hippocampal neurogenesis and antidepressive therapy: shocking relations
TRANSCRIPT

Review ArticleHippocampal Neurogenesis and Antidepressive TherapyShocking Relations
Peter Rotheneichner1 Simona Lange2 Anna OrsquoSullivan1 Julia Marschallinger2
Pia Zaunmair1 Christian Geretsegger3 Ludwig Aigner2 and Sebastien Couillard-Despres1
1 Paracelsus Medical University Institute of Experimental Neuroregeneration Spinal Cord Injury andTissue Regeneration Center Salzburg (SCI-TReCS) Strubergasse 22 5020 Salzburg Austria
2 Paracelsus Medical University Institute of Molecular Regenerative Medicine Spinal Cord Injury andTissue Regeneration Center Salzburg (SCI-TReCS) Strubergasse 22 5020 Salzburg Austria
3 University Clinics of Psychiatry and Psychotherapy I Paracelsus Medical UniversityIgnaz-Harrer-Straszlige 79 5020 Salzburg Austria
Correspondence should be addressed to Sebastien Couillard-Despres scouillard-desprespmuacat
Received 31 January 2014 Accepted 25 April 2014 Published 22 May 2014
Academic Editor Paul Lucassen
Copyright copy 2014 Peter Rotheneichner et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Speculations on the involvement of hippocampal neurogenesis a form of neuronal plasticity in the aetiology of depression and themode of action of antidepressive therapies started to arisemore than a decade ago But still conclusive evidence that adult neuroge-nesis contributes to antidepressive effects of pharmacological and physical therapies has not been generated yetThis review revisitsrecent findings on the close relation between the mode(s) of action of electroconvulsive therapy (ECT) a powerful interventionused as second-line treatment of major depression disorders and the neurogenic response to ECT Following application of electro-convulsive shocks intricate interactions between neurogenesis angiogenesis andmicroglia activation the hypothalamic-pituitary-adrenal axis and the secretion of neurotrophic factors have been documented Furthermore considering the fact that neurogenesisstrongly diminishes along agingwe investigated the response to electroconvulsive shocks in young aswell as in aged cohorts ofmice
1 Electroconvulsive Therapy and MajorDepression Disorders
Major depression is one of themost common forms ofmentaldisorders in humans Hereditary and environment-triggeredforms of this disease lead to a severe decline of life qualityand might elicit life-threating events like suicide attemptsDespite the constant development and improvement ofantidepressive drugs a compound providing fast and long-lasting pharmacological efficacy in most patients is still lack-ing Although the binding specificity for recently developeddrug-classes for example selective serotonin noradrenalinreuptake inhibitors (SSNRIs) has been improved furtherconcerns regarding their side effects such as the increasedrisk of suicide must be heeded
Electroconvulsive therapy (ECT) as an alternative treat-ment offers a safe rapid and potent therapy to severely
depressed patients ECT-associated side effects like cognitiveimpairments are transient and may slightly vary from oneECT-protocol to the other [1 2] ECT was firstly performedby Ugo Cerletti and Lucio Bini in 1938 [3] and inducesrapid improvements on mental disorders In contrast stan-dard pharmacological antidepressive treatments such asthe administration of selective serotonin reuptake inhibitors(SSRIs) commonly show their first efficacy weeks after initi-ation and are dependent on individual features such as sextreatment or disease pattern [4 5] Since the introductionof ECT in the clinic several parameters such as electrodeplacement pulse width and pulse shape were improved inorder to optimize efficacy and minimize side effects [6ndash8] Nowadays ECT has several indications for instancepsychotic and melancholic depressions malignant catatoniaand delirious mania [9] Nevertheless ECT is predominantly
Hindawi Publishing CorporationNeural PlasticityVolume 2014 Article ID 723915 14 pageshttpdxdoiorg1011552014723915
2 Neural Plasticity
used in severe cases ofmajor depression as a second line treat-ment following failure of pharmacological approaches [10]Overall according to meta-analyses of clinical studies ECTwas reported to be significantly more effective in depressivedisorders compared to pharmacological treatments [2 11]
2 Are Neurogenesis and AntidepressiveActivity Linked
Adult neurogenesis the generation of new neurons takesplace in numerous mammalian species including humans[12 13] Neurogenesis occurs in at least two specific areas ofthe adult brain namely the dentate gyrus of the hippocampusand the subventricular zone (SVZ) of the lateral ventricles Inthese so-called neurogenic niches neural stem cells (NSCs)can on the one hand divide symmetrically leading to twodaughter stem cells and on the other hand perform anasymmetric division resulting in a daughter stem cell and arapid amplifying progenitor cell Depending on the microen-vironment and surrounding factors these progenitor cellscan over a period of three to four weeks in rodents matureinto neuronswhich integrate into existing neuronal networksAlong aging the number of neural stem and progenitor cellsdeclines significant and various steps of neurogenesis (egproliferation survival or maturation) get decelerated [14ndash16] The cause for the decreased levels of proliferation inold individuals is still under debate Putative mechanismscould lengthen cell cycles deplete progenitor cells in the hip-pocampus or cause their quiescence [14] Additionally eventssuch as depression or stress strongly reduce the hippocampalpotential for plasticity [17] Environmental changes howevercan counteract this decline by stimulating neurogenesis andprobably lead to healthy aging [18 19]
To elucidate the mechanisms by which ECT leads toantidepressive effects preclinical research has primarilymade use of experimental electroconvulsive shock (ECS) theanimal model counterpart of ECT A strong enhancementof neurogenesis has been observed in various species fol-lowing experimental ECS treatments [20 21] Several studiesindicated a close relation between hippocampal function andmood regulation The observation of an antidepressive-likeeffect and an upregulation of hippocampal cell proliferationupon experimental ECS raised speculations on the partici-pation of neurogenesis in the antidepressive mode of actionHowever evidence for a direct participation of neurogenesisin antidepressivemechanisms still remains to be convincinglydemonstrated [17]
Based on various correlations observed in experimentalECS animal studies several hypotheses like the stimulation ofneurogenesis the restoration of hippocampal volume mod-ulations of neurotransmitter and hormone levels changes inangiogenesis and cerebral blood flow have been formulatedto account for the antidepressive action of ECT In this reportwe revisited the impact of experimental ECS on neurogenesisand speculate on the mechanisms leading to its upregulationwhich may be also involved in ECTrsquos antidepressive effectsin patients Moreover the influence of aging on the impactof ECTexperimental ECS and neurogenesis has been scruti-nized in more detail
3 Enhanced Neurogenesis followingAntidepressive Treatment Effect or SideEffect of the Therapy
Numerous groups reported a close relation between adultborn hippocampal neurons and antidepressive effects [22]Chronic administration of antidepressants which counteractthe negative behavioural effects of chronic stress leadsreportedly to an upregulation of neurogenesis [23] Addi-tionally evidence supporting the involvement of neurogen-esis in the mode of antidepressant action has subsequentlybeen revealed by Santarelli and colleagues showing thatdeletion of neurogenesis in mice via hippocampal X-rayirradiation ablated the behavioural response to the antide-pressants imipramine and fluoxetine [24] However furtherexperiments displayed that the ablation of hippocampalneurogenesis per se does not elicit a depressive phenotype[25ndash28] Besides it could be shown by Sah and colleaguesusing a mouse model of increased anxiety that rather thelevels of neuronal activity in the dentate gyrus than the ratesof neurogenesis correlate with the depression-like behaviours[29]
To evaluate studies addressing experimental ECSworkingmechanisms it is noteworthy that most studies have beenperformed in healthy animals and only occasionally in animalmodels based on chronic stress These situations certainlydiffer from the complex conditions pertaining in patientswith mental disorders like major depression Despite thisbiological effects of experimental ECS on the central nervoussystem (CNS) in animals can be monitored properly Takentogether the accumulated evidence suggests that depressiondoes not result from an impaired neurogenesis but that theaddition of new neurons in the hippocampal circuitry mayfacilitate the action of antidepressants
It is intriguing that the delayed behavioural response tocommon antidepressant drugs for example SSRIs in rodentscoincides with the delayed upregulation of neurogenesis thatis after approximately three weeks [30] Similarly the rapidantidepressive action of experimental ECS is associatedwith arapid stimulation of neurogenesis which follows after the firsttreatment [31] In this context the enlargement of the imma-ture neuron population may be involved in the behaviouralresponseThese immature cells have been demonstrated to behighly excitable compared tomature neurons [32]Thereforestimulation by lower input intensity could lead to a higherinput sensitivity of the whole surrounding network
Nonetheless one should not get the impression outof these speculations that depression is exclusively a hip-pocampal disorder The involvement of additional cerebralstructures such as the amygdala the frontal cortex and thethalamus has been well documented in studies revealingstructural abnormalities and disbalances of neurotransmit-ters [33ndash35] Hence ECT is likely to act on numerous cerebralstructures possibly stimulating their neuronal plasticity Inthis respect pharmacological antidepressive treatments wereshown to restore the mechanism of long term potentiation(LTP) [36 37] which is impaired in depressed patients andanimalmodels with depressive-like behaviours As previously
Neural Plasticity 3
Day 30Day 0 Differentiation and maturation
ECSPharmacologicaltreatments (eg SSRIs)
Neural stemcellradial gliaScarce symmetric andasymmetric division
Marker NestinGFAPSox2Musashi-1
Neuronal progenitor
Rapid symmetric andasymmetric division
Marker DCXPSA-NCAMNeuroD
Neuroblast
Neuronal fateDifferentiationMigrationSurvivalMarkerDCXPSA-NCAMTUC-4
Young immatureneuronNetwork integration
MarkerDCXNeuNcalretinin
Mature neuron
Network activity
MarkerNeuNcalbindin
Figure 1 Differentiation and maturation of adult neural stem cells The process from stem cell division to the fully integrated matureneuron in the dentate gyrus takes approximately 30 days in rodents Specific markers characterize the various maturation stages (eg DCXfor neuroblasts) Experimental ECS or pharmacological treatments can influence neurogenesis at various stages experimental ECS is alsoreported to activate quiescent stem cells
demonstrated for pharmacological treatments ECT mightstop dendritic atrophy [38 39] or increase dendritic spinedensity in various cortical and limbic structures [40 41]In the hippocampus plasticity is not restricted to synapticplasticity but also involves cellular plasticity that is neuroge-nesis Asmentioned above an enhancednumber of immatureneurons may increase excitability of the hippocampus andputatively associated areas of the limbic system as wellAlternatively or in addition ECT might rewire existingneuronal networks for example via sprouting of mossyfibers that is axonal projections of the granular neurons tothe CA3 pyramidal neurons in the hippocampus restoringthereby functionality of limbic and cortical circuits Althoughit remains to be definitely demonstrated that upregulationof neurogenesis contributes to the antidepressive actionlevels of neurogenesis could become a surrogate marker topredict andmonitor the patientsrsquo response to treatment Eventhough adequate protocols for in vivomeasurement of humanneurogenesis are not existent yet efforts have been recentlydeployed using magnetic resonance spectroscopy (MRS) [42
43] and positron emission tomography (PET) using labelledtracer [44]
4 Differences in ECS and PharmacologicalTreatment on Neural Stem Cells
The finding that experimental ECS strongly induces hip-pocampal neurogenesis in various mammals includingrodents and nonhuman primates triggered the idea of usingECS tomodulate the pool of neural stem and progenitor cellsPharmacological antidepressive treatments like fluoxetineadministration act mainly on amplifying neural progenitors(ANPs) [45] In contrast it has been postulated that experi-mental ECS can in addition shove quiescent neural progeni-tors (QNPs) from their resting phase to a proliferative statusthereby increasing the pool of active stem cells generatingnew progenitor cells [46 47] (Figure 1) Importantly newneurons generated upon experimental ECS-treatment do notdiffer in fate or phenotype as compared to neurons generatedunder physiological conditions [31] Hence new neurons
4 Neural Plasticity
generated following experimental ECS mature and integratein the existing neuronal network and form appropriatesynapses [48 49]
5 Age-Dependent Response to ECT
To date most experimental ECS studies were performed inyoung animals even though depressive episodes must beaddressed also in aged individuals As a group in whichmajor depression disorders are increased and often under-diagnosed the elderly could especially benefit from ECT Onthe one hand this population is often poly-medicated whichincreases the risk of adverse drug interaction upon additionalpharmacological interventions On the other hand ECToffers an alternative to classical pharmacological antide-pressants which fail as first line treatment in the agedpopulation in up to 77 [50] This lack of response toantidepressant drugs might be exacerbated by age-relatedphysiological changes such as decreased concentrations ofacetylcholine and dopamine increased level of monoamineoxidase activity or increased concentration of cortisol due toa dysregulation of the hypothalamic-pituitary-adrenal (HPA)axis [50] Remarkably repeated ECT treatments have shownto be as effective as pharmacological antidepressive therapy inavoiding relapse in aged individuals [51] Nevertheless bothpharmacological and ECT-based approaches benefit from amultimodal therapy concept complemented by physical andmental exercises psychological therapy and resocialisation
So far the mechanisms by which experimental ECS andantidepressant drugs interfere with cells of the neurogenicniches could not be fully deciphered Both types of antide-pressive treatments have been reported to increase the num-ber of proliferating cells in the hippocampus and promote thematuration of neuronal progenitors [5 20 30 52] Under ourexperimental ECS paradigm we could demonstrate a highlysignificant increase of cell proliferation in the dentate gyrus ofmice from the young and aged groups (for details please seesection Material and Methods) (Figure 2) In the 20-month-old mice ECS increased the number of BrdU-labelled cellsby more than sixfold as compared to the aged sham-treatedmice whereas in the 2-month-old mice the increase wasroughly a doubling Most of the BrdU+ cells were located inthe subgranular zone of the dentate gyrus the place whereneural stem cells are resident Intriguingly those BrdU+cells in the 20-month-old group expressed neither Nestina marker for neural stem cells nor Doublecortin (DCX) amarker for neuronal progenitors It is possible that in agedindividuals the maturation steps from stem cells and earlyprogenitors to immature neurons expressing DCX are sloweddown Further studies need to address kinetics and fate ofnewly generated cells after experimental ECS especially inaged individuals
6 How Neurogenesis May Be Supported byModulation of Microenvironment
Neural stem and progenitor cells are embedded in a complexcellular niche with tight regulatory interactions between the
(a)
(b)
5000
4000
3000
2000
1000
0
lowastlowastlowast
lowastlowastlowast
ShamECS
2 20
(months)
BrdU
+ce
lls
(c)
Figure 2 Representative micrographs showing the detection ofBrdU-labelled cells in the dentate gyrus of a 2-month-old mousefrom (a) the sham group or (b) the experimental ECS group Scalebar 200 120583m (c) The number of BrdU+ cells was determined toestimate the rates of cell proliferation in the dorsal DG of 2- and 20-month-old mice Both groups showed a highly significant enhance-ment in cell proliferation following experimental ECS-treatmentTwo months 119875 lt 00001 (lowastlowastlowast) 20 months 119875 = 00004 (lowastlowastlowast)
