indirect elisa-based approach for comparative measurement of high-affinity cohesin-dockerin...
TRANSCRIPT

Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use.
This chapter was originally published in the book Methods in Enzymology, Vol. 510 published by Elsevier, and the attached copy is provided by Elsevier for the author's benefit and for the benefit of the author's institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier's permissions site at:
http://www.elsevier.com/locate/permissionusematerial
From: Michal Slutzki, Yoav Barak, Dan Reshef, Ora Schueler-Furman, Raphael Lamed, and Edward A. Bayer, Measurements of Relative Binding of Cohesin and Dockerin Mutants
Using an Advanced ELISA Technique for High-Affinity Interactions. In Harry J. Gilbert, editor: Methods in Enzymology, Vol. 510,
Burlington: Academic Press, 2012, pp. 417-428. ISBN: 978-0-12-415931-0
© Copyright 2012 Elsevier Inc. Academic Press

Author's personal copy
C H A P T E R T W E N T Y - T W O
M
IS
*{
{
}
ethods
SN 0
DepaChemDepaHadaDepa
Measurements of Relative Binding
of Cohesin and Dockerin Mutants
Using an Advanced ELISA Technique
for High-Affinity Interactions
Michal Slutzki,* Yoav Barak,† Dan Reshef,‡
Ora Schueler-Furman,‡ Raphael Lamed,§ and Edward A. Bayer*
Contents
1. In
in
076
rtmicartmssahrtme
troduction
Enzymology, Volume 510 # 2012
-6879, DOI: 10.1016/B978-0-12-415931-0.00022-7 All rig
ent of Biological Chemistry, The Weizmann Institute of Science, Rehovot, Israell Research Support, The Weizmann Institute of Science, Rehovot, Israelent of Microbiology and Molecular Genetics, Institute for Medical Research IsMedical School, The Hebrew University, Jerusalem, Israelnt of Molecular Microbiology and Biotechnology, Tel Aviv University, Ramat Aviv
Else
hts
rael
, Is
418
2. P
reparation of Constructs for ELISA Procedure 4202
.1. C BM-fused cohesins 4212
.2. E xpression and purification of xylanase-fused dockerins 4223. D
etection of Free Dockerins Using iELISA 4223
.1. C oating 4243
.2. C ohesin–dockerin equilibrated interaction 4243
.3. D etection of unbound dockerin concentration by iELISA 4244. D
ata Analysis 4255. N
otes 4266. S
ummary 427Refe
rences 427Abstract
The cellulosome is a large bacterial extracellular multienzyme complex able to
degrade crystalline cellulosic substrates. The complex contains catalytic and
noncatalytic subunits, interconnected by high-affinity cohesin–dockerin inter-
actions. In this chapter, we introduce an optimized method for comparative
binding among different cohesins or cohesin mutants to the dockerin partner.
This assay offers advantages over other methods (such as ELISA, cELIA, SPR,
and ITC) for particularly high-affinity binding interactions. In this approach, the
high-affinity interaction of interest occurs in the liquid phase during the
vier Inc.
reserved.
-Canada,
rael
417

418 Michal Slutzki et al.
Author's personal copy
equilibrated binding step, whereas the interaction with the immobilized phase
is used only for detection of the unbound dockerins that remain in the solution
phase. Once equilibrium conditions are reached, the change in free energy of
binding (DDGbinding), as well as the affinity constant of mutants, can be esti-
mated against the known affinity constant of the wild-type interaction. In light
of the above, we propose this method as a preferred alternative for the relative
quantification of high-affinity protein interactions.
