interaction of chk1 with treslin negatively regulates the
TRANSCRIPT

Article
Interaction of Chk1 with T
reslin NegativelyRegulates the Initiation of Chromosomal DNAReplicationGraphical Abstract
Highlights
d Chk1 binds to a C-terminal region of Treslin called the TRCT
domain
d Mutations in the TRCT domain abolish phosphorylation of
Treslin by Chk1
d Disruption of the Treslin-Chk1 interaction increases initiation
of DNA replication
d Chk1 regulates Treslin during an apparently normal S phase
Guo et al., 2015, Molecular Cell 57, 492–505February 5, 2015 ª2015 Elsevier Inc.http://dx.doi.org/10.1016/j.molcel.2014.12.003
Authors
Cai Guo, Akiko Kumagai, ...,
Andrej Shevchenko, William G. Dunphy
In Brief
In vertebrates, Treslin is necessary for
activation of the replicative helicase and
the ensuing initiation of DNA replication.
Previous studies have indicated that S
phase cyclin-dependent kinase activity
positively regulates the action of Treslin.
Guo et al. now demonstrate that the
checkpoint-regulatory kinase Chk1
negatively regulates the initiation-
promoting function of Treslin.

Molecular Cell
Article
Interaction of Chk1 with TreslinNegatively Regulates the Initiationof Chromosomal DNA ReplicationCai Guo,1 Akiko Kumagai,1 Katharina Schlacher,2,3,5 Anna Shevchenko,4 Andrej Shevchenko,4 and William G. Dunphy1,*1Division of Biology and Biological Engineering, California Institute of Technology, Pasadena, CA 91125, USA2Department of Molecular and Medical Pharmacology, University of California, Los Angeles, CA 90095, USA3Developmental Biology Program, Memorial Sloan-Kettering Cancer Center, New York, NY 10065, USA4Max Planck Institute of Molecular Cell Biology and Genetics, 01307 Dresden, Germany5Present Address: Department of Cancer Biology, MD Anderson Cancer Center, Houston, TX 77054, USA
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.molcel.2014.12.003
SUMMARY
Treslin helps to trigger the initiation of DNA replica-tion by promoting integration of Cdc45 into the repli-cative helicase. Treslin is a key positive-regulatorytarget of cell-cycle control mechanisms; activationof Treslin by cyclin-dependent kinase is essentialfor the initiation of replication. Here we demonstratethat Treslin is also a critical locus for negative regula-tory mechanisms that suppress initiation. We foundthat the checkpoint-regulatory kinase Chk1 associ-ates specifically with a C-terminal domain of Treslin(designated TRCT). Mutations in the TRCT domainabolish binding of Chk1 to Treslin and thereby elimi-nate Chk1-catalyzed phosphorylation of Treslin.Significantly, abolition of the Treslin-Chk1 interactionresults in elevated initiation of chromosomal DNAreplication during an unperturbed cell cycle, whichreveals a function for Chk1 during a normal S phase.This increase is due to enhanced loading of Cdc45onto potential replication origins. These studies pro-vide important insights into how vertebrate cellsorchestrate proper initiation of replication.
INTRODUCTION
In eukaryotic cells, duplication of the genome depends upon the
intricate, stepwise assembly of protein complexes onto origins
of DNA replication (Sclafani and Holzen, 2007; Siddiqui et al.,
2013; Tanaka and Araki, 2013). Initially, the origin recognition
complex (ORC) and Cdc6 associate with potential origins and
thereupon recruit Cdt1 and the mini-chromosome maintenance
(MCM) complex. The MCM complex serves as the core of the
replicative helicase that unwinds the DNA strands for replication.
A key regulatory juncture in replication involves the concerted
binding of additional proteins to the MCM complex to form the
mature, activated version of the helicase. In particular, the
Cdc45 and GINS proteins associate with the MCM proteins
and thereby form the CMG (Cdc45-MCM-GINS) complex, which
corresponds to the fully constituted helicase.
In vertebrates, integration of Cdc45 and GINS with the MCMs
depends upon TopBP1 and a recently discovered TopBP1-bind-
ing protein called Treslin (also known as Ticrr) (Kumagai et al.,
2010; Sansam et al., 2010). Importantly, formation of the
TopBP1-Treslin complex requires phosphorylation of Treslin by
the S phase cyclin-dependent kinase (S-CDK) (Boos et al.,
2011; Kumagai et al., 2011). Consequently, this phosphorylation
helps to explain how the cell-cycle control system dictates the
timing of S phase. An analogous situation exists in budding yeast
where phosphorylation of the Treslin homolog Sld3 by S-CDK is
also critical for replication (Labib, 2010; Siddiqui et al., 2013; Ta-
naka and Araki, 2013).
Treslin is a relatively large protein (220 kD) that is approxi-
mately three times bigger than yeast Sld3. Thus, Treslin may
have acquired newproperties that allow it tomeet themore com-
plex demands of higher eukaryotes. To obtain further insight into
established and potentially novel functions of Treslin, we have
engaged in a search for Treslin-interacting proteins in human
cells. These studies resulted in the identification of Chk1, an
effector kinase in checkpoint control mechanisms. Chk1 is
best known for its role in blocking activation of CDKs in cells
with incompletely replicated or damaged DNA (Perry and Korn-
bluth, 2007; Toledo et al., 2011).
Significantly, further studies have indicated that Chk1 also
plays a role during a seemingly normal cell cycle. For
example, a number of observations have implicated Chk1 in
the control of replication during an unperturbed S phase
(Maya-Mendoza et al., 2007; McIntosh and Blow, 2012;
Miao et al., 2003; Syljuasen et al., 2005). The mechanism by
which Chk1 exerts these effects is obscure. Chk1 also partic-
ipates in suppressing endoreplication in trophoblast stem (TS)
cells (Ullah et al., 2011). Moreover, Chk1 appears to function
in the system that monitors correct attachment of chromo-
somes to the mitotic spindle (Zachos et al., 2007). Overall,
these observations suggest that Chk1 has diverse roles in
cell-cycle regulation, which may help to explain why it is
essential for viability (Liu et al., 2000). Therefore, it will be
important to understand how cells control the participation
of Chk1 in these varied functions.
492 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.
In this report, we have investigated the molecular mechanism
and functional consequences of the Treslin-Chk1 interaction in
both human cells and Xenopus egg extracts. We show that
Chk1 negatively regulates the Treslin-mediated loading of
Cdc45 onto chromatin and thereby serves to antagonize the
initiation of replication. These studies provide an important
perspective on how vertebrate cells control the initiation of
DNA replication through opposing negative and positive regula-
tory mechanisms. Furthermore, these experiments reveal the
mechanistic basis for a critical function of Chk1 apart from its
role in checkpoint responses to damaged DNA.
RESULTS
Chk1 Is a Treslin-Interacting ProteinTo search for Treslin-interacting proteins, we expressed various
tagged versions of Treslin in human 293T cells, reisolated these
polypeptides, and then analyzed associated proteins by mass
spectrometry (Figure 1A). For these experiments, we produced
recombinant full-length human Treslin with both S peptide and
3X-FLAG tags at the C-terminal end (designated Treslin-SF).
We also prepared tagged fragments corresponding to residues
1–1,257 and 1,253–1,909 of the protein. The 1–1,257 fragment
can restore DNA replication to Treslin-depleted cells (Kumagai
A B
S-protein pull-down 5% Input
TopBP1
Chk1
FLAG
Chk1
Treslin
C
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8 9 10 11
pcDNA5/T
O
Tresli
n 1-19
09
1-125
7
1253
-1909
pcDNA5/T
O
Tresli
n 1-19
09
1-125
7
1253
-1909
InputContro
l IP
Chk1 IP
InputContro
l IP
Tresli
n IP
InputContro
l IP
Chk1 IP
, left o
n ice
Chk1 IP
+ buffe
r
Chk1 IP
+ λP
Pase
Treslin-1-1909-SF
293T
FLAG IP
Elute with 3X-FLAG peptide
S-protein pull-down
SDS-PAGE
transfect
48 hr later, harvest
Treslin-1-1257-SFTreslin-1253-1909-SF
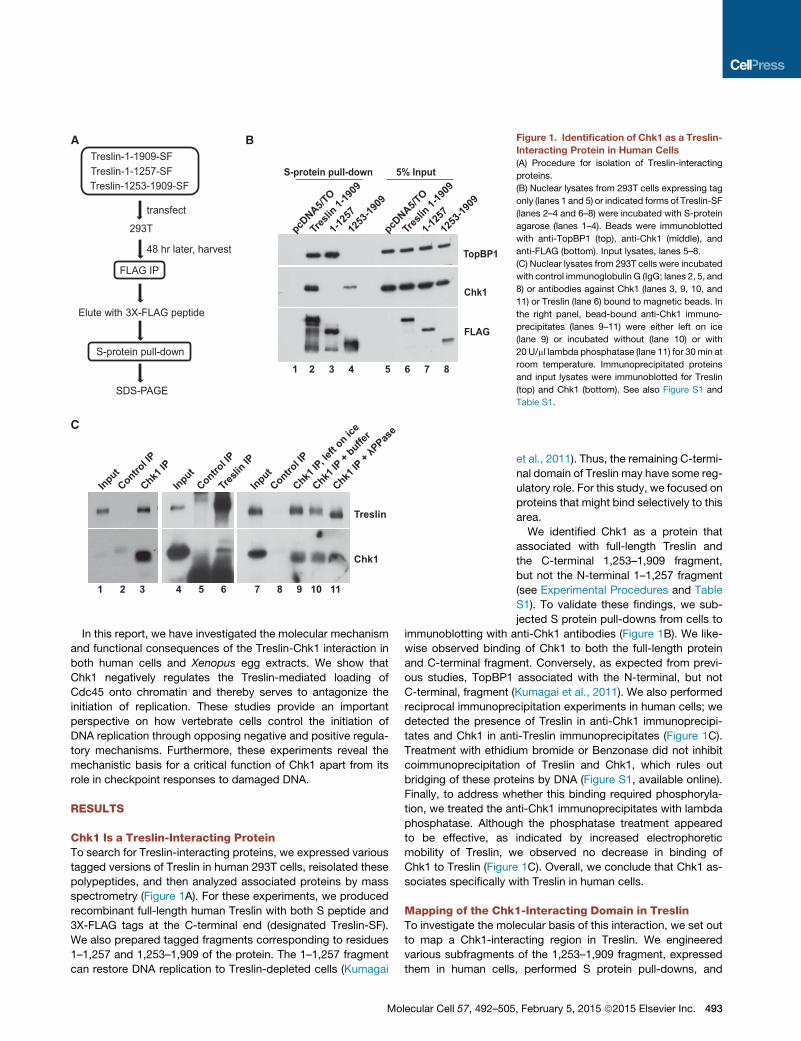
Figure 1. Identification of Chk1 as a Treslin-
Interacting Protein in Human Cells
(A) Procedure for isolation of Treslin-interacting
proteins.
(B) Nuclear lysates from 293T cells expressing tag
only (lanes 1 and 5) or indicated forms of Treslin-SF
(lanes 2–4 and 6–8) were incubated with S-protein
agarose (lanes 1–4). Beads were immunoblotted
with anti-TopBP1 (top), anti-Chk1 (middle), and
anti-FLAG (bottom). Input lysates, lanes 5–8.
(C) Nuclear lysates from 293T cells were incubated
with control immunoglobulin G (IgG; lanes 2, 5, and
8) or antibodies against Chk1 (lanes 3, 9, 10, and
11) or Treslin (lane 6) bound to magnetic beads. In
the right panel, bead-bound anti-Chk1 immuno-
precipitates (lanes 9–11) were either left on ice
(lane 9) or incubated without (lane 10) or with
20 U/ml lambda phosphatase (lane 11) for 30 min at
room temperature. Immunoprecipitated proteins
and input lysates were immunoblotted for Treslin
(top) and Chk1 (bottom). See also Figure S1 and
Table S1.
et al., 2011). Thus, the remaining C-termi-
nal domain of Treslin may have some reg-
ulatory role. For this study, we focused on
proteins that might bind selectively to this
area.
We identified Chk1 as a protein that
associated with full-length Treslin and
the C-terminal 1,253–1,909 fragment,
but not the N-terminal 1–1,257 fragment
(see Experimental Procedures and Table
S1). To validate these findings, we sub-
jected S protein pull-downs from cells to
immunoblotting with anti-Chk1 antibodies (Figure 1B). We like-
wise observed binding of Chk1 to both the full-length protein
and C-terminal fragment. Conversely, as expected from previ-
ous studies, TopBP1 associated with the N-terminal, but not
C-terminal, fragment (Kumagai et al., 2011). We also performed
reciprocal immunoprecipitation experiments in human cells; we
detected the presence of Treslin in anti-Chk1 immunoprecipi-
tates and Chk1 in anti-Treslin immunoprecipitates (Figure 1C).
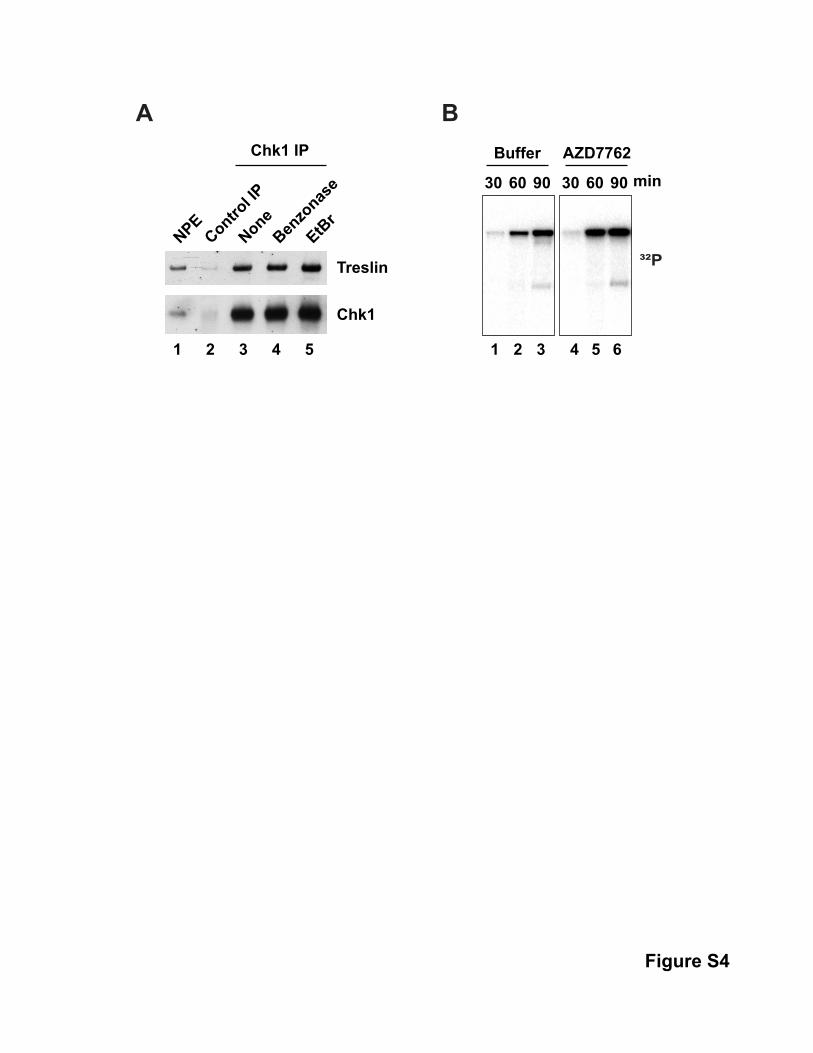
Treatment with ethidium bromide or Benzonase did not inhibit
coimmunoprecipitation of Treslin and Chk1, which rules out
bridging of these proteins by DNA (Figure S1, available online).
Finally, to address whether this binding required phosphoryla-
tion, we treated the anti-Chk1 immunoprecipitates with lambda
phosphatase. Although the phosphatase treatment appeared
to be effective, as indicated by increased electrophoretic
mobility of Treslin, we observed no decrease in binding of
Chk1 to Treslin (Figure 1C). Overall, we conclude that Chk1 as-
sociates specifically with Treslin in human cells.
Mapping of the Chk1-Interacting Domain in TreslinTo investigate the molecular basis of this interaction, we set out
to map a Chk1-interacting region in Treslin. We engineered
various subfragments of the 1,253–1,909 fragment, expressed
them in human cells, performed S protein pull-downs, and
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 493
A
B
D E
1484
-1909
1484
-1818
1585
-1909
1585
-1818
1810
-1909
1810
-1872
1253
-1909
1253
-1702
pcDNA5/T
O
1484
-1909
1484
-1818
1585
-1909
1585
-1818
1810
-1909
1810
-1872
1253
-1909
1253
-1702
pcDNA5/T
O
Binding to Chk1Human Treslin1-19091-12571253-17021253-19091484-18181484-19091585-18181585-19091810-18721810-19091870-1909
--+-
+
+-+-+-
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
S-protein pull-down
C
TopBP1
Chk1
1-125
7
1-190
9 7A
1-190
9 WT
pcDNA5/T
O
1-125
7
1-190
9 7A
1-190
9 WT
pcDNA5/T
O
5% In
put
GST18
10-19
09
S1887
A
S1893
A
T1897
A
7A 1810
-1872
1870
-1909
Chk1
CBB
S-protein pull-downGST pull-down
Human 1810 AWQ--LPSTGDEEVFVSGSTPPPSCAVRSCLSASALQALTQSPLLFQGKTPSSQSKDPRDXenopus 1876 SYLSSSQQSICDDVFNMSDFTPPSKVPKNPLSACGLLTLTQSPLLYKGKTPSSKRKEKIQZebrafish 1747 ------------EQFAWMGRKVDTPKVKKQVSASGIFALTQSPLLYKKSAVIKEAT----
* * *Human 1868 EDVD--------------VLPSTV-EDSPFSRAFSRRRPISRTYTRKKLMGTWLEDLXenopus 1936 DVFSDGDSDHGTPTLKRPTNPAAVSDDSPF-RKVNPLRSISKTYSRKKLIT------Zebrafish 1791 -QFSGSKSE---------------LEISPL-CQPRRRRTPSRTYSRKKLLD------
Input
Input
FLAG
1 2 3 4 5 6 7 81 2 3 4 5 6 7 8 9
Chk1
FLAG
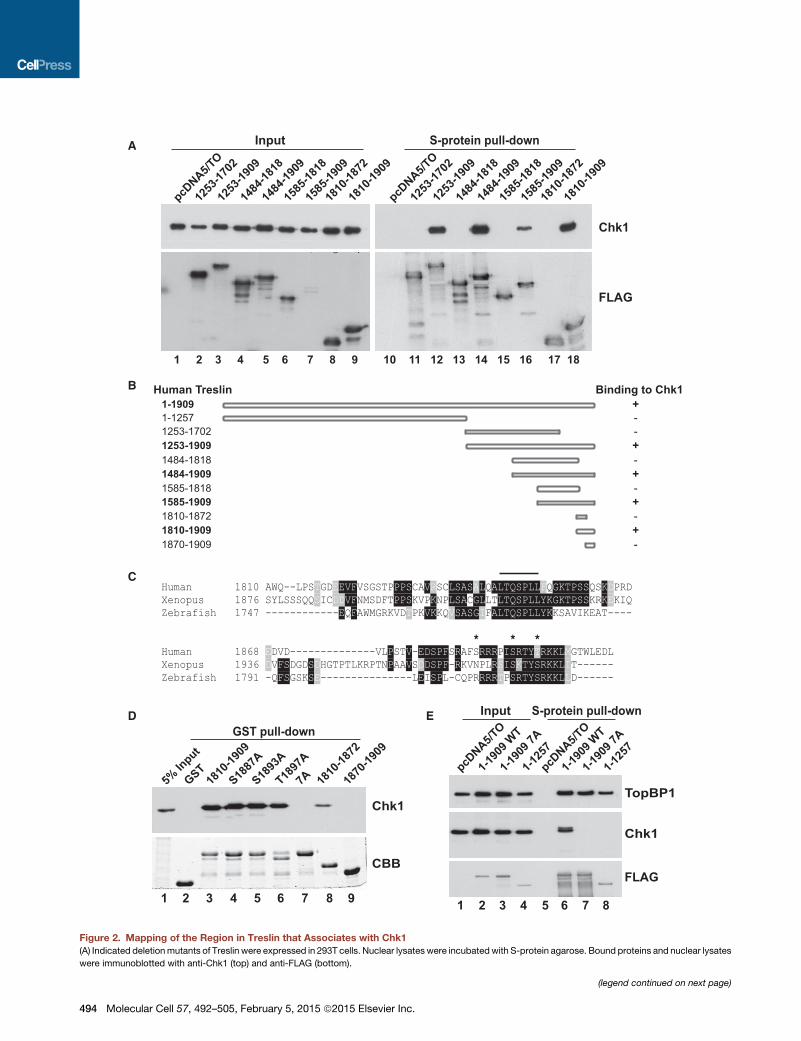
Figure 2. Mapping of the Region in Treslin that Associates with Chk1
(A) Indicated deletionmutants of Treslin were expressed in 293T cells. Nuclear lysates were incubatedwith S-protein agarose. Bound proteins and nuclear lysates
were immunoblotted with anti-Chk1 (top) and anti-FLAG (bottom).
(legend continued on next page)
494 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.

