interactions among oscillatory pathways in nf-κb signalling
TRANSCRIPT

RESEARCH ARTICLE Open Access
Interactions among oscillatory pathways inNF-kappa B signalingYunjiao Wang1,4*, Pawel Paszek2, Caroline A Horton2, Douglas B Kell3, Michael RH White2, David S Broomhead4,Mark R Muldoon4*
Abstract
Background: Sustained stimulation with tumour necrosis factor alpha (TNF-alpha) induces substantial oscillations—observed at both the single cell and population levels—in the nuclear factor kappa B (NF-kappa B) system.Although the mechanism has not yet been elucidated fully, a core system has been identified consisting of anegative feedback loop involving NF-kappa B (RelA:p50 hetero-dimer) and its inhibitor I-kappa B-alpha. Manyauthors have suggested that this core oscillator should couple to other oscillatory pathways.
Results: First we analyse single-cell data from experiments in which the NF-kappa B system is forced by shorttrains of strong pulses of TNF-alpha. Power spectra of the ratio of nuclear-to-cytoplasmic concentration ofNF-kappa B suggest that the cells’ responses are entrained by the pulsing frequency. Using a recent model of theNF-kappa B system due to Caroline Horton, we carried out extensive numerical simulations to analyze the responsefrequencies induced by trains of pulses of TNF-alpha stimulation having a wide range of frequencies andamplitudes. These studies suggest that for sufficiently weak stimulation, various nonlinear resonances should beobservable. To explore further the possibility of probing alternative feedback mechanisms, we also coupled themodel to sinusoidal signals with a wide range of strengths and frequencies. Our results show that, at least insimulation, frequencies other than those of the forcing and the main NF-kappa B oscillator can be excited viasub- and superharmonic resonance, producing quasiperiodic and even chaotic dynamics.
Conclusions: Our numerical results suggest that the entrainment phenomena observed in pulse-stimulatedexperiments is a consequence of the high intensity of the stimulation. Computational studies based on currentmodels suggest that resonant interactions between periodic pulsatile forcing and the system’s natural frequenciesmay become evident for sufficiently weak stimulation. Further simulations suggest that the nonlinearities of theNF-kappa B feedback oscillator mean that even sinusoidally modulated forcing can induce a rich variety ofnonlinear interactions.
BackgroundNuclear factor kappa B (NF-�B) transcription factors arecritical to the control of response to cellular stress andare also involved in the regulation of cell-cycle/growth,survival, apoptosis, inflammation and immunity [1-5].They are dimeric molecules, composed of either homo-or hetero-dimers, with the most common form beingthe RelA:p50 hetero-dimer.In resting cells, NF-�B dimers are sequestered in the
cytoplasm by members of a family of molecules, theinhibitors of �B or I�Bs. When the cells are stimulated
with tumour necrosis factor alpha (TNFa) certainkinases, the IKKs, are activated and phosphorylate I�Bs,marking them for degradation. The degraded I�Bs thenrelease NF-�B dimers which are free to translocate intothe nucleus, where they bind to specific sequences inthe promoter or enhancer regions of target genes,including those for I�Ba and the zinc finger proteinA20 [6,7]. Newly synthesized I�Ba migrates to thenucleus, binds to NF-�B dimers and removes themfrom the nucleus, while A20 protein stays in cytoplasmand represses the activity of TNFa receptors [8]. Hencethe NF-�B system includes at least two negative feed-back loops, one involving cytoplasmic sequestrationmediated by I�Ba and another involving A20.* Correspondence: [email protected]; [email protected]
Full list of author information is available at the end of the article
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
© 2011 Wang et al; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative CommonsAttribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction inany medium, provided the original work is properly cited.

Coupled negative feedback loops are known to havethe potential to support oscillations [9,10]. Biologicalevidence suggesting NF-�B oscillations was reported byHoffmann et al. [11], who did population-level studiesof I�Ba-/-embryonic fibroblasts using electro-mobilityshift assays, and also observed by Nelson et al. [12] inthe cellular concentration and nuclear: cytoplasmic loca-lisation of I�Ba and RelA-fluorescent fusion proteins insingle SK-N-AS cells. Single cell time-lapse imaging datashowed persistent cycling of NF-�B localisation betweenthe cytoplasm and nucleus of SK-N-AS cells in responseto continual TNFa stimulation [12,13].The expression of genes regulated by NF-�B is tightly
coordinated with the activities of many other signallingand transcription-factor pathways [14-18] including thep53 signalling pathway. Though the canonical NF-�Bsignalling pathway has been studied extensively, theexistence and mechanisms of the interactions betweenthe NF-�B pathway and other signalling pathways areunclear.Ashall et al. [19] reported that different patterns of
pulsatile stimulation of the NF-�B system lead to differ-ent patterns of NF-�B dependent gene expression, sup-porting the view that the frequency of the oscillationsmay have a functional role. It is therefore important tounderstand how the system’s response frequencies areinfluenced by interactions with other oscillatory path-ways. If both the NF-�B network and its couplings toother other oscillatory pathways were purely linear, thenit would be straightforward to use the machinery oftransfer functions to characterise their interactions. Inparticular, one would expect the power spectrum of aperiodically stimulated system to have its power concen-trated at the forcing signal’s frequency and its harmo-nics. But the NF-�B network is a highly nonlinearsystem of coupled chemical reactions and hence, as wewill demonstrate in modelling studies below, itsresponse to periodic stimulation can include complexinteractions between the intrinsic negative feedbackoscillator and the stimulus. In what follows we workwith a deterministic model first described in [19] andexamine the power spectral densities of the time coursesof NF-�B localisation when the model is subjected totwo types of periodic signals: trains of rectangular pulsesand sinusoidal signals. Both sorts of stimulus areintended as a proxies for the influence of other oscilla-tory pathways on the core NF-�B feedback loop and wefind that such interactions can produce a rich variety ofnonlinear dynamical behaviour. Our results on sinusoi-dal forcing are in broad agreement with those of Fonsletet al. [20], who applied a related family of sinusoidal sti-mulation protocols to a reduced model of the core NF-�B oscillator originally developed in [21].