Abbreviations
BSA
bovine serum albumin CBM carbohydrate-binding module family 3a fromC. thermocellum
cELIA competitive enzyme-linked interaction assay Coh cohesin Doc dockerin DSC differential scanning calorimetry ELISA enzyme-linked immunosorbent assay HRP horseradish peroxidase IPTG isopropyl-1-thio-b-D-galactoside ITC isothermal titration calorimetry MES 2-(N-morpholino) ethanesulfonic acid SDS-PAGE polyacrylamide gel electrophoresis in sodium dode-cyl sulfate
SPR surface-plasmon resonance TBS Tris-buffered saline WT wild type Xyn xylanase1. Introduction
The ultra-tight interactions between cohesins and dockerins deter-mine the organization of cellulosome (Bayer et al., 1983), the multienzymecomplex responsible for efficient degradation of cellulose in selected anaer-obic bacteria. Cohesins and dockerins are known to interact with oneanother in a highly specific manner, depending on their function (i.e.,binding to enzymatic, structural, or cell-surface anchoring subunits) andtheir microbial origin (e.g., dockerins from one bacterial species usuallyfail to bind cohesins from another). The nature of the specificity and

Indirect ELISA for Cohesin–Dockerin Interaction 419
Author's personal copy
promiscuity, which simultaneously exist in this system, is a very intriguingsubject for scientific research. To investigate the possible role of differentresidues in recognition patterns, target amino acids can either be mutatedto alanine or be modified to the corresponding residue from anothercohesin/dockerin pair (usually from another species; Fierobe et al., 2001;Handelsman et al., 2004; Mechaly et al., 2000, 2001; Nakar et al., 2004;Pinheiro et al., 2008).
Several methods, described in the above publications, have beendeveloped to measure the strength of the various cohesin–dockerin interac-tions, such as enzyme-linked immunosorbent assay (ELISA), competitiveenzyme-linked interaction assay (cELIA), surface-plasmon resonance (SPR;e.g., BIAcore), isothermal titration calorimetry (ITC), and differential scan-ning calorimetry (DSC). Each of these methods, however, has limitations,either in general or in specific terms, for measuring high-affinity interactions.Moreover, some of these methods are not amenable for comparative analysisof multiple samples. Here, we specify the disadvantages of each method ascompared to the indirect ELISA (iELISA)-based method that we describe indetail in this chapter.
(1) Standard affinity-based ELISA (Barak et al., 2005): In this method, theinteraction occurs between two different phases: solid (immobilized)and liquid (protein in solution). Thus, we can control concentrationsof only one binding partner (the one that is in solution), since duringcoating the immobilized protein undergoes partial denaturation.Clearly, binding constants cannot be determined using this technique,since concentrations of immobilized proteins are unknown, andthe level of immobilization of different mutants on the plate is hardto compare.
(2) cELIA (Handelsman et al., 2004): At first glance, it seems as if theinteraction between components occurs in the soluble phase in thisassay. However, since the components (mutant cohesin and dockerin)are added simultaneously to the well coated with wild-type (WT)cohesin, the binding between the mutant cohesin and dockerin isaccompanied by competition with the binding to the immobilizedprotein. Thus, not only is the immobilized protein involved in estimat-ing the status of the interaction, but there is an uncertainty with respectto the equilibrium conditions, which interferes with calculations of thechange in free energy of binding (DDGbinding).
(3) SPR: This methodology is limited by very slow dissociation rates forthe cohesin–dockerin complex (undetectable, i.e., <10�4 s�1), cou-pled with very high affinity. Therefore, low protein concentrations(below the sensitivity limits) had to be used to determine the associa-tion constants in a precise manner. In addition, since one of the inter-acting proteins is immobilized, the binding constants are apparently

420 Michal Slutzki et al.
Author's personal copy
overestimated relative to solution-based approaches, for example,ITC (Fierobe et al., 1999; Mechaly et al., 2001) vs. SPR (Carvalhoet al., 2003, 2007; Miras et al., 2002), respectively.
(4) ITC: The binding affinity that can be accurately measured by thismethod is limited to a maximum of Ka�109 M�1 (Velazquez-Campoy et al., 2004). To estimate higher affinities, very low levels ofprotein concentrations are required, which are out of the sensitivity limitof the instrument. Alternatively, competitive binding can be measured in“displacement titration” (Velazquez-Campoy and Freire, 2006), but thiswould require an available ligand of lower binding affinity.