immunoblotted for Chk1. We found that approximately 100
amino acids at the C-terminal end of the protein were necessary
for binding to Chk1 (Figures 2A and 2B). We proceeded to show
that a 100 amino acid fragment from theC-terminal end (residues
1,810–1,909) was also sufficient for binding. The sequence of
this region is well conserved in the Xenopus and zebrafish homo-
logs of Treslin (Figure 2C). Accordingly, we named this region the
TRCT (Treslin C-terminal) domain.
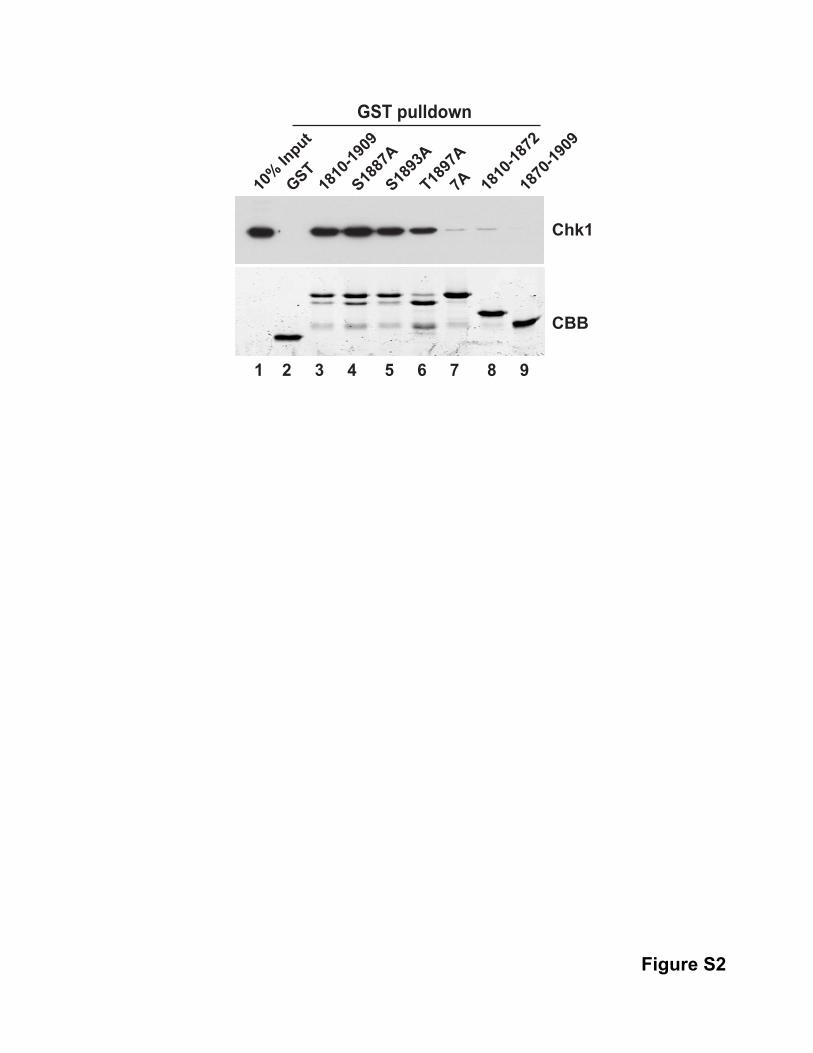
To characterize the TRCT, we prepared this domain as a GST
fusion protein (Figure 2D). The purified GST-TRCT bound well
to Chk1 in human cell lysates. By contrast, GST-tagged subfrag-
ments of this region (e.g., residues 1,810–1,872 and 1,870–1,909)
did not associate with Chk1 significantly, which suggests that the
whole domain is necessary for binding. The most conserved
stretch within the human TRCT corresponds to the sequence
LTQSPLL at positions 1,846–1,852. This sequence is identical
in Xenopus and zebrafish homologs of Treslin but is not present
in budding yeast Sld3.Wemutated each residue in this sequence
toalanine (to create the7Amutant) in the context of both theGST-
TRCT construct and the full-length Treslin-SF protein. We found
that 7A mutant versions of these polypeptides were completely
defective for binding to Chk1 (Figures 2D and 2E). Notably, the
7A mutant of full-length Treslin bound TopBP1 normally. We
also identified three residues within the TRCT (S1887, S1893,
and T1987) that sit in consensus sequences for phosphorylation
by Chk1 (RXXS/T). However, mutation of any one of these resi-
dues to alanine had no effect on the binding to Chk1 (Figure 2D).
Finally, we assessed whether Treslin associates with Chk1
directly. For this purpose, we incubated the TRCT domain with
purified, recombinant Chk1 (Figure S2). We observed that the
wild-type (WT) TRCT bound to isolated Chk1 very efficiently,
whereas there was virtually no binding of the 7A mutant. Since
no phosphorylation could occur under these conditions, this
observation reinforces the concept that phosphorylation of the
TRCT is not necessary for binding to Chk1. Overall, these results
indicate that Chk1 associates directly in a highly specific manner
with sequences in the C-terminal region of Treslin.
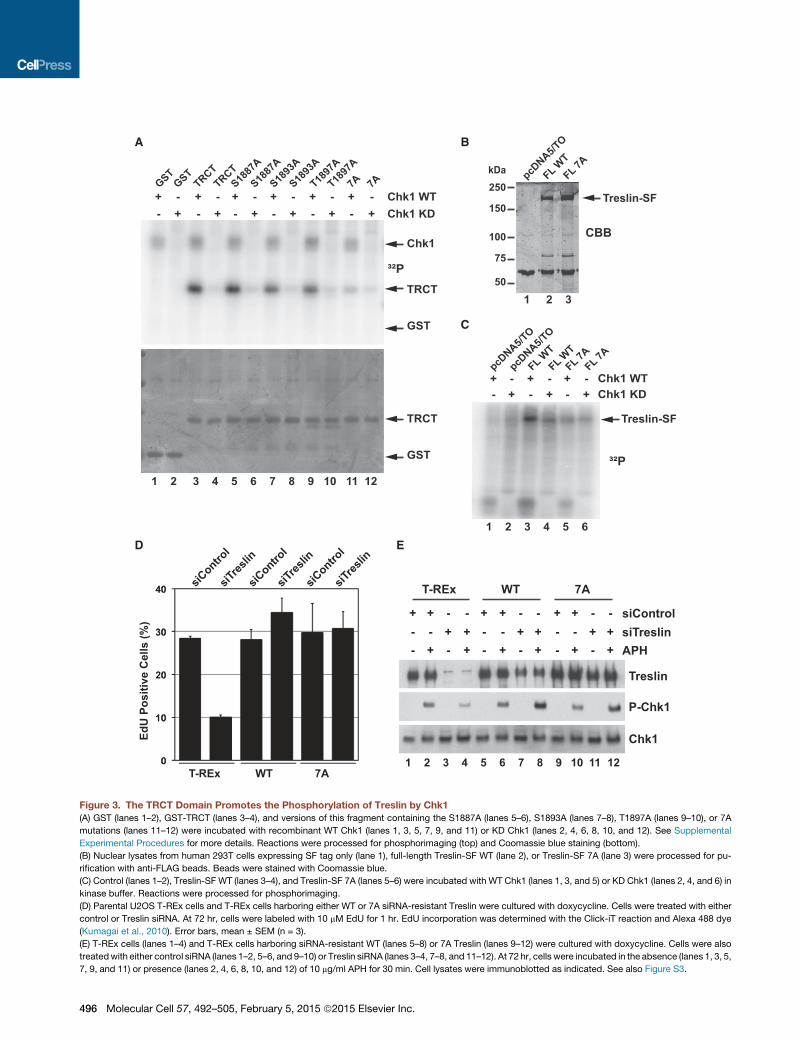
The TRCT Domain Promotes Chk1-CatalyzedPhosphorylation of TreslinOne explanation for the binding of Chk1 to the TRCT would be
that it facilitates phosphorylation of Treslin by Chk1. To address
this possibility, we first examined whether Treslin could serve as
a substrate of Chk1. We initially tested the GST-TRCT construct
and found that WT recombinant Chk1, but not kinase-dead (KD)
Chk1, could phosphorylate this fragment well (Figure 3A). By
contrast, there was no phosphorylation of the mutant GST-
TRCT-7A fragment by Chk1 above background levels. The
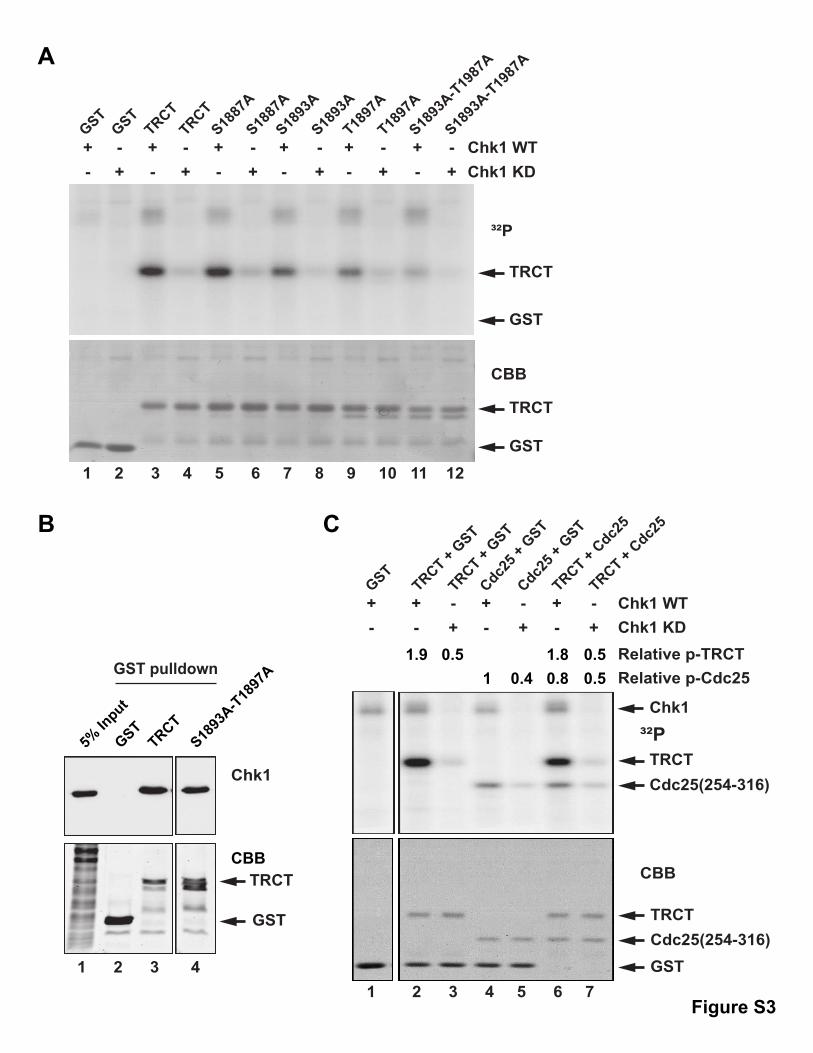
S1893A and T1897A mutants of the TRCT domain showed
reduced phosphorylation by Chk1, while the S1887A mutant
was still an equally good substrate. A combined S1893A/
T1897A mutant displayed near-background levels of phosphor-
ylation by Chk1 (Figure S3A), which suggests that these posi-
tions are the two main in vitro phosphorylation sites within this
domain. As anticipated, this mutant also still bound normally to
Chk1 (Figure S3B). Phosphorylation of the TRCT domain by
Chk1 appears to be quite efficient. For comparison, phosphory-
lation of a GST-tagged peptide fromCdc25 that contains a single
well-documented site for Chk1 (Kumagai et al., 1998) was about
2-fold lower (Figure S3C). Finally, we likewise examined full-
length Treslin and found that the WT, but not 7A mutant protein,
could serve as good substrate for Chk1 (Figures 3B and 3C).
Taken together, these results indicate that docking of Chk1
onto the TRCT domain strongly stimulates phosphorylation of
Treslin by Chk1.
We employedmass spectrometry to identify in vitro phosphor-
ylation sites for Chk1 on full-length Treslin. This analysis resulted
in the identification of numerous sites throughout much of the
protein (not shown). For these studies, we decided to focus on
using the 7A mutant to investigate the functional significance
of the Treslin-Chk1 interaction. This mutant is not an effective
substrate for Chk1 in vitro and most likely in vivo. Furthermore,
this approach would also address the possibility that the phys-
ical association of Chk1 with Treslin may also have a regulatory
impact apart from phosphorylation.
Binding of Treslin Is Not Essential for Activation of Chk1We previously demonstrated that ablation of Treslin from human
cells compromises both DNA replication and phosphorylation of
Chk1 upon treatment with aphidicolin (APH) (Kumagai et al.,
2010). In principle, the latter defect could reflect a direct role
for Treslin in the activation of Chk1. Alternatively, this effect could
be indirectly due to the absence of replication forks in cells
without Treslin. Because Treslin associates with Chk1, we asked
whether Treslin is directly necessary for activation of Chk1 in
APH-treated cells. For this purpose, we utilized lines of U2OS
cells in which expression of siRNA-resistant versions of WT
and 7A Treslin was under the control of a doxycycline-inducible
promoter (see Experimental Procedures). To characterize these
cell lines, we first used labeling with EdU to assess whether the
7Amutant could rescue DNA replication in Treslin siRNA-treated
cells. We found that the percentages of EdU-positive nuclei were
similar for Treslin-ablated cells expressing WT or 7A Treslin
(Figure 3D).
Next, we examined phosphorylation of Chk1 in cells treated
with APH. As expected, treatment of the parental U2OS cell
(B) Abilities of various fragments of Treslin to interact with Chk1. Data from (A) and (D); interacting fragments shown in bold.
(C) Amino acids 1,810–1,909 of human Treslin and corresponding portions of the X. laevis and zebrafish proteins were aligned with the Clustal Omega program.
Amino acids with asterisks (S1887, S1893, and T1897) or a line (1846-LTQSPLL-1852) were subjected to mutagenesis.
(D) GST only (lane 2), GST-tagged TRCT (lane 3), TRCT containing mutations S1887A (lane 4), S1893A (lane 5), T1897A (lane 6), or 7A (lane 7), and GST-tagged
forms of residues 1,810–1,872 (lane 8) and 1,870–1,909 from Treslin (lane 9) were isolated from bacteria with glutathione agarose. Bead-bound fragments were
incubated with 293T nuclear lysates. Beads were processed for immunoblotting with anti-Chk1 (top) and staining with Coomassie blue (bottom). Lane 1, input
nuclear lysate.
(E) Nuclear lysates from 293T cells expressing SF tag only (lanes 1 and 5) or indicated forms of Treslin-SF (lanes 2–4 and 6–8) were incubated with S-protein
agarose. Input lysates (lanes 1–4) and retrieved bead fractions (lanes 5–8) were immunoblotted as indicated. See also Figure S2.
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 495
Chk1 WT Chk1 KD
GSTGST
TRCTTRCT
S1887
A
S1887
A
S1893
A
S1893
A
T1897
A
T1897
A
7A 7A
³²P
+- +
- +- +
- +- +
- +- +
- +- +
- +- +
-
pcDNA5/T
O
FL WT
FL 7AFL 7A
FL WT
pcDNA5/T
O
Chk1 WT Chk1 KD
+- +
- +- +
- +- +
-
pcDNA5/T
O
³²P
CBB
BA
FL WTFL 7A
1 2 3 4 5 6 7 8 9 10 11 12
1 2 3 4 5 6
Chk1
TRCT
GST
TRCT
GST
C
T
Treslin-SF
Treslin
P-Chk1
Chk1
1 2 3 4 5 6 7 8 9 10 11 12
- + - + - + - + - + - + APHsiTreslinsiControl
- + +- - + +- - + +-+ + -- + + -- + + --
T-REx WT 7A
1 2 3
kDa250
150
100
75
50
ED
T-REx WT 7A
siContro
l
siTres
lin
siContro
l
siTres
lin
siContro
l
siTres
lin
EdU
Pos
itive
Cel
ls (%
)
Figure 3. The TRCT Domain Promotes the Phosphorylation of Treslin by Chk1
(A) GST (lanes 1–2), GST-TRCT (lanes 3–4), and versions of this fragment containing the S1887A (lanes 5–6), S1893A (lanes 7–8), T1897A (lanes 9–10), or 7A
mutations (lanes 11–12) were incubated with recombinant WT Chk1 (lanes 1, 3, 5, 7, 9, and 11) or KD Chk1 (lanes 2, 4, 6, 8, 10, and 12). See Supplemental
Experimental Procedures for more details. Reactions were processed for phosphorimaging (top) and Coomassie blue staining (bottom).
(B) Nuclear lysates from human 293T cells expressing SF tag only (lane 1), full-length Treslin-SF WT (lane 2), or Treslin-SF 7A (lane 3) were processed for pu-
rification with anti-FLAG beads. Beads were stained with Coomassie blue.
(C) Control (lanes 1–2), Treslin-SF WT (lanes 3–4), and Treslin-SF 7A (lanes 5–6) were incubated with WT Chk1 (lanes 1, 3, and 5) or KD Chk1 (lanes 2, 4, and 6) in
kinase buffer. Reactions were processed for phosphorimaging.
(D) Parental U2OS T-REx cells and T-REx cells harboring either WT or 7A siRNA-resistant Treslin were cultured with doxycycline. Cells were treated with either
control or Treslin siRNA. At 72 hr, cells were labeled with 10 mM EdU for 1 hr. EdU incorporation was determined with the Click-iT reaction and Alexa 488 dye
(Kumagai et al., 2010). Error bars, mean ± SEM (n = 3).
(E) T-REx cells (lanes 1–4) and T-REx cells harboring siRNA-resistant WT (lanes 5–8) or 7A Treslin (lanes 9–12) were cultured with doxycycline. Cells were also
treatedwith either control siRNA (lanes 1–2, 5–6, and 9–10) or Treslin siRNA (lanes 3–4, 7–8, and 11–12). At 72 hr, cells were incubated in the absence (lanes 1, 3, 5,
7, 9, and 11) or presence (lanes 2, 4, 6, 8, 10, and 12) of 10 mg/ml APH for 30 min. Cell lysates were immunoblotted as indicated. See also Figure S3.
496 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.