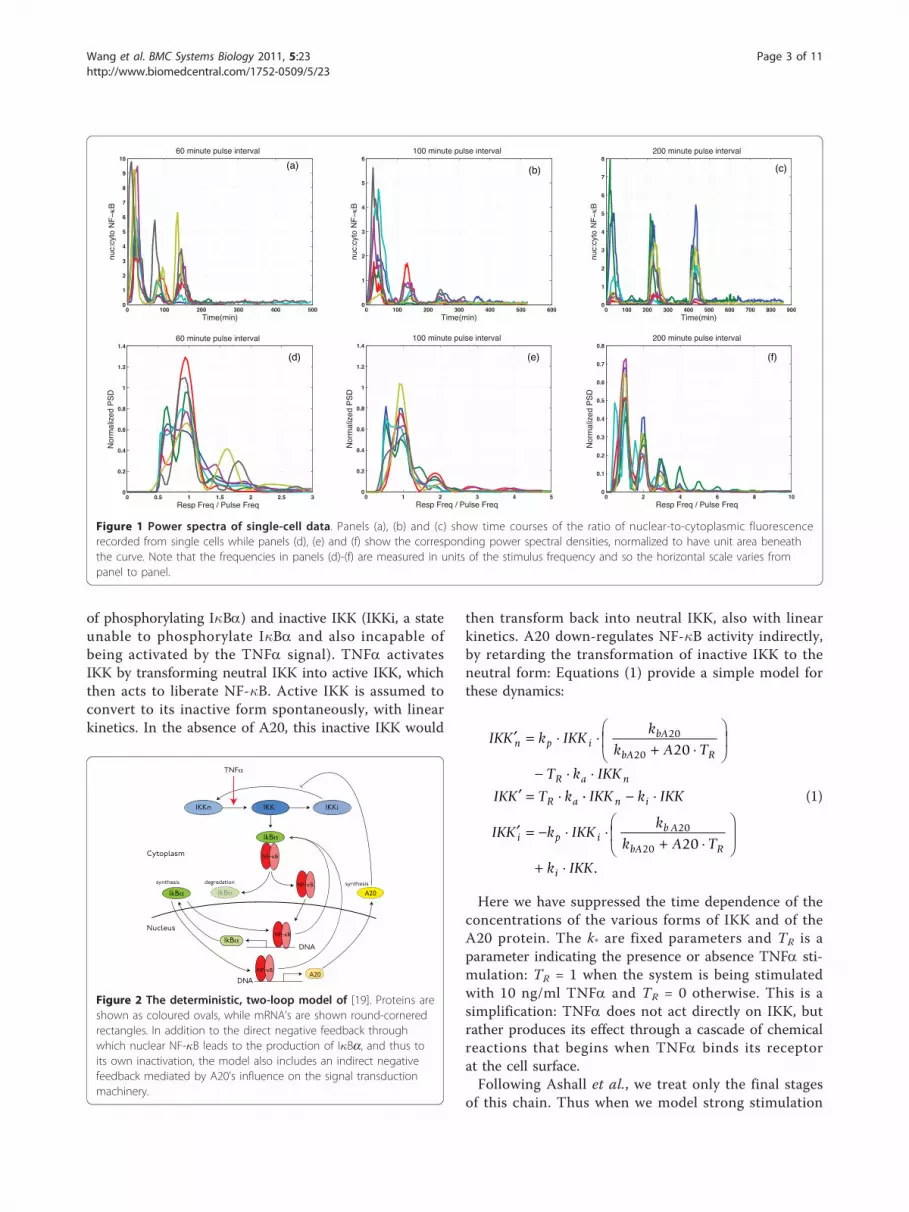
Results and DiscussionExperimental phenomena and time series analysisWe begin by analyzing the time courses of the localisa-tion of fluorescently tagged NF-�B in single cells sub-jected to TNFa stimulation (see [22] for details). Thepanels in the top row of Figure 1 show the ratio ofnuclear-to-cytoplasmic fluorescence for three patterns ofstimulation. In all cases the cells received three strong(10 ng/ml) pulses of TNFa, each of five minute dura-tion. These pulses were separated by various inter-pulseintervals—55 minutes for panel (a),95 minutes for panel(b) and 195 minutes for panel (c)—resulting in net sti-mulus periods of 60, 100 and 200 minutes. The first ofthese is rather shorter than the observed period of theoscillations induced by constant TNFa stimulation,which is close to 100 minutes.The bottom row of Figure 1 shows the corresponding
power spectral densities and makes the point that theintrinsic oscillation—the one induced by constant stimu-lation—does not appear to have been excited by thesepulsed stimulation protocols: in all cases the peaks inthe power spectral density appear at multiples of the sti-mulus frequency. In mathematical terms, the cellsrespond as though they have been perturbed away froma stable resting state and do not offer evidence of anymore complex internal dynamics. Nonetheless, thesedata provided crucial input to the development of themodels in [19]. In particular, note that the amplitude ofthe response to the second and third pulses of stimula-tion varies as a function of the inter-pulse period, beingsomewhat reduced in panels (a) and (b), but not in (c):this constrains the rate at which the model should relaxtoward equilibrium.One possibility is that the pulsing signal is too strong
and suppresses the response at the natural frequency.In the following section we report in numerical ex-periments, based on the deterministic model in [19],through which we investigate this possibility by simulat-ing the system’s response to a periodic train of squarepulses with strength varying from 0 to 10 ng/ml andinter-pulse intervals ranging from 20 to 295 minutes.