(5) DSC: This method can also measure very high-affinity interactions, butit cannot be used if the proteins are not soluble enough under the highconcentrations required for this approach, or if thermal denaturationis not reversible. Another protein is frequently required as a carrier toimprove solubility and expression of the target protein, which mayinterfere in distinguishing the peak of energy input contributed by theprotein of interest.
Owing to the above-mentioned drawbacks, we therefore developed analternative approach, based on iELISA, to overcome the above limitations.Our approach incorporates the previously described cELIA and ELISAmethods, but modified by addition of a necessary equilibration step forinteraction in solution. In this modified method, the estimated interactiontakes place in the soluble phase (opposite to that of ELISA), which alsorenders this method more sensitive than cELIA, since the procedure isperformed under conditions of much lower dockerin concentrations. Theapproach allows estimates of the change in free energy of binding(DDGbinding) between the test protein (e.g., mutant cohesin or dockerinmodule) and the reference protein (e.g., wild type). The present approach issimilar to a method reported earlier (Friguet et al., 1985), with moreaccurate data analysis.
2. Preparation of Constructs for
ELISA Procedure
When dockerins and cohesins are produced in their free form, theyusually exhibit low expression levels, low solubilities, and a tendency toaggregate (Adams et al., 2005; Fierobe et al., 1999; Lytle et al., 2001).Therefore, we chose to fuse them to carrier proteins that are known fortheir good expression levels and solubility, when expressed in Escherichia coli(Barak et al., 2005). These carrier proteins were also selected from theoriginal molecular context that is relevant to cellulose degradation, that is,the dockerin is fused to a particularly high-expressing xylan-hydrolyzing

Indirect ELISA for Cohesin–Dockerin Interaction 421
Author's personal copy
enzyme (xylanase)—a thermostable enzyme originating from the bacteriumGeobacillus stearothemophilus, and the cohesin is expressed as a fusion proteintogether with a carbohydrate-binding module (CBM)—derived from thethermostable cellulosomal scaffoldin protein of Clostridium thermocellum(Morag et al., 1995). Using such protein fusions, we achieve standardizationin purification procedures and in the assay itself. By adding a xylanasemodule on all dockerins, we were able to use the same antibodies to detectall the dockerins.
2.1. CBM-fused cohesins
Reagents
LB (Luria–Bertani) broth supplemented with 50 mg/ml kanamycinIPTG (isopropyl-1-thio-b-D-galactoside) (FermentasUAB, Vilnuis, Lithuania)Amorphous cellulose: 7.5 g phosphoric acid-treated Avicel/liter DDWWashing buffer #1: Tris-buffered saline (TBS), pH 7.4 with 1 M NaClWashing buffer #2: TBS, pH 7.4Triethylamine: 1% (v/v)2-(N-Morpholino) ethanesulfonic acid (MES): 1 M, pH 5.5
Protocol
The CBM-Coh gene cassette (Barak et al., 2005) consists of a family 3aCBM from the C. thermocellum CipA scaffoldin (Morag et al., 1995)cloned into plasmid pET28a (Novagen Inc., Madison, WI, USA), intowhich any cohesin gene can be introduced between BamHI and XhoIrestriction sites of the plasmid. For demonstration purposes, the Coh2,the second cohesin from the C. thermocellum CipA scaffoldin, was used inthis chapter.
CBM-Cohs were thus expressed and purified, in a manner similar tothe description of Barak et al. (2005). A cell culture was grown in LBmedium with kanamycin. Upon induction of recombinant proteinexpression with IPTG, the culture was grown for 3 h at 37 �C. Aftersonication, 15 ml of amorphous cellulose was added to the supernatantand used for affinity purification. Binding between cellulose and CBMoccurred during overnight incubation on a rotator at 4 �C. The amor-phous cellulose was pelleted by centrifugation at 4000 rpm at 4 �C for5 min and washed three times with 45 ml washing buffer #1, and threetimes with washing buffer #2. After each step, the suspension waspelleted again and the supernatant fluids were discarded. Finally, theCBM-Cohs were eluted with triethylamine and neutralized using MESuntil pH 7 was reached. Purity was verified using SDS-PAGE. Theproteins were stored in 50% glycerol at �20 �C.