line with APH in the presence of control siRNA efficiently
induced the phosphorylation of Chk1 on S345 (Figure 3E). As
described previously, treatment with Treslin siRNA resulted in
markedly reduced phosphorylation of Chk1. We proceeded to
show that expression of either WT or 7A siRNA-resistant Treslin
could efficiently rescue phosphorylation of Chk1 in APH-treated
cells that had been treated with the Treslin siRNA. These obser-
vations indicate that the binding of Chk1 to the TRCT domain is
not essential for activation of Chk1 in response to APH.
The Isolated TRCT Domain of Treslin Stimulates DNAReplication in Xenopus Egg ExtractsWe next considered the possibility that Chk1 regulates DNA
replication by associating with the TRCT domain. As one
approach to examine this question, we utilized various types of
extracts from Xenopus eggs that recapitulate DNA replication
in a cell-free reaction. In particular, we utilized the nucleoplasmic
extract (NPE) system, in which replication occurs in a soluble nu-
clear fraction lacking membranes (Walter and Newport, 2000).
We also used whole-egg extracts in which DNA replication takes
place in reconstituted nuclei.
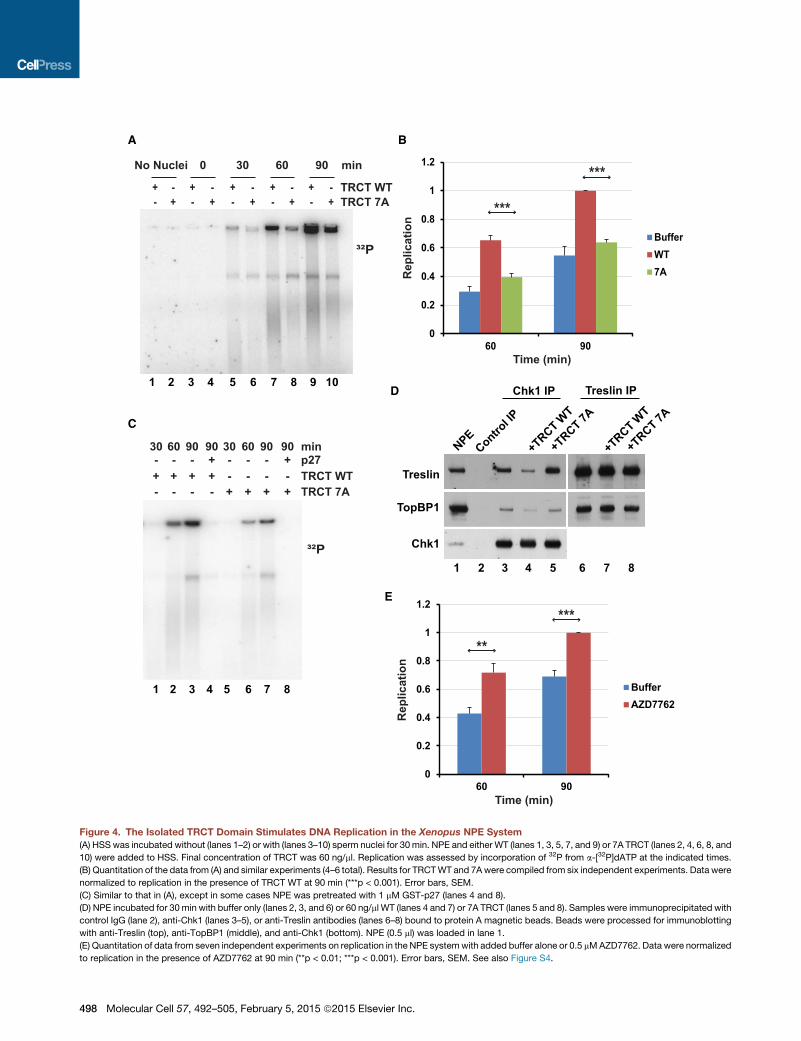
We reasoned that the isolated TRCT domain might act as a
competitor of the interaction between endogenous Treslin and
Chk1 in egg extracts. To explore this possibility, we added the
GST-TRCT to the NPE system and then monitored the time
course of DNA replication. For this purpose, we assessed incor-
poration of radioactive phosphate from a-[32P]dATP into chro-
mosomal DNA. We observed that addition of the TRCT fragment
elicited a significant increase in DNA replication in comparison
with samples treated with control buffer alone or the mutant
TRCT-7A fragment (Figures 4A and 4B).
To assess whether this replication occurs by the normal CDK-
mediated mechanism, we utilized the CDK inhibitor p27
(Figure 4C). We noted that there was no DNA replication in p27-
treated extracts in either the presence or absence of the TRCT
domain. To gauge whether the TRCT does actually prevent the
binding of Chk1 to endogenous Treslin, we performed immuno-
precipitation experiments with anti-Chk1 antibodies (Figure 4D).
We could readily detect Treslin in anti-Chk1 immunoprecipitates
from NPE fractions. Association of Treslin with Chk1 was not in-
hibited by treatmentwith ethidiumbromide or Benzonase, agents
that would prohibit bridging by DNA (Figure S4A). We observed
that addition of the WT TRCT fragment to NPE fractions caused
a severe reduction in the binding of Treslin to Chk1, whereas
the 7A mutant fragment had no effect. By contrast, the TRCT
domain had no effect on the binding of Treslin to TopBP1.Overall,
these results indicated that blockage of the binding of Chk1 to
Treslin results in elevated DNA replication.
Chk1 Regulates DNA Replication in Xenopus EggExtractsAs another means to investigate the regulation of DNA replica-
tion by Chk1, we utilized the Chk1 inhibitor AZD7762. We found
that this compound also elicited an increase in DNA replication
comparable to that induced by the TRCT-WT fragment (Figures
4E and S4B). We also attempted to immunodeplete Chk1 from
the NPE system, which entails separate immunodepletion from
the HSS and NPE fractions that are necessary for these experi-
ments. However, these immunodepletion procedures resulted
in a nonspecific reduction of replication, which confounded
this approach.
Asanalternative,weattempted todepleteChk1 fromwhole-egg
extracts (Kumagai et al., 1998).We first needed to assesswhether
the TRCT affects replication in whole-egg extracts, which would
require transport of the TRCT into reconstituted nuclei in these
extracts. Since our initial GST construct lacked a nuclear localiza-
tion sequence (NLS), we prepared a new construct with this
sequence (GST-TRCT-NLS) (Figure 5A). We verified that both
WT and 7A versions of the TRCT-NLS could be found in lysates
of reconstituted nuclei from whole-egg extracts. Unexpectedly,
the WT, but not 7A, version of the TRCT-NLS also caused a dra-
matic accumulation of Chk1 in the nuclei (Figure 5B). A plausible
explanation is that the TRCT-NLS, because of its robust binding
to Chk1, may promote nuclear entry of Chk1.
Next, we added the WT TRCT-NLS to whole-egg extracts
containing reconstituted nuclei and found that this fragment
also elicited an increase in DNA replication relative to incuba-
tions containing control buffer or a 7A mutant version of the
TRCT-NLS (Figures 5C and 5D). Finally, we removed Chk1
from whole-egg extracts with anti-Chk1 antibodies (Figure 5E).
We observed a significant acceleration of DNA replication in
Chk1-depleted extracts (Figures 5F and 5G). Furthermore,
addition of recombinant Chk1 to the depleted extracts restored
replication to the lower level found in mock-depleted extracts.
Taken together, these experiments provide multiple lines of
evidence that Chk1 regulates DNA replication in the Xenopus
system.
The Isolated TRCT Domain Promotes Initiation ofReplication in Egg ExtractsA key function of Treslin involves the loading of Cdc45 onto
chromatin (Kumagai et al., 2010). It has been well established
that the loading of Cdc45 is critical for the initiation of DNA repli-
cation in both yeast and vertebrates (Sclafani and Holzen, 2007;
Siddiqui et al., 2013; Tanaka and Araki, 2013). To explore the
basis of the TRCT-mediated stimulation of replication, we exam-
ined the loading of Cdc45 onto chromatin in the NPE system in
the presence of this fragment. We observed that addition of
the TRCT-WT protein stimulated a large increase in the loading
of Cdc45 onto chromatin in comparison with samples containing
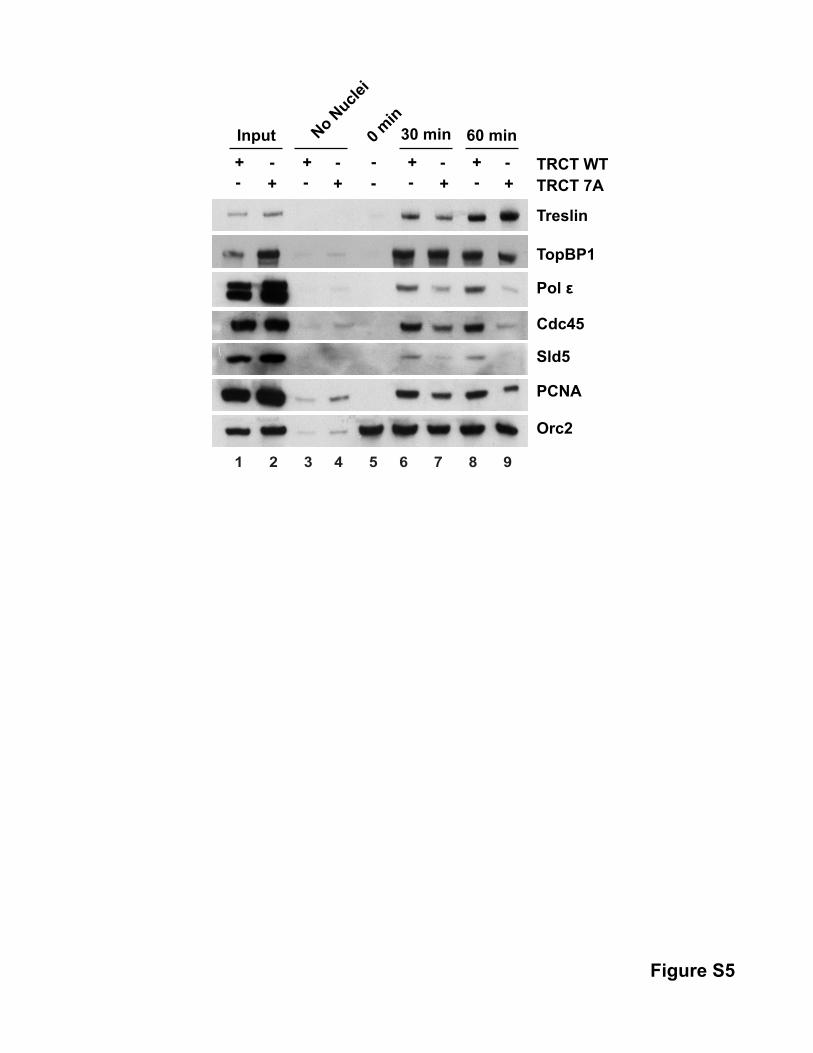
the TRCT-7A protein (Figure 6A). There was a similar increase in
the loading of Sld5 (a component of GINS), PCNA, and DNA po-
lymerase epsilon (Figure S5). The TRCT domain typically did not
affect the binding of the ORC and MCM complexes to the DNA.
These experiments suggest that the isolated TRCT domain pro-
motes initiation by stimulating the loading of the helicase activa-
tors Cdc45 and GINS onto chromatin. Furthermore, we also
observed an increase in other replication fork proteins, such as
PCNA and DNA polymerase epsilon, which participate directly
in the ensuing DNA synthesis.
As another method to characterize this phenomenon, we at-
tempted to block replication just after initiation. Actinomycin D
blocks replication in the NPE system shortly after initiation but
before significant unwinding of the DNA (Pacek and Walter,
2004). Accordingly, we added actinomycin D to the NPE system
in the absence and presence of the TRCT domain. We found that
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 497
+- +
- +- +
- +- +
- +- +
- +- +
-
³²P
+ + + +-- --+ + + + -- --- -- + - -- +
TRCT WTTRCT 7A
p27
A B
C
30 60 90 90 30 60 90 90 min
D
E
³²P
TRCT WTTRCT 7A
min30 60 90 0No Nuclei
Rep
licat
ion
Rep
licat
ion
0
0.2
0.4
0.6
0.8
1
1.2
60 90
BufferWT7A
***
***
0
0.2
0.4
0.6
0.8
1
1.2
60 90
BufferAZD7762
Time (min)
**
***
Time (min)
1 2 3 4 5 6 7 8
NPEContro
l IP
Chk1 IP
Treslin
Chk1
+TRCT W
T
+TRCT 7A
TopBP1
+TRCT W
T
+TRCT 7A
Treslin IP
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8 9 10
Figure 4. The Isolated TRCT Domain Stimulates DNA Replication in the Xenopus NPE System
(A) HSS was incubated without (lanes 1–2) or with (lanes 3–10) sperm nuclei for 30 min. NPE and either WT (lanes 1, 3, 5, 7, and 9) or 7A TRCT (lanes 2, 4, 6, 8, and
10) were added to HSS. Final concentration of TRCT was 60 ng/ml. Replication was assessed by incorporation of 32P from a-[32P]dATP at the indicated times.
(B) Quantitation of the data from (A) and similar experiments (4–6 total). Results for TRCTWT and 7Awere compiled from six independent experiments. Data were
normalized to replication in the presence of TRCT WT at 90 min (***p < 0.001). Error bars, SEM.
(C) Similar to that in (A), except in some cases NPE was pretreated with 1 mM GST-p27 (lanes 4 and 8).
(D) NPE incubated for 30 min with buffer only (lanes 2, 3, and 6) or 60 ng/ml WT (lanes 4 and 7) or 7A TRCT (lanes 5 and 8). Samples were immunoprecipitated with
control IgG (lane 2), anti-Chk1 (lanes 3–5), or anti-Treslin antibodies (lanes 6–8) bound to protein A magnetic beads. Beads were processed for immunoblotting
with anti-Treslin (top), anti-TopBP1 (middle), and anti-Chk1 (bottom). NPE (0.5 ml) was loaded in lane 1.
(E) Quantitation of data from seven independent experiments on replication in the NPE systemwith added buffer alone or 0.5 mMAZD7762. Data were normalized
to replication in the presence of AZD7762 at 90 min (**p < 0.01; ***p < 0.001). Error bars, SEM. See also Figure S4.
498 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.
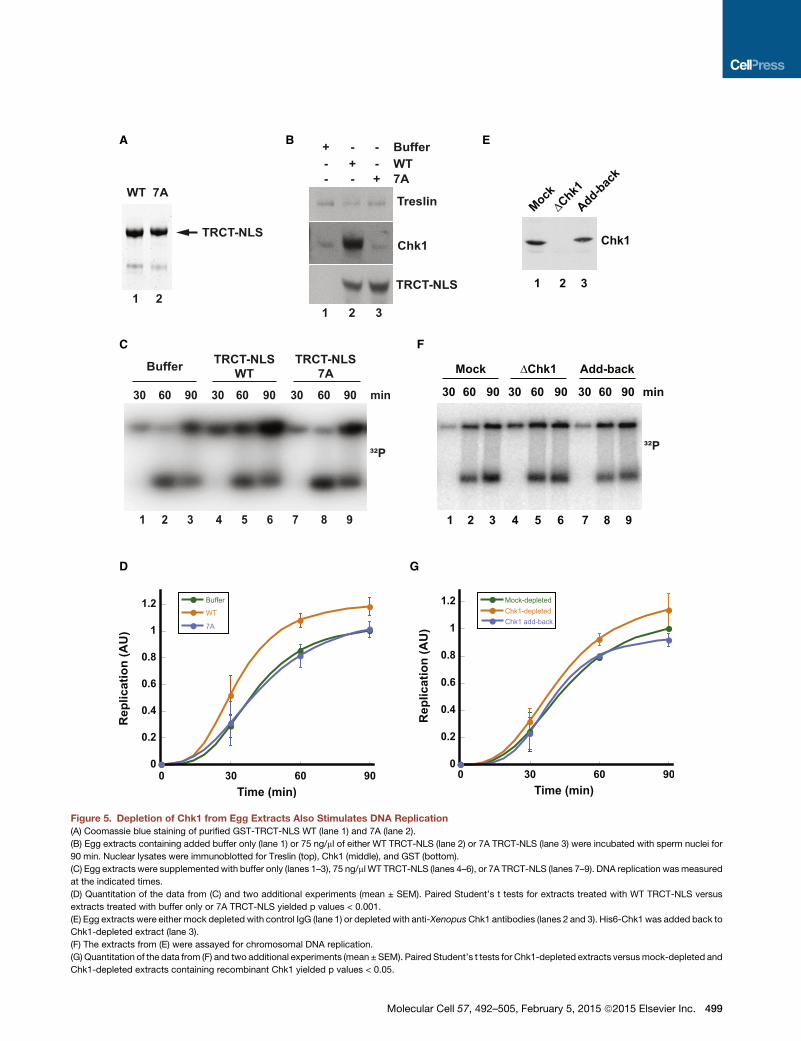
A B E
C F
D G
Figure 5. Depletion of Chk1 from Egg Extracts Also Stimulates DNA Replication
(A) Coomassie blue staining of purified GST-TRCT-NLS WT (lane 1) and 7A (lane 2).
(B) Egg extracts containing added buffer only (lane 1) or 75 ng/ml of either WT TRCT-NLS (lane 2) or 7A TRCT-NLS (lane 3) were incubated with sperm nuclei for
90 min. Nuclear lysates were immunoblotted for Treslin (top), Chk1 (middle), and GST (bottom).
(C) Egg extracts were supplemented with buffer only (lanes 1–3), 75 ng/ml WT TRCT-NLS (lanes 4–6), or 7A TRCT-NLS (lanes 7–9). DNA replication wasmeasured
at the indicated times.
(D) Quantitation of the data from (C) and two additional experiments (mean ± SEM). Paired Student’s t tests for extracts treated with WT TRCT-NLS versus
extracts treated with buffer only or 7A TRCT-NLS yielded p values < 0.001.
(E) Egg extracts were either mock depleted with control IgG (lane 1) or depleted with anti-XenopusChk1 antibodies (lanes 2 and 3). His6-Chk1 was added back to
Chk1-depleted extract (lane 3).
(F) The extracts from (E) were assayed for chromosomal DNA replication.
(G) Quantitation of the data from (F) and two additional experiments (mean ± SEM). Paired Student’s t tests for Chk1-depleted extracts versusmock-depleted and
Chk1-depleted extracts containing recombinant Chk1 yielded p values < 0.05.
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 499
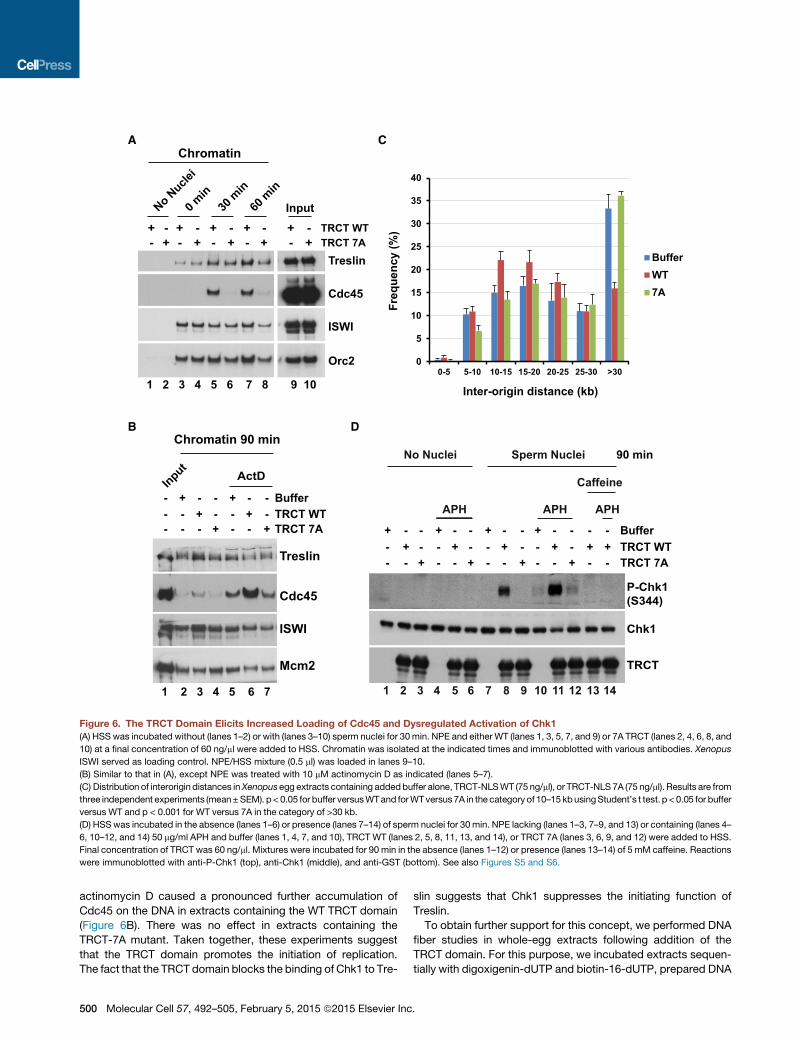
actinomycin D caused a pronounced further accumulation of
Cdc45 on the DNA in extracts containing the WT TRCT domain
(Figure 6B). There was no effect in extracts containing the
TRCT-7A mutant. Taken together, these experiments suggest
that the TRCT domain promotes the initiation of replication.
The fact that the TRCT domain blocks the binding of Chk1 to Tre-
slin suggests that Chk1 suppresses the initiating function of
Treslin.
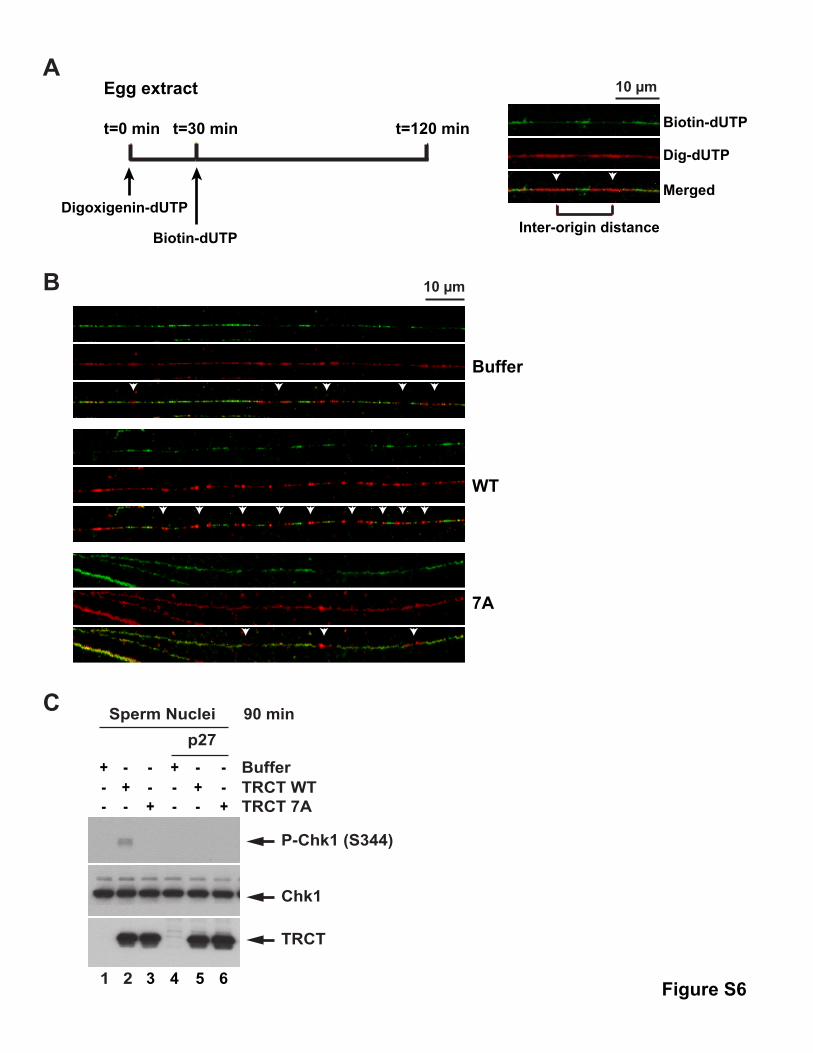
To obtain further support for this concept, we performed DNA
fiber studies in whole-egg extracts following addition of the
TRCT domain. For this purpose, we incubated extracts sequen-
tially with digoxigenin-dUTP and biotin-16-dUTP, prepared DNA
- - + - - + - - + - - +- + - - + - - + - - + -
-+ - - + - - + - - + -
- -+ +- -
TRCT
Chk1
P-Chk1 (S344)
D
90 min
Caffeine
TRCT WTTRCT 7A
Buffer
No Nuclei Sperm Nuclei
APH APH APH
1 2 3 4 5 6 7 8 9 10 11 12 13 14
B
TRCT WTTRCT 7A
--
+- +
--- +
- +-
Buffer+ - - + - -
---
ISWI
Cdc45
Treslin
Mcm2
ActD
Chromatin 90 min
1 2 3 4 5 6 7
Input
TRCT WTTRCT 7A
+- +
- +- +
- +- +
- +- +
-No N
uclei
0 min
30 m
in
60 m
in
+- +
Input
Orc2
ISWI
Cdc45
Treslin
A
-
1 2 3 4 5 6 7 8 9 10
Chromatin
Freq
uenc
y (%
)Inter-origin distance (kb)
C
0
5
10
15
20
25
30
35
40
0-5 5-10 10-15 15-20 20-25 25-30 >30
BufferWT7A
Figure 6. The TRCT Domain Elicits Increased Loading of Cdc45 and Dysregulated Activation of Chk1
(A) HSS was incubated without (lanes 1–2) or with (lanes 3–10) sperm nuclei for 30 min. NPE and either WT (lanes 1, 3, 5, 7, and 9) or 7A TRCT (lanes 2, 4, 6, 8, and
10) at a final concentration of 60 ng/ml were added to HSS. Chromatin was isolated at the indicated times and immunoblotted with various antibodies. Xenopus
ISWI served as loading control. NPE/HSS mixture (0.5 ml) was loaded in lanes 9–10.
(B) Similar to that in (A), except NPE was treated with 10 mM actinomycin D as indicated (lanes 5–7).
(C) Distribution of interorigin distances inXenopusegg extracts containing addedbuffer alone, TRCT-NLSWT (75 ng/ml), or TRCT-NLS 7A (75 ng/ml). Results are from
three independent experiments (mean±SEM). p< 0.05 for buffer versusWTand forWTversus7A in the category of 10–15kbusingStudent’s t test. p <0.05 for buffer
versus WT and p < 0.001 for WT versus 7A in the category of >30 kb.
(D) HSSwas incubated in the absence (lanes 1–6) or presence (lanes 7–14) of sperm nuclei for 30 min. NPE lacking (lanes 1–3, 7–9, and 13) or containing (lanes 4–
6, 10–12, and 14) 50 mg/ml APH and buffer (lanes 1, 4, 7, and 10), TRCT WT (lanes 2, 5, 8, 11, 13, and 14), or TRCT 7A (lanes 3, 6, 9, and 12) were added to HSS.
Final concentration of TRCT was 60 ng/ml. Mixtures were incubated for 90 min in the absence (lanes 1–12) or presence (lanes 13–14) of 5 mM caffeine. Reactions
were immunoblotted with anti-P-Chk1 (top), anti-Chk1 (middle), and anti-GST (bottom). See also Figures S5 and S6.
500 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.