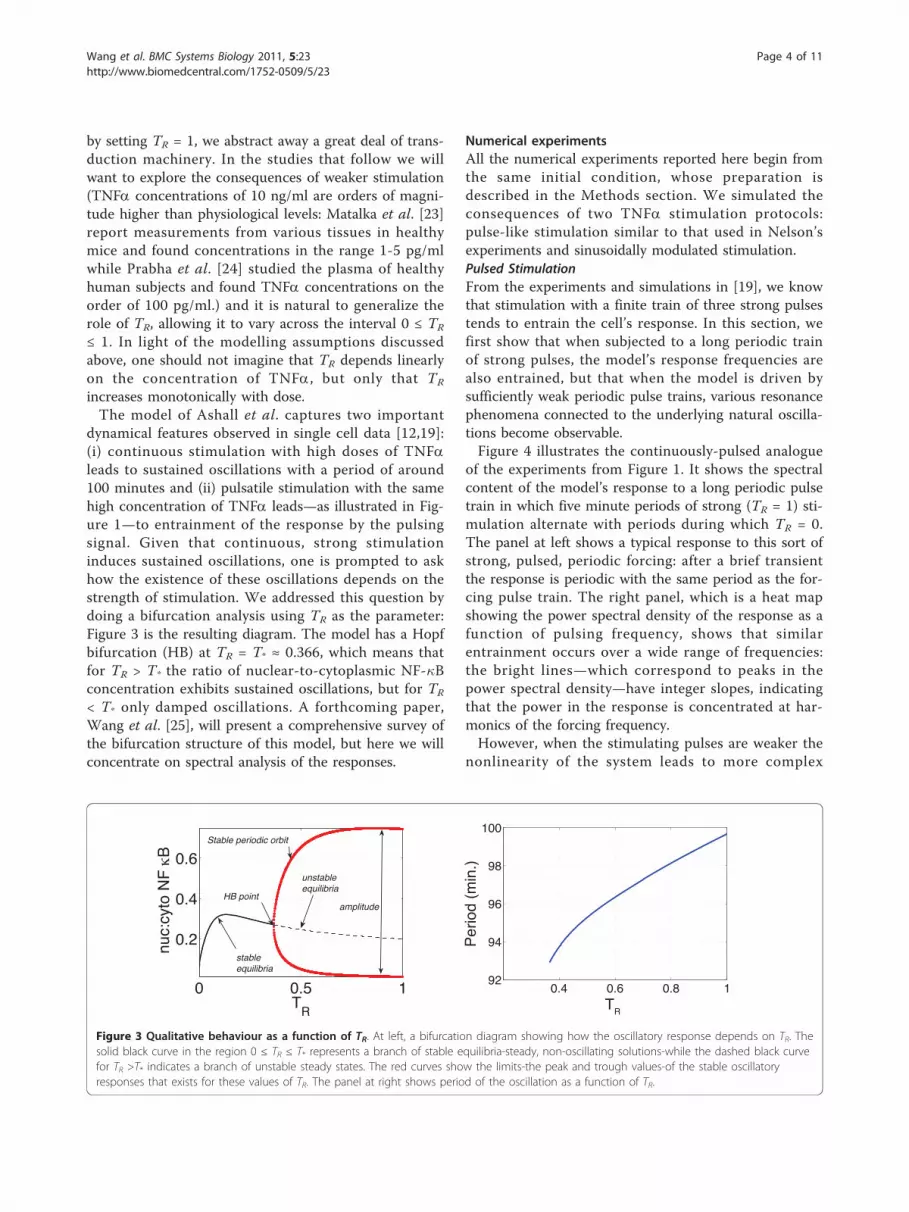
Model Introduction and Bifurcation AnalysisIn this section, we introduce a deterministic mathemati-cal model developed in [19]. It includes two couplednegative feedback loops—one for NF-�B-I�Ba interac-tions and another that models the dynamics of A20—asis illustrated in Figure 2. Here A20 acts by modifyingthe activity of I�B kinase (IKK), which transduces theTNFa signal by phosphorylating I�Ba, thus initiatingthe production of free NF-�B. IKK is assumed to existin one of three states: neutral IKK (denoted IKKn, astate ready for activation), active IKK (the state capable
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 2 of 11
of phosphorylating I�Ba) and inactive IKK (IKKi, a stateunable to phosphorylate I�Ba and also incapable ofbeing activated by the TNFa signal). TNFa activatesIKK by transforming neutral IKK into active IKK, whichthen acts to liberate NF-�B. Active IKK is assumed toconvert to its inactive form spontaneously, with linearkinetics. In the absence of A20, this inactive IKK would
then transform back into neutral IKK, also with linearkinetics. A20 down-regulates NF-�B activity indirectly,by retarding the transformation of inactive IKK to theneutral form: Equations (1) provide a simple model forthese dynamics:
IKK k IKKk
k A T
T k IKK
IKK T k
n p ibA
bA R
R a n
R a
20
20 20
IKK k IKK
IKK k IKKk
k A T
k IKK
n i
i p ib A
bA R
i
20
20 20
..
(1)
Here we have suppressed the time dependence of theconcentrations of the various forms of IKK and of theA20 protein. The k* are fixed parameters and TR is aparameter indicating the presence or absence TNFa sti-mulation: TR = 1 when the system is being stimulatedwith 10 ng/ml TNFa and TR = 0 otherwise. This is asimplification: TNFa does not act directly on IKK, butrather produces its effect through a cascade of chemicalreactions that begins when TNFa binds its receptorat the cell surface.Following Ashall et al., we treat only the final stages
of this chain. Thus when we model strong stimulation
0 100 200 300 400 5000
1
2
3
4
5
6
7
8
9
10
Time(min)
nuc:
cyto
NF
−κB
60 minute pulse interval
0 100 200 300 400 500 6000
1
2
3
4
5
6
Time(min)
nuc:
cyto
NF
−κB
100 minute pulse interval
0 100 200 300 400 500 600 700 800 9000
1
2
3
4
5
6
7
8
Time(min)
nuc:
cyto
NF
−κB
200 minute pulse interval
0 0.5 1 1.5 2 2.5 30
0.2
0.4
0.6
0.8
1
1.2
1.4
Resp Freq / Pulse Freq
Nor
mal
ized
PS
D
60 minute pulse interval
0 1 2 3 4 50
0.2
0.4
0.6
0.8
1
1.2
1.4
Resp Freq / Pulse Freq
Nor
mal
ized
PS
D100 minute pulse interval
0 2 4 6 8 100
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
Resp Freq / Pulse Freq
Nor
mal
ized
PS
D
200 minute pulse interval
(a) (b) (c)
(d) (e) (f)
Figure 1 Power spectra of single-cell data. Panels (a), (b) and (c) show time courses of the ratio of nuclear-to-cytoplasmic fluorescencerecorded from single cells while panels (d), (e) and (f) show the corresponding power spectral densities, normalized to have unit area beneaththe curve. Note that the frequencies in panels (d)-(f) are measured in units of the stimulus frequency and so the horizontal scale varies frompanel to panel.
Nucleus
synthesis synthesis
NF−κB
NF−κB
NF−κB
IkBα
IkBα IkBα
Cytoplasm
IKKn IKK IKKi
NF−κBA20
DNA
DNA
TNFα
IkBα
A20
Figure 2 The deterministic, two-loop model of [19]. Proteins areshown as coloured ovals, while mRNA’s are shown round-corneredrectangles. In addition to the direct negative feedback throughwhich nuclear NF-�B leads to the production of I�Ba, and thus toits own inactivation, the model also includes an indirect negativefeedback mediated by A20’s influence on the signal transductionmachinery.