different cellular partners Not only is this microenvironmentexposed to factors delivered by the blood system but alsostem and progenitor cells are in close contact with cellsforming and surrounding the blood vessels such as pericytesand astrocytes [15 53] This organization referred to as theneurovascular unit is involved in the precise regulation ofneurogenesis [54] It is well documented that endothelialcells astrocytes and microglia modulate the behaviour ofstem and progenitor cells [55ndash57] The impact of experimen-tal ECS on these cell populations was therefore examined Inmice and nonhuman primates histological staining revealedthat experimental ECS increases the expression level of GFAPas well as the number of cells expressing this astrocytemarkerin the hippocampus [20 47 58] In our own experiments we
Neural Plasticity 5
(a)
(b)
ShamECS
2 20
(months)
lowast
ns
80
60
40
20
0
GFA
P+su
rface
in D
G (
)
(c)
Figure 3 Detection of GFAP in the dentate gyrus of (a) sham-treated or (b) ECS-treatedmouse of the 2months of age cohort scalebar in (a) 100120583m (c) Graph showing percentage of area covered byGFAP-expressing cells in the granular layer of dentate gyrus in 2-and 20-month-old mice following ECS or sham treatment A two-way ANOVA detected a significant increase of the GFAP expressionupon experimental ECS (119875 = 00043 (lowastlowast)) Bonferroni post hoctest revealed that the significance resulted from differences in the2-month-old group (119875 lt 005)
focused on GFAP+ cells in the granular layer of the dentategyrus of 2- and 20-month-old mice Overall experimentalECS-treatment increases the expression ofGFAP significantly(Figure 3) A detailed analysis of our data showed an agedependent effect of this treatment hence only the 2-month-old group of mice showed a significant increase of GFAP-expression Some recent studies demonstrated that GFAP-expressing cells in the cortex could be triggered to becomemultipotent neural progenitors [59] It remains to be eluci-dated if experimental ECS and ECT can recruit a pool ofquiescent progenitors within the hippocampus and therebyincrease neurogenesis
Microglia the immune cells of the CNS have been shownto influence neurogenesis [60] Within the granular layer in
the dentate gyrus of young and old mice we detected cellsexpressing the ionized calcium-binding adaptor molecule 1(Iba1) a marker for microglia cells With our experimentalsetup we observed no increase in the amount of Iba1+ cells inthe dentate gyrus (2 months sham 13885 plusmn 1606 cellsmm3versus ECS 12656 plusmn 2480 cellsmm3 119875 = 0511 20months sham 10753 plusmn 955 cellsmm3 versus ECS 11750 plusmn1696 cellsmm3 119875 = 0385) however the experimental ECSenhanced the volume of Iba1+ cells significantly (Figure 4)These enlarged microglia namely hyper-ramified constitutean intermediate state between resting and activated [61] Suchenlarged microglia have been previously reported to appearafter experimental ECS-treatment in rodents [62] Further-more Jansson et al reported an upregulation of MHCII inmicroglia following experimental ECS which also suggestsan activation of these immune cells The group proposedthat theseMHCII-expressing microglia play a significant rolefor maintenance of neurogenesis in the hippocampus Takentogether these observations indicate that experimental ECSmodulates the microglial activation state to the benefit ofneurogenesis and therefore potentially contributes also to theantidepressive effect
7 Blood System and Its Role in ECT
The antidepressive action of ECT can most likely not bereduced to a single mechanism but results from the syner-gistic cooperation of several processes and factors involvingneurogenesis modulation of blood supply and angiogenesiscorticosteroids brain-derived neurotrophic factor (BDNF)vascular endothelial growth factor (VEGF) and so forthIndeed a closer look at these putativemodes of action revealsclose interconnection between the different systems Forinstance proliferation of stem cells located in the neurogenicniches can be modulated by factors delivered via the closelyassociated vasculature [63] In aged mice we showed thatneurogenesis is downregulated by blood derived signallingmolecules that are delivered through the vessels Howeverneurogenesis can be reactivated by the introduction of youngblood into the circulation via direct serum injection orheterochronic parabiosis [64]
Considering that neurogenic niches are closely apposedon blood vessels delivering nutrients and numerous fac-tors it is noteworthy that comorbidities between cardio-vascular diseases and depression have been reported innumerous studies (eg see [65 66]) Moreover PET imag-ing revealed that ECT treatment transiently changes localcerebral blood flow leading to an increase in the brainstem diencephalon and basal ganglia for instance [67]Yet the so-called blood-brain barrier (BBB) constitutes abarrier controlling the molecular exchange between thecirculating blood and the brain parenchyma (reviewed egby Ilbay and colleagues [68]) Roughly speaking the BBBprevents under physiological conditions the diffusion of largeandor hydrophilic molecules but allows for the passage ofhydrophobic molecules and substances having their specifictransporters for example glucose Neuropathological distur-bances like stroke or epileptic seizure can lead to leakage of
6 Neural Plasticity
(a)
ShamECS
2 20
(months)
20
15
10
5
0
Iba1
+su
rface
in D
G (
)(b)
Figure 4 (a)Detection ofmicroglia in the granular layer (GL) of a 2-month-oldmouse based on the expression of Iba1 (white) (MLmolecularlayer) Scale bar 50120583m (b) Microglia volume was estimated according to the percentage of the surface covered by Iba1+ labelling in thegranular layer of 2- and 20-month-old mice following ECS or sham treatment Two-way ANOVA revealed upon ECS a significant increaseof Iba1-labeled surface in the dentate gyrus of both age groups (119875 = 00475)
this barrier [68] Under these conditions various circulatingmolecules can diffuse out of the blood vessels and therebychange the microenvironment of the brain parenchyma andof the neurogenic niches
It has been proposed that experimental ECS leads to agreater permeability of the BBB enabling an influx of blood-derived factors into the parenchyma However leakage of theBBB has not been observed as an acute effect of experimentalECS but was rather seen following several experimental ECSsessions in rodent models [69] For instance no increasedpermeability to Evans Blue was reported by Oztas and collab-orators following a single experimental ECS treatment how-ever BBB leakage could be recognized after ten experimentalECS-sessions [70] The potential BBB leakage after ECT wasfurthermore analysed in humans using different tracers forPET imaging by Bolwig and colleagues who concluded thatdue to changes in local cerebral blood flow and potentiallyfrom the leakage of newly formed capillaries proteins couldpass more easily out of the blood vessels following ECT [71]In a subsequent study on the experimental ECS-induced BBBleakage in rats Sartorius and colleagues reported elevatedBDNF levels in the serum as well as the prefrontal cortexand the hippocampus after five experimental ECS sessionsAccording to their hypothesis a fraction of the parenchymalBDNF originated from the blood circulation and crossedthe BBB after experimental ECS [72] Hence enhanced BBBpermeability might contribute to the beneficial impact ofrepeated ECT session yet the rapid induction of neurogenesisseen after a single experimental ECS session is unlikely linkedto an early failure of the BBB integrity
Strikingly Hattiangady and colleagues reported that thenumber of neural stem cells in the dentate gyrus is notdecreasing with age but only their activation state which wasshown to be closely dependant on the distance of stem cells to
the next blood vessel [73]Therefore experimental ECSmightupregulate neurogenesis by promoting a closer association ofstem cells to the blood vessels Angiogenesis the generationof new blood vessels is tightly regulated during developmentand in the adult brain by numerous factors in a similarfashion to neurogenesis [74] In the healthy brain the densityof blood vessels remains relatively stable during adulthood[75] However increased angiogenesis (and neurogenesis)was observed in SSRI-treated patients suffering from majordepression [74] Similarly experimental ECS treatment wasreported to promote the elongation of existing vessels and toincrease their density in the stratum lacunosum moleculare(SLM) a subregion of the hippocampal molecular layer [7677] However within the granular layer of the dentate gyrusthat is in the vicinity of stem and progenitor cell somaswe did not detect an increase of the blood vessel densityupon experimental ECS treatments (Figure 5) Neverthelessan enhanced vascularization in the molecular layer couldintensify the delivery factors to the tip of radial stem cellsor to the dendritic trees of granular neurons for retrogradetransport and signalling Alternatively factors secreted in themolecular layer could act on other cells types capable of neu-rogenesis modulation for example astrocytes and microgliaFurthermore Newton et al proposed that endothelial cellsper se can influence neurogenesis by secreting factors actingon neural stem and progenitor cells [77]
8 Neurogenesis and the Normalisation ofthe HPA-Axis
The crosstalk between neurogenesis and the ECT putativemodes of action is further exemplified by the modula-tion of the hypothalamic-pituitary-adrenal-axis (HPA-axis)signalling during depression and its interaction with the
Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

2 Neural Plasticity
used in severe cases ofmajor depression as a second line treat-ment following failure of pharmacological approaches [10]Overall according to meta-analyses of clinical studies ECTwas reported to be significantly more effective in depressivedisorders compared to pharmacological treatments [2 11]
2 Are Neurogenesis and AntidepressiveActivity Linked
Adult neurogenesis the generation of new neurons takesplace in numerous mammalian species including humans[12 13] Neurogenesis occurs in at least two specific areas ofthe adult brain namely the dentate gyrus of the hippocampusand the subventricular zone (SVZ) of the lateral ventricles Inthese so-called neurogenic niches neural stem cells (NSCs)can on the one hand divide symmetrically leading to twodaughter stem cells and on the other hand perform anasymmetric division resulting in a daughter stem cell and arapid amplifying progenitor cell Depending on the microen-vironment and surrounding factors these progenitor cellscan over a period of three to four weeks in rodents matureinto neuronswhich integrate into existing neuronal networksAlong aging the number of neural stem and progenitor cellsdeclines significant and various steps of neurogenesis (egproliferation survival or maturation) get decelerated [14ndash16] The cause for the decreased levels of proliferation inold individuals is still under debate Putative mechanismscould lengthen cell cycles deplete progenitor cells in the hip-pocampus or cause their quiescence [14] Additionally eventssuch as depression or stress strongly reduce the hippocampalpotential for plasticity [17] Environmental changes howevercan counteract this decline by stimulating neurogenesis andprobably lead to healthy aging [18 19]
To elucidate the mechanisms by which ECT leads toantidepressive effects preclinical research has primarilymade use of experimental electroconvulsive shock (ECS) theanimal model counterpart of ECT A strong enhancementof neurogenesis has been observed in various species fol-lowing experimental ECS treatments [20 21] Several studiesindicated a close relation between hippocampal function andmood regulation The observation of an antidepressive-likeeffect and an upregulation of hippocampal cell proliferationupon experimental ECS raised speculations on the partici-pation of neurogenesis in the antidepressive mode of actionHowever evidence for a direct participation of neurogenesisin antidepressivemechanisms still remains to be convincinglydemonstrated [17]
Based on various correlations observed in experimentalECS animal studies several hypotheses like the stimulation ofneurogenesis the restoration of hippocampal volume mod-ulations of neurotransmitter and hormone levels changes inangiogenesis and cerebral blood flow have been formulatedto account for the antidepressive action of ECT In this reportwe revisited the impact of experimental ECS on neurogenesisand speculate on the mechanisms leading to its upregulationwhich may be also involved in ECTrsquos antidepressive effectsin patients Moreover the influence of aging on the impactof ECTexperimental ECS and neurogenesis has been scruti-nized in more detail
3 Enhanced Neurogenesis followingAntidepressive Treatment Effect or SideEffect of the Therapy
Numerous groups reported a close relation between adultborn hippocampal neurons and antidepressive effects [22]Chronic administration of antidepressants which counteractthe negative behavioural effects of chronic stress leadsreportedly to an upregulation of neurogenesis [23] Addi-tionally evidence supporting the involvement of neurogen-esis in the mode of antidepressant action has subsequentlybeen revealed by Santarelli and colleagues showing thatdeletion of neurogenesis in mice via hippocampal X-rayirradiation ablated the behavioural response to the antide-pressants imipramine and fluoxetine [24] However furtherexperiments displayed that the ablation of hippocampalneurogenesis per se does not elicit a depressive phenotype[25ndash28] Besides it could be shown by Sah and colleaguesusing a mouse model of increased anxiety that rather thelevels of neuronal activity in the dentate gyrus than the ratesof neurogenesis correlate with the depression-like behaviours[29]
To evaluate studies addressing experimental ECSworkingmechanisms it is noteworthy that most studies have beenperformed in healthy animals and only occasionally in animalmodels based on chronic stress These situations certainlydiffer from the complex conditions pertaining in patientswith mental disorders like major depression Despite thisbiological effects of experimental ECS on the central nervoussystem (CNS) in animals can be monitored properly Takentogether the accumulated evidence suggests that depressiondoes not result from an impaired neurogenesis but that theaddition of new neurons in the hippocampal circuitry mayfacilitate the action of antidepressants
It is intriguing that the delayed behavioural response tocommon antidepressant drugs for example SSRIs in rodentscoincides with the delayed upregulation of neurogenesis thatis after approximately three weeks [30] Similarly the rapidantidepressive action of experimental ECS is associatedwith arapid stimulation of neurogenesis which follows after the firsttreatment [31] In this context the enlargement of the imma-ture neuron population may be involved in the behaviouralresponseThese immature cells have been demonstrated to behighly excitable compared tomature neurons [32]Thereforestimulation by lower input intensity could lead to a higherinput sensitivity of the whole surrounding network
Nonetheless one should not get the impression outof these speculations that depression is exclusively a hip-pocampal disorder The involvement of additional cerebralstructures such as the amygdala the frontal cortex and thethalamus has been well documented in studies revealingstructural abnormalities and disbalances of neurotransmit-ters [33ndash35] Hence ECT is likely to act on numerous cerebralstructures possibly stimulating their neuronal plasticity Inthis respect pharmacological antidepressive treatments wereshown to restore the mechanism of long term potentiation(LTP) [36 37] which is impaired in depressed patients andanimalmodels with depressive-like behaviours As previously
Neural Plasticity 3
Day 30Day 0 Differentiation and maturation
ECSPharmacologicaltreatments (eg SSRIs)
Neural stemcellradial gliaScarce symmetric andasymmetric division
Marker NestinGFAPSox2Musashi-1
Neuronal progenitor
Rapid symmetric andasymmetric division
Marker DCXPSA-NCAMNeuroD
Neuroblast
Neuronal fateDifferentiationMigrationSurvivalMarkerDCXPSA-NCAMTUC-4
Young immatureneuronNetwork integration
MarkerDCXNeuNcalretinin
Mature neuron
Network activity
MarkerNeuNcalbindin
Figure 1 Differentiation and maturation of adult neural stem cells The process from stem cell division to the fully integrated matureneuron in the dentate gyrus takes approximately 30 days in rodents Specific markers characterize the various maturation stages (eg DCXfor neuroblasts) Experimental ECS or pharmacological treatments can influence neurogenesis at various stages experimental ECS is alsoreported to activate quiescent stem cells