fibers from nuclear fractions, and then measured interorigin
distances (Bellelli et al., 2014; Marheineke et al., 2009). We
observed that the WT TRCT fragment caused a significant in-
crease in shorter interorigin distances (e.g., 10–15 kb) relative
to incubations containing added buffer alone or the 7A fragment
(Figures 6C, S6A, and S6B). Concomitantly, there was a
decrease in larger interorigin distances (e.g., >30 kb) in incuba-
tions containing the WT fragment. These findings directly
support an increase in the firing of origins, which fits well with
the observations that the TRCT domain enhances formation of
the activated replicative helicase.
Dysregulated Origin Firing in the Absence of the Treslin-Chk1 Interaction Induces Activation of Chk1The elevated loading of Cdc45 onto chromatin and the ensuing
increase in replication in the presence of the TRCT domain raised
the possibility that this fragment might elicit replication stress.
To address this issue, we examined phosphorylation of Xenopus
Chk1 on S344 (Kumagai et al., 1998). We observed that the WT
TRCT domain induced phosphorylation of Chk1 in NPE fractions
containing sperm chromatin even in the absence of APH (Fig-
ure 6D). The TRCT domain also caused a substantial further in-
crease in phosphorylation of Chk1 in extracts containing APH.
By contrast, there was no increase in extracts treated with con-
trol buffer or the 7A mutant TRCT domain. Furthermore, the
increase in the presence of the WT TRCT domain was abolished
by caffeine, an inhibitor of the ATR-catalyzed phosphorylation of
Chk1. Therefore, the TRCT-stimulated activation of Chk1 in-
volves the ATR-mediated pathway. This increase also depended
on the presence of sperm chromatin as a template for replica-
tion, which suggests that the TRCT domain does not somehow
directly activate Chk1 by a DNA-independent mechanism. In
further support of this concept, we found that inhibition of DNA
replication with p27 abolished the TRCT-stimulated increase
in the phosphorylation of Chk1 in extracts lacking APH
(Figure S6C). Overall, these experiments indicate that the iso-
lated TRCT domain causes a pronounced derangement of
DNA replication in the Xenopus egg extract system.
Expression of the Treslin-7A Mutant in Human CellsLeads to Increased Origin FiringAs another strategy to examine the function of the Treslin-Chk1
interaction, we utilized human cells that overexpress the Treslin-
7Amutant. In particular, we employed U2OS T-REx cells that ex-
press WT and 7A Treslin in a doxycycline-inducible manner. We
induced the expression of Treslin in these cells by addition of
doxycycline and later examined origin firing by DNA fiber anal-
ysis (Jackson and Pombo, 1998; Schlacher et al., 2011). For
these studies, we incubated the cells with CldU for 20 min and
then with IdU for 20 min (Figure 7A). Next, we prepared DNA
fibers from the cells by standard methods and detected incorpo-
ration of the modified nucleotides with fluorescently tagged
antibodies that detect CldU (green tracks) and IdU (red tracks).
Finally, we quantitated the frequency of singly labeled IdU (red)
tracks as a measure of new origin firing during the second label-
ing period. We noted that there was a significant increase in
origin firing in cells expressing the full-length Treslin-7A mutant
in comparison with cells expressing the WT protein (Figures
7B–7D). The induced levels of theWT and 7A proteins were quite
similar in the different cell lines. Moreover, overexpression of WT
Treslin did not have a significant effect on origin firing. We also
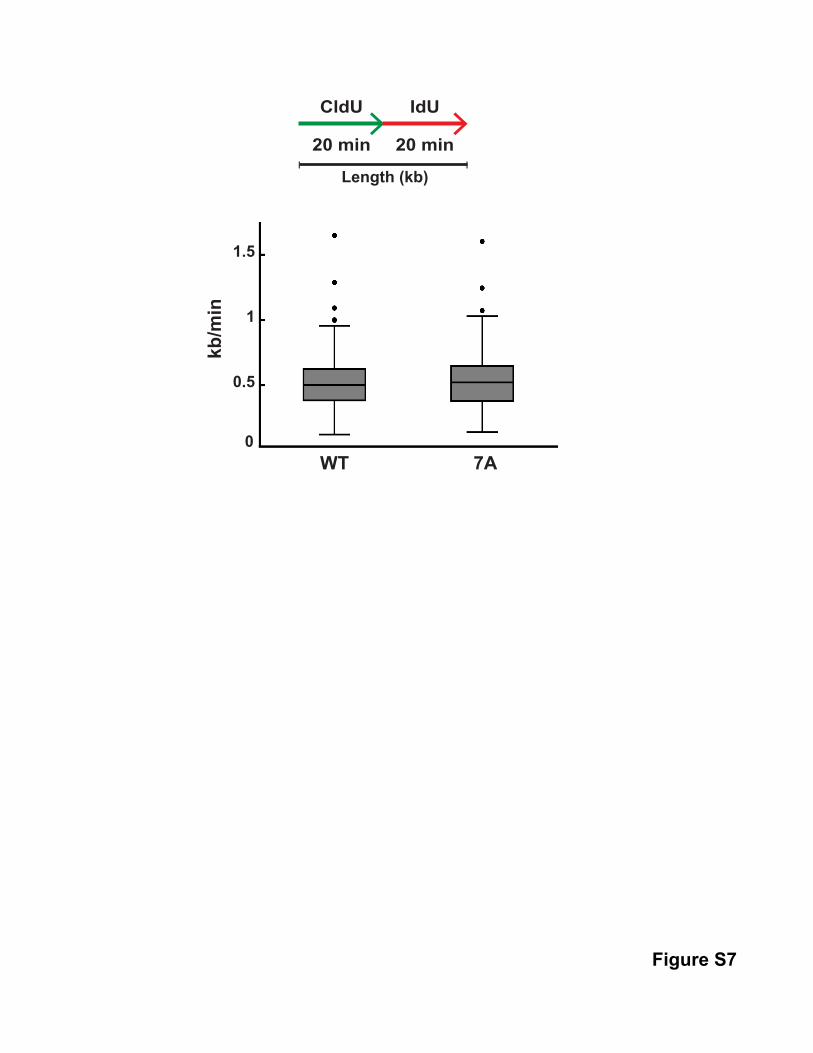
examined replication fork speed in these cells but could not
discern a difference between cells expressing WT or 7A Treslin
(Figure S7). Overall, these experiments further support our find-
ings with Xenopus egg extracts that disruption of the Treslin-
Chk1 interaction leads to increased origin firing. The fact that
the Treslin-Chk1 interaction is functionally important in both Xen-
opus and humans suggests that this regulatory mechanism is a
conserved feature of DNA replication in vertebrates.
DISCUSSION
In this report, we have identified an interaction between Treslin
and the key checkpoint-effector kinase Chk1. The high speci-
ficity of this binding suggested that there is a significant regula-
tory relationship between these two proteins. Indeed, we have
found that Chk1 negatively regulates the replication-initiating
function of Treslin (see Figure 7E). To reach this conclusion, we
utilized both Xenopus egg extracts and human cells. In one
approach, we added the TRCT domain from Treslin to Xenopus
egg extracts as a competitor of the Treslin-Chk1 interaction.
Strikingly, this peptide elicited a pronounced increase in DNA
replication.
In searching for the basis of this phenomenon, we found that
the TRCT fragment strongly stimulated the binding of Cdc45 to
chromatin. The loading of Cdc45 onto chromatin in egg extracts
is rate limiting for origin firing (Mimura et al., 2000; Walter and
Newport, 2000). Thus, the TRCT domain appears to act by
enhancing initiation. To provide further evidence, we have
dissected this process with actinomycin D, a drug that blocks
replication in egg extracts just after initiation but before signifi-
cant unwinding of the DNA (Pacek and Walter, 2004). Treatment
with both the TRCT domain and actinomycin D led to a dramatic
and sustained accumulation of Cdc45 on chromatin. Finally, we
utilized DNA fiber studies to show that the addition of the TRCT
domain to egg extracts leads to an overall decrease in interorigin
distances in replicating chromatin.
In a complementary approach, we also employed human cells.
In particular, we also usedDNA fiber analysis to show that human
cells overexpressing the full-length Treslin-7A mutant display
increasedorigin firingduringanunperturbedSphase. This obser-
vation is consistent with the fact that human cells with compro-
mised function of Chk1 display elevated firing of replication
origins during a normal cell cycle (Maya-Mendoza et al., 2007;
McIntosh and Blow, 2012; Miao et al., 2003; Syljuasen et al.,
2005). Hence, our studies have revealed a mechanism by which
Chk1 controls replication in cells without overtly damaged
DNA. It has also been reported that Chk1-deficient cells display
decreased replication fork speed (see Petermann et al., 2010).
We have not observed anobvious effect on fork speed in cells ex-
pressing the Treslin-7A mutant. This mutant may not cause
enough origin firing to disrupt fork progression. Moreover, Chk1
could have distinct targets for initiation versus elongation.
Utilization of replication origins places a number of regulatory
demands on cells. The firing of origins depends upon positive
regulation of Treslin by S-CDK, but there may also have to be
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 501
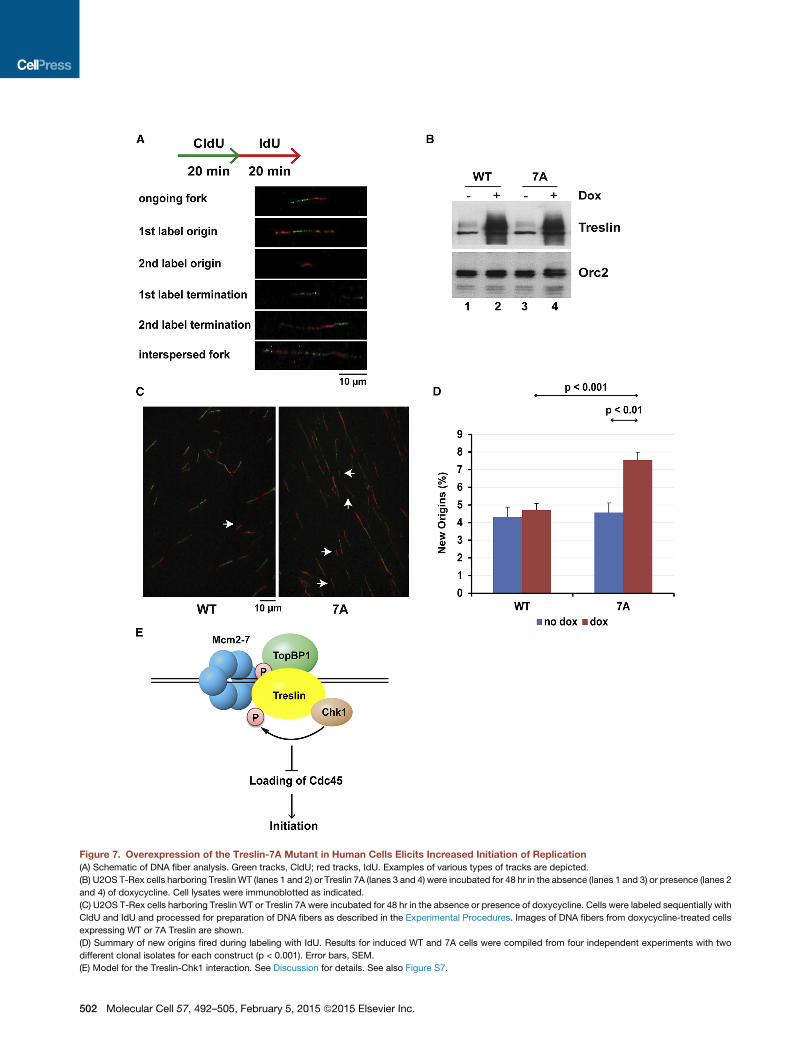
Figure 7. Overexpression of the Treslin-7A Mutant in Human Cells Elicits Increased Initiation of Replication
(A) Schematic of DNA fiber analysis. Green tracks, CldU; red tracks, IdU. Examples of various types of tracks are depicted.
(B) U2OS T-Rex cells harboring Treslin WT (lanes 1 and 2) or Treslin 7A (lanes 3 and 4) were incubated for 48 hr in the absence (lanes 1 and 3) or presence (lanes 2
and 4) of doxycycline. Cell lysates were immunoblotted as indicated.
(C) U2OS T-Rex cells harboring Treslin WT or Treslin 7A were incubated for 48 hr in the absence or presence of doxycycline. Cells were labeled sequentially with
CldU and IdU and processed for preparation of DNA fibers as described in the Experimental Procedures. Images of DNA fibers from doxycycline-treated cells
expressing WT or 7A Treslin are shown.
(D) Summary of new origins fired during labeling with IdU. Results for induced WT and 7A cells were compiled from four independent experiments with two
different clonal isolates for each construct (p < 0.001). Error bars, SEM.
(E) Model for the Treslin-Chk1 interaction. See Discussion for details. See also Figure S7.
502 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.

