increased expression of protease-activated receptor 1 (par1) in human leukemias
TRANSCRIPT

Justin R. Hamilton, Albert G. Frauman and Thomas M. CocksPAR2 and PAR4 Agonists
toCoronary Artery by Inflammatory Stimuli Unveils Endothelium-Dependent Relaxations Increased Expression of Protease-Activated Receptor-2 (PAR2) and PAR4 in Human
Print ISSN: 0009-7330. Online ISSN: 1524-4571 Copyright © 2001 American Heart Association, Inc. All rights reserved.is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation Research
doi: 10.1161/hh1301.0926612001;89:92-98; originally published online June 21, 2001;Circ Res.
http://circres.ahajournals.org/content/89/1/92World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circres.ahajournals.org//subscriptions/
is online at: Circulation Research Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer about this process is available in the
located, click Request Permissions in the middle column of the Web page under Services. Further informationEditorial Office. Once the online version of the published article for which permission is being requested is
can be obtained via RightsLink, a service of the Copyright Clearance Center, not theCirculation Researchin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from

Increased Expression of Protease-Activated Receptor-2(PAR2) and PAR4 in Human Coronary Artery by
Inflammatory Stimuli Unveils Endothelium-DependentRelaxations to PAR2 and PAR4 Agonists
Justin R. Hamilton, Albert G. Frauman, Thomas M. Cocks
Abstract—Protease-activated receptor (PAR)1 and PAR2 are expressed on vascular endothelial cells and mediateendothelium-dependent relaxation in several species, and PAR4 agonists cause similar responses in rat aortas. To date,only PAR1 has been reported to mediate relaxation of human arteries despite endothelial cell expression of both PAR1and PAR2 in these tissues. Because inflammatory stimuli increase PAR2 expression in human endothelial cells inculture, the present study investigated the effect of similar stimuli on PARs in human isolated coronary arteries (HCAs).In HCA ring segments suspended for isometric tension measurements, the selective PAR1-activating peptide, TFLLR(0.01 to 10mmol/L), caused endothelium-dependent relaxation of precontracted preparations. Little or no change invascular tension was elicited by either the PAR2- or PAR4-activating peptides, SLIGKV and GYPGQV, respectively(up to 100mmol/L). Exposure of HCAs to interleukin (IL)-1a (1 ng/mL, 12 hours) or tumor necrosis factor-a (3 nmol/L,12 hours) did not affect PAR1 expression but increased PAR2 and PAR4 mRNA levels by'5- and 4-fold, respectively,as determined by quantitative polymerase chain reaction. Similar IL-1a treatment did not affect TFLLR-inducedrelaxations but revealed significant endothelium-dependent relaxations to SLIGKV (100mmol/L, 61.466.7%) andGYPGQV (100mmol/L, 34.866.4%). These studies are the first to demonstrate functional PAR2 and PAR4 in humanarteries in situ. The selective upregulation of PAR2 and PAR4 expression and the increased vascular response in HCAsafter exposure to inflammatory stimuli suggest a role for these endothelial receptors during inflammation.(Circ Res.2001;89:92-98.)
Key Words: endotheliumn human coronary arteriesn protease-activated receptors
Protease-activated receptors (PARs) located on vascularsmooth muscle and/or endothelial cells have been impli-
cated in the control of vascular tone and hemostasis, partic-ularly during inflammatory states (see reviews1–3). PARs areG-protein–coupled receptors activated by particular serineproteases via a unique mechanism as follows: site-specificproteolysis of the amino-terminal exodomain of the receptorexposes a neo-N-terminal, which acts as a self-activating“tethered” ligand.4,5 Four PARs have been cloned. PAR14 andPAR36 are essentially thrombin receptors. PAR27 is activatedmost sensitively by trypsin and can be activated to a lesserextent by blood-borne enzymes, such as mast cell tryptase8,9
and coagulation factor VIIa.10 The known enzyme activatorsof PAR4 include thrombin, trypsin, and neutrophil-derivedcathepsin G.11,12 Intriguingly, at least for mouse receptorsexpressed in COS-7 cells, activation of PAR4 by thrombinrequires PAR3 as a thrombin-binding cofactor.13 Becauseeach cloned PAR is activated by trypsin (PAR2) or boththrombin and trypsin (PAR1, PAR3, and PAR4), it is fortu-
nate that PARs, with the exception of PAR3,6 can beselectively activated by synthetic peptides corresponding tothe distinct tethered ligand sequences of each receptor.4,6,7,12
Emerging literature suggests that endothelial PARs playimportant roles during vascular inflammation and tissueinjury.1,2,14–16PAR1 and PAR2 are present on smooth muscleand/or endothelial cells, where they mediate direct contrac-tion and NO-mediated endothelium-dependent relaxation,respectively, depending on the preparation, species, anddisease state of the tissue. For example, activation of PAR1(with thrombin or the PAR1-activating peptide, SFLLRN) orPAR2 (with trypsin or the PAR2-activating peptide, SLI-GRL) induces endothelium-dependent relaxation of the pigcoronary artery17 and rat aorta.18 However, in endothelium-denuded pig coronary arteries, none of these PAR agonistsalters vascular tension.17 By contrast, activators of PAR1, butnot PAR2, cause contraction of the endothelium-denuded rataorta.18 This differs again from the mouse renal artery, inwhich activators of PAR2 not only cause typical endotheli-
Original received April 10, 2000; resubmission received April 4, 2001; revised resubmission received May 10, 2001; accepted May 10, 2001.From the Department of Pharmacology (J.R.H., T.M.C.), University of Melbourne, Victoria, Australia, and the Clinical Pharmacology and Therapeutics
Unit (A.G.F.), Department of Medicine, Austin and Repatriation Medical Centre, Heidelberg, Victoria, Australia.Correspondence to Tom Cocks, PhD, Department of Pharmacology, University of Melbourne, Victoria 3010, Australia. E-mail
[email protected]© 2001 American Heart Association, Inc.