422 Michal Slutzki et al.
Author's personal copy
2.2. Expression and purification of xylanase-fused dockerins
Reagents
LB broth supplemented with 50 mg/ml kanamycinIPTG (Fermentas UAB)TBS, pH 7.4Binding buffer: TBS, supplemented with 5 mM imidazole (Merck KGaA,
Darmstadt, Germany)Washing buffer: TBS, supplemented with 20 mM imidazoleElution buffer #1: TBS, supplemented with 100 mM imidazoleElution buffer #2: TBS, supplemented with 250 mM imidazoleNi-NTA agarose (Qiagen GmbH, Hiden, Germany)
Protocol
The Xyn-Doc gene cassette (Barak et al., 2005) consists of xylanase T6 fromG. stearothemophilus with an N-terminal His-tag (Handelsman et al., 2004;Lapidot et al., 1996) cloned into plasmid pET9d (Novagen Inc.), intowhichany dockerin encoding sequence can be introduced between the KpnI andBamHI restriction sites of the plasmid. For demonstration purposes, thedockerin from cellulase S (DocS) ofC. thermocellumwas used in this chapter.
The dockerin-bearing xylanases (Xyn-Docs) were thus expressed and pur-ified, in a manner similar to the description of Barak et al. (2005). A cellculture was grown in LBmediumwith kanamycin. Upon induction withIPTG, the culture was grown for 3 h at 37 �C. After sonication, thesupernatant was applied to a Ni-NTA column, equilibrated with bindingbuffer. The column was washed with washing buffer on a rotator at 4 �C,and the Xyn-Docs were then released with elution buffers #1 and #2,sequentially. Purity was verified using SDS-PAGE. The proteins werestored in 50% glycerol at �20 �C.
3. Detection of Free Dockerins Using iELISA
Reagents
WT CBM-Coh (10 nM, the precise working concentration is determinedby calibration as described in Barak et al., 2005)
Coating buffer: 0.1 M Na2CO3, pH 9Blocking buffer: TBS, 10 mM CaCl2, 0.05% Tween 20, 2% BSAWashing buffer: TBS, 10 mM CaCl2, 0.05% Tween 20Xyn-Doc: 600 pM (should be roughly in the range of KD)Polyclonal rabbit antibodies against Xyn-T6 (anti-Xyn antibody; Lapidot
et al., 1996): 1:10,000 (or according to a predetermined calibration)
Indirect ELISA for Cohesin–Dockerin Interaction 423
Author's personal copy
Secondary antibody–enzyme conjugate (HRP-labeled goat anti-rabbit IgG)( Jackson ImmunoResearch Laboratories Inc., West Grove, PA, USA):1:10,000 (or according to your predetermined calibration)
