histological analysis of the egg capsule of the ovoviviparous polystomatid monogenean,...
TRANSCRIPT

Pergamon InrernotionalJournalfor Parasitology, Vol. 27, No. 9, pp 1075-1080, 1997
!Q 1997 Australian Society for Parasitology. Pubbshed by Elsevier Science Ltd Printed in Great Britain
PII: SOO20-7519(97)00068-4 002tL7519197 617.00+0.00
Histological Analysis of the Egg Capsule of the Ovoviviparous Polystomatid Monogenean,
Pseudodiplorchis americanus
J. CABLE,*8 K. TOCQUET and R. C. TINSLEY*
‘School of Biological Sciences, University of Bristol, Bristol BS8 I UG, U.K. j-Department of Epidemiology, Public Health Laboratory Service, Fazakerle-y Hospital,
Lower Lane, Liverpool L9 7AL, U.K.
(Received 29 January 1997; accepted 5 May 1997)
Abstract-Cable J., Tocque K. & Tinsley R. C. 1997. Histological analysis of the egg capsule of the ovoviviparous polystomatid monogenean, Pseudodiplorchis americanus. International Journalfor Parasitology 27: 1075-1080. Transmission of Pseudodiplorchis americanus is restricted to the brief period when its host, a desert toad, enters water to spawn. The parasite accumulates its entire annual reproductive output within an elongated uterus during the lO-11-month period of host hibernation. Embryos of P. americanus, at all stages of development, are retained within the uterus which eventually becomes packed with around 150 encapsulated infective larvae. Recently formed eggs, which comprise a fertilized ovum and 2-3 vitelline cells, are closely surrounded by a primary eggshell which stains positively for acidic proteins and keratin. Initially, during passage along the proximal uterus, the egg capsule is only 6Ogm in diameter, but as it passes to the distal uterus it expands to 800~ in diameter to accommodate the growing larva. Due to chemical alterations or complete replacement of the shell, the final (secondary) egg capsule is a large sac-like structure composed of elastin. The flexible nature of this shell maximizes the numbers of infective larvae which can be stored in utero. TEM studies have revealed this capsule to be composed of multi-laminate membranes with a specialized cytoplasmic lining involved in a unique mechanism for embryo nutrition. This is the first report of an elastin- type eggshell within the Monogenea. (Q 1997 Australian Society for Parasitology. Published by Elsevier Science Ltd.
Key words: Monogenea; Polystomatidae; eggshell; elastin; keratin; histochemistry.
INTRODUCTION The exact form of the “phenol” within the vitellaria
Within the platyhelminths, 3 main types of eggshell is unknown; presumably it is not free phenol as this
have been identified, composed of sclerotin. keratin would be highly toxic to the cells. Tanning (oxidation
and elastin. The majority of eggshells in turbellarians, of phenol via phenolase to form a quinone-bonded
monogeneans, digeneans and pseudophyllidean ces- protein) occurs gradually after egg formation, but is
todes are formed from the tanned protein, sclerotin generally completed before oviposition (Smyth &
(Huggins & Waite, 1993; Kearn, 1986; Smyth & Clegg, 1959). Helminths with keratin eggshells contain
Halton. 1983; Wharton, 1983). Precursors of sclerotin the precursors of quinone tanning in the vitellaria but
eggshells (basic protein, “phenol” and phenolase) are the eggshell protein is stabilized by -SS- linkages
stored within vitelline droplets. Vitelline cells are trig- and dityrosine. These keratin-based eggs are colour-
gered via intracellular calcium to release the shell pre- less and transparent, but they retain the rigidity of
cursors within the ootype (Wells & Cordingley, 1991). tanned sclerotin shells (Arfin & Nizami, 1986; Kal- antan & Arfin. 1984; Kalantan et al.. 1994; Rama- lingham, 1973). To date, elastin eggshells, which are
$To whom correspondence should be addressed. Tel: 0117 thin and flexible, have only been identified in a ces- 9287473; Fax: 0117 9257374; E-mail: [email protected]. todarian and a digenean (Coil, 1987; Madhavi & Rao,
1075

1076 J. Cable et al.
1971). Extremely thin and elastic egg capsules have been described in a few other digeneans and cestodes, but the chemical composition of these shells has not been identified (Allison & Coakley, 1973; Madhavi & Rao, 1971; McMichael-Phillips et al., 1992; Nollen, 1971; Taft, 1986).