Circulation Researchis available at http://www.circresaha.org
92
Clinical Research
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from

um-dependent relaxation in endothelium-intact vessels butalso induce smooth muscle contraction of endothelium-denuded preparations.19 In addition to PAR1 and PAR2,functional PAR3 and PAR4 have been reported on endothe-lial cells of rat brain capillaries and rat aorta, respectively.20,21
Although our insight into the roles of vascular PARs inanimal models is progressing, little is known about PARs inhuman vasculature. Endothelial localization of PAR1 mRNAvia in situ hybridization has been described in nonatheroscle-rotic hepatic arteries and in both endothelial and smoothmuscle cells of atherosclerotic arteries.22 This finding issupported by functional studies on human isolated coronaryarteries (HCAs), which demonstrated PAR1-mediated endo-thelium-dependent relaxation of nonatherosclerotic arteriesand a lack of contraction to PAR1 agonists in artery prepa-rations with or without functional endothelium23,24 but anincreased contraction associated with an increased degree ofatheroma in endothelium-disrupted artery segments.23 ForPAR2, immunoreactivity has been reported in the endotheli-um and smooth muscle of HCAs.25 Functionally, PAR2-activating peptides induce calcium fluxes in human umbilicalvein endothelial cells (HUVECs)8 and mitogenic responses inhuman aortic smooth muscle cells in culture.26 However,PAR2 agonists cause neither contraction nor relaxation ofHCAs.24
We wondered whether the discrepancy between PAR2expression and the ability of PAR2 agonists to cause changesin human vascular tone is due to low receptor number.Because the inflammatory cytokines interleukin (IL)-1a andtumor necrosis factor (TNF)-a increase PAR2 expression inHUVECs27 and because similar inflammatory stimuli ame-liorate the hypotensive response to PAR2 activation in the ratin vivo,28 the present study investigated whether exposure ofHCAs to inflammatory stimuli, in vitro, would unveil anendothelium-dependent relaxation to PAR2 activation in thistissue. Also, because there are no reports of PAR3 or PAR4in human arteries in situ, we investigated the presence ofthese receptors in HCAs.
The present study detected PAR1, PAR2, and PAR4mRNAs in HCAs by polymerase chain reaction (PCR) andlocalized immunoreactivities for PAR1 and PAR2 to HCAendothelial cells. Despite the expression of three PARs, onlyagonists of PAR1 induced changes in vascular tone in HCAsin the absence of inflammatory stimuli. Exposure of HCAs toeither IL-1a or TNF-a significantly increased the expressionof both PAR2 and PAR4, as determined by quantitative PCR,which unveiled endothelium-dependent relaxations in re-sponse to agonists of these receptors. The present studyprovides the first evidence of functional PAR2 and PAR4 inhuman arteries in situ and suggests a role for these endothelialcell receptors during vascular inflammation.
Materials and MethodsMaterialsPAR1 and PAR2 antisera were gifts from Prof Lawrence Brass(University of Pennsylvania, Philadelphia). Anti-mouse antiserumand FITC-streptavidin were from Amersham Pharmacia Biotech;U46619 and nifedipine, from Sapphire Bioscience; (2)-isoprenaline,substance P (acetate salt), anda-thrombin (bovine serum), from
Sigma Chemical Co; IL-1a and TNF-a (both recombinant, human),from Endogen; and trypsin (bovine pancreas), from WorthingtonBiochemical. TFLLR, SLIGKV, and GYPGQV were synthesizedwith carboxyl-terminal amidation and purified to 95% via high-per-formance liquid chromatography by Auspep.
Human Coronary ArteriesWith informed patient consent and ethics approval from The AlfredHospital and University of Melbourne Ethics Committees, epicardialcoronary arteries were dissected from explanted hearts of 12 patients(9 men and 3 women, aged 45.365.1 years, range 28 to 61 years)undergoing transplantation at The Alfred Hospital (Melbourne,Australia) because of dilated congestive cardiomyopathy (n56),restrictive cardiomyopathy (n53), ischemic heart disease (n52), orseptal defects (n51). Three-millimeter-long arterial ring segmentsthat were macroscopically free of atheromatous plaques (endotheli-um was mechanically removed in some) were prepared and (1)frozen in liquid nitrogen and stored at270°C for RNA extraction,(2) fixed in 4% paraformaldehyde (4°C overnight) for immunohis-tochemistry, (3) incubated at 37°C/5% CO2 for 12 hours (CO2
incubator, Forma Scientific) in 5 mL Krebs solution in the absenceor presence of IL-1a (1 ng/mL) or TNF-a (3 nmol/L), or (4)immediately mounted in organ chambers for isometric tensionmeasurement.
Polymerase Chain ReactionTotal RNA was isolated and subjected to DNase digestion by usingthe RNeasy Mini Kit (Qiagen). cDNA synthesis was performed at42°C for 1 hour with 100 ng total RNA, 1mmol/L poly(dT)12–16
primers, and 200 U Moloney murine leukemia virus reverse tran-scriptase with use of the First Strand Synthesis Kit (Promega). For“conventional” PCR, cDNA was amplified by a “touchdown” PCRmethod29 to detect transcripts under high-stringency hybridizationconditions, with the use of 5 ng cDNA, 0.5mmol/L specific primers(Table 1), 0.2 mmol/L dNTPs, 2.5 U Taq DNA polymerase,10 mmol/L Tris HCl (pH 8.3), 50 mmol/L KCl, and 1.5 mmol/LMgCl2 and with heating to 94°C for 3 minutes, followed by 5 cyclesof 94°C, 70°C, and 72°C (each for 30 seconds) and then 35 cycles of94°C, 55°C, and 72°C (each for 30 seconds), and, finally, 72°C for5 minutes. Quantitative PCR was performed with an ABI Prism 7700Sequence Detection System (Applied Biosystems) according to themanufacturer’s instructions. Specific primer sequences were used incombination with internal probes, each labeled at the 59end with thereporter dye 6-carboxyfluorescein and at the 39end with thequencher dye 6-carboxytetramethylrhodamine (Table 2). Amplifica-tion was performed for 40 cycles of 95°C for 15 seconds and 60°Cfor 60 seconds.
