in vitro cytotoxicity against different human cancer cell lines of laticifer proteins of calotropis...
TRANSCRIPT

Available online at www.sciencedirect.com
www.elsevier.com/locate/toxinvit
Toxicology in Vitro 21 (2007) 1563–1573
In vitro cytotoxicity against different human cancer cell linesof laticifer proteins of Calotropis procera (Ait.) R. Br
Jefferson Soares de Oliveira a, Daniel Pereira Bezerra b, Cleverson Diniz Teixeira de Freitas a,Jose Delano Barreto Marinho Filho b, Manoel Odorico de Moraes b, Claudia Pessoa b,
Letıcia Veras Costa-Lotufo b,*, Marcio Viana Ramos a,*
a Departamento de Bioquımica e Biologia Molecular, Universidade Federal do Ceara, Campus do Pici, Caixa Postal 6033,
CEP 60.451-970, Fortaleza, Ceara, Brazilb Departamento de Fisiologia e Farmacologia, Universidade Federal do Ceara, Caixa Postal 3157,
CEP 60.430-270, Fortaleza, Ceara, Brazil
Received 5 March 2007; accepted 11 May 2007Available online 23 May 2007
Abstract
This work evaluated the in vitro cytotoxic activity of laticifer proteins (LP) recovered from the latex of the medicinal plant Calotropis
procera. The LP displayed considerable cytotoxicity with IC50 values ranging from 0.42 to 1.36 lg/ml to SF295 and MDA-MB-435 celllines, respectively. In healthy peripheral blood mononuclear cells exposed to LP (10 lg/ml) for 72 h, no noticeable effects on viability orcell morphology were seen. The fractionating of LP on an ion exchange chromatography gave rise to a new fraction (PI) that retainedalmost all cytotoxicity. The cytotoxic effects of both LP and PI were diminished when previously treated with pronase, or 2-mercap-toethanol, suggesting a protein nature of active molecules, however, pre-incubation with dithiothreitol (DTT) only reduced PI activity.PI did not exhibit cysteine proteinase activity, indicating that cysteine proteinases, abundantly found in LP, are not implicated in LPcytotoxicity. In this study, using HL-60 cell as a model, LP was shown to inhibit DNA synthesis. This is probably due to alterationsin the topology of DNA, since it was observed that LP is able to interfere in topoisomerase I activity by somehow acting uponDNA. LP provoked reduction in cell number but it did not cause any significant increase in the number of non-viable cells. These find-ings corroborated with the morphologic analysis, where cells treated with LP showed morphology of apoptotic process with abundantvacuoles, chromatin condensation and fragmentation of the nuclei. The results of this study suggests that LP is a target for DNA topo-isomerase I triggering apoptosis in cancer cell lines.� 2007 Elsevier Ltd. All rights reserved.
Keywords: Apoptosis; Cytotoxic activity; Latex; Topoisomerase I inhibitor
1. Introduction
The circulating fluid named latex is found in more than12,000 plant species and the most abundant chemical struc-ture found is the polymeric hydrocarbon cis-poly-isoprene
0887-2333/$ - see front matter � 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.tiv.2007.05.007
Abbreviations: LP, (laticifer proteins); PBMC, (peripheral blood mono-nuclear cells).
* Corresponding authors. Fax: +55 85 3366 9789 (M.V. Ramos).E-mail addresses: [email protected] (L.V. Costa-Lotufo), vramos
@ufc.br (M.V. Ramos).
that is implicated in the formation of rubber (Yeang et al.,2002). A wide range of proteins has been described asoccurring in latex fluids, including soluble proteinsinvolved in the biosynthesis of rubber, plant defence andoxidative metabolism (Azarkan et al., 1997; Nallamsettyet al., 2003). Laticifer plants have been proven to be aninteresting source of biologically active molecules and theireffectiveness is frequently supported by their previous usein folk medicine. This is the case with Calotropis procera.Pharmacological activities attributed to C. procera latexinclude anti-inflammation, antioxidant and antinociceptive

1564 J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573
among many others (Arya and Kumar, 2005; Roy et al.,2005; Soares et al., 2005). Recently, the plant has attractedgreater attention because of distinct and new relevant activ-ities found to be present in its latex (Choedon et al., 2006;Ramos et al., 2006a; Chitme et al., 2004).
Although there is convincing evidence of the pharmaco-logical potential of C. procera latex, the whole latex is usu-ally taken as a test sample. Thus the nature of the activemolecule for each specific effect remains unknown. This isbecause latex is not a homogeneous fluid and latex proteinsare not homogeneously dispersed. Accordingly, few pro-teins have been completely isolated from C. procera latex(Dubey and Jagannadham, 2003). Indian groups havemade relevant contributions to the understanding of phar-macological properties of the latex of C. procera. Theyhave recently shown that the whole latex of C. procera pos-sesses anticancer and cytotoxic activity against hepatocel-lular carcinoma (Choedon et al., 2006). Therefore, thepresence of poly-isoprene (rubber) is undesirable, as itmay be potentially responsible for other undesired effectsdescribed to occur in the latex (Al-Mezaine et al., 2005;Kumar et al., 2006).
In this work, the fresh latex of C. procera was fraction-ated to eliminate the rubber fraction and small molecularsize compounds as described recently (Alencar et al.,2004). The water-soluble protein fraction of the latex wasevaluated to determine the potential cytotoxic activityagainst different human cancer cell lines.
2. Materials and methods
2.1. Latex and extraction of laticifer proteins (LP)
Healthy plants of C. procera (Asclepiadaceae) growingin the field around the beaches of Fortaleza, Brazil wereused as the source of fresh latex. The plant material wasidentified by Prof. Edson Paula Nunes and voucher N.32663 was deposited at the Prisco Bezerra Herbarium ofthe Universidade Federal do Ceara, Brazil.
The latex was collected and placed in distilled water togive a dilution rate of 1:2 (v/v). The mixture was gently agi-tated during collection to overcome the natural coagula-tion effect of the material. Later, the samples werecentrifuged at 5000g for 10 min at 25 �C. The rubber-richprecipitate was removed and the supernatant was exhaus-tively dialyzed in distilled water using membranes of 8000molecular weight cut-off. The dialysis water was renewedevery 4 h to flow out ions, salts and other small organicmetabolites and newly centrifuged using the conditionsdescribed above. The clean, rubber-free supernatant waslyophilized and used for further experiments.
2.2. Biochemical analysis
2.2.1. Ion exchange chromatography (CM-Sepharose)LP (10 mg/ml) was submitted to ion exchange chroma-
tography in a CM-Sepharose fast flow column (Amersham
Biosciences, Brazil) previously equilibrated with 50 mMacetate-Na buffer pH 5.0. Samples of 30 mg were assayed.After washing the column with acetate buffer to elute theunbound proteins, the column was washed with 0.2 Mand 0.3 M NaCl sequentially. Three distinct protein peaksnamed PI (unbound), PII and PIII were then recovered,dialyzed against distilled water and lyophilized. They werefurther checked by PAGE-SDS and tested for cytotoxicityand enzymatic activity.
2.2.2. Digestion of LP and chromatographic fraction by
pronase
LP and PI were submitted to enzymatic digestion usingprotease from Streptomyces griseus (SIGMA P-5147) for24 h at 37 �C in 100 mM PBS pH 7.4. The enzyme samplerate was 1:100 (w/w) and the enzyme was added three timescorresponding to zero, 12 and 18 h of digestion. Afterbeing digested samples were heated at 60 �C for 10 minto inactivate the enzyme and lyophilized. Proteolysis wasfurther checked by electrophoresis.
2.2.3. Treatment of LP and chromatographic fraction by
dithiothreitol (DTT) and 2-mercaptoethanolSamples of LP and peaks of CM-Sepharose were dis-
solved in phosphate-buffered saline (PBS) pH 7.5 contain-ing 3 mM DTT or 1% 2-mercaptoethanol. These sampleswere further used in different assays to investigate the pos-sible involvement of cysteine-containing proteins in thecytotoxicity of latex protein fractions.
