helicobacter pylori disulphide reductases: role in metronidazole reduction
TRANSCRIPT

www.fems-microbiology.org
FEMS Immunology and Medical Microbiology 44 (2005) 137–142
Helicobacter pylori disulphide reductases: role inmetronidazole reduction
Nadeem O. Kaakoush, George L. Mendz *
School of Biotechnology and Biomolecular Sciences, The University of New South Wales, Biological Science Builiding, Sydney, NSW 2052, Australia
Received 26 June 2004; received in revised form 27 October 2004; accepted 22 November 2004
First published online 2 December 2004
Abstract
Disulphide reductases play an important role in maintaining intracellular redox potential. Three disulphide reductase activities
were identified in Helicobacter pylori, which used dithiobis-2-nitrobenzoic acid, glutathione or LL-cystine and ferredoxin as sub-
strates. The kinetic parameters of these activities were determined and it was demonstrated that the reductase activities were inhib-
ited by the presence of metronidazole. Substrate competition experiments served to show inhibition of metronidazole reduction by
dithiobis-2-nitrobenzoic acid, glutathione and ferredoxin in lysates from metronidazole susceptible and resistant matched pairs of
strains. The study demonstrated that the activities of three disulphide reductases were modulated by the presence of metronidazole,
and that metronidazole reduction was inhibited by the presence of disulphide reductase substrates.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Helicobacter pylori; Disulphide reductases; Metronidazole; Resistance; Redox potential
1. Introduction
Metronidazole (Mtr) is an important component oftherapeutic regimes currently used to treat many micro-
bial pathogens. This 5-nitroimidazole is activated via
interactions with redox systems capable of reducing
the low potential (�415 mV) nitro group in position 5
of the imidazole ring [1]. This property makes metroni-
dazole effective against organisms of low intracellular re-
dox state, such as anaerobic bacteria and protozoa, as
well as some microaerophiles, such as Campylobacter
spp. and Helicobacter pylori [2].
The frequent use of metronidazole has resulted in in-
creased resistance to the antibiotic by many pathogens
including H. pylori. The emergence of resistant isolates
that do not respond to the drug fostered a keen interest
0928-8244/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsim.2004.11.007
* Corresponding author. Tel.: +61 2 9385 2042; fax: +61 2 9385
1483.
E-mail address: [email protected] (G.L. Mendz).
to understand the primary causes of resistance to metro-
nidazole in this bacterium. Extensive investigations on
H. pylori established that main causes of metronidazoleresistance are mutations in the genes rdxA or frxA [3–6].
However, insufficient data correlating RdxA and/or
FrxA with the resistant phenotype, and the fact that a
small percentage of resistant strains do not have muta-
tions in either rdxA or frxA indicated that the molecular
basis of H. pylori resistance to Mtr has not been charac-
terised completely.
Early studies showed that oxygen tensions have alarge impact on the resistance of H. pylori to Mtr [7–
10], and several investigations have linked the activities
of specific oxidoreductases to the Mtr-susceptible phe-
notype of the bacterium [10–12]. These results suggested
a possible role in Mtr activation of enzymes catalysing
redox reactions which modulate the intracellular redox
status. A type of such enzymes are disulphide reductases
whose reactions contribute to the redox balance of thecell.
. Published by Elsevier B.V. All rights reserved.

138 N.O. Kaakoush, G.L. Mendz / FEMS Immunology and Medical Microbiology 44 (2005) 137–142
In the present study, ferredoxin (Fdx) oxidoreductase
and two other enzyme activities which use oxidised glu-
tathione (GSSG), LL-cystine (Cys-Cys) and dithiobis-2-
nitrobenzoic acid (DTNB) as substrates were identified
in H. pylori, and the effects of metronidazole on their
activities were characterised. The potential involvementof these disulphide reductases in metronidazole activa-
tion was investigated by measuring the effects of their
substrates on the rates of metronidazole reduction.
2. Materials and methods
2.1. Chemicals and reagents
Blood agar base No. 2, brain heart infusion (BHI),
defibrinated horse blood and horse serum were from
Oxoid (Heidelberg West, VIC, Australia). Amphotericin
(Fungizone�), bicinchoninic acid, bovine serum albu-
min, copper II sulphate, LL-cystine, DTNB, Fdx, GSSG,
Cys-Cys, metronidazole, mineral oil, b-nicotinamide
adenine dinucleotide reduced form (NADH), polymixinB and trimethoprim were from Sigma (Castle Hill,
NSW, Australia). Vancomycin was from Eli Lilly
(North Ryde, NSW, Australia). Deuterium oxide was
from Cambridge isotope laboratories (Cambridge, Eng-
land). Tris base was from Amersham Biosciences (Mel-
bourne, VIC, Australia). All other reagents were of
analytical grade.
2.2. Bacterial cultures and preparation
The strains used in this study were J99 (with anno-
tated genome), NCTC 11639, N6 (Institut Pasteur,
Paris), and P10 and SS1 (University of New South
Wales collection). The isolates LC11 and LC20 were ob-
tained recently from patients with gastritis, and the Mtr
susceptible and resistant matched pairs 10593/2 andRIG 117 were obtained before and after therapy from
patients treated unsuccessfully with metronidazole.
The SS1 resistant strains were constructed by sequential
passing on plates containing increasing concentrations
of Mtr. In the clinical isolates as well as in the SS1 sus-
ceptible and resistant strains, the matched pairs have the
same genetic background [13]. Bacteria were grown on
Campylobacter selective agar supplemented with defi-brinated horse blood, 2.0 lg ml�1 Amphotericin, 5.0
mg ml�1 vancomycin, 1250 U ml�1 polymixin B, and
2.5 mg ml�1 trimethoprim. Cultures were incubated at
37 �C under the microaerobic conditions 5% CO2, 5%
O2 and 90% N2. The purity of the cultures was con-
firmed as H. pylori by positive urease and catalase tests,
and motility and morphology observed under phase
contrast microscopy.H. pylori cells were harvested in 150 mM sodium
chloride (NaCl) and centrifuged at 16,000g at 4 �C for
10 min. The pellet was collected and the supernatant dis-
carded. The pellet was resuspended in 150 mM NaCl
solution and washed three times. Cells were lysed by
thrice freezing in liquid nitrogen and thawing. Cell-free
extracts were obtained by centrifuging lysates at
16,000g at 4 �C for 20 min and collecting the solublefraction.
Protein concentrations were estimated by the bicinch-
oninic acid method based on microtitre plate protocol
[12].
2.3. Nuclear magnetic resonance spectroscopy
Suspensions of bacterial lysates or cell-free extractswere placed in 5 or 10 mm tubes (Wilmad, Buena, NJ,
USA), the appropriate substrates added, and measure-
ments of enzyme activities were carried out at 37 �C.Proton nuclear magnetic resonance spectroscopy (1H
NMR) and nitrogen-14 nuclear magnetic resonance
(14N NMR) free induction decays were collected using
a Bruker DMX-600 or a Bruker DMX-500 spectrome-
ter, respectively, operating in the pulsed Fourier trans-form mode with quadrature detection. The
instrumental parameters for the DMX-600 spectrometer
were: operating frequency 600.13 MHz, spectral width
6009.61 Hz, memory size 16 K, acquisition time 1.36 s,
number of transients 64, pulse angle 50� (3 ls) and relax-
ation delay with solvent presaturation 1.7 s. Spectral res-
olution was enhanced by Gaussian multiplication with
line broadening of �0.7 Hