in vitro effects of native human acute myelogenous leukemia blasts on fibroblasts and osteoblasts
TRANSCRIPT

IN VITRO EFFECTS OF NATIVE HUMAN ACUTE MYELOGENOUS LEUKEMIABLASTS ON FIBROBLASTS AND OSTEOBLASTSNils GLENJEN*, Elisabeth ERSVÆR, Anita RYNINGEN and Øystein BRUSERUD
Section for Hematology, Department of Medicine, Haukeland University Hospital and University of Bergen, Bergen, Norway
Bone marrow stromal cells constitute a heterogeneouspopulation, and in the present study we investigated inter-cellular crosstalk via release of soluble mediators betweennative human AML blasts and fibroblasts/osteoblasts. Cocul-ture of nonleukemic stromal cells and AML blasts separatedby a semipermeable membrane decreased proliferation ofthe fibroblast line HFL1, and the inhibition was maintainedwhen HFL1 and AML cells were cultured in direct contact. Asimilar inhibitory effect was observed for osteoblastic sar-coma cell lines (Cal72, SJSA-1) and normal osteoblasts. GM-CSF was released by both nonleukemic cells and a subset ofAML blast populations, and increased levels of GM-CSF weredetected in AML cocultures with fibroblasts and osteoblasticsarcoma cells when testing AML cell populations with con-stitutive GM-CSF release. Furthermore, constitutive IL-1�secretion by AML blasts was detected only for a subset ofpatients, whereas relatively high levels of IL-1RA were ob-served for all patients; coculture of AML blasts with HFL1fibroblasts and osteoblastic sarcoma cells increased IL-1�levels for patients with constitutive IL-1� secretion, whereasIL-1RA levels were slightly decreased but still generallyhigher than IL-1� levels (tested only for HFL1 fibroblasts).The bidirectional crosstalk between AML blasts and stromalcells with increased release of AML growth factors may beimportant in leukemogenesis, whereas the decreased stro-mal cell proliferation combined with the persistent release ofIL-1RA may in addition inhibit remaining normal hematopoi-esis and thereby contribute to bone marrow failure in AML.© 2004 Wiley-Liss, Inc.
Key words: acute myelogenous leukemia; fibroblast; osteoblast;granulocyte-macrophage colony-stimulating factor; IL-1
Bone marrow stromal cells constitute a very heterogeneouspopulation (i.e., including fibroblasts/reticular cells, adipocytes,endothelial cells, macrophages and osteogenic cells) that is impor-tant in normal as well as leukemic hematopoiesis, including theneoplastic proliferation of immature myeloid cells in AML.1–9
Firstly, normal myeloid cells as well as AML blasts expressadhesion molecules that bind to stromal elements. These moleculesinclude �1 (CD29) and �2 (CD18) integrins, the immunoglobulinsPECAM-1 (CD31) and LFA-3 (CD58), L-selectin and the proteo-glycan CD44.6,10–13 Secondly, stromal cells release a wide rangeof cytokines, including hematopoietic growth factors.5,6,14–16
Thirdly, stromal cells produce extracellular matrix, e.g., differentcollagens, proteoglycans (e.g., chondroitin sulfate, heparan sulfate,dermatan sulfate), hyaluronan and glycoproteins like fibronectin,laminin and thrombospondin.5,10,17 One function of these mole-cules is to bind growth factors and thereby create a local extracel-lular reservoir.17 The stromal cells together with their matrixcomponents thereby constitute an interacting network with AMLcells.11,18
The proliferative capacity of in vitro cultured AML blasts variesbetween patients: spontaneous or autocrine in vitro proliferation isdetected only for a subset of patients and associated with poorprognosis.19 Furthermore, only a subpopulation of the hierarchi-cally organized AML clone appears to proliferate during in vitroculture, whereas the majority of the cells often undergo spontane-ous in vitro apoptosis.20 Both the presence of hematopoieticgrowth factors alone as well as coculture with stromal cells (e.g.,fibroblast cell lines) can then increase AML blast proliferation andinhibit apoptosis.16,19–22 However, previous studies have not char-acterized in detail the effects of leukemia cells on various stromal
cells. AML-induced modulation of different stromal cells may beimportant for the regulation of both leukemic and remaining nor-mal hematopoiesis.23 We have therefore used standardized exper-imental in vitro models to compare the effects on fibroblasts andosteoblasts by native human AML cells derived from a large groupof consecutive patients. Bone marrow stromal cells are heteroge-neous, and even fibroblasts may differ in their functional pheno-type;14 for this reason, we investigated AML effects on 2 well-characterized fibroblast lines, 3 different osteosarcoma cell lines (2having an osteoblastic phenotype), normal osteoblasts and a het-erogeneous population of normal bone marrow stromal cells.
MATERIAL AND METHODS
Acute leukemia cellsThe clinical and biologic characteristics of 55 consecutive AML
patients are presented in Table I. Cells from 11 patients with ALLwere also examined; 10 patients had B-cell disease and one patient,T-cell disease (Table II).24 All patients had high peripheral bloodblast counts; leukemic PBMCs were therefore isolated by densitygradient separation alone (Ficoll-Hypaque, specific density 1.077;NyCoMed, Oslo, Norway). Cells were stored in liquid nitrogen.25
The percentage of leukemic blasts among PBMCs exceeded 95%for all patients as judged by light microscopy of May-Grunwald/Giemsa-stained cytospin smears.26–28
Nonleukemic cellsFibroblast cell lines. HFL1 cells (ATCC, Manassas, VA; ATCC
number CCL-153), derived from the lungs of a fetus (16–18 weeksold) have a diploid karyotype; the recommended medium is Ham’sF12K with 10% FCS. Hs27 cells (ATCC number CRL-1634),derived from the normal foreskin of a newborn, have a normalmale karyotype; the recommended culture medium is DMEM with10% FCS. Both lines were expanded in the recommended mediaand stored in frozen aliquots.25
Human osteosarcoma cell lines. Cal72 cells (DeutscheSammlung von Zellkulturen und Mikroorganismen, Braun-
Abbreviations: ALL, acute lymphoblastic leukemia; AML, acute my-elogenous leukemia; bFGF, basic fibroblast growth factor; FAB, French–American–British; GM-CSF, granulocyte-macrophage colony-stimulatingfactor; HGF, hepatocyte growth factor; IL-1RA, IL-1 receptor antagonist;MAb, monoclonal antibody; PBMC, peripheral blood mononuclear cell;SCF, stem cell factor; TNF, tumor necrosis factor; VEGF, vascular endo-thelial growth factor.
Grant sponsor: Norwegian Cancer Society; Grant sponsor: Rachel andOtto Christian Bruun’s Foundation; Grant sponsor: Olaf Runshaugen’sFoundation.
*Correspondence to: Department of Medicine, Haukeland UniversityHospital, N-5021 Bergen, Norway. Fax: �47-55-97-29-50.E-mail: [email protected]
Received 13 February 2004; Accepted after revision 15 March 2004
DOI 10.1002/ijc.20353Published online 20 May 2004 in Wiley InterScience (www.interscience.
wiley.com).
Int. J. Cancer: 111, 858–867 (2004)© 2004 Wiley-Liss, Inc.