TABLE 1. Gene-Specific Primer Sequences Used inConventional PCR Amplifications
GenePrimer Sequence*
(59–39)PCR Product
Size, bp
PAR1 Sense 2422TGTGAACTGATCATGTTTATG2442 708
Antisense 3129TTCGTAAGATAAGAGATATGT3109
PAR2 Sense 44GCAGGTGAGAGGCTGACTTT63 334
Antisense 377CAGTCGGTTCCGGTCTAACCGG356
PAR3 Sense 147GAAAGCCCTCATCTTTGCAG166 599
Antisense 745AGGTGAAAGGATGGACGATG726
PAR4 Sense 1121GGCAACCTCTATGGTGCCTA1140 244
Antisense 1364TTCGACCCAGTACAGCCTTC1345
ecNOS Sense 1152ACCCTCACCGCTACAACATC1171 744
Antisense 1895AGATGCTGTTGAAGCGGATC1876
*Numbers refer to base positions on the known human gene sequence.
Hamilton et al PAR2 and PAR4 Upregulation in Human Coronary Artery 93
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from

ImmunohistochemistryParaformaldehyde-fixed HCA was washed in PBS, placed in sucrosesolution (15% [wt/vol] in PBS) at 4°C overnight, embedded in OCTcompound (Sakura Finetek), and frozen in liquid nitrogen. PAR1 andPAR2 were localized by using previously characterized monoclonalantibodies produced in mice immunized with peptides corresponding tothe amino-terminus of human PAR1 (S42FLLRNPNDKYEPF55, 1:100)or human PAR2 (S37LIGKVDGTSHVTG50, 1:20) (see Molino et al8).Primary antisera were labeled with a biotinylated sheep anti-mouseantiserum (1:100, 1 hour) and FITC-conjugated streptavidin (1:20, 1hour).
Isometric Tension MeasurementsHCAs were mounted for isometric tension measurement as previ-ously described.24 After a standard equilibration procedure,24 includ-ing treatment with nifedipine (0.3mmol/L) to inhibit characteristicspontaneous contractile activity,30 HCAs were contracted to'50%of their maximum contraction to 125 mmol/L KCl with U46619, andcumulative half-log increases in the concentration of PAR agonistswere added. The maximum endothelium-dependent and-independent relaxations were determined by adding substance P (3nmol/L) and isoprenaline (1 mmol/L), respectively.
Statistical AnalysisChanges in isometric tension were normalized as percent tissueresponse to 1mmol/L isoprenaline. pEC50 values represent thenegative logarithm of the molar concentration that caused 50% of themaximum response. mRNA levels are expressed as fold increase inPAR mRNA normalized against 18S rRNA as the within-tissuecontrol. All data are mean6SEM. Differences in mean relaxationsand mRNA levels were tested for significance (P,0.05) byANOVA, with post hoc analyses for multiple comparisons.
ResultsPolymerase Chain ReactionConventional PCR analyses on mRNA extracted from endo-thelium-intact HCAs resulted in amplification of a singlefragment of predicted size (Table 1) for PAR1, PAR2, andPAR4 (n54 artery segments, from four patients) (Figure 1).By contrast, no PCR fragment was detected for PAR3 in thesetissues (n54 artery segments, from four patients) (Figure 1).When human cardiac muscle was used as a positive controlfor PAR3 expression,6 a single PCR product of predicted sizewas detected (n52 artery segments, from two patients)(Figure 1).
In similar PCR analyses on mRNA extracted from HCApreparations from which the endothelium had been removed,we again observed the expected PCR products for PAR1 andPAR4 (n52 artery segments, from two patients) (Figure 1).By contrast, the PAR2 PCR fragment detected in endotheli-um-intact HCAs was no longer evident (n52 artery segments,from two patients) (Figure 1). We confirmed the effectiveremoval of endothelial cells in these “endothelium-denuded”HCA segments by using specific primers for endothelialconstitutive NO synthase (ecNOS), which amplified a prod-uct of the expected size in endothelium-intact HCA segmentsbut not in endothelium-denuded HCA segments (n52 arterysegments, from two patients) (Figure 1).
Because PAR1, PAR2, and PAR4 are expressed in thevasculature, it was difficult to obtain negative tissue controlsfor the expression of these receptors. Thus, PCR products forPAR1, PAR2, and PAR4 were detected in vascularizedhuman tissues, such as testis, thyroid, and muscle of cardiac,ocular, and abdominal origin (data not shown). However, noproducts were observed if cDNA from rat heart was used inthe PCR reaction (data not shown). The possibility of con-tamination by genomic DNA was unlikely because isolatedRNA was subjected to DNase treatment before transcription.Furthermore, no PCR products were detected when PCR wasperformed on untranscribed RNA (data not shown).
TABLE 2. Gene-Specific Primer Sequences and InternalProbes Used in Quantitative Real-Time PCR Amplifications
GenePrimer/Probe Sequence*
(59–39)
PAR1 Sense 2379AGTAGGCTATTCCTGAGAGCTGCAT2403
Antisense 2454ATGGCCCTGGCATGTGTCT2436
Probe† 2405TGTCCGCCCCCGATGGAGG2423
PAR2 Sense 43GGCAGGTGAGAGGCTGACTT62
Antisense 107CGCCACCGATTCGAAACT90
Probe† 64CTCTCGGTGCGTCCAGTGGAGCTCT88
PAR4 Sense 1194GGGTCCCTTCCCCCACTT1211
Antisense 1216GACAGTTGTAACAACCCTATTTCCAAA1242
Probe† 1272TCCTTCCTGGGACCTCAGAATGTGACC1246
*Numbers refer to base positions on the known human gene sequence.†PAR probes were labeled at the 59 end with 6-carboxyfluorescein and at
the 39 end with 6-carboxytetramethylrhodamine.
Figure 1. Shade-inverted digitized photographs ofUV-illuminated agarose gels (1.5%, plus 0.5 mg/mL ethidiumbromide) showing molecular weight markers (M) and PCR prod-ucts for PAR1, PAR2, PAR3 (absent), PAR4, and ecNOS in en-dothelium-intact and endothelium-denuded HCAs (A) andmolecular weight markers (M) and PCR products for PAR3 andPAR4 in human prostate and cardiac muscle (B). Numbers rep-resent product sizes (bp).
94 Circulation Research July 6, 2001
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from
ImmunohistochemistryPunctate PAR1 and PAR2 immunoreactivities were localizedto the endothelium with little, if any, staining with eitherantisera detectable in the smooth muscle (Figure 2).
Quantitative PCRTreatment of endothelium-intact HCA segments with IL-1a(1 ng/mL, 12 hours) caused a significant (P,0.05)'5- and4-fold increase in mRNA expression for PAR2 and PAR4,respectively, as measured by real-time quantitative PCR(n53 artery segments, from three patients) (Figure 3). By
contrast, mRNA levels for PAR1 were virtually unaffected('1.3-fold increase) by similar treatment with IL-1a (n53artery segments, from three patients) (Figure 3). Treatment ofHCAs with TNF-a (3 nmol/L) caused a similar increase inPAR2 ('4-fold) and PAR4 ('3-fold) mRNA levels but,again, had little effect on PAR1 expression ('1.3-foldincrease in mRNA; n53 artery segments, from three patients)(Figure 3). The effects of IL-1a and TNF-a were likelyspecific because boiling these cytokines before treatment ofHCAs abolished their effect (data not shown).
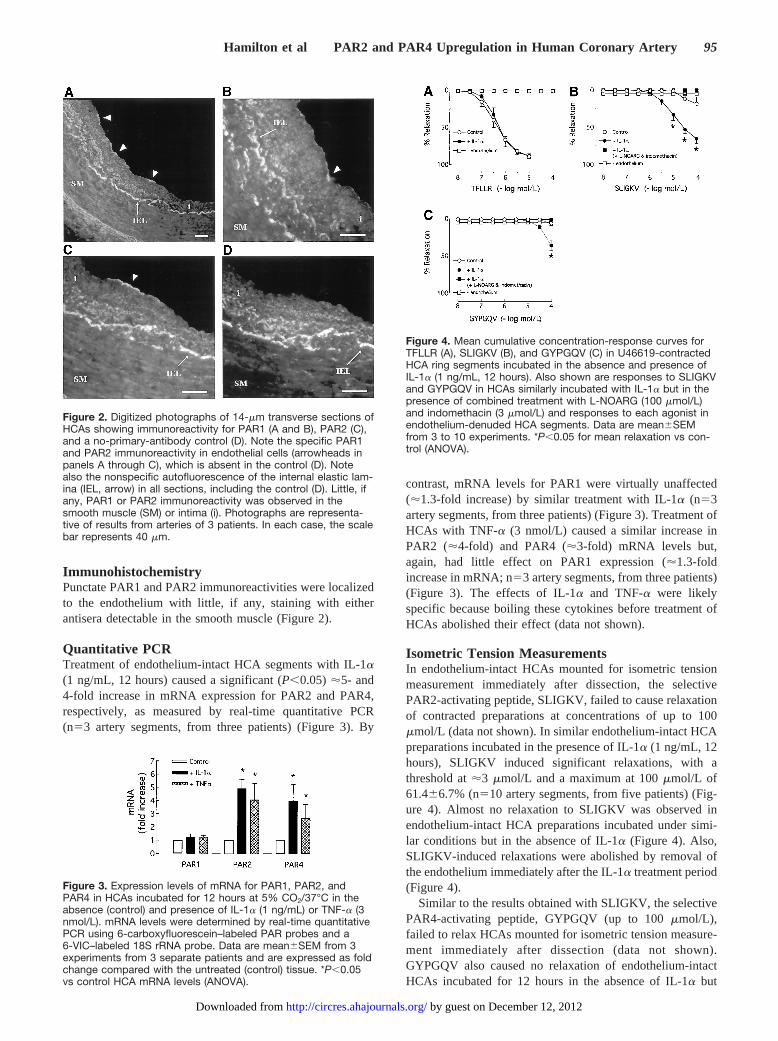
Isometric Tension MeasurementsIn endothelium-intact HCAs mounted for isometric tensionmeasurement immediately after dissection, the selectivePAR2-activating peptide, SLIGKV, failed to cause relaxationof contracted preparations at concentrations of up to 100mmol/L (data not shown). In similar endothelium-intact HCApreparations incubated in the presence of IL-1a (1 ng/mL, 12hours), SLIGKV induced significant relaxations, with athreshold at'3 mmol/L and a maximum at 100mmol/L of61.466.7% (n510 artery segments, from five patients) (Fig-ure 4). Almost no relaxation to SLIGKV was observed inendothelium-intact HCA preparations incubated under simi-lar conditions but in the absence of IL-1a (Figure 4). Also,SLIGKV-induced relaxations were abolished by removal ofthe endothelium immediately after the IL-1a treatment period(Figure 4).
Similar to the results obtained with SLIGKV, the selectivePAR4-activating peptide, GYPGQV (up to 100mmol/L),failed to relax HCAs mounted for isometric tension measure-ment immediately after dissection (data not shown).GYPGQV also caused no relaxation of endothelium-intactHCAs incubated for 12 hours in the absence of IL-1a but
Figure 2. Digitized photographs of 14-mm transverse sections ofHCAs showing immunoreactivity for PAR1 (A and B), PAR2 (C),and a no-primary-antibody control (D). Note the specific PAR1and PAR2 immunoreactivity in endothelial cells (arrowheads inpanels A through C), which is absent in the control (D). Notealso the nonspecific autofluorescence of the internal elastic lam-ina (IEL, arrow) in all sections, including the control (D). Little, ifany, PAR1 or PAR2 immunoreactivity was observed in thesmooth muscle (SM) or intima (i). Photographs are representa-tive of results from arteries of 3 patients. In each case, the scalebar represents 40 mm.