demonstrated for pharmacological treatments ECT mightstop dendritic atrophy [38 39] or increase dendritic spinedensity in various cortical and limbic structures [40 41]In the hippocampus plasticity is not restricted to synapticplasticity but also involves cellular plasticity that is neuroge-nesis Asmentioned above an enhancednumber of immatureneurons may increase excitability of the hippocampus andputatively associated areas of the limbic system as wellAlternatively or in addition ECT might rewire existingneuronal networks for example via sprouting of mossyfibers that is axonal projections of the granular neurons tothe CA3 pyramidal neurons in the hippocampus restoringthereby functionality of limbic and cortical circuits Althoughit remains to be definitely demonstrated that upregulationof neurogenesis contributes to the antidepressive actionlevels of neurogenesis could become a surrogate marker topredict andmonitor the patientsrsquo response to treatment Eventhough adequate protocols for in vivomeasurement of humanneurogenesis are not existent yet efforts have been recentlydeployed using magnetic resonance spectroscopy (MRS) [42
43] and positron emission tomography (PET) using labelledtracer [44]
4 Differences in ECS and PharmacologicalTreatment on Neural Stem Cells
The finding that experimental ECS strongly induces hip-pocampal neurogenesis in various mammals includingrodents and nonhuman primates triggered the idea of usingECS tomodulate the pool of neural stem and progenitor cellsPharmacological antidepressive treatments like fluoxetineadministration act mainly on amplifying neural progenitors(ANPs) [45] In contrast it has been postulated that experi-mental ECS can in addition shove quiescent neural progeni-tors (QNPs) from their resting phase to a proliferative statusthereby increasing the pool of active stem cells generatingnew progenitor cells [46 47] (Figure 1) Importantly newneurons generated upon experimental ECS-treatment do notdiffer in fate or phenotype as compared to neurons generatedunder physiological conditions [31] Hence new neurons
4 Neural Plasticity
generated following experimental ECS mature and integratein the existing neuronal network and form appropriatesynapses [48 49]
5 Age-Dependent Response to ECT
To date most experimental ECS studies were performed inyoung animals even though depressive episodes must beaddressed also in aged individuals As a group in whichmajor depression disorders are increased and often under-diagnosed the elderly could especially benefit from ECT Onthe one hand this population is often poly-medicated whichincreases the risk of adverse drug interaction upon additionalpharmacological interventions On the other hand ECToffers an alternative to classical pharmacological antide-pressants which fail as first line treatment in the agedpopulation in up to 77 [50] This lack of response toantidepressant drugs might be exacerbated by age-relatedphysiological changes such as decreased concentrations ofacetylcholine and dopamine increased level of monoamineoxidase activity or increased concentration of cortisol due toa dysregulation of the hypothalamic-pituitary-adrenal (HPA)axis [50] Remarkably repeated ECT treatments have shownto be as effective as pharmacological antidepressive therapy inavoiding relapse in aged individuals [51] Nevertheless bothpharmacological and ECT-based approaches benefit from amultimodal therapy concept complemented by physical andmental exercises psychological therapy and resocialisation
So far the mechanisms by which experimental ECS andantidepressant drugs interfere with cells of the neurogenicniches could not be fully deciphered Both types of antide-pressive treatments have been reported to increase the num-ber of proliferating cells in the hippocampus and promote thematuration of neuronal progenitors [5 20 30 52] Under ourexperimental ECS paradigm we could demonstrate a highlysignificant increase of cell proliferation in the dentate gyrus ofmice from the young and aged groups (for details please seesection Material and Methods) (Figure 2) In the 20-month-old mice ECS increased the number of BrdU-labelled cellsby more than sixfold as compared to the aged sham-treatedmice whereas in the 2-month-old mice the increase wasroughly a doubling Most of the BrdU+ cells were located inthe subgranular zone of the dentate gyrus the place whereneural stem cells are resident Intriguingly those BrdU+cells in the 20-month-old group expressed neither Nestina marker for neural stem cells nor Doublecortin (DCX) amarker for neuronal progenitors It is possible that in agedindividuals the maturation steps from stem cells and earlyprogenitors to immature neurons expressing DCX are sloweddown Further studies need to address kinetics and fate ofnewly generated cells after experimental ECS especially inaged individuals
6 How Neurogenesis May Be Supported byModulation of Microenvironment
Neural stem and progenitor cells are embedded in a complexcellular niche with tight regulatory interactions between the
(a)
(b)
5000
4000
3000
2000
1000
0
lowastlowastlowast
lowastlowastlowast
ShamECS
2 20
(months)
BrdU
+ce
lls
(c)
Figure 2 Representative micrographs showing the detection ofBrdU-labelled cells in the dentate gyrus of a 2-month-old mousefrom (a) the sham group or (b) the experimental ECS group Scalebar 200 120583m (c) The number of BrdU+ cells was determined toestimate the rates of cell proliferation in the dorsal DG of 2- and 20-month-old mice Both groups showed a highly significant enhance-ment in cell proliferation following experimental ECS-treatmentTwo months 119875 lt 00001 (lowastlowastlowast) 20 months 119875 = 00004 (lowastlowastlowast)
different cellular partners Not only is this microenvironmentexposed to factors delivered by the blood system but alsostem and progenitor cells are in close contact with cellsforming and surrounding the blood vessels such as pericytesand astrocytes [15 53] This organization referred to as theneurovascular unit is involved in the precise regulation ofneurogenesis [54] It is well documented that endothelialcells astrocytes and microglia modulate the behaviour ofstem and progenitor cells [55ndash57] The impact of experimen-tal ECS on these cell populations was therefore examined Inmice and nonhuman primates histological staining revealedthat experimental ECS increases the expression level of GFAPas well as the number of cells expressing this astrocytemarkerin the hippocampus [20 47 58] In our own experiments we
Neural Plasticity 5
(a)
(b)
ShamECS
2 20
(months)
lowast
ns
80
60
40
20
0
GFA
P+su
rface
in D
G (
)
(c)
Figure 3 Detection of GFAP in the dentate gyrus of (a) sham-treated or (b) ECS-treatedmouse of the 2months of age cohort scalebar in (a) 100120583m (c) Graph showing percentage of area covered byGFAP-expressing cells in the granular layer of dentate gyrus in 2-and 20-month-old mice following ECS or sham treatment A two-way ANOVA detected a significant increase of the GFAP expressionupon experimental ECS (119875 = 00043 (lowastlowast)) Bonferroni post hoctest revealed that the significance resulted from differences in the2-month-old group (119875 lt 005)
focused on GFAP+ cells in the granular layer of the dentategyrus of 2- and 20-month-old mice Overall experimentalECS-treatment increases the expression ofGFAP significantly(Figure 3) A detailed analysis of our data showed an agedependent effect of this treatment hence only the 2-month-old group of mice showed a significant increase of GFAP-expression Some recent studies demonstrated that GFAP-expressing cells in the cortex could be triggered to becomemultipotent neural progenitors [59] It remains to be eluci-dated if experimental ECS and ECT can recruit a pool ofquiescent progenitors within the hippocampus and therebyincrease neurogenesis
Microglia the immune cells of the CNS have been shownto influence neurogenesis [60] Within the granular layer in
the dentate gyrus of young and old mice we detected cellsexpressing the ionized calcium-binding adaptor molecule 1(Iba1) a marker for microglia cells With our experimentalsetup we observed no increase in the amount of Iba1+ cells inthe dentate gyrus (2 months sham 13885 plusmn 1606 cellsmm3versus ECS 12656 plusmn 2480 cellsmm3 119875 = 0511 20months sham 10753 plusmn 955 cellsmm3 versus ECS 11750 plusmn1696 cellsmm3 119875 = 0385) however the experimental ECSenhanced the volume of Iba1+ cells significantly (Figure 4)These enlarged microglia namely hyper-ramified constitutean intermediate state between resting and activated [61] Suchenlarged microglia have been previously reported to appearafter experimental ECS-treatment in rodents [62] Further-more Jansson et al reported an upregulation of MHCII inmicroglia following experimental ECS which also suggestsan activation of these immune cells The group proposedthat theseMHCII-expressing microglia play a significant rolefor maintenance of neurogenesis in the hippocampus Takentogether these observations indicate that experimental ECSmodulates the microglial activation state to the benefit ofneurogenesis and therefore potentially contributes also to theantidepressive effect
7 Blood System and Its Role in ECT
The antidepressive action of ECT can most likely not bereduced to a single mechanism but results from the syner-gistic cooperation of several processes and factors involvingneurogenesis modulation of blood supply and angiogenesiscorticosteroids brain-derived neurotrophic factor (BDNF)vascular endothelial growth factor (VEGF) and so forthIndeed a closer look at these putativemodes of action revealsclose interconnection between the different systems Forinstance proliferation of stem cells located in the neurogenicniches can be modulated by factors delivered via the closelyassociated vasculature [63] In aged mice we showed thatneurogenesis is downregulated by blood derived signallingmolecules that are delivered through the vessels Howeverneurogenesis can be reactivated by the introduction of youngblood into the circulation via direct serum injection orheterochronic parabiosis [64]
Considering that neurogenic niches are closely apposedon blood vessels delivering nutrients and numerous fac-tors it is noteworthy that comorbidities between cardio-vascular diseases and depression have been reported innumerous studies (eg see [65 66]) Moreover PET imag-ing revealed that ECT treatment transiently changes localcerebral blood flow leading to an increase in the brainstem diencephalon and basal ganglia for instance [67]Yet the so-called blood-brain barrier (BBB) constitutes abarrier controlling the molecular exchange between thecirculating blood and the brain parenchyma (reviewed egby Ilbay and colleagues [68]) Roughly speaking the BBBprevents under physiological conditions the diffusion of largeandor hydrophilic molecules but allows for the passage ofhydrophobic molecules and substances having their specifictransporters for example glucose Neuropathological distur-bances like stroke or epileptic seizure can lead to leakage of
6 Neural Plasticity
(a)
ShamECS
2 20
(months)
20
15
10
5
0
Iba1
+su
rface
in D
G (
)(b)
Figure 4 (a)Detection ofmicroglia in the granular layer (GL) of a 2-month-oldmouse based on the expression of Iba1 (white) (MLmolecularlayer) Scale bar 50120583m (b) Microglia volume was estimated according to the percentage of the surface covered by Iba1+ labelling in thegranular layer of 2- and 20-month-old mice following ECS or sham treatment Two-way ANOVA revealed upon ECS a significant increaseof Iba1-labeled surface in the dentate gyrus of both age groups (119875 = 00475)
this barrier [68] Under these conditions various circulatingmolecules can diffuse out of the blood vessels and therebychange the microenvironment of the brain parenchyma andof the neurogenic niches
It has been proposed that experimental ECS leads to agreater permeability of the BBB enabling an influx of blood-derived factors into the parenchyma However leakage of theBBB has not been observed as an acute effect of experimentalECS but was rather seen following several experimental ECSsessions in rodent models [69] For instance no increasedpermeability to Evans Blue was reported by Oztas and collab-orators following a single experimental ECS treatment how-ever BBB leakage could be recognized after ten experimentalECS-sessions [70] The potential BBB leakage after ECT wasfurthermore analysed in humans using different tracers forPET imaging by Bolwig and colleagues who concluded thatdue to changes in local cerebral blood flow and potentiallyfrom the leakage of newly formed capillaries proteins couldpass more easily out of the blood vessels following ECT [71]In a subsequent study on the experimental ECS-induced BBBleakage in rats Sartorius and colleagues reported elevatedBDNF levels in the serum as well as the prefrontal cortexand the hippocampus after five experimental ECS sessionsAccording to their hypothesis a fraction of the parenchymalBDNF originated from the blood circulation and crossedthe BBB after experimental ECS [72] Hence enhanced BBBpermeability might contribute to the beneficial impact ofrepeated ECT session yet the rapid induction of neurogenesisseen after a single experimental ECS session is unlikely linkedto an early failure of the BBB integrity
Strikingly Hattiangady and colleagues reported that thenumber of neural stem cells in the dentate gyrus is notdecreasing with age but only their activation state which wasshown to be closely dependant on the distance of stem cells to
the next blood vessel [73]Therefore experimental ECSmightupregulate neurogenesis by promoting a closer association ofstem cells to the blood vessels Angiogenesis the generationof new blood vessels is tightly regulated during developmentand in the adult brain by numerous factors in a similarfashion to neurogenesis [74] In the healthy brain the densityof blood vessels remains relatively stable during adulthood[75] However increased angiogenesis (and neurogenesis)was observed in SSRI-treated patients suffering from majordepression [74] Similarly experimental ECS treatment wasreported to promote the elongation of existing vessels and toincrease their density in the stratum lacunosum moleculare(SLM) a subregion of the hippocampal molecular layer [7677] However within the granular layer of the dentate gyrusthat is in the vicinity of stem and progenitor cell somaswe did not detect an increase of the blood vessel densityupon experimental ECS treatments (Figure 5) Neverthelessan enhanced vascularization in the molecular layer couldintensify the delivery factors to the tip of radial stem cellsor to the dendritic trees of granular neurons for retrogradetransport and signalling Alternatively factors secreted in themolecular layer could act on other cells types capable of neu-rogenesis modulation for example astrocytes and microgliaFurthermore Newton et al proposed that endothelial cellsper se can influence neurogenesis by secreting factors actingon neural stem and progenitor cells [77]
8 Neurogenesis and the Normalisation ofthe HPA-Axis
The crosstalk between neurogenesis and the ECT putativemodes of action is further exemplified by the modula-tion of the hypothalamic-pituitary-adrenal-axis (HPA-axis)signalling during depression and its interaction with the
Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

Neural Plasticity 3
Day 30Day 0 Differentiation and maturation
ECSPharmacologicaltreatments (eg SSRIs)
Neural stemcellradial gliaScarce symmetric andasymmetric division
Marker NestinGFAPSox2Musashi-1
Neuronal progenitor
Rapid symmetric andasymmetric division
Marker DCXPSA-NCAMNeuroD
Neuroblast
Neuronal fateDifferentiationMigrationSurvivalMarkerDCXPSA-NCAMTUC-4
Young immatureneuronNetwork integration
MarkerDCXNeuNcalretinin
Mature neuron
Network activity
MarkerNeuNcalbindin
Figure 1 Differentiation and maturation of adult neural stem cells The process from stem cell division to the fully integrated matureneuron in the dentate gyrus takes approximately 30 days in rodents Specific markers characterize the various maturation stages (eg DCXfor neuroblasts) Experimental ECS or pharmacological treatments can influence neurogenesis at various stages experimental ECS is alsoreported to activate quiescent stem cells