Publication of the International Union Against Cancer

TABLE I – CLINICAL AND BIOLOGIC CHARACTERISTICS OF AML PATIENTS
Patient Sex Age (years)Previous malignant
or premalignantdisease
FABclassification
Membrane molecule1 expressionCytogenetic abnormality Flt3
abnormality2WBC
counts3CD13 CD14 CD15 CD33 CD34
1 M 29 AML-M4 � � � � � 46,XY ITD, PM 18.62 M 82 AML-M5 � � � � � 45X — 198.03 M 33 AML-M1 � � � � � 47,XY,�4/46,XY ITD 41.04 F 64 MDS AML-M1 � � � � � 46,XX,del(11)(q14)/46,XX, del
(11)(q14),del(20)(q11)/50, XX,del(11)(q14), �21,�21,�21,�21
— 15.5
5 F 61 AML-M4 � � � � � 46,XX — 29.26 F 51 AML-M2 � � � � � 46,XX ITD, wt� 154.07 F 75 AML-M5 � � � � � 46,XX ITD 104.08 F 45 AML-M4 � � � � � 46,XX PM 66.99 F 70 AML AML-M1 � � � � � nt ITD 142.010 M 75 AML-M0 � � nt nt � nt — 16.111 F 57 MDS AML-M2 � � � � � 47,XX,�21(4)/46, XX(11) — 69.212 F 63 AML-M1 � � � � � nt nt 75.513 F 74 AML-M2 nt nt nt nt nt nt — 28.314 M 36 CML AML-M2 � nt nt � � 46,XY,(9)(ins22q12;q34) — 202.015 M 43 AML-M2 � � � � � 46, XY — 90.416 F 79 AML-M5 � � � � nt 46,XX,del(12)(p11)/46,XX, del
(12)(p11),add(11)(p15)PM 201.0
17 M 82 AML-M1 � � nt � � nt — 80.018 M 79 AML-M1 � � � � � 46,XY ITD 5.619 F 78 AML-M4 � � � � � 47–49,XX,�4,�5,�der(8)
t(8;?)(q21;?),�20,�21— 92.6
20 M 74 MDS AML-M5 � � � � � nt ITD 73.421 F 59 AML-M2 � � � � � 45,XX,�7 — 41.922 F 63 AML-M4 � � � � � 46,XX ITD 85.723 M 56 AML-M2 � � � � � 46,XY ITD 50.524 M 65 CML AML-M2 � � � � � 47,XY,t(13;15)(q10;q10),�17,
�21?,�22?— 46.6
25 M 69 MDS AML-M1 � nt � � � nt — 1.026 M 74 AML-M0 � � � � � 90–94,XXYY — 123.027 M 33 AML-M5 � � 46,XY ITD 131.028 F 34 AML-M5 � � � � � 46XX,t(9;11)(p22;q23) PM 286.029 M 64 AML-M4 � � � � � 46,XY ITD, wt� 135.030 F 36 AML-M5 � � � � � 46,XX,t(9;11)(p21;q23) — 12.731 M 72 AML-M5 � � � � � 46,XY — 41.832 F 58 AML-M2 � � � � � 46,XX ITD, wt� 40.733 F 64 AML-M1 � � � � � 43,XX,der(5)t(5;?8)(q14;q21), �7,
�der(7)i(7)(p10)ins(7;11)(p11?;q23q?), der(8)t(8;?)(q21;?), der(11)t(5?;11)(q31;q25?), der(14)t(13?;14)(p10;p10)del(13)(q32?),�16,�17,�21
— 13.2
34 F 66 AML-M1 � � nt � nt nt — 59.635 M 56 AML-M4 � � � � � 46,Y — 10.836 F 55 AML-M0 � � � � � 46,X ITD 38.637 M 68 MDS AML-M2 � � nt � � 46,Y nt 146.038 F 54 Breast cancer AML-M1 � � nt � nt nt ITD 35.339 M 77 HD, MDS AML-M2 � � � � � del(7), �20 nt 61.640 F 45 Ovarian
carcinomaAML-M4 � � � � � 46,X — 69.9
41 M 48 AML-M5 � � nt � � 46,Y ITD 63.542 M 81 MDS AML-M4 � � nt � � nt — 60.543 F 55 MDS AML-M1 � � � � � 46,X,t(3;3)(q21;q26) PM 42.344 M 80 MDS AML-M4 � � nt � � 46,Y nt 10.545 M 83 AML-M2 � � nt � � nt — 49.346 M 79 AML-M4 � � � � � nt ITD 105.047 M 79 AML-M0 � � � � � nt — 60.548 F 45 AML-M2 � � � � � 46,X ITD, PM 123.549 M 40 MDS AML-M6 � � � � � 46,Y ITD 27.850 M 64 MDS AML-M1 � � � � � 37-46XY PM 11.951 M 69 AML-M2 � nt nt � � 46,XY,inv(16) — 67.252 M 43 AML-M5 � � � � � 46,XY,inv(16)(p13q22) PM 351.053 F 38 AML-M4 � � � � � 46,XX 182.054 F 44 AML-M1 � � � � � 46,XX,del(7)(q22) — 28.955 F 48 AML-M5 � � � � � 46,XX,t(9;11)(p22;q23(16)/47,
XX,t(9;11)p22;q23),�8(9)— 33.1
1Patients were regarded as positive when �20% of blast cells stained positive, as judged by flow-cytometric analysis.–2Flt3 abnormalities wereinternal tandem duplications (ITD), point mutations (PM) and loss of wild-type (wt�). nt, not tested.–3White blood cell (WBC) counts inperipheral blood are expressed as � 109/l (normal range 3.5–11.0 � 109/l). WBC included at least 80% leukemia blasts.
MDS, myelodysplastic syndrome; HD, Hodgkin’s disease; CML, chronic myelogenous leukemia.
859AML CELL EFFECTS ON FIBROBLASTS/OSTEOBLASTS

schweig, Germany) have been characterized previously and have aphenotype close to normal osteoblasts with an adherent growthpattern and a broad cytokine release profile.29,30 Pilot experimentsdemonstrated that this cell line released high levels of IL-6, VEGF,HGF and MCP-1. The SJSA-1 cell line (ATCC number CRL-2098) has a similar growth pattern and cytokine profile, whereasthe lines Saos-2 (ATCC number HTB-85), SK-ES-1 (ATTC num-ber HTB-86), U2OS (ATCC number HTB-96), 143.98.2 (ATCCnumber CCL-11226) and KHOS-32IH (ATCC numberCRL-1546) showed either an epithelial growth pattern or a differ-ent cytokine release profile.
Normal human osteoblasts and bone marrow stromal cells.These cells were delivered in frozen vials (Clonetics-BioWhit-taker, Walkersville, MA), stored in liquid nitrogen until thawedand used directly in the coculture assay. Both cell populationsshowed a purity of 95% and tested negative for mycoplasma,HIV-1, hepatitis B and hepatitis C (PCR). The culture medium inthese experiments was osteoblast growth medium containing 10%FCS (Clonetics-BioWhittaker).
Normal human osteoblasts were derived from a healthy 16-year-old male Caucasian. Cells were derived by explant, and charac-terization after in vitro redifferentiation showed positivity afterstaining for alkaline phosphatase and bone mineralization (vonKossa stain). Normal bone marrow stromal cells were derivedfrom a healthy 20-year-old female Caucasian. Bone marrow mono-nuclear cells were then derived by gradient separation (specificdensity 1.077) and cultured in Myelocult growth medium (Clonet-ics-BioWhittaker) for 4 weeks. Stromal cells represent the adher-ent cell population of these in vitro cultured cells and are aheterogeneous population of fibroblasts, reticular cells, endothelialcells, macrophages and fat cells.
Reagents
Culture media. The following culture media were used: Ham’sF12K medium, DMEM, McCoy’s medium (all from ATCC),RPMI-1640 (GIBCO, Paisley, UK), Stem Span SFEM (Stem SpanTechnologies, Vancouver, Canada), X-vivo 10, X-vivo 15 andX-vivo 20 (all from Clonetics-BioWhittaker). X-vivo and StemSpan media can be used for serum-free cell culture; the othermedia were supplemented with 10% heat-inactivated FCS (Clo-netics-BioWhittaker). Unless otherwise stated, we used Stem Spanmedium supplemented with 10% FCS and gentamicin 100 �g/mlbecause (i) pilot experiments demonstrated that FCS was requiredfor optimal growth of fibroblasts and osteosarcoma cell lines, (ii)all sarcoma cell lines and HFL1 fibroblasts showed strong prolif-eration in this medium (Hs27 fibroblasts showed 50% reduced
proliferation compared to the recommended medium) and (iii) themedium should be regarded as suitable for culture of native humanAML cells.31 Proliferation showed wide variation between celllines even when using this medium, the highest proliferation beingobserved for the 143.98.2 sarcoma cell line.