Histochemical analyses of rigid monogenean egg- shells have identified sclerotin and keratin-type capsules. The precursors for sclerotin have been dem- onstrated in several species: Gastrocotyle trachuri,
Entobdella soleae, Diclidophora luscae, D. merlangi,
Octornacrum lanceatum, Obegyrodactylus farlowellae,
Protomicrocotyle sp.. Rajonchocotyle batis, Calicotyle
sp. and in the polystomatids Protopolvstoma xeno-
podis, Polystoma integerrimum and Polystomoides sp. (reviewed by Smyth & Halton, 1983). Keratin-type eggshells occur in Heteromicrocotyle carangis, H.
indicus, Pricea multae and Pseudomicrocotyle sp. (Kalantan & Arfin, 1984; Kalantan et al., 1994; Ramalingham, 1973).
Previous studies on the polystomatids Pseudo-
diplorchis americanus and Neodiplorchis scaphiopodis
have shown that their egg capsules are composed of flexible rather than rigid material. However, the chemical composition of these eggshells is unknown. In both parasites, large numbers of encapsulated eggs may be stored in an elongate uterus and the egg cap- sules increase enormously in size during development (Tinsley, 1983). Encapsulated embryos at all stages of development are maintained in the uterus of gravid adults. The uterus is an elongated loop; the proximal descending limb extends into the haptor from where the ascending limb meanders to the anterior genital atrium. Previous TEM studies have shown that the structure of the egg capsule alters during passage along the uterus (Cable & Tinsley, 1991). Vitelline glands in both P. americanus (see Lamothe-Argu- medo, 1986) and N. scaphiopodis (see Rodgers, 1941) are rudimentary. In these monogeneans, the usual dual role of the vitellaria in providing eggshell materials and nutrients for embryo development has been largely taken over by the uterus (Cable & Tinsley, 1991). The aim of this study was to follow the devel- opment of the egg during passage along the uterus and to determine, histochemically, the nature of the egg capsule in P. americanus.
MATERIALS AND METHODS
Adult Pseudodiplorchis americamu were recovered from the urinary bladder of Scaphiopus couchii collected in south- eastern Arizona, U.S.A. The specimens were processed for histochemistry as described in the references listed in Table 1, Control tests for quinone tanning precursors were carried out on adult Protopolystoma senopodis recovered from the bladder of Xenopus lawis, imported from South Africa.
Human skin and hair were used as controls for elastin and keratin, respectively. An additional control for keratin was provided by the marginal haptoral hooks (see Lyons, 1966) of P. americanus adults and encapsulated larvae.
RESULTS
Vitelline system
The vitelline system of Pseudodiplorchis americanus
is greatly restricted in distribution: there are 2 small symmetrically arranged vitelline glands, level with the ovary, which consist of about 15 cells each. In Pro-
topolystoma xenopodis, hundreds of vitelline follicles extend throughout the body proper and this pattern is more typical of the Monogenea. Histochemistry gave contrasting results in these 2 species. Tests for basic protein, phenol and phenolase gave intense reac- tions in the vitelline follicles of P. xenopodis, but did not stain the equivalent cells in P. americanus (Table 1). Only the orcein-aniline blue-orange G test gave a slight reaction for keratin in the vitelline droplets of P. americanus, and an intense reaction in the vitellaria of P. xenopodis.
Mehlis ’ gland
The secretory cells of the Mehlis’ gland of P. amer-
icanus stained positively with light green, indicating acidic material (Gurr, 1963). Despite an examination of several hundred specimens, no parasites were fixed which had developing eggs in the ootype. However, clusters of 4 or 5 vitelline cells were observed occasion- ally in a small triangular reservoir at the junction of the vagino-vitelline canals.
