impact of seasonal changes on fungal diversity of a semiarid ecosystem revealed by 454...
TRANSCRIPT

FEMS Microbiology Ecology, 91, 2015, fiv044
doi: 10.1093/femsec/fiv044Advance Access Publication Date: 15 April 2015Research Article
RESEARCH ARTICLE
Impact of seasonal changes on fungal diversity of asemi-arid ecosystem revealed by 454 pyrosequencingLluvia Vargas-Gastelum1, Adriana L. Romero-Olivares2, Ana E. Escalante3,Axayacatl Rocha-Olivares4, Carlos Brizuela5 and Meritxell Riquelme1,∗
1Department of Microbiology, Center for Scientific Research and Higher Education of Ensenada (CICESE), Ctra.Ensenada-Tijuana No. 3918, Ensenada, Baja California, 22860, Mexico, 2Department of Ecology andEvolutionary Biology, University of California-Irvine, 321 Steinhaus Hall, Irvine, CA 92697, USA, 3NationalLaboratory of Sustainability Sciences (LANCIS) Ecology Institute, National Autonomous University of Mexico(UNAM), Tercer Circ. Ext. next to Jardın Botanico, Ciudad Universitaria, 70275, Mexico, 4Department ofBiological Oceanography, Center for Scientific Research and Higher Education of Ensenada (CICESE), Ctra.Ensenada-Tijuana No. 3918, Ensenada, Baja California, 22860, Mexico and 5Department of Computer Sciences,Center for Scientific Research and Higher Education of Ensenada (CICESE), Ctra. Ensenada-Tijuana No. 3918,Ensenada, Baja California, 22860, Mexico∗Corresponding author: Department of Microbiology, Center for Scientific Research and Higher Education of Ensenada (CICESE), Ctra. Ensenada-TijuanaNo. 3918, Ensenada, Baja California, 22860, Mexico. Tel: +51-646-1750500. E-mail: [email protected] sentence summary: The fungal diversity and soil physicochemical characteristics of a semi-arid ecosystem in Baja California, Mexico, endemic forValley Fever, were analyzed.Editor: Ian C. Anderson
ABSTRACT
Fungi play fundamental ecological roles in terrestrial ecosystems. However, their distribution and diversity remain poorlydescribed in natural communities, particularly in arid and semi-arid ecosystems. In order to identify environmental factorsdetermining fungal community structure in these systems, we assessed their diversity in conjunction with soilphysicochemical characteristics in a semi-arid ecosystem in Baja California, Mexico, endemic for Coccidioidomycosis(Valley Fever). Two different microhabitats, burrows (influenced by rodent activity) and topsoil, were compared in winterand summer. Using a metagenomic approach, the ITS1 region of nuclear ribosomal DNA was used as barcode. A total of1940 Operational Taxonomic Units (OTUs) were identified from 362 332 ITS1 sequences obtained by 454 pyrosequencing.Differences in fungal composition between seasons were clearly identified. Moreover, differences in composition betweenmicrohabitats were mainly correlated to significant differences in environmental factors, such as moisture and clay contentin topsoil samples, and temperature and electrical conductivity in burrow samples. Overall, the fungal communitystructure (dominated by Ascomycota and Basidiomycota) was less variable between seasons in burrow than in topsoilsamples. Coccidioides spp. went undetected by pyrosequencing. However, a nested PCR approach revealed its higherprevalence in burrows.
Keywords: mycobiota; molecular ecology; metagenomics; ITS1 region
Received: 12 December 2014; Accepted: 2 April 2015C© FEMS 2015. All rights reserved. For permissions, please e-mail: [email protected]
1

2 FEMS Microbiology Ecology, 2015, Vol. 91, No. 5
INTRODUCTION
Arid and semi-arid ecosystems cover one-third of the terrestrialsurface (Reynolds et al. 2007; Collins et al. 2008) and about 40% ofthe Mexican territory (Magana et al. 2004). The coverage of theseecosystems is increasing due to erosion resulting from anthro-pogenic activities, such as land use changes (Foley et al. 2005),as well as a consequence of climate change (Challenger andSoberon 2008). Despite its extensive coverage of the terrestrialsurface, little attention has been given to the study of arid andsemi-arid ecosystems, specifically to the soil processes and mi-crobial diversity of fungi and prokaryotes (Giri et al. 2005; Bates,Nash and Garcia-Pichel 2012).
The spatial and temporal distributions of microorganisms insoils are related to the nutritional and physicochemical featuresof their habitat, such as organic matter content (Buee et al. 2007;Burke et al. 2009), pH (Fierer and Jackson 2006), aggregate size(Huxman et al. 2004; Noguez et al. 2008; Lauenroth and Brad-ford 2012), water content and temperature, which also influencetheir survival and dispersal (Whitford 1996). Specifically, the ac-tivity and diversity of fungi in arid and semi-arid ecosystemsare highly influenced by exposure to optimal temperature andthe frequency and intensity of precipitation; even small pre-cipitation events can influence primary productivity, increas-ing carbon input and thus, decomposition (Belnap et al. 2005;Zak 2005). Climate change forecast models suggest that arid andsemi-arid ecosystems will experience temperature increases of0.5–2◦C (IPCC 2008) as well as fewer precipitation events in thefollowing century (Easterling et al. 2000; Bell et al. 2009). Thiscould affect the diversity and function of fungal communities inarid and semi-arid soils, since water dependence ismore criticalin these dry ecosystems (Zak 2005). In fact, recent studies havedemonstrated that in Mexico, temperatures are rising (Pavia,Graef and Reyes 2009) and precipitation patterns are changing(Arriaga-Ramırez and Cavazos 2010), but the response of fungalcommunities to these changes is not yet well documented. Pre-vious studies have suggested that variation in soil fungal diver-sity of several arid and semi-arid ecosystems of Baja Californiais related to differences in climatic conditions (Romero-Olivareset al. 2013), suggesting that community structure and diversityare under environmental control.
In microbial communities, the fungal composition has beenfar less studied than the bacterial composition (Tedersooet al. 2014). Classic fungal taxonomic characterization is highlyunattainable, partly because many species require specializedsampling strategies (Cannon 1999); because culturing of indi-vidual species has been difficult or impossible for some groupsand also because of the existence of a large number of cryp-tic species (Jones and Richards 2011). More recently, the use ofmolecular techniques has enabled culture-independent studiesof the fungal component of microbial communities by directisolation of genomic DNA from soil samples followed by am-plification and sequencing of fungal ITS (Internal TranscribedSpacer) variable regions contained in the ribosomal DNA (rDNA).Furthermore, the increasing progress and application of mas-sive next-generation sequencing techniques, has provided thenecessary tools for the characterization of fungal communitiesfrom complex soil samples. Currently, fungal ecological stud-ies based on next-generation sequencing have significantly in-creased the detection power to estimate more accurately the ac-tual levels of diversity of microbial communities in soil ecosys-tems. For instance, studies based on pyrosequencing have iden-tified over ‘1130 potentially novel taxa’ (Hibbett, Ohman andKirk2009), which exceeds the total number of species of Dikarya and
Glomeromycota described in the Index of Fungi up to 2008(Center for Agricultural Bioscience International) (Buee et al.2009; Hibbett, Ohman and Kirk 2009; Jumpponen and Jones 2009;Opik et al. 2009). This suggests that a vast number of fungalspecies remain undescribed.