2.2.4. Electrophoresis analysis of LP and chromatographicfractions
LP, products of LP and PI digestion by pronase andpeaks recovered from CM-Sepharose were examined by12.5% polyacrylamide gel electrophoresis in the presenceof sodium dodecyl sulphate (SDS) according to Laemmli(1970). Samples were treated with 0.0625 M tris bufferpH 6.8 containing 2% SDS in the presence or absence of3 mM Dithiothreitol (DTT) or 1% 2-mercaptoethanoland applied into the gel. Runs were performed at 40 mAat room temperature. Gels were stained with 0.1% coomas-sie brilliant blue (R-350) solution in water:acetic acid:meth-anol (8:1:3.5). Unbound dye was removed with the samesolution without the dye.
2.2.5. Evaluation of proteolytic activity of LP and
chromatographic fractionsAssays for cysteine proteinases were produced using
BANA (SIGMA Product N. B-4750) as the substrate.Since it is already known that such an activity occurs inLP of C. Procera (Dubey and Jagannadham, 2003), theobjective was to measure this activity in an attempt todetermine its possible involvement in cytotoxic action ofLP.
Aliquots of 50 ll (1 mg/ml in 50 mM PBS pH 6.0) of LPand peaks recovered from the CM-Sepharose column werepre-incubated with 40 ll of an activation solution with

J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573 1565
3 mM DTT and 2 mM EDTA for 10 min at 37 �C and200 ll of BANA (1 mM in 50 mM PBS pH 6.0) wereadded. After 30 min, the reaction was stopped by adding500 ll of 2% HCl in ethanol and 500 ll of 0.06% 4-(dimethyl-amino)cinnamaldehyde. After 40 min, the colourdeveloped was measured by absorbance at 540 nm (Abeet al., 1992). One unit of enzymatic activity was definedas the amount of enzyme that increases the absorbance at540 nm by 0.01. All assays were performed in triplicate.The results of each series were expressed as the meanvalue ± SEM.
2.2.6. Proteolytic activity of LP and chromatographic
fractions on gelatin-containing gels
Electrophoresis were carried out according to themethod used by Laemmli (1970). The sample was separatedby electrophoresis through 12.5% polyacrylamide gels con-taining 0.1% gelatin at 25 �C (Macedo et al., 2004). Afterelectrophoresis, the gels were immersed in water containing2.5% of Triton X-100 (re-naturing solution) and gently sha-ken for 30 min at room temperature to wash out the SDSand re-nature proteins. The gels were then incubated in50 mM PBS pH 6.0, for 2 h at 37 �C containing 3 mMDTT and 2 mM EDTA. The incubated gels were stainedand revealed as before. Proteolytic activity was detectedas a transparent band that was not stained by CoomassieBrilliant Blue.
2.3. Cytotoxicity assays
2.3.1. Cell line and cell cultureThe cell lines used in this work were HL-60 (promyelo-
cytic leukemia), HCT-8 (colon), MDA-MB-435 (breast)and SF295 (brain), all of which were obtained from TheNational Cancer Institute (Bethesda, MD, USA). The cellswere maintained in RPMI 1640 medium supplementedwith 10% fetal bovine serum, 2 mM glutamine, 100 U/mlpenicillin, 100 lg/ml streptomycin at 37 �C with 5% CO2.
Heparinized blood from healthy, non-smoker donorswho had not taken any drugs for at least 15 days priorto sampling was collected and peripheral blood mononu-clear cells (PBMC) were isolated by a standard methodof density-gradient centrifugation over Ficoll-Hypaque.PBMC were washed and re-suspended at a concentrationof 3 · 105 cells/ml in RPMI 1640 medium supplementedwith 20% fetal bovine serum, 2 mM glutamine, 100 U/mlpenicillin, 100 lg/ml streptomycin at 37 �C with 5% CO2.Phytohemagglutinin (4%) was added at the beginning ofculture. After 24 h of culture, cells were treated with thetested compounds.
2.3.2. Proliferation and viability assays
2.3.2.1. Inhibition of tumor cell proliferation. The cytotoxic-ity of LP and CM-Sepharose peaks (PI, PII and PIII) wastested against HL-60 (promyelocytic leukemia), HCT-8(colon), MDA-MB-435 (breast) and SF295 (brain). InAddition, pronase digest, DTT or 2-mercaptoethanol-trea-
ted LP and PI were evaluated. For all experiments, cellswere plated in 96-well plates (105 cells/well for adherentcells or 0.5 · 105 cells/well for suspended cells in 100 ll ofmedium). After 24 h, LP (0.02–25.0 lg/ml) dissolved inwater was added to each well (using the high-throughputscreening biomek 3000 laboratory automation workstationin a biosafety hood, Baker Company, Maine) and incu-bated for 72 h. Doxorubicin (Doxolem�, Zodiac ProdutosFarmaceuticos S/A, Brazil) was used as positive control.The tumor cell growth was quantified by the ability of liv-ing cells to reduce the yellow dye 3-(4,5-dimethyl-2-thiaz-olyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) to apurple formazan product (Mosmann, 1983). At the endof the incubation, the plates were centrifuged and the med-ium was then replaced by fresh medium (200 ll) containingMTT (0.5 mg/ml). After 3 h, the MTT formazan productwas dissolved in 150 ll DMSO, and the absorbance wasmeasured using a multiplate reader (Spectra Count, Pack-ard, Ontario, Canada). The drug effect was quantified asthe percentage of control absorbance of reduced dye at595 nm.
2.3.2.2. Inhibition of PBMC proliferation. The antiprolifer-ative activity of LP and doxorubicin (10 or 25 lg/ml) onPBMC was assessed by culturing cells (3 · 105 cells/ml) inthe presence of each drug for 24, 48 and 72 h. After eachperiod, aliquots were removed from the cultures, and cellproliferation and viability were determined by the trypanblue dye exclusion test.
2.3.3. Analysis of mechanisms involved in the cytotoxic
activity
In order to elucidate the mechanisms involved in thecytotoxic action of LP, a second set of experiments wereperformed using HL-60 cells as a model essentially asdescribed by Rocha et al. (2006). The LP was tested atthe concentration of 0.25, 0.5 and 1 lg/ml. The trypan blueexclusion test was also performed before each experimentdescribed below to assess cell viability. Doxorubicin(0.3 lg/ml) was used as a positive control.
2.3.3.1. Inhibition of DNA synthesis. HL-60 cells (3 ·105 cells/ml) were plated onto 24-well tissue culture plates(2 ml/well) and treated with LP. Doxorubicin (0.3 lg/ml)was used as a positive control. Ten microliters of 5-bromo-2 0-deoxyuridine (BrdU, 10 mM) were added to eachwell and incubated for 3 h at 37 �C before completing the24 h period of drug exposure. To assay the amount ofBrdU incorporated into DNA, cells were harvested, trans-ferred to cytospin slides, and allowed to dry for 2 h at roomtemperature. Cells that had incorporated BrdU werelabeled by direct peroxidase immunocytochemistry usingchromogen diaminobenzidine. Slides were counterstainedwith hematoxylin, mounted, and coverslipped. Evaluationof BrdU positivity was performed by light microscopy(Olympus, Tokyo, Japan). Two hundred cells were countedper sample to determine the percentage of positive cells.

1566 J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573
2.3.3.2. Morphological analysis with hematoxylin–eosin
staining. Untreated or LP-treated HL-60 cells were exam-ined for morphological changes by light microscopy(Olympus, Tokyo, Japan). To evaluate nuclear morphol-ogy, cells from cultures were harvested after 24 h, trans-ferred to cytospin slides, fixed with methanol for 30 seg,and stained with hematoxylin–eosin.
