor growth. Thus, IL-la directly and indirectly stimulates the growth and differentiation of purified progenitors through the type I IL-1R but not the type
This is a US government work. There are no restrictions on its use.
II IL-1R.
1-induced production of CSFs from hematopoietic acces- sory cells in vitro and the production of CSFs in vivo, and block the radioprotective effect of IL-1 in vivo.”-” In addi- tion, we have shown that MoAb 35F5 inhibits the IL-la- mediated upmodulation of both IL- la and CSF receptors on purified BMCs in V~VO.’~,’~ Taken together, the indirect ef- fects of IL-la are most likely mediated through the type I
Therefore, to determine the type of IL-1Rs that mediates the direct proliferative effect of IL-1 on hematopoietic pro- genitors in vitro, we examined the synergistic effects of IL- la on BMCs in colony-formation assays and in single-cell assays in the presence or absence of the MoAbs 35F5 and 4E2.
IL- 1 R.
MATERIALS AND METHODS
Growth factors and anti-IL-IR antibodies. Purified recornbi- nant human IL-la, IL-10, and the MoAbs against type I and type
From the Laboratory of Molecular Immunoregulation, Biological Response Modifiers Program, and Biological Carcinogenesis and Development Program, Program Resources, Inc/DynCorp, National Cancer Institute-Frederick Cancer Research and Development Cen- ter, Frederick, MD; and the Department of Immunophannacology and Molecular Genetics, Hoffmann-LaRoche, Nutley, NJ.
Submitted September 16, 1993; accepted March 11, 1994. The content of this publication does not necessarily refect the
views or poiicies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organiza- tions imply endorsement by the US Government.
Address reprint requests to Jonathan R. Keller, PhD, Biological Carcinogenesis and Development Program, PRI/DynCorp, PO Box B, Frederick, MD 21702-1201.
The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 V.S.C. section 1734 solely to indicate this fact.
This is a US government work. There are M restrictions on its use. 0006-4971/94/8401-07$0.00/0
125

126 HESTDAL ET AL
I1 IL-IRs (35F5 and 4E2; both monoclonal rat IgG) were prepared at Hoffmann-LaRoche (Nutley, NJ). The activity of IL-la and IL- Io was 3 X 1 OB U/mg determined by thymocyte proliferation assay. Purified recombinant murine granulocyte macrophage (GM)-CSF was kindly provided by Dr Ian K. McNiece (Amgen Corp, Thousand Oaks, CA) and recombinant murine IL-3 was purchased from R& D Systems (Minneapolis, MN). Recombinant human CSF-I was a generous gift from Dr Michael Geier (Cetus Corp, Emeryville, CA).
BMCs. Normal murine BMCs were obtained by aspirating fe- murs of BALB/c mice with RPMI and then washed before resuspen- sion in Iscove's modified Dulbecco's Medium (IMDM) supple- mented with 10% fetal calf serum (FCS; Inovar, Gaithersburg, MD), 1% L-glutamine, and antibiotics (complete IMDM). The Lin- BMCs were purified by immunomagnetic bead separation according to a previously described protocol.35 Briefly, light-density BMCs were obtained by separating BMCs on lymphocyte separation medium (Organon Teknika Corp, Durham, NC). The light-density BMCs were incubated in complete IMDM at 4°C for 30 minutes with a cocktail of antibodies: B220 (CD45; Pharmingen, San Diego, CA), RB6-8C5 ((2-1; gift from Dr R.L. Coffman, DNAXCorp, Palo Alto, CA), MAC-l (Boeringer Mannheim, Indianapolis, IN), Lyt-2 (CD8), and L3T4 (CD4) (both purchased from Becton Dickinson & CO, Sunnyvale, CA). The cells were washed and resuspended in complete IMDM before incubation with immunomagnetic beads (Dynal, Great Neck, NY) at a ratio 40:l (beads/cell) for 30 minutes at 4°C. The Lint cells were removed by magnetic particle concentrator (Dynal), and Lin- cells were collected and washed in complete IMDM. Fur- ther purification of Lin- cells into Lin-Thy- l + and Lin-Thy- l ~ cells was performed by flow cytometric activated cell sorting (FACS) using biotinylated MoAbs anti-Thy-1.2 (Becton Dickinson). Lin cells were labeled with biotinylated anti-Thy-1.2 or isotype- matched control IgG (Pharmingen) and washed before streptavidin- fluorescein isothiocyanate were added. Lin-Thy-l+ and Lin-Thy-l- cells were then sorted using FACStar (Becton Dickinson).
Thymocyte proliferation assay. Thymus cells were obtained from 4- to 6-week-old normal C3H/HeN mice. The cells were washed in RPMI before being resuspended in complete IMDM. The cells, I X IO6 celldwell, were seeded in 96-well flat microtiter plates (Costar, Cambridge, MA) and incubated with phytohemagglutinin ( 1 yg/mL) and IL-la ( 1 ng/mL) in the presence or absence of MoAb 35F5 or control IgG for 72 hours at 37°C 5% CO?. The cells were pulsed with 1 yCi of 'H-thymidine (6.7 Ci/mmol; New England Nuclear, Boston, MA) for the last 12 hours. Cell cultures were harvested with a multiple-sample automated cell harvester on glass fiber filters, individual filters were placed in scintillation fluid, and radioactivity was assessed by liquid scintillation counting.
Colony growth. Lin-, Lin-Thy-l+, or Lin-Thy- BMCs were plated in 35-mm Lux Petri dishes (Miles Laboratories, Inc, Naper- ville, IL) in 1 mL of complete IMDM and 0.3% Seaplaque agarose ( M C Bioproducts, Rockland, ME) and supplemented with predeter- mined optimal concentrations of indicated HGFs. Dishes were incu- bated at 37°C in 5% CO2 for 6 to 8 days before scoring for colony growth (>50 cells). Single-cell proliferation assays were performed in Terasaki plates (Nunc, Kamstrup, Denmark) at a concentration of 1 cell per well in 20 pL of complete IMDM and indicated growth factors. For each determination a total of 300 cells were examined. The frequency number represents the number of responding progeni- tors divided by the number of single cells plated (300 cells). Wells were scored for proliferation (> I O cells/well) after 6 days of incuba- tion at 37°C and 5% CO2. MoAbs against IL-I -binding proteins or control IgG were preincubated with target cells for 45 minutes at 37°C before HGFs were added.