demonstrated for pharmacological treatments ECT mightstop dendritic atrophy [38 39] or increase dendritic spinedensity in various cortical and limbic structures [40 41]In the hippocampus plasticity is not restricted to synapticplasticity but also involves cellular plasticity that is neuroge-nesis Asmentioned above an enhancednumber of immatureneurons may increase excitability of the hippocampus andputatively associated areas of the limbic system as wellAlternatively or in addition ECT might rewire existingneuronal networks for example via sprouting of mossyfibers that is axonal projections of the granular neurons tothe CA3 pyramidal neurons in the hippocampus restoringthereby functionality of limbic and cortical circuits Althoughit remains to be definitely demonstrated that upregulationof neurogenesis contributes to the antidepressive actionlevels of neurogenesis could become a surrogate marker topredict andmonitor the patientsrsquo response to treatment Eventhough adequate protocols for in vivomeasurement of humanneurogenesis are not existent yet efforts have been recentlydeployed using magnetic resonance spectroscopy (MRS) [42
43] and positron emission tomography (PET) using labelledtracer [44]
4 Differences in ECS and PharmacologicalTreatment on Neural Stem Cells
The finding that experimental ECS strongly induces hip-pocampal neurogenesis in various mammals includingrodents and nonhuman primates triggered the idea of usingECS tomodulate the pool of neural stem and progenitor cellsPharmacological antidepressive treatments like fluoxetineadministration act mainly on amplifying neural progenitors(ANPs) [45] In contrast it has been postulated that experi-mental ECS can in addition shove quiescent neural progeni-tors (QNPs) from their resting phase to a proliferative statusthereby increasing the pool of active stem cells generatingnew progenitor cells [46 47] (Figure 1) Importantly newneurons generated upon experimental ECS-treatment do notdiffer in fate or phenotype as compared to neurons generatedunder physiological conditions [31] Hence new neurons
4 Neural Plasticity
generated following experimental ECS mature and integratein the existing neuronal network and form appropriatesynapses [48 49]
5 Age-Dependent Response to ECT
To date most experimental ECS studies were performed inyoung animals even though depressive episodes must beaddressed also in aged individuals As a group in whichmajor depression disorders are increased and often under-diagnosed the elderly could especially benefit from ECT Onthe one hand this population is often poly-medicated whichincreases the risk of adverse drug interaction upon additionalpharmacological interventions On the other hand ECToffers an alternative to classical pharmacological antide-pressants which fail as first line treatment in the agedpopulation in up to 77 [50] This lack of response toantidepressant drugs might be exacerbated by age-relatedphysiological changes such as decreased concentrations ofacetylcholine and dopamine increased level of monoamineoxidase activity or increased concentration of cortisol due toa dysregulation of the hypothalamic-pituitary-adrenal (HPA)axis [50] Remarkably repeated ECT treatments have shownto be as effective as pharmacological antidepressive therapy inavoiding relapse in aged individuals [51] Nevertheless bothpharmacological and ECT-based approaches benefit from amultimodal therapy concept complemented by physical andmental exercises psychological therapy and resocialisation
So far the mechanisms by which experimental ECS andantidepressant drugs interfere with cells of the neurogenicniches could not be fully deciphered Both types of antide-pressive treatments have been reported to increase the num-ber of proliferating cells in the hippocampus and promote thematuration of neuronal progenitors [5 20 30 52] Under ourexperimental ECS paradigm we could demonstrate a highlysignificant increase of cell proliferation in the dentate gyrus ofmice from the young and aged groups (for details please seesection Material and Methods) (Figure 2) In the 20-month-old mice ECS increased the number of BrdU-labelled cellsby more than sixfold as compared to the aged sham-treatedmice whereas in the 2-month-old mice the increase wasroughly a doubling Most of the BrdU+ cells were located inthe subgranular zone of the dentate gyrus the place whereneural stem cells are resident Intriguingly those BrdU+cells in the 20-month-old group expressed neither Nestina marker for neural stem cells nor Doublecortin (DCX) amarker for neuronal progenitors It is possible that in agedindividuals the maturation steps from stem cells and earlyprogenitors to immature neurons expressing DCX are sloweddown Further studies need to address kinetics and fate ofnewly generated cells after experimental ECS especially inaged individuals
6 How Neurogenesis May Be Supported byModulation of Microenvironment
Neural stem and progenitor cells are embedded in a complexcellular niche with tight regulatory interactions between the
(a)
(b)
5000
4000
3000
2000
1000
0
lowastlowastlowast
lowastlowastlowast
ShamECS
2 20
(months)
BrdU
+ce
lls
(c)
Figure 2 Representative micrographs showing the detection ofBrdU-labelled cells in the dentate gyrus of a 2-month-old mousefrom (a) the sham group or (b) the experimental ECS group Scalebar 200 120583m (c) The number of BrdU+ cells was determined toestimate the rates of cell proliferation in the dorsal DG of 2- and 20-month-old mice Both groups showed a highly significant enhance-ment in cell proliferation following experimental ECS-treatmentTwo months 119875 lt 00001 (lowastlowastlowast) 20 months 119875 = 00004 (lowastlowastlowast)
different cellular partners Not only is this microenvironmentexposed to factors delivered by the blood system but alsostem and progenitor cells are in close contact with cellsforming and surrounding the blood vessels such as pericytesand astrocytes [15 53] This organization referred to as theneurovascular unit is involved in the precise regulation ofneurogenesis [54] It is well documented that endothelialcells astrocytes and microglia modulate the behaviour ofstem and progenitor cells [55ndash57] The impact of experimen-tal ECS on these cell populations was therefore examined Inmice and nonhuman primates histological staining revealedthat experimental ECS increases the expression level of GFAPas well as the number of cells expressing this astrocytemarkerin the hippocampus [20 47 58] In our own experiments we
Neural Plasticity 5
(a)
(b)
ShamECS
2 20
(months)
lowast
ns
80
60
40
20
0
GFA
P+su
rface
in D
G (
)
(c)
Figure 3 Detection of GFAP in the dentate gyrus of (a) sham-treated or (b) ECS-treatedmouse of the 2months of age cohort scalebar in (a) 100120583m (c) Graph showing percentage of area covered byGFAP-expressing cells in the granular layer of dentate gyrus in 2-and 20-month-old mice following ECS or sham treatment A two-way ANOVA detected a significant increase of the GFAP expressionupon experimental ECS (119875 = 00043 (lowastlowast)) Bonferroni post hoctest revealed that the significance resulted from differences in the2-month-old group (119875 lt 005)
focused on GFAP+ cells in the granular layer of the dentategyrus of 2- and 20-month-old mice Overall experimentalECS-treatment increases the expression ofGFAP significantly(Figure 3) A detailed analysis of our data showed an agedependent effect of this treatment hence only the 2-month-old group of mice showed a significant increase of GFAP-expression Some recent studies demonstrated that GFAP-expressing cells in the cortex could be triggered to becomemultipotent neural progenitors [59] It remains to be eluci-dated if experimental ECS and ECT can recruit a pool ofquiescent progenitors within the hippocampus and therebyincrease neurogenesis
Microglia the immune cells of the CNS have been shownto influence neurogenesis [60] Within the granular layer in
the dentate gyrus of young and old mice we detected cellsexpressing the ionized calcium-binding adaptor molecule 1(Iba1) a marker for microglia cells With our experimentalsetup we observed no increase in the amount of Iba1+ cells inthe dentate gyrus (2 months sham 13885 plusmn 1606 cellsmm3versus ECS 12656 plusmn 2480 cellsmm3 119875 = 0511 20months sham 10753 plusmn 955 cellsmm3 versus ECS 11750 plusmn1696 cellsmm3 119875 = 0385) however the experimental ECSenhanced the volume of Iba1+ cells significantly (Figure 4)These enlarged microglia namely hyper-ramified constitutean intermediate state between resting and activated [61] Suchenlarged microglia have been previously reported to appearafter experimental ECS-treatment in rodents [62] Further-more Jansson et al reported an upregulation of MHCII inmicroglia following experimental ECS which also suggestsan activation of these immune cells The group proposedthat theseMHCII-expressing microglia play a significant rolefor maintenance of neurogenesis in the hippocampus Takentogether these observations indicate that experimental ECSmodulates the microglial activation state to the benefit ofneurogenesis and therefore potentially contributes also to theantidepressive effect
7 Blood System and Its Role in ECT
The antidepressive action of ECT can most likely not bereduced to a single mechanism but results from the syner-gistic cooperation of several processes and factors involvingneurogenesis modulation of blood supply and angiogenesiscorticosteroids brain-derived neurotrophic factor (BDNF)vascular endothelial growth factor (VEGF) and so forthIndeed a closer look at these putativemodes of action revealsclose interconnection between the different systems Forinstance proliferation of stem cells located in the neurogenicniches can be modulated by factors delivered via the closelyassociated vasculature [63] In aged mice we showed thatneurogenesis is downregulated by blood derived signallingmolecules that are delivered through the vessels Howeverneurogenesis can be reactivated by the introduction of youngblood into the circulation via direct serum injection orheterochronic parabiosis [64]
Considering that neurogenic niches are closely apposedon blood vessels delivering nutrients and numerous fac-tors it is noteworthy that comorbidities between cardio-vascular diseases and depression have been reported innumerous studies (eg see [65 66]) Moreover PET imag-ing revealed that ECT treatment transiently changes localcerebral blood flow leading to an increase in the brainstem diencephalon and basal ganglia for instance [67]Yet the so-called blood-brain barrier (BBB) constitutes abarrier controlling the molecular exchange between thecirculating blood and the brain parenchyma (reviewed egby Ilbay and colleagues [68]) Roughly speaking the BBBprevents under physiological conditions the diffusion of largeandor hydrophilic molecules but allows for the passage ofhydrophobic molecules and substances having their specifictransporters for example glucose Neuropathological distur-bances like stroke or epileptic seizure can lead to leakage of
6 Neural Plasticity
(a)
ShamECS
2 20
(months)
20
15
10
5
0
Iba1
+su
rface
in D
G (
)(b)
Figure 4 (a)Detection ofmicroglia in the granular layer (GL) of a 2-month-oldmouse based on the expression of Iba1 (white) (MLmolecularlayer) Scale bar 50120583m (b) Microglia volume was estimated according to the percentage of the surface covered by Iba1+ labelling in thegranular layer of 2- and 20-month-old mice following ECS or sham treatment Two-way ANOVA revealed upon ECS a significant increaseof Iba1-labeled surface in the dentate gyrus of both age groups (119875 = 00475)
this barrier [68] Under these conditions various circulatingmolecules can diffuse out of the blood vessels and therebychange the microenvironment of the brain parenchyma andof the neurogenic niches
It has been proposed that experimental ECS leads to agreater permeability of the BBB enabling an influx of blood-derived factors into the parenchyma However leakage of theBBB has not been observed as an acute effect of experimentalECS but was rather seen following several experimental ECSsessions in rodent models [69] For instance no increasedpermeability to Evans Blue was reported by Oztas and collab-orators following a single experimental ECS treatment how-ever BBB leakage could be recognized after ten experimentalECS-sessions [70] The potential BBB leakage after ECT wasfurthermore analysed in humans using different tracers forPET imaging by Bolwig and colleagues who concluded thatdue to changes in local cerebral blood flow and potentiallyfrom the leakage of newly formed capillaries proteins couldpass more easily out of the blood vessels following ECT [71]In a subsequent study on the experimental ECS-induced BBBleakage in rats Sartorius and colleagues reported elevatedBDNF levels in the serum as well as the prefrontal cortexand the hippocampus after five experimental ECS sessionsAccording to their hypothesis a fraction of the parenchymalBDNF originated from the blood circulation and crossedthe BBB after experimental ECS [72] Hence enhanced BBBpermeability might contribute to the beneficial impact ofrepeated ECT session yet the rapid induction of neurogenesisseen after a single experimental ECS session is unlikely linkedto an early failure of the BBB integrity
Strikingly Hattiangady and colleagues reported that thenumber of neural stem cells in the dentate gyrus is notdecreasing with age but only their activation state which wasshown to be closely dependant on the distance of stem cells to
the next blood vessel [73]Therefore experimental ECSmightupregulate neurogenesis by promoting a closer association ofstem cells to the blood vessels Angiogenesis the generationof new blood vessels is tightly regulated during developmentand in the adult brain by numerous factors in a similarfashion to neurogenesis [74] In the healthy brain the densityof blood vessels remains relatively stable during adulthood[75] However increased angiogenesis (and neurogenesis)was observed in SSRI-treated patients suffering from majordepression [74] Similarly experimental ECS treatment wasreported to promote the elongation of existing vessels and toincrease their density in the stratum lacunosum moleculare(SLM) a subregion of the hippocampal molecular layer [7677] However within the granular layer of the dentate gyrusthat is in the vicinity of stem and progenitor cell somaswe did not detect an increase of the blood vessel densityupon experimental ECS treatments (Figure 5) Neverthelessan enhanced vascularization in the molecular layer couldintensify the delivery factors to the tip of radial stem cellsor to the dendritic trees of granular neurons for retrogradetransport and signalling Alternatively factors secreted in themolecular layer could act on other cells types capable of neu-rogenesis modulation for example astrocytes and microgliaFurthermore Newton et al proposed that endothelial cellsper se can influence neurogenesis by secreting factors actingon neural stem and progenitor cells [77]
8 Neurogenesis and the Normalisation ofthe HPA-Axis
The crosstalk between neurogenesis and the ECT putativemodes of action is further exemplified by the modula-tion of the hypothalamic-pituitary-adrenal-axis (HPA-axis)signalling during depression and its interaction with the
Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013
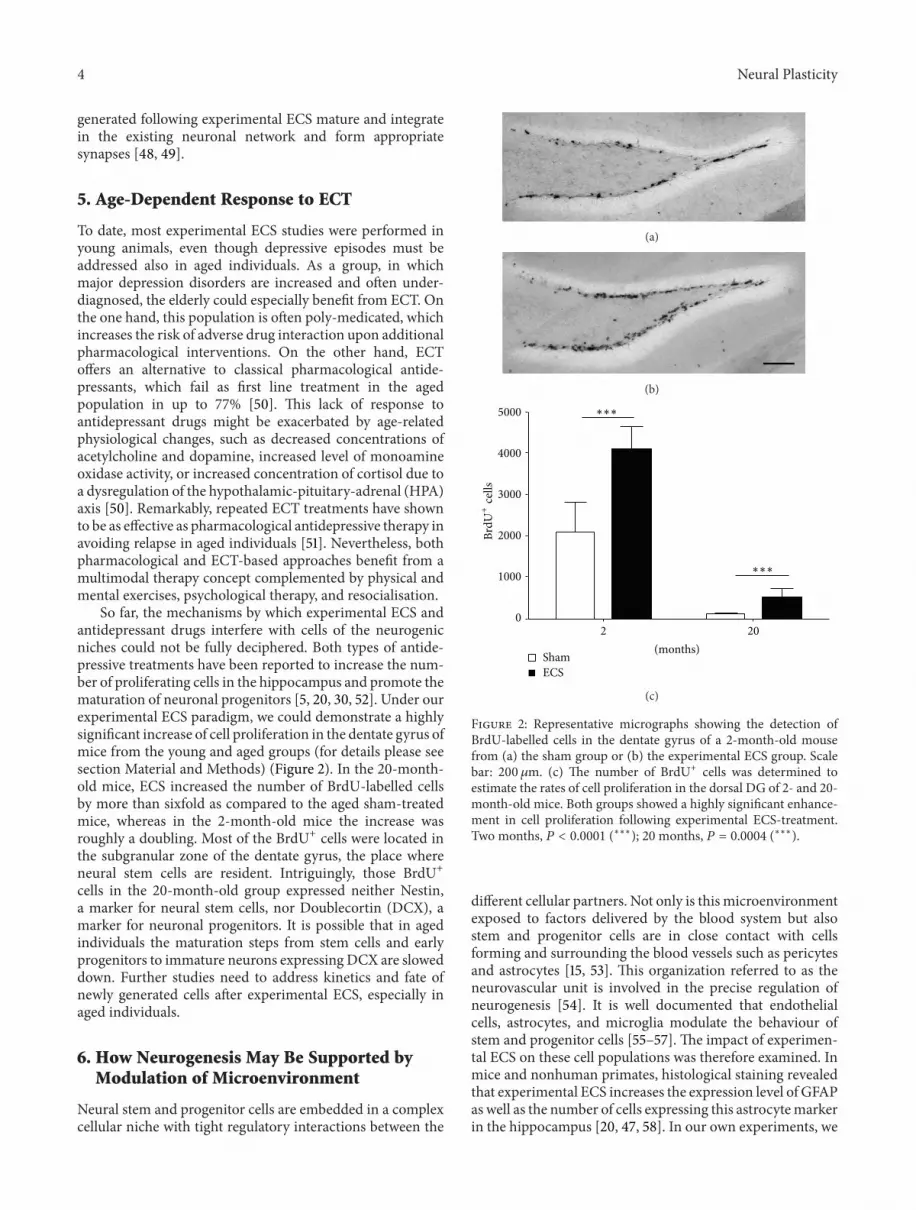
4 Neural Plasticity
generated following experimental ECS mature and integratein the existing neuronal network and form appropriatesynapses [48 49]
5 Age-Dependent Response to ECT
To date most experimental ECS studies were performed inyoung animals even though depressive episodes must beaddressed also in aged individuals As a group in whichmajor depression disorders are increased and often under-diagnosed the elderly could especially benefit from ECT Onthe one hand this population is often poly-medicated whichincreases the risk of adverse drug interaction upon additionalpharmacological interventions On the other hand ECToffers an alternative to classical pharmacological antide-pressants which fail as first line treatment in the agedpopulation in up to 77 [50] This lack of response toantidepressant drugs might be exacerbated by age-relatedphysiological changes such as decreased concentrations ofacetylcholine and dopamine increased level of monoamineoxidase activity or increased concentration of cortisol due toa dysregulation of the hypothalamic-pituitary-adrenal (HPA)axis [50] Remarkably repeated ECT treatments have shownto be as effective as pharmacological antidepressive therapy inavoiding relapse in aged individuals [51] Nevertheless bothpharmacological and ECT-based approaches benefit from amultimodal therapy concept complemented by physical andmental exercises psychological therapy and resocialisation