High-throughput amplicon pyrosequencing has been used toanalyze fungal diversity of forest samples (Buee et al. 2009; Jump-ponen and Jones 2009; Opik et al. 2009; Monard, Gantner andStenlid 2013) tundra (Lentendu et al. 2011), compost (De Gannes,Eudoxie and Hickey 2013), grassland (Dumbrell et al. 2011) andMediterranean soils of different land-use backgrounds (Orgiazziet al. 2012), but our understanding of the soil fungal diversity inarid and semi-arid ecosystems is more limited (Fierer et al. 2007;Porras-Alfaro et al. 2011; Oliveira et al. 2013; Romero-Olivares et al.2013).
In the present study, we describe the fungal diversity andcomposition of soil samples from a semi-arid ecosystem in Vallede las Palmas (VDP), Baja California, Mexico, identified as en-demic for Coccidioidomycosis or Valley Fever (Cairns et al. 2000).Previous studies in VDP have found the causal agent of the dis-ease, Coccidioides spp. in soil samples collected from two differ-ent microhabitats defined by their physical structure: one in-fluenced by rodent activity (burrows; Baptista-Rosas et al. 2012)and a second undisturbedmicrohabitat (topsoil; Catalan-Dibeneet al. 2014).
Here, we characterize the soil fungal community of VDP us-ing 454 pyrosequencing of the ITS region. One of the main goalsof this study was to test whether differences in environmentaland soil physicochemical variables of two microhabitats (bur-rows and topsoil) sampled in summer and winter would cor-relate with significant shifts in the diversity and compositionof the fungal communities. An additional goal of this metage-nomic approach was to investigate if a specific fungal commu-nity profile associated to Coccidioides spp. endemic sites could beidentified.
MATERIALS AND METHODSStudy site and soil sampling
VDP is a semi-arid ecosystemwith Mediterranean climate, char-acterized by hot and dry summers and cold winters, duringwhich most annual precipitation takes place (total annual pre-cipitation: 209 mm; Romero-Olivares et al. 2013), and with atemperature ranging between 4–35◦C (minimum and maximummean annual air temperatures).
Sampling was carried out in summer (S; July 2012) and win-ter (W; January 2013). Sampling sites were selected randomly,but were directed to two microhabitats: one influenced by ro-dent activity (burrow—B), and the other undisturbed (topsoil—T)(Fig. 1, Fig. S1, (Supporting Information)). These points were geo-referenced with the help of a handheld GPS (Global PositioningSystem, precision of ± 14.7 m). The sampled area correspondsto 2.77 Ha (Catalan-Dibene et al. 2014), and the minimum andmaximum distance between samples was 40 and 145 m, respec-tively. Both summer and winter samples were collected fromthe same GPS positions; except for a few summer burrows thatwere closed or no longer active in winter. In those instances,new active burrows located as close as possible to the originalswere sampled. In order to obtain enough geneticmaterial to con-duct fungal diversity analyses and to characterize soil proper-ties, in each season 20 samples of approximately 500 g were col-lected, 10 samples for eachmicrohabitat. (i.e. TS topsoil summer,BS burrow summer, TW topsoil winter and BW burrow winter).

Vargas-Gastelum et al. 3
Figure 1. Study site: Valle de las Palmas. (a) Location of the study site in Baja California (Google earth 7.1, 2014). (b)Study site in summer; note burrow and topsoilmicrohabitats. (c) Study site in winter; note burrow and topsoil microhabitats.
Burrow samples were collected near the burrow entrance (at adepth of about 15 cm inside the burrow), while topsoil sampleswere collected from 10–15 cm below the surface. For each con-dition (microhabitat and season), each sample was considereda replicate (e.g. 1SS, 2TS . . . 10TS), and the 500 g was dividedinto two sub-samples for physicochemical and molecular anal-yses, which were stored at ambient temperature and at −80◦C,respectively.
Soil physicochemical analyses
For each sample, several parameters were determined either insitu or in the lab. Soil temperaturewasmeasured in situwith sen-sors (Theta Probe type ML2x) coupled to an Infra Red Gas Ana-lyzer (IRGA, LI-8100, Licor, Lincoln, NE). Using 10 g of soil, totalmoisture was determined by gravimetric analysis. Organic mat-ter content was determined by ignition loss weight using 10 g ofsoil and placed in a muffle at 500◦C for 8 h until obtaining con-stant weight. pH was determined using 20 ml of water and 10 gsoil with a pHmeter (Corning Pinnacle M530 pHmeter, Woburn,MA USA; precision ± 0.01). Electrical conductivity wasmeasuredusing 100 g of soil and the Field Scout Direct Soil EC Meter (Spec-trum Technologies, Inc.). Finally, texture was determined by theBouyoucos method using 60 g of soil (Bouyoucos 1936).
To visualize spatial variation in temperature and moistureacross sample points, contour maps were drawn with the pro-gram surfer 10.1.561 (Golden Software Inc., Golden, Colorado,USA).
DNA extraction, pyrosequencing and data processing
Genomic DNA was extracted from 0.25 g of soil using the PowerSoil DNA Extraction Kit (Mo Bio Laboratories Inc., Carlsbad, CA,
USA) and following the manufacturer´s instructions. To maxi-mize extraction yield, DNA was eluted in 50 μl of elution buffer.Genomic DNA was adjusted to 20 ng μl−1, and sent to Re-search and Testing Laboratory (Lubbock, Texas, USA) for ampli-fication of the entire ITS region (comprising partial 18S rRNA,ITS1, 5.8 S rRNA, ITS2, and partial 28S rRNA genes) and pyrose-quencing. The ITS region was amplified using primers ITS1-F(5´-CTTGGTCATTTAGAGGAAGTAA-3´; Gardes and Bruns 1993)and ITS4 (5´-TCCTCCGCTTATTGATATGC-3´; White et al. 1990)and sequenced by Roche 454 FLX with an average coverageof 5X. Each sample was de-multiplexed from the resulting sfffile based upon the eight-base barcode attached to each of thesamples during PCR, using the Roche sff software. De-noisingand chimera analysis were conducted with the AmpliconNoise(Quince et al. 2011) and UCHIME algorithms (Edgar et al. 2011).
After sequence clean up, ITS1 sequences were obtained byremoving unwanted sequences (18S, 5.8S, ITS2 and 28S) usingthe Fungal ITS Extractor (Nilsson et al. 2010). Resulting ITS1 se-quences were further processed using the Quantitative Insightsinto Microbial Ecology (QIIME; Caporaso et al. 2010). Quality con-trol involved keeping sequences no shorter than 100 bp and nolonger than 250 bp with a quality score higher than 25. The re-maining sequences were used for subsequent analysis.
A 97% identity threshold was used to cluster sequences intoOperational Taxonomic Units (OTUs) using the UCLUST algo-rithm (Edgar 2010). The OTUs represented by one sequence (sin-gletons) were removed. The taxonomic assignment (to specieslevel) was carried out using the BLAST algorithm (Altschul et al.1990) against the UNITE fungal database v5.0 (Abarenkov et al.2010). In order to establish differences between samples, a fre-quency OTU table including the number of times that an OTU isfound in each sample (e.g. 1TS, 2TS, . . . , 10TS) and the taxonomicidentification of each OTU, was obtained (data not shown).