Radioiodinated cytokines and binding assay. Radioiodination of GM-CSF was performed by a modification of the chloramine-T method as described previously for IL-3.36 The specific radioactivi-
ties of '"1-GM-CSF ranged from 4.0 to 7.0 x 10' cpdpmol. There was no significant loss in biologic activity of cytokines after the iodination procedure. The binding assays were performed by a pre- viously described phthalate oil separation meth~d.~ ' Lin- cells were washed in RPMI 1640 and resuspended in 50 nmol/L glycine-hydro- chloric acid for 1 minute to release bound ligands. The cells were washed in binding medium (RPMI 1640 containing 2% bovine serum albumin, 20 mmol/L HEPES, and 0.1 % sodium azide) before radio- iodinated GM-CSF was added to 2 X IOh cells. After 75 minutes of incubation at 20"C, cell-bound rahoactivity was separated from unbound radioiodinated ligands by centrifugation of samples through a mixture of dibutyl phthalate and bis(2-ethyl-hexyl) phthalate oil (ratio 1 S : l ) (Eastman Kodak, Rochester, NY). Specific binding was measured as the difference between counts in the presence and ab- sence of 50-fold excess of cold competitor.
RESULTS
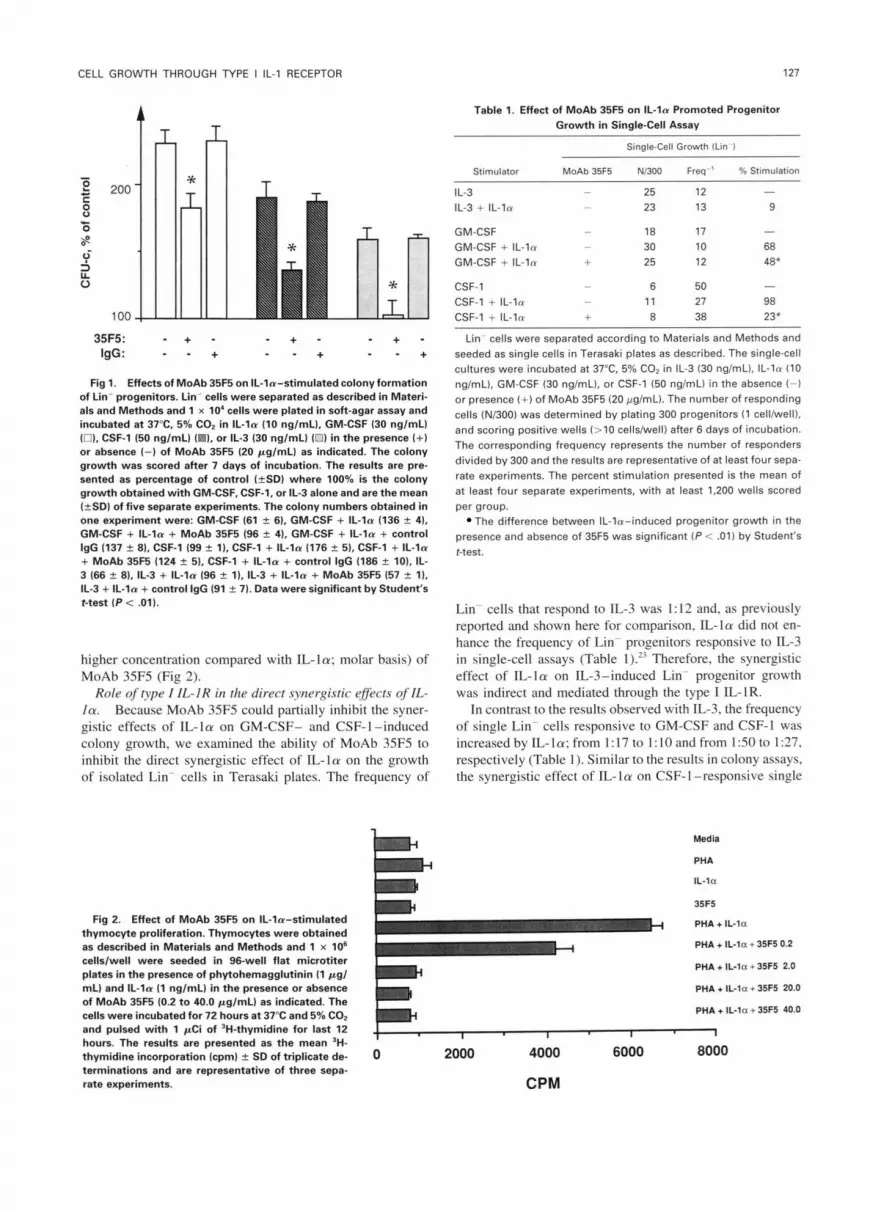
Role of type I IL-IR in the synergistic effects of IL-la on BMC growth in soft-agar colony assays. Although the synergistic effects of IL-1 on hematopoietic progenitor growth have been proposed to be indirect, possibly through the induction of HGF production from hematopoietic acces- sory cells, we have also shown that IL-la in combination with HGFs can directly promote the growth of purified pro- genitor cells in single-cell assay.23 In particular, the growth of Lin- progenitors responsive to GM-CSF was enhanced by IL-la in single-cell assays." However, using '251-IL-la we have been unable to detect and define the type of IL- 1 R(s) constitutively expressed on purified progenitors. Therefore, using MoAb 35F5 we examined whether the synergistic ef- fect of IL-La in colony assays was mediated through IL-1R I. First, Lin- BMCs were plated in soft-agar assays with IL- la (10 ng/mL) in combination with either IL-3 (30 ng/mL), GM-CSF (30 ng/mL), or CSF-1 (50 ng/mL). As previously shown, IL-la enhanced the CSF-induced colony formation of all three CSFs examined (Fig l). '4 The highest synergy was observed between IL-la and GM-CSF (140%), whereas IL-la enhanced the CSF-1- and IL-3-induced colony for- mation by 90% and 60%, respectively (Fig I). The ability of MoAb 35F5 (20 pg/mL) to block IL-la-enhanced CSF- induced colony growth varied between the CSFs examined (Fig l). Specifically, MoAb 35F5 inhibited 90% to 100% of the synergistic effect of IL- la on IL-3-induced colony formation, whereas the synergistic effect of IL-la on CSF- 1- and GM-CSF-induced progenitor growth was reduced 55% to 60% and 40% to 50%, respectively (Fig 1). Finally, 35F5-mediated inhibition of the synergistic effects of IL-la was dose dependent, with maximal inhibition observed at 20 ,ug/mL of antibody (data not shown).