Soluble mediators. The following soluble mediators were usedat 50 ng/ml (unless otherwise stated, cytokines were from Prepro-tech, Rocky Hill, NJ): angiogenin (R&D Systems, Abingdon, UK),angiopoietin-2 (R&D Systems), angiostatin (Calbiochem, Darm-stadt, Germany), bFGF, endostatin (Calbiochem), Flt3 ligand(Flt3-L), G-CSF (Roche, Basel, Switzerland), GM-CSF (Sandoz,Basel, Switzerland), IFN-�, IL-1�, IL-1�, IL-2, IL-3, IL-4, IL-9,IL-10, IL-13, IL-15, IL-16, IL-17, M-CSF, SCF, TNF-�, VEGF,IL-1RA (R&D Systems).
Antibodies. The following cytokine-specific neutralizing anti-bodies (R&D Systems, neutralization data reported by distributor)were used: (i) antihuman GM-CSF: mouse IgG1 MAb 3209.01,0.3–0.5 �g/ml of this antibody neutralizing 50% of the biologicactivity of GM-CSF 0.5 ng/ml; (ii) antihuman HGF: mouse IgG1MAb 24612.111, 0.1–0.3 �g/ml neutralizing 50% of the biologicactivity of HGF 100 ng/ml; (iii) antihuman VEGF: mouse IgG2bMAb 26503, 0.04–0.08 �g/ml neutralizing 50% of the biologicactivity of VEGF 10 ng/ml; (iv) antihuman SCF: polyclonal goatantiserum, 0.05–0.1 �g/ml of this preparation neutralizing 50% ofthe biologic activity of SCF 10 ng/ml. Anti-GM-CSF and anti-HGF were used at 10 �g/ml; anti-SCF at 2 �g/ml and anti-VEGFwere used at 1 �g/ml. Control cultures were prepared using cor-responding normal mouse IgG1 (anti-GM-CSF, anti-HGF) andIgG2b (anti-VEGF) antibodies or normal goat antiserum (anti-SCF) (all supplied by R&D Systems) at the same concentrations asthe specific antibodies.
Tissue culture
Preparation of AML supernatants. AML blasts (2 � 106 cells in2 ml Stem Span/well) were cultured in 24-well tissue culture plates(Costar 3524; Costar, Cambridge, MA) for 48 hr before superna-tants were harvested
In vitro culture of nonleukemic cells alone. Frozen nonleukemiccells were precultured in the recommended medium in a 75 cm2
culture flask until they were confluent, then trypsinized and dis-tributed into flat-bottomed microtiter wells (104 cells/well culturedin 200 �l medium; Costar 3596 96-well culture plates). After 3days 37 kBq of 3H-thymidine (TRA 310; Amersham, Aylesbury,UK) were added in 20 �l saline, and nuclear radioactivity wasdetermined 18 hr later by liquid scintillation counting.
TABLE II – CLINICAL AND BIOLOGIC CHARACTERISTICS OF ALL PATIENTS
Patient Sex Age (years) Previous disease or chemotherapy ALL subclassification1 Cytogenetic analysis2 WBC count3
1 M 82 B-ALL nt 1252 F 23 Previous chemotherapy Pro-B-ALL nt (bcr/abl�) 47.13 M 24 Previous chemotherapy for
testicular carcinomaPro-B-ALL t(9;22) (bcr/abl�) 89
4 M 21 B-ALL 46XY, dic(7;9)(p11;p11) 15.25 F 28 T-ALL nt (bcr/abl�) 686 M 74 Common-B-ALL nt 787 F 54 Common-B-ALL t(9;22) (bcr/abl�) 5608 F 22 Common-B-ALL nt (bcr/abl�) 3.69 F 58 Pre-B-ALL nt 34
10 F 32 ALL relapse Pre-B-ALL nt 3511 F 14 ALL relapse Common-B-ALL nt 25
1ALL blasts were regarded as positive for membrane molecules when �20% of cells stained positive by flow-cytometric analysis.Classification was based on the guidelines given by the European Group for the Immunological Classification, of Acute Leukemias.18 Accordingto this classification, B-lineage ALL blasts are positive for at least 2 of the 3 markers CD19, CD22 and CD79a. Patients classified as pro-B-ALL(also referred to as B-I or null ALL) express no other B-cell differentiation antigens, common ALL (also referred to as c-ALL, pre-pre-B-ALLor B-II) express CD10, pre-B-ALL (B-III) express cytoplasmic lg and mature-B-ALL (also referred to as B-ALL or B-IV) express surfacemembrane Ig.–2Routine screening for chromosomal abnormalities was done by analysis of cells in mitosis. The abbreviation “nt” (not tested)means that cells were not available for testing or did not proliferate in vitro; for 3 of these patients, the presence of the bcr/abl translocation(Philadelphia chromosome) was analyzed by the FISH technique.–3Peripheral blood white blood cell (WBC) counts are expressed as � 109/l.
860 GLENJEN ET AL.

Coculture of AML and nonleukemic cells separated by a semi-permeable membrane. Cultures were prepared in Transwell cultureplates (Costar, Transwell 3401), where cells in the lower, largercompartment were separated from cells in the upper, smaller wellby a semipermeable membrane with pore size 0.4 �m. Nonleuke-mic cells were initially cultured for 3 days in the lower chamber(104 cells in 1 ml medium) before 106 native human AML blastswere added in 0.5 ml medium to the upper well. Cocultures wereincubated for 6 days before 3H-thymidine (280 kBq/well) wasadded; cultures were thereafter incubated for an additional 18 hrbefore AML cells were removed. All cultures were observed undera microscope before harvesting to ensure that cells/osteoblasts/fibroblasts did not form a confluent layer. Adherent cells were thenwashed in isotonic saline before 300 �l/well of trypsin-EDTAsolution (Stem Cell Technologies) were added and nuclear radio-activity was assayed in 70 �l aliquots.
Coculture of AML cells in direct contact with fibroblasts. HFL1and Hs27 fibroblasts were precultured for 3 days (104 cells/well in1 ml medium, Costar 3524 culture plates) before AML cells wereadded (2 � 106 leukemia cells in 0.5 ml medium). Cocultures wereincubated for 6 days before 3H-thymidine (280 kBq/well) wasadded. Nonadherent cells were removed 18 hr later; light micros-copy then demonstrated that the remaining cells formed a uniformlayer with a morphologic appearance consistent with fibroblastsand that they did not form a confluent layer in any culture.Adherent cells were trypsinized and nuclear radioactivity wasassayed as described above.
Analysis of GM-CSF, IL-1� and IL-1RA levels. GM-CSF, IL-1�and IL-1RA levels were determined by ELISA (Quantikine ELISAkit, R&D Systems) performed strictly according to the manufac-turer’s instruction. Standard curves were prepared as recom-mended, and differences between duplicates were generally 10%of the mean. Minimal detectable levels were as follows: GM-CSF,3 pg/ml; IL-1�, 2.8 pg/ml; and IL-1RA, 14 pg/ml.
Presentation of data3H-thymidine incorporation. The mean response of triplicate
determinations was used in all calculations and statistical compar-isons. Significant proliferation was defined as 3H-thymidine incor-poration exceeding 1,000 cpm and exceeding the mean counts perminute of the negative control by at least 3 SD. Incrementalresponses (stimulated response in cpm – negative control) wereused for all calculations. A significant alteration of proliferationwas defined as (i) a difference exceeding 2,000 cpm and (ii) thisdifference being �20% of the control.
GM-CSF and IL-1RA supernatant levels. A significant alterationin GM-CSF and IL-1RA supernatant levels was defined as adifference exceeding 20% of the control when paired samples werecompared. Incremental GM-CSF levels were defined as the levelsin cocultures minus the summarized levels for control culturescontaining corresponding AML cells and nonleukemic cells alone.
Statistical analysis. Wilcoxon’s test for paired samples and theMann-Whitney U-test/Wilcoxon test for 2 independent sampleswere used for comparison of continuous data. Spearman’s testwas used for correlation analysis. Differences were regarded asstatistically significant at p 0.05.