4 FEMS Microbiology Ecology, 2015, Vol. 91, No. 5
Statistical analyses
Environmental variables. In order to identify significant differ-ences of soil physicochemical characteristics between micro-habitats and seasons, a repeated measure ANOVA, followed bymultiple pairwise comparison using Tukey’s test at α = 0.05 withStatistica 10 software (Statsoft; Tulsa, OK, USA) were conducted.
OTU diversity analyses. Diversity was estimated as theShannon–Wiener (H´) and Simpson (D´) indexes as implementedin EstimateS v9 (Colwell 2013). To compare levels of OTU rich-ness among samples, we used the non-parametric Chao 1 esti-mator. Due to the different sequencing depth per sample andto distinguish differences in OTUs richness between sampletypes and sequencing effort, a rarefaction analysis was carriedout using Ecosim v7 (Entsminger 2012) to an even sequenc-ing depth of 48 305, which represents the total number of se-quences obtained for TS samples (lower than the total numberof sequences obtained for TW, BS and BW; Table S2, SupportingInformation).
OTU composition analysis. To visually identify patterns ofcommunity structure among the samples, we constructed aheatmap based on the abundance of ordinal taxa, with dou-ble hierarchical dendrograms computed usingWard’s clusteringalgorithm applied to matrices of Euclidean distances of sam-ples across taxa and taxa across samples. A repeated measuresMANOVA followed by Bonferroni-corrected P-test (P < 0.05 prob-ability level) was conductedwith SYSTAT 13.1 (Evanston, Illinois,USA) in order to identify differences in community compositionamong microhabitats. To identify the influence of soil physic-ochemical characteristics on the fungal community of VDP, aCanonical Correspondence Analysis (CCA) was performed by us-ing the OTU table and the soil physicochemical characteristicsper replicate (e.g. 1BS, 2BS, 3BS, containing presence/absencedata). Only the variables that displayed significant differencesamong treatments (P < 0.05) were included in the ordinationplot. Both analyses were carried out using the BiodiversityRpackage in R v.3.01 (R Development Core Team, 2008).
To analyze the fungal community structure among samples,a non-metric multidimensional scaling (NMDS) analysis and ananalysis of variance (ANOVA) using the ANOSIM function wereperformed, as implemented in MASS (Ripley et al. 2013) and Ve-gan (Oksanen et al. 2015) packages in R v.3.01 (R DevelopmentCore Team, 2008), respectively.
Finally, we used a Mantel test to assess the influence of spa-tial distance on the similarity of fungal communities by corre-latingmatrices of geographic and ecological distances; the anal-yses was performed out by using the Ade4 package (Dray andDufour 2007) in R v.3.01 (R Development Core Team, 2008).
Molecular detection of Coccidioides spp.
Nested PCR conditions. The molecular detection of Coccidioidesspp. in soil samples required a nested PCR approach. First,the entire ITS1–5.8S-ITS2 (≈900 bp) rDNA region was amplifiedusing primers specific for the fungal sub-kingdom Dikarya,forward NSI1: 5′-GATTGAATGGCTTAGTGAGG-3′ and reverseNLB4: 5′-GGATTCTCACCCTCTATGAC-3′ as previously described(Martin and Rygiewicz 2005; Baptista-Rosas et al. 2012). Neu-rospora crassa DNA and HPLC water were used as positive andnegative controls, respectively. Thermal cycling (MJ MiniTM Gra-dient, Bio Rad) consisted of an initial denaturation at 95◦C for 2min, 35 cycles of 95◦C for 30 s, 62◦C for 40 s and 72◦C for 1 min,followed by a final elongation to 72◦C for 5 min. Second, 1 μl of a1:10 dilution of the previous PCR product was used as template
of a nested PCR, testing two sets of primers. For the amplificationof the ITS2 region, we used the primers of Binnicker et al. (2007)(forward ITS2F: 5′-CGAGGTCAAACCGGATA-3′ and reverse ITS2R:5′-CCTTCAAGCACGGCTT-3′) as described in previous studies(Baptista-Rosas et al. 2012; Catalan-Dibene et al. 2014). In addi-tion, we used newly designed primers to amplify the ITS1 regionin order to improve the specificity toward Coccidioides spp. (for-ward ITS1CF: 5′-GTGGCGTCCGGCTGCGCACCTCCCCCGCGG-3and reverse ITS1CR: 5′-GCGCAAGGCGGGCGATCCCCGGCAGCC-3) (Fig. S4, Supporting Information). Thermal cycling consistedof an initial denaturation at 95◦C for 2 min, followed by 30 cyclesof 95◦C for 30 s, 70◦C for 40 s and 72◦C for 1 min and a finalelongation step at 72◦C for 5 min.
Nested PCR products were confirmed by 2% agarose gel elec-trophoresis. DNA bands of the expected size were purified byQiaquick Gel Extraction Kit (Qiagen, German-town, MD, USA)before sequencing (Eton Bioscience Inc., San Diego, CA, USA).BLASTn (NCBI, USA) searches were used to determine the tax-onomic identity of ITS sequences.
RESULTS
The composition of the fungal community showed seasonalvariation with marked differences in the topsoil microhabi-tat, while in burrows the fungal community was more stablethrough seasons. No remarkable differences betweenmicrohab-itats within the same season were found.
Soil temperature and moisture differed betweenseasons but not between microhabitats
Significant differences in soil temperature and moisture werefound between summer (TS and BS) and winter (TW and BW)samples, with the highest temperature and lowest soil moisturein summer samples, and the lowest temperature and highestsoil moisture in winter samples (Table 1; Table S1; Figs S2 and 3,Supporting Information). In contrast, no significant differencesin organic matter content, pH, or electrical conductivity werefound among soil samples (i.e. TS, TW, BS and BW) (Table 1; TableS1, Supporting Information).
OTU richness and diversity is highest in winter
A total of 385 329 sequences were obtained through pyrose-quencing. After ITS1 extraction and quality control, 362 332 se-quences remained for subsequent analyses (Table S2, Support-ing Information). The number of sequences per sample variedfrom 1165 to 55 944 (Table S2, Supporting Information). A total of3057 OTUs were obtained at the 97% similarity threshold; fromthose, 1117 singletons were removed, and the remaining 1940OTUs were used for subsequent analysis.
Observed richness varied among microhabitats and seasons(Table 2). Using an even sequencing depth, rarefaction curvesshowed that BW samples presented the highest richness, whileno significant differences were observed among the TW, TS andBS curves (Fig. 2). Chao’s asymptotic richness estimator washigher for burrow than for topsoil in summer samples, whereasit was lower for burrow than for topsoil in winter samples andlarger differences between the number of observed OTUs, andthe Chao 1 values were observed for samples from winter thanfor summer (Table 2). No marked differences were observed forSimpson’s diversity between sample types. Samples collectedin winter displayed the highest values of Simpson’s diversity,with burrows being more diverse (Table 2). In contrast, topsoil

Vargas-Gastelum et al. 5
Table 1. Mean values (n = 10) of physicochemical characteristics of the soil samples.