To further control for the amount of antibody required, we examined the concentration of MoAb 35F5 needed to recognize the type I IL-1R and inhibit IL-la-induced T- cell proliferation. The proliferation of thymocytes (l X 10' cells/well) was determined by 'H-thymidine incorporation after 72 hours of treatment with phytohemagglutinin (1 pg/ mL) in combination with IL-la (1 ng/mL) (Fig 2). When compared with the effect of MoAb 35F5 on IL-la-induced progenitor cell growth, MoAb 35F5 inhibited the IL-la- induced T-cell proliferation in an dose-dependent way with maximal inhibition ( 1 00%) obtained at 2.0 pg/mL (200-fold
CELL GROVVTH THROUGH TYPE I IL-l RECEPTOR
t , T
127
Table l. Effect of MoAb 35F5 on IL-la Promoted Progenitor Growth in Single-Cell Assay
35F5: - + - IgG:
- + - + " + "
;o + -
- +
Fig 1. Effects of MoAb 35R on IL-la-stimulated colony formation of Lin- progenitors. Lin- cells were separated as described in Materi- als and Methods and 1 x 10' cells were plated in soft-agar assay and incubated at 37°C. 5% CO, in IL- la (10 nglmL), GM-CSF (30 nglmL) (01. CSF-1 (50 nglmL) I 1, or IL-3 (30 nglmLl (0) in the presence (+) or absence (-1 of MoAb 35F5 (20 pglmL) as indicated. The colony growth was scored after 7 days of incubation. The results are pre- sented as percentage of control (fSD) where 100% is the colony growth obtained with GM-CSF, CSF-1, or IL-3 alone and are the mean (+SD) of five separate experiments. The colony numbers obtained in one experiment were: GM-CSF (61 -L 6). GM-CSF + IL- la 1136 % 4). GM-CSF + IL- la + MoAb 35F5 196 f 41, GM-CSF + IL-la + control
+ MoAb 35F5 (124 2 5) . CSF-1 + IL- la + control lgG (186 f 101, IL-
IL-3 + IL-la + control IgG (91 f 7). Data were significant by Student's t-test ( P i .01).
IgG (137 f 81, CSF-1 (99 k l), CSF-1 + IL-la (176 f 5). CSF-1 + IL- la
3 (66 % 8), 11-3 + IL-la (96 2 l), IL-3 + IL- la + MoAb 35F5 (57 2 l),
higher concentration compared with IL-la; molar basis) of MoAb 35F5 (Fig 2).
Role of hpe I IL-IR in the direct synergistic efects of IL- la. Because MoAb 35F5 could partially inhibit the syner- gistic effects of IL-la on GM-CSF- and CSF-l-induced colony growth, we examined the ability of MoAb 35F5 to inhibit the direct synergistic effect of IL-la on the growth of isolated Lin- cells in Terasaki plates. The frequency of
Fig 2. Effect of MoAb 35- on IL-la-stimulated thymocyte proliferation. Thymocytes were obtained as described in Materials and Methods and l x l o6 cellslwell were seeded in 96-well flat microtiter plates in the presence of phytohemagglutinin (1 p g l mLI and IL-la (1 nglmLl in the presence or absence of MoAb 35F5 (0.2 to 40.0 pglmL) as indicated. The cells were incubated for 72 hours at 37°C and 5% COn and pulsed with 1 pCi of 'H-thymidine for last 12 hours. The results are presented as the mean 3H- thymidine incorporation (cpm) 2 SD of triplicate de- terminations and are representative of three sepa- rate experiments.
Single-Cell Growth (Lin )
Stimulator MoAb 35F5 NB00 Frea ' % Stimulation
IL-3 - 25 12 IL-3 + IL-la - 23 13 9
-
GM-CSF - 18 17 GM-CSF + IL-la - 30 10 68
-
GM-CSF + IL-la + 25 12 48*
CSF-1 - 6 50 CSF-1 + IL-la - 11 27 98 CSF-1 + IL-la + 8 38 23'
-
Lin- cells were separated according to Materials and Methods and seeded as single cells in Terasaki plates as described. The single-cell cultures were incubated at 37°C. 5% CO2 in IL-3 (30 nglmL), IL-la (10 ng/mL), GM-CSF (30 nglmL1, or CSF-1 (50 nglmL) in the absence ( - )
or presence (+ ) of MoAb 35F5 (20 pglmL). The number of responding cells (N/300) was determined by plating 300 progenitors (1 cell/well). and scoring positive wells (>l0 cells/well) after 6 days of incubation. The corresponding frequency represents the number of responders divided by 300 and the results are representative of at least four sepa- rate experiments. The percent stimulation presented is the mean of at least four separate experiments, with at least 1,200 wells scored per group.
The difference between IL-la-induced progenitor growth in the presence and absence of 35F5 was significant (P < .01) by Student's t-test.
Lin- cells that respond to IL-3 was I : 12 and. as previously reported and shown here for comparison, IL-la did not en- hance the frequency of Lin- progenitors responsive to IL-3 in single-cell assays (Table 1 ).'3 Therefore, the synergistic effect of IL-la on IL-3-induced Lin- progenitor growth was indirect and mediated through the type I IL-IR.
In contrast to the results observed with IL-3. the frequency of single Lin- cells responsive to GM-CSF and CSF-I was increased by IL- 1 a; from 1 : 17 to 1 : I O and from 1 :50 to 1 :27, respectively (Table 1). Similar to the results in colony assays, the synergistic effect of IL-la on CSF-I -responsive single
Medla
PHA
IL-la
35F5
PHA + IL-la
PHA + IL-la + 35F5 0.2
PHA + IL-la + 35F5 2.0
PHA + IL-la + 35F5 20.0
PHA + IL-la + 35F5 40.0
I I I
0 2000 4000 6000 8000

128 HESTDAL ET AL
Table 2. Effect of MoAb 35F5 on the Direct Synergistic Response of IL-la on Progenitor Proliferation
Single-Cell Growth
Thy-l' Thy-l
MoAb % % Stimulator 35F5 Frea ' Stimulation Frea" Stimulation
IL-3 - 6 IL-3 + IL-la - 6 10 12 7
- 12 -
GM-CSF - 75 - 10 GM-CSF + IL-la - 18 316 10 -8 GM-CSF + IL-la + 20 261 ND ND
-
CSF-1 - 300 - 50 CSF-1 + IL-la - 27 713 17 83
CSF-1 + IL-la + 42 638 43 13
-
Lin-Thy-l+ and Lin-Thy-1- cells were separated according to Mate- rials and Methods and seeded as single cells in Terasaki plates as described. The single-cell cultures were incubated at 37"C, 5% CO, in IC-3 130 ng/mL), IL-la (10 ng/rnL), GM-CSF (30 ng/rnL), or CSF-1 (50 ng/mL) in the absence (-) or presence ( + l of MoAb 35F5 (20 pg/ mL). The frequency of responding progenitors ( > l 0 cells/well) was determined after 6 days of incubation. The frequency of responding progenitors was obtained as described in Materials and Methods and Table 1. The results are representative of at least four separate experi- ments. The percent stimulation presented is the mean of at least four separate experiments, with at least 1,200 wells scored per group.