2.3.3.3. Morphological analysis with fluorescence micro-
scope. HL-60 cells were treated with LP for 24 h, pelletedand re-suspended in 25 ll phosphate-buffered saline(PBS). Afterwards, 1 ll of aqueous solution of acridineorange/ethidium bromide (AO/EB, 100 lg/ml) was addedand the cell types were observed under a fluorescencemicroscope (Olympus, Tokyo, Japan). Three hundred cellswere counted per sample, being classified as follows: viablecells, apoptotic cells and necrotic cells.
2.3.4. DNA relaxation assay
The inhibitory effects of LP (5 or 10 lg/ml) on humanTopoisomerase I (Topo I) were measured using Topo IDrug Screening Kit (TopoGEN, Inc.). Supercoiled (FormI) plasmid DNA (250 ng or 500 ng) was incubated withhuman Topo I (4 units) at 37 �C for 30 min in relaxationbuffer (10 mM Tris buffer pH 7.9; 1 mM EDTA; 0.15 MNaCl; 0.1% BSA; 0.1 mM spermidine and 5% glycerol)with or without LP (5 or 10 lg/ml, final 20 ll). The reac-tion was stopped by adding of 10% SDS (2 ll) and proteaseK (50 lg/ml) and incubated at 37 �C for 30 min. DNAsamples were added to the loading dyes (2 ll) and subjected
Table 1Cytotoxic activity on tumoral cell lines of LP and PI untreated and after dige
Treatment Drug HL-60IC50 (lg/ml)
– Doxorubicin 0.020.01–0.03
Sole LP 0.860.72–1.04
PI 2.051.68–2.53
Pronase LP 5.70*
4.96–6.54PI 10.60**
8.81–12.75
DTT LP 1.801.57–2.07
PI 10.41**
7.09–15.29
2-Mercaptoethanol LP 3.87*
2.95–2.08PI 21.69**
18.82–23.50
Doxorubicin was used as positive control. Data are presented as IC50 values andduplicate, for leukemia (HL-60), colon (HCT-8), breast (MDA-MB-435) and bIC50 between different treatments.
* p < 0.05 comparing data obtained with LP sole with different treatments.** p < 0.05 comparing data obtained with PI sole with different treatments.
to electrophoresis on a 1% agarose gel for 90 min at roomtemperature, and visualized with ethidium bromide.
2.3.5. Statistical analysis
Data are presented as mean ± S.E.M from at least twoindependent experiments, performed in duplicate. TheIC50 values and their 95% confidence intervals (CI 95%)were obtained by a non-linear regression using a sigmoidaldose–response equation (variable slope). F-test was used tocompare the best-fit value of IC50 between different treat-ments (p < 0.05). Data from other experiments (PMBCproliferation, DNA synthesis and AO/BE staining) werecompared by one-way analysis of variance (ANOVA) fol-lowed by Student Newman–Keuls (p < 0.05). All analyseswere performed using the GRAPHPAD program (IntuitiveSoftware for Science, San Diego, CA).
3. Results
3.1. LP induces selective cytotoxicity on human tumor cell
lines
Several tumor cell lines were treated with increasing con-centrations of LP for 72 h and analyzed by the MTT assay.A significant suppression of cell growth was detected in thepresence of the proteins of C. procera latex, which occurredin a dose-dependent manner. Values of IC50 obtained after72 h of exposure are summarized in Table 1. Cytotoxicactivity was only weakly detected in PBMC after being trea-ted with LP. In PBMC exposed to LP (10 lg/ml) for 24, 48
stion by pronase or treatment with DTT or 2-mercaptoethanol
HCT-8 SF295 MDA-MD-435IC50 (lg/ml) IC50 (lg/ml) IC50 (lg/ml)
0.01 0.24 0.480.01–0.02 0.17–0.36 0.34–0.66
1.12 0.42 1.360.96–1.31 0.37–0.48 0.98–1.902.91 1.43 2.992.63–3.23 1.30–1.86 2.56–3.13
3.71* 7.27* 18.33*
2.54–5.42 5.43–9.73 15.75–21.328.24** 2.83** 10.64**
6.05–11.21 2.13–3.77 8.32–13.60
1.82 1.00 1.571.53–2.16 0.90–1.10 1.25–2.008.17** 7.27** 18.33**
6.27–10.65 5.43–9.73 15.75–21.32
2.90* 1.27* 5.12*
1.19–3.84 1.01–1.59 3.55–7.4015.00** 10.56** >2511.97–18.79 8.50–12.87
95% confidence interval from two independent experiments, performed inrain (SF295) cancer cells. F-test was used to compare the best-fit value of
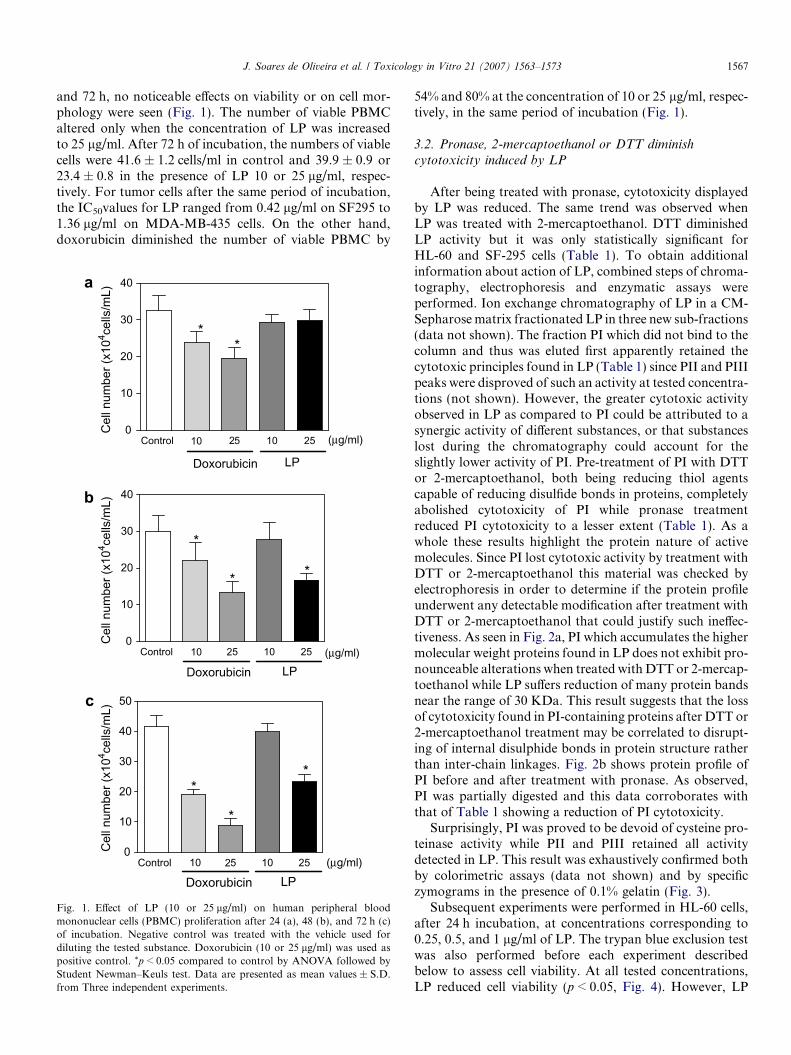
J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573 1567
and 72 h, no noticeable effects on viability or on cell mor-phology were seen (Fig. 1). The number of viable PBMCaltered only when the concentration of LP was increasedto 25 lg/ml. After 72 h of incubation, the numbers of viablecells were 41.6 ± 1.2 cells/ml in control and 39.9 ± 0.9 or23.4 ± 0.8 in the presence of LP 10 or 25 lg/ml, respec-tively. For tumor cells after the same period of incubation,the IC50values for LP ranged from 0.42 lg/ml on SF295 to1.36 lg/ml on MDA-MB-435 cells. On the other hand,doxorubicin diminished the number of viable PBMC by
Control 5 10 250
10
20
30
40
**
Doxorubicin
(μg/ml)
LP
Cel
l num
ber (
x104 ce
lls/m
L)C
ell n
umbe
r (x1
04 cells
/mL)
Cel
l num
ber (
x104 ce
lls/m
L)
Control 5 10 250
10
20
30
40
(μg/ml)
Doxorubicin
*
* *
Control 5 10 250
10
20
30
40
50
*
*
*
(μg/ml)
Doxorubicin LP
10 2
a
b
LP10
c
2
10 2
Fig. 1. Effect of LP (10 or 25 lg/ml) on human peripheral bloodmononuclear cells (PBMC) proliferation after 24 (a), 48 (b), and 72 h (c)of incubation. Negative control was treated with the vehicle used fordiluting the tested substance. Doxorubicin (10 or 25 lg/ml) was used aspositive control. *p < 0.05 compared to control by ANOVA followed byStudent Newman–Keuls test. Data are presented as mean values ± S.D.from Three independent experiments.