Sample Moisture (%) Temperature (◦C)Organicmatter (%) pH
Electricalconductivity (mS) Texture
Clay (%) Silt (%) Sand (%)
TS 0.80 ± 0.38a 40.46 ± 4.60a 2.03 ±0.49a
5.92 ± 0.17a 0.39 ± 0.09a 23.08 20.68 56.24
BS 0.71 ± 0.73a 36.81 ± 4.18a 1.9 ± 1.12a 5.81 ± 0.29a 0.72 ± 0.61a 22.96 8.61 68.42TW 7.68 ± 3.39b 14.88 ± 1.62b 3.27 ±
2.16a5.82 ± 0.67a 1.05 ± 0.98a 15.12 14.13 71.02
BW 8.61 ± 2.62b 15.26 ± 3.29b 2.04 ±0.50a
5.75 ± 0.61a 1.24 ± 1.11a 10.2 18.73 70.57
Abbreviations: TS topsoil summer, BS burrow summer, TW topsoil winter and BW burrow winter.
Different letters (a and b) indicate significant differences between samples (P < 0.05) after a Tukey post hoc test.
Table 2. Richness and diversity indexes.
Index Samples
TS BS TW BW
Number of OTUs (Observed richness) 760 1088 962 974Chao 1 (estimated asymptotic richness)confidence intervals 95% (lower, upper)
1039 (1007, 1085) 1470 (1451, 1499) 1940 (1940, 1940) 1747 (1739, 1761)
Shannon—-Wiener 4.1 ± 0.32 4.33 ± 0.24 4.56 ± 0 4.49 ± 0.16Simpson 0.94 0.91 0.95 0.97
Abbreviations: TS topsoil summer, BS burrow summer, TW topsoil winter and BW burrow winter.
Figure 2. Rarefaction curves describing observed fungal richness among sample types (BW, TW, TS and BS). Graph displays the number of ITS1 OTUs to 97% similarityas a function of the number of sequences obtained. Error bars represent 95% confidence intervals. TS topsoil summer, BS burrow summer, TW topsoil winter and BWburrow winter.
samples were more diverse than burrows in summer sampling.Unlike the Simpson index, the Shannon–Wiener diversity indexdid not differ among the samples (Table 2).
Fungal community composition differed mostlythrough seasons
The taxonomic composition of the fungal community differedbetween seasons for eachmicrohabitat (Fig. 3; Table S3, Support-ing Information). The Ascomycota was the most abundant phy-lum in both microhabitats. The Chytridiomycota and the Mu-coromycotina were more prevalent in burrows than in topsoilsamples, whereas the Basidiomycota were more abundant in
topsoil samples than in burrows, and the Glomeromycota didnot show any specific pattern at the microhabitat level (Fig. 3a;Table S3, Supporting Information). The Ascomycota were moreabundant in summer than in winter. In contrast, the Basidiomy-cota, Chytridiomycota, Glomeromycota and Mucoromycotinawere more abundant in winter than in summer. Within the As-comycota (Fig. 3b; Table S3, Supporting Information), the pre-dominant orders in topsoil samples were Pleosporales, Helo-tiales and unidentified Ascomycota, whereas the Hypocrealesand Sordariales were more abundant in burrows. The Pleospo-rales and an unidentified order were themost abundant in sum-mer; whereas the Helotiales, Hypocreales and Sordariales wereso in winter.
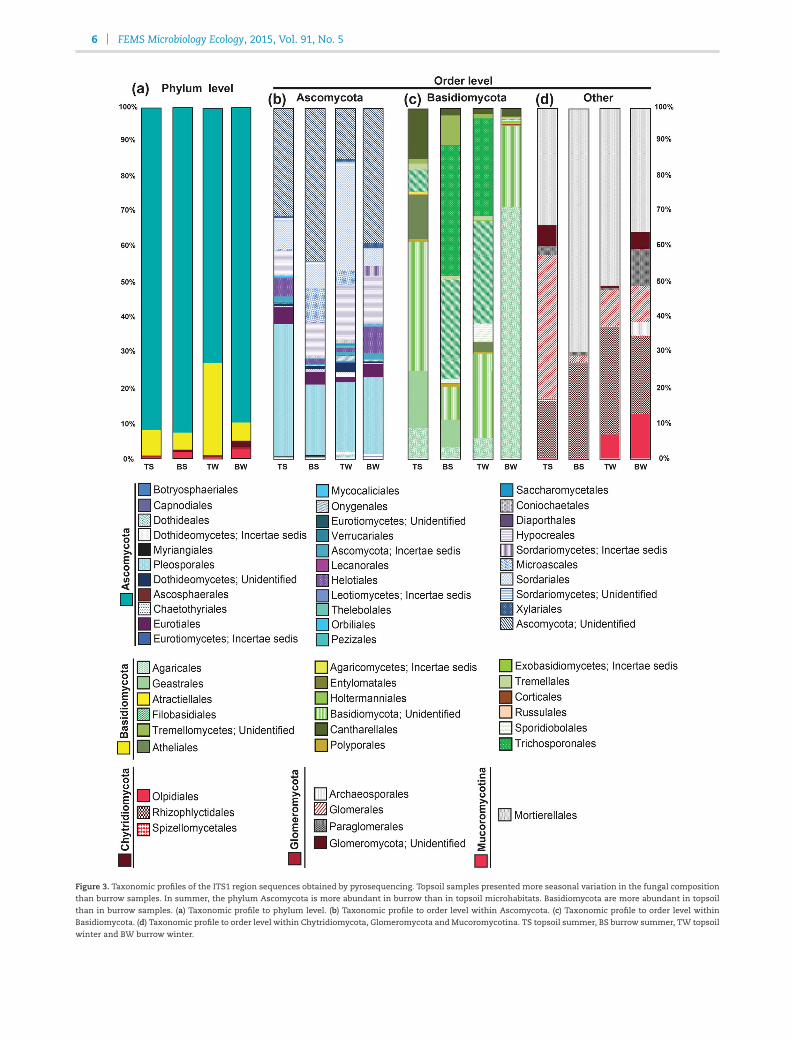
6 FEMS Microbiology Ecology, 2015, Vol. 91, No. 5
Figure 3. Taxonomic profiles of the ITS1 region sequences obtained by pyrosequencing. Topsoil samples presented more seasonal variation in the fungal compositionthan burrow samples. In summer, the phylum Ascomycota is more abundant in burrow than in topsoil microhabitats. Basidiomycota are more abundant in topsoilthan in burrow samples. (a) Taxonomic profile to phylum level. (b) Taxonomic profile to order level within Ascomycota. (c) Taxonomic profile to order level withinBasidiomycota. (d) Taxonomic profile to order level within Chytridiomycota, Glomeromycota and Mucoromycotina. TS topsoil summer, BS burrow summer, TW topsoil
winter and BW burrow winter.