Abbreviation: ND, not determined.
progenitors was reduced 75% to 80% by MoAb 35F5 (Table l), and the synergistic effect on GM-CSF-induced single progenitor growth was only partially reduced (20%) by MoAb 35F5 (Table 1). Thus, the synergistic effect of IL- 1 a on CSF- 1 -induced growth was direct and mainly mediated through the type I IL-lR, and the synergistic effect of IL- l a on GM-CSF-responsive Lin- progenitors was direct and only partially mediated through the type I IL-1R.
Because the Lin- cells represent a partially purified popu- lation of progenitors, this might account for the differential effects of MoAb 35F5 on the synergistic effect of IL-la; therefore, we further separated the Lin- cells on the basis of Thy-l antigen. Lin- cells expressing the Thy- 1 antigen have previously been shown to be enriched for multipotential hematopoietic stem cells.'s The direct proliferative effect of IL-la (10 ng/mL) on CSF-l- (50 ng/mL), GM-CSF- (30 ng/mL), and IL-3-induced (30 ng/mL) growth was exam- ined on Lin-Thy-l+ (Thy-l') and Lin-Thy-l- (Thy-I-) cells in Terasaki plates. The frequency of progenitors responsive to IL-3 in single-cell assays was higher in Thy-l' cells com- pared with Thy-l- cells, 1:6 versus 1:12 (Table 2), and similar to the results with Lin- cells no direct synergy be- tween IL-la and IL-3 was observed in either population (Table 2).
The proliferation of isolated progenitors responsive to CSF-l was enhanced by IL-la; 85% to 95% for Thy-l- cells and approximately 700% for Thy-l+ (Table 2). Although the direct synergistic effect of IL-la on CSF-l-induced growth of Thy-1- cells was inhibited 85% to 100% by MoAb 35F5 (Table 2), the direct synergistic effect of IL-la on the prolif-
eration of CSF-I -induced by Thy-l + single cells was mar- ginally reduced (10% to 15%) in the presence of MoAb 35F5 (Table 2). GM-CSF-induced proliferation of isolated Thy- 1 ' cells was enhanced (300%) by IL-la, but IL-la did not affect GM-CSF-induced progenitor growth of single Thy- 1 progenitors (Table 2). Similar to the results obtained with CSF-I on the Thy-l + population, the frequency of sin- gle Thy-l + cells responsive to the combination of GM-CSF and IL-la was partially reduced (15% to 25%) by the MoAb against the type I IL-IR (Table 2). Thus, IL-la and CSF- 1 directly synergized through the type I IL-1R on Thy-l progenitors, whereas the direct synergistic response of IL-la on both CSF- 1 and GM-CSF-responsive Thy- 1 + progenitors was only marginally inhibited by MoAb 35F5.
Role of type I1 IL-lR in the growth of Lin- Thy-]+ cells. These results suggested that the direct synergistic effect of IL- la on Lin.-Thy- 1 + progenitors could be mediated through the type I1 IL-IR. Therefore, the effect of MoAb 4E2 on IL- la-induced progenitor growth was examined on Thy-l' cells in single-cell assays. The MoAb 4E2 has been shown to block the binding of radioiodinated IL-la to the type I1 IL-IR but not the type I IL-1R; however, the affinity of this MoAb for the IL- 1 R I1 is lower than the affinity of MoAb 35F5 for the type I IL-1R (R.C., unpublished observation, October 1992). Preliminary experiments showed that the binding of radioiodinated IL- l a on both unseparated BMCs and NFS-60 cells (murine myeloid progenitor cell line with high numbers of IL- 1 R) was completely inhibited by MoAb 4E2 (200-fold higher concentration compared with '2sI-IL-1 on molar basis) (data not shown). Because MoAb 4E2 has a reduced affinity for type I1 IL-lR, we also reduced the concentration of IL- 1 a in the single-cell assays; therefore, we used IL-la at 0.5 ng/mL and MoAbs 4E2 and 35F5 at 20 pg/mL (4,000-fold higher concentration of MoAb compared with IL-la; molar basis). As a result of using 0.5 ng/mL IL- l a , the direct synergistic effect of IL-la on CSF-l and GM- CSF-induced proliferation of Thy- 1 + cells was reduced 50% to 60% compared with the optimal dose of IL-la (10 ng/ mL). Similar to the results obtained with IL-la at 10 ng/ mL, the addition of the MoAb 35F5 reduced the frequency of Thy- 1 + single progenitor responding to GM-CSF plus IL- la (0.5 ng/mL) by approximately 20% (Fig 3). However, in some experiments we could detect up to 60% to 70% inhibi- tion. The synergistic effect of IL-la (0.5 ng/mL) on CSF- l-induced Thy-]' progenitor growth in Terasaki plates was inhibited 100% by MoAb 35F5 (20 pg/mL) (Fig 3). In com- parison, the MoAb 4E2 at 4,000-fold higher concentration than IL-la did not decrease the direct synergistic effect of IL-la on GM-CSF responsive Thy-l + progenitors or sig- nificantly reduce the frequency of single Thy-l' cells re- sponsive to IL-la (0.5 ng/mL) and CSF-l (Fig 3).
Taken together, the direct synergistic effect of IL-la on CSF- 1 -responsive progenitors, both Thy-l + and Thy- 1 -, OC- curred through the type I IL-I R and not the type 11 IL- 1R. GM-CSF and IL-la directly synergize to promote the proliferation of Thy- 1 but not Thy- 1 progenitors, partially through the type I IL-1R but not the type I1 1L- 1R.