TMB (Dako 3,30,5,50-tetramethylbenzidine)þSubstrate-Chromogen (DakoCorp., Carpinteria, CA, USA)
Protocol
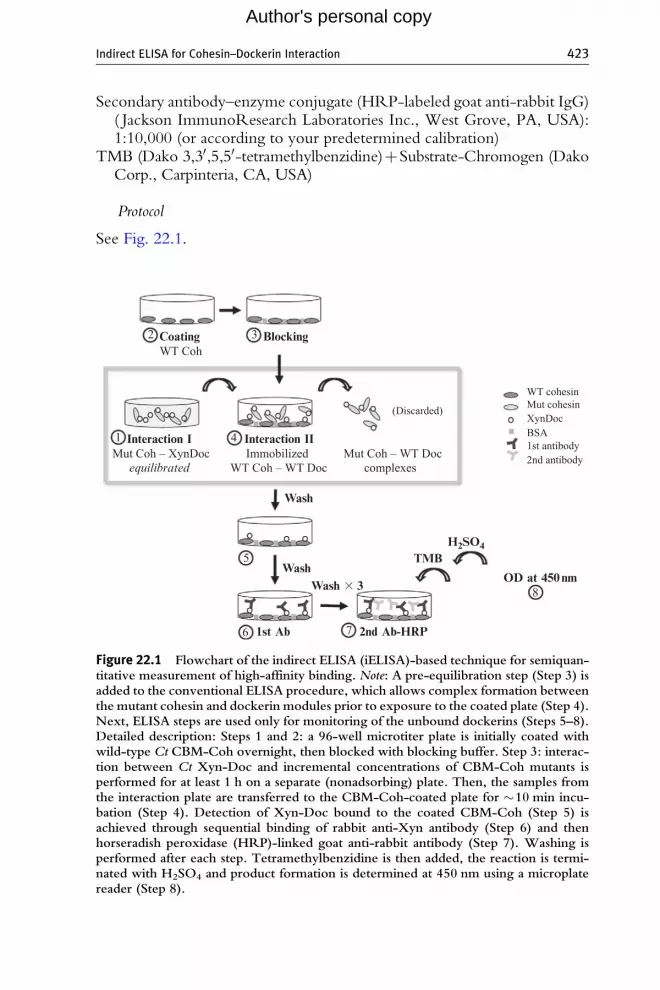
See Fig. 22.1.
Coating
Interaction I Interaction II
Wash
WashTMB
OD at 450 nm
H2SO4
1st Ab 2nd Ab-HRP
Wash � 3
Blocking2
1 4
5
7
8
6
3WT Coh
Mut Coh – XynDoc Mut Coh – WT DocImmobilizedcomplexes
(Discarded)
WT cohesinMut cohesinXynDocBSA1st antibody2nd antibody
WT Coh – WT Docequilibrated
Figure 22.1 Flowchart of the indirect ELISA (iELISA)-based technique for semiquan-titative measurement of high-affinity binding. Note: A pre-equilibration step (Step 3) isadded to the conventional ELISA procedure, which allows complex formation betweenthe mutant cohesin and dockerin modules prior to exposure to the coated plate (Step 4).Next, ELISA steps are used only for monitoring of the unbound dockerins (Steps 5–8).Detailed description: Steps 1 and 2: a 96-well microtiter plate is initially coated withwild-type Ct CBM-Coh overnight, then blocked with blocking buffer. Step 3: interac-tion between Ct Xyn-Doc and incremental concentrations of CBM-Coh mutants isperformed for at least 1 h on a separate (nonadsorbing) plate. Then, the samples fromthe interaction plate are transferred to the CBM-Coh-coated plate for �10 min incu-bation (Step 4). Detection of Xyn-Doc bound to the coated CBM-Coh (Step 5) isachieved through sequential binding of rabbit anti-Xyn antibody (Step 6) and thenhorseradish peroxidase (HRP)-linked goat anti-rabbit antibody (Step 7). Washing isperformed after each step. Tetramethylbenzidine is then added, the reaction is termi-nated with H2SO4 and product formation is determined at 450 nm using a microplatereader (Step 8).

424 Michal Slutzki et al.
Author's personal copy
3.1. Coating
In the first stage of this method, we expose the surface of the plates to ourcoating protein (Steps 1 and 2), in alkaline solution to promote its immobi-lization onto the wells of microtiter plates.
1. Dispense WT CBM-Coh in coating buffer (100 ml/well) into MaxiSorp96-well plates.
2. Incubate overnight at 4 �C.3. Discard the liquid contents of the well.4. Block the wells with 100 ml/well of blocking buffer for 1 h at 37 �Cwith
shaking.