Eggshell formation
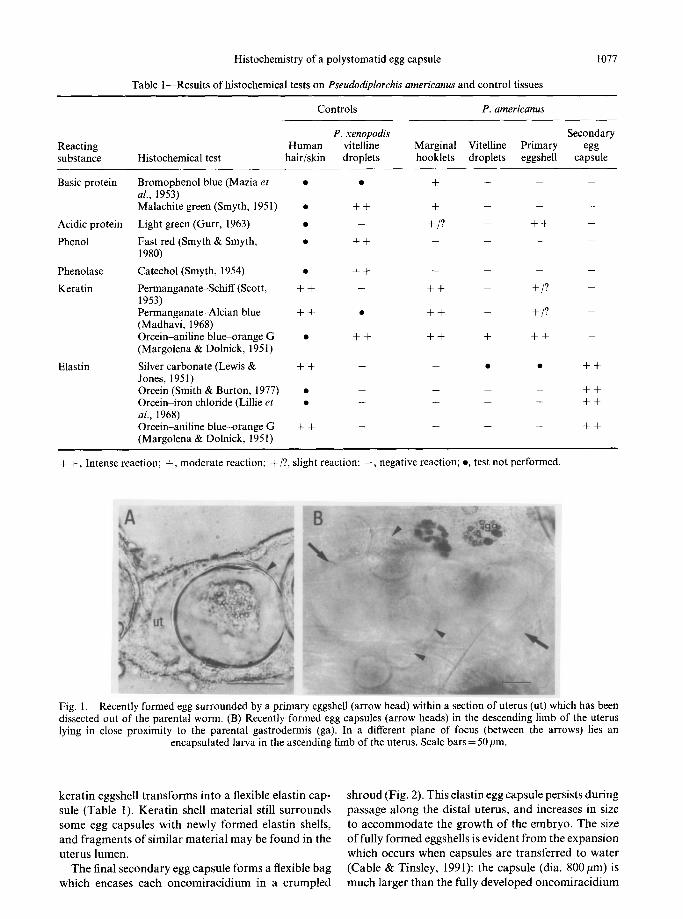
Recently formed eggs comprise a fertilized ovum surrounded by granular material and 2 (occasionally 3) vitelline cells. These vitelline cells are either intact or have no distinguishable cell membrane. The first 5 or 6 embryos in the proximal uterus are spherical and surrounded by a tightly fitting translucent egg capsule. This relatively thin shell gives positive reactions for acidic protein and keratin (Table 1; Fig. 1A). As the embryos pass along the uterus, the shell appears to thicken and the reactions for protein and keratin intensify. The shape of the eggshell also becomes less uniform. This shell persists as the embryos pass along the descending limb of the uterus into the middle third of the body (Fig. 1B).
At later stages of development. as egg capsules pass into the posterior body of the adult worms, there is a transition in the chemical composition of the eggshells. The primary tightly fitting acidic protein/
Histochemistry of a polystomatid egg capsule
Table l-Results of histochemical tests on Pseudodiplorchis americanus and control tissues
1077
Reacting substance Histochemical test
Controls P. americanus
P. xenopodis Secondary Human vitelline Marginal Vitelline Primary egg
hair/skin droplets hooklets droplets eggshell capsule
Basic protein
Acidic protein
Phenol
Phenolase Keratin
Elastin
Bromophenol blue (Mazia et al., 1953) Malachite green @myth, 1951)
Light green (Gurr, 1963)
Fast red (Smyth & Smyth, 1980)
Catechol (Smyth, 1954)
Permanganate-Schiff (Scott, 1953) Perrnanganate-Alcian blue (Madhavi, 1968) Orcein-aniline blue-orange G (Margolena & Dolnick, 1951)
Silver carbonate (Lewis & Jones, 1951) Orcein (Smith & Burton, 1977) Orcein-iron chloride (Lillie et al., 1968) Orcein-aniline blue-orange G (Margolena & Dolnick, 1951)
.
.
.
.
.
++
++
.
++
.
.
++
. +
++ + - +I?
++ -
++ - - ++
. ++
++ ++
- - - -
- - - - ++ - - - -
- - -
- +I? -
- +I? -
+ ++ -
. . ++
- - ++ - - ++
- - ++
+ + , Intense reaction; + , moderate reaction; + /?, slight reaction; - , negative reaction; ., test not performed.
Fig. 1 dissec lying
:tel in
Recently formed egg surrounded by a primary eggshell (arrow head) within a section of uterus (ut) which has been d out of the parental worm. (B) Recently formed egg capsules (arrow heads) in the descending limb of the uterus
close proximity to the parental gastrodermis (ga). In a different plane of focus (between the arrows) lies an encapsulated larva in the ascending limb of the uterus. Scale bars = 50 pm.