To show that we could completely inhibit the direct syner- gistic response of 1L- 1 on GM-CSF-responsive progenitors

CELL GROWTH THROUGH TYPE I IL-l RECEPTOR
600
500 E
100
T T
35F5: - + - 4E2: - - +
- + - - - +
Fig 3. Effect of MoAb 35R and MoAb 4E2 on the direct synergistic response of IL-ln on Lin-Thy-l' progenitor growth. Lin-Thy-l' cells were separated according t o Materials and Methods and seeded as single cells in Terasaki plates as described. The single-cell cultures were incubated at 37°C. 5% CO2 in IL-ln (0.5 ng/mL), GM-CSF (30 ng/mL) (0). or CSF-1 (50 nglmLl (0) in the presence (+l or absence (-1 of MoAb 4E2 (20 pglmL) or MoAb 35F5 (20 pglmL) as indicated. The frequency of responding progenitors (>l0 cellslwell) was deter- mined after 6 days of incubation. The results are presented as per- centage of control (GM-CSF or CSF-l alone) and are the mean (?SDI of four separate experiments.
in our assay system, we examined the effect of MoAb 4CS on IL-la- and IL-lp-induced proliferation of Thy-l' pro- genitors in single assays. MoAb 4CS recognizes a 96-kD cell membrane protein expressed by all cells with IL- I Rs (R. Chizzonite, P. Nunes, T. Truitt: manuscript in preparation, October 1993). The cDNA for this protein has been recently cloned and shows homology with the type I IL-I R (approxi- mately 65%) but is distinct from both type I and type I1 IL- IRs. The 4CS antibody binds to recombinant 4CS protein expressed in COS cells. MoAb 4CS has been shown to spe- cifically inhibit the proliferative effect of IL-Ip on lympho- cytes but had no effect on the proliferative response of 1L-la (R.C., unpublished observation, October 1992). Al- though IL-la-induced proliferation was not affected by MoAb 4CS, the direct synergistic effect of IL-10 on GM- CSF-responsive progenitors was completely (100%) inhib- ited by MoAb 4CS (Table 3).
Finally, because MoAb 4E2 has been shown to have an agonistic effect on the growth of lymphocytes ( R C , unpub- lished observation, January 1991),.we examined the effect of MoAb 4E2 on the growth of isolated Thy-l' cells in the presence of CSF-I or GM-CSF. The MoAb 4E2 (25 pg/mL) did not enhance GM-CSF- or CSF-I -induced progenitor cell growth in single-cell assays (Table 4).
IL-la indirectly enhanced the IL-3-induced progenitor growth of Lin-Thy-I' cells. Although 1L-la did not di- rectly synergize with IL-3 to promote the growth of Lin-, Lin-Thy-l', or Lin-Thy-l- cells in Terasaki plates, it did
129
Table 3. Effect of MoAb 4C5 on 1L-la- and IL-lp-Induced Progenitor Growth
Stimulator Single-Cell Growth (N/3001
GM-CSF 5 GM-CSF + IL-la 10 GM-CSF + IL-la + MoAb 4C5 13
GM-CSF + IL-10 13 GM-CSF + IL-10 + MoAb 4C5 3
Lin Thy-l ' cells were separated according to Materials and Meth- ods and seeded as single cells in Terasaki plates as described. The single-cell cultures were incubated at 37°C. 5% CO2 in IL-la (0.5 ngl mL), IL-10 (0.5 ng/mL), GM-CSF (30 nglmL), or MoAb 4C5 (20 pg/mL) as indicated. The growth of single cells are presented as the number (N) of responding progenitors per 300 cells (1 celllwell) and after 6 days of incubation scoring positive wells (> l0 cellslwell). The results are one representative of two separate experiments.
synergize with IL-3 to promote the growth of Lin- cells in colony assays (Fig l ) . Therefore, we examined whether IL- la could also indirectly promote the growth of FACS-sepa- rated Thy-l " or Thy-l 'Lin- progenitors in colony assays. Lin-Thy- 1 and Lin-Thy- I ' cells were plated in soft-agar assays with IL-3 (30 ng/mL) alone or in the combination with IL-la ( I O ng/mL). Interestingly, although IL- l a ( I O ng/mL) did not enhance IL-3-induced colony growth of Thy- 1 cells, it did enhance the growth of IL-3-responsive Thy-l' cells (2.5 X 10' cells/plate) by 60% (Fig 4). The synergistic effect of IL-la on IL-3-responsive Thy-l + pro- genitors in colony assay was completely inhibited by MoAb 3SFS (20 mg/mL) (Fig 4).
IL-la induces GM-CSF binding on Lin- cells through Cpe I IL-IR. This laboratory has previously proposed that upregulation of CSF receptors may be one mechanism for the synergistic interaction between HGFs.'~ Since we have shown that IL-la enhanced GM-CSF-induced colony for- mation of Lin- cells and IL-la upmodulated the expression of GM-CSF receptors on these cells, we examined whether
Table 4. Agonistic Effect of MoAb 4E2 on Progenitor Growth
Stimulator Single-Cell Growth (Thy-l .) Freq
GM-CSF 33 GM-CSF + IL-la 15 GM-CSF + MoAb 4E2 37 GM-CSF + control IgG 37
CSF-1 100 CSF-1 + IL-la 21 CSF-1 + MoAb 4E2 100 CSF-1 + control IgG 100
Lin' Thy-l + cells were separated according to Materials and Meth- ods and seeded as single cells in Terasaki plates as described. The single-cell cultures were incubated at 37°C. 5% CO2 in IL-la (0.5 ngl mL), GM-CSF (30 nglmL), CSF-1 (50 nglmL), MoAb 4E2 (25 pglmL). or control IgG (25 pg/mL) as indicated. The frequency of responding progenitors (> l0 cells/well) was determined after 6 days of incuba- tion. The frequency of responding progenitors was obtained as de- scribed in Materials and Methods and Table l. The results are one representative of two separate experiments.