negative regulatorymechanisms that suppresspremature or inap-
propriate action of Treslin at origins. The ability of Chk1 to inhibit
the Treslin-mediated loading of Cdc45 would provide such a
mechanism. This process could operate generally at all origins or
come into play at a subset of origins under certain circumstances.
Typically, the number of loaded MCM complexes on chro-
matin at the onset of S phase greatly exceeds the number of or-
igins that actually fire in a given cell cycle. This ‘‘MCM paradox’’
has suggested that there are a large number of dormant origins in
the genome (see McIntosh and Blow, 2012). It has been pro-
posed that utilization of these dormant origins would allow cells
to cope with replication stress by increasing the likelihood that
replication in stressed areas would reach completion. The ques-
tion arises, however, about how the cell would regulate the firing
of such dormant origins under both unstressed and stressed
conditions. It has been postulated that these origins might fire
stochastically such that passive replication from nearby origins
would typically occur under unstressed conditions. However,
under stressed conditions of slowed or blocked replication, there
would be more time for the dormant origins to undergo firing.
Another type of explanation would incorporate the existence
of an inhibitory mechanism that suppresses firing of dormant or-
igins. In principle, inhibition of Treslin-mediated loading of
Cdc45 by basally active Chk1 under unstressed conditions
could correspond to such a mechanism. Our DNA fiber studies
with egg extracts do not suggest that there is rampant firing of
dormant origins upon addition of the TRCT domain. However,
more limited firing of dormant origins may contribute to the
accelerated replication that we have observed under these
conditions.
The Treslin-Chk1 interactionmight also help to explain the tem-
poral order in firing of origins during S phase in somatic cells. In
eukaryotic cells, there is typically a pattern wherein ‘‘early’’ and
‘‘late’’ origins at distinct chromosomal regions fire at different
times (McIntosh andBlow, 2012; Siddiqui et al., 2013). In budding
yeast, regulation of the Treslin homolog Sld3 has a role in the
distinction between early and late origins (Lopez-Mosqueda
et al., 2010; Zegerman and Diffley, 2010). However, the mecha-
nisms that regulate suchdifferential firing invertebrates are largely
unknown.Anotherpotential explanation for the inhibitionof Treslin
by Chk1 is that this process could be necessary for the suppres-
sion of later-firing origins during the earlier parts of S phase.
Further studieswill be required to identify thecharacteristics of or-
igins that are subject to Chk1-dependent inhibition of Treslin.
It is well accepted that the ATR-dependent activation of Chk1
in response to incompletely replicated and damaged DNA leads
to inhibition of origin firing at chromosomal regions that are not
already engaged in replication (McIntosh and Blow, 2012; Søren-
sen and Syljuasen, 2012; Yekezare et al., 2013). The activation of
Chk1 results in decreased function of Cdc25 and increased func-
tion of Wee1 (Perry and Kornbluth, 2007). In turn, these effects
promote the inhibitory phosphorylation of Cdk2 (as well as
Cdk1) on T14 and Y15. The loading of Cdc45 onto chromatin
requires phosphorylation of Treslin by Cdk2, which mediates
binding of Treslin to TopBP1 (Boos et al., 2011; Kumagai et al.,
2011). Therefore, it might be anticipated that reduced activity
of Cdk2 would compromise formation of the Treslin-TopBP1
complex. Indeed, treatment of human cells with hydroxyurea re-
duces the binding of Treslin to TopBP1, and the Chk1 inhibitor
AZD7762 reverses this effect (Boos et al., 2011).
We have not been able to detect an override of the hydroxy-
urea-induced block to replication in U2OS cells expressing the
Treslin-7A mutant (not shown). This mutation would presumably
not affect the ability of Chk1 to downregulate Cdk2. We have
also observed that both the WT and 7A forms of Treslin bind
equally well to TopBP1. Moreover, the Treslin-7A mutant dis-
plays normally regulated binding to TopBP1 (i.e., decreased
binding in the presence of hydroxyurea) (not shown). Finally,
the TRCT domain does not affect binding of Treslin to TopBP1
in Xenopus NPE fractions. It would be interesting to examine
the effect of the Treslin-7A mutant in replication-stressed cells
in which the inhibition of Cdk2 had been overridden.
Although inhibition of Cdk2 could account for the effect of
Chk1 on DNA replication in stressed cells, it is unclear that this
mechanism could explain the role of Chk1 in unstressed cells.
The activity of Cdk2 rises substantially at S phase in
unperturbed cells. It is possible that there could be some local
inhibition of Cdk2 in certain regions of the genome, but the
mechanistic basis for such an effect is unknown. Another expla-
nation is that Chk1 could control replication in some other
manner besides regulation of Cdk2 under conditions where there
is not a strong, exogenous threat to the DNA. The regulation of
Treslin by Chk1 may be such a mechanism. It should also be
noted that our studies do not rule out the possibility that Chk1
could have an additional target(s) besides Treslin during an un-
perturbed S phase. Overall, inhibition of Treslin by Chk1 could
suppress relevant origins during an unperturbed S phase and
perhaps even supplement the inhibition of Cdk2 in replication-
stressed cells.
A previous study in the Xenopus system indicated that deple-
tion of Chk1 did not increase DNA replication in either untreated
or APH-treated egg extracts (Luciani et al., 2004). For the
untreated extracts, these investigators quantitated DNA replica-
tion at a single late time point (120 min) when replication would
have reached completion. Hence, this analysis would not have
detected the early acceleration of replication that we have
observed in this study. Another group found that the Chk1 inhib-
itor UCN-01 did not increase replication in egg extracts, but
actually inhibited replication to some extent (Murphy and
Michael, 2013). However, UCN-01 also inhibits Cdk2 at the con-
centrations used in this study (Kawakami et al., 1996), which
complicates interpretation of the results.
In principle, phosphorylation of Treslin by Chk1 may alter its
conformation or directly affect its interactions with other pro-
teins to preclude helicase activation. Moreover, the physical
association of Chk1 might also have such effects on Treslin.
Regulation by Chk1 appears not to reduce binding of Treslin
to TopBP1, but Chk1 may nonetheless alter the helicase-acti-
vating properties of this complex. Chk1 may also inhibit the
ability of Treslin and TopBP1 to recognize Cdc45/GINS effec-
tively or deliver Cdc45/GINS to the MCM complex. Finally,
Chk1 may influence the interaction of Treslin with some other
component(s) of the replication apparatus. We will need to
understand better the exact molecular mechanism by which
Treslin promotes helicase activation in order to evaluate which,
if any, of these possibilities is correct.
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 503