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 3 of 11
by setting TR = 1, we abstract away a great deal of trans-duction machinery. In the studies that follow we willwant to explore the consequences of weaker stimulation(TNFa concentrations of 10 ng/ml are orders of magni-tude higher than physiological levels: Matalka et al. [23]report measurements from various tissues in healthymice and found concentrations in the range 1-5 pg/mlwhile Prabha et al. [24] studied the plasma of healthyhuman subjects and found TNFa concentrations on theorder of 100 pg/ml.) and it is natural to generalize therole of TR, allowing it to vary across the interval 0 ≤ TR
≤ 1. In light of the modelling assumptions discussedabove, one should not imagine that TR depends linearlyon the concentration of TNFa, but only that TR
increases monotonically with dose.The model of Ashall et al. captures two important
dynamical features observed in single cell data [12,19]:(i) continuous stimulation with high doses of TNFaleads to sustained oscillations with a period of around100 minutes and (ii) pulsatile stimulation with the samehigh concentration of TNFa leads—as illustrated in Fig-ure 1—to entrainment of the response by the pulsingsignal. Given that continuous, strong stimulationinduces sustained oscillations, one is prompted to askhow the existence of these oscillations depends on thestrength of stimulation. We addressed this question bydoing a bifurcation analysis using TR as the parameter:Figure 3 is the resulting diagram. The model has a Hopfbifurcation (HB) at TR = T* ≈ 0.366, which means thatfor TR > T* the ratio of nuclear-to-cytoplasmic NF-�Bconcentration exhibits sustained oscillations, but for TR
< T* only damped oscillations. A forthcoming paper,Wang et al. [25], will present a comprehensive survey ofthe bifurcation structure of this model, but here we willconcentrate on spectral analysis of the responses.
Numerical experimentsAll the numerical experiments reported here begin fromthe same initial condition, whose preparation isdescribed in the Methods section. We simulated theconsequences of two TNFa stimulation protocols:pulse-like stimulation similar to that used in Nelson’sexperiments and sinusoidally modulated stimulation.Pulsed StimulationFrom the experiments and simulations in [19], we knowthat stimulation with a finite train of three strong pulsestends to entrain the cell’s response. In this section, wefirst show that when subjected to a long periodic trainof strong pulses, the model’s response frequencies arealso entrained, but that when the model is driven bysufficiently weak periodic pulse trains, various resonancephenomena connected to the underlying natural oscilla-tions become observable.Figure 4 illustrates the continuously-pulsed analogue
of the experiments from Figure 1. It shows the spectralcontent of the model’s response to a long periodic pulsetrain in which five minute periods of strong (TR = 1) sti-mulation alternate with periods during which TR = 0.The panel at left shows a typical response to this sort ofstrong, pulsed, periodic forcing: after a brief transientthe response is periodic with the same period as the for-cing pulse train. The right panel, which is a heat mapshowing the power spectral density of the response as afunction of pulsing frequency, shows that similarentrainment occurs over a wide range of frequencies:the bright lines—which correspond to peaks in thepower spectral density—have integer slopes, indicatingthat the power in the response is concentrated at har-monics of the forcing frequency.However, when the stimulating pulses are weaker the
nonlinearity of the system leads to more complex
0 0.5 1
0.2
0.4
0.6
TR
nuc:
cyto
NF
κB
HB point
stableequilibria
unstableequilibria
Stable periodic orbit
amplitude
0.4 0.6 0.8 192
94
96
98
100
TR
Per
iod
(min
.)
Figure 3 Qualitative behaviour as a function of TR. At left, a bifurcation diagram showing how the oscillatory response depends on TR. Thesolid black curve in the region 0 ≤ TR ≤ T* represents a branch of stable equilibria-steady, non-oscillating solutions-while the dashed black curvefor TR >T* indicates a branch of unstable steady states. The red curves show the limits-the peak and trough values-of the stable oscillatoryresponses that exists for these values of TR. The panel at right shows period of the oscillation as a function of TR.
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 4 of 11

patterns of resonance. In particular, both subharmonicresonance—when periodic forcing excites a response ata rationally-related lower frequency—and superharmonicresonance—in which a pure sinusoid excites responsescontaining higher harmonics—are possible. Note thatthese terms are defined with reference to the forcing fre-quency, a convention used in, for example, [26]. Super-harmonic resonance is harder to identify when, as withthe rectangular pulses used here, the periodic forcingalready has power at higher harmonics, but subharmo-nic resonance occurs when 0.01 < TR < 0.2, as is evidentin the power spectral densities summarized in Figure 5.In addition to the bright lines with integer slopes, lines
with slopes 12
and 32
also appear. The first of these
provides evidence that subharmonic resonance occursover a wide range of pulsing frequencies.When TR ≥ 0.2 the power spectral density of the
response shows power only at integer multiples of thepulsing frequency, indicating that the response is fullyentrained by the forcing. Given that the pulse-strengthused in the experiments corresponds to TR = 1, thesemodelling results are in qualitative agreement with theexperimental data in Figure 1: the stimulation was sostrong that we should have expected it to have entrainedthe responses completely.Sinusoidally modulated stimulationIn this section we study the response of Horton’s modelto sinusoidally modulated stimulation of the form
T t tR( ) ( sin( )). 1 2 (2)
Here 0 ≤ ε ≤ 1/(1 + h) is the time average of the sti-mulus strength while 0 ≤ h ≤ 1 and ν > 0 are, respec-tively, the relative amplitude and the frequency of thesinusoidal modulation. With this parameterization TR (t)has period τ = 1/ν and range
0 1 1 1 ( ) ( ) ( ) .T tR
The roles of the parameters are illustrated in Figure 6.The case with h = 1 is the straightforward substitution
of pulse trains with sinusoidal waves having peak-to-trough amplitude 2ε. Our motivations for this formula-tion are twofold: firstly, provided h < 1 we have TR(t) >0 at all times and so expect the system more readily to
0 200 400 600 800 1000 1200 1400 1600 18000
0.5
1
1.5
2
2.5
3
3.5
4
Time(min)
nuc:
cyto
NF
−κB
Time interval 60 minutes
Figure 4 Periodic stimulation with strong pulses. The panel at left shows the model’s response to sustained periodic stimulation in whichstrong (TR = 1) pulses of 5 minute duration alternate with stimulus-free intervals of 55 minute duration. The panel at right shows the powerspectral density (PSD) of the response as a function of pulsing frequency: a vertical strip cut from this panel provides a colour-coded version ofthe PSD of the models’s response to pulsed forcing at the corresponding frequency.