130
160 f
l! IL-3 IL-3 11-3
IL-a IL-la 35F5
(4 +I- 4)
IL-3 IL-3 IL-l a
Fig 4. The indirect synergistic effect of IL-la on 11-3 promoted Lin-Thy-l' (0) or Lin-Thy-1- ( I l l progenitor growth. Lin-Thy-l' and Lin-Thy-l- cells were separated according t o Materials and Methods and plated in soft-agar assay at 2,500 cellslplate or 10,000 cellslplate, respectively. The cultures were incubated at 37"C, 5% COz in IL- ln (10 nglmLl and 11-3 (30 nglmL) in the presence ( + l or absence ( - 1 of MoAb 35F5 (20 pglmL1 as indicated. The colony growth (>50 cells) was scored after 7 to 10 days of incubation. The results are presented as the mean 2 SD of duplicate determinations and are representative of four separate experiments. In the parentheses are shown the mean percent stimulation (+Sol from four separate experiments.
the upregulation of GM-CSF binding occurred through the type I IL-IR. Lin- cells were incubated with media, IL- la ( I O ng/mL), or IL-3 (30 ng/mL) with or without MoAb 3SFS (20 mg/mL). The specific binding of radio-iodinated GM- CSF on Lin- cells was examined after 24-hour incubation. As previously shown," IL-la as well as IL-3 enhanced the specific binding of "'I-GM-CSF on Lin- cells compared with control (2.3-fold and 3.S-fold, respectively) (Fig S). Although MoAb 3SFS did not reduce the IL-3-induced upregulation of "'I-GM-CSF-specific binding on Lin- cells, the enhanced binding observed after IL-la treatment was completely blocked by incubating the Lin- cells with MoAb 3SFS (Fig S). Thus, although MoAb 3SFS only partially inhibited the synergistic effect of IL-la on Lin- progenitor growth (Fig l ) , it blocked the upmodulation of GM-CSF binding on Lin- cells by 1L-la.
DISCUSSION
The biologic effects of IL- I are mediated through specific cell-surface receptors, and two distinct types have been shown: type I and I1 IL-IR.'4.2s.37.3X The two IL- 1 Rs represent different gene products and also show different immunoreac-
fibrohlasts, endothelial cells, keratinocytes, hepatocytes, chondrocytes, and synovial cells, whereas IL- I R I1 (p68) has heen described only on B cells, macrophages, and neutro- phils.2"'x.3X Although IL-la can directly synergize with other HGFs to promote hematopoietic progenitor growth, we have been unable to detect the expression of IL-IR I or IL-IR I1
tivity.?J.2S.W.J~l The IL-IR I (p80) has been found on T cells,
HESTDAL ET AL
using radioiodinated IL-la on purified progenitor cells.'" Because it was unknown whether type 1, type 11. or other IL-l -binding proteins mediated the proliferative signal of IL-la on purified progenitor cells. we used the difference in immunoreactivity between type I and type I1 IL-IR to better assess the role of the different IL- I Rs in hematopoietic progenitor growth. We present clear experimental evidence that IL- la directly and indirectly can stimulate the growth and differentiation of highly purified progenitors through the type I IL-IR but not the type I I IL-l R.
The synergistic effect. indirect or direct. observed with IL- la was dependent on the CSF used. In particular. although progenitor growth induced by GM-CSF or CSF-I was di- rectly stimulated by IL- I a. the growth of progenitors respon- sive to IL-3 was only indirectly enhanced through the type 1 IL-IR. Interestingly. the synergistic effect between IL-la and IL-3 was observed in colony assays using both Lin and Lin-Thy-l' cells at a low-plating density (2.500 Lin-Thy- 1 + cells/plate). Thus. the use of purified progenitors plated at low-cell density in colony assays does not exclude the possibility of indirect effects.
IL-l has been shown to induce the production of CSFs both in vitro and in vi~o '~ ' ' ' ; thus. the synergistic effect of IL-la on IL-.?-responsive progenitors could be indirectly mediated through HGF(s) production by Lin . However. we were unable to detect any synergistic effect of conditioned medium obtained from IL-la-stimulated Lin- cells (IL-l - conditioned medium) on IL-3-responsive progenitor growth (K.H. et al. unpublished observation, May 1993). Further- more, IL-l -conditioned medium did not stirnulate the prolif- eration of cell lines responsive to IL-3. GM-CSF. IL-6. G-
12
c1
x 5 ._
8
0 z m 0 k
a m 4
V W
U m
Y CI
0
35F5: - + + +
Fig 5. Effect of MoAb 35F5 on IL-le-induced GM-CSF binding t o Lin- progenitor cells. Lin- cells were incubated at 37°C for 24 hours in media K3, IL-la (10 nglmL) (0). or IL-3 (30 nglmL) ( presence (+) or absence (-1 of MoAb 35F5 (20 pglmL1 as indicated. Equilibrium binding with '%GM-CSF was performed as described in Materials and Methods. Data presented are the mean of duplicate determinations (+SDI and are representative of at least three sepa- rate experiments.

CELL GROWTH THROUGH TYPE I IL-l RECEPTOR 131
CSF, CSF-1, IL-4, and SCF (c-kit ligand) (K.H. and J.R.K., unpublished observation). However, low levels of HGF pro- duction, effects of combined HGFs, or membrane-bound forms of HGFs cannot be ruled out to explain the observed indirect effects.
We and others have shown that MoAb 35F5 inhibits the biologic effects of IL-la on cells expressing type I IL-1R both in vitro and in v ~ v o . ~ * ~ ~ ~ ~ ” Because IL-lR I is expressed on hematopoietic accessory cells such as T cells and stromal cells but not detectable on progenitor cells, the effects of IL- la on hematopoiesis were proposed to be indirectly medi- ated through type I IL-lR. However, in this study we show that the effect of IL-la on the proliferation of CSF-l -re- sponsive Thy-l+ and Thy-l- progenitors is direct and blocked by antibody to the type I receptor.
In contrast to the direct effect of IL-la on CSF-l-respon- sive progenitors, the direct synergistic effect of IL-la on GM-CSF-induced progenitor growth was only partially in- hibited by MoAb 35F5. However, in some experiments at lower doses of IL-la we could inhibit as much as 60% to 70% of the direct synergistic effects of IL-la and GM-CSF. In this regard, we have recently shown that GM-CSF can induce IL- IR expression on hematopoietic progenitor cells in therefore, upmodulation of IL-1Rs by GM-CSF may explain the incomplete inhibition of the direct syner- gistic effect of IL-la on GM-CSF-responsive progenitors observed by both MoAbs 35F5 and 4E2. Finally, IL-la- binding protein(s) other than type I and type I1 IL-lR, as previously de~cribed,2~~”~’~ may be important in the direct synergistic effect of IL-la in hematopoietic progenitor growth.