Vargas-Gastelum et al. 7
Among the Basidiomycota (Fig. 3c; Table S3, Supporting In-formation), the topsoil community was dominated by Agari-cales, Atheliales, Cantharellales, Geastrales and unidentified Ba-sidiomycota, while burrows were dominated by Filobasidiales,Trichosporonales and unidentified Tremellomycetes. The or-ders Atheliales, Geastrales, Filobasidiales, Trichosporonales, theclass Tremellomycetes (unidentified order) and other uniden-tified Basidiomycota were the most abundant in summer;whereas the Agaricales and Cantharellales were the most abun-dant in winter.
Within the Chytridiomycota, Glomeromycota and Mu-coromycotina (Fig. 3d; Table S3, Supporting Information), Olpidi-ales, Glomerales, Paraglomerales and unidentified Glomeromy-cota were the most abundant in topsoil samples, while theRhizophyctidales andMortierellales were themost abundant or-ders in burrows. The most abundant orders in summer weretheGlomerales, unidentifiedGlomeromycota andMortierellales;the Olpidiales, Rhizophyctidales and Paraglomerales were moreabundant in winter.
The repeated measures MANOVA indicated that there wereno significant differences in fungal composition between sea-sons (summer and winter) (P = 0.855) nor between microhabi-tats (burrow and topsoil) (P = 0.276). However, the Bonferroni-corrected P-test showed significant differences between topsoilsamples across seasons (P= 0.026) and no significant differencesbetween burrow samples (P = 0.136).
The heatmap shows the abundances of fungi in the differ-ent samples (Fig. 4). Apart from a few exceptions, the summer(TS and BS) and winter (TW and BW) samples appeared overallclustered in distinct groups, whose differences are given primar-ily by the relative abundance of the Ascomycota, especially by anunidentified order, and the orders of Pleosporales and Sordari-ales.
Moisture, temperature, clay content and electricalconductivity influenced fungal community structure
We observed significant correlations between fungal commu-nity structure and soil moisture, temperature, electrical con-ductivity and clay content (P < 0.05; Fig. 5). These variables ex-plained 16.9% of the variation (CCA1 = 12.1 and CCA2 = 4.8).Moisture and clay content appeared more associated with top-soil samples, whereas temperature and electrical conductivitywith burrow samples (Fig. 5). Variation along the CCA1 axis dif-ferentiates the microhabitat: topsoil samples are located to theright and burrow samples to the left of the scatter plot (BS andBW) (Fig. 5). Moisture was highly positively correlated with CCA1(r = 0.94) and to a lesser extent with the CCA2 (r = 0.34); temper-ature was highly negatively correlated with CCA1 (r = −0.99) andmuch lesswith CCA2 (r= −0.07); electrical conductivitywas neg-atively correlated with the CCA1 (r = −0.98) and correlated posi-tively with CCA2 (r = 0.19) and the clay content is positively cor-related with CCA1 (r = 0.87) and negatively correlated with CCA2(r = 0.47).
Fungal community composition across samples variedprimarily depending on the season
The fungal community composition across samples varied sig-nificantly (r = 0.3403, P = 0.009; ANOSIM test). However, NMDS(stress = 3.39 after 100 iterations) showed that that even thoughsamples are scattered in the Cartesian plane they segregate bymicrohabitat (topsoil left and burrow right) and season (summerbottom andwinter top) (Fig. 6). TheMantel test revealed a lack of
significant correlation (P> 0.05) between geographic and ecolog-ical distances.
Coccidioides spp. was more frequent in burrow samples
All 11 ITS1 amplicons obtained were identified as Coccidioidesspp. (Table S4, Supporting Information). Nine of them corre-sponded to C. posadasii and two to C. immitis. Nine of these sam-ples were from burrows (summer and winter), while two werefrom topsoil (winter) (Table S4, Supporting Information). As forthe ITS2 amplicons, nine were identified as C. immitis, two asAphanoascus canadensis, one as Penicillium cyclopium and one as P.dipodomyicola (Table S4, Supporting Information).
DISCUSSION
We characterized the soil mycobiota in two microhabitats withcontrasting levels of biogenic disturbance (topsoil and burrows)and differing in soil properties. To our knowledge, this is thefirst study in Mexico that uses next generation sequencing forthe analysis of soil fungal diversity in a semi-arid ecosystem.The fungal community of VDP was diverse and included speciesranging from parasites and pathogens to lichenized fungi. Thenumber of ITS1 sequences obtained in this study (362 332 se-quences) was larger than reported in other studies conducted inforest (166 350 sequences; Buee et al. 2009), spruce forest (18 000sequences; Wallander et al. 2010) and alpine tundra (205 131 se-quences; Lentendu et al. 2011).
The pyrosequencing technology allowed us to find differ-ences in the fungal community composition between seasonsthat weremore evident in topsoil microhabitats. Species belong-ing to the phyla Basidiomycota, Glomeromycota and Chytrid-iomycota and the sub-phylum Mucoromycotina, were moreabundant in winter than in summer, suggesting that changes insoil characteristics (influenced by the season), have an effect onthe growth of those fungal taxa. Fungal groups that are not ther-mophilic (Porras-Alfaro et al. 2011), xerophilic (Dix and Webster1995), halophilic or osmophilic (Magan 2007), and some sapro-trophic fungi (Bell, McIntyre and Cox 2008) are presumably moresusceptible to environmental changes. When temperature in-creases, groups of fungi like yeast and molds (free-living sapro-trophs, including representatives of the Eurotiales, Mucorales,Mortieralles, Saccharomycetales and Tremellales) increase theirabundance, but when temperature decreases, there is a declinein groups of litter-associated fungi (litter degraders) and root-associated fungi (Skouw 2013).
Within each microhabitat significant differences in tem-perature and moisture between seasons were revealed. Thesedifferences are typical of seasonal variation in semi-arid ecosys-tems with Mediterranean climate such as VDP, with characteris-tic dry and hot summers and cooler andmoist winters. Seasonalchanges in these variables could influence the fungal commu-nity structure and function (Bell et al. 2009). The identified differ-ences in fungal compositionwere correlatedwith changes in soilmoisture and clay content in topsoil samples, and with changesin temperature and electrical conductivity in burrow samples.
Our rarefaction analyses suggested that the sequencing andsampling efforts were insufficient to reach a clear-cut asymp-totic plateau, indicating a better sampling coverage in summerthan in winter. The Chao 1 index depends on sampling size;therefore, we could obtain a higher number of OTUs by increas-ing the number of samples (Mohamed and Martiny 2011) orthe sequencing effort (Jumpponen, Jones and Blair 2010). TheShannon–Wiener values obtained in this study were similar to

8 FEMS Microbiology Ecology, 2015, Vol. 91, No. 5
Figure 4. Heatmap of the relative abundance of fungal orders in each sample and dual hierarchical dendrogram based on fungal orders. Top dendrogram: samples
dissimilarity. Left dendrogram: classes dissimilarity, not phylogenetic. Ecological distance: Euclidean. Hierarchical clustering algorithm: ward. TS topsoil summer, BSburrow summer, TW topsoil winter and BW burrow winter.