3.2. Cohesin–dockerin equilibrated interaction
The equilibration step (Fig. 22.1, Step 3) is a critically important componentof the iELISA approach.
1. Prepare 8–12 different concentrations (ranging from pM to mM) of thedesired cohesins (e.g., both mutants and the wild-type reference) inblocking buffer (after addition of the dockerin (Xyn-Doc) the finalconcentration will be two-thirds of the original). Final volume in thewell should be 100 ml.
2. Add 50 ml of 600 pM Xyn-Doc to each well (upon addition, will bediluted threefold to a final concentration of 200 pM), mix and incubateto reach equilibrium (different times should be checked in preliminaryexperiments until no change is observed. In this case approximately 1 hincubation is sufficient).
3.3. Detection of unbound dockerin concentration by iELISA
1. After the critical equilibrium step is achieved (Fig. 22.1, Step 3), themixture is transferred to the coated plate (Step 4). The interaction atthis stage should be as short as possible (e.g., 10 min). Only unbounddockerins are expected to react here, while those in the complex arekept intact due to the extremely low dissociation rates and very shortexposure time.
2. Wash wells by adding washing buffer (Step 5). Previously formedcohesin–dockerin complexes will be washed out. No more intensivewashing is needed until the last step since each of the subsequent steps isessentially a wash for the previous one.
3. Add 100 ml anti-Xyn antibody to each well (Step 6).4. Incubate 40 min at 37 �C with shaking.5. Wash wells by filling the wells with washing buffer and discarding it.

Indirect ELISA for Cohesin–Dockerin Interaction 425
Author's personal copy
6. Add 100 ml goat anti-rabbit antibody–enzyme conjugate to each well(Step 7).
7. Incubate 40 min at 37 �C shaking.8. Wash three times with washing buffer.9. Add 100 ml TMB. Wait until the color develops (about 1–3 min).
10. Stop the reaction by addition of 50 ml 1 N H2SO4.11. Read the optical density at 450 nm using a microplate reader.
4. Data Analysis
For each mutant, at least three iELISA experiments are carried out induplicate. Absorbance data are plotted as a semilogarithmic graph as a functionof the Xyn-Doc concentration and analyzed using GraphPad Prism (version5.00 for Windows, GraphPad Software, San Diego, CA, USA). Curves arenormalized to a scale of 0–1 indicating relative binding. Results from differentexperiments are standardized by dividing them by thewild-type values in eachparticular experiment and then multiplying the product by the average of allwild-type values. The results are fitted to the sigmoidal (dose–response) curve(Motulsky and Christopoulos, 2003), assuming 1:1 binding model:
Y ¼Bottomþ Top�Bottom
1þ10X�logIC50
;
where Y is the relative binding of dockerin (Xyn-Doc) to the coated WTcohesin (CBM-Coh), “Bottom” is minimal and “Top” is maximal relativebinding, X is the concentration of test cohesin in solution, and IC50 is theconcentration of test cohesin that causes 50% inhibition of binding to thecoated WT cohesin.
Changes in free energy of binding (DDGbinding) were calculated relativeto wild type according to the equation:
DDGbinding ¼�RT lnIC50WT
IC50mut
� �¼�RT ln
KDWT
KDmut
� �;
where R is the gas constant, T is the temperature, IC50 is the concentrationof the test cohesin that causes 50% inhibition of binding, and KD is thedissociation constant of either the wild type or the mutant. If the dissocia-tion constant is known for the wild-type module (i.e., has been determinedusing an appropriate technique, such as SPR or ITC), then the value for themutant is attained using the above equation.
Examples of calculation for change in free energy of binding and KDs formutant from Fig. 22.2 are shown in Table 22.1. When DDGbinding between

0.0
-12 -10 -8 -6
0.5
WTmut1
mut2Negative controlR
elat
ive
bind
ing
log[Coh] (M)
1.0
Figure 22.2 Cohesin–dockerin binding profiles, measured using iELISA by determin-ing the amount of dockerin molecules which failed to bind to the cohesin mutant inthe equilibration step. Cohesin mutants 1 and 2 show different binding affinities relativeto the wild type.