In conclusion, we have used both the Xenopus and human
systems to provide perspectives on the function of Chk1 and
the regulation of early steps in DNA replication. Further analysis
of this process may yield additional insights into how vertebrate
cells ensure the faithful propagation of their genomes.
EXPERIMENTAL PROCEDURES
Plasmids
The cDNA for human Treslin (GenBank ADC30133.1) was described before (Ku-
magai et al., 2010). pcDNA5/TO-Treslin-SF (encoding the SV40 NLS, an S-pep-
tide tag, and a 3X-FLAG tag at the C-terminal end of Treslin) was created from
pcDNA5/TO-Treslin-Myc by usingPCR to exchange tags (Kumagai et al., 2011).
Human Tissue Culture Cells
293T and U2OS cells were cultured in Dulbecco’s modified Eagle’s medium
(DMEM) containing 10% fetal bovine serum. 293T cells were transfected using
FuGENE6. Large-scale transfections as well as preparation of nuclear lysates
are described in the Supplemental Experimental Procedures.
Antibodies
Anti-Treslin antibodies were previously described (Kumagai et al., 2010). For
other antibodies, see the Supplemental Experimental Procedures.
Mass Spectrometry Analysis
Plasmids encoding different versions of Treslin-SF were transfected into 293T
cells. Tagged proteins were reisolated and analyzed by mass spectrometry as
described in the Supplemental Experimental Procedures. Binding of Chk1 to
full-length Treslin was established bymatching 9 unique peptides that covered
24% of the sequence of Chk1. For binding to the 1,253–1,909 C-terminal frag-
ment, we detected 10 unique peptides also covering 24% of the sequence.
There was no binding of Chk1 to the 1–1,257 N-terminal fragment.
Recombinant GST Fusion Proteins
To produce GST and GST-NLS fusion proteins containing versions of the hu-
man TRCT, appropriate DNAwas amplified by PCR and cloned into pGEX4T-3
or pGEX-NLS, respectively. Proteins were produced as described in the Sup-
plemental Experimental Procedures.
Production of U2OS Cell Lines Expressing Treslin and Its Mutants
U2OS T-REx cells were maintained, transfected with pcDNA5/TO encoding
siRNA-resistant Treslin, and selected as described (Kumagai et al., 2011). Sin-
gle colonies that expressed the desired protein upon addition of doxycycline
were isolated for analysis.
siRNA Experiments
Stealth siRNA specific for Treslin (#2, AGGACACAUUCUGCCUCCUUCUAUU)
and control siRNA (low GC) were obtained from Invitrogen. The siRNA (30 nM)
was transfected into U2OS cells with Lipofectamine RNAiMAX (Invitrogen). For
rescue experiments, pcDNA5/TO-Treslin-SF was rendered resistant to siRNA
#2. We obtained more efficient rescue by using siRNA #2 instead of the previ-
ous siRNA #1 (Kumagai et al., 2010).
Xenopus Egg Extracts
Whole extracts from Xenopus eggs were prepared as before (Kumagai et al.,
2010). Preparation of nuclear lysates, immunodepletion of Chk1, and DNA
replication assays were also described previously (Kumagai et al., 1998,
2010). Further details are provided in the Supplemental Experimental
Procedures.
Xenopus NPE System
Nucleoplasmic extract (NPE) and high-speed supernatant (HSS) from Xenopus
eggs were prepared as described (Lebofsky et al., 2009). Details on the use of
these extracts for assays of DNA replication and binding to chromatin are
described in the Supplemental Experimental Procedures.
DNA Fiber Assays
Xenopus egg extracts were incubated sequentially with digoxigenin-11-dUTP
and biotin-16-dUTP. Labeled DNA fibers were prepared as described (Bellelli
et al., 2014; Marheineke et al., 2009). Human U2OS cells were labeled with
40 mMCldU, washed with phosphate-buffered saline (PBS), and finally labeled
with 50 mM IdU as indicated in the figures. Preparation of DNA fiber spreads
was reported previously (Jackson and Pombo, 1998; Schlacher et al., 2011).
Further details on these protocols are provided in the Supplemental Experi-
mental Procedures.
SUPPLEMENTAL INFORMATION
Supplemental Information includes Supplemental Experimental Procedures,
seven figures, and one table and can be found with this article online at
http://dx.doi.org/10.1016/j.molcel.2014.12.003.
ACKNOWLEDGMENTS
We are grateful to laboratory members for comments on the manuscript. We
also thank Juan Ramırez-Lugo for anti-ISWI antibodies. This work was sup-
ported by NIH grants GM043974 and GM070891 to W.G.D.
Received: July 15, 2014
Revised: October 16, 2014
Accepted: November 24, 2014
Published: December 31, 2014
REFERENCES
Bellelli, R., Castellone, M.D., Guida, T., Limongello, R., Dathan, N.A., Merolla,
F., Cirafici, A.M., Affuso, A., Masai, H., Costanzo, V., et al. (2014). NCOA4 tran-
scriptional coactivator inhibits activation of DNA replication origins. Mol. Cell
55, 123–137.
Boos, D., Sanchez-Pulido, L., Rappas, M., Pearl, L.H., Oliver, A.W., Ponting,
C.P., and Diffley, J.F. (2011). Regulation of DNA replication through Sld3-
Dpb11 interaction is conserved fromyeast tohumans.Curr. Biol.21, 1152–1157.
Jackson, D.A., and Pombo, A. (1998). Replicon clusters are stable units of
chromosome structure: evidence that nuclear organization contributes to the
efficient activation and propagation of S phase in human cells. J. Cell Biol.
140, 1285–1295.
Kawakami, K., Futami, H., Takahara, J., and Yamaguchi, K. (1996). UCN-01,
7-hydroxyl-staurosporine, inhibits kinase activity of cyclin-dependent kinases
and reduces the phosphorylation of the retinoblastoma susceptibility gene
product in A549 human lung cancer cell line. Biochem. Biophys. Res.
Commun. 219, 778–783.
Kumagai, A., Guo, Z., Emami, K.H., Wang, S.X., and Dunphy, W.G. (1998). The
XenopusChk1 protein kinasemediates a caffeine-sensitive pathway of check-
point control in cell-free extracts. J. Cell Biol. 142, 1559–1569.
Kumagai, A., Shevchenko, A., Shevchenko, A., and Dunphy, W.G. (2010).
Treslin collaborates with TopBP1 in triggering the initiation of DNA replication.
Cell 140, 349–359.
Kumagai, A., Shevchenko, A., Shevchenko, A., and Dunphy, W.G. (2011).
Direct regulation of Treslin by cyclin-dependent kinase is essential for the
onset of DNA replication. J. Cell Biol. 193, 995–1007.
Labib, K. (2010). How do Cdc7 and cyclin-dependent kinases trigger the initia-
tion of chromosome replication in eukaryotic cells? Genes Dev. 24, 1208–1219.
Lebofsky, R., Takahashi, T., and Walter, J.C. (2009). DNA replication in nu-
cleus-free Xenopus egg extracts. Methods Mol. Biol. 521, 229–252.
Liu, Q., Guntuku, S., Cui, X.S., Matsuoka, S., Cortez, D., Tamai, K., Luo, G.,
Carattini-Rivera, S., DeMayo, F., Bradley, A., et al. (2000). Chk1 is an essential
kinase that is regulated by Atr and required for the G(2)/MDNA damage check-
point. Genes Dev. 14, 1448–1459.
Lopez-Mosqueda, J., Maas, N.L., Jonsson, Z.O., Defazio-Eli, L.G.,
Wohlschlegel, J., and Toczyski, D.P. (2010). Damage-induced phosphoryla-
tion of Sld3 is important to block late origin firing. Nature 467, 479–483.
504 Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc.

Luciani, M.G., Oehlmann, M., and Blow, J.J. (2004). Characterization of a novel
ATR-dependent, Chk1-independent, intra-S-phase checkpoint that sup-
presses initiation of replication in Xenopus. J. Cell Sci. 117, 6019–6030.
Marheineke, K., Goldar, A., Krude, T., andHyrien, O. (2009). Use of DNA comb-
ing to study DNA replication in Xenopus and human cell-free systems.
Methods Mol. Biol. 521, 575–603.
Maya-Mendoza, A., Petermann, E., Gillespie, D.A., Caldecott, K.W., and
Jackson, D.A. (2007). Chk1 regulates the density of active replication origins
during the vertebrate S phase. EMBO J. 26, 2719–2731.
McIntosh, D., and Blow, J.J. (2012). Dormant origins, the licensing checkpoint,
and the response to replicative stresses. Cold Spring Harb. Perspect. Biol. 4,
a012955.
Miao, H., Seiler, J.A., and Burhans, W.C. (2003). Regulation of cellular and
SV40 virus origins of replication by Chk1-dependent intrinsic and UVC radia-
tion-induced checkpoints. J. Biol. Chem. 278, 4295–4304.
Mimura, S., Masuda, T., Matsui, T., and Takisawa, H. (2000). Central role for
cdc45 in establishing an initiation complex of DNA replication in Xenopus
egg extracts. Genes Cells 5, 439–452.
Murphy, C.M., andMichael, W.M. (2013). Control of DNA replication by the nu-
cleus/cytoplasm ratio in Xenopus. J. Biol. Chem. 288, 29382–29393.
Pacek, M., and Walter, J.C. (2004). A requirement for MCM7 and Cdc45 in
chromosome unwinding during eukaryotic DNA replication. EMBO J. 23,
3667–3676.
Perry, J.A., and Kornbluth, S. (2007). Cdc25 and Wee1: analogous opposites?
Cell Div. 2, 12.
Petermann, E., Woodcock, M., and Helleday, T. (2010). Chk1 promotes repli-
cation fork progression by controlling replication initiation. Proc. Natl. Acad.
Sci. USA 107, 16090–16095.
Sansam, C.L., Cruz, N.M., Danielian, P.S., Amsterdam, A., Lau, M.L., Hopkins,
N., and Lees, J.A. (2010). A vertebrate gene, ticrr, is an essential checkpoint
and replication regulator. Genes Dev. 24, 183–194.
Schlacher, K., Christ, N., Siaud, N., Egashira, A., Wu, H., and Jasin, M. (2011).
Double-strand break repair-independent role for BRCA2 in blocking stalled
replication fork degradation by MRE11. Cell 145, 529–542.
Sclafani, R.A., and Holzen, T.M. (2007). Cell cycle regulation of DNA replica-
tion. Annu. Rev. Genet. 41, 237–280.
Siddiqui, K., On, K.F., and Diffley, J.F. (2013). Regulating DNA replication in eu-
karya. Cold Spring Harb. Perspect. Biol. 5, a012930.
Sørensen, C.S., and Syljuasen, R.G. (2012). Safeguarding genome integrity:
the checkpoint kinases ATR, CHK1 and WEE1 restrain CDK activity during
normal DNA replication. Nucleic Acids Res. 40, 477–486.
Syljuasen, R.G., Sørensen, C.S., Hansen, L.T., Fugger, K., Lundin, C.,
Johansson, F., Helleday, T., Sehested, M., Lukas, J., and Bartek, J. (2005).
Inhibition of human Chk1 causes increased initiation of DNA replication, phos-
phorylation of ATR targets, and DNA breakage. Mol. Cell. Biol. 25, 3553–3562.
Tanaka, S., and Araki, H. (2013). Helicase activation and establishment of repli-
cation forks at chromosomal origins of replication. Cold Spring Harb.
Perspect. Biol. 5, a010371.
Toledo, L.I., Murga, M., and Fernandez-Capetillo, O. (2011). Targeting ATR
and Chk1 kinases for cancer treatment: a new model for new (and old) drugs.
Mol. Oncol. 5, 368–373.
Ullah, Z., de Renty, C., and DePamphilis, M.L. (2011). Checkpoint kinase 1 pre-
vents cell cycle exit linked to terminal cell differentiation. Mol. Cell. Biol. 31,
4129–4143.
Walter, J., and Newport, J. (2000). Initiation of eukaryotic DNA replication:
origin unwinding and sequential chromatin association of Cdc45, RPA, and
DNA polymerase alpha. Mol. Cell 5, 617–627.
Yekezare, M., Gomez-Gonzalez, B., and Diffley, J.F. (2013). Controlling DNA
replication origins in response to DNA damage - inhibit globally, activate
locally. J. Cell Sci. 126, 1297–1306.
Zachos, G., Black, E.J., Walker, M., Scott, M.T., Vagnarelli, P., Earnshaw,
W.C., and Gillespie, D.A. (2007). Chk1 is required for spindle checkpoint func-
tion. Dev. Cell 12, 247–260.
Zegerman, P., and Diffley, J.F. (2010). Checkpoint-dependent inhibition of
DNA replication initiation by Sld3 and Dbf4 phosphorylation. Nature 467,
474–478.
Molecular Cell 57, 492–505, February 5, 2015 ª2015 Elsevier Inc. 505