So far the mechanisms by which experimental ECS andantidepressant drugs interfere with cells of the neurogenicniches could not be fully deciphered Both types of antide-pressive treatments have been reported to increase the num-ber of proliferating cells in the hippocampus and promote thematuration of neuronal progenitors [5 20 30 52] Under ourexperimental ECS paradigm we could demonstrate a highlysignificant increase of cell proliferation in the dentate gyrus ofmice from the young and aged groups (for details please seesection Material and Methods) (Figure 2) In the 20-month-old mice ECS increased the number of BrdU-labelled cellsby more than sixfold as compared to the aged sham-treatedmice whereas in the 2-month-old mice the increase wasroughly a doubling Most of the BrdU+ cells were located inthe subgranular zone of the dentate gyrus the place whereneural stem cells are resident Intriguingly those BrdU+cells in the 20-month-old group expressed neither Nestina marker for neural stem cells nor Doublecortin (DCX) amarker for neuronal progenitors It is possible that in agedindividuals the maturation steps from stem cells and earlyprogenitors to immature neurons expressing DCX are sloweddown Further studies need to address kinetics and fate ofnewly generated cells after experimental ECS especially inaged individuals
6 How Neurogenesis May Be Supported byModulation of Microenvironment
Neural stem and progenitor cells are embedded in a complexcellular niche with tight regulatory interactions between the
(a)
(b)
5000
4000
3000
2000
1000
0
lowastlowastlowast
lowastlowastlowast
ShamECS
2 20
(months)
BrdU
+ce
lls
(c)
Figure 2 Representative micrographs showing the detection ofBrdU-labelled cells in the dentate gyrus of a 2-month-old mousefrom (a) the sham group or (b) the experimental ECS group Scalebar 200 120583m (c) The number of BrdU+ cells was determined toestimate the rates of cell proliferation in the dorsal DG of 2- and 20-month-old mice Both groups showed a highly significant enhance-ment in cell proliferation following experimental ECS-treatmentTwo months 119875 lt 00001 (lowastlowastlowast) 20 months 119875 = 00004 (lowastlowastlowast)
different cellular partners Not only is this microenvironmentexposed to factors delivered by the blood system but alsostem and progenitor cells are in close contact with cellsforming and surrounding the blood vessels such as pericytesand astrocytes [15 53] This organization referred to as theneurovascular unit is involved in the precise regulation ofneurogenesis [54] It is well documented that endothelialcells astrocytes and microglia modulate the behaviour ofstem and progenitor cells [55ndash57] The impact of experimen-tal ECS on these cell populations was therefore examined Inmice and nonhuman primates histological staining revealedthat experimental ECS increases the expression level of GFAPas well as the number of cells expressing this astrocytemarkerin the hippocampus [20 47 58] In our own experiments we
Neural Plasticity 5
(a)
(b)
ShamECS
2 20
(months)
lowast
ns
80
60
40
20
0
GFA
P+su
rface
in D
G (
)
(c)
Figure 3 Detection of GFAP in the dentate gyrus of (a) sham-treated or (b) ECS-treatedmouse of the 2months of age cohort scalebar in (a) 100120583m (c) Graph showing percentage of area covered byGFAP-expressing cells in the granular layer of dentate gyrus in 2-and 20-month-old mice following ECS or sham treatment A two-way ANOVA detected a significant increase of the GFAP expressionupon experimental ECS (119875 = 00043 (lowastlowast)) Bonferroni post hoctest revealed that the significance resulted from differences in the2-month-old group (119875 lt 005)
focused on GFAP+ cells in the granular layer of the dentategyrus of 2- and 20-month-old mice Overall experimentalECS-treatment increases the expression ofGFAP significantly(Figure 3) A detailed analysis of our data showed an agedependent effect of this treatment hence only the 2-month-old group of mice showed a significant increase of GFAP-expression Some recent studies demonstrated that GFAP-expressing cells in the cortex could be triggered to becomemultipotent neural progenitors [59] It remains to be eluci-dated if experimental ECS and ECT can recruit a pool ofquiescent progenitors within the hippocampus and therebyincrease neurogenesis
Microglia the immune cells of the CNS have been shownto influence neurogenesis [60] Within the granular layer in
the dentate gyrus of young and old mice we detected cellsexpressing the ionized calcium-binding adaptor molecule 1(Iba1) a marker for microglia cells With our experimentalsetup we observed no increase in the amount of Iba1+ cells inthe dentate gyrus (2 months sham 13885 plusmn 1606 cellsmm3versus ECS 12656 plusmn 2480 cellsmm3 119875 = 0511 20months sham 10753 plusmn 955 cellsmm3 versus ECS 11750 plusmn1696 cellsmm3 119875 = 0385) however the experimental ECSenhanced the volume of Iba1+ cells significantly (Figure 4)These enlarged microglia namely hyper-ramified constitutean intermediate state between resting and activated [61] Suchenlarged microglia have been previously reported to appearafter experimental ECS-treatment in rodents [62] Further-more Jansson et al reported an upregulation of MHCII inmicroglia following experimental ECS which also suggestsan activation of these immune cells The group proposedthat theseMHCII-expressing microglia play a significant rolefor maintenance of neurogenesis in the hippocampus Takentogether these observations indicate that experimental ECSmodulates the microglial activation state to the benefit ofneurogenesis and therefore potentially contributes also to theantidepressive effect
7 Blood System and Its Role in ECT
The antidepressive action of ECT can most likely not bereduced to a single mechanism but results from the syner-gistic cooperation of several processes and factors involvingneurogenesis modulation of blood supply and angiogenesiscorticosteroids brain-derived neurotrophic factor (BDNF)vascular endothelial growth factor (VEGF) and so forthIndeed a closer look at these putativemodes of action revealsclose interconnection between the different systems Forinstance proliferation of stem cells located in the neurogenicniches can be modulated by factors delivered via the closelyassociated vasculature [63] In aged mice we showed thatneurogenesis is downregulated by blood derived signallingmolecules that are delivered through the vessels Howeverneurogenesis can be reactivated by the introduction of youngblood into the circulation via direct serum injection orheterochronic parabiosis [64]
Considering that neurogenic niches are closely apposedon blood vessels delivering nutrients and numerous fac-tors it is noteworthy that comorbidities between cardio-vascular diseases and depression have been reported innumerous studies (eg see [65 66]) Moreover PET imag-ing revealed that ECT treatment transiently changes localcerebral blood flow leading to an increase in the brainstem diencephalon and basal ganglia for instance [67]Yet the so-called blood-brain barrier (BBB) constitutes abarrier controlling the molecular exchange between thecirculating blood and the brain parenchyma (reviewed egby Ilbay and colleagues [68]) Roughly speaking the BBBprevents under physiological conditions the diffusion of largeandor hydrophilic molecules but allows for the passage ofhydrophobic molecules and substances having their specifictransporters for example glucose Neuropathological distur-bances like stroke or epileptic seizure can lead to leakage of
6 Neural Plasticity
(a)
ShamECS
2 20
(months)
20
15
10
5
0
Iba1
+su
rface
in D
G (
)(b)
Figure 4 (a)Detection ofmicroglia in the granular layer (GL) of a 2-month-oldmouse based on the expression of Iba1 (white) (MLmolecularlayer) Scale bar 50120583m (b) Microglia volume was estimated according to the percentage of the surface covered by Iba1+ labelling in thegranular layer of 2- and 20-month-old mice following ECS or sham treatment Two-way ANOVA revealed upon ECS a significant increaseof Iba1-labeled surface in the dentate gyrus of both age groups (119875 = 00475)
this barrier [68] Under these conditions various circulatingmolecules can diffuse out of the blood vessels and therebychange the microenvironment of the brain parenchyma andof the neurogenic niches
It has been proposed that experimental ECS leads to agreater permeability of the BBB enabling an influx of blood-derived factors into the parenchyma However leakage of theBBB has not been observed as an acute effect of experimentalECS but was rather seen following several experimental ECSsessions in rodent models [69] For instance no increasedpermeability to Evans Blue was reported by Oztas and collab-orators following a single experimental ECS treatment how-ever BBB leakage could be recognized after ten experimentalECS-sessions [70] The potential BBB leakage after ECT wasfurthermore analysed in humans using different tracers forPET imaging by Bolwig and colleagues who concluded thatdue to changes in local cerebral blood flow and potentiallyfrom the leakage of newly formed capillaries proteins couldpass more easily out of the blood vessels following ECT [71]In a subsequent study on the experimental ECS-induced BBBleakage in rats Sartorius and colleagues reported elevatedBDNF levels in the serum as well as the prefrontal cortexand the hippocampus after five experimental ECS sessionsAccording to their hypothesis a fraction of the parenchymalBDNF originated from the blood circulation and crossedthe BBB after experimental ECS [72] Hence enhanced BBBpermeability might contribute to the beneficial impact ofrepeated ECT session yet the rapid induction of neurogenesisseen after a single experimental ECS session is unlikely linkedto an early failure of the BBB integrity
Strikingly Hattiangady and colleagues reported that thenumber of neural stem cells in the dentate gyrus is notdecreasing with age but only their activation state which wasshown to be closely dependant on the distance of stem cells to
the next blood vessel [73]Therefore experimental ECSmightupregulate neurogenesis by promoting a closer association ofstem cells to the blood vessels Angiogenesis the generationof new blood vessels is tightly regulated during developmentand in the adult brain by numerous factors in a similarfashion to neurogenesis [74] In the healthy brain the densityof blood vessels remains relatively stable during adulthood[75] However increased angiogenesis (and neurogenesis)was observed in SSRI-treated patients suffering from majordepression [74] Similarly experimental ECS treatment wasreported to promote the elongation of existing vessels and toincrease their density in the stratum lacunosum moleculare(SLM) a subregion of the hippocampal molecular layer [7677] However within the granular layer of the dentate gyrusthat is in the vicinity of stem and progenitor cell somaswe did not detect an increase of the blood vessel densityupon experimental ECS treatments (Figure 5) Neverthelessan enhanced vascularization in the molecular layer couldintensify the delivery factors to the tip of radial stem cellsor to the dendritic trees of granular neurons for retrogradetransport and signalling Alternatively factors secreted in themolecular layer could act on other cells types capable of neu-rogenesis modulation for example astrocytes and microgliaFurthermore Newton et al proposed that endothelial cellsper se can influence neurogenesis by secreting factors actingon neural stem and progenitor cells [77]
8 Neurogenesis and the Normalisation ofthe HPA-Axis
The crosstalk between neurogenesis and the ECT putativemodes of action is further exemplified by the modula-tion of the hypothalamic-pituitary-adrenal-axis (HPA-axis)signalling during depression and its interaction with the
Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013
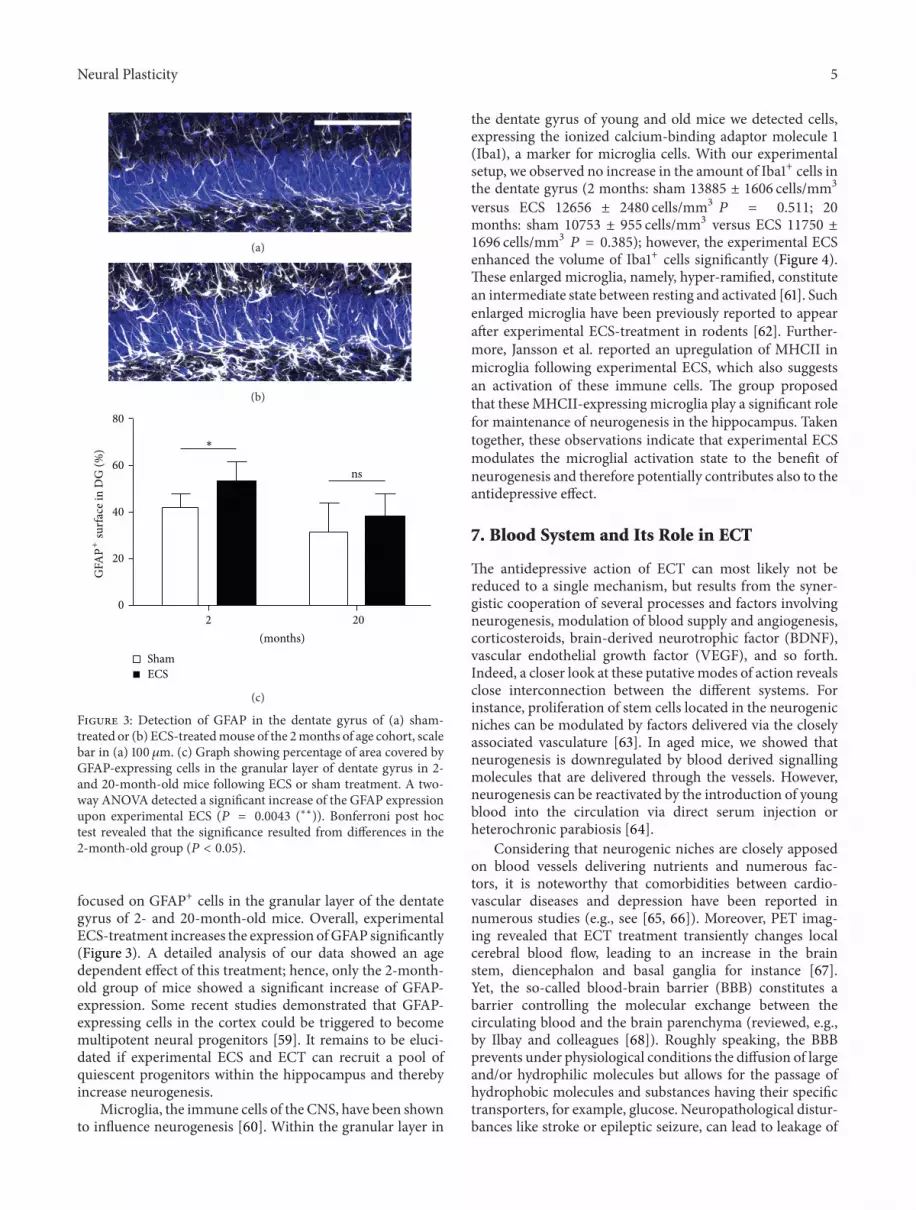
Neural Plasticity 5
(a)
(b)
ShamECS
2 20
(months)
lowast
ns
80
60
40
20
0
GFA
P+su
rface
in D
G (
)
(c)
Figure 3 Detection of GFAP in the dentate gyrus of (a) sham-treated or (b) ECS-treatedmouse of the 2months of age cohort scalebar in (a) 100120583m (c) Graph showing percentage of area covered byGFAP-expressing cells in the granular layer of dentate gyrus in 2-and 20-month-old mice following ECS or sham treatment A two-way ANOVA detected a significant increase of the GFAP expressionupon experimental ECS (119875 = 00043 (lowastlowast)) Bonferroni post hoctest revealed that the significance resulted from differences in the2-month-old group (119875 lt 005)
focused on GFAP+ cells in the granular layer of the dentategyrus of 2- and 20-month-old mice Overall experimentalECS-treatment increases the expression ofGFAP significantly(Figure 3) A detailed analysis of our data showed an agedependent effect of this treatment hence only the 2-month-old group of mice showed a significant increase of GFAP-expression Some recent studies demonstrated that GFAP-expressing cells in the cortex could be triggered to becomemultipotent neural progenitors [59] It remains to be eluci-dated if experimental ECS and ECT can recruit a pool ofquiescent progenitors within the hippocampus and therebyincrease neurogenesis
Microglia the immune cells of the CNS have been shownto influence neurogenesis [60] Within the granular layer in
the dentate gyrus of young and old mice we detected cellsexpressing the ionized calcium-binding adaptor molecule 1(Iba1) a marker for microglia cells With our experimentalsetup we observed no increase in the amount of Iba1+ cells inthe dentate gyrus (2 months sham 13885 plusmn 1606 cellsmm3versus ECS 12656 plusmn 2480 cellsmm3 119875 = 0511 20months sham 10753 plusmn 955 cellsmm3 versus ECS 11750 plusmn1696 cellsmm3 119875 = 0385) however the experimental ECSenhanced the volume of Iba1+ cells significantly (Figure 4)These enlarged microglia namely hyper-ramified constitutean intermediate state between resting and activated [61] Suchenlarged microglia have been previously reported to appearafter experimental ECS-treatment in rodents [62] Further-more Jansson et al reported an upregulation of MHCII inmicroglia following experimental ECS which also suggestsan activation of these immune cells The group proposedthat theseMHCII-expressing microglia play a significant rolefor maintenance of neurogenesis in the hippocampus Takentogether these observations indicate that experimental ECSmodulates the microglial activation state to the benefit ofneurogenesis and therefore potentially contributes also to theantidepressive effect
7 Blood System and Its Role in ECT
The antidepressive action of ECT can most likely not bereduced to a single mechanism but results from the syner-gistic cooperation of several processes and factors involvingneurogenesis modulation of blood supply and angiogenesiscorticosteroids brain-derived neurotrophic factor (BDNF)vascular endothelial growth factor (VEGF) and so forthIndeed a closer look at these putativemodes of action revealsclose interconnection between the different systems Forinstance proliferation of stem cells located in the neurogenicniches can be modulated by factors delivered via the closelyassociated vasculature [63] In aged mice we showed thatneurogenesis is downregulated by blood derived signallingmolecules that are delivered through the vessels Howeverneurogenesis can be reactivated by the introduction of youngblood into the circulation via direct serum injection orheterochronic parabiosis [64]
Considering that neurogenic niches are closely apposedon blood vessels delivering nutrients and numerous fac-tors it is noteworthy that comorbidities between cardio-vascular diseases and depression have been reported innumerous studies (eg see [65 66]) Moreover PET imag-ing revealed that ECT treatment transiently changes localcerebral blood flow leading to an increase in the brainstem diencephalon and basal ganglia for instance [67]Yet the so-called blood-brain barrier (BBB) constitutes abarrier controlling the molecular exchange between thecirculating blood and the brain parenchyma (reviewed egby Ilbay and colleagues [68]) Roughly speaking the BBBprevents under physiological conditions the diffusion of largeandor hydrophilic molecules but allows for the passage ofhydrophobic molecules and substances having their specifictransporters for example glucose Neuropathological distur-bances like stroke or epileptic seizure can lead to leakage of
6 Neural Plasticity
(a)
ShamECS
2 20
(months)
20
15
10
5
0
Iba1
+su
rface
in D
G (
)(b)
Figure 4 (a)Detection ofmicroglia in the granular layer (GL) of a 2-month-oldmouse based on the expression of Iba1 (white) (MLmolecularlayer) Scale bar 50120583m (b) Microglia volume was estimated according to the percentage of the surface covered by Iba1+ labelling in thegranular layer of 2- and 20-month-old mice following ECS or sham treatment Two-way ANOVA revealed upon ECS a significant increaseof Iba1-labeled surface in the dentate gyrus of both age groups (119875 = 00475)
this barrier [68] Under these conditions various circulatingmolecules can diffuse out of the blood vessels and therebychange the microenvironment of the brain parenchyma andof the neurogenic niches