Table 22.1 Change in free energy of binding (DDG) and dissociation constants (KD)calculated from the half-maximal inhibitory concentration (IC50)
IC50 (nM) KDmut/KDWT DDG binding (kcal/mol)
WT 1 – –
mut 1 5 5 1.0
mut 2 93 93 2.8
426 Michal Slutzki et al.
Author's personal copy
the wild-type and the mutant exceeds a threshold value (usually fixed at1.5–2.0 kcal/mol), the mutated residue is considered to participate in thedesignated binding interaction (in this case, between the cohesin and thedockerin).
5. Notes
The description provided in this chapter focuses on mutant versuswild-type cohesin modules. If there is a need to compare many dockerinmutants, the procedure can be reversed, taking into account that immobi-lized Xyn-Docs provide a reduced response compared to the CBM-Cohs(Barak et al., 2005), presumably due to steric interference of the xylanasewhich may restrict access to the dockerin. These two modules have

Indirect ELISA for Cohesin–Dockerin Interaction 427
Author's personal copy
significant differences in their sizes: the molecular weight of the xylanase isapproximately 44 versus 10 kDa of the dockerin. Alternatively, the differ-ence may be due to the nature of the dockerin, such as its hydrophobicityand tendency to aggregate.
6. Summary
The ELISA-based method, iELISA, described in this chapter is pro-posed to overcome several limitations of other methods used to determinebinding affinity. It was optimized for the high-affinity cohesin–dockerinsystem to measure the effect of different mutations on cohesins or forcomparison between affinities of different cohesins (e.g., from the samescaffoldin) toward the same dockerin, but can easily be adopted for measur-ing other high-affinity interactions.
REFERENCES
Adams, J. J., Webb, B. A., Spencer, H. L., and Smith, S. P. (2005). Structural characteriza-tion of type II dockerin module from the cellulosome of Clostridium thermocellum:Calcium-induced effects on conformation and target recognition. Biochemistry 44,2173–2182.
Barak, Y., Handelsman, T., Nakar, D., Mechaly, A., Lamed, R., Shoham, Y., andBayer, E. A. (2005). Matching fusion protein systems for affinity analysis of two inter-acting families of proteins: The cohesin-dockerin interaction. J. Mol. Recognit. 18,491–501.
Bayer, E. A., Kenig, R., and Lamed, R. (1983). Adherence of Clostridium thermocellum tocellulose. J. Bacteriol. 156, 818–827.
Carvalho, A. L., Dias, F. M., Prates, J. A., Nagy, T., Gilbert, H. J., Davies, G. J.,Ferreira, L. M., Romao, M. J., and Fontes, C. M. (2003). Cellulosome assembly revealedby the crystal structure of the cohesin-dockerin complex. Proc. Natl. Acad. Sci. USA 100,13809–13814.
Carvalho, A. L., Dias, F. M. V., Nagy, T., Prates, J. A. M., Proctor, M. R., Smith, N.,Bayer, E. A., Davies, G. J., Ferreira, L. M. A., Romao, M. J., Fontes, C. M. G. A., andGilbert, M. J. (2007). Evidence for a dual binding mode of dockerin modules to cohesins.Proc. Natl. Acad. Sci. USA 104, 3089–3094.
Fierobe, H. P., Pages, S., Belaich, A., Champ, S., Lexa, D., and Belaich, J. P. (1999).Cellulosome from Clostridium cellulolyticum: Molecular study of the dockerin/cohesininteraction. Biochemistry 38, 12822–12832.
Fierobe, H. P., Mechaly, A., Tardif, C., Belaich, A., Lamed, R., Shoham, Y., Belaich, J. P.,and Bayer, E. A. (2001). Design and production of active cellulosome chimeras. Selectiveincorporation of dockerin-containing enzymes into defined functional complexes.J. Biol. Chem. 276, 21257–21261.