54% and 80% at the concentration of 10 or 25 lg/ml, respec-tively, in the same period of incubation (Fig. 1).
3.2. Pronase, 2-mercaptoethanol or DTT diminish
cytotoxicity induced by LP
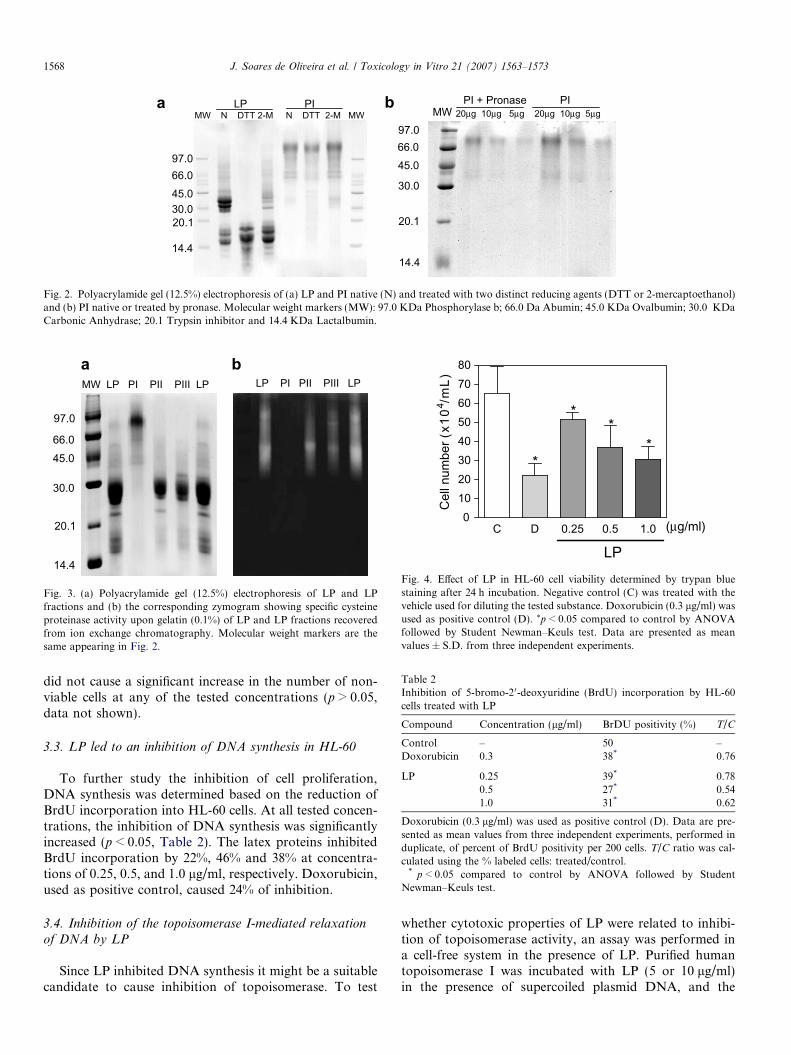
After being treated with pronase, cytotoxicity displayedby LP was reduced. The same trend was observed whenLP was treated with 2-mercaptoethanol. DTT diminishedLP activity but it was only statistically significant forHL-60 and SF-295 cells (Table 1). To obtain additionalinformation about action of LP, combined steps of chroma-tography, electrophoresis and enzymatic assays wereperformed. Ion exchange chromatography of LP in a CM-Sepharose matrix fractionated LP in three new sub-fractions(data not shown). The fraction PI which did not bind to thecolumn and thus was eluted first apparently retained thecytotoxic principles found in LP (Table 1) since PII and PIIIpeaks were disproved of such an activity at tested concentra-tions (not shown). However, the greater cytotoxic activityobserved in LP as compared to PI could be attributed to asynergic activity of different substances, or that substanceslost during the chromatography could account for theslightly lower activity of PI. Pre-treatment of PI with DTTor 2-mercaptoethanol, both being reducing thiol agentscapable of reducing disulfide bonds in proteins, completelyabolished cytotoxicity of PI while pronase treatmentreduced PI cytotoxicity to a lesser extent (Table 1). As awhole these results highlight the protein nature of activemolecules. Since PI lost cytotoxic activity by treatment withDTT or 2-mercaptoethanol this material was checked byelectrophoresis in order to determine if the protein profileunderwent any detectable modification after treatment withDTT or 2-mercaptoethanol that could justify such ineffec-tiveness. As seen in Fig. 2a, PI which accumulates the highermolecular weight proteins found in LP does not exhibit pro-nounceable alterations when treated with DTT or 2-mercap-toethanol while LP suffers reduction of many protein bandsnear the range of 30 KDa. This result suggests that the lossof cytotoxicity found in PI-containing proteins after DTT or2-mercaptoethanol treatment may be correlated to disrupt-ing of internal disulphide bonds in protein structure ratherthan inter-chain linkages. Fig. 2b shows protein profile ofPI before and after treatment with pronase. As observed,PI was partially digested and this data corroborates withthat of Table 1 showing a reduction of PI cytotoxicity.
Surprisingly, PI was proved to be devoid of cysteine pro-teinase activity while PII and PIII retained all activitydetected in LP. This result was exhaustively confirmed bothby colorimetric assays (data not shown) and by specificzymograms in the presence of 0.1% gelatin (Fig. 3).
Subsequent experiments were performed in HL-60 cells,after 24 h incubation, at concentrations corresponding to0.25, 0.5, and 1 lg/ml of LP. The trypan blue exclusion testwas also performed before each experiment describedbelow to assess cell viability. At all tested concentrations,LP reduced cell viability (p < 0.05, Fig. 4). However, LP
LP PI MW N DTT 2-M N DTT 2-M MW
14.4
20.130.045.066.097.0
PI + Pronase PI MW
14.4
20.1
30.0
45.0 66.097.0
20μg 10μg 5μg 20μg 10μg 5μga b
Fig. 2. Polyacrylamide gel (12.5%) electrophoresis of (a) LP and PI native (N) and treated with two distinct reducing agents (DTT or 2-mercaptoethanol)and (b) PI native or treated by pronase. Molecular weight markers (MW): 97.0 KDa Phosphorylase b; 66.0 Da Abumin; 45.0 KDa Ovalbumin; 30.0 KDaCarbonic Anhydrase; 20.1 Trypsin inhibitor and 14.4 KDa Lactalbumin.
14.4
20.1
30.0
97.0
45.066.0
MW LP PI PII PIII LP LP PI PII PIII LP
Fig. 3. (a) Polyacrylamide gel (12.5%) electrophoresis of LP and LPfractions and (b) the corresponding zymogram showing specific cysteineproteinase activity upon gelatin (0.1%) of LP and LP fractions recoveredfrom ion exchange chromatography. Molecular weight markers are thesame appearing in Fig. 2.