Figure 3. Expression levels of mRNA for PAR1, PAR2, andPAR4 in HCAs incubated for 12 hours at 5% CO2/37°C in theabsence (control) and presence of IL-1a (1 ng/mL) or TNF-a (3nmol/L). mRNA levels were determined by real-time quantitativePCR using 6-carboxyfluorescein–labeled PAR probes and a6-VIC–labeled 18S rRNA probe. Data are mean6SEM from 3experiments from 3 separate patients and are expressed as foldchange compared with the untreated (control) tissue. *P,0.05vs control HCA mRNA levels (ANOVA).
Figure 4. Mean cumulative concentration-response curves forTFLLR (A), SLIGKV (B), and GYPGQV (C) in U46619-contractedHCA ring segments incubated in the absence and presence ofIL-1a (1 ng/mL, 12 hours). Also shown are responses to SLIGKVand GYPGQV in HCAs similarly incubated with IL-1a but in thepresence of combined treatment with L-NOARG (100 mmol/L)and indomethacin (3 mmol/L) and responses to each agonist inendothelium-denuded HCA segments. Data are mean6SEMfrom 3 to 10 experiments. *P,0.05 for mean relaxation vs con-trol (ANOVA).
Hamilton et al PAR2 and PAR4 Upregulation in Human Coronary Artery 95
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from

caused significant relaxations in tissues incubated under thesame conditions but in the presence of IL-1a (1 ng/mL,34.866.4% at 100mmol/L; n54 artery segments, from twopatients) (Figure 4). GYPGQV-induced relaxations wereabolished by removal of the endothelium after the incubationperiod (Figure 4).
In contrast to SLIGKV- and GYPGQV-induced responses,relaxations caused by the selective PAR1-activating peptide,TFLLR, were not different between endothelium-intactHCAs incubated in the absence (pEC50 6.460.2, maximalrelaxation 89.062.6%) and presence (pEC50 6.360.1, maxi-mal relaxation 87.862.5%) of IL-1a (n58 artery segments,from four patients) (Figure 4).
A similar upregulation of endothelium-dependent relax-ations to SLIGKV and GYPGQV, but not TFLLR, wasobserved after treatment of HCAs with TNF-a (3 nmol/L)(n54 artery segments, from two patients) (data not shown).
Relaxations to both SLIGKV and GYPGQV in IL-1a–treatedHCAs were abolished by combined treatment with the ecNOSinhibitor,NG-nitro-L-arginine (L-NOARG, 100mmol/L), and thecyclooxygenase inhibitor, indomethacin (3mmol/L) (n54 arterysegments, from two patients) (Figure 4). Also, TFLLR,SLIGKV, and GYPGQV all failed to cause relaxation ofendothelium-denuded HCAs incubated with either IL-1a orTNF-a (data not shown).
Finally, TFLLR, SLIGKV, and GYPGQV caused no sig-nificant contraction under any of the conditions investigatedin the present study. Thus, none of the PAR-activatingpeptides, at concentrations of up to 100mmol/L, contractedendothelium-intact or -denuded HCAs incubated with orwithout IL-1a or TNF-a, either in the absence or presence ofU46619-induced tone (data not shown). Furthermore, thefailure of TFLLR, SLIGKV, and GYPGQV to contract HCAspersisted in the absence of nifedipine (data not shown).
DiscussionThe main finding of the present study was that exposure ofHCAs to inflammatory stimuli selectively increased PAR2and PAR4 expression, which unveiled endothelium-dependent relaxations to agonists of these receptors. To ourknowledge, the present study is the first to report functionalPAR2 and PAR4 in human arteries and, indeed, is the first todescribe the presence of PAR4 in human vasculature in situ.
We previously reported endothelium-dependent relaxationof HCAs by thrombin and the PAR1-activating peptide,SFLLRN.24 The present study first confirmed that the moreselective PAR1-activating peptide, TFLLR,31 caused similarrelaxation of HCAs. By contrast with TFLLR, the selectivePAR2-activating peptide, SLIGKV, failed to relax contractedHCAs, a finding that we have previously reported.24 Despitethis, two findings from our present study indicated that PAR2was expressed on HCA endothelium. First, PAR2 immuno-reactivity was localized specifically to the endothelial celllayer. Second, PAR2 mRNA was readily detected in endo-thelium-intact HCAs but not in segments of artery fromwhich the endothelium had been removed. Importantly, weused PCR for ecNOS, the expression of which is limited toendothelial cells in the vasculature,32 to confirm the absenceof contaminating endothelium in HCA segments from which
the endothelium was presumed to have been effectivelyremoved. Our finding that PAR2 is expressed only in theendothelium of HCAs differs from the study of Molino et al,25
who reported PAR2 immunoreactivity on smooth muscle aswell as endothelial cells in this tissue. The reason for thediscrepancy between the present study and that of Molino etal is unclear, although one possibility may involve differencesin the disease state of the tissue used in each study. Forexample, disease-related differences in receptor expressionhave been reported for PAR1. Thus, PAR1 expression waslimited to the endothelium of nonatherosclerotic arteries butwas observed in both endothelial and smooth muscle cells inatheromatous arteries.22 Regardless of these differences in thedetection of PAR2 expression on smooth muscle, the presentstudy and that of Molino et al have clearly shown that unlikethe expression in arteries of other species, the expression ofPAR2 on the endothelium of HCAs does not necessarilytranslate to PAR2-mediated endothelium-dependent relax-ation of this tissue.