Friguet, B., Chaffotte, A. F., Djavadi-Ohaniance, L., and Goldberg, M. E. (1985). Measure-ments of the true affinity constant in solution of antigen-antibody complexes by enzyme-linked immunosorbent assay. J. Immunol. Methods 77, 305–319.

428 Michal Slutzki et al.
Author's personal copy
Handelsman, T., Barak, Y., Nakar, D., Mechaly, A., Lamed, R., Shoham, Y., andBayer, E. A. (2004). Cohesin-dockerin interaction in cellulosome assembly: A singleAsp-to-Asn mutation disrupts high-affinity cohesin-dockerin binding. FEBS Lett. 572,195–200.
Lapidot, A., Mechaly, A., and Shoham, Y. (1996). Overexpression and single-step purifica-tion of a thermostable xylanase from Bacillus stearothermophilus T-6. J. Biotechnol. 51,259–264.
Lytle, B. L., Volkman, B. F., Westler, W. M., Heckman, M. P., and Wu, J. H. D. (2001).Solution structure of a type I dockerin domain, a novel prokaryotic, extracellularcalcium-binding domain. J. Mol. Biol. 307, 745–753.
Mechaly, A., Yaron, S., Lamed, R., Fierobe, H.-P., Belaich, A., Belaich, J.-P., Shoham, Y.,and Bayer, E. A. (2000). Cohesin-dockerin recognition in cellulosome assembly: Exper-iment versus hypothesis. Proteins 39, 170–177.
Mechaly, A., Fierobe, H. P., Belaich, A., Belaich, J. P., Lamed, R., Shoham, Y., andBayer, E. A. (2001). Cohesin-dockerin interaction in cellulosome assembly: A singlehydroxyl group of a dockerin domain distinguishes between nonrecognition and highaffinity recognition. J. Biol. Chem. 276, 9883–9888.
Miras, I., Schaeffer, F., Beguin, P., and Alzari, P. M. (2002). Mapping by site-directedmutagenesis of the region responsible for cohesin-dockerin interaction on the surfaceof the seventh cohesin domain of Clostridium thermocellum CipA. Biochemistry 41,2115–2119.
Morag, E., Lapidot, A., Govorko, D., Lamed, R., Wilchek, M., Bayer, E. A., andShoham, Y. (1995). Expression, purification and characterization of the cellulose-bindingdomain of the scaffoldin subunit from the cellulosome of Clostridium thermocellum. Appl.Environ. Microbiol. 61, 1980–1986.
Motulsky, H. J., and Christopoulos, A. (2003). Fitting Models to Biological Data UsingLinear and Nonlinear Regression. A Practical Guide to Curve Fitting. GraphPad Soft-ware Inc., San Diego, CA, www.graphpad.com.
Nakar, D., Handelsman, T., Shoham, Y., Fierobe, H.-P., Belaich, J. P., Morag, E.,Lamed, R., and Bayer, E. A. (2004). Pinpoint mapping of recognition residues on thecohesin surface by progressive homologue swapping. J. Biol. Chem. 279, 42881–42888.
Pinheiro, B. A., Protctor, M. R., Martinez-Fleites, C., Prates, J. A., Money, V. A.,Davies, G. J., Bayer, E. A., Fontesm, C. M., Fierobe, H. P., and Gilbert, H. J. (2008).The Clostridium cellulolyticum dockerin displays a dual binding mode for its cohesinpartner. J. Biol. Chem. 283, 18422–18430.
Velazquez-Campoy, A., and Freire, E. (2006). Isothermal titration calorimetry to determineassociation constants for high-affinity ligands. Nat. Protoc. 1, 186–191.
Velazquez-Campoy, A., Ohtaka, H., Nezami, A., Muzammil, S., and Freire, E. (2004).Isothermal titration calorimetry. Curr. Protoc. Cell Biol. 17.8, 1–24.