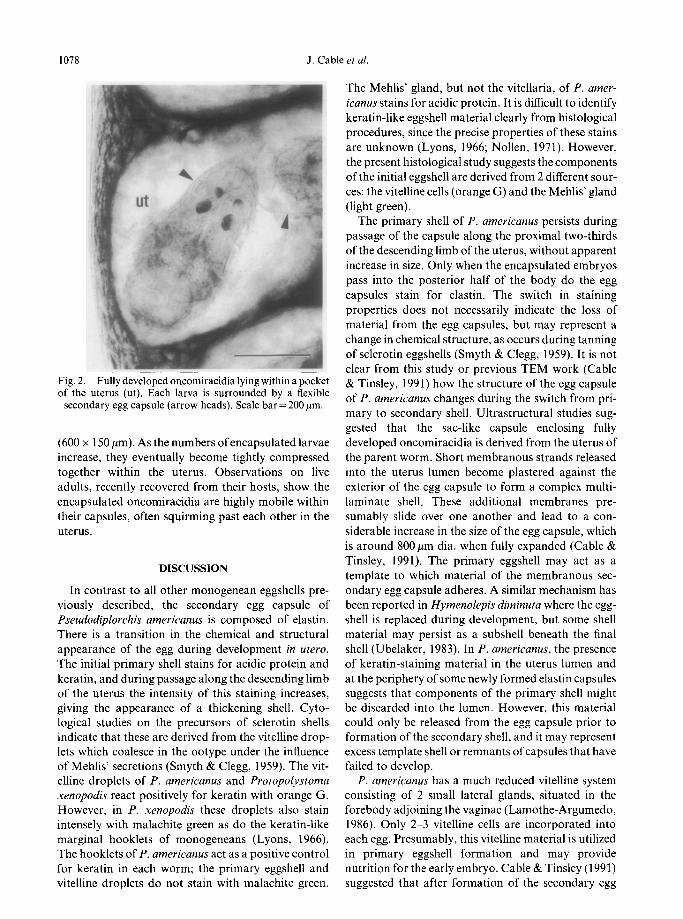
keratin eggshell transforms into a flexible elastin cap- shroud (Fig. 2). This elastin egg capsule persists during sule (Table 1). Keratin shell material still surrounds passage along the distal uterus, and increases in size some egg capsules with newly formed elastin shells, to accommodate the growth of the embryo. The size and fragments of similar material may be found in the of fully formed eggshells is evident from the expansion uterus lumen. which occurs when capsules are transferred to water
The final secondary egg capsule forms a flexible bag (Cable & Tinsley, 199 1): the capsule (dia. 800 pm) is which encases each oncomiracidium in a crumpled much larger than the fully developed oncomiracidium
J. Cable et al.
Fig. 2. Fully developed oncomiracidia lying within a pocket of the uterus (ut). Each larva is surrounded by a flexible
secondary egg capsule (arrow heads). Scale bar = 200 pm.
(600 x 150 pm). As the numbers of encapsulated larvae increase, they eventually become tightly compressed together within the uterus. Observations on live adults, recently recovered from their hosts, show the encapsulated oncomiracidia are highly mobile within their capsules, often squirming past each other in the uterus.
DISCUSSION
In contrast to all other monogenean eggshells pre- viously described, the secondary egg capsule of Pseudodiplorchis americanus is composed of elastin. There is a transition in the chemical and structural appearance of the egg during development in utero.
The initial primary shell stains for acidic protein and keratin, and during passage along the descending limb of the uterus the intensity of this staining increases, giving the appearance of a thickening shell. Cyto- logical studies on the precursors of sclerotin shells indicate that these are derived from the vitelline drop- lets which coalesce in the ootype under the influence of Mehlis’ secretions (Smyth & Clegg, 1959). The vit- elline droplets of P. americanus and Protopolystoma
senopodis react positively for keratin with orange G. However, in P. xenopodis these droplets also stain intensely with malachite green as do the keratin-like marginal hooklets of monogeneans (Lyons, 1966). The hooklets of P. americanus act as a positive control for keratin in each worm; the primary eggshell and vitelline droplets do not stain with malachite green.
The Mehlis’ gland, but not the vitellaria, of P. amer-
icanus stains for acidic protein. It is difficult to identify keratin-like eggshell material clearly from histological procedures, since the precise properties of these stains are unknown (Lyons, 1966; Nollen, 1971). However, the present histological study suggests the components of the initial eggshell are derived from 2 different sour- ces: the vitelline cells (orange G) and the Mehlis’ gland (light green).