RESULTS
Native human AML cells inhibit proliferation of HFL1, but notHs27, fibroblasts
AML blasts were cultured together with HFL1 fibroblasts (Ta-ble I, patients 1–44) in transwell cultures prepared in Stem Spanmedium with FCS. HFL1 fibroblasts cultured alone showed rela-tively strong proliferation, which was decreased during coculturewith AML blasts (Fig. 1, n � 44, p � 0.007); this effect wasreproduced in repeated experiments for 11 patients. The number ofHFL1 fibroblasts was also counted after coculture with AMLblasts derived from 16 consecutive patients (Table I, patients 1, 3,7, 9, 10, 17–24, 26, 47, 54), and the reduced numbers of HFL1
fibroblasts after cocultures confirmed that the decreased 3H-thy-midine incorporation during coculture reflected a true reduction inthe number of fibroblasts (mean decrease 38%, Wilcoxon’s test forpaired samples p � 0.048). Decreased HFL1 proliferation was alsodetected in the presence of exogenous hematopoietic growth fac-tors (transwell cultures with GM-CSF � SCF � Flt3-L, 23 pa-tients examined; data not shown) and when AML cells and fibro-blasts were cultured in direct contact (Table I, patients 1–33;Wilcoxon’s test for paired samples p � 0.01).
HFL1 fibroblasts were also cultured in the recommended FCS-containing medium supplemented with 50 �l/well (final dilution1:4) of AML cell supernatants (Table I, patients; 1, 3–5, 7, 10,14–16, 19–25, 27–29). These selected patients represent a con-secutive group. Supernatants increased fibroblast proliferation sig-nificantly (Fig. 2, p 0.001). This enhancement was maintained inthe presence of exogenous angiogenin, angiopoietin-2, angiostatin,SCF and VEGF, whereas the enhancement was not detected in thepresence of exogenous IL-1�, bFGF and endostatin (data notshown). Thus, the AML-associated inhibition of HFL1 prolifera-tion during in vitro culture depends on the crosstalk between the 2cell populations.
The effects of AML blasts on Hs27 fibroblasts differed fromthose on HFL1 fibroblasts: (i) Hs27 cells showed relatively low invitro proliferation; (ii) proliferation was increased during coculturewith AML blasts in transwell cultures (Fig. 1, n � 33, p � 0.001);and (iii) proliferation of Hs27 cells was not altered during cocul-ture in direct contact with AML blasts (data not shown).
The effects of AML blasts on HFL1 and Hs27 fibroblast pro-liferation showed no correlation with peripheral blood counts(platelets, hemoglobin levels) at the time of diagnosis, signs ofAML cell differentiation (FAB classification and CD34 expres-sion) or cytogenetic or genetic Flt3 abnormalities of the AML cells(data not shown).
Native human AML blasts inhibit proliferation of osteoblasticsarcoma cells
Native AML blasts were cultured in transwell cultures togetherwith Cal72 (Table I, patients 1–10, 12, 13, 16–23, 26, 28, 29, 31,34–36, 38, 42, 43, 45–47, 49, 50–54), SJSA-1 and 143.98.2osteosarcoma cells (patients 1–10, 12–14, 16–26, 28–36, 38, 40,42, 43, 45–54). Proliferation of osteoblastic Cal72 (Fig. 1; Wil-coxon’s test for paired samples n � 39, p 0.001) and SJSA-1(Fig. 1; n � 47, p � 0.01) sarcoma cells was significantly de-creased during coculture. The effects on Cal72 proliferation werereproduced in repeated experiments for 11 patients, and the growthinhibition of osteoblastic sarcoma cells showed no association withkaryotype or Flt3 abnormalities in AML cells (data not shown).Numbers of Cal72 and SJSA-1 cells were also compared forAML-free cultures and cocultures with AML blasts derived froma consecutive subset of 15 patients (Table I, patients 1, 3, 7, 9, 10,17–23, 26, 47, 54), and decreased numbers of sarcoma cells werethen detected after coculture both for Cal72 (median decrease47%, Wilcoxon’s test for paired samples p � 0.048) and SJSA-1(median decrease 59%, p 0.0005) cells.
In contrast to the effects on Cal72 and SJSA-1 cells, AMLblasts had only minor and divergent effects on the proliferationof the epithelioid cell line 143.98.2 (data not shown) and of the4 other osteosarcoma cell lines (11 patients examined, data notshown).
The effects of AML blasts on osteoblastic sarcoma cells showedno association with signs of AML cell differentiation (FAB clas-sification and CD34 expression), cytogenetic abnormalities or ge-netic Flt3 abnormalities of AML cells (data not shown).
GM-CSF is involved in crosstalk between fibroblasts and nativehuman AML cells
To further characterize the molecular mechanisms involvedin crosstalk between AML blasts and nonleukemic cells, we
861AML CELL EFFECTS ON FIBROBLASTS/OSTEOBLASTS

prepared transwell cultures with and without cytokine-inhibi-tory mediators. AML blasts derived from 12 randomly selectedpatients (patients 7, 9, 10, 17, 21, 23, 29, 31, 42, 51, 54, 55)were cultured with HFL1 fibroblasts and Cal72 osteoblasticsarcoma cells in transwell cultures in the presence of neutral-izing anti-GM-CSF, anti-HGF, anti-VEGF, anti-SCF or corre-sponding control antibodies. Pilot experiments demonstratedthat these mediators did not alter the proliferation of fibroblastscultured alone (data not shown). Fibroblast proliferation wasincreased by anti-GM-CSF for a majority of patients (10 of 12,Wilcoxon’s test for paired samples p 0.005), though exoge-nous GM-CSF alone did not alter proliferation of fibroblastscultured alone (data not shown). Furthermore, an increase cor-responding to �2,000 cpm and exceeding the control by at least
20% was also observed for certain patients in the presence ofanti-HGF (2/12) and anti-SCF (2/12). Proliferation of Cal72osteoblastic sarcoma cells was altered (i.e., the difference being�2,000 cpm and �20% of control) only for a minority ofpatients by the neutralizing antibodies (anti-GM-CSF 1/12,anti-HGF 0/12, anti-SCF 1/12, anti-VEGF 0/12). Furthermore,leukemia cells derived from 11 other randomly selected patients(Table I, patients 1, 9, 10, 17–21, 23, 25, 26) were cultured withCal72 and HFL1 cells in medium alone and in the presence ofIL-1RA 50 ng/ml. Exogenous IL-1RA increased nonleukemiccell proliferation (i.e., a difference corresponding to �2,000cpm and �20% of controls) for only a minority of patientswhen testing HFL1 (2/11) and Cal72 cells (3/11). We concludethat (i) GM-CSF is usually involved in crosstalk between fi-
FIGURE 1 – AML induced effectson the proliferation of fibroblasts(Hs27 and HFL1 cells) and osteo-blastic sarcoma cells (Cal72 andSJSA-1). Native human AMLblasts were cultured together withnonleukemic cells separated by asemipermeable membrane. Theculture medium was serum-freeStem Span with 10% FCS, andproliferation was assayed as 3H-thymidine incorporation after 7days of coculture. AML cells wereexamined in separate experiments,each including 6–8 patients, andthe results are therefore presentedas the percentage increase/decreasein fibroblast proliferation (incre-mental responses used for the cal-culation). Proliferation in AML-free controls showed widevariation between the cell lines: (i)Hs27 (upper left), proliferation inAML-free controls in separate ex-periments corresponding to 3,520–11,737 cpm (median of triplicatedeterminations); (ii) HFL1 (upperright), AML-free controls gener-ally exceeding 30,000 cpm; (iii)Cal72 (lower left), AML-free con-trols exceeding 30,000 cpm in allexperiments; and (iv) SJSA-1(lower right), AML-free controlsalways exceeding 80,000 cpm.
862 GLENJEN ET AL.

broblasts and native human AML cells but (ii) additional mech-anisms that differ between patients and may involve HGF, SCFand IL-1� also appear to be involved in the AML-associatedinhibition of fibroblasts and osteoblasts.