C D 0.25 0.5 1.00
1020304050607080
*
**
*
(μg/ml)
LP
Cel
l num
ber (
x104 /m
L)
Fig. 4. Effect of LP in HL-60 cell viability determined by trypan bluestaining after 24 h incubation. Negative control (C) was treated with thevehicle used for diluting the tested substance. Doxorubicin (0.3 lg/ml) wasused as positive control (D). *p < 0.05 compared to control by ANOVAfollowed by Student Newman–Keuls test. Data are presented as meanvalues ± S.D. from three independent experiments.
Table 2Inhibition of 5-bromo-20-deoxyuridine (BrdU) incorporation by HL-60cells treated with LP
Compound Concentration (lg/ml) BrDU positivity (%) T/C
Control – 50 –Doxorubicin 0.3 38* 0.76
LP 0.25 39* 0.780.5 27* 0.541.0 31* 0.62
Doxorubicin (0.3 lg/ml) was used as positive control (D). Data are pre-sented as mean values from three independent experiments, performed induplicate, of percent of BrdU positivity per 200 cells. T/C ratio was cal-culated using the % labeled cells: treated/control.
* p < 0.05 compared to control by ANOVA followed by StudentNewman–Keuls test.
1568 J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573
did not cause a significant increase in the number of non-viable cells at any of the tested concentrations (p > 0.05,data not shown).
3.3. LP led to an inhibition of DNA synthesis in HL-60
To further study the inhibition of cell proliferation,DNA synthesis was determined based on the reduction ofBrdU incorporation into HL-60 cells. At all tested concen-trations, the inhibition of DNA synthesis was significantlyincreased (p < 0.05, Table 2). The latex proteins inhibitedBrdU incorporation by 22%, 46% and 38% at concentra-tions of 0.25, 0.5, and 1.0 lg/ml, respectively. Doxorubicin,used as positive control, caused 24% of inhibition.
3.4. Inhibition of the topoisomerase I-mediated relaxation
of DNA by LP
Since LP inhibited DNA synthesis it might be a suitablecandidate to cause inhibition of topoisomerase. To test
whether cytotoxic properties of LP were related to inhibi-tion of topoisomerase activity, an assay was performed ina cell-free system in the presence of LP. Purified humantopoisomerase I was incubated with LP (5 or 10 lg/ml)in the presence of supercoiled plasmid DNA, and the
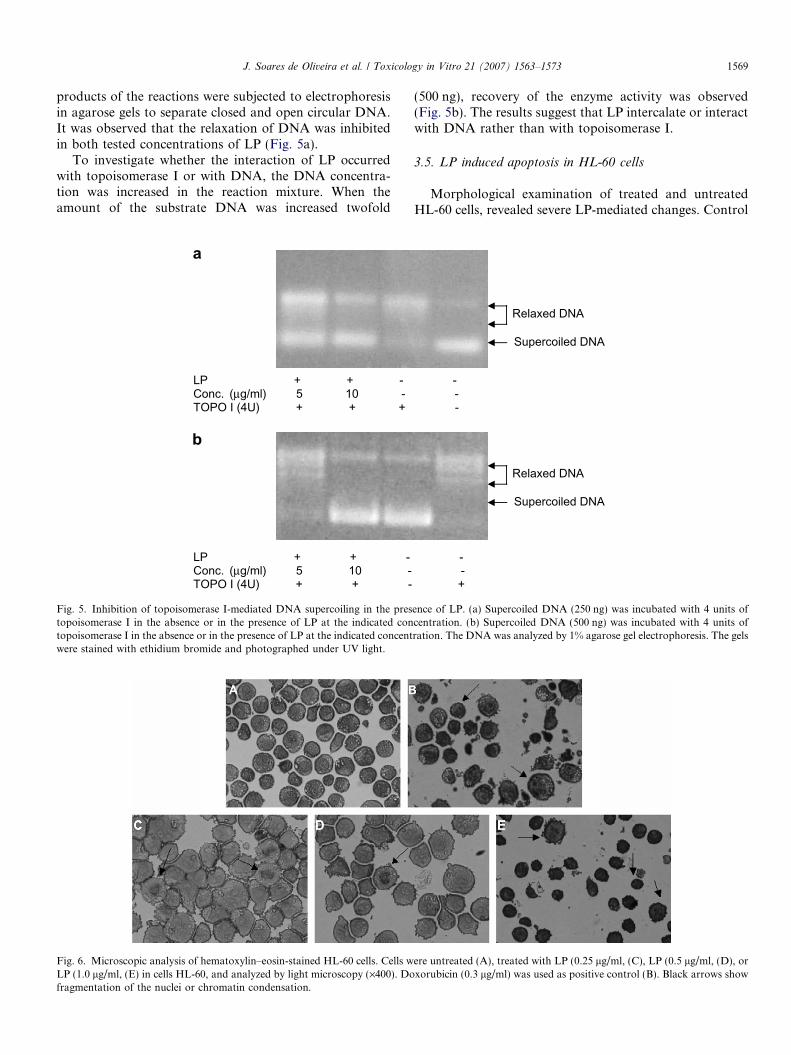
J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573 1569
products of the reactions were subjected to electrophoresisin agarose gels to separate closed and open circular DNA.It was observed that the relaxation of DNA was inhibitedin both tested concentrations of LP (Fig. 5a).
To investigate whether the interaction of LP occurredwith topoisomerase I or with DNA, the DNA concentra-tion was increased in the reaction mixture. When theamount of the substrate DNA was increased twofold
LP + + - Conc. (μg/ml) 5 10 - TOPO I (4U) + + +
LP + + - Conc. (μg/ml) 5 10 -TOPO I (4U) + + -
a
b
Fig. 5. Inhibition of topoisomerase I-mediated DNA supercoiling in the prestopoisomerase I in the absence or in the presence of LP at the indicated contopoisomerase I in the absence or in the presence of LP at the indicated concentwere stained with ethidium bromide and photographed under UV light.
Fig. 6. Microscopic analysis of hematoxylin–eosin-stained HL-60 cells. Cells wLP (1.0 lg/ml, (E) in cells HL-60, and analyzed by light microscopy (·400). Dofragmentation of the nuclei or chromatin condensation.
(500 ng), recovery of the enzyme activity was observed(Fig. 5b). The results suggest that LP intercalate or interactwith DNA rather than with topoisomerase I.