Vargas-Gastelum et al. 9
Figure 5. Canonical Correlation Analysis (CCA). Ordination diagram of the fungal community data together with soil variables: moisture and temperature (Temp),electrical conductivity (EC) and clay content. Only the vectors that were significant for the distribution of soil fungal community were indicated. TS topsoil summer,BS burrow summer, TW topsoil winter and BW burrow winter.
those obtained in other studies of temperate ecosystems (Jump-ponen, Jones and Blair 2010; Monard, Gantner and Stenlid 2013).Even though the Shannon–Wiener diversity values were not sig-nificantly different between seasons for both microhabitats, theSimpson index results suggest that diversity could be driven byseasonal differences in burrows.
The taxonomic composition of the fungal community in bur-rows presented less seasonal variation, than topsoil samples.The stability of the fungal burrow community could be furtherevidenced in the NMDS, where burrow samples cluster together,indicating that the fungal composition is stable independentlyof the sampling season. The most abundant phylotypes repre-sent 72% of total sequences (Table S5, Supporting Information).The fungal diversity found in this study expands previous re-sults for VDP, whereby utilizing a traditional cloning methodfollowed by Sanger sequencing (directed to the sub-kingdomDikarya), and 18 OTUs were identified (Romero-Olivares et al.2013). Most of those OTUswere assigned to the phylumAscomy-cota, including five classes and 12 orders, with Pleosporales be-ing the most abundant order. In this study, we obtained a totalof 1940 OTUs, most of them belonging to the phyla Ascomycota(10 classes and 32 orders), with unidentified Ascomycota andPleosporales being the most abundant, and Basidiomycota (sixclasses and 16 orders).
In general, the fungal composition of the different mi-crohabitats included lichenized members (e.g. Lecanorales),pathogens (e.g. Capnodiales and Microascales), saprobes (e.g.Saccharomycetales, Dothideales, Chaetothyriales, Eurotiales,
Helotiales, Xylariales and Filobasidiales), mutualistic root sym-bionts (e.g. Glomerales, Paraglomerales and Archaeosperales)and parasites (Pleosporales, Pezizales, Saccharomycetales,Hypocreales and Atheliales). In almost all samples, we identi-fied abundant phylotypes of plant pathogens such as Fusariumoxysporum, Colletotrichum circinans and Alternaria solani, as wellas human pathogens such as Trichosporon asahii and Aspergillusflavus (Table S6, Supporting Information). Given our samplingand sequencing strategies, definite assignment of ecologicalroles to specific taxa is not possible.
The CCA suggests that the fungal community of the topsoilsamples is particularly sensitive to changes inmoisture and claycontent. These soil characteristics are correlated, sincewater in-filtration depends on soil porosity (Pumpanen, Ilvesniemi andHari 2003). For example, topsoil samples, which are more ex-posed to wind than burrows, could present more variability intheir clay content because of the commonwind erosion in semi-arid ecosystems (Ravi et al. 2004). The decrease of clay contentturns the soil into a porousmicrohabitat, whichwould allowwa-ter to infiltrate faster influencing changes in soil temperature. Inburrow samples, temperature and electrical conductivity influ-ence, apparently, fungal composition. Mammals of active bur-rows maintain a constant temperature (Siminski 2014); there-fore, this biogenic source of heat may provide a more stable en-vironment enabling the proliferation and permanence of fungaldiversity.
There are no studies that specify the effect of biotic left-overs on microbial diversity or how this factor can contribute

10 FEMS Microbiology Ecology, 2015, Vol. 91, No. 5
Figure 6. NMDS ordination of the similarities in fungal composition across samples. Fungal composition differed significantly between sample types (r2 = 0.3403,P = 0.009 901). Samples corresponding to the same type are shown with 95% confidence interval ellipses. TS topsoil summer, BS burrow summer, TW topsoil winter
and BW burrow winter.
to the microbial composition in burrowmicrohabitats. However,in desert ecosystems, plant litter can be considered as a con-tributing factor for fungal species occurrence and abundance.Through the activities of small mammals, plant litter can ac-cumulate, therefore promoting fungal colonization (Zak 2005).A study on fungal diversity associated to litter buried by smallmammals like kangaroo rats in arid regions has suggested thatthere is a symbiotic or mutualistic relationship between kanga-roo rats and fungi that colonize the litter (Hawkins 1999); fungibenefit from the environment that the rodents provide, with abi-otic conditions different than those in the soil surface. Althoughthe effect of fungi on rodents is not well understood, genera offungi that grow in buried litter, such as Penicillium, may producemycotoxins, as well as antibiotics with potential protection forthe rodents.
Differences in soil fungal communities were found betweenseasons. Seasonal changes of soil fungal communities are im-portant to understand the role of fungi in ecosystem function-ing as well as their responses to global climate changes and howthis may affect their role in nutrient cycling and decompositionof organic matter. The dendrograms coupled to the heatmapclearly differentiated samples from winter and summer. Anunidentified Ascomycota order and Pleosporales drive these dif-ferences. This is consistent with seasonal changes in the fungalcommunity of a temperate forest, in which the fungal commu-
nity structure was dynamic, with significant differences in theirrelative abundance depending on the sampling time (Vorıskovaet al. 2014) but not on a geographic point as suggested by theMantel test. In a temperate grassland, temporal changes inthe community structure of arbuscular mycorrhizal fungi werefound to correlate with environmental variables, especially tem-perature (Dumbrell et al. 2011).
No Coccidioides spp. were identified by pyrosequencing in anyof the samples. Probably rare taxa may go undetected in thiskind of analysis. In previous studies in VDP, the detection of thispathogen directly from soil samples required a nested PCR of theDikarya ITS region followed by a third PCR with specific primersthat allowed the amplification of Coccidioides spp. ITS2 region(Baptista-Rosas et al. 2012; Catalan-Dibene et al. 2014). In thisstudy, in order to avoid non-specific PCR products, we performedonly a nested PCR that included a first PCR for the amplificationof the Dikarya ITS region, followed by a second PCR with specificnewly designed primers that allowed the amplification of Coc-cidioides spp. ITS1 region. So far, amplification of Coccidioides spp.sequences directly from soil samples by using universal fun-gal primers in a direct PCR reaction has not been possible. Thissuggests that there are very low amounts of Coccidioides’ DNAin the samples or/and that very little Coccidioides’ DNA can berecovered from the arthrospores present in the soil samples.The results obtained for the amplification of ITS regions of

Vargas-Gastelum et al. 11
Coccidioides spp., suggest that the ITS1 region is a more specificbarcode to identify Coccidioides spp. than the ITS2, shown to bemore highly conserved among different fungal species (Catalan-Dibene et al. 2014). In addition, the ITS1 region allowed us to dif-ferentiate between C. immitis and C. posadasii, the two sympatricspecies presumably inhabiting VDP.
The higher rate of detection of Coccidioides spp. in burrowsamples suggests that the presence/absence of this fungus inthe soil in VDP could be associated to thatmicrohabitat (Table S4,Supporting Information). The fungal community structure wasalso found to bemore seasonally stable in burrow than in topsoilsamples. Arguably, this stability in the fungal community couldfavor the maintenance of Coccidioides spp. in this microhabitat.However, more studies are needed to understand the ecologyof Coccidioides. A previous study in VDP (Catalan-Dibene et al.2014) identified species of rodents that build burrows, includingkangaroo rats (Dipodomys merriami). Studies in an arid grasslandon Socorro County, New Mexico, showed that the burrows con-structed by kangaroo rats (D. spectabilis) are a favorablemoist andtemperatemicrohabitat for fungi (Hawkins 1996). Kangaroo rats,as well as other rodents and animals, such as snakes, skunks,among others, are able to inhabit the burrows, where there isproduction of urine, feces and food waste that alter the compo-sition of the organic matter. In this study, no differences in or-ganic matter quantity were found between burrows and topsoil,but there could be qualitative differences in the organic mat-ter available, whichwere notmeasured. These qualitative differ-ences of the organic material inside the burrows could favor thegrowth and development of some fungal taxa and contribute tothe fungal community stability inside burrows through seasons.