GM-CSF levels can be increased during coculture of AMLblasts with HFL1 fibroblasts and osteoblastic sarcoma cells
Constitutive GM-CSF secretion by native AML blasts wasinvestigated for patients 1–54 (Table I), and detectable release wasdefined as detectable supernatant levels in at least 2 of 3 experi-ments when AML blasts were cultured alone for 7 days. DetectableGM-CSF release was then observed for 40 patients (median level26.1 pg/ml, variation range 2.3–1,260 pg/ml). Constitutive GM-CSF release was also detected for fibroblast and sarcoma cell lines,the levels in control cultures of each coculture experiment beingfor HFL1 fibroblasts always 41.0 pg/ml, for Hs27 fibroblasts35.0 pg/ml, for SJSA-1 sarcoma cells 32.0 pg/ml, for Cal72sarcoma cells 5.6 pg/ml and for 143.98.2 sarcoma cells �1,000pg/ml.
Native human AML blasts were cocultured in transwell cul-tures with HFL1 and Hs27 fibroblasts (Table I, patients 1–34)
and with the osteoblastic sarcoma cell lines Cal72, SJSA-1 and143.98.2 (Table I, patients 1–10, 12, 13, 16 –23, 26, 28, 29, 31,34 –36, 38, 42, 43, 45– 47, 49, 50 –54). Increased GM-CSFlevels were detected when AML blasts with constitutive GM-CSF secretion (20 of the 34 patients) were cocultured withHFL1 fibroblasts both in transwell cultures (Fig. 3, Wilcoxon’stest for paired samples p � 0.004) and when cells were culturedin direct contact (p � 0.008, data not shown). Increased levelswere also observed during coculture with Cal72 and SJSA-1cells (Fig. 3, p 0.001 for both) when testing AML blasts withconstitutive GM-CSF release (31 of the 39 patients). GM-CSFlevels for AML cells cultured alone showed strong correlationswith the levels in cocultures both when testing HFL1, Cal72 andSJSA-1 nonleukemic cells (Spearman’s test, correlation co-efficients �0.89 and p 0.001 for all 3 cell lines). In contrast,an increase was not observed in experiments including (i) AMLblasts with low/undetectable GM-CSF release (Fig. 3); (ii)coculture of AML blasts with Hs27 fibroblasts, these levelsbeing significantly lower than for HFL1 cells (Wilcoxon’s testfor paired samples p 0.001); and (iii) cocultures with theepithelial sarcoma cell line 143.98.2, decreased GM-CSF levelsbeing detected (p � 0.002, data not shown).
GM-CSF levels in cultures containing AML cells alone andAML cells cocultured with any of the nonleukemic cells showedno correlation with differentiation (FAB classification or CD34expression), AML cell karyotype or genetic Flt3 abnormalities(data not shown).
IL-1� levels (but not levels of IL-1RA) are increased duringcoculture of AML blasts with HFL1 fibroblasts and osteoblasticsarcoma cells
Both normal as well as leukemic hematopoiesis can be stimu-lated by IL-1, and native human AML blasts can release both IL-1and IL-1RA.32 The balance between IL-1 and IL-1RA release willprobably determine the effect on neighboring normal hematopoi-esis. IL-1� is usually released at higher levels than IL-1� by nativehuman AML blasts,26 and we therefore compared levels of IL-1�in cultures containing AML blasts alone, osteoblasts/fibroblastsalone and cocultures of AML blasts with HFL1 fibroblasts (TableI, patients 1–33) with the osteoblastic sarcoma cell lines Cal72 andSJSA-1 (Table I, patients 1–10, 12, 13, 16–23, 26, 28, 29, 31,34–36, 38, 42, 43, 45–47, 49, 50–54). Constitutive IL-1� releasewas detected only for a subset of patients (median level 21.6pg/ml, range 4.8–416 pg/ml). Increased IL-1� levels were ob-served when AML blasts with constitutive IL-1� release werecocultured with HFL1 fibroblasts and Cal72 and SJSA-1 osteo-blastic sarcoma cells (Fig. 4, Wilcoxon’s test for paired samplesp 0.001 for all 3 cell lines); this increase was also maintainedwhen AML cells and HFL1 fibroblast were cultured in directcontact. IL-1� levels for AML cells cultured alone showed strongcorrelations with the levels in cocultures when testing HFL1,Cal72 and SJSA-1 nonleukemic cells (Spearman’s test correla-tion coefficients �0.90 and p 0.001 for all 3 cell lines). Incontrast, IL-1� was not detected for culture containing only non-leukemic cells and for a small minority of cocultures containingAML cells without constitutive IL-1� secretion (undetectable lev-els 2.8 pg/ml).
We analyzed IL-1RA levels after in vitro culture of AML blastsderived from 50 patients (Table I, patients 1–16, 18–36, 38–43,46–54). IL-1RA was detected in the supernatants for all patients(median level 1,100 pg/ml, range 20–2,800 pg/ml), and patientswith AML-M4/M5 had increased levels (Mann-Whitney U-testp � 0.02). AML blasts from 26 randomly selected patients wereavailable for additional studies of IL-1RA levels after coculturewith HFL1 fibroblasts (patients 1–7, 9–13, 18–30, 33). DetectableIL-1RA levels were then observed for 25 patients (median 305pg/ml, range 14.0–2,700 pg/ml). IL-1RA levels for AML cul-tures and corresponding cocultures showed a significant correla-tion (Fig. 5, Spearman’s test, n � 26, correlation coefficient 0.53,p � 0.005), though levels were lower in cocultures (Wilcoxon’s
FIGURE 2 – Effect of AML culture supernatants on proliferation ofHFL1 fibroblasts. Native human AML blasts derived from 19 patientswere cultured in serum-free Stem Span medium for 48 hr beforesupernatants were harvested. HFL1 fibroblasts were cultured in therecommended Ham’s F12K medium with 10% FCS in the presence ofthe supernatants (final dilution 1:4). 3H-Thymidine incorporation wasassayed after 4 days of culture. Results are presented as the percentageincrease/decrease in fibroblast proliferation (incremental responsesused for the calculation).
863AML CELL EFFECTS ON FIBROBLASTS/OSTEOBLASTS

test for paired samples p � 0.049). Furthermore, a significantincrease in the IL-1�/IL-1RA ratio was observed for HFL1 trans-well cultures (median 0.065, range 0.001–4.762) compared tocorresponding control cultures containing AML blasts alone (me-dian 0.020, range 0.002–0.151, Wilcoxon’s test for paired samplesp � 0.002).
FIGURE 3 – GM-CSF levels during coculture of native human AMLblasts and nonleukemic cells (osteoblastic Cal72 and SJSA-1 osteo-sarcoma cells, HFL1 fibroblasts) in transwell cultures. Results forAML cell populations with (closed symbols) and without (open orhalf-open symbols) constitutive GM-CSF release. At left, we compareGM-CSF levels when AML cells derived from 39 patients werecultured alone (0) and cocultured with either Cal72 or SJSA-1 osteo-blastic sarcoma cells. AML cell populations with constitutive secretion(closed symbols) showed increased GM-CSF levels in coculturescompared to the summarized levels for corresponding control culturescontaining AML cells and nonleukemic cells alone. For 8 patients,AML cell populations did not show constitutive GM-CSF release(open symbols). Levels in these cocultures were generally low; forcertain patients, levels were even lower than those for control culturescontaining Cal72 and SJSA-1 nonleukemic cells alone (open symbols),whereas coculture levels exceeded control levels for the other patients(half-open symbols). Results for AML cells cultured alone and cocul-tured with HFL1 fibroblasts are presented in the same way at right.Increased GM-CSF levels were detected when AML cells were cocul-tured with HFL1 fibroblasts.
FIGURE 4 – IL-1� levels during coculture of native human AMLblasts with osteoblastic sarcoma cell lines and HFL1 fibroblasts intranswell cultures. IL-1� levels were determined by ELISA for AMLcells cultured alone and cocultured with Cal-72, SJSA-1 (39 patients)and HFL1 (33 patients) cells. Nonleukemic cells showed low/unde-tectable IL-1� release corresponding to 2.8 pg/ml. ConstitutiveIL-1� release was observed only for a subset of patients when cultur-ing AML cells alone, and the figure presents the results only for thissubset of patients. For patients with nonsecreting AML blasts, unde-tectable IL-1� levels (2.8 pg/ml) were usually observed both whenAML cells were cultured alone and in all cocultures (only 4 excep-tions, marked with open symbols).