3.5. LP induced apoptosis in HL-60 cells
Morphological examination of treated and untreatedHL-60 cells, revealed severe LP-mediated changes. Control
- - -
Relaxed DNA
Supercoiled DNA
- - +
Relaxed DNA
Supercoiled DNA
ence of LP. (a) Supercoiled DNA (250 ng) was incubated with 4 units ofcentration. (b) Supercoiled DNA (500 ng) was incubated with 4 units ofration. The DNA was analyzed by 1% agarose gel electrophoresis. The gels
ere untreated (A), treated with LP (0.25 lg/ml, (C), LP (0.5 lg/ml, (D), orxorubicin (0.3 lg/ml) was used as positive control (B). Black arrows show
0
20
40
60
80
100
Viable cellsApoptotic cellsNecrotic cells
C D
*
*
*
*
*
*
(μg/ml)0.25 0.5 1.0
LP
Cel
l num
ber (
%)
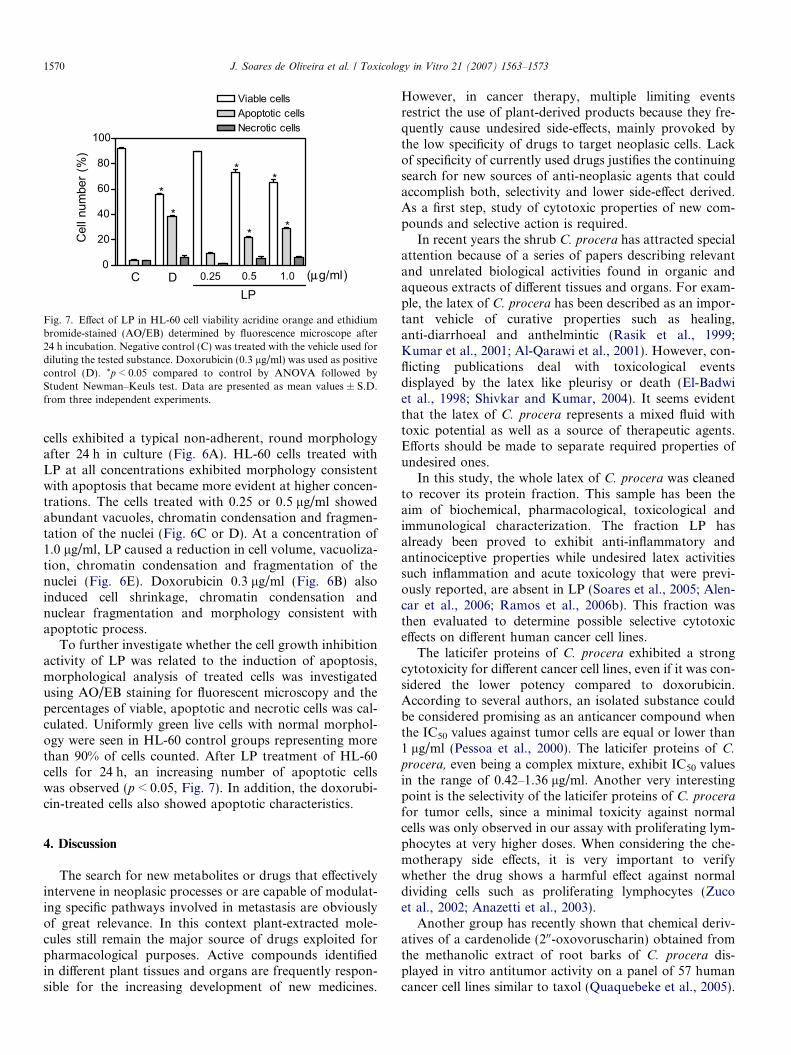
Fig. 7. Effect of LP in HL-60 cell viability acridine orange and ethidiumbromide-stained (AO/EB) determined by fluorescence microscope after24 h incubation. Negative control (C) was treated with the vehicle used fordiluting the tested substance. Doxorubicin (0.3 lg/ml) was used as positivecontrol (D). *p < 0.05 compared to control by ANOVA followed byStudent Newman–Keuls test. Data are presented as mean values ± S.D.from three independent experiments.
1570 J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573
cells exhibited a typical non-adherent, round morphologyafter 24 h in culture (Fig. 6A). HL-60 cells treated withLP at all concentrations exhibited morphology consistentwith apoptosis that became more evident at higher concen-trations. The cells treated with 0.25 or 0.5 lg/ml showedabundant vacuoles, chromatin condensation and fragmen-tation of the nuclei (Fig. 6C or D). At a concentration of1.0 lg/ml, LP caused a reduction in cell volume, vacuoliza-tion, chromatin condensation and fragmentation of thenuclei (Fig. 6E). Doxorubicin 0.3 lg/ml (Fig. 6B) alsoinduced cell shrinkage, chromatin condensation andnuclear fragmentation and morphology consistent withapoptotic process.
To further investigate whether the cell growth inhibitionactivity of LP was related to the induction of apoptosis,morphological analysis of treated cells was investigatedusing AO/EB staining for fluorescent microscopy and thepercentages of viable, apoptotic and necrotic cells was cal-culated. Uniformly green live cells with normal morphol-ogy were seen in HL-60 control groups representing morethan 90% of cells counted. After LP treatment of HL-60cells for 24 h, an increasing number of apoptotic cellswas observed (p < 0.05, Fig. 7). In addition, the doxorubi-cin-treated cells also showed apoptotic characteristics.
4. Discussion
The search for new metabolites or drugs that effectivelyintervene in neoplasic processes or are capable of modulat-ing specific pathways involved in metastasis are obviouslyof great relevance. In this context plant-extracted mole-cules still remain the major source of drugs exploited forpharmacological purposes. Active compounds identifiedin different plant tissues and organs are frequently respon-sible for the increasing development of new medicines.
However, in cancer therapy, multiple limiting eventsrestrict the use of plant-derived products because they fre-quently cause undesired side-effects, mainly provoked bythe low specificity of drugs to target neoplasic cells. Lackof specificity of currently used drugs justifies the continuingsearch for new sources of anti-neoplasic agents that couldaccomplish both, selectivity and lower side-effect derived.As a first step, study of cytotoxic properties of new com-pounds and selective action is required.
In recent years the shrub C. procera has attracted specialattention because of a series of papers describing relevantand unrelated biological activities found in organic andaqueous extracts of different tissues and organs. For exam-ple, the latex of C. procera has been described as an impor-tant vehicle of curative properties such as healing,anti-diarrhoeal and anthelmintic (Rasik et al., 1999;Kumar et al., 2001; Al-Qarawi et al., 2001). However, con-flicting publications deal with toxicological eventsdisplayed by the latex like pleurisy or death (El-Badwiet al., 1998; Shivkar and Kumar, 2004). It seems evidentthat the latex of C. procera represents a mixed fluid withtoxic potential as well as a source of therapeutic agents.Efforts should be made to separate required properties ofundesired ones.
In this study, the whole latex of C. procera was cleanedto recover its protein fraction. This sample has been theaim of biochemical, pharmacological, toxicological andimmunological characterization. The fraction LP hasalready been proved to exhibit anti-inflammatory andantinociceptive properties while undesired latex activitiessuch inflammation and acute toxicology that were previ-ously reported, are absent in LP (Soares et al., 2005; Alen-car et al., 2006; Ramos et al., 2006b). This fraction wasthen evaluated to determine possible selective cytotoxiceffects on different human cancer cell lines.
The laticifer proteins of C. procera exhibited a strongcytotoxicity for different cancer cell lines, even if it was con-sidered the lower potency compared to doxorubicin.According to several authors, an isolated substance couldbe considered promising as an anticancer compound whenthe IC50 values against tumor cells are equal or lower than1 lg/ml (Pessoa et al., 2000). The laticifer proteins of C.
procera, even being a complex mixture, exhibit IC50 valuesin the range of 0.42–1.36 lg/ml. Another very interestingpoint is the selectivity of the laticifer proteins of C. procera
for tumor cells, since a minimal toxicity against normalcells was only observed in our assay with proliferating lym-phocytes at very higher doses. When considering the che-motherapy side effects, it is very important to verifywhether the drug shows a harmful effect against normaldividing cells such as proliferating lymphocytes (Zucoet al., 2002; Anazetti et al., 2003).
Another group has recently shown that chemical deriv-atives of a cardenolide (200-oxovoruscharin) obtained fromthe methanolic extract of root barks of C. procera dis-played in vitro antitumor activity on a panel of 57 humancancer cell lines similar to taxol (Quaquebeke et al., 2005).

J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573 1571
These findings suggest that active anticancer moleculespresent in the latex of C. procera are products of secondarymetabolism. The results described here show that proteinsof latex of C. procera separated from low molecular sub-stances (lower than 8000 Da) and lacking rubber still exhi-bit very selective cytotoxic effects. Additional evidence forthis was obtained by observing the diminishing of cytotoxicactivity after proteolytic digestion with a non-specific pro-tease (pronase) and also treatment with reducing agents(DTT and 2-mercaptoethanol) which disrupt disulphidebonds involving cysteine residues in protein structure.The fact that LP was still effective after DTT treatmentwhile 2-mercaptoethanol reduced LP cytotoxicity, couldbe explained at least in part by the presence of cysteine pro-teinases that are very avid to DTT and could sequester itavoiding action of DTT upon proteins actually involvedin cytotoxicity as observed with PI. Kinetic of treatmentof LP with different concentrations of DTT prior to analy-sing cytotoxicity will be required to clarify this point.Another hypothesis takes in account the presence of morethan one molecule involved in cytotoxicity and this is cur-rently under investigation.