It has been proposed that experimental ECS leads to agreater permeability of the BBB enabling an influx of blood-derived factors into the parenchyma However leakage of theBBB has not been observed as an acute effect of experimentalECS but was rather seen following several experimental ECSsessions in rodent models [69] For instance no increasedpermeability to Evans Blue was reported by Oztas and collab-orators following a single experimental ECS treatment how-ever BBB leakage could be recognized after ten experimentalECS-sessions [70] The potential BBB leakage after ECT wasfurthermore analysed in humans using different tracers forPET imaging by Bolwig and colleagues who concluded thatdue to changes in local cerebral blood flow and potentiallyfrom the leakage of newly formed capillaries proteins couldpass more easily out of the blood vessels following ECT [71]In a subsequent study on the experimental ECS-induced BBBleakage in rats Sartorius and colleagues reported elevatedBDNF levels in the serum as well as the prefrontal cortexand the hippocampus after five experimental ECS sessionsAccording to their hypothesis a fraction of the parenchymalBDNF originated from the blood circulation and crossedthe BBB after experimental ECS [72] Hence enhanced BBBpermeability might contribute to the beneficial impact ofrepeated ECT session yet the rapid induction of neurogenesisseen after a single experimental ECS session is unlikely linkedto an early failure of the BBB integrity
Strikingly Hattiangady and colleagues reported that thenumber of neural stem cells in the dentate gyrus is notdecreasing with age but only their activation state which wasshown to be closely dependant on the distance of stem cells to
the next blood vessel [73]Therefore experimental ECSmightupregulate neurogenesis by promoting a closer association ofstem cells to the blood vessels Angiogenesis the generationof new blood vessels is tightly regulated during developmentand in the adult brain by numerous factors in a similarfashion to neurogenesis [74] In the healthy brain the densityof blood vessels remains relatively stable during adulthood[75] However increased angiogenesis (and neurogenesis)was observed in SSRI-treated patients suffering from majordepression [74] Similarly experimental ECS treatment wasreported to promote the elongation of existing vessels and toincrease their density in the stratum lacunosum moleculare(SLM) a subregion of the hippocampal molecular layer [7677] However within the granular layer of the dentate gyrusthat is in the vicinity of stem and progenitor cell somaswe did not detect an increase of the blood vessel densityupon experimental ECS treatments (Figure 5) Neverthelessan enhanced vascularization in the molecular layer couldintensify the delivery factors to the tip of radial stem cellsor to the dendritic trees of granular neurons for retrogradetransport and signalling Alternatively factors secreted in themolecular layer could act on other cells types capable of neu-rogenesis modulation for example astrocytes and microgliaFurthermore Newton et al proposed that endothelial cellsper se can influence neurogenesis by secreting factors actingon neural stem and progenitor cells [77]
8 Neurogenesis and the Normalisation ofthe HPA-Axis
The crosstalk between neurogenesis and the ECT putativemodes of action is further exemplified by the modula-tion of the hypothalamic-pituitary-adrenal-axis (HPA-axis)signalling during depression and its interaction with the
Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013
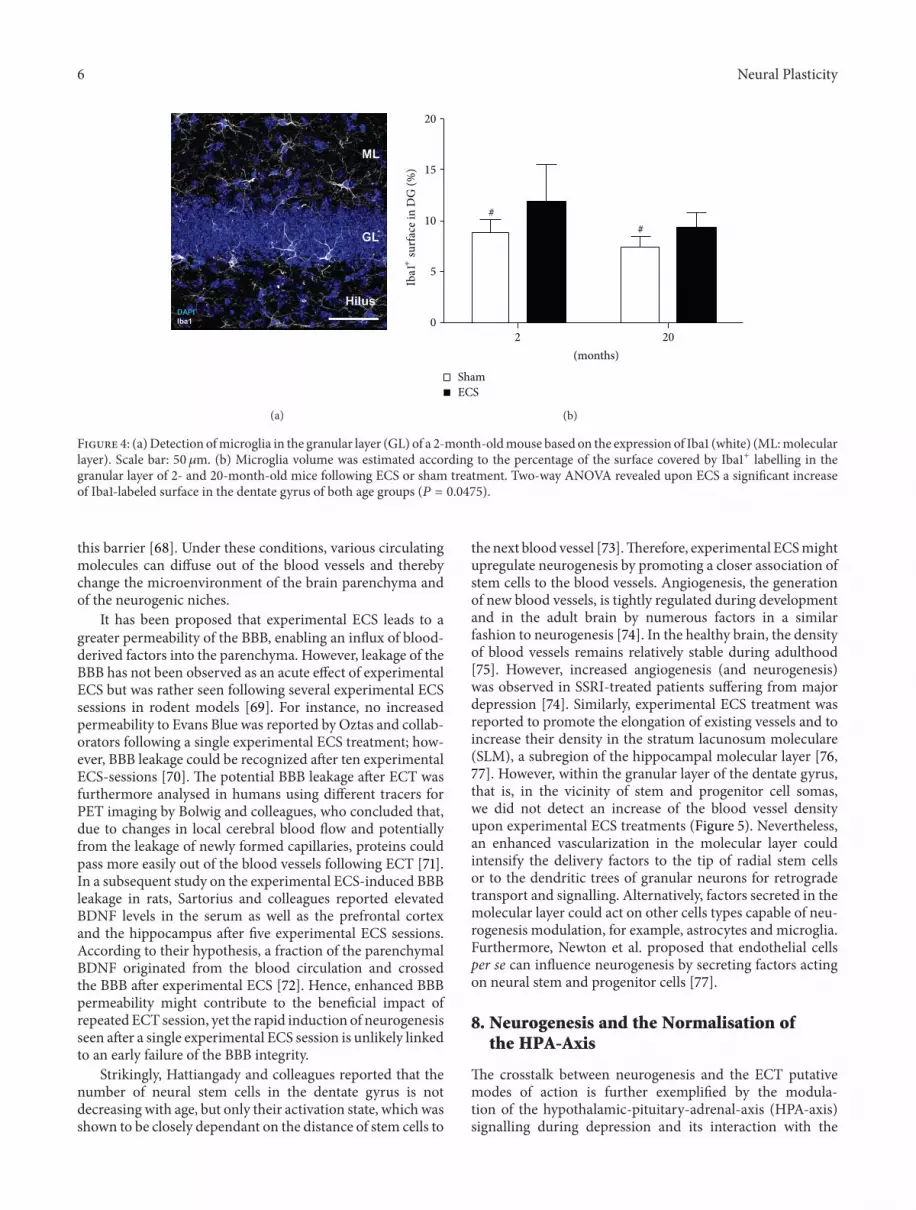
6 Neural Plasticity
(a)
ShamECS
2 20
(months)
20
15
10
5
0
Iba1
+su
rface
in D
G (
)(b)
Figure 4 (a)Detection ofmicroglia in the granular layer (GL) of a 2-month-oldmouse based on the expression of Iba1 (white) (MLmolecularlayer) Scale bar 50120583m (b) Microglia volume was estimated according to the percentage of the surface covered by Iba1+ labelling in thegranular layer of 2- and 20-month-old mice following ECS or sham treatment Two-way ANOVA revealed upon ECS a significant increaseof Iba1-labeled surface in the dentate gyrus of both age groups (119875 = 00475)
this barrier [68] Under these conditions various circulatingmolecules can diffuse out of the blood vessels and therebychange the microenvironment of the brain parenchyma andof the neurogenic niches
It has been proposed that experimental ECS leads to agreater permeability of the BBB enabling an influx of blood-derived factors into the parenchyma However leakage of theBBB has not been observed as an acute effect of experimentalECS but was rather seen following several experimental ECSsessions in rodent models [69] For instance no increasedpermeability to Evans Blue was reported by Oztas and collab-orators following a single experimental ECS treatment how-ever BBB leakage could be recognized after ten experimentalECS-sessions [70] The potential BBB leakage after ECT wasfurthermore analysed in humans using different tracers forPET imaging by Bolwig and colleagues who concluded thatdue to changes in local cerebral blood flow and potentiallyfrom the leakage of newly formed capillaries proteins couldpass more easily out of the blood vessels following ECT [71]In a subsequent study on the experimental ECS-induced BBBleakage in rats Sartorius and colleagues reported elevatedBDNF levels in the serum as well as the prefrontal cortexand the hippocampus after five experimental ECS sessionsAccording to their hypothesis a fraction of the parenchymalBDNF originated from the blood circulation and crossedthe BBB after experimental ECS [72] Hence enhanced BBBpermeability might contribute to the beneficial impact ofrepeated ECT session yet the rapid induction of neurogenesisseen after a single experimental ECS session is unlikely linkedto an early failure of the BBB integrity
Strikingly Hattiangady and colleagues reported that thenumber of neural stem cells in the dentate gyrus is notdecreasing with age but only their activation state which wasshown to be closely dependant on the distance of stem cells to
the next blood vessel [73]Therefore experimental ECSmightupregulate neurogenesis by promoting a closer association ofstem cells to the blood vessels Angiogenesis the generationof new blood vessels is tightly regulated during developmentand in the adult brain by numerous factors in a similarfashion to neurogenesis [74] In the healthy brain the densityof blood vessels remains relatively stable during adulthood[75] However increased angiogenesis (and neurogenesis)was observed in SSRI-treated patients suffering from majordepression [74] Similarly experimental ECS treatment wasreported to promote the elongation of existing vessels and toincrease their density in the stratum lacunosum moleculare(SLM) a subregion of the hippocampal molecular layer [7677] However within the granular layer of the dentate gyrusthat is in the vicinity of stem and progenitor cell somaswe did not detect an increase of the blood vessel densityupon experimental ECS treatments (Figure 5) Neverthelessan enhanced vascularization in the molecular layer couldintensify the delivery factors to the tip of radial stem cellsor to the dendritic trees of granular neurons for retrogradetransport and signalling Alternatively factors secreted in themolecular layer could act on other cells types capable of neu-rogenesis modulation for example astrocytes and microgliaFurthermore Newton et al proposed that endothelial cellsper se can influence neurogenesis by secreting factors actingon neural stem and progenitor cells [77]
8 Neurogenesis and the Normalisation ofthe HPA-Axis
The crosstalk between neurogenesis and the ECT putativemodes of action is further exemplified by the modula-tion of the hypothalamic-pituitary-adrenal-axis (HPA-axis)signalling during depression and its interaction with the
Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

Neural Plasticity 7
(a)
ShamECS
2 20
(months)
ns
ns
6
4
2
0
Lect
in+
surfa
ce o
f DG
()
(b)
Figure 5 (a) Detection of blood vessels in the granular layer of the dorsal dentate gyrus with B simplicifolia lectin (red) (inset overview ofthe dentate gyrus with position of the field of view ML molecular layer GL granular layer) Scale bar 50 120583m (b) No significant differencesin blood vessel density could be detected following ECS Interestingly aged brains had a slightly decreased vessel density compared to theyounger group (119875 = 00122)
neurogenesis [78ndash81] In depressed patients the regulationof the HPA-axis is heavily disturbed The malfunction ofthe negative feedback mechanisms results in an excessivesecretion of the hormone cortisol Interestingly increasedlevels of cortisol and defective HPA-axis response were alsoreported in aged individuals [82] In general the level ofglucocorticoids negatively correlates with the proliferationrates of neural stem and progenitor cells which possess thereceptors for glucocorticoid and mineralocorticoids [83 84]Surget and colleagues established an important link betweenneurogenesis and the antidepressive effect of fluoxetine byidentifying a direct correlation between newly generatedneurons and the restoration of adequate control on theHPA stress response system [85] The report demonstratednamely that the generation of new neurons is required forthe function of the negative feedback loop of the HPA-axis while others critically discuss a direct correlation [86]Interestingly high levels of cortisol have also antiangiogenicproperties [87] Long-lasting elevated cortisol levels suchas in the aged individual have been proposed to induceepigenetic alterations in neural stem and progenitor cellswhich could lead to their long term quiescence and thereforeto a significant reduction of neurogenesis [88]
Preclinical research on experimental ECS has been per-formed often in young and healthy animals which do notshow signs of depressive behaviour For a better under-standing of the processes taking place in the depressedbrain and the impact of ECT on the CNS adequate ani-mal models showing depressive-like behaviour with ele-vated corticosteroid-levels should be selected This could beachieved artificially by direct injection of corticosteroidsApplication of large doses of corticosteroids was reportedto inhibit neurogenesis in a rat model In this model cellproliferation in the dentate gyrus could be restored to
physiological levels by experimental ECS treatment [89]Hence experimental ECS may be indirectly involved inthe normalization of the HPA-axis regulation through theactivation of neurogenesis
9 Release of Signalling Factors followingExperimental ECS and ECT
In animal models experimental ECS activates additionallyneurons of the hypothalamic-pituitary-system namely theparaventricular nucleus supraoptic nucleus and ventrome-dial nucleus (PVN SON and VMH) which regulate thesecretion of diverse depression-relevant HPA-axis-relatedhormones such as adrenocorticotropin neuropeptide Y(NPY) prolactin and vasopressin [62 90ndash94] It has beenreported that NPY concentrations were decreased in thecerebrospinal fluid (CSF) of depressed patients but throughECT treatments the level could be normalized again [95ndash97]The link between NPY and depression was further validatedin rodents in which a direct administration of NPY into thelateral cerebral ventricles led to an antidepressive effect [94]
In addition to the modulation of HPA-related neuropep-tides experimental ECS treatment leads to higher concentra-tion of several neurogenicneurotrophic factors like BDNFVEGF neuritin nerve growth factor (NGF) and fibroblastgrowth factor 2 (FGF2) within the CNS [46 98ndash100] Astronger secretion of these factors and the induction of cellproliferation can be observed even after a single experimentalECS however the proliferation level will further increasewith multiple experimental ECS sessions [21 101 102] Thebiological activities of factors induced by experimental ECSare manifold For instance the neurotrophic factor neuritinwhich is one of the most upregulated genes in the granule
8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

8 Neural Plasticity
(a)
ShamECS
2 20
(months)
04
03
02
01
00
Volu
me (
mm
3)
nslowastlowast
(b)
Figure 6 (a) Overview of a murine hippocampus with its substructures cornu ammonis (CA) 1ndash3 lateral ventricle (LV) stratum lacunosummoleculare (SLM) molecular layer (ML) granular layer (GL) and the hilus Scale bar 200 120583m (b) Volume of the dorsal dentate gyrus of2- and 20-month-old mice following ECS or sham-treatment Young mice showed a significant increase of volume after ECS (2 months119875 = 0002 (lowastlowast) 20 months (ns))
cell layer of the dentate gyrus following experimental ECSinduces neurite outgrowth [103] Other upregulated factorslike FGF-2 possess mitogenic neurotrophic and neuro-protective capabilities [104] Finally VEGF besides beingone of the most potent angiogenic factors also promotesneurogenesis after experimental ECS [46 63 105 106]
Importantly experimental ECS treatments increase thelevels of BDNF in areas of the brain relevant to depressionsuch as the entorhinal cortex and the hippocampus [99 107]The observation that the infusion of BDNF into the midbrainor hippocampus led to an antidepressive effect substantiatedthe role of BDNF in experimental ECS mode of action [108109] BDNF was also shown to increase synaptic strengthneuronal survival and integration but not the proliferation[110 111] In animal models receiving chronic administrationof antidepressant drugs the appearance of the antidepressiveactivity coincided with the increase of BDNF concentrationsand relied on TrkB-signalling [112] Interestingly secretion ofproBDNF the precursor of mature BDNF was increased inrat hippocampal synaptosomes even after one experimentalECS session and could be further enhanced by repeatedsessions [113]
The neurogenic impact of BDNF not only is dependenton its net concentration in the brain but also is determinedby the BDNF subtypes expressed BDNF has at least 34different transcripts which can be translated tomature formsof BDNF [114] Alternative and age dependent regulation invarious brain regions and involvement of BDNF in diversefunctions complicated the analysis of BDNF signalling [115]It is noteworthy that a polymorphic variant of BDNF (G169A)might associate with major depression and carriers of thispolymorphismwere observed to have a reduced hippocampalvolume Nevertheless these individuals do not differ in their
response to ECT in comparison to other depressed patients[116]
10 Gain of Hippocampal Volume followingExperimental ECS and ECT
Several studies pointed out a significant hippocampal atrophyin depressed patients and animal models of depression[117ndash121] This decline of hippocampal volume could bedirectly correlated with the duration of untreated depression[120] Postmortem studies revealed that the hippocampalvolume reduction originated from a reduction of neuronalvolume and of the glial density [122ndash124] Nevertheless anormal hippocampal volume could be regained followingECT-treatment of depressed patients [125] Interestingly thehippocampal volume could also be significantly increased byECS session in nondepressed rodent and non-human primatemodels [20]
As mentioned above the brains of young and old differin several aspects including changes in neurotransmitterconcentrations decline of stem and progenitor cell prolifer-ation or alterations of hormone levels In our experimentsexperimental ECS treatments could slightly but significantlyincrease the volume of the dentate gyrus of the youngercohort of mice (Figure 6) In the 20-month-old mice thevolume increase failed to reach significance Hence themechanisms leading to the volumetric changes followingexperimental ECS appear to be dampenedwith ageing whichmaydisclose a reduced plasticity and correlatewith the higherrates of depression recurrence observed in the elderly popula-tion [126] Although the validity of volumetricmeasurementsas a biomarker for the monitoring of depression remains to
Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

Neural Plasticity 9
be established the rapid volumetric response to experimentalECS treatment may reflect the responsiveness of mechanismscontrolling plasticity of the brain
11 Epilepsy and ExperimentalECSECT-Induced Plasticity
ECT shares some similarities with epileptic seizures forinstance a massive excitation of neuronal networks and anenhanced cell proliferation in dentate gyrus It is noteworthythat experimental ECS in contrast to epileptic seizuresresults neither in neurotoxicity and apoptosis nor in theappearance of ectopic neurons in the hilus of the hip-pocampus [21 76] A proper wiring between the dentategyrus and its target area namely the CA3 is important forcorrect processing of information entering the hippocampalcircuitry This connectivity has been reported to be aberrantin the brains of patients suffering of temporal lobe epilepsyor following experimental seizures in rodents [127] On theother hand experimental ECS sessions promote sproutingof mossy fibers without generation of abnormal connectivityand therefore do not lead to the appearance of spontaneousepileptic activities [128 129] This is in clear contrast with thestimulation of hippocampal neurogenesis through epilepticseizures which results in the generation of new neuronscharacterized by structural and functional abnormalities[130]