864 GLENJEN ET AL.
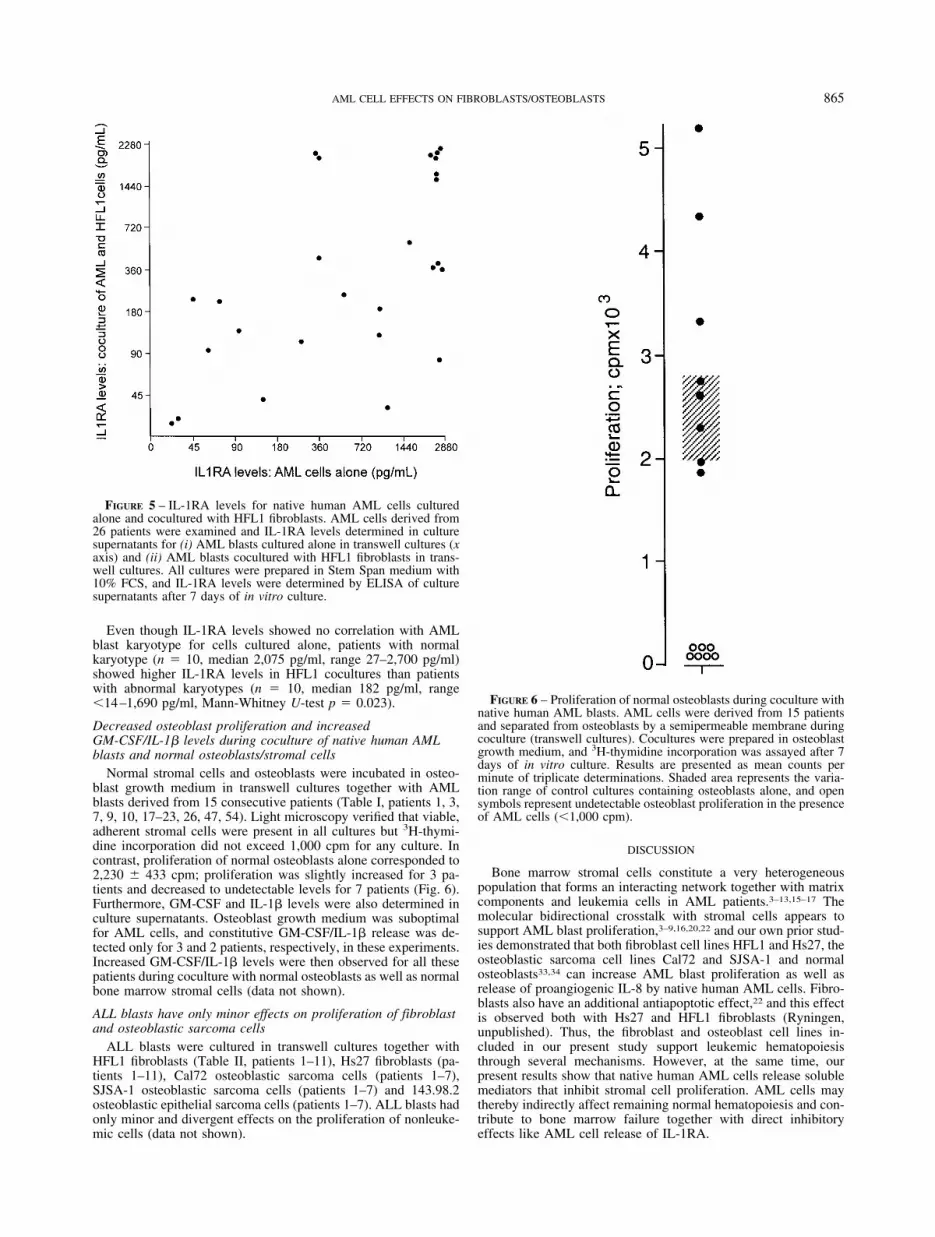
Even though IL-1RA levels showed no correlation with AMLblast karyotype for cells cultured alone, patients with normalkaryotype (n � 10, median 2,075 pg/ml, range 27–2,700 pg/ml)showed higher IL-1RA levels in HFL1 cocultures than patientswith abnormal karyotypes (n � 10, median 182 pg/ml, range14–1,690 pg/ml, Mann-Whitney U-test p � 0.023).
Decreased osteoblast proliferation and increasedGM-CSF/IL-1� levels during coculture of native human AMLblasts and normal osteoblasts/stromal cells
Normal stromal cells and osteoblasts were incubated in osteo-blast growth medium in transwell cultures together with AMLblasts derived from 15 consecutive patients (Table I, patients 1, 3,7, 9, 10, 17–23, 26, 47, 54). Light microscopy verified that viable,adherent stromal cells were present in all cultures but 3H-thymi-dine incorporation did not exceed 1,000 cpm for any culture. Incontrast, proliferation of normal osteoblasts alone corresponded to2,230 � 433 cpm; proliferation was slightly increased for 3 pa-tients and decreased to undetectable levels for 7 patients (Fig. 6).Furthermore, GM-CSF and IL-1� levels were also determined inculture supernatants. Osteoblast growth medium was suboptimalfor AML cells, and constitutive GM-CSF/IL-1� release was de-tected only for 3 and 2 patients, respectively, in these experiments.Increased GM-CSF/IL-1� levels were then observed for all thesepatients during coculture with normal osteoblasts as well as normalbone marrow stromal cells (data not shown).
ALL blasts have only minor effects on proliferation of fibroblastand osteoblastic sarcoma cells
ALL blasts were cultured in transwell cultures together withHFL1 fibroblasts (Table II, patients 1–11), Hs27 fibroblasts (pa-tients 1–11), Cal72 osteoblastic sarcoma cells (patients 1–7),SJSA-1 osteoblastic sarcoma cells (patients 1–7) and 143.98.2osteoblastic epithelial sarcoma cells (patients 1–7). ALL blasts hadonly minor and divergent effects on the proliferation of nonleuke-mic cells (data not shown).
DISCUSSION
Bone marrow stromal cells constitute a very heterogeneouspopulation that forms an interacting network together with matrixcomponents and leukemia cells in AML patients.3–13,15–17 Themolecular bidirectional crosstalk with stromal cells appears tosupport AML blast proliferation,3–9,16,20,22 and our own prior stud-ies demonstrated that both fibroblast cell lines HFL1 and Hs27, theosteoblastic sarcoma cell lines Cal72 and SJSA-1 and normalosteoblasts33,34 can increase AML blast proliferation as well asrelease of proangiogenic IL-8 by native human AML cells. Fibro-blasts also have an additional antiapoptotic effect,22 and this effectis observed both with Hs27 and HFL1 fibroblasts (Ryningen,unpublished). Thus, the fibroblast and osteoblast cell lines in-cluded in our present study support leukemic hematopoiesisthrough several mechanisms. However, at the same time, ourpresent results show that native human AML cells release solublemediators that inhibit stromal cell proliferation. AML cells maythereby indirectly affect remaining normal hematopoiesis and con-tribute to bone marrow failure together with direct inhibitoryeffects like AML cell release of IL-1RA.
FIGURE 5 – IL-1RA levels for native human AML cells culturedalone and cocultured with HFL1 fibroblasts. AML cells derived from26 patients were examined and IL-1RA levels determined in culturesupernatants for (i) AML blasts cultured alone in transwell cultures (xaxis) and (ii) AML blasts cocultured with HFL1 fibroblasts in trans-well cultures. All cultures were prepared in Stem Span medium with10% FCS, and IL-1RA levels were determined by ELISA of culturesupernatants after 7 days of in vitro culture.