Figure 5 Response to periodic trains of weak pulses. A heatmap summarising the power spectral density of NF-�B localisationwhen the model is subjected to pulsed forcing with TR = 0.1. Forforcing frequencies in the range 1.5-3.0 × 10-4. Hz. the lowest-lyingline segment, which has slope ≈ 1/2, provides evidence ofsubharmonic resonance.
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 5 of 11

exhibit oscillations similar to those induced by constantstimulation. Secondly, periodic forcing of the form (2)converts our model into a sinusoidally forced nonlinearoscillator, a class of systems that has been studied veryextensively (see, for example, Pikovsky et al. [27], Wig-gins [26] or Nayfeh and Mook [28]).When h = 0 the forcing (2) reduces to a constant stimu-
lus with TR (t) = ε and the bifurcation analysis illustratedin Figure 3 leads us to expect stable oscillations when ε >0.366 and a stable steady-state otherwise. But for h > 0 thesystem’s response depends delicately on the relationshipbetween the forcing frequency ν and the natural frequencyν0. When the forcing frequency ν and natural frequency ν0are close (the requisite degree of closeness depends on h)the response will become mode locked, or synchronizedwith the forcing: it will then have the same frequency ν asthe forcing and its power spectral density will be concen-trated around the harmonics of ν. When the difference (ν-ν0) is larger—when it lies just outside of a critical intervalaround zero whose size depends on h —the responsebecomes quasiperiodic and its power spectrum has fea-tures at frequencies f of the form
f p q 0 (3)
where p and q are integers.If pν and qν0 are close, the system’s nonlinearity will
permit what is called synchronization of order p/q: therewill be periodic responses in which the intrinsic oscilla-tor goes through p cycles for every q periods of the for-cing, so that the response has frequency
f p q 0 (4)
This idea allows us to give precise definitions for theterms sub-and superharmonic resonance used above:the former corresponds to resonances where p = 1 andq > 1 in (4), while the latter corresponds to q = 1 and
p > 1. Finally, when both h and the difference (ν -ν0)are large, the system’s response can become chaotic, sothat the features in its power spectral density bear nosimple relationship to the frequencies ν and ν0.Figure 7 which is a heat map of the power spectral
density of the response generated by relatively strongsinusoidal forcing of the from (2), illustrates many ofthe behaviours described above. Consider first the verti-cal strip with ν/ν0 ≈ 1. In this region the sinusoidalmodulation entrains the NF-�B system essentially com-pletely and so the heat map resembles the correspond-ing region in the left panel Figure 4: the power in theresponse is concentrated along lines corresponding toharmonics of the forcing frequency. Away from theregion ν/ν0 ≈ 1 the power spectrum of the response isconsiderably more complex: the strong horizontal bandsin Figure 7 which occur at integer multiples of ν0, showthat there is substantial power at the NF-�B system’snatural frequency and its harmonics. Additionally, thenonlinearity of the system means that the response haspower at frequencies given by
f q 0 ,
which gives rise to the network of lines with slope of±1. Finally, Figure 7 also exhibits sub-harmonic reso-nance: vertical strips for which ν ≈ qν0 (with q a wholenumber) show power concentrated at the forcing fre-quency, but also at frequencies f = ν/p, where p is awhole number. This is perhaps clearest in the strip ν ≈2ν0, where the strongest spectral feature lies along theline f = ν/2 ≈ ν0.
0.0 0.5 1.0 1.5 2.0 2.5 3.0
0.2
0.4
0.8
0.6
1.0
TR(t)
t
ε
ε(1 + η)
ε
εη
ε(1 - η)
εη
Figure 6 Sinusoidal forcing function. The function TR (t) for theparameter values ν = 1, h = 0.5 and ε = 0.6.
Figure 7 Nonlinear resonances to sinusoidal forcing. A heatmap showing the power spectral density of the response ofHorton’s model to forcing of the form (2) with ε = h = 0.5 andfrequencies in the range 0 ≤ ν ≤ 4.5 × ν0.
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 6 of 11

The two panels of Figure 8 are analogues of Figure 7but with especially strong (h = 1, left panel) or weak (h =0.1, right panel) modulation. The qualitative features aremuch the same, though it is interesting to note that inthe limit of very strong modulation—when h = 1 and soTR (t) vanishes once per forcing period—the system isvery strongly entrained by the forcing and does not showmuch power at its natural frequency ν0 or its harmonicsuntil the forcing frequency ν > 1.5 ν0. By contrast, whenthe modulation is weak evidence of modal interaction is
also weak, with very narrow mode-locking regions nearforcing frequencies of the form ν ≈ mν0.As Figures 7 and 8 illustrate, the susceptibility of
our model NF-�B system to resonance with sinusoidalmodulations depends strongly on the amplitude of themodulation: Figure 9 provides a quantitative surveyof this phenomenon. The shaded regions are examplesof Arnol’d tongues: their precise shapes can be calcu-lated with the methods outlined in the Appendix.When the modulation frequency and amplitude (ν, h)lie inside these tongues, the response of the systemwill be periodic, with a period τ that is an integermultiple of the modulation period 1/ν and thatalso lies close to an integer multiple of the naturalPeriod 1/ν0:
q p
0(5)
where p and q are whole numbers. Generally speakingthese periodic responses—which are a nonlinear gener-alization of the familiar phenomenon of resonance inlinear systems—are easiest to excite when the numbersp and q are small: the tongues in Figure 9 are labelledby ratios q:p where p and q are as in (5). Although Fig-ure 9 illustrates this story for the specific family of mod-ulations with ε = 0.5, the qualitative picture is essentiallythe same for all values in the range of 0.366 < ε < 0.5:in all cases there will be narrow tongues of parametercombinations (ν, h) for which the response is periodicwith a period given by a resonance relation like (5). Formodulations whose parameters lie outside the tonguesthe qualitative behaviour of the response will be more
Figure 8 Weak and strong modulations. Heat maps showing the power spectral density of the responses to forcing of the form (2) with ε =0.5 and h = 1 (left) or h = 0.1 (right).
0 1.0 2.0 3.0 4.00
0.2
0.4
0.6
0.8
1
η 1:2
1:1
3:22:1
3:1 4:1
ν/ν0
Figure 9 Arnol’d tongues. The qualitative behaviour of Horton’sNF- �B system when subjected to forcing of the from 2 with ε =0.5: when the modulation frequency ν and amplitude h lie insidethe shaded regions the response is periodic with a period that isrelated, by a relation like (5), to both the modulation’s period andthe system’s natural period.