The primary shell of P. americanus persists during passage of the capsule along the proximal two-thirds of the descending limb of the uterus, without apparent increase in size. Only when the encapsulated embryos pass into the posterior half of the body do the egg capsules stain for elastin. The switch in staining properties does not necessarily indicate the loss of material from the egg capsules, but may represent a change in chemical structure, as occurs during tanning of sclerotin eggshells (Smyth & Clegg, 1959). It is not clear from this study or previous TEM work (Cable & Tinsley. 1991) how the structure of the egg capsule of P. americanus changes during the switch from pri- mary to secondary shell. Ultrastructural studies sug- gested that the sac-like capsule enclosing fully developed oncomiracidia is derived from the uterus of the parent worm. Short membranous strands released into the uterus lumen become plastered against the exterior of the egg capsule to form a complex multi- laminate shell. These additional membranes pre- sumably slide over one another and lead to a con- siderable increase in the size of the egg capsule, which is around 800 pm dia. when fully expanded (Cable & Tinsley, 1991). The primary eggshell may act as a template to which material of the membranous sec- ondary egg capsule adheres. A similar mechanism has been reported in Hymenolepis diminuta where the egg- shell is replaced during development, but some shell material may persist as a subshell beneath the final shell (Ubelaker, 1983). In P. americanus, the presence of keratin-staining material in the uterus lumen and at the periphery of some newly formed elastin capsules suggests that components of the primary shell might be discarded into the lumen. However, this material could only be released from the egg capsule prior to formation of the secondary shell, and it may represent excess template shell or remnants of capsules that have failed to develop.
P. americanus has a much reduced vitelline system consisting of 2 small lateral glands, situated in the forebody adjoining the vaginae (Lamothe-Argumedo, 1986). Only 2-3 vitelline cells are incorporated into each egg. Presumably, this vitelline material is utilized in primary eggshell formation and may provide nutrition for the early embryo. Cable & Tinsley (1991) suggested that after formation of the secondary egg

Histochemistry of a polystomatid egg capsule 1079
capsule, nutrients pass directly from the parent to the developing embryo through fine cytoplasmic exten- sions of the egg capsule lining. The ontogeny of P.
anwricanus egg capsules and mechanism of embryo nutrition is similar to that which occurs in Mesostoma
ehrenbergii. In this turbellarian, subitaneous egg cap- sules are produced when the parental vitelline system is small and undeveloped (Gremigni & Domenici, 1976). The eggs receive nutrients directly across the uterus wall. In contrast, dormant eggs of M. ehren-
bergii which develop in the external environment are
provided with large reserves of vitelline material.
The resistant properties of rigid eggshells, whether formed from sclerotin or keratin, provide protection from damage in the host gut or the external environ- ment (Wharton, 1983). The development of thin, non- tanned egg capsules in Gorgoderina attenuata, Mega-
lodiscus temperatus and Philophthalmus megalurus was linked by Nollen (1971) to lack of exposure to the host digestive tract. Similarily, the elastin egg capsules of Orchispirium heterozGtellatum are protected from the hazards of the external environment as they develop in the gills of the fish (Madhavi & Rao, 1971). Flexible egg capsules have also been associated with the con- servation of space and intra-uterine embryonic devel- opment (Tinsley. 1983). The thin eggshell of Sanguinicolu inermis affords little or no protection, but its structure may provide a route for transfer of nutrients from host to developing miracidium (McMichael-Phillips et al., 1992). Like P. americanus,
the eggshell of Sunguinicola idahoensis expands as the embryo absorbs nutrients (Schell. 1974). A variety of ecological situations appear to have favoured the evolution of flexible egg capsules (Allison & Coakley, 1973: Fairweather & Threadgold. 1981; Madhavi & Rao, 1971; Tinsley, 1978a, 1983).