In an attempt to reconcile the apparent discrepancy be-tween PAR2 expression and function on HCA endothelialcells, we sought to investigate the effects of inflammatorystimuli on this receptor. Previously, Nystedt et al27 showedthat treatment of HUVECs with IL-1a, TNF-a, or lipopoly-saccharide increased mRNA and cell-surface protein levelsfor PAR2, but not PAR1. In addition, Cicala et al28 reportedthat treatment of rats in vivo with lipopolysaccharide simi-larly increased PAR2 expression in the aorta and jugular vein,which was manifested as an increased sensitivity to thedilator response of PAR2 agonists in rat vasculature in vitroand in vivo. Again, this upregulation in response to aninflammatory stimulus was selective for PAR2 over PAR1.28
A similar selective upregulation of PAR2 mRNA in HCAstreated with either IL-1a or TNF-a was observed in thepresent study with the use of quantitative PCR. The 4- to5-fold increase in PAR2 mRNA observed in the present studyoccurred at the same concentrations of the two cytokines (3nmol/L and 1 ng/mL for TNF-a and IL-1a, respectively) andwithin the same time frame (12 hours) that gave an'3-foldincrease in PAR2 mRNA in the study of Nystedt et al.Although Nystedt et al found that 1 ng/mL IL-1a wassufficient to cause maximum increase in PAR2 mRNA('13-fold increase), this response occurred after 48 hours ofIL-1a treatment. Unfortunately, such an incubation periodwas not possible in the present study, inasmuch as prelimi-nary experiments determined that endothelium-dependentrelaxation of HCA ring preparations declined with incuba-tions beyond 12 hours. This is an important experimentallimitation of incubating isolated tissues, because we alsoobserved that a 10-fold increase in IL-1a concentration (to 10ng/mL) caused no greater induction of PAR2 mRNA expres-sion or SLIGKV-induced vascular relaxation (not shown).Therefore, the'60% relaxation caused by 100mmol/LSLIGKV appears to be the maximum response that we caninduce in this tissue with IL-1a. However, on the basis of thetime course studies of Nystedt et al in cultured HUVECs, itseems likely that prolonged (ie,.12-hour) exposure toinflammatory cytokines, such as that occurring during patho-
96 Circulation Research July 6, 2001
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from

physiological conditions in vivo, will further increase thesensitivity to PAR2 agonists.
A pattern of vascular reactivity similar to that observed forPAR2 was seen with PAR4. As was the case with SLIGKV,the selective PAR4-activating peptide, GYPGQV, failed tocause relaxation of contracted HCAs at concentrations of upto 100 mmol/L. As with PAR2, PAR4 mRNA was readilydetected in HCAs, although in contrast to PAR2, PAR4mRNA was also detected in endothelium-denuded HCAsegments. Because antibodies directed against PAR4 werenot available at the time of the present study, definitiveevidence for PAR4 expression on HCA endothelial cellscould not be provided. However, our finding (ie, that theupregulation of vascular relaxation to GYPGQV after expo-sure of HCAs to IL-1a or TNF-a was entirely dependent onthe presence of functional endothelium) suggests that likePAR2, PAR4 was expressed in HCA endothelial cells. To ourknowledge, this is the first demonstration of the in situlocalization and functional description of PAR4 in the humanvasculature. This result supports an earlier study in which anNO-mediated endothelium-dependent relaxation was ob-served in rat aortas in response to PAR4-activating peptides.21
Our hypothesis that relaxations of HCAs by agonists ofPAR2 or PAR4 are due to upregulation of these receptors onendothelial cells is supported by our observations that bothSLIGKV- and GYPGQV-induced relaxations were entirelydependent on an intact endothelium and were abolished in thepresence of inhibitors of the synthesis of NO (L-NOARG)and prostacyclin (indomethacin), the primary mediators ofendothelium-dependent relaxation in human arteries.30 Inaddition, the effects of IL-1a and TNF-a are likely specificbecause they were (1) blocked by boiling the cytokines beforethe treatment period, (2) concentration-dependent and oc-curred at the same concentration previously reported to causesimilar effects, (3) dependent on the presence of an intactendothelial cell layer, and (4) selective for SLIGKV andGYPGQV, inasmuch as relaxations to TFLLR, thrombin, andsubstance P were unaffected by similar treatment regimes(not shown).
Upregulation of endothelial cell PAR2 in response toinflammatory stimuli has been taken to indicate that PAR2 isinvolved in the progression of vascular inflammatory diseas-es.3 However, as we have previously pointed out,1 theopposite could apply. Thus, PAR2 (and PAR4) upregulationmay increase the protective barrier function of the endothe-lium and limit, for example, clotting and disseminatingintravascular coagulation in proinflammatory conditions,such as sepsis and hypertension. It is interesting to note thatin hypertension, although most other endothelium-dependentvasodilator function is diminished33 and although there is anincreased potential for thrombosis,34 PAR2-mediated vasodi-latation is preserved.35 Further evidence that endothelialPAR2 is anti-inflammatory, albeit indirect, comes from ourearlier observation in porcine isolated coronary arteries. Wefound that proinflammatory enzymes, such as bacterial ther-molysin, which disable PAR2 without causing receptor acti-vation, trigger a novel translation-dependent mechanism forreplenishment of the cell surface receptor, which is rapid andspecific for PAR2 over PAR1.17 Therefore, vascular endothe-
lial PAR2 (and possibly PAR4) is not only upregulated ininflammation, but novel mechanisms appear to exist tomaintain PAR2 responsiveness during such conditions.
The inability of agonists of PAR1, PAR2, or PAR4 toinduce contraction of HCAs under any of the conditionsinvestigated in the present study suggests that smooth musclecell PARs play a limited role in contracting human arteries.Although PAR1-mediated contractions occur in HCAs dis-playing signs of atheroma,23 smooth muscle cell PARs maybe involved in functions other than the regulation of vasculartone, such as smooth muscle growth and differentiation,36 inthe absence of atheroma.