Despite the inability of anti-IL-1R 11 and anti-IL-RI to completely inhibit the direct synergistic effects of IL-la on GM-CSF-responsive Lin-Thy- l+ progenitors, we showed that MoAb 4C5 completely inhibited the synergistic response of IL-Io in combination with GM-CSF on these progenitors, whereas the synergistic effect of IL-la was not inhibited. This is comparable with the observation using MoAb 4C5 on IL- 1 -induced B-cell growth (R.C., unpublished observation, January 1992). This indicates that blocking IL-l binding and signal transduction inhibits the direct synergistic effects of IL- I on GM-CSF-responsive progenitors.
We also show that IL-la enhanced GM-CSF-induced colony formation of Lin- cells by 140% (Fig 1) and that this was reduced to 80% in the presence of MoAb 35F5 com- pared with GM-CSF alone (Fig 1). In comparison, IL-la enhanced GM-CSF-induced growth of isolated Lin- cells by 70% in single-cell assays that were partially inhibited by MoAb 35F5 (Table 1). This indicates that a portion of the synergistic response between IL- la and GM-CSF in colony assay is indirectly mediated through the type I IL-1R whereas the remainder of the synergistic response is direct and partially mediated through the type I IL-1R.
We have proposed that one mechanism for the synergistic effect of IL-la on GM-CSF-induced progenitor growth could be IL-la-induced upmodulation of GM-CSF recep- tors on Lin- progenitor cells.23 We show here that pretreat- ment of Lin- cells with MoAb 35F5 inhibits IL-la-induced GM-CSF receptor upmodulation but only partially reduces
IL-la-induced progenitor growth. Thus, the marginal inhi- bition of the direct synergistic effect of IL-la on GM-CSF- induced progenitor growth by MoAb 35F5, together with the observed inhibition of GM-CSF receptor upmodulation by anti-IL-1R I, indicates that IL-la may directly synergize with GM-CSF on progenitor cells by mechanism(s) other than receptor modulation.
IL-l is important in the regulation of hematopoietic growth and differentiation. In particular, it has proven to be essential in the stimulation of very immature progenitor cells, such as high proliferative potential colony-forming cells.”4 We have shown that IL-la can enhance progenitor growth both indirectly and directly through the type I IL-1R. Be- cause IL-1R I1 has been shown to be expressed on BMCs the role of this receptor in the regulation of hematopoietic development remains to be determined.
REFERENCES 1. Jubinsky PT, Stanley ER: Purification of hemopoietin 1 : A
multi-lineage hemopoietic growth factor. Proc Natl Acad Sci USA 82:2764, 1985
2. Stanley ER, Bartocci A, Patinkin D, Rosendaal M, Bradley TR: Regulation of very primitive multipotent, hemopoietic cells by hemopoietin-l. Cell 45:667, 1986
3. Mochizuki DY, Eisenman JR, Conlon PJ, Larsen AD, Tushin- ski RJ: Interleukin 1 regulates hematopoietic activity, a role pre- viously ascribed to hemopoietin 1. Proc Natl Acad Sci USA 845267, 1987
4. Bartelmez SH, Bradley TR, Bertoncello I, Mochizuki DY, Tus- hinski RI, Stanley ER, Hapel A J , Young IG, Kriegler AB, Hodgson GS: Interleukin 1 plus interleukin 3 plus colony-stimulation factor 1 are essential for clonal proliferation of primitive myeloid bone marrow cells. Exp Hematol 17:240, 1989
5. Hoang T, Haman A, Gonsalves 0, Letendre F, Mathieu M, Wong GC, Clark SC: Interleukin 1 enhances growth factor-depen- dent proliferation of the clonogenic cells in acute myeloblastic leuke- mia and of normal human primitive hemopoietic precursors. J Exp Med 168:463, 1988
6. Moore MAS, Warren DJ: Synergy of interleukin 1 and granulo- cyte colony-stimulating factor: In vivo stimulation of stem cell recov- ery and hematopoietic regeneration following 5-fluorouracil treat- ment of mice. Proc Natl Acad Sci USA 84:7134, 1987
7. Neta R, Sztein MB, Oppenheim JJ, Gillis S , Douches SD: The in vivo effect of interleukin 1 : Bone marrow cells are induced to cycle after administration of interleukin 1. J Immunol 139: 1861, 1987
8. Stork LC, Peterson VM, Rundus CH, Robinson WA: Interleu- kin-l enhances murine granulopoiesis in vivo. Exp Hematol 16:163, 1988
9. Benjamin WR, Tare NS, Hayes TJ, Becker JM, Anderson TD: Regulation of hemopoiesis in myelosuppressed mice by human re- combinant IL-la. J Immunol 142:792, 1989
10. Fibbe WE, van der Meer JWM, Falkenburg JHF, Hamilton MS, Kluin PM, Dinarello CA: A single low dose of human recombi- nant interleukin I accelerates the recovery of neutrophils in mice with cyclophosphamide-induced neutropenia. Exp Hematol 17:805, 1989
11. Johnson CJ, Douglas JK, Topper MI, Braunschweiger PG, Furmanski P In vivo hematopoietic effects of recombinant interleu- kin-la in mice: Stimulation of granulocytic, monocytic, megakaryo- cytic, and early erythroid progenitors, suppression of late-stage eryth- ropoiesis, and reversal of erythroid suppression with erythropoietin. Blood 73:678, 1989

132 HESTDAL ET AL
12. Moore MAS: Role of interleukin-l in hematopoiesis. Immu- no1 Res 8: 165, 1989
13. Bartelmez SH, Stanley ER: Synergism between hemopoietic growth factors (HGFs) detected by their effects on cells bearing receptors for lineage specific HGF: Assay for hemopoietin- 1. J Cell Physiol 122:370, 1985
14. Warren DJ, Moore MAS: Synergism among interleukin I , interleukin 3 and interleukin 5 in the production of eosinophils from primitive hemopoietic stem cells. J Immunol 140:94, 1988
15. Bradley TR, Hodgson GS: Detection of primitive macrophage progenitor cells in bone marrow. Blood 54:1446, 1979