Ovoviviparity is a common reproductive strategy amongst polystomatid monogeneans which have exploited hosts whose ecology and behaviour present a restricted opportunity for transmission (Tinsley, 1983). Diplorchis ranae. Eupolystorna anterorchis,
Neodiplorchis scaphiopodis and P. americanus all pro- duce immediately infective larvae and can store up to 300 egg capsules in utero. However, there is no obvious association between the mode of embryonic devel- opment and eggshell structure. D. ranae produces
rigid operculate eggs which are orange in colour, indicative of a tanned protein: this species has exten- sive vitelline glands which presumably provide a large part of the nutrients for embryonic development (Ozaki, 1935). On the other hand, the larvae of P.
americanus, N. scaphiopodis and Eupol.vstonza ante-
rorchis (see Tinsley. 1978b) are enclosed within thin almost transparent eggshells, which increase in size during passage along the uterus. Preliminary obser-
vations on N. scaphiopodis (which is almost identical in its development and life-cycle to P. americanus)
suggest that its secondary eggshell is also composed of elastin (unpublished data). Both P. americanus and N. scuphiopodis have a reduced vitelline system associ- ated with an almost complete dependence on a uterine provision of nutrients. These unusual features of the reproductive system are adaptations to the unpre- dictable opportunity for transmission. The data of Tocque & Tinsley (1991) suggest that oncomiracidia can be maintained in an infective state for 2-3 months within the parent worm by means of the continuous intra-uterine supply of nutrients. The highly folded, thin and extremely flexible eggshells allow the maximum number of larvae to be packed into the uterus. The larvae hatch as soon as they are expelled from their parent, so there is no requirement for the egg capsules to provide protection.
Acknowledgements-We are grateful for financial support from the Natural Environment Research Council, U.K., and for fieldwork facilities at the Southwestern Research Station of the American Museum of Natural History, Portal, AZ.
REFERENCES
Allison F. R. & Coakley A. 1973. Two species of Gyrocotyle in the elephant fish, Callorhynchus milii (Bory). Journal of the Royal Societv of New Zealand 3: 381-392.
Arfin M. & Nizami W. A. 1986. Chemical nature and mode of stabilization of egg-shell proteins of some digenetic trematodes. Helminthologia 23: 157-164.
Cable J. & Tinsley R. C. 1991. Intrauterine larval devel- opment of the polystomatid monogeneans, Pseudo- diplorchis americanus and Neodiplorchis scaphiopodis. Parasitology 103: 253-266.
Coil W. H. 1987. The early egg of Austramphilina elongata (Cestodaria). Parasitology Research 73: 451457.
Fairweather I. & Threadgold L. T. 1981. Hymenolepis nana: the fine structure of the embryonic envelopes. Parasitolog), 82: 429443.
Gremigni V. & Domenici L. 1976. Uptake of maternal nutri- ents during embryonic development in the subitaneous eggs of the turbellarian Mesostoma ehrenbergii (Focke). Monitore Zoologico Italian0 IO: 2299238.
Gurr G. T. 1963. Biological Staining-Method.s, 7th edition. G. T. Gurr Ltd. London.
Huggins L. G. & Waite J. H. 1993. Eggshell formation in Bdelloura candida, an ectoparasitic turbellarian of the horseshoe crab Limrrluspolvphemtrs. The Journal of Exper- imental Zoology 265: 5499557.
Kalantan A. M. N. & At-fin M. 1984. Stabilization of egg- shell in Heteromicrocotyle indicus Ramalingham, 1960 (Monogenea: Microcotylidae). Proceedings of the Saudi Biological Society 7: 415427.
Kalantan A. M. N., Arfin M., Al-Yousif A. L. & Nizami W. A. 1994. Histochemical nature of egg-shell of some trematodes. Japanese Journal of Parasitology 43: 110-l 17.
Kearn G. C. 1986. The eggs of monogeneans. Adzlances in Parasitology 25: 1755273.
Lamothe-Argumedo R. 1986. Monogeneos de 10s anfibios

1080 J. Cable : et al.
de Mexico VII. Hallazgo de Pseudodiplorchis americanus (Rodgers y Kuntz, 1940) Yamaguti, 1963 en Baja Cal- ifornia SW, Mexico. Annales de Institute de Biologia Univ- ersidad de National Autanoma Mexico Zoologia 56: 291L 300.
Lewis A. L. & Jones R. S. 1951. A combined connective tissue stain for elastin, reticulin and collagen. Stain Tech- nology 26: 85-87.
Lillie R. D., Gutierrez A., Madden D. & Henderson R. 1968. Acid orcein-iron and acid orceincopper stains for elastin. Stain Technology 43: 203-206.
Lyons K. M. 1966. The chemical nature and evolutionary significance of monogenean attachment sclerites. Para- sitology 56: 63-100.