FIGURE 6 – Proliferation of normal osteoblasts during coculture withnative human AML blasts. AML cells were derived from 15 patientsand separated from osteoblasts by a semipermeable membrane duringcoculture (transwell cultures). Cocultures were prepared in osteoblastgrowth medium, and 3H-thymidine incorporation was assayed after 7days of in vitro culture. Results are presented as mean counts perminute of triplicate determinations. Shaded area represents the varia-tion range of control cultures containing osteoblasts alone, and opensymbols represent undetectable osteoblast proliferation in the presenceof AML cells (1,000 cpm).
865AML CELL EFFECTS ON FIBROBLASTS/OSTEOBLASTS

The major part of our experiments was performed with well-characterized cell lines; this approach allows comparison of AML-induced effects between patients in standardized experimentalmodels. Both HFL1 and Hs27 fibroblasts support leukemic hema-topoiesis.33,35 The sarcoma cell line Cal72 has an osteoblasticphenotype and supports normal hematopoiesis,29,30 and SJSA-1cells had a similar phenotype. To further evaluate the biologicrelevance of our experimental results, we also included studies ofnormal osteoblasts and a heterogeneous population of normal bonemarrow stromal cells.
Osteoblasts and fibroblasts are closely related,36 and the effectsof native human AML cells on HFL1 fibroblasts, osteoblasticsarcoma cells (Cal72 and SJSA-1) and normal osteoblasts showedseveral similarities. Firstly, AML cells inhibited proliferation of allthese cells. This growth inhibition was detected by a 3H-thymidineincorporation assay. An alternative explanation for this decreased3H-thymidine incorporation after 7 days of in vitro coculture couldbe increased proliferation with formation of confluent layers be-fore day 7 and thereby decreased 3H-thymidine at the end of theculture period. However, several observations make this last ex-planation unlikely: (i) cell counting confirmed that cultures con-tained a decreased number of nonleukemic cells; (ii) all cultureswere observed under a microscope before harvesting, and adherentcells were not confluent in either control cultures or AML cocul-tures; (iii) 143.98.2 cells showed the strongest in vitro proliferationwhen cultured alone or in the presence of native human AML cells,but for this cell line no AML-associated growth inhibition wasdetected. We therefore conclude that the decreased 3H-thymidineincorporation by HFL1, Cal72 and SJSA-1 cells represents a trueAML-associated growth inhibition. Secondly, the presence offibroblasts, osteoblastic cells and normal osteoblasts could in-crease GM-CSF and IL-1� levels when these cells are cocul-tured with constitutive GM-CSF/IL-1�-secreting AML cells.These increased cytokine levels are probably caused by in-creased release from AML cells because the levels for AMLcells cultured alone showed a wide variation and significantcorrelations with the corresponding cocultures. Thirdly, theseeffects showed no associations with FAB classification, cyto-genetic abnormalities or genetic Flt3 abnormalities, therebysuggesting that the growth inhibition and increased GM-CSF/IL-1� levels are common functional characteristics of otherwiseheterogeneous AML cell populations.
The AML-associated growth inhibitory and GM-CSF/IL-1�-increasing effects were detected only for HFL1 fibroblasts, osteo-blastic sarcoma cells and normal osteoblasts/stromal cells, whereasthese effects were not observed when testing other osteosarcomacells (the epithelial line 143.98.2 was included in all experimentstogether with Cal72 and SJSA-1) and the Hs27 fibroblast cell line.Taken together, these observations suggest that the AML-associ-ated effects on sarcoma cells depend on the osteoblastic phenotyperather than on osteosarcoma-associated characteristics. Fibroblastsmay also show different phenotypic characteristics,14 and ourresults suggest that the AML-associated effects are observed onlyfor certain fibroblast subsets. However, our in vitro culture condi-tions appeared to be suboptimal for Hs27 cells, and this may alsocontribute to the differences between HFL1 and Hs27 fibroblasts.
In a separate series of experiments, we compared the effects onHFL1 fibroblasts cocultured with native human AML cells and inthe presence of culture supernatants harvested from AML cells thatwere cultured alone. The divergence between supernatant (in-
creased HFL1 proliferation) and coculture (decreased prolifera-tion) experiments clearly demonstrated that the AML-associatedgrowth inhibition is dependent on bidirectional crosstalk betweenthe cells.
GM-CSF was released by both nonleukemic cells as well asmost AML blast populations, and GM-CSF was detected in mostcocultures with fibroblasts and osteoblastic sarcoma cells. Exper-iments with GM-CSF neutralizing antibodies demonstrated thatGM-CSF was important for the crosstalk between AML cells andfibroblasts, but exogenous GM-CSF alone did not alter fibroblastproliferation. Our hypothesis is therefore that the increased GM-CSF levels mainly have effects on AML cells during coculture;AML blasts are usually GM-CSF-responsive, and GM-CSF can bean autocrine growth factor for native human AML cells.26–28
GM-CSF effects on AML blasts may then affect the cytokinerelease profile and thereby contribute indirectly to the effects onfibroblasts. Furthermore, AML blasts derived from different pa-tients vary in cytokine release profile,26,27,37 and other mediatorsmay also contribute to the growth inhibition in certain patients, assuggested by our cytokine neutralization studies.
Our present results confirmed that IL-1RA was released byall AML cell populations, especially by AML-M4/M5 cells,38
whereas IL-1� levels were generally lower and detectable lev-els were observed for only a subset of patients. Increased IL-1�levels were detected during coculture of AML blasts with HFL1fibroblasts and osteoblastic sarcoma cells but only for the subsetof patients with constitutive IL-1� release. Although release ofIL-1RA was decreased during coculture, the remaining levelswere still relatively high. IL-1 is important for normal hema-topoiesis,32 and the persistent local release of IL-1RA by AMLcells may thereby contribute to decreased normal hematopoiesisand bone marrow failure in AML patients, at least for the largesubset of patients with low/undetectable IL-1 release. IL-1� andIL-1RA levels for AML blasts cultured alone were significantlycorrelated to the corresponding levels in cocultures, suggestingthat AML blasts are the major source of both mediators duringcoculture.
Osteoblasts are a part of the stem cell niche, i.e., the bonemarrow microcompartment where hematopoietic stem cells arelocated.39,40 Previous studies have described reduced numbers ofcolony-forming fibroblasts in AML bone marrow,6 and our presentresults support this conclusion and suggest that the number ofother important stromal cells may also be reduced in AML. Al-though the local levels of certain AML-derived cytokines areincreased during coculture, the molecular mechanisms behind theAML-induced antiproliferative effect on nonleukemic cells areprobably heterogeneous and not caused by a single mediator. Agrowth-inhibitory effect on osteoblasts mediated via local cytokinenetworks may thereby represent a mechanism for AML-inducedinhibition of the remaining normal hematopoiesis. This inhibitionmay be further increased by similar inhibitory effects on fibroblastsubsets and the release of direct inhibitory mediators like IL-1RA(IL-1�/IL-1RA ratio 1.00 is usually maintained even in fibro-blast/AML cell cocultures). Taken together with previous results(Ryningen, unpublished),5,7,22,34,37 we therefore suggest that non-leukemic stromal cells may be important in disease development inAML patients through several mechanisms, including enhance-ment of leukemic hematopoiesis, stimulation of bone marrowangiogenesis and reduced stimulation of remaining normal hema-topoiesis.
REFERENCES
1. Lowenberg B, Downing JR, Burnett A. Acute myeloid leukemia.N Engl J Med 1999;341:1051–62.
2. Bruserud O� , Gjertsen BT. New strategies for the treatment of acutemyelogenous leukemia: differentiation induction-present use and fu-ture possibilities. Stem Cells 2000;18:157–65.
3. Dilly SA, Jagger CJ. Bone marrow stromal cell changes in haemato-logical malignancies. J Clin Pathol 1990;43:942–6.
4. Mayani H, Guilbert LJ, Clark SC, Belch AR, Janowska-Wieczorek A.
Composition and functional integrity of the in vitro hemopoieticmicroenvironment in acute myelogenous leukemia: effect of macro-phage colony-stimulating factor. Exp Hematol 1992;20:1077–84.
5. Mayani H. Composition and function of the hemopoietic microenvi-ronment in human myeloid leukemia. Leukemia 1996;10:1041–7.