Molecular Cell, Volume 57
Supplemental Information
Interaction of Chk1 with Treslin Negatively Regulates the Initiation of Chromosomal DNA
Replication
Cai Guo, Akiko Kumagai, Katharina Schlacher, Anna Shevchenko, Andrej Shevchenko, and William G.
Dunphy

Control IP
InputChk1
IP
Control IP
Chk1 IP
Control IP
Chk1 IP
-- +- + -
+- + -
---- EtBr
Benzonase
Treslin
Chk1
1 2 3 4 5 6 7
Figure S1
10% In
put
GST18
10-19
09
S1887
A
S1893
A
T1897
A
7A 1810
-1872
1870
-1909
Chk1
CBB
1 2 3 4 5 6 7 8 9
GST pulldown
Figure S2
Chk1
GST pulldown
Chk1 WT Chk1 KD
GSTGST
TRCT
TRCT
S1887
A
S1887
A
S1893
A
S1893
A
T1897
A
T1897
A
S1893
A-T1987
A
+- +
- +- +
- +- +
- +- +
- +- +
- +- +
-S18
93A-T19
87A
³²P
TRCT
GST
TRCT
GST
CBB
1 2 3 4
1 2 3 4 5 6 7 8 9 10 11 12
GSTTRCT
S1893
A-T1897
A
5% In
put
CBB
A
B
GST
TRCT
Chk1 WT Chk1 KD
GSTTRCT + G
ST
TRCT + GST
+-
+- +
- +- +
- +- +
-
Cdc25 +
GST
Cdc25 +
GST
TRCT + Cdc2
5
TRCT + Cdc2
5
³²PTRCT Cdc25(254-316)
GST
TRCT Cdc25(254-316)
Chk1
CBB
1 2 3 4 5 6 7
Relative p-TRCT Relative p-Cdc25
1.9 0.5 1.8 0.51 0.4 0.8 0.5
C
Figure S3
Control IP
Chk1 IP
NPE
1 2 3 4 5
Benzo
nase
EtBr
Treslin
Chk1
None
A
30 60 90 30 60 90 min
1 2 3 4 5 6
Buffer AZD7762
B
³²P
Figure S4
Treslin
TopBP1
Pol ε
Cdc45
Sld5
PCNA
Orc2
TRCT WTTRCT 7A
Input No Nucle
i
0 min30 min 60 min
+ -- +
+ -- +
+ -- +
+ -- +
--
1 2 3 4 5 6 7 8 9
Figure S5
t=0 min t=30 min t=120 min
Digoxigenin-dUTP
Biotin-dUTP
Egg extract
10 µm
Buffer
WT
7A
Biotin-dUTP
Dig-dUTP
Merged
Inter-origin distance
10 µmA
B
Figure S6
TRCT
Chk1
P-Chk1 (S344)
p27Sperm Nuclei 90 min
1
BufferTRCT WTTRCT 7A+- - +- -
+ +- - - -+- - - + -
2 3 4 5 6
C
WT 7A
1
0
1.5
kb/m
in
0.5
CldU IdU
20 min 20 min
Length (kb)
Figure S7

Figure S1. Treatment with Ethidium Bromide or Benzonase Does Not Inhibit Co-immunoprecipitation of Treslin and Chk1. Related to Figure 1. 293T nuclear lysates were treated with 50 µg/ml ethidium bromide (lanes 4 and 5) or 1 U/µl Benzonase (lanes 6 and 7) for 30 min on ice before immunoprecipitation was performed as in Figure 1C using control IgG (lanes 2, 4, and 6) or antibodies against Chk1 (lanes 3, 5, and 7). Figure S2. The TRCT Domain Binds Directly to Chk1. Related to Figure 2. GST only (lane 2), GST-tagged TRCT (lane 3), TRCT containing mutations S1887A (lane 4), S1893A (lane 5), T1897A (lane 6), or 7A (lane 7), and GST-tagged forms of residues 1810-1872 (lane 8) and 1870-1909 from Treslin (lane 9) were isolated from bacteria with glutathione agarose. Bead-bound proteins were incubated with recombinant His6-Chk1 in binding buffer at 4ºC for 90 min. Beads were retrieved, washed, and subjected to immunoblotting with anti-Chk1 (top) and staining with Coomassie blue (bottom). Recombinant Chk1 alone was loaded lane 1. Figure S3. Further Characterization of the Phosphorylation of Treslin by Chk1. Related to Figure 3. (A) GST only (lanes 1-2), GST-TRCT (lanes 3-4), and versions of this fragment containing the S1887A (lanes 5-6), S1893A (lanes 7-8), T1897A (lanes 9-10), or S1893A and T1897A mutations (lanes 11-12) were incubated with recombinant WT Chk1 (lanes 1, 3, 5, 7, 9, and 11) or KD Chk1 (lanes 2, 4, 6, 8, 10, and 12). Reactions were processed for phosphorimaging (top) and Coomassie blue staining (bottom). (B) GST only (lane 2), GST-tagged form of wild-type TRCT (lane 3), and a version of this fragment containing both the S1893A and T1897A mutations (lane 4) were expressed in bacteria and isolated with glutathione agarose. Bead-bound fragments were incubated with 293T nuclear lysates. Beads were retrieved, washed, and processed for immunoblotting with anti-Chk1 (top) and Coomassie blue staining (bottom). 293T nuclear lysate was loaded in lane 1 as input. (C) GST only (lane 1), GST-TRCT plus GST (lanes 2-3), GST-Cdc25(254-316) plus GST (lanes 4-5), and GST-TRCT plus GST-Cdc25(254-316) (lanes 6-7) were incubated with recombinant WT Chk1 (lanes 1, 2, 4, and 6) or KD Chk1 (lanes 3, 5, and 7). Reactions were processed for phosphorimaging (top) and Coomassie blue staining (bottom). Relative levels of phosphorylation normalized to protein amount are shown. Phosphorylation of Cdc25(254-316) in lane 4 was denoted as 1. Figure S4. Further Characterization of the Effect of Chk1 on DNA Replication. Related to Figure 4. (A) NPE was treated with 1 U/µl Benzonase (lane 4) or 50 µg/ml ethidium bromide (lane 5) for 30 min on ice before immunoprecipitation was performed as in Figure 4D. (B) HSS was incubated for 30 min at room temperature. NPE containing buffer (lanes 1-3), or AZD7762 (lanes 4-6) was added to HSS. The final concentration of AZD7762 was 0.5 µM.

Samples were incubated in the presence of [α-³²P]dATP to determine chromosomal DNA replication. Quantitation of the results is presented in Figure 4E. Figure S5. The TRCT Domain Stimulates the Loading of Numerous Replication Proteins onto Chromatin. Related to Figure 6. HSS was incubated without (lanes 3-4) or with sperm nuclei (lanes 5-9) for 30 min. NPE and either WT (lanes 1, 3, 6, and 8) or 7A TRCT (lanes 2, 4, 7, and 9) at a final concentration of 60 ng/µl were added to HSS. Chromatin was isolated at indicated times and immunoblotted with the indicated antibodies. NPE/HSS mixture was loaded in lanes 1-2. Figure S6. Determination of Inter-origin Distances in Egg Extracts Treated with the TRCT Domain. Related to Figure 6. (A) Schematic of DNA fiber assay in Xenopus egg extracts and fiber visualization by immunofluorescence. (B) Representative images of fibers from egg extracts containing buffer alone, TRCT WT (75 ng/µl), or TRCT 7A (75 ng/µl) are shown. Distributions of inter-origin distances are shown in Figure 6C. (C) HSS was incubated in the presence of sperm nuclei (lanes 1-6) for 30 min. NPE either lacking (lanes 1-3) or containing 1 µM GST-p27 (lanes 4-6) and buffer (lanes 1 and 4), TRCT WT (lanes 2 and 5), or TRCT 7A (lanes 3 and 6) were added to HSS. Final concentration of TRCT was 60 ng/µl. Mixtures were incubated for 90 min and immunoblotted with anti-P-Chk1 (top), anti-Chk1 (middle), and anti-GST (bottom). Figure S7. Replication-fork Speed in Cells Expressing WT or 7A Treslin. Related to Figure 7. U2OS T-Rex cells harboring Treslin WT or 7A were cultured in the presence of doxycycline for 48 hr. DNA fibers were prepared and replication-fork speed was determined.
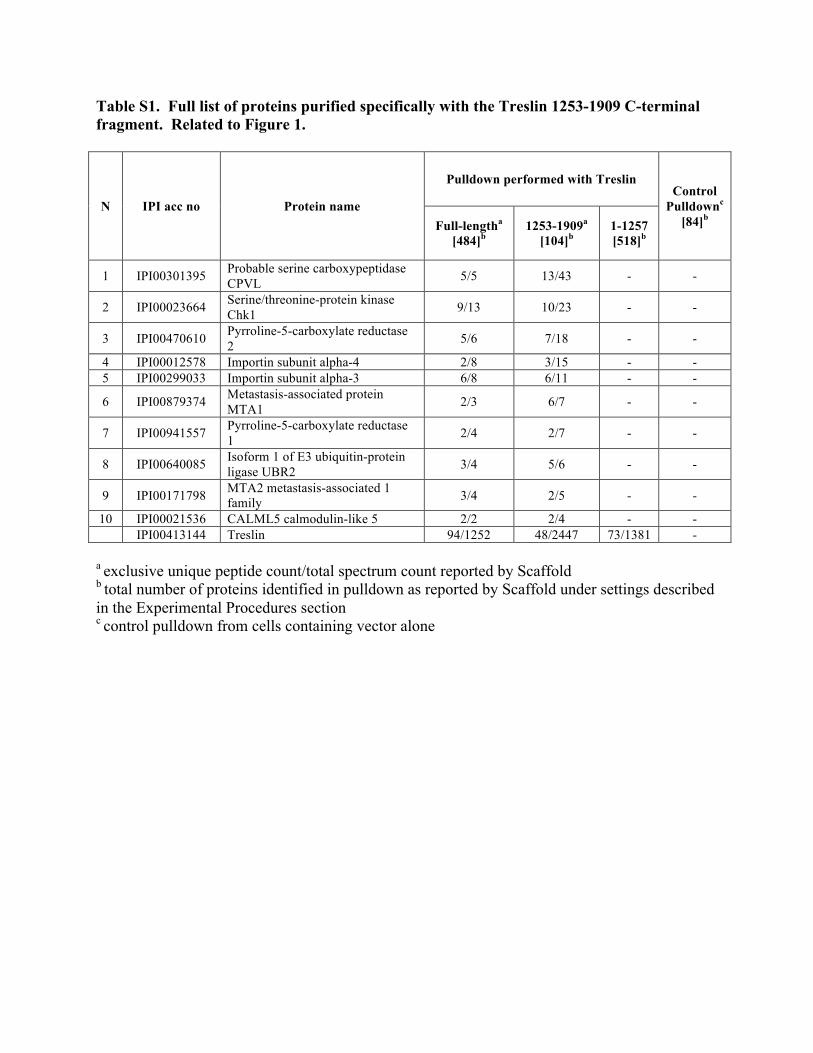
Table S1. Full list of proteins purified specifically with the Treslin 1253-1909 C-terminal fragment. Related to Figure 1.
N IPI acc no Protein name
Pulldown performed with Treslin Control
Pulldownc [84]b Full-lengtha
[484]b 1253-1909a
[104]b 1-1257 [518]b
1 IPI00301395 Probable serine carboxypeptidase CPVL 5/5 13/43 - -
2 IPI00023664 Serine/threonine-protein kinase Chk1 9/13 10/23 - -
3 IPI00470610 Pyrroline-5-carboxylate reductase 2 5/6 7/18 - -
4 IPI00012578 Importin subunit alpha-4 2/8 3/15 - - 5 IPI00299033 Importin subunit alpha-3 6/8 6/11 - -
6 IPI00879374 Metastasis-associated protein MTA1 2/3 6/7 - -
7 IPI00941557 Pyrroline-5-carboxylate reductase 1 2/4 2/7 - -
8 IPI00640085 Isoform 1 of E3 ubiquitin-protein ligase UBR2 3/4 5/6 - -
9 IPI00171798 MTA2 metastasis-associated 1 family 3/4 2/5 - -
10 IPI00021536 CALML5 calmodulin-like 5 2/2 2/4 - - IPI00413144 Treslin 94/1252 48/2447 73/1381 -
a exclusive unique peptide count/total spectrum count reported by Scaffold b total number of proteins identified in pulldown as reported by Scaffold under settings described in the Experimental Procedures section c control pulldown from cells containing vector alone

SUPPLEMENTAL EXPERIMENTAL PROCEDURES
Plasmids
Deletion mutants were created by PCR-based methods. Point mutations were created
with the QuikChange kit using Pfu DNA polymerase (Agilent Technologies). All mutations
were confirmed by DNA sequencing.
Additional Antibodies
Anti-FLAG (M2) and anti-human TopBP1 (A300-111A) antibodies were obtained from
Agilent Technologies and Bethyl Laboratories, respectively. An anti-human mouse monoclonal
antibody that also recognizes Xenopus PCNA and anti-human Orc2 antibodies were purchased
from BD Biosciences. Anti-human-phospho-Chk1 antibodies (S345, 133D3), which also
recognize S344-phosphorylated Xenopus Chk1, were purchased from Cell Signaling
Technology. Antibodies against human Chk1 (G-4) and GST (Z-5) were obtained from Santa
Cruz Biotechnology. Antibodies against Xenopus Chk1, TopBP1, Cdc45, p60 of DNA
polymerase epsilon, and Orc2 were described previously (Kumagai et al., 1998; Kumagai et al.,
2006; Lee et al., 2005; Lee et al., 2003). Antisera against Xenopus Sld5 were generously
supplied by H. Takisawa (Osaka University, Osaka, Japan). Antibodies against Xenopus ISWI
were prepared as described (Demeret et al., 2002). An anti-human Mcm2 mouse monoclonal
antibody (BM28) that cross-reacts with Xenopus Mcm2 was purchased from EM Biosciences.
For the DNA fiber assays, anti-CldU (anti-BrdU BU1/75(ICR1)) and anti-IdU antibodies (anti-
BrdU, clone B44) were obtained from Novus Biologicals and BD Biosciences, respectively.
Alexa Fluor 594 goat anti-mouse and Alexa Fluor 488 donkey anti-rat antibodies were purchased
from Invitrogen. NeutrAvidin Oregon Green 488 was purchased from Life Technologies.