McMichael-Phillips D. F., Lewis J. W. & Thorndyke M. C. 1992. Ultrastructure of the egg of Sanguinicola inermis Plehn, 1905 (Digenea: Sanguinicolidae). JournalofNatural History 26: 895-904.
Madhavi R. 1968. Diplodiscus mehrai: chemical nature of eggshell. Experimental Parasitology 23: 392-397.
Madhavi R. & Rao K. H. 1971. Orchispirium hetero- oitellatum: chemical nature of the eggshell. Experimental Parasitology 30: 345-348.
Margolena L. A. & Dolnick E. H. 1951. A differential stain- ing method for elastic fibers, collagenic fibres and keratin. Stain Technology> 26: 119-121.
Mazia D., Brewer P. A. & Alfert M. 1953. The cytochemical staining and measurement of protein with mercuric bro- mophenol blue. Biological Bulletin 104: 5747.
Nollen P. M. 1971. Digenetic trematodes: quinone tanning system in eggshells. Experimental Parasito1og.v 30: 6472.
Ozaki Y. 1935. Studies on the frog trematode Diplorchis ranae. I. Morphology of the adult worm with a review of the family Polystomatidae. Journal of Science of the Hiroshima Unit~ersity, Series B 3: 193-225.
Ramalingham K. 1973. Chemical nature of the eggshell in helminths: II. Mode of stabilization of eggshells of mono- genetic trematodes. Experimental Parasitology 34: 115- 122.
Rodgers L. 0.1941. Diplorchisscaphiopi, a new polystomatid monogenean fluke from the spadefoot toad. Journal of Parasitology 27: 153-l 57.
Schell S. C. 1974. The life history of Sanguinicola idahoensis sp. n. (Trematoda: Sanguinicolidae), a blood parasite of steelhead trout, Salmo gairdneri Richardson. Journal of Parasitology 60: 561-566.
Scott H. R. 1953. Demonstration of keratin with aldehyde- fuchsin. Nature 172: 674675.
Smith A. & Burton J. 1977. A Colour Atlas of Histological Staining Techniques. Wolfe Medical Atlases 18. Wolfe Medical Publications, London.
Smyth J. D. 1951. Specific staining of egg-shell material in trematodes and cestodes. Stain Technology 26: 255-256.
Smyth J. D. 1954. A technique for the histological dem- onstration ‘of polyphenol oxidase and its application to egg-shell formation in helminths and byssus formation in Mvtilus. Quarterly Journal of Microscopical Science 95: 139-152.
Smyth J. D. & Clegg J. A. 1959. Egg-shell formation in trematodes and cestodes. Experimental Parasitology 8: 286323.
Smyth J. D. & Halton D. W. 1983. The Physiology of Trema- todes. Cambridge University Press, Cambridge.
Smyth J. D. & Smyth M. M. 1980. Frogs as Host-Parasite Systems, Vol. I, An Introduction to Parasitology through the Parasites of Rana temporaria. R. esculenta and R. pipiens. Macmillan Press, London.
Taft S. J. 1986. Aspects of larval development and histo- chemistry of Ophthalmophagus singularis. Journal of Para- sitology 72: 13&141.
Tinsley R. C. 1978a. The morphology and distribution of Eupolystoma species (Monogenoidea) in Africa, with a description of E. anterorchis sp. n. from Bufo pardalis at the Cape. Journal of Helminthology 52: 291-302.
Tinsley R. C. 1978b. Oviposition, hatching and the onco- miracidium of Eupolystoma anterorchis (Monogenoidea). Parasitology 77: 121-132.
Tinsley R. C. 1983. Ovoviviparity in platyhelminth life- cycles. Parasitology 86: 161-196.
Tocque K. & Tinsley R. C. 1991, Asymmetric reproductive output by the monogenean Pseudodiplorchis americanus. Parasitology 102: 213-220.
Ubelaker J. E. 1983. The morphology, development and evolution of tapeworm larvae. In: Bio1og.v of the Euces- toda, Vol. I (Edited by Arme C. &Pappas P. W.). pp. 235- 296. Academic Press, London.
Wells K. E. & Cordingley J. S. 1991. Schistosoma mansoni: Eggshell formation is regulated by pH and calcium. Exper- imental Parasitology 73: 295-3 10.
Wharton D. A. 1983. The production and functional mor- phology of helminth egg-shells. Parasitology 86: 85597.