Although it is not yet possible to identify the putativeprotein involved in such cytotoxic activity, combined anal-ysis of chromatography, electrophoresis and cysteine pro-teinase activity have offered consistent information todiscuss the present data. Despite the elevated endogenouscysteine proteinase activity present in LP, evidence suggeststhat these proteinases are not involved in cytotoxic activityobserved, since PI from CM-Sepharose has retained almostcytotoxicity while it was rigorously absent of such an enzy-matic activity. The inhibition of cytotoxicity found aftertreatment of PI with DTT and 2-mercaptoethanol suggeststhat DTT and 2-mercaptoethanol induce structural disar-rangement in some proteins leading to loss of activity.According to the electrophoresis analysis of PI treatedand non-treated with DTT or 2-mercaptoethanol as shownin Fig. 2a, similar protein pattern seem to be present. Thissuggests that proteins involved in cytotoxic activity possessinternal (intra-chain) disulfide bonds instead of inter-chainassociation mediated by disulfide bonds. Additional purifi-cation steps are now being carried out in an attempt toidentify proteins involved in cytotoxicity.
Such antiproliferative effects were further investigated toreveal the mechanism of cytotoxic action exhibited by LP.DNA synthesis was affected after treatment with LP in allconcentrations, resulting in a lower number of cell divisionsthat corroborated with trypan blue exclusion. LP promptlyinhibits DNA synthesis in cancer cells likely due to inhibi-tion of topoisomerse I. Topoisomerse I acts by making atransient break in one DNA strand, allowing the DNAto swivel and release torsional strain, changing the linkingnumber by steps of one (Wang, 1996; Pommier et al.,1998). This enzyme is crucial for cellular genetic processessuch as DNA replication, transcription, recombination,and chromosome segregation at mitosis. It has long beenaccepted that topos are valuable targets of cancer chemo-
therapeutics (Chen and Liu, 1994; Wang, 1996; Pommieret al., 1998). Several classes of topo inhibitors have beenintroduced into cancer clinics as potent anticancer drugs,including camptothecin (CPT) derivatives (e.g., irinotecanand topotecan) (Pommier et al., 1998).
The inhibition mechanism of topoisomerase I by LP wasthe next issue to be investigated. The data presented herefavour an indirect interaction between LP and topoisomer-ase I enzyme. Consistent with this fact, the inhibition oftopoisomerase I activity by LP was reversed by increasingthe amount of DNA in the reaction mixture. This indicatesthat other mechanisms, such as DNA intercalating or inter-act with the DNA may be involved in LP activity.
Many studies have suggested that topoisomerase Iinhibitor induces cancer cell apoptosis (Wood and New-comb, 1999; Tabata et al., 2001). In fact, the treatment ofHL-60 cells with LP induced morphological alterationstypical of the apoptotic process, including reduction in cellvolume, vacuolization, chromatin condensation and DNAfragmentation (Hu and Kavanagh, 2003)
Inducing apoptosis in tumor cells is extremely desirablefor cancer chemotherapy (Los et al., 2003). Apoptosis isprogrammed cell death that involves genetically controlledmorphological and biochemical events, including external-izing phosphatidylserine, cytochrome c leakage from themitochondria, activating caspase, reducing cell and nuclearvolume, condensing chromatin, and fragmenting internu-cleosomal DNA (Schultz and Harrington, 2003).
Choedon and co-workers reported that the methanolicextract prepared of the dry latex of C. procera inducesextensive cell death accompanied by extensive fragmenta-tion of DNA while no changes in markers of apoptosis likeBcl2 and caspase 3 were observed. These parameters wereseen in a hepatoma (Huh-7) and non-hepatoma (COS-1)cell lines while non-cancerous (AML12) cells suffered onlya marginal effect, suggesting a high degree of selectivity(Choedon et al., 2006). The authors also reported thatthe aqueous extract of the dry latex of C. procera exhibiteda chemoprotective effect on transgenic mouse affected byhepatocellular carcinoma when oral doses correspondingto 400 mg/Kg were used for a period of 15 weeks.
LP represents a protein fraction and at the first momentthis could be a disadvantageous point if considering possi-ble use of LP as anticancer agent. However, accumulatingresults have reinforced very positive points associated tothis material. Regarding its toxicology, preliminary labora-tory results obtained suggest that LP does not produceacute effects on rats when administered by the oral routeeven with doses of 2500 mg/Kg. Rats given LP continu-ously for a period of 35 days did not exhibit behaviouralchanges. Death was not documented and the presence ofLP in faecal material was not detected by immunochemicalmethods (Ramos et al., 2006b). Furthermore, LP is free ofpro-inflammatory activity contradicting other literaturewhich report latex-mediating inflammation (Alencar et al.,2006). Instead, LP was shown to join anti-inflammatoryand antinociceptive activities (Alencar et al., 2004; Soares

1572 J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573
et al., 2005). These latter activities of LP now associated tothe selective in vitro cytotoxicity highlight a new status ofthe material in view of its use as a phytotherapic agent.As a last point, we have shown that LP does not induceallergy in rats when doses are offered orally (Ramos et al.,2006c). LP and PI are now been evaluated to their possiblein vivo anticancer activity with promising results.
After all, it should be emphasized that use of this rub-ber-free fraction as well as the investigation of the mecha-nism of action and molecules involved in the selectivecytotoxic activity represent considerable advances in char-acterizing sample and active molecules.
Acknowledgements
Studies of biochemical, functional and applied proper-ties of the latex from C. procera have been supported bygrants of the following Agencies: MCT/PADCT, CNPq,FUNCAP, RENORBIO, FINEP and CAPES from Braziland International Foundation for Science – IFS fromSweden (M.V.R.).
References
Abe, M., Abe, K., Kuroda, M., Arai, S., 1992. Corn kernel cysteineproteinase inhibitor as a novel cystatin superfamily member of plantorigin. Molecular cloning and expression studies. European Journal ofBiochemistry 209, 933–937.
Alencar, N.M.A., Figueiredo, I.S.T., Vale, M.R., Bitencurt, F.S., Oliveira,J.S., Ribeiro, R.A., Ramos, M.V., 2004. Antiinflammatory effect of thelatex from Calotropis procera in three different experimental models:peritonitis, paw edema and hemorrhagic cystitis. Planta Medica 70,1144–1149.
Alencar, N.M.N., Oliveira, J.S., Mesquita, R.O., Lima, M.W., Vales,M.R., Freitas, J.P., Etchells, C.D.T., Ramos, M.V., 2006. Pro- andanti-inflammatory activities of the latex from Calotropis procera (Ait.)R. Br. are trigged by compounds fractionated by dialysis. Inflamma-tion Research 55, 559–564.
Al-Mezaine, H.S., Al-Rajhi, A.A., Al-Assiri, A., Wagoner, M.D., 2005.Calotropis procera (ushaar) keratitis. American Journal of Ophthal-mology 139, 199–202.
Al-Qarawi, A.A., Mahmoud, O.M., Sobaih, Haroun, E.M., Adam, S.E.,2001. A preliminary study on the anthelmintic activity of Calotropis
procera latex against Haemonchus contortus infection in Najdi sheep.Veterinary Research Communication 25, 61–70.
Anazetti, M.C., Melo, P.S., Duran, N., Haun, M., 2003. Comparativecytotoxicity of dimethylamide–crotonin in the promyelocytic leukemiacell line (HL60) and human peripheral blood mononuclear cells.Toxicology 188, 261–274.
Arya, S., Kumar, V.L., 2005. Antiinflammatory efficacy of extracts oflatex of Calotropis procera against different mediators of inflammation.Mediators of Inflammation 2005, 228–232.
Azarkan, M., Amrani, A., Nijs, M., Vandermeers, A., Zerhouni, S.,Smolders, N., Looze, Y., 1997. Carica papaya latex is a rich source of aclass II chitinase. Phytochemistry 46, 1319–1325.
Chen, A.Y., Liu, L.F., 1994. DNA topoisomerases: essential enzymes andlethal targets. Annual Review of Pharmacology and Toxicology 34,191–218.