Finally, the present study raises the possibility of a lack ofPAR3 in the human vasculature. The inability of syntheticPAR3-activating peptides to activate this receptor6 prohibitsus from investigating PAR3 function in this way. However,we were unable to detect PAR3 mRNA in HCAs. Further-more, desensitization to the PAR1-activating peptides,SFLLRN and TFLLR (which cannot activate PAR3 at theconcentrations used in the present study6,12), abolished sub-sequent responses to thrombin (the most potent knownactivator of PAR36) (J.R. Hamilton, T.M. Cocks, unpublisheddata, 2000). These findings contrast with those of previousreports that showed functional PAR3 in HUVECs via mea-surement of intracellular calcium concentration changes.37
The discrepancy between our results and those obtained inHUVECs remains unexplained but may simply reflect differ-ences between HUVEC and native human endothelial cells ordifferences in PAR3 expression between venous and arterialendothelial cells, such as that reported for other PARs.25
In conclusion, the present study is the first to demonstratefunctional PAR2 and PAR4 in human vasculature in situ. Wesuggest that under physiological conditions, circulatingthrombin activates PAR1 to relax underlying smooth muscle.In contrast with PAR1, endothelial PAR2 and PAR4 areexpressed in low levels and do not contribute to serineprotease regulation of vasodilator tone in the coronary bed.However, in pathological conditions in which inflammatorycytokines are released, such as ischemia/reperfusion injury,38
atherosclerosis,39 and, perhaps, hypertension,40 PAR2 andPAR4 are upregulated and may now respond to (patho)phys-iological concentrations of activating enzymes and contributeto endothelium-dependent vasodilatation. PAR2 is insensitiveto thrombin, and at least for the mouse receptor, PAR4 cannotbe activated by thrombin in the absence of PAR3.13 BecausePAR3 was not expressed in HCAs, PAR2 and PAR4 arelikely nonthrombin PARs in this tissue. On this assumption,we suggest that vascular inflammation promotes the switch-ing of endothelial cell PARs from being thrombin sensitiveunder physiological conditions to being thrombin insensitiveduring vascular inflammation. As such, the inflammatorystimuli–induced upregulation of PAR2 and PAR4 in HCAsmay indicate that these receptors are protective when ex-pressed on endothelial cells and that they regulate vasculartone in inflammation and/or vascular injury.
AcknowledgmentsThis study was supported by the National Health and MedicalResearch Council of Australia (Dr Cocks). We gratefully acknowl-
Hamilton et al PAR2 and PAR4 Upregulation in Human Coronary Artery 97
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from

edge the assistance of the Heart and Lung Transplant Unit at TheAlfred Hospital, Melbourne, Australia, for their generous assistancein obtaining the human tissue used in this study. The authors alsowish to thank Lerna Gulluyan for assistance with the quantitativePCR studies and Dr Alastair Stewart for providing the IL-1a andTNF-a.
References1. Cocks TM, Moffatt JD. Protease-activated receptors: sentries for inflam-
mation?Trends Pharmacol Sci. 2000;21:103–108.2. Coughlin SR. Thrombin signalling and protease-activated receptors.
Nature. 2000;407:258–264.3. Déry O, Corvera CU, Steinhoff M, Bunnett NW. Proteinase-activated
receptors: novel mechanisms of signaling by serine proteases.Am JPhysiol. 1998;274:C1429–C1452.
4. Vu T-KH, Hung DT, Wheaton VI, Coughlin SR. Molecular cloning of afunctional thrombin receptor reveals a novel proteolytic mechanism ofreceptor activation.Cell. 1991;64:1057–1068.
5. Chen J, Ishii M, Wang L, Ishii K, Coughlin SR. Thrombin receptoractivation: confirmation of the intramolecular tethered ligandinghypothesis and discovery of an alternative liganding mode.J Biol Chem.1994;269:16041–16045.
6. Ishihara H, Connolly AJ, Zeng D, Kahn ML, Zheng YW, Timmons C,Tram T, Coughlin SR. Protease-activated receptor 3 is a second thrombinreceptor in humans.Nature. 1997;386:502–506.
7. Böhm SK, Kong W, Brömme D, Smeekens SP, Anderson DC, ConnollyA, Kahn M, Nelken NA, Coughlin SR, Payan DG, Bunnett NW.Molecular cloning, expression and potential functions of the humanproteinase-activated receptor-2.Biochem J. 1996;314:1009–1016.
8. Molino M, Barnathan ES, Numerof R, Clark J, Dreyer M, Cumashi A,Hoxie JA, Schechter N, Woolkalis M, Brass LF. Interactions of mast celltryptase with thrombin receptors and PAR-2.J Biol Chem. 1997;272:4043–4049.
9. Fox MT, Harriott P, Walker B, Stone SR. Identification of potentialactivators of proteinase-activated receptor-2.FEBS Lett. 1997;417:267–269.
10. Camerer E, Huang W, Coughlin SR. Tissue factor- and factorX-dependent activation of protease-activated receptor 2 by factor VIIa.Proc Natl Acad Sci U S A. 2000;97:5255–5260.
11. Sambrano GR, Huang W, Faruqi T, Mahrus S, Craik C, Coughlin SR.Cathepsin G activates protease-activated receptor-4 in human platelets.J Biol Chem. 2000;275:6819–6823.
12. Xu W, Andersen H, Whitmore TE, Presnell SR, Yee DP, Ching A,Gilbert T, Davie EW, Foster DC. Cloning and characterization of humanprotease-activated receptor 4.Proc Nat Acad Sci U S A. 1998;95:6642–6646.
13. Nakanishi-Matsui M, Zheng Y-W, Sulciner DJ, Weiss EJ, Ludeman MJ,Coughlin SR. PAR3 is a cofactor for PAR4 activation by thrombin.Nature. 2000;404:609–613.
14. Damiano BP, D’Andrea MR, de Garavilla L, Cheung W-M, Andrade-Gordon P. Increased expression of protease-activated receptor-2 (PAR-2)in balloon-injured rat carotid artery.Thromb Haemost. 1999;81:808–814.
15. Vergnolle N. Proteinase-activated receptor-2-activating peptides induceleukocyte rolling, adhesion, and extravasation in vivo.J Immunol. 1999;163:5064–5069.