16. Bradley TR, Hodgson GS, Bertoncello l: Characteristics of macrophage progenitor cells with high proliferative potential. Their relationships to cells with marrow repopulating ability in 5-FU treated mouse bone marrow, in Baum SJ, Ledney GD, Van Bekkum DW (eds): Experimental Hematology Today. New York, NY, Karger, 1979, p 285
17. Zucali JR, Dinarello CA, Oblon DJ, Gross MA, Anderson L, Weiner RS: Interleukin 1 stimulates fibroblasts to produce granulo- cyte-macrophage colony-stimulating activity and prostaglandin E2. J Clin Invest 77:1857, 1986
18. Bagby GC Jr, Dinarello CA, Wallance P, Wagner C, Hefe- neider S, McCall E: Interleukin 1 stimulates granulocyte macrophage colony-stimulating activity release by vascular endothelial cells. J Clin Invest 78: 13 16, 1986
19. Fibbe WE, Van Damme J, Billiau A, Duinkerken N, Lurvink E, Ralph P, Altrock BW, Kaushansky K, Willemze R, Falenburg JHF: Human fibroblasts produce granulocyte-CSF, macrophage CSF, and granulocyte-macrophage-CSF following stimulation by in- terleukin-l and poly(rI).poly(rC). Blood 72:860, 1988
20. Kaushansky K, Lin N, Adamson JW: Interleukin 1 stimulates fibroblasts to synthesize granulocyte-macrophage and granulocyte colony-stimulating factors. J Clin Invest 81:92, 1988
21. Fibbe WE, Van Damme J, Billiau A, Goselink HM, Voogt PJ, Van Eeden G, Ralph P, Altrock BW, Falkenburg JHF Interleukin-l induces human marrow stromal cells in long-term culture to produce granulocyte-colony-stimulating factor and macrophage-colony-stim- ulating factor. Blood 7 I :430, 1988
22. Vogel SN, Douches SD, Kaufman EN, Neta R: Induction of colony stimulating factor in vivo by recombinant interleuhn-l a and recombinant tumor necrosis factor a. J Immunol 138:2143, 1987
23. Jacobsen SEW, Ruscetti FW, Dubois CM, Wine J, Keller JR: Induction of colony-stimulating factor receptor expression on hematopoietic progenitor cells: Proposed mechanism for growth fac- tor synergism. Blood 80:678, 1992
24. Bomsztyk K, Sims JE, Stanton TH, Slack J, McMahan CJ, Valentine MA, Dower SK: Evidence for different interleukin 1 re- ceptors in murine B- and T-cell lines. Proc Natl Acad Sci USA 86:8034, 1989
25. Chizzonite R, Truitt T, Kilian PL, Stem AS, Nunes P, Parker KP, Kaffka KL, Chua AO, Lugg DK, Gubler U: Two high-affinity interleukin 1 receptors represent separate gene product. Proc Natl Acad Sci USA 86:8029, 1989
26. Dower SK, Kronheim SR, March CJ, Conlon PJ, Hopp TJ,
Gillis S, Urdal DL: Detection and characterization of high affinity plasma membrane receptors for human interleukin-l. J Exp Med 162:501, 1985
27. Uhl J , Newton RC, Giri JG, Sandlin G, Horuk R: Identifica- tion of IL-l receptors on human monocytes. J Immunol 142: 1576, 1988
28. Matsushima K, Akahosi T, Yamada M, Furutani Y, Oppen- heim JJ: Properties of a specific interleukin-l receptor on human Epstein-Barr virus-transformed B lymphocytes: Identity of the recep- tors for IL-la and [L-10. J Immunol 136:4496, 1986
29. Dubois CM, Ruscetti F W , Jacobsen SEW, Oppenheim JJ, Keller JR: Hematopoietic growth factors upregulate the p65 type l1 interleukin-l receptor on bone marrow progenitor cells in vitro. Blood 80:600, 1992
30. Dubois CM, Ruscetti F W , Keller JR, Oppenheim JJ, Hestdal K, Chizzonite R, Neta R: In vivo interleukin-l (IL-l) administration indirectly promotes type I1 IL-l receptor expression on hematopoi- etic bone marrow cells: Novel mechanism for the hematopoietic effects of IL-I. Blood 78:2841, 1991
3 1 . Benjamin WR, Chizzonite RA, Tare NS, Plocinski JM, Kaf- fka KL, McIntyre KW, Faherty DA, Kilian PL: Hematopoietic activ- ities of interleukin-l alpha: In vitro and in vivo modulation by an anti-IL-l receptor antibody, in Melli M, Parente L (eds): Cytokines and Lipocortins in Inflammation and Differentiation. New York, NY, Wiley-Lis, 1990, p 355
32. Neta R, Vogel SN, Plocinski JM, Tare NS, Benjamin W, Chizzonite R, Pilcher M: In vivo modulation with anti-interleukin- 1 (IL-I) receptor (p80) antibody 35F5 of the response to IL-I. The relationship of radioprotection, colony-stimulating factor, and IL-6. Blood 7657, 1990
33. Sims JE, Gayle MA, Slack JL, Alderson MR, Bird TA, Gin JG, Colotta F, Re F, Mantovani A, Shanebeck K, Grabstein KH, Dower SK: Interleukin 1 signaling occurs exclusively via the type I receptor. Proc Natl Acad Sci USA 906155, 1993
34. Hestdal K, Jacobsen SEW, Ruscetti FW, Dubois CM, Longo DL, Chizzonite R, Oppenheim JJ, Keller JR: In vivo effect of in- terleukin- la on hematopoiesis: Role of colony-stimulating factor receptor modulation. Blood 80: 1 , 1992
35. Spangrude GJ, Heimfeld S, Weissman IL: Purification and characterization of mouse hematopoietic stem cells. Science 241 58, 1988
36. Palaszynsh EW, lhle JN: Evidence for specific receptors for interleukin 3 on lymphokine-dependent cell lines established from long-term bone marrow cultures. J Immunol 132:1872, 1984
37. Dower SK, Ozato K, Segal DM: The interaction of mono- clonal antibodies with MHC class I antigens on mouse spleen cells 1. Analysis of the mechanism of binding. J Immunol 132:751, 1984
38. Dinarello CA: Interleukin-] and interleukin-l antagonism. Blood 77:1627, 1991
39. Kroggel R, Martin M, Pingoud V, Dayer J-M, Resch K: Two- chain structure of the interleukin-l receptor. FEBS Lett 22959, 1988
40. Savage N, Puren AJ, Orencole SF, Ikejima T, Clark BD, Dinarello CA: Studies on IL-l receptors on DlOS T-helper cells: Demonstration of two molecularly and antigenically distinct IL-l binding proteins. Cytolune 1:23, 1989