Mouse anti-digoxigenin Alexa Fluor 594 was purchased from Jackson ImmunoResearch.
Biotinylated anti-avidin was obtained from Thermo Scientific.
Protein Purification and Mass Spectrometry Analysis
Plasmids encoding different versions of Treslin-SF were each transfected into a total of
six 15-cm dishes of 293T cells. DNA (17 µg) was first diluted in 2.5 ml OPTI-MEM, which was
then supplemented with 20 µg/ml polyethyleneimine. After 15 min, the mixture was added to
each dish. After 48 hr, cells were harvested and lysed in 5.4 ml hypotonic buffer (10 mM Hepes-
KOH, pH 7.9, 1.5 mM MgCl2, 10 mM KCl, 0.5 mM DTT, 0.5 mM PMSF, and 0.6 µM
tautomycin) for 10 min on ice. NP-40 was added to 0.05% and lysates were centrifuged for 10
min at 330 g at 4ºC. The pellet was resuspended in 6.75 ml high salt buffer containing 20 mM
Hepes-KOH, pH 7.5, 500 mM NaCl, 0.5% Triton X-100, 5 mM EDTA, 1 mM DTT, 10 mM β-
glycerolphosphate, 1 mM NaF, 0.1 mM vanadate, 0.6 µM tautomycin, 1 mM PMSF, 10 µg/ml
each of pepstatin, chymostatin, and leupeptin. The supernatant was collected by centrifugation at
16,000 g for 10 min and diluted with 20 mM Hepes-KOH (pH 7.5) to 150 mM NaCl. The
supernatant was again collected by centrifugation at 16,000g for 5 min and incubated with 180 µl
anti-FLAG M2 agarose (Sigma) at 4ºC for 4 hr. After washing, Treslin-associated proteins were
eluted overnight in 0.9 ml elution buffer containing 20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5
mM EDTA, 1 mM DTT, 0.5 mM PMSF, and 120 ng/µl 3X-FLAG peptide. The eluate was
collected by centrifugation and the anti-FLAG resin was eluted twice more with 1.35 ml 20 mM
Tris-HCl, pH 7.5, 150 mM NaCl, 0.13% Triton-X-100, and 0.5 mM PMSF. All eluates were
combined and incubated with 260 µl S protein agarose at 4ºC for 4 hr. The samples were
washed with 20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Triton-X-100, and 0.5 mM PMSF
and eluted in SDS sample buffer.

The Treslin-associated proteins were resolved by SDS-PAGE, stained with Coomassie,
and subjected to GeLC MS/MS analysis. Briefly, complete gel lanes were sliced into 6-8 slabs,
each of which was in-gel digested with trypsin (Shevchenko et al., 2006). Tryptic peptides
recovered from the gel matrix were subjected to LC-MS/MS at the Ultimate 3000 nanoLC
system (Thermo Dionex, Idstein, Germany) interfaced to a LTQ Orbitrap Velos hybrid mass
spectrometer (Thermo Fisher Scientific, Bremen, Germany) as described (Vasilj et al., 2012).
The acquired spectra were searched against an IPI Human sequence database (version 3.86)
using Mascot software (MatrixScience, London, UK) under the following settings: tryptic
enzyme specificity; mass tolerance of 5 ppm and 0.5 Da for precursor and fragment ions,
respectively; up to two mis-cleavages allowed; variable modifications: oxidation of methionine;
deamidation of glutamines and asparagines; propionamidation of cysteines, protein acetylation at
the N-terminus; and, where specified, serine / threonine / tyrosine phosphorylation. Protein hits
were further validated using Scaffold software (Proteome Software, Portland, USA, version
2.04) under the following settings: two or more matched peptides required under 95% and 99%
peptide and protein statistical significance thresholds, respectively. False Discovery Rate (FDR)
as calculated by the Scaffold software was below 0.1.
Preparation and Processing of Nuclear Lysates from Human Cells
Cells were lysed in high salt buffer containing 20 mM Hepes-KOH, pH 7.5, 500 mM
NaCl, 0.5% Triton X-100, 5 mM EDTA, 1 mM DTT, 10 mM β-glycerolphosphate, 1 mM NaF,
0.1 mM vanadate, 0.6 µM tautomycin, 1 mM PMSF, 10 µg/ml each of pepstatin, chymostatin,
and leupeptin. The supernatant was collected by centrifugation at 16,000 g for 10 min and
diluted with 20 mM Hepes-KOH (pH 7.5) to 150 mM NaCl. The supernatant was again
collected by centrifugation at 16,000 g for 5 min.

For pulldowns or immunoprecipitations, lysates were incubated for 2 hr at 4ºC with S
protein agarose, anti-FLAG agarose, or antibodies bound to protein A or protein G magnetic
beads. The samples were washed four times with 20 mM Hepes-KOH, pH 7.5, 150 mM NaCl,
0.15% Triton X-100, 1.5 mM EDTA, 1 mM DTT, 10 mM β-glycerolphosphate, 1 mM NaF, and
0.1 mM vanadate. For kinase assays with Treslin-SF proteins, anti-FLAG beads were collected,
washed twice with buffer containing 20 mM Hepes-KOH, pH 7.5, 150 mM NaCl, 0.15% Triton
X-100, 1.5 mM EDTA, and 0.3 mM DTT; twice with high salt buffer containing 20 mM Hepes-
KOH, pH 7.5, 450 mM NaCl, and 1 mM DTT; twice with RIPA buffer containing 20 mM Tris-
HCl, pH 7.5, 150 mM NaCl, 2 mM EDTA, 1% NP40, and 0.1% SDS; and twice with HBS
buffer containing 20 mM Hepes-KOH, pH 7.5, and 150 mM NaCl. After washing, bead-bound
proteins were either stored at -80˚C or used immediately.
Production of GST Fusion Proteins
Plasmids encoding GST-tagged versions of the TRCT were introduced into Escherichia
coli Rosetta (DE3)pLysS cells (EMD). E. coli cells expressing the fragments were resuspended
in 0.2 M sodium borate, pH 8.2, containing 0.5 M NaCl, 0.5% NP-40, 5 mM EGTA, and 1 mM
PMSF, and then were sonicated. The lysates were clarified by centrifugation for 10 min at
20,000 g and incubated with glutathione agarose at 4ºC for 2 hr. The proteins were eluted with
50 mM Tris-HCl (pH 8.0) containing 10 mM reduced glutathione. Typically, the expressed
proteins were dialyzed into ELB. GST-p27 was purified as described previously (Kumagai et
al., 2011).
In Vitro Direct Binding Assay
GST-tagged human Treslin fragments were expressed in bacteria and isolated with
glutathione agarose beads. Bead-bound fragments (1 µg) were incubated with 200 ng His6-

tagged Xenopus Chk1 (Kumagai et al., 1998) in 0.5 ml of binding buffer containing 20 mM Tris-
HCl, pH 7.5, 150 mM NaCl, 0.5% NP40, 1 mM EDTA, 1 mM DTT, and 10% glycerol at 4ºC for
90 min. The beads were retrieved, washed, and subjected to immunoblotting and staining with
Coomassie brilliant blue.
Kinase Assays
Full-length human Treslin-SF proteins (WT and 7A) bound to anti-FLAG beads or
soluble GST-tagged human TRCT fragments were incubated with WT or KD (N135A) Xenopus
His6-Chk1 (Kumagai et al., 1998) in 40 µl of kinase buffer containing 10 mM Tris-HCl, pH 7.5,
10 mM MgCl2, 1 mM DTT, 2 µCi [γ-³²P] ATP, and 10 µM ATP at room temperature for 30 min.
Reactions were terminated with SDS sample buffer. For assays with Cdc25, we utilized a GST-
tagged version of residues 254-316 from Xenopus Cdc25 in which S287 is the sole site of
phosphorylation for Chk1 (Kumagai et al., 1998).
Use of the Xenopus NPE System
HSS (33 µl) was mixed with 1.32 µl ATP regeneration mix (10 µl 1 M phosphocreatine,
10 µl 100 mM ATP, and 0.5 µl 5 mg/ml creatine kinase), 0.5 µl 0.2 mg/ml nocodazole, and
sperm nuclei (4,000 or 10,000 per µl for replication and chromatin-binding assays, respectively).
At 15 min, 20 µl NPE was diluted with 46 µl ELB and supplemented with 2.64 µl ATP
regeneration mix and 2.64 µl 50 mM DTT. For replication assays, 5 µCi [α-³²P]dATP was also
added to NPE. At 30 min, TRCT fragments or AZD7762 were added to the HSS. One volume
of HSS was then immediately mixed with two volumes of NPE and incubated as indicated. For
gel analysis of reactions, incubations were terminated with SDS sample buffer. For preparation
of chromatin fractions, reactions were mixed with 60 µl ice cold ELB containing 0.2% Triton X-

100 and transferred onto 180 µl ELB containing 0.5 M sucrose. Chromatin was collected by
centrifugation at 16,000 g and washed twice with ELB containing 0.25 M sucrose. For
replication assays, aliquots from reactions were mixed with an equal volume of 2X stop solution
(2% SDS, 75 mM EDTA, 25 mM Tris-HCl, pH 8) and digested with proteinase K. Samples
were electrophoresed in 0.8% agarose gels. Gels were dried and subjected to phosphorimaging.
Preparation of Nuclear Lysates from Egg Extracts
Methods have been described previously (Kumagai et al., 2010). In brief, egg extracts
containing 3,000 demembranated sperm nuclei per µl were incubated at room temperature for 90
min. Nuclei were isolated by centrifugation for 5 min at 6,100 g through a sucrose cushion (1M
sucrose dissolved in 20 mM Hepes-KOH, pH 7.5, 80 mM KCl, 2.5 mM potassium gluconate,
and 10 mM magnesium gluconate). Pellets were resuspended in the sucrose solution and
centrifuged again to collect nuclei. The nuclei were resuspended in 450 mM NaCl dissolved in
20 mM Hepes-KOH, pH 7.5, 10 mM β-glycerolphosphate, 6 mM DTT, 2.5 mM EGTA, 1 mM
NaF, 0.1 mM sodium vanadate, 0.6 µM tautomycin, 0.1% NP-40, 0.1 mM PMSF, and 10 µg/ml
each of pepstatin, chymostatin, and leupeptin. The lysates were centrifuged at 11,000 g for 10
min to remove particulate material and chromatin. The supernatant was diluted with 20 mM
Hepes-KOH (pH 7.5) to adjust the concentration of NaCl to 75 mM and centrifuged again. The
salt-extracted nuclear lysates were subject to SDS-PAGE and immunoblotting.
DNA Replication Assays with Whole Egg Extracts
For replication assays in whole egg extracts, 1 µCi [α-³²P]dATP was added to 10 µl egg
extract containing 0.4 mM CaCl2 and 3,000 sperm nuclei per µl. At indicated time points,
reactions were stopped by addition of 10 µl 2X sample buffer (80 mM Tris-HCl, pH 8.0, 8 mM

EDTA, 0.13% phosphoric acid, 10% Ficoll, 5% SDS and 0.2% bromophenol blue). Samples
were digested with 1 mg/ml proteinase K at 37ºC for 1 hr and loaded onto gels containing 1%
agarose. Gels were dried and subjected to phosphorimaging.
DNA Fiber Assays with Xenopus Egg Extracts
Methods were described previously with some modifications (Bellelli et al., 2014;
Marheineke et al., 2009). Sperm nuclei (3000/µl) were incubated in egg extracts containing 20
µM digoxigenin-11-dUTP (Roche) in the presence of buffer, TRCT WT (75 ng/µl), or TRCT 7A
(75 ng/µl). At 30 min, biotin-16-dUTP (Biotium) was added to extracts at a final concentration
20 µM. At 120 min, nuclei were isolated by centrifugation for 5 min at 6,100 g through a
sucrose cushion containing 1M sucrose dissolved in 20 mM HEPES-KOH, pH 7.5, 80 mM KCl,
2.5 mM potassium gluconate, and 10 mM magnesium gluconate. Pellets were resuspended
gently in sucrose cushion and centrifuged again to collect nuclei. Nuclei were resuspended in
PBS. The nuclear suspension was spotted onto one end of a microscope slide (Fisher Scientific)
and mixed with six volumes of lysis buffer (0.5% SDS, 50 mM EDTA, 200 mM Tris-HCl, pH
7.4) for 10 min. The slide was carefully tilted 15º to allow the fibers to spread by gravity. Fibers
were then air dried, fixed in methanol and acetic acid (3:1) for 3 min, and subsequently
denatured with 2 N HCl for 1 hr. Slides were neutralized and washed with PBS (once at pH 8.0
and 3 times at pH 7.4), and blocked with 1% nucleic acid blocking reagent (Roche) in PBS at
room temperature for 1 hr. Slides were incubated with NeutrAvidin Oregon Green 488 (1/50)
and mouse anti-digoxigenin Alexa Fluor 594 (1/50) in blocking buffer at room temperature for 1
hr. Slides were washed and incubated with biotinylated anti-avidin and goat anti-mouse Alexa
Fluor 594 in blocking buffer at room temperature for 1 hr. Slides were again washed and
incubated with NeutrAvidin Oregon Green 488 (1/50) in blocking buffer for 45 min. Slides were

washed and mounted in Vectashield mounting medium (Vector Laboratories) prior to analysis
using an Axioplan microscope (Carl Zeiss) equipped with a Plan Neofluar (63x/0.75 NA)
objective. Inter-origin distances were measured using ImageJ software (National Institutes of
Health; http://rsbweb.nih.gov/ij/) and kb values were obtained by multiplying micrometers by
2.59 (Jackson and Pombo, 1998).
DNA Fiber Assays with Human Cells
After labeling with CldU and IdU, cells were harvested and resuspended in cold PBS.
The cell suspension was spotted onto one end of a microscope slide (Fisher Scientific) and mixed
with six volumes of lysis buffer (0.5% SDS, 50 mM EDTA, 200 mM Tris-HCl, pH 7.4) for 5
min. The slide was carefully tilted 15º to allow the fibers to spread by gravity. Fibers were then
fixed in methanol and acetic acid (3:1) for 3 min and subsequently denatured with 2 N HCl for 1
hr. Slides were neutralized and washed with PBS (once at pH 8.0 and 3 times at pH 7.4) and
blocked with 10% goat serum and 0.1% Triton-X in PBS for 1 hr. Slides were incubated with
primary antibodies against IdU (1:100 in blocking buffer) and CldU (1:200 in blocking buffer)
and secondary antibodies (Alexa Fluor 488 donkey anti-rat and Alexa Fluor 594 goat anti-mouse,
1:300 in blocking buffer) for 1 hr each. Slides were mounted in Vectashield mounting medium
(Vector Laboratories) prior to analysis using an Axioplan microscope (Carl Zeiss) equipped with
a Plan Neofluar (63x/0.75 NA) objective.

SUPPLEMENTAL REFERENCES
Bellelli, R., Castellone, M.D., Guida, T., Limongello, R., Dathan, N.A., Merolla, F., Cirafici, A.M., Affuso, A., Masai, H., Costanzo, V., et al. (2014). NCOA4 transcriptional coactivator inhibits activation of DNA replication origins. Mol. Cell 55, 123-137.
Demeret, C., Bocquet, S., Lemaitre, J.M., Francon, P., and Mechali, M. (2002). Expression of ISWI and its binding to chromatin during the cell cycle and early development. Journal of structural biology 140, 57-66.
Jackson, D.A., and Pombo, A. (1998). Replicon clusters are stable units of chromosome structure: evidence that nuclear organization contributes to the efficient activation and propagation of S phase in human cells. J. Cell Biol. 140, 1285-1295.
Kumagai, A., Guo, Z., Emami, K.H., Wang, S.X., and Dunphy, W.G. (1998). The Xenopus Chk1 protein kinase mediates a caffeine-sensitive pathway of checkpoint control in cell-free extracts. J. Cell Biol. 142, 1559-1569.
Kumagai, A., Lee, J., Yoo, H.Y., and Dunphy, W.G. (2006). TopBP1 activates the ATR-ATRIP complex. Cell 124, 943-955.
Kumagai, A., Shevchenko, A., Shevchenko, A., and Dunphy, W.G. (2010). Treslin collaborates with TopBP1 in triggering the initiation of DNA replication. Cell 140, 349-359.
Kumagai, A., Shevchenko, A., Shevchenko, A., and Dunphy, W.G. (2011). Direct regulation of Treslin by cyclin-dependent kinase is essential for the onset of DNA replication. J. Cell Biol. 193, 995-1007.
Lee, J., Gold, D.A., Shevchenko, A., Shevchenko, A., and Dunphy, W.G. (2005). Roles of replication fork-interacting and Chk1-activating domains from Claspin in a DNA replication checkpoint response. Mol. Biol. Cell 16, 5269-5282.
Lee, J., Kumagai, A., and Dunphy, W.G. (2003). Claspin, a Chk1-regulatory protein, monitors DNA replication on chromatin independently of RPA, ATR, and Rad17. Mol. Cell 11, 329-340.
Marheineke, K., Goldar, A., Krude, T., and Hyrien, O. (2009). Use of DNA combing to study DNA replication in Xenopus and human cell-free systems. Methods Mol. Biol. 521, 575-603.
Shevchenko, A., Tomas, H., Havlis, J., Olsen, J.V., and Mann, M. (2006). In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 1, 2856-2860.
Vasilj, A., Gentzel, M., Ueberham, E., Gebhardt, R., and Shevchenko, A. (2012). Tissue proteomics by one-dimensional gel electrophoresis combined with label-free protein quantification. J. Proteome Res. 11, 3680-3689.