The addition of new neurons and rewiring of hippocam-pal networks likely contribute to the antidepressive activityof experimental ECS although the mechanisms are still to beelucidated Recent evidence suggests that the antidepressiveeffect is not proportional to the number of newly generatedneurons but rather to the activation of the whole neuronalpopulation [86] Substantiating this hypothesis is the obser-vation that c-fos a marker for cellular activity is stronglyupregulated in mature neurons of the dentate gyrus afterexperimental ECS [110] Sah and colleagues further reportedthat the antidepressive effects of fluoxetine in a mousemodel of anxiety disorder resulted from the restorationof the hypoactive dentate gyrus back to a normal activitylevel and not from an elevation of the neurogenesis rate[29] Nevertheless taken that newly generated neurons aresignificantly more excitable than their mature counterpartsincreasing neurogenesis may also result in the increase ofglobal dentate gyrus activity [32 131 132]
12 Outlook
There is still an important debate on the antidepressive modeof actions of ECT but undoubtedly this treatment is a fast andpowerful therapy with few side effects and simultaneouslya very potent enhancer of hippocampal neurogenesis Evena decade after the report that antidepressants increase therate of hippocampal neurogenesis relation between the latterand the antidepressive activity could not be fully decipheredNevertheless evidence for a functional relevance of adulthippocampal neurogenesis in processes of neuronal plasticityand behaviours is slowly emerging
13 Material and Methods
In the experiment described in this review we involvedC57Bl6 mice of two and twenty months of age in orderto reflect the various age groups receiving antidepressivetreatments Number of mice were 11 (sham) and 11 (ECS)in the 2-month-old group and 8 (sham) and 9 (ECS) in the20-month-old group male and female equally distributedExperiments were performed in conformity with the Direc-tive (201063EU) of the European Parliament and of theCouncil and were approved by the local animal healthcommission Animals were bred in the central animal facilityof the Paracelsus Medical University Salzburg Austria Allanimals were housed under standard conditions of a 12-hourlightdark cycle with food and water ad libitum
These mice were submitted to a daily session of exper-imental ECS for five consecutive days Anaesthesia rightbefore shock application was obtained using 25 isofluranein pure oxygen ECS parameters applied were as followsfrequency 100 pulsessec pulse width 03ms shock duration1 sec and current 18mA using an Ugo Basil 57800 ECTUnit via ear clips moisten with Ringerrsquos Solution Sham-treated mice were only anaesthetized daily with isofluraneIn addition dividing cells were marked by intraperitoneallyBrdU injection (50mgkg bodyweight) daily at the end ofevery ECS or sham treatment
For histological analysis mice were perfused intracar-dially on day 6 with phosphate buffered 4 paraformalde-hyde pH74 Thereafter brains were further post-fixedovernight in the same solution and then cryoprotected with a30 sucrose solution for several days Brains were cut sagittalinto 40 120583m thick sections using a sliding microtome on dryice (Leica Nuszligloch Germany)
Immunohistological analyses were performed as previ-ously described [133] using the following antibodies andkits dividing cells were labelled with rat anti-BrdU (1500Serotec Puchheim Germany) and detected with a biotin-conjugated rabbit anti-rat (11000 Vector Burlingame USA)followed by the VECTASTAIN ABC System and DAB Per-oxidase Substrate kit (Vector Burlingame USA) Microgliawere labelled with rabbit anti-Iba1 (1300 Wako NeussGermany) and detected with donkey anti-rabbit Alexa 647(11000 Milipore Billerica USA) For blood-vessel stainingbiotinylated Lectin (1500 Sigma-Aldrich Vienna Austria)from B simplicifolia was used followed by Streptavidin Alexa488 (1400 Invitrogen Lofer Austria) Nuclei in fluorescencestainings were marked with 410158406-diamidino-2-phenylindole(DAPI 05 120583gmL Sigma-Aldrich Vienna Austria)
Extrapolation for one brain hemisphere of the totalnumber of BrdU or Iba1-labelled cells located in the dentategyrus was performed by detection of BrdU in every tenthsection or Iba1 in 3 randomly selected fields of view Pictureswere acquired with an Olympus IX81 (Olympus ViennaAustria) microscope using the Volocity Software (PerkinElmer)
The surface labelled with anti-GFAP anti-Iba1 or Lectinwas estimated as previously described [134 135] Briefly z-stack pictures with a resolution of 1024times1024 pixels coveringthe whole 40 120583m slice were acquired using a LSM 700
10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

10 Neural Plasticity
confocal microscope (Carl Zeiss Jena Germany) with a 20xobjective and Zeiss ZEN 2011 software Stacks were mergedusing ImageJ Software 146r (National Institutes of HealthUSA) The granular layer of dentate gyrus was outlined andconverted to grey scale (Adobe Photoshop CS2 Adobe SanJose USA) prior to the selection of a detection thresholdThepercentage of pixels containing labelling within the dentategyrus was quantified
For statistical analysis GraphPad Prism 50 (GraphPadSoftware Inc) was used with two-tailed two-way ANOVAand Bonferroni post hoc test Graphs show mean values withstandard deviation as error bars Significance was 119875 lt 005lowast119875 lt 001
lowastlowast and 119875 lt 0001lowastlowastlowast
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work has been made possible through the support fromthe State Government of Salzburg (Austria) the founda-tion Propter Homines (Liechtenstein) and the FWF SpecialResearch Program (SFB) F44 ldquoCell Signaling in Chronic CNSDisordersrdquo The research leading to these results has receivedfunding from the European Unionrsquos Seventh FrameworkProgramme for research technological development anddemonstration under Grant agreement no 279288 (IDEA)and no 278850 (INMiND) We acknowledge the support ofthe Core Facility Microscopy at SCI-TReCS
References
[1] A Ingram M M Saling and I Schweitzer ldquoCognitive sideeffects of brief pulse electroconvulsive therapy a reviewrdquoJournal of ECT vol 24 no 1 pp 3ndash9 2008
[2] T U E Group ldquoEfficacy and safety of electroconvulsive therapyin depressive disorders a systematic review and meta-analysisrdquoThe Lancet vol 361 no 9360 pp 799ndash808 2003
[3] T Baghai R Frey S Kasper and H-J Moller Elektrokonvul-sionstherapie Klinische und Wissenschaftliche Aspekte vol 483Springer Vienna Austria 2004
[4] J R Epp C L Beasley and L A Galea ldquoIncreased hippocampalneurogenesis and p21 expression in depression dependent onantidepressants sex age and antipsychotic exposurerdquo Neu-ropsychopharmacology vol 38 no 11 pp 2297ndash2306 2013
[5] S Couillard-Despres C Wuertinger M Kandasamy et alldquoAgeing abolishes the effects of fluoxetine on neurogenesisrdquoMolecular Psychiatry vol 14 no 9 pp 856ndash864 2009
[6] B Cronholm and J O Ottosson ldquoUltrabrief stimulus tech-nique in electroconvulsive therapy II Comparative studiesof therapeutic effects and memory disturbances in treatmentof endogenous depression with the elther es electroshockapparatus and siemens konvulsator IIIrdquoThe Journal of Nervousand Mental Disease vol 137 pp 268ndash276 1963
[7] L R Squire and P C Slater ldquoBilateral and unilateral ECTeffects on verbal and nonverbal memoryrdquo American Journal ofPsychiatry vol 135 no 11 pp 1316ndash1320 1978
[8] L R Squire and J A Zouzounis ldquoECT andmemory brief pulseversus sine waverdquo American Journal of Psychiatry vol 143 no5 pp 596ndash601 1986
[9] M Fink ldquoElectroconvulsive therapy resurrected its successesand promises after 75 yearsrdquoCanadian Journal of Psychiatry vol56 no 1 pp 3ndash4 2011
[10] C Salzman E Wong and B C Wright ldquoDrug and ECTtreatment of depression in the elderly 1996ndash2001 a literaturereviewrdquo Biological Psychiatry vol 52 no 3 pp 265ndash284 2002
[11] S Sarkar and S Grover ldquoA systematic review and meta-analysisof trials of treatment of depression from Indiardquo Indian Journalof Psychiatry vol 56 no 1 pp 29ndash38 2014
[12] P S Eriksson E Perfilieva T Bjork-Eriksson et al ldquoNeurogen-esis in the adult human hippocampusrdquo Nature Medicine vol 4no 11 pp 1313ndash1317 1998
[13] I Amrein K Isler andH-P Lipp ldquoComparing adult hippocam-pal neurogenesis in mammalian species and orders influenceof chronological age and life history stagerdquo European Journal ofNeuroscience vol 34 no 6 pp 978ndash987 2011
[14] E Drapeau and D N Abrous ldquoStem cell review series role ofneurogenesis in age-related memory disordersrdquo Aging Cell vol7 no 4 pp 569ndash589 2008
[15] C J Miranda L Braun Y Jiang et al ldquoAging brain microen-vironment decreases hippocampal neurogenesis through Wnt-mediated survivin signalingrdquo Aging Cell vol 11 no 3 pp 542ndash552 2012
[16] VMHeine SMaslamM Joels and P J Lucassen ldquoProminentdecline of newborn cell proliferation differentiation and apop-tosis in the aging dentate gyrus in absence of an age-relatedhypothalamus-pituitary-adrenal axis activationrdquo Neurobiologyof Aging vol 25 no 3 pp 361ndash375 2004
[17] P J Lucassen M W Stumpel Q Wang and E AronicaldquoDecreased numbers of progenitor cells but no response toantidepressant drugs in the hippocampus of elderly depressedpatientsrdquo Neuropharmacology vol 58 no 6 pp 940ndash949 2010
[18] G Kempermann H G Kuhn and F H Gage ldquoExperience-induced neurogenesis in the senescent dentate gyrusrdquo Journalof Neuroscience vol 18 no 9 pp 3206ndash3212 1998
[19] H G Kuhn H Dickinson-Anson and F H Gage ldquoNeurogen-esis in the dentate gyrus of the adult rat age-related decrease ofneuronal progenitor proliferationrdquo Journal of Neuroscience vol16 no 6 pp 2027ndash2033 1996
[20] T D Perera J D Coplan S H Lisanby et al ldquoAntidepressant-induced neurogenesis in the hippocampus of adult nonhumanprimatesrdquo Journal of Neuroscience vol 27 no 18 pp 4894ndash49012007
[21] T M Madsen A Treschow J Bengzon T G Bolwig OLindvall andA Tingstrom ldquoIncreased neurogenesis in amodelof electroconvulsive therapyrdquo Biological Psychiatry vol 47 no12 pp 1043ndash1049 2000
[22] A J Eisch and D Petrik ldquoDepression and hippocampal neu-rogenesis a road to remissionrdquo Science vol 338 no 6103 pp72ndash75 2012
[23] R S Duman S Nakagawa and J Malberg ldquoRegulation of adultneurogenesis by antidepressant treatmentrdquo Neuropsychophar-macology vol 25 no 6 pp 836ndash844 2001
[24] L Santarelli M Saxe C Gross et al ldquoRequirement of hip-pocampal neurogenesis for the behavioral effects of antidepres-santsrdquo Science vol 301 no 5634 pp 805ndash809 2003
[25] K A Holick D C Lee R Hen and S C Dulawa ldquoBehavioraleffects of chronic fluoxetine in BALBcJ mice do not require
Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

Neural Plasticity 11
adult hippocampal neurogenesis or the serotonin 1A receptorrdquoNeuropsychopharmacology vol 33 no 2 pp 406ndash417 2008
[26] C ZhaoW Deng and F H Gage ldquoMechanisms and functionalimplications of adult neurogenesisrdquoCell vol 132 no 4 pp 645ndash660 2008
[27] A Sahay and R Hen ldquoAdult hippocampal neurogenesis indepressionrdquo Nature Neuroscience vol 10 no 9 pp 1110ndash11152007
[28] P Jedynak P Jaholkowski GWozniak C Sandi L Kaczmarekand R K Filipkowski ldquoLack of cyclin D2 impairing adult brainneurogenesis alters hippocampal-dependent behavioral taskswithout reducing learning abilityrdquo Behavioural Brain Researchvol 227 no 1 pp 159ndash166 2012
[29] A Sah C Schmuckermair S B Sartori et al ldquoAnxiety- ratherthan depression-like behavior is associated with adult neuro-genesis in a female mouse model of higher trait anxiety- andcomorbid depression-like behaviorrdquo Translational psychiatryvol 2 article e171 2012
[30] J E Malberg A J Eisch E J Nestler and R S DumanldquoChronic antidepressant treatment increases neurogenesis inadult rat hippocampusrdquo Journal of Neuroscience vol 20 no 24pp 9104ndash9110 2000
[31] K Nakamura M Ito Y Liu T Seki T Suzuki and H AraildquoEffects of single and repeated electroconvulsive stimulation onhippocampal cell proliferation and spontaneous behaviors inthe ratrdquo Brain Research vol 1491 pp 88ndash97 2013
[32] S Couillard-Despres B Winner C Karl et al ldquoTargetedtransgene expression in neuronal precursors watching youngneurons in the old brainrdquo European Journal of Neuroscience vol24 no 6 pp 1535ndash1545 2006
[33] G Sanacora G F Mason D L Rothman et al ldquoIncreasedcortical GABA concentrations in depressed patients receivingECTrdquo American Journal of Psychiatry vol 160 no 3 pp 577ndash579 2003
[34] J J Mann ldquoNeurobiological correlates of the antidepressantaction of electroconvulsive therapyrdquo Journal of ECT vol 14 no3 pp 172ndash180 1998
[35] M Markianos J Hatzimanolis and L Lykouras ldquoRelation-ship between prolactin responses to ECT and dopaminergicand serotonergic responsivity in depressed patientsrdquo EuropeanArchives of Psychiatry and Clinical Neuroscience vol 252 no 4pp 166ndash171 2002
[36] W Li L Liu Y-Y Liu et al ldquoEffects of electroconvulsivestimulation on long-termpotentiation and synaptophysin in thehippocampus of rats with depressive behaviorrdquo Journal of ECTvol 28 no 2 pp 111ndash117 2012
[37] K G Bath D Q Jing I Dincheva et al ldquoBDNF Val66Metimpairs fluoxetine-induced enhancement of adult hippocam-pus plasticityrdquo Neuropsychopharmacology vol 37 no 5 pp1297ndash1304 2012
[38] J M Bessa D Ferreira I Melo et al ldquoThe mood-improvingactions of antidepressants do not depend on neurogenesis butare associatedwith neuronal remodelingrdquoMolecular Psychiatryvol 14 no 8 pp 764ndash773 2009
[39] A M Magarinos and B S McEwen ldquoStress-induced atrophy ofapical dendrites of hippocampal CA3c neurons involvement ofglucocorticoid secretion and excitatory amino acid receptorsrdquoNeuroscience vol 69 no 1 pp 89ndash98 1995
[40] R S Duman and G K Aghajanian ldquoSynaptic dysfunction indepression potential therapeutic targetsrdquo Science vol 338 no6103 pp 68ndash72 2012
[41] E Ampuero F J Rubio R Falcon et al ldquoChronic fluoxetinetreatment induces structural plasticity and selective changesin glutamate receptor subunits in the rat cerebral cortexrdquoNeuroscience vol 169 no 1 pp 98ndash108 2010
[42] L N Manganas X Zhang Y Li et al ldquoMagnetic resonancespectroscopy identifies neural progenitor cells in the live humanbrainrdquo Science vol 318 no 5852 pp 980ndash985 2007
[43] P Ramm S Couillard-Despres S Plotz et al ldquoA nuclearmagnetic resonance biomarker for neural progenitor cells is itall neurogenesisrdquo Stem Cells vol 27 no 2 pp 420ndash423 2009
[44] M A Rueger H Backes M Walberer et al ldquoNoninvasiveimaging of endogenous neural stem cell mobilization in vivousing positron emission tomographyrdquo Journal of Neurosciencevol 30 no 18 pp 6454ndash6460 2010
[45] J M Encinas A Vaahtokari and G Enikolopov ldquoFluoxetinetargets early progenitor cells in the adult brainrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 103 no 21 pp 8233ndash8238 2006
[46] E Segi-Nishida J L Warner-Schmidt and R S DumanldquoElectroconvulsive seizure and VEGF increase the proliferationof neural stem-like cells in rat hippocampusrdquo Proceedings of theNational Academy of Sciences of the United States of Americavol 105 no 32 pp 11352ndash11357 2008
[47] T Weber V Baier K Lentz et al ldquoGenetic fate mapping oftype-1 stem cell-dependent increase in newborn hippocampalneurons after electroconvulsive seizuresrdquoHippocampus vol 23no 12 pp 1321ndash1330 2013
[48] F Chen T M Madsen G Wegener and J R NyengaardldquoRepeated electroconvulsive seizures increase the total numberof synapses in adult male rat hippocampusrdquo European Neu-ropsychopharmacology vol 19 no 5 pp 329ndash338 2009
[49] O S Jorgensen ldquoSynaptic proteins after electroconvulsivestimulationrdquo Science vol 205 no 4407 pp 705ndash707 1979
[50] L Stanton and R Kohn ldquoDepression and the aging brainrdquoMedicine and Health Rhode Island vol 95 no 7 pp 210ndash2112012
[51] A M Van Schaik H C Comijs C M Sonnenberg A TBeekman P Sienaert and M L Stek ldquoEfficacy and safetyof continuation and maintenance electroconvulsive therapyin depressed elderly patients a systematic reviewrdquo AmericanJournal of Geriatric Psychiatry vol 20 no 1 pp 5ndash17 2012