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 7 of 11

complex, having a power spectrum qualitatively similarto those in Figure 7, with features at harmonics of themodulation frequency, at harmonics of the natural fre-quency ν0 (which varies with ε) and at near-resonantcombinations (3) of the two.When TR < 0.366, the unforced system is a damped
oscillator and so, when subjected to periodic forcing, mayexhibit resonant phenomena. By using the same numeri-cal simulation and power spectral density analyses as wedid for the case ε = 0.5, we find that the responses of thepathway to forcing with with TR < 0.366 can be dividedinto two groups: those in which harmonic, sub-andsuperharmonic resonances can be observed (in the region0 < ε ≤ 0.01), and those for which only harmonic andsubharmonic resonance can be observed.We have focussed here on periodic and quasiperiodic
oscillatory interactions amenable to power-spectral ana-lyses of the sort illustrated in Figures 4, 5, 6, 7, 8 and,for single-cell recordings, in the lower panels of Figure1. But there is every reason to expect a much richerrange of dynamical behaviour: Fonslet et al. [20], whoapplied forcing of the form (2) with h = 1 to a simpli-fied NF-�B model, saw evidence of a period-doublingcascade as well as chaos and strange attractors. Com-plete analysis of these more complex dynamical regimesrequires tools beyond the power spectrum, and so wedefer their exploration to a future paper.
ConclusionsGiven that intrinsically nonlinear chemical kineticsunderpin cell-signalling networks, one shouldn’t expectthese systems to be linear and any analysis of experimen-tal time series, whether in the time or frequency domain,must take this intrinsic nonlinearity into account. Ourmodelling studies suggest that coupling even the simplest,sinusoidal signal into the NF-�B network can give rise toa host nonlinear phenomena, including harmonic, sub-harmonic and superharmonic resonances as well as qua-siperiodic and even chaotic behaviour.The simulation studies reported here used an exter-
nally-imposed oscillatory forcing as a proxy for interac-tions between the core NF-�B feedback loop and otheroscillatory networks. Our results suggest that interac-tions between the NF-�B oscillator and other oscillatorypathways can give rise to extremely rich temporal sig-nalling programs and so, perhaps, to many distinct pat-terns of expression for target genes.
MethodsDifferential EquationsThe model studied in this paper is specified by the fol-lowing ODEs. External forcing via TNFa stimulation isrepresented by the function Tr (t), whose value ranges
between 0 and 1, indicating the strength of stimulationas a fraction of the maximum possible.
NFkB IkBaNFkB IkBa FkB
NFkB IkBaNFkB
k k
k k
k
d a
i degc
1 1
1
ee degpin
d a
f k
k k1
1 1
nNFkB pIkBaNFkB;
IkBa IkBaNFkB IkBa N
FFkB
IkBa nIkBa
IkBa tIkBa
IKK IkB
k k
k k
k
i e
degf tria
c
2 2
2
*
aa;
IkBaNFkB IkBa NFkB IkBaNFkB
nIkBaNFkB
k k
k ka d
elc de
1 1* * *
ggc
c
d a
k
k k
k aNFkB
IKK IkBaNFkB;
nNFkB * nIkBaNFkB nIk1
3
1 * BBa nhNFkB
NFkB nIkBaNFkB
nNFkB
nIkB
*
* * *
* * ;
k k k
k kv i degcn
v e f
1
1
a nIkBaNFkB nIkBa n FkB
IkBa nIkBa
kd a
v i v e
k
k k k k1 1
2 2
* * *
k f
k k
k
def
a d
v
*
* *
nIkBa;
nIkBaNFkB nIkBa nNFkB nIkBaNFkB1 1 kk
k
k k
elc
degcn
ctria degtia
*
;
*
nIkBaNFkB
nIkBaNFkB
tIkBa tIk
BBa
nNFkB
nNFkB
IKKn IKKi
kk
kk
k
irtia
h
h h
pbA
bA
* ;
* * 20
200
A20
IKKn
IKK IKKn IKK;
IKKi
*
* * ;
* * *
T
T k
T k k
k
r
r a
r a i
i **
* **
;
*
IKK
IKKiA20
tA20 t
kk
k T
k k
pbA
bA r
ctra degta
20
20
AA20
*nNFkB
nNFkB
A20 tA20 A20
kk
k k
itra
h
h h
tra da
;
* * ;
ppIkBa IKK IkBa pIkBa;
pIkBaNfkB IKK IkBaN
k k
kc degpi
c
2
3
* * *
* * FFkB
pIkBaNFkB; kdegpin *
Initial conditionsAll the numerical experiments reported above beganwith the initial concentrations
IkBaNFkB 0.1042424 and IKKn 0.1042424
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 8 of 11

while all others start at zero. We then integrated theODEs through 2000 minutes of simulated time and usedthe result as an initial condition for the various TNFa-stimulation studies.
NumericsThe bifurcation diagram in Figure 3 and the plot of theArnol’d tongues in Figure 9 were prepared using BardErmentrout’s XPPAUT [29], a front end for the power-ful numerical bifurcation package AUTO [30]. All ourother numerical work used MATLAB© to integrate theODEs, compute the power spectra and plot the figures.