6. Duhrsen U, Hossfeld DK. Stromal abnormalities in neoplastic bonemarrow disease. Ann Hematol 1996;73:53–70.
7. Zhang W, Knieling G, Vohwinkel G, Martinez T, Kuse R, Hossfeld
866 GLENJEN ET AL.

DK, Duhrsen U. Origin of stroma cells in long-term bone marrowcultures from patients with acute myeloid leukemia. Ann Hematol1999;78:305–14.
8. Allen TD, Dexter TM. The essential cells of the hemopoietic micro-environment. Exp Hematol 1984;12:517–24.
9. Bianco P, Riminucci M, Gronthos S, Robey PG. Bone marrow stro-mal stem cells: nature, biology, and potential applications. Stem Cells2001;19:180–92.
10. Wilson JG. Adhesive interactions in hemopoiesis. Acta Haematol1997;97:6–12.
11. Liesveld JE. Expression and function of adhesion receptors in acutemyelogenous leukemia: parallels with normal erythroid and myeloidprogenitors. Acta Hematol 1997;97:53–62.
12. Bendall LJ, Kortlepel K, Gottlieb DJ. Human acute myeloid leukemiacells bind to bone marrow stroma via a combination of �-1 and �-2integrin mechanisms. Blood 1993;82:3125–32.
13. Bendall LJ, Gottlieb DJ. CD44 and adhesion of normal and leukemicCD34� cells to bone marrow stroma. Leuk Lymphoma 1999;32:427–39.
14. Rougier F, Dupuis F, Denizot Y. Human bone marrow fibroblasts—anoverview of their characterization, proliferation and inflammatorymediator production. Hematol Cell Ther 1996;38:241–6.
15. Kittler EL, McGrath H, Temeles D, Crittenden RB, Kister VK,Quesenberry PJ. Biologic significance of constitutive and subliminalgrowth factor production by bone marrow stoma. Blood 1992;79:3168–78.
16. Stolze B, Emmendorffer A, Corbacioglu S, Konig A, Welte K, EbellW. Effects of bone marrow fibroblasts on the proliferation and dif-ferentiation of myeloid leukemic cell lines. Exp Hematol 1995;23:1378–87.
17. Klein G. The extracellular matrix of the hematopoietic microenviron-ment. Experientia 1995;51:914–26.
18. Santucci MA, Lemoli SM, Tura S. Peripheral blood mobilization ofhematopoietic stem cells: cytokine-mediated regulation of adhesiveinteractions within the hematopoietic microenvironment. ActaHaematol 1997;97:90–6.
19. Lowenberg B, van Putten W, Touw IP, Delwel R, Satini V. Auton-omous proliferation of leukemic cells in vitro as a determinant ofprognosis in adult acute myeloid leukemia. N Engl J Med 1993;328:614–9.
20. Bendall LJ, Daniel A, Kortlepel K, Gottlieb DJ. Bone marrow adher-ent layers inhibits apoptosis of acute myeloid leukemia cells. ExpHematol 1994;22:1252–60.
21. Murohashi I, Yoshida K, Handa A, Murayoshi M, Yoshida S, JinnaiI, Bessho M, Hirashima K. Differential regulation by hematopoieticgrowth factors of apoptosis and mitosis in acute myeloblastic leuke-mia cells. Exp Hematol 1997;25:1042–50.
22. Garrido SM, Appelbaum FR, Willman CL, Banker DE. Acute my-eloid leukemia cells are protected from spontaneous and drug-inducedapoptosis by direct contact with a human bone marrow stromal cellline (HS-5). Exp Hematol 2001;29:448–57.
23. Lichtman MA. Interrupting the inhibition of normal hematopoiesis inmyelogenous leukemia: a hypothetical approach to therapy. StemCells 2000;18:304–6.
24. Bene MC, Castoldi G, Knapp W, Ludwig WD, Matutes E, Orfao A,van’t Veer MB. Proposals for the immunological classification ofacute leukemias. Leukemia 1995;9:1783–6.
25. Bruserud O� . Effects of dipyridamole, theophyllamine and verapamil
on spontaneous proliferation of myelogenous leukemia cells. ActaOncol 1992;31:53–8.
26. Bruserud O� , Gjertsen BT, Brustugun OT, Bassoe CF, Nesthus I,Akselsen PE, Buhring HJ, Pawelec G. Effect of interleukin 10 on blastcells derived from patients with acute myelogenous leukemia. Leu-kemia 1995;9:1910–20.
27. Bruserud O� , Gjertsen BT, Foss B, Huang TS. New strategies in thetreatment of acute myelogenous leukemia (AML): in vitro culture ofAML cells—the present use in experimental studies and the possibleimportance for future therapeutic strategies. Stem Cells 2001;19:1–11.
28. Bruserud O� , Huang TS, Glenjen N, Gjertsen BT, Foss B. Leptin inhuman acute myelogenous leukemia: studies of in vivo levels and invitro effects on native functional leukemic blasts. Haematologica2002;87:584–95.
29. Rochet N, Leroy P, Far DF, Ollier L, Loubat A, Rossi B. CAL72: ahuman osteosarcoma cell line with unique effects on hematopoieticcells. Eur J Haematol 2003;70:43–52.
30. Rochet N, Dubousset J, Mazeau C, Zanghellini E, Farges MF, deNovion HS, Chompret A, Delpech B, Cattan N, Frenay M, Gioanni J.Establishment, characterization and partial cytokine expression profileof a new human osteosarcoma cell line (Cal72). Int J Cancer 1999;82:282–5.
31. Bruserud O� , Gjertsen BT, von Volkman HL. In vitro culture of humanacute myelogenous leukemia (AML) cells in serum-free media: stud-ies of native AML blasts and AML cell lines. J Hematother Stem CellRes 2000;9:923–32.
32. Dinarello CA. Interleukin-1, interleukin-1 receptors and interleukin-1receptor antagonist. Int Rev Immunol 1998;16:457–99.
33. Bruserud O� , Glenjen N, Ryningen A. Effects of angiogenic regulatorson in vitro proliferation and cytokine secretion by native human acutemyelogenous leukemia blasts. Eur J Haematol 2003;71:9–17.
34. Bruserud O� , Ryningen A, Wergeland L, Glenjen NI, Gjertsen BT.Osteoblasts increase proliferation and release of proangiogenic inter-leukin 8 by native human acute myelogenous leukemia blasts. Haema-tologica 2004;89:391–402.
35. Bruserud O� , Glenjen N, Ryningen A, Ulvestad E. In vitro culture ofhuman acute myelogenous leukemia (ALL) cells in serum-free media:a comparison of native ALL blasts, ALL cell lines and virus-trans-formed B cell lines. Leukemia Res 2003;27:455–64.
36. Ducy P, Schinke T, Karsenty G. The osteoblast: a sophisticatedfibroblast under central surveillance. Science 2000;289:1501–4.
37. Glenjen N, Hovland R, Wergeland L, Wendelbo O, Ernst P, BruserudO. The angioregulatory phenotype of native human acute myeloge-nous leukemia cells: influence of karyotype, Flt3 abnormalities anddifferentiation status. Eur J Haematol 2003;71:163–73.
38. Bruserud O, Aasen I, Akselsen PE, Bergheim J, Rasmussen G, Nes-thus I. Interleukin 1 receptor antagonist (IL-1RA) in acute leukaemia:IL-1RA is both secreted spontaneously by myelogenous leukaemiablasts and is a part of the acute phase reaction in patients withchemotherapy-induced leucopenia. Eur J Haematol 1996;57:87–95.
39. Nilson SK, Johnston HM, Coverdale JA. Spatial localization of trans-planted hematopoietic cells: inferences for the localization of stemcell niches. Blood 2001;97:2293–9.
40. Calvi LM, Adams GB, Weibrecht KW, Weber JM, Olson DP, KnightMC, Martin RP, Schipani E, Divieti P, Bringhurst FR, Milner LA,Kronenberg HM, Scadden DT. Osteoblastic cells regulate the hema-topoietic stem cell niche. Nature 2003;23:778–9.
867AML CELL EFFECTS ON FIBROBLASTS/OSTEOBLASTS