16. Lindner JR, Kahn ML, Coughlin SR, Sambrano GR, Schauble E,Bernstein D, Foy D, Hafezi-Moghadam A, Ley K. Delayed onset ofinflammation in protease-activated receptor-2-deficient mice.J Immunol.2000;165:6504–6510.
17. Hamilton JR, Chow JM, Cocks TM. Protease-activated receptor-2turnover stimulated independently of receptor activation in porcine cor-onary endothelial cells.Br J Pharmacol. 1999;127:617–622.
18. Magazine HI, King JM, Srivastava KD. Protease activated receptorsmodulate aortic vascular tone.Int J Cardiol. 1996;53:S75–S80.
19. Moffatt JD, Cocks TM. Endothelium-dependent and -independentresponses to protease-activated receptor-2 (PAR-2) activation in mouseisolated renal arteries.Br J Pharmacol. 1998;125:591–594.
20. Bartha K, Domotor E, Lanza F, Adam-Vizi V, Machovich M. Identifi-cation of thrombin receptors in rat brain capillary endothelial cells.J Cereb Blood Flow Metab. 2000;20:175–182.
21. Hollenberg MD, Saifeddine M, Al-Ani B, Gui Y. Proteinase-activatedreceptor 4 (PAR4): action of PAR4-activating peptides in vascular andgastric tissue and lack of cross-reactivity with PAR1 and PAR2.CanJ Physiol Pharmacol. 1999;77:458–464.
22. Nelken NA, Soifer SJ, O’Keefe J, Vu T-K, Charo IF, Coughlin SR.Thrombin receptor expression in normal and atherosclerotic humanarteries.J Clin Invest. 1992;90:1614–1621.
23. Ku DD, Dai J. Expression of thrombin receptors in human atheroscleroticcoronary arteries leads to an exaggerated vasoconstrictory response invitro. J Cardiovasc Res. 1997;30:649–657.
24. Hamilton JR, Nguyen PB, Cocks TM. Atypical protease-activatedreceptor mediates endothelium-dependent relaxation of human coronaryarteries.Circ Res. 1998;82:1306–1312.
25. Molino M, Raghunath PN, Kuo A, Ahuja M, Hoxie JA, Brass LF,Barnathan ES. Differential expression of functional protease-activatedreceptor-2 (PAR-2) in human vascular smooth muscle cells.ArteriosclerThromb Vasc Biol. 1998;18:825–832.
26. Bono F, Lamarche I, Herbert JM. Induction of vascular smooth musclecell growth by selective activation of the proteinase activated receptor-2.Biochem Biophys Res Commun. 1997;241:762–764.
27. Nystedt S, Ramakrishnan V, Sundelin J. The proteinase-activatedreceptor 2 is induced by inflammatory mediators in human endothelialcells.J Biol Chem. 1996;271:14910–14915.
28. Cicala C, Pinto A, Bucci M, Sorrentino R, Walker B, Harriot P, CruchleyA, Kapas S, Howells GL, Cirino G. Protease-activated receptor-2involvement in hypotension in normal and endotoxemic rats in vivo.Circulation. 1999;99:2590–2597.
29. Major BJ, Cures A, Frauman AG. The full length and splice variantthyrotropin receptor is expressed exclusively in skeletal muscle ofextraocular origin: a link to the pathogenesis of Graves’ ophthalmopathy.Biochem Biophys Res Commun. 1997;230:493–496.
30. Stork AP, Cocks TM. Pharmacological reactivity of human epicardialcoronary arteries: characterization of relaxation responses to endotheli-um-derived relaxing factor.Br J Pharmacol. 1994;113:1099–1104.
31. Kawabata A, Saifeddine M, Al-Ani B, Leblond L, Hollenberg MD.Evaluation of proteinase-activated receptor-1 (PAR1) agonists and antag-onists using a cultured cell receptor desensitization assay: activation ofPAR2 by PAR1-targeted ligands.J Pharmacol Exp Ther. 1999;288:358–370.
32. Pollock JS, Nakane M, Buttery LDK, Martinez A, Springall D, Polak JM,Förstermann U, Murad F. Characterization and localization of endothelialnitric oxide synthase using specific monoclonal antibodies.Am J Physiol.1993;265:C1379–C1387.
33. Vanhoutte PM, Boulanger CM. Endothelium-dependent responses inhypertension.Hypertens Res. 1995;18:87–98.
34. Tomiyama H, Kimura Y, Mitsuhashi H, Kinouchi T, Yoshida H, KushiroT, Doba N. Relationship between endothelial function and fibrinolysis inearly hypertension.Hypertension. 1998;31:321–327.
35. Sobey CG, Moffatt JD, Cocks TM. Evidence for selective effects ofchronic hypertension on cerebral artery vasodilatation to protease-acti-vated receptor-2 activation.Stroke. 1999;30:1933–1941.
36. Ahn H-S, Foster C, Boykow G, Stamford A, Manna M, Graziano M.Inhibition of cellular action of thrombin byN3-cyclopropyl-7-{[4-(1-methylethyl)phenyl]methyl}-7H-pyrrolo[3,2-f]quinazoline-1,3-diamine(SCH 79797), a nonpeptide thrombin receptor antagonist.BiochemPharmacol. 2000;60:1425–1434.
37. Schmidt VA, Nierman WC, Maglott DR, Cupit LD, Moskowitz KA,Wainer JA, Bahou WF. The human proteinase-activated receptor-3(PAR-3) gene: identification within a PAR gene cluster and character-ization in vascular endothelial cells and platelets.J Biol Chem. 1998;273:15061–15068.
38. Nose PS. Cytokines and reperfusion injury.J Card Surg. 1993;8:305–308.
39. Nilsson J. Cytokines and smooth muscle cells in atherosclerosis.Car-diovasc Res. 1993;27:1184–1190.
40. Voelkel NF, Tuder RM. Cellular and molecular mechanisms in thepathogenesis of severe pulmonary hypertension.Eur Respir J. 1995;8:2129–2138.
98 Circulation Research July 6, 2001
by guest on December 12, 2012http://circres.ahajournals.org/Downloaded from