Chitme, H.R., Chandra, M., Kaushik, S., 2004. Studies on anti-diarrhoealactivity of Calotropis gigantea R. Br. in experimental animals. Journalof Pharmacy and Pharmaceutical Sciences 7, 70–75.
Choedon, T., Mathan, G., Arya, S., Kumar, V.L., Kumar, V., 2006.Anticancer and cytotoxic properties of the latex of Calotropis procera
in a transgenic mouse model of hepatocellular carcinoma. WorldJournal of Gastroenterology 12, 2517–2522.
Dubey, V.K., Jagannadham, M.V., 2003. Procerain, a stable cysteineproteinase from the latex of Calotropis procera. Phytochemistry 62,1057–1071.
El-Badwi, Samia, M.A., Adam, S.E., Shigidi, M.T., Hapke, H.J., 1998.Studies on laticiferous plants: toxic effects in goats of Calotropis
procera latex given by different routes of administration. DtschTierarztl Wochenschr 105, 425–427.
Hu, W., Kavanagh, J.J., 2003. Anticancer therapy targeting the apoptoticpathway. The Lancet Oncology 4, 721–729.
Kumar, S., Dewan, S., Sangraula, H., Kumar, V.L., 2001. Anti-diarrhoealactivity of the latex of Calotropis procera. Journal of Ethnopharma-cology 76, 115–118.
Kumar, V.L., Roy, S., Sehgal, R., Padhy, B.M., 2006. A comparativestudy on the efficacy of rofecoxib in monoarticular arthritis induced bylatex of Calotropis procera and Freund’s complete adjuvant. Inflam-mopharmacology 14, 17–21.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assemblyof the bacteriophage t4. Nature 227, 680–685.
Los, M., Burek, C.J., Stroh, C., Benedyk, K., Hug, H., Mackiewicz, A.,2003. Anticancer drugs of tomorrow: apoptotic pathways as target fordrug design. Drug Discovery Today 8, 67–77.
Macedo, M.L., Freire, M.D., Parra Jr., 2004. A Kunitz-type inhibitor ofcoleopteran proteinases, isolated from Adenanthera pavonina L. seedsand its effect on Callosobruchus maculatus. Journal of Agricultural andFood Chemistry 52, 2533–2540.
Mosmann, T., 1983. Rapid colorimetric assay for cellular growth andsurvival: application to proliferation and cytotoxicity assays. Journalof Immunological Methods 16, 55–63.
Nallamsetty, S., Kundu, S., Jagannadham, M.V., 2003. Purification andbiochemical characterization of a highly active cysteine proteinaseErvatamin A from latex of Eryvatamia coronaria. Journal of ProteinChemistry 22, 1–13.
Pessoa, C., Silveira, E.R., Lemos, T.G.L., Wetmore, L.A., Moraes, M.O.,Leyva, A., 2000. Antiproliferative effects of compounds derived fromplants of northeast Brazil. Phytotherapy Research 14, 187–191.
Pommier, Y., Pourquier, P., Fan, Y., Strumberg, D., 1998. Mechanism ofaction of eukaryotic DNA topoisomerase I and drugs targeted to theenzymes. Biochimica et Biophysica Acta 1400, 83–105.
Quaquebeke, E.V., Simon, G., Andre, A., Dewelle, J., El Yazidi, M.,Bruyneel, F., Tuti, J., Nacoulma, O., Guissou, P., Decaestecker, C.,Braekman, J.C., Kiss, R., Darro, F., 2005. Identification of a novelcardenolide (200-oxovoruscharin) from Calotropis procera and thehemisynthesis of novel derivatives displaying potent in vitro antitumoractivities and high in vivo tolerance: structure–activity relationshipanalyses. Journal of Medical Chemistry 10, 849–856.
Ramos, M.V., Bandeira, G.P., Freitas, C.D.T., Nogueira, N.A.P.,Alencar, N.M.N., Sousa, P.A.S., Carvalho, A.F.U., 2006a. Latexconstituents from Calotropis procera (R. Br.) display toxicity upon egghatching and larvae of Aedes aegypti (Linn). Memorias do InstitutoOswaldo Cruz 101, 503–510.
Ramos, M.V., Aguiar, V.C., Xavier, A.A.S., Lima, M.W., Bandeira, G.P.,Etchells, J.P., Nogueira, N.A.P., Alencar, N.M.N., 2006b. Latexproteins from the plant Calotropis procera are partially digested uponin vitro enzymatic action and are not immunologically detected in fecalmaterial. Fitoterapia 77, 251–256.
Ramos, M.V., Aguiar, V.C., Melo, V.M.M., Mesquita, R.O., Silvestre,P.P., Oliveira, J.S., Oliveira, R.S.B., Macedo, N.M.R., Alencar,N.M.N., 2006c. Immunological and allergenic responses induced bylatex fractions of Calotropis procera (Ait.) R. Br. Journal of Ethno-pharmacology 2006c. doi:10.1016/j.jep.2006.10.03.
Rasik, A.M., Raghubir, R., Gupta, A., Shukla, A., Dubey, M.P.,Srivastava, S., Jain, H.K., Kulshrestha, D.K., 1999. Healing potentialof Calotropis procera on dermal wounds in guinea pigs. Journal ofEthnopharmacology 15, 261–266.
Rocha, D.D., Militao, G.C.G., Veras, M.L., Pessoa, O.D.L., Silveira,E.R., Alves, A.P.N.N., Moraes, M.O., Pessoa, C., Costa-Lotufo, L.V.,

J. Soares de Oliveira et al. / Toxicology in Vitro 21 (2007) 1563–1573 1573
2006. Seletive cytotoxicitiy of withaphysalins in myeloid leukemia celllines versus peripheral blood mononuclear cells. Life Science 79, 1692–1701.
Roy, S., Sehgal, R., Padhy, B.M., Kumar, V.L., 2005. Antioxidant andprotective effect of latex of Calotropis procera against alloxan-induceddiabetes in rats. Journal of Ethnopharmacology 102, 470–473.
Schultz, D.R., Harrington, W.J., 2003. Apoptosis: programmed cell deathat molecular level. Seminars in Arthritis and Rheumatism 32, 345–369.
Shivkar, Y.M., Kumar, V.L., 2004. Effect of anti-inflammatory drugs onpleurisy induced by latex of Calotropis procera in rats. Pharmacolog-ical Research 50, 335–340.
Soares, P.M., Lima, S.R., Matos, S.G., Andrade, M.M., Patrocinio, M.C.,de Freitas, C.D., Ramos, M.V., Criddle, D.N., Cardi, B.A., Carvalho,K.M., Assreuy, A.M., Vasconcelos, S.M., 2005. Antinociceptiveactivity of Calotropis procera latex in mice. Journal of Ethnopharma-cology 99, 125–129.
Tabata, M., Tabata, R., Grabowski, D.R., Bukowski, R.M., Ganapathi,M.K., Ganapathi, R., 2001. Role of NF-kB and 26 S proteosome inapoptotic cell death induced by topoisomerase I and II poisons inhuman nonsmall cell lung carcinoma. Journal of Biological Chemistry276, 8029–8036.
Wang, J.C., 1996. DNA topoisomerases. Annual Review of Biochemistry65, 635–692.
Wood, D.E., Newcomb, E.W., 1999. Caspase-dependent activation ofcalpain during drug-induced apoptosis. Journal of Biological Chem-istry 274, 8309–8315.
Yeang, H.Y., Arif, S.A.M., Yusof, F., Sunderasan, E., 2002. Allergenicproteins of natural rubber latex. Methods 27, 32–45.
Zuco, V., Supino, R., Righetti, S.C., Cleris, L., Marchesi, E., Passerine-Gambacorti, C., Formelli, F., 2002. Selective cytotoxicity of betulinicacid on tumor cell lines, but not on normal cells. Cancer Letters 175,17–25.