SUPPLEMENTARY DATA
Supplementary data is available at FEMSEC online.
ACKNOWLEDGEMENTS
We thank the Consejo Nacional de Ciencia y Tecnologıa (CONA-CyT) of Mexico for providing scholarship no. 263435 to LVG. Weare grateful to J. Stajich from the University of California, River-side for enriching this work with helpful discussions. We thankS. Bullock, R. Vargas and S. Ibarra from CICESE, for providing in-strumentation for soil analyses and G. Gonzalez for technicalfield assistance.
FUNDING
This research was supported by the Fund for Scientific Researchand Technological Development of CICESE to AR-O, MR and CB.
Conflict of interest. None declared.
REFERENCES
Abarenkov K, Nilsson RH, Larsson KH, et al. The UNITE databasefor molecular identification of fungi—recent updates and fu-ture perspectives. New Phytol 2010;186:281–5.
Altschul SF, Gish W, Miller W, et al. Basic local alignment searchtool. J Mol Biol 1990;215:403–10.
Arriaga-Ramırez S, Cavazos T. Regional trends of daily precip-itation indices in northwest Mexico and southwest UnitedStates. J Geophys Res-Atmos 2010;115:1–10.
Baptista-Rosas RC, Catalan-Dibene J, Romero-Olivares AL, et al.Molecular detection of Coccidioides spp. from environmental
samples in Baja California: linking Valley Fever to soil andclimate conditions. Fungal Ecol 2012;5:177–90.
Bates ST, Nash TH, III, Garcia-Pichel F. Patterns of diversity forfungal assemblages of biological soil crusts from the south-western United States. Mycologia 2012;104:353–61.
Bell C, Acosta-Martinez V, McIntyre N, et al. Linking microbialcommunity structure and function to seasonal differences insoilmoisture and temperature in a Chihuahuan desert grass-land. Microb Ecol 2009;58:827–42.
Bell C, McIntyre N, Cox S. Soil microbial responses to tempo-ral variations of moisture and temperature in a Chihuahuandesert grassland. Microb Ecol 2008;56:153–67.
Belnap J, Welter JR, GrimmNB, et al. Linkages between microbialand hydrologic processes in arid and semiarid watersheds.Ecology 2005;86:298–307.
Binnicker MJ, Buckwalter SP, Eisberner JJ, et al. Detection of Coc-cidioides species in clinical specimens by real-time PCR. J ClinMicrobiol 2007;45:173–8.
Bouyoucos GJ. Directions for making mechanical analyses ofsoils by the hydrometer method. Soil Sci 1936;42:225–30
Buee M, Courty P, Mignot D, et al. Soil niche effect on species di-versity and catabolic activities in an ectomycorrhizal fungalcommunity. Soil Biol Biochem2007;39:1947–55.
Buee M, Reich M, Murat C, et al. 454 Pyrosequencing analyses offorest soils reveal an unexpectedly high fungal diversity.NewPhytol 2009;184:449–56.
Burke DJ, Lopez-Gutierrez JC, Smemo KA, et al. Vegetation andsoil environment influence the spatial distribution of root-associated fungi in amature beech-maple forest.Appl EnvironMicrob 2009;75:7639–48.
Cairns L, BlytheD, KaoA, et al.Outbreak of coccidioidomycosis inWashington state residents returning fromMexico. Clin InfectDis 2000;30:61–4.
Cannon PF. Options and constraints in rapid diversity analysisof fungi in natural ecosystems. Fungal Divers 1999;2:1–15.
Caporaso JG, Kuczynski J, Stombaugh J, et al. QIIME allows anal-ysis of high-throughput community sequencing data. NatMethods 2010;7:335–6.
Catalan-Dibene J, Johnson SM, Eaton R, et al. Detection of coc-cidioidal antibodies in serum of a small rodent communityin Baja California, Mexico. Fungal Biol 2014;118:330–9.
Challenger A, Soberon J. Los ecosistemas terrestres. In: CapitalNatural de Mexico, Vol. 1. Mexico: Comision Nacional para elConocimiento y Uso de la Biodiversidad, 2008, 87–108.
Collins SL, Sinsabaugh RL, CrenshawC, et al. Pulse dynamics andmicrobial processes in aridland ecosystems. Journal of Ecol2008;96:413–20.
Colwell RK. Estimates: statistical estimation of species rich-ness and shared species from samples, 2013. Version 8.2.http://purl.oclc.org/estimates (13 April 2015, date last ac-cessed).
De Gannes V, Eudoxie G, Hickey WJ. Insights into fungal com-munities in composts revealed by 454-pyrosequencing: im-plications for humanhealth and safety. FrontMicrobiol 2013;4:1–9.
Dix NJ, Webster J. Fungal Ecology. Chapman & Hall, London, UK,1995, 39–84.
Dray S, Dufour A-B. The ade4 Package: implementing the dualitydiagram for ecologists. J Stat Softw 2007;22:1–20.
Dumbrell AJ, Ashton PD, Aziz N, et al. Distinct seasonal assem-blages of arbuscularmycorrhizal fungi revealed bymassivelyparallel pyrosequencing. New Phytol 2011;190:794–804.
Easterling DR, Meehl G, Parmesan C, et al. Climate extremes: ob-servations, modeling, and impacts. Science 2000;289:2068–74.

12 FEMS Microbiology Ecology, 2015, Vol. 91, No. 5
Edgar RC. Search and clustering orders of magnitude faster thanBLAST. Bioinformatics 2010;26:2460–1.
Edgar RC, Haas BJ, Clemente JC, et al. UCHIME improvessensitivity and speed of chimera detection. Bioinformatics2011;27:2194–200.
Entsminger GL. ECOSIM: null modeling software for ecologist,2012. Version 7. http://homepages.together.net/∼gentsmin/ecosim.htm (13 April 2015, date last accessed).
Fierer N, Breitbart M, Nulton J, et al. Metagenomic and small-subunit rRNA analyses reveal the genetic diversity of bac-teria, archaea, fungi, and viruses in soil. Appl Environ Microb2007;73:7059–66.
Fierer N, Jackson RB. The diversity and biogeography of soil bac-terial communities. P Natl Acad Sci USA 2006;103:626–31.
Foley JA, Defries R, Asner GP, et al. Global consequences of landuse. Science 2005;309:570–4.
Gardes M, Bruns TD. ITS primers with enhanced specificity forbasidiomycetes-application to the identification of mycor-rhizae and rusts. Mol Ecol 1993;2:113–8.
Giri B, Giang PH, Kumari R Microbial diversity in soils. In: BuscotF, Varma A, et al. (eds). Microorganisms in Soils: Roles in Genesisand Functions, Vol. 3. Berlin: Springer, 2005, 19–55.