Appendix: circle mapsHere we discuss a standard mathematical tool, the circlemap, used to study the response of a nonlinear oscilla-tor subjected to periodic forcing. The idea is to considersufficiently weak forcing that the response is close tothat of the unforced system, and then define a phaseangle j that is close to the phase of the correspondingunforced oscillator.If ω0 is the frequency of the unperturbed oscillator
and ω is the forcing frequency then, after one period ofthe forcing, j will have advanced by
0
0 2
G
G
( )
/ ( )
where τ = 2π/ω is the period of the forcing, ε is ameasure of its amplitude and G(j) is a phase-dependentfunction that characterizes the response.The simplest circle map, studied here by way of illus-
tration, is given by a function F : [0, 2π) ® [0, 2π) ofthe form of
F( )
sin( )
sin( ),
0
2
(6)
where a = (ω0/ω), and 0 < ε ≪ 2π. The dynamics of acircle map can be characterized with a single parametercalled the rotation number. For a given initial point j0,the rotation number is defined as the long-term averagephase shift per one iteration (that is, per one period ofthe forcing):
( ) lim ,0
0
2
n
n
n(7)
where
nn n nF F F F F F ( ) ( ) ....0
10
20
One can show that the limit in the definition of therotation number (7) exists and does not depend on theinitial point j0 [31], p.102. The qualitative dynamics ofthe forced system are thus of two types: motions withrational rotation numbers and those with irrationalones. Further, the rotation number is retional-say
pq, with p Î N and q Î ℤ+ —if and only if there
exists some j0 Î [0, 2π) such that
0 02 p F q . (8)
That is, rational rotation numbers correspond to peri-odic dynamics: the orbit of j0 is called a (p, q) cycle. Onthe other hand, irrational rotation numbers correspondto quasiperiodic dynamics [27,31,32].According to a theorem of Denjoy [33], if the rotation
number is irrational there is a continuous, invertiblechange of coordinates h: [0, 2π) ® [0, 2π), say, h(j) = θ,such that
h F h F 2 2or
Where F h F h 1 . This implies that
forced oscillators whose corresponding circle maps haveirrational rotation number never repeat periodically.Results of Arnold [31] show that the qualitative beha-
viour of the circle map (6) is stable against arbitrarysmall perturbations if and only if the rotation number isrational. We can thus expect that if F has a rational
rotation numberpq, there exists a region of parameter
values (a, ε) such that the all the forced systems whoseparameters lie in this region share the same rationalrotation number: that is, they all have the same sorts ofperiodic response. Such regions of parameter values arecalled Arnol’d tongues.
A similar story holds for the sinusoidally forced NF-�B system: when the time-average of the stimulation issufficiently strong that the system supports periodicoscillations with frequency ω0, and when the amplitudeof the forcing is sufficiently weak that it is sensible tomeasure a phase j with respect to that of the intrinsicoscillator, then one can construct numerically a circlemap of the form
F T G 0 , (9)
where T = 2π/ω is the period of the forcing. Althoughthe function G’ in (9) is not as simple as the the G (j)in (6), the map F’ still has (p, q) periodic responses and
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 9 of 11

a corresponding system of Arnol’d tongues: Figure 9shows examples.We’ll conclude this Appendix by explaining why, at
least for the simple circle map (6), the Arnol’d tonguesare wedge-shaped. Consider the case where F has
rational rotation number pq. Then, as mentioned
above, there exists a periodic point j0 satisfying (8). Ifwe expand Fq (j0) in powers of ε we find
F qq
q
0 0
0 1 1
2
2
sin sin sin
Consider the case where a is very close to a rational
number: pq
with |b| ≪ 1/q. Then
F p Gqp q 0 0 02 / , , ,
Where
G
q
q
p q
q
/ , ,
sin sin
sin
0
0 1
0
2
2
ssin F q
10
(10)
It is not hard to show that GP/q is bounded and contin-uous when regarded as a function of j0 and so attains itsmaximum and minimum values on [0, 2π]. Thus, foreach fixed ε, there is an interval of b on which GP/q = 0for some j Î [0, 2π]. Now consider the way in which theend points of this interval depend on ε. The case q = 1and p = 0 is especially clear: Eqn. (10) becomes
Gp q/ , , sin 2 0
which has some solutions j Î [0, 2π] provided that |2πb| ≤ ε or |b| ≤ ε/2π.The values q = 1 and p = 1 correspond to the case where
the periodic response has the same frequency as the forcedsystem and thus undergoes one complete phase-rotationper period of the forcing. The analysis sketched aboveshows that this particular form of mode-locking will per-tain for (ε, a) in a wedge-shaped region whose widthincreases linearly with forcing amplitude ε. For more gen-eral mode-locking resonances, where q ≥ 2, Arnol’dshowed in [31] that the width of the tongue satisfies:
q.
AcknowledgementsCAH and YW were supported by BBSRC grant BBD0088081. PP and MRHWwere supported by MRC Grant G0500346. PP, MRM, DSB and MRHW weresupported by BBSRC grants BBF005381 & BBF00561X1.
Author details1Mathematical Biosciences Institute, The Ohio State University, Jennings Hall,Columbus, Ohio 43210, USA. 2Centre for Cell Imaging, School of BiologicalSciences, Bioscience Research Building, Crown Street, Liverpool, L69 7ZB, UK.3School of Chemistry and The Manchester Interdisciplinary Biocentre,University of Manchester, 131 Princess Street, Manchester M1 7DN, UK.4School of Mathematics, Alan Turing Building, University of Manchester,Manchester M13 9PL, UK.
Authors’ contributionsYW did the numerical experiments reported here, basing her work on codeoriginally written by CAH, who also developed the underlying model. PPand CAH provided biologically-informed advice on modelling, especially onhow best to couple the model to an external signal. MRHW, DBK and DSBprovided critical readings of the manuscript, as well scientific guidancethroughout the project. MRM worked closely with YW on the computationalside and, with YW, wrote the first draft and prepared the figures. All authorsreviewed and approved the final draft.
Received: 26 April 2010 Accepted: 3 February 2011Published: 3 February 2011
References1. Hayden M, Ghosh S: Signaling to NF-κB. Genes & Development 2004,
18:2195-2224[http://dx.doi.org/10.1101/gad.1228704].2. Bonizzi G, Karin M: The two NF-κB activation pathways and their role in
innate and adaptive immunity. Trends in Immunology 2004, 25(6):280-288.3. Pasparakis M, Luedde T, Schmidt-Supprian M: Dissection of the NF-κB
signalling cascade in transgenic and knockout mice. Cell Death &Differentiation 2006, 13:861-872.
4. Pahl HL: Activators and target genes of Rel/NF-κB transcription factors.Oncogene 1999, 18(49):6853-6866[http://www.nature.com/onc/journal/v18/n49/abs/1203239a.html].
5. Ghosh S, May M, Kopp E: NF-κB and rel proteins: Evolutionarily conservedmediators of immune responses. Annual Review of Immunology 1998,16:225-260.
6. Scott ML, Fujita T, Liou HC, Nolan GP, Baltimore D: The p65 subunit of NF-kappa B regulates I kappa B by two distinct mechanisms. Genes &Development 1993, 7(7a):1266-1276[http://genesdev.cshlp.org/content/7/7a/1266.abstract].
7. Song HY, Rothe M, Goeddel DV: The tumor necrosis factor-inducible zincfinger protein A20 interacts with TRAF1/TRAF2 and inhibits NF-kappaBactivation. Proceedings of the National Academy of Sciences of the UnitedStates of America 1996, 93(13):6721-6725.