[52] J-W Wang D J David J E Monckton F Battaglia and RHen ldquoChronic fluoxetine stimulates maturation and synapticplasticity of adult-born hippocampal granule cellsrdquo Journal ofNeuroscience vol 28 no 6 pp 1374ndash1384 2008
[53] S Lange A Trost H Tempfer et al ldquoBrain pericyte plasticityas a potential drug target in CNS repairrdquoDrug Discovery Todayvol 18 no 9-10 pp 456ndash463 2013
[54] J S Goldberg and K K Hirschi ldquoDiverse roles of the vascula-ture within the neural stem cell nicherdquo Regenerative Medicinevol 4 no 6 pp 879ndash897 2009
[55] C Gemma andA D Bachstetter ldquoThe role of microglia in adulthippocampal neurogenesisrdquo Frontiers in Cellular Neurosciencevol 7 article 229 2013
[56] Q Shen S K Goderie L Jin et al ldquoEndothelial cells stimulateself-renewal and expand neurogenesis of neural stem cellsrdquoScience vol 304 no 5675 pp 1338ndash1340 2004
[57] H Song C F Stevens and F H Gage ldquoAstroglia induceneurogenesis from adult neural stem cellsrdquo Nature vol 417 no6884 pp 39ndash44 2002
12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

12 Neural Plasticity
[58] A J Dwork V Arango M Underwood et al ldquoAbsence ofhistological lesions in primate models of ECT and magneticseizure therapyrdquo American Journal of Psychiatry vol 161 no 3pp 576ndash578 2004
[59] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[60] C T Ekdahl Z Kokaia and O Lindvall ldquoBrain inflammationand adult neurogenesis the dual role of microgliardquo Neuro-science vol 158 no 3 pp 1021ndash1029 2009
[61] W J Streit ldquoMicroglial response to brain injury a briefsynopsisrdquo Toxicologic Pathology vol 28 no 1 pp 28ndash30 2000
[62] L Jansson M Wennstrom A Johanson and A TingstromldquoGlial cell activation in response to electroconvulsive seizuresrdquoProgress in Neuro-Psychopharmacology and Biological Psychia-try vol 33 no 7 pp 1119ndash1128 2009
[63] A Schanzer F-P Wachs D Wilhelm et al ldquoDirect stimulationof adult neural stem cells in vitro and neurogenesis in vivo byvascular endothelial growth factorrdquo Brain Pathology vol 14 no3 pp 237ndash248 2004
[64] S A Villeda J Luo K I Mosher et al ldquoThe ageing systemicmilieu negatively regulates neurogenesis and cognitive func-tionrdquo Nature vol 477 no 7362 pp 90ndash96 2011
[65] K E Joynt D JWhellan andCMOrsquoConnor ldquoDepression andcardiovascular disease mechanisms of interactionrdquo BiologicalPsychiatry vol 54 no 3 pp 248ndash261 2003
[66] S Malhotra G E Tesar and K Franco ldquoThe relationshipbetween depression and cardiovascular disordersrdquo CurrentPsychiatry Reports vol 2 no 3 pp 241ndash246 2000
[67] H Takano N Motohashi T Uema et al ldquoChanges in regionalcerebral blood flow during acute electroconvulsive therapyin patients with depression positron emission tomographicstudyrdquo British Journal of Psychiatry vol 190 pp 63ndash68 2007
[68] G Ilbay C Dalcik M Yardimoglu H Dalcik and E DUbeyli ldquoThe blood-brain barrier and epilepsyrdquo in EpilepsymdashHistological Electroencephalographic and Psychological AspectsD D Stevanovic Ed pp 31ndash47 InTech Rijeka Croatia 2012
[69] T G Bolwig M M Hertz and J Holm-Jensen ldquoBlood brainbarrier permeability during electroshock seizures in the ratrdquoEuropean Journal of Clinical Investigation vol 7 no 2 pp 95ndash100 1977
[70] B Oztas M Kaya and S Camurcu ldquoAge related changes inthe effect of electroconvulsive shock on the blood brain barrierpermeability in ratsrdquo Mechanisms of Ageing and Developmentvol 51 no 2 pp 149ndash155 1990
[71] T G BolwigMMHertz andO B Paulson ldquoThe permeabilityof the blood brain barrier during electrically induced seizures inmanrdquo European Journal of Clinical Investigation vol 7 no 2 pp87ndash93 1977
[72] A Sartorius R Hellweg J Litzke et al ldquoCorrelations anddiscrepancies between serum and brain tissue levels of neu-rotrophins after electroconvulsive treatment in ratsrdquo Pharma-copsychiatry vol 42 no 6 pp 270ndash276 2009
[73] B Hattiangady and A K Shetty ldquoAging does not alter thenumber or phenotype of putative stemprogenitor cells in theneurogenic region of the hippocampusrdquo Neurobiology of Agingvol 29 no 1 pp 129ndash147 2008
[74] M Boldrini R Hen M D Underwood et al ldquoHippocampalangiogenesis and progenitor cell proliferation are increasedwith antidepressant use in major depressionrdquo Biological Psychi-atry vol 72 no 7 pp 562ndash571 2012
[75] A Eichmann and J L Thomas ldquoMolecular parallels betweenneural and vascular developmentrdquo Cold Spring Harbor Perspec-tives in Medicine vol 3 no 1 Article ID a006551 2013
[76] J Hellsten M J West A Arvidsson et al ldquoElectroconvulsiveseizures induce angiogenesis in adult rat hippocampusrdquo Biolog-ical Psychiatry vol 58 no 11 pp 871ndash878 2005
[77] S S Newton M J Girgenti E F Collier and R S DumanldquoElectroconvulsive seizure increases adult hippocampal angio-genesis in ratsrdquo European Journal of Neuroscience vol 24 no 3pp 819ndash828 2006
[78] B S McEwen ldquoStress and hippocampal plasticityrdquo AnnualReview of Neuroscience vol 22 pp 105ndash122 1999
[79] B S McEwen D Albeck H Cameron et al ldquoStress and thebrain a paradoxical role for adrenal steroidsrdquo Vitamins andHormones vol 51 pp 371ndash402 1995
[80] B A Samuels and R Hen ldquoNeurogenesis and affective disor-dersrdquo European Journal of Neuroscience vol 33 no 6 pp 1152ndash1159 2011
[81] P Hu C Oomen A-M van Dam et al ldquoA single-day treatmentwith mifepristone is sufficient to normalize chronic glucocor-ticoid induced suppression of hippocampal cell proliferationrdquoPLoS ONE vol 7 no 9 Article ID e46224 2012
[82] N R Nichols M Zieba and N Bye ldquoDo glucocorticoidscontribute to brain agingrdquo Brain Research Reviews vol 37 no1ndash3 pp 273ndash286 2001
[83] C Mirescu and E Gould ldquoStress and adult neurogenesisrdquoHippocampus vol 16 no 3 pp 233ndash238 2006
[84] A Garcia B Steiner G Kronenberg A Bick-Sander and GKempermann ldquoAge-dependent expression of glucocorticoid-andmineralocorticoid receptors on neural precursor cell popu-lations in the adult murine hippocampusrdquoAging Cell vol 3 no6 pp 363ndash371 2004
[85] A Surget A Tanti E D Leonardo et al ldquoAntidepressantsrecruit new neurons to improve stress response regulationrdquoMolecular Psychiatry vol 16 no 12 pp 1177ndash1188 2011
[86] P J Lucassen C P Fitzsimons A Korosi M Joels C Belzungand D N Abrous ldquoStressing new neurons into depressionrdquoMolecular Psychiatry vol 18 no 4 pp 396ndash397 2013
[87] S P Jung B Siegrist M R Wade C T Anthony and E AWoltering ldquoInhibition of human angiogenesis with heparin andhydrocortisonerdquo Angiogenesis vol 4 no 3 pp 175ndash186 2001
[88] X Yang E R Ewald Y Huo et al ldquoGlucocorticoid-inducedloss of DNA methylation in non-neuronal cells and potentialinvolvement of DNMT1 in epigenetic regulation of Fkbp5rdquoBiochemical andBiophysical ResearchCommunications vol 420no 3 pp 570ndash575 2012
[89] J Hellsten M Wennstrom P Mohapel C T Ekdahl J Beng-zon and A Tingstrom ldquoElectroconvulsive seizures increasehippocampal neurogenesis after chronic corticosterone treat-mentrdquo European Journal of Neuroscience vol 16 no 2 pp 283ndash290 2002
[90] J E Smith KWilliams S BurkettHNicholson PGlue andDJNutt ldquoOxytocin and vasopressin responses to ECTrdquoPsychiatryResearch vol 32 no 2 pp 201ndash202 1990
[91] R Ohman J Balldin and L Abrahamsson ldquoProlactin responseto electroconvulsive therapyrdquo The Lancet vol 2 no 7992 pp936ndash937 1976
[92] J P Allen D Denney J W Kendall and P H BlachlyldquoCorticotropin release during ECT in manrdquo American Journalof Psychiatry vol 131 no 11 pp 1225ndash1228 1974
Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

Neural Plasticity 13
[93] S Soelberg P Sorensen M Hammer and T G BolwigldquoVasopressin release during electroconvulsive therapyrdquo Psy-choneuroendocrinology vol 7 no 4 pp 303ndash308 1982
[94] J P Redrobe Y Dumont A Fournier and R Quirion ldquoTheneuropeptide Y (NPY) Y1 receptor subtype mediates NPY-induced antidepressant-like activity in the mouse forced swim-ming testrdquo Neuropsychopharmacology vol 26 no 5 pp 615ndash624 2002
[95] A A Mathe ldquoNeuropeptides and electroconvulsive treatmentrdquoJournal of ECT vol 15 no 1 pp 60ndash75 1999
[96] G Nikisch and A A Mathe ldquoCSF monoamine metabolites andneuropeptides in depressed patients before and after electro-convulsive therapyrdquo European Psychiatry vol 23 no 5 pp 356ndash359 2008
[97] M Heilig O Zachrisson A Thorsell et al ldquoDecreasedcerebrospinal fluid neuropeptide Y (NPY) in patients withtreatment refractory unipolar major depression preliminaryevidence for association with preproNPY gene polymorphismrdquoJournal of Psychiatric Research vol 38 no 2 pp 113ndash121 2004
[98] M Nibuya S Morinobu and R S Duman ldquoRegulation ofBDNFand trkBmRNA in rat brain by chronic electroconvulsiveseizure and antidepressant drug treatmentsrdquo Journal of Neuro-science vol 15 no 11 pp 7539ndash7547 1995
[99] S S Newton E F Collier J Hunsberger et al ldquoGene profileof electroconvulsive seizures induction of neurotrophic andangiogenic factorsrdquo Journal of Neuroscience vol 23 no 34 pp10841ndash10851 2003
[100] C A Altar P Laeng L W Jurata et al ldquoElectroconvulsiveseizures regulate gene expression of distinct neurotrophic sig-naling pathwaysrdquo Journal of Neuroscience vol 24 no 11 pp2667ndash2677 2004
[101] J Hellsten M Wennstrom J Bengzon P Mohapel and ATingstrom ldquoElectroconvulsive seizures induce endothelial cellproliferation in adult rat hippocampusrdquo Biological Psychiatryvol 55 no 4 pp 420ndash427 2004
[102] M Wennstrom J Hellsten and A Tingstrom ldquoElectroconvul-sive seizures induce proliferation of NG2-expressing glial cellsin adult rat amygdalardquo Biological Psychiatry vol 55 no 5 pp464ndash471 2004
[103] G S Naeve M Ramakrishnan R Kramer D Hevroni Y Citriand L ETheill ldquoNeuritin a gene induced by neural activity andneurotrophins that promotes neuritogenesisrdquo Proceedings of theNational Academy of Sciences of theUnited States of America vol94 no 6 pp 2648ndash2653 1997
[104] D J Bonthius B Karacay D Dai and N J Pantazis ldquoFGF-2NGF and IGF-1 but not BDNF utilize a nitric oxide pathway tosignal neurotrophic and neuroprotective effects against alcoholtoxicity in cerebellar granule cell culturesrdquoDevelopmental BrainResearch vol 140 no 1 pp 15ndash28 2003
[105] J L Warner-Schmidt and R S Duman ldquoVEGF is an essentialmediator of the neurogenic and behavioral actions of antide-pressantsrdquoProceedings of the National Academy of Sciences of theUnited States of America vol 104 no 11 pp 4647ndash4652 2007
[106] K Jin Y Zhu Y Sun X O Mao L Xie and D A Green-berg ldquoVascular endothelial growth factor (VEGF) stimulatesneurogenesis in vitro and in vivordquo Proceedings of the NationalAcademy of Sciences of the United States of America vol 99 no18 pp 11946ndash11950 2002
[107] T G Bolwig ldquoHow does electroconvulsive therapy workTheories on itsmechanismrdquoCanadian Journal of Psychiatry vol56 no 1 pp 13ndash18 2011
[108] Y Shirayama A C-H Chen S Nakagawa D S Russell andR S Duman ldquoBrain-derived neurotrophic factor producesantidepressant effects in behavioral models of depressionrdquoJournal of Neuroscience vol 22 no 8 pp 3251ndash3261 2002
[109] J A Siuciak D R Lewis S J Wiegand and R M LindsayldquoAntidepressant-like effect of brain-derived neurotrophic factor(BDNF)rdquoPharmacology Biochemistry and Behavior vol 56 no1 pp 131ndash137 1997
[110] E Segi-Nishida ldquoExploration of newmolecularmechanisms forantidepressant actions of electroconvulsive seizurerdquo Biologicaland Pharmaceutical Bulletin vol 34 no 7 pp 939ndash944 2011
[111] M Sairanen G Lucas P Ernfors M Castren and E CastrenldquoBrain-derived neurotrophic factor and antidepressant drugshave different but coordinated effects on neuronal turnoverproliferation and survival in the adult dentate gyrusrdquo Journalof Neuroscience vol 25 no 5 pp 1089ndash1094 2005
[112] T Saarelainen P Hendolin G Lucas et al ldquoActivation of theTrkB neurotrophin receptor is induced by antidepressant drugsand is required for antidepressant-induced behavioral effectsrdquoJournal of Neuroscience vol 23 no 1 pp 349ndash357 2003
[113] O Kampman and E Leinonen ldquoEfficacy of electroconvulsivetherapy is it in the BDNF generdquo Pharmacogenomics vol 14no 12 pp 1365ndash1368 2013
[114] P Pruunsild A Kazantseval T Aid K Palm and T TimmuskldquoDissecting the human BDNF locus bidirectional transcrip-tion complex splicing and multiple promotersrdquo Genomics vol90 no 3 pp 397ndash406 2007
[115] D Suri V Veenit A Sarkar et al ldquoEarly stress evokes age-dependent biphasic changes in hippocampal neurogenesisBdnf expression and cognitionrdquo Biological Psychiatry vol 73no 7 pp 658ndash666 2013
[116] H Kaija A Sami H Martti et al ldquoBrain-derived neurotrophicfactor (BDNF) polymorphisms G196A and C270T are notassociated with response to electroconvulsive therapy in majordepressive disorderrdquo European Archives of Psychiatry and Clini-cal Neuroscience vol 257 no 1 pp 31ndash35 2007
[117] S Campbell M Marriott C Nahmias and G M MacQueenldquoLower hippocampal volume in patients suffering from depres-sion a meta-analysisrdquo American Journal of Psychiatry vol 161no 4 pp 598ndash607 2004
[118] R S Duman G R Heninger and E J Nestler ldquoAmolecular andcellular theory of depressionrdquo Archives of General Psychiatryvol 54 no 7 pp 597ndash606 1997
[119] E J Nestler M Barrot R J DiLeone A J Eisch S J Gold andLMMonteggia ldquoNeurobiology of depressionrdquoNeuron vol 34no 1 pp 13ndash25 2002
[120] Y I Sheline M H Gado and H C Kraemer ldquoUntreateddepression and hippocampal volume lossrdquo American Journal ofPsychiatry vol 160 no 8 pp 1516ndash1518 2003
[121] P Videbech and B Ravnkilde ldquoHippocampal volume anddepression a meta-analysis of MRI studiesrdquo American Journalof Psychiatry vol 161 no 11 pp 1957ndash1966 2004
[122] D Cotter D Mackay S Landau R Kerwin and I EverallldquoReduced glial cell density and neuronal size in the anteriorcingulate cortex in major depressive disorderrdquo Archives ofGeneral Psychiatry vol 58 no 6 pp 545ndash553 2001
[123] G Rajkowska J J Miguel-Hidalgo JWei et al ldquoMorphometricevidence for neuronal and glial prefrontal cell pathology inmajor depressionrdquo Biological Psychiatry vol 45 no 9 pp 1085ndash1098 1999
14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013

14 Neural Plasticity
[124] D Ongur W C Drevets and J L Price ldquoGlial reduction in thesubgenual prefrontal cortex in mood disordersrdquo Proceedings ofthe National Academy of Sciences of the United States of Americavol 95 no 22 pp 13290ndash13295 1998
[125] I Tendolkar M van Beek I van Oostrom et al ldquoElectrocon-vulsive therapy increases hippocampal and amygdala volumein therapy refractory depression a longitudinal pilot studyrdquoPsychiatry Research vol 214 no 3 pp 197ndash203 2013
[126] M G Cole F Bellavance and A Mansour ldquoPrognosis ofdepression in elderly community and primary care populationsa systematic review and meta-analysisrdquo American Journal ofPsychiatry vol 156 no 8 pp 1182ndash1189 1999
[127] J M Parent and G G Murphy ldquoMechanisms and functionalsignificance of aberrant seizure-induced hippocampal neuroge-nesisrdquo Epilepsia vol 49 supplement 5 pp 19ndash25 2008
[128] S R Lamont A Paulls and C A Stewart ldquoRepeated electro-convulsive stimulation but not antidepressant drugs inducesmossy fibre sprouting in the rat hippocampusrdquo Brain Researchvol 893 no 1-2 pp 53ndash58 2001
[129] V A Vaidya J A Siuciak F Du and R S Duman ldquoHippocam-pal mossy fiber sprouting induced by chronic electroconvulsiveseizuresrdquo Neuroscience vol 89 no 1 pp 157ndash166 1999
[130] J M Parent and D H Lowenstein ldquoSeizure-induced neuroge-nesis are more new neurons good for an adult brainrdquo Progressin Brain Research vol 135 pp 121ndash131 2002
[131] C Schmidt-Hieber P Jones and J Bischofberger ldquoEnhancedsynaptic plasticity in newly generated granule cells of the adulthippocampusrdquo Nature vol 429 no 6988 pp 184ndash187 2004
[132] S Ge C-H Yang K-S Hsu G-L Ming and H SongldquoA critical period for enhanced synaptic plasticity in newlygenerated neurons of the adult brainrdquoNeuron vol 54 no 4 pp559ndash566 2007
[133] S Couillard-Despres B Winner S Schaubeck et al ldquoDou-blecortin expression levels in adult brain reflect neurogenesisrdquoEuropean Journal of Neuroscience vol 21 no 1 pp 1ndash14 2005
[134] V M Heine J Zareno S Maslam M Joels and P J LucassenldquoChronic stress in the adult dentate gyrus reduces cell pro-liferation near the vasculature and VEGF and Flk-1 proteinexpressionrdquo European Journal of Neuroscience vol 21 no 5 pp1304ndash1314 2005
[135] A H Nagahara M Mateling I Kovacs et al ldquoEarly BDNFtreatment ameliorates cell loss in the entorhinal cortex of APPtransgenic micerdquoThe Journal of Neuroscience vol 33 no 39 pp15596ndash15602 2013