Hawkins LK. Burrows of kangaroo rats are hotspots for desertsoil fungi. J Arid Environ 1996;32:239–49.
Hawkins LK. Microfungi associated with the banner-tailedkangaroo-rat, Dypodomys spectabilis.Mycologia 1999;91:735–41.
Hibbett DS, Ohman A, Kirk PM. Fungal ecology catches fire. NewPhytol 2009;184:279–82.
Huxman TE, Snyder KA, Tissue D, et al. Precipitation pulsesand carbon fluxes in semiarid and arid ecosystems. Oecolo-gia 2004;141:254–68.
IPCC. Climate Change and Water. Technical Paper of the Intergovern-mental Panel on Climate Change, Geneva: IPCC Secretariat, 2008.
Jones MDM, Richards TA. Environmental DNA analysis andthe expansion of the fungal tree of life. In: Poggeler S,Wostemeyer J (eds). Evolution of Fungi and Fungal-Like Organ-isms, Vol. 14. Heidelberg: Springer, 2011, 37–54.
Jumpponen A, Jones KL. Massively parallel 454 sequencingindicates hyperdiverse fungal communities in temperateQuercus macrocarpa phyllosphere. New Phytol 2009;184:438–48.
Jumpponen A, Jones KL, Blair J. Vertical distribution offungal communities in tallgrass prairie soil. Mycologia2010;102:1027–41.
Lauenroth WK, Bradford JB. Ecohydrology of dry regions of theUnited States: water balance consequences of small precipi-tation events. Ecohydrology 2012;5:46–53.
Lentendu G, Zinger L, Manel S, et al. Assessment of soil fungaldiversity in different alpine tundra habitats by means of py-rosequencing. Fungal Divers 2011;49:113–23.
Magan N. Fungi in extreme environments. In: Kubicek CPD, IrinaS (eds). Environmental andMicrobial Relationships, Vol. 4. Berlin,Heidelberg: Springer, 2007, 85–103.
Magana V, Mendez JM, Morales R Consecuencias presentes y fu-turas de la variabilidad y el cambio climatico en Mexico. In:Martinez J, Bremauntz AF, et al. (eds). Secretarıa del MedioAmbiente y Recursos Naturales: Una Vision Desde Mexico,Instituto Nacional de Ecologıa, 2004;1:203–13.
Martin KJ, Rygiewicz PT. Fungal-specific PCR primers developedfor analysis of the ITS region of environmental DNA extracts.BMC Microbiol 2005;5:28.
Mohamed DJ, Martiny JBH. Patterns of fungal diversity and com-position along a salinity gradient. ISME J 2011;5:379–88.
Monard C, Gantner S, Stenlid J. Utilizing ITS1 and ITS2 to studyenvironmental fungal diversity using pyrosequencing. FEMSMicrobiol Ecol 2013;84:165–75.
Nilsson RH, Veldre V, Hartmann M, et al. An open source soft-ware package for automated extraction of ITS1 and ITS2from fungal ITS sequences for use in high-throughput com-munity assays and molecular ecology. Fungal Ecol 2010;3:284–7.
Noguez AM, Escalante AE, Forney LJ, et al. Soil aggregates ina tropical deciduous forest: effects on C and N dynamics,and microbial communities as determined by t-RFLPs. Bio-geochemistry 2008;89:209–20.
Oksanen J, Blanchet FJ, Kindt R, et al. Vegan: Community Ecol-ogy Package. R package version 2.2-1, 2015. Available online at:http://CRAN.R-project.org/package=vegan.
Oliveira LG, Cavalcanti MAQ, Fernandes MJS, et al. Diversity offilamentous fungi isolated from the soil in the semiarid area,Pernambuco, Brazil. J Arid Environ 2013;95:49–54.
Opik M, Metsis M, Daniell TJ, et al. Large-scale parallel 454 se-quencing reveals host ecological group specificity of arbus-cular mycorrhizal fungi in a boreonemoral forest. New Phytol2009;184:424–37.
Orgiazzi A, Lumini E, Nilsson RH, et al. Unravelling soil fungalcommunities from different Mediterranean land-use back-grounds. PloS one 2012;7:1–9.
Pavia EG, Graef F, Reyes J. Annual and seasonal surface air tem-perature trends in Mexico. Int J Climatol 2009;29:1324–9.
Porras-Alfaro A, Herrera J, Natvig DO, et al. Diversity and distri-bution of soil fungal communities in a semiarid grassland.Mycologia 2011;103:10–21.
Pumpanen J, Ilvesniemi H, Hari P. A process-based model forpredicting soil carbon dioxide efflux and concentration. SoilSci Soc Am J 2003;67:402–13.
Quince C, Lanzen A, Davenport RJ, et al. Removing noise frompyrosequenced amplicons. BMC Bioinformatics 2011;12:38.
Ravi S, D’Odorico P, Over TM, et al. On the effect of air humidityon soil susceptibility towind erosion: the case of air-dry soils.Geophys Res Lett 2004;31:1–4.
Reynolds JF, Smith DM, Lambin EF, et al. Global desertifica-tion: building a science for dryland development. Science2007;316:847–51.
Ripley B, Venables B, Hornik K, et al. MASS: support func-tions and datasets for Venables and Ripley’s MASS. 2013,http://cran.r-project.org/web/packages/MASS/ (13 April 2015,date last accessed).
Romero-Olivares AL, Baptista-Rosas RC, Escalante AE, et al. Dis-tribution patterns of Dikarya in arid and semiarid soils ofBaja California, Mexico. Fungal Ecol 2013;6:92–101.
Siminski P. The desert adaptations of birds & mammals (2014)Arizona-Sonora desert museum, Tucson, Arizona. 2014,https://www.desertmuseum.org/books/nhsd adaptationsbirds.php (13 April 2015, date last accesed).
Skouw M. Soil fungal community responses to global changes.Ph.D.Thesis. University of Copenhagen 2013.
Tedersoo L, Bahram M, Polme S, et al. Global diversity and geog-raphy of soil fungi. Science 2014;346:1256688.
Vorıskova J, Brabcova V, Cajthaml T, et al. Seasonal dynamics offungal communities in a temperate oak forest soil. New Phy-tol 2014;201:269–78.
Wallander H, Johansson U, Sterkenburg E, et al. Production ofectomycorrhizal mycelium peaks during canopy closure inNorway spruce forests. New Phytol 2010;187:1124–34.
White TJ, Bruns T, Lee S Amplification and direct sequencingof fungal ribosomal RNA genes for phylogenetics. In: Innin

Vargas-Gastelum et al. 13
N, Gelfand D, Sninski J, et al. (eds). PCR Protocols: A Guide toMethods and Applications, Vol. 18. New York: Academic Press,1990, 315–22.
Whitford WG. The importance of the biodiversity of soil biota inarid ecosystems. Biodivers Conserv 1996;5:185–95.
Zak J. Fungal communities of desert ecosystems: links toclimate change. In: Dighton J, White JF, Oudemans P(eds). The Fungal Community: Its Organization and Role inthe Ecosystem, Vol. 23. Mycology: CRC Press, 2005, 659–81.