8. Wertz IE, O’Rourke KM, Zhou H, Eby M, Aravind L, Seshagiri S, Wu P,Wiesmann C, Baker R, Boone DL, Ma A, Koonin EV, Dixit VM: De-ubiquitination and ubiquitin ligase domains of A20 downregulate NF-κBsignalling. Nature 2004, 430:694-699.
9. Pigolotti S, Krishna S, Jensen MH: Oscillation patterns in negativefeedback loops. Proceedings of the National Academy of Sciences 2007,104(16):6533-6537.
10. Thomas R: Logical analysis of systems comprising feedback loops. Journalof Theoretical Biology 1978, 73(4):631-656.
11. Hoffmann A, Levchenko A, Scott M, Baltimore D: The I kappa B-NF-kappaB signaling module: Temporal control and selective gene activation.Science 2002, 298(5596):1241-1245.
12. Nelson DE, Ihekwaba AEC, Elliott M, Johnson JR, Gibney CA, Foreman BE,Nelson G, See V, Horton CA, Spiller DG, Edwards SW, McDowell HP, Unitt JF,Sullivan E, Grimley R, Benson N, Broomhead DS, Kell DB, White MRH:Oscillations in NF-kappa B signaling control the dynamics of geneexpression. Science 2004, 306(5696):704-708.
13. Horton CA: Computational Modelling of Cell Signalling Pathway Dynamics.PhD thesis School of Biological Sciences, University of Liverpool; 2006.
14. Garside H, Stevens A, Farrow S, Normand C, Houle B, Berry A, Maschera B,Ray D: Glucocorticoid ligands specify different interactions with NF-
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 10 of 11

kappa B by allosteric effects on the glucocorticoid receptor DNAbinding domain. Journal of Biological Chemistry 2004, 279(48):50050-50059.
15. Ikeda A, Sun X, Li Y, Zhang Y, Eckner R, Doi T, Takahashi T, Obata Y,Yoshioka K, Yamamoto K: p300/CBP-dependent and -independenttranscriptional interference between NF-kappa B RelA and p53.Biochemical and Biophysical Research Communications 2000, 272(2):375-379.
16. Perkins ND: Integrating cell-signalling pathways with NF-κkB and IKKfunction. Nature Reviews Molecular Cell Biology 2007, 8:49-62.
17. Salminen A, Ojala J, Huuskonen J, Kauppinen A, Suuronen T, Kaarniranta K:Interaction of aging-associated signaling cascades: Inhibition of NF-kappa B signaling by longevity factors FoxOs and SIRT1. Cellular AndMolecular Life Sciences 2008, 65(7-8):1049-1058.
18. Wadgaonkar R, Phelps K, Haque Z, Williams A, Silverman E, Collins T: CREB-binding protein is a nuclear integrator of nuclear factor-kappa B andp53 signaling. Journal of Biological Chemistry 1999, 274(4):1879-1882.
19. Ashall L, Horton CA, Nelson DE, Paszek P, Harper CV, Sillitoe K, Ryan S,Spiller DG, Unitt JF, Broomhead DS, Kell DB, Rand DA, See V, White MRH:Pulsatile stimulation determines timing and specificity of NF-kappaB-dependent transcription. Science 2009, 324(5924):242-246.
20. Fonslet J, Rud-Petersen K, Krishna S, Jensen MH: Pulses and Chaos:Dynamical Response in a Simple Genetic Oscillator. International Journalof Modern Physics B 2007, 21(23-24):4083-4090.
21. Krishna S, Jensen MH, Sneppen K: Minimal model of spiky oscillations inNF-κB signaling. PNAS 2006, 103(29):10840-10845.
22. Nelson DE: Charaterisation and Manipulation of NF-κB Signal Oscillationsin Single Living Cells. PhD thesis School of Biological Sciences, University ofLiverpool; 2005.
23. Matalka KZ, Tutunji MF, Abu-Baker M, Abu Baker Y: Measurement ofprotein cytokines in tissue extracts by enzyme-linked immunosorbentassays: Application to lipopolysaccharide-induced differential milieu ofcytokines. Neuroendocrinology Letters 2005, 26(3):231-236.
24. Prabha C, Jalapathy KV, Matsa RP, Das SD: Role of TNF-alpha in hostimmune response in tuberculous pleuritis. Current Science 2003,85(5):639-642.
25. Wang Y, Paszek P, Horton CA, Hong Y, White MRH, Kell DB, Muldoon MR,Broomhead DS: A systematic survey of the response of a model NF-κBsignalling pathway to TNFα stimulation. MIMS EPrint 2009.104 ManchesterInstitute for Mathematical Sciences, The University of Manchester; 2010[http://eprints.ma.man.ac.uk/1404/].
26. Wiggins S: Introduction to Applied Nonlinear Dynamical Systems and ChaosNew York; London: Springer Verlag; 1990.
27. Pikovsky A, Rosenblum M, Kurths J: Synchronization : A Universal Concept inNonlinear Sciences Cambridge, UK: Cambridge University Press; 2003.
28. Nayfeh AH, Mook DT: Nonlinear Oscillators New York: John Wiley & Sons;1979.
29. Ermentrout B: Simulating, Analyzing, and Animating Dynamical Systems: AGuide to XPPAUT for Researchers and Students Philadelphia: SIAM; 2002.
30. Doedel EJ, Paffenroth RC, Champneys AR, Fairgrieve TF, Kuznetsov YA,Oldeman BE, Sandstede B, Wang XJ: AUTO2000 : Software forcontinuation and bifurcation problems in ordinary differential equations.Tech rep California Institute of Technology, Department of Applied andComputational Mathematics, Pasadena CA 91125; 2000.
31. Arnold VI: Geometrical Methods in the Theory of Ordinary DifferentialEquations. 2 edition. New York; London: Springer Verlag; 1988.
32. Arrowsmith DK, Place CM: An introduction to Dynamical Systems Cambridge,UK: Cambridge University Press; 1990.
33. Denjoy A: Sur les courbes definies par les equations differentielles a lasurface du tore. J Math 1932, 17(IV):333-375.
doi:10.1186/1752-0509-5-23Cite this article as: Wang et al.: Interactions among oscillatory pathwaysin NF-kappa B signaling. BMC Systems Biology 2011 5:23.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
Wang et al. BMC Systems Biology 2011, 5:23http://www.biomedcentral.com/1752-0509/5/23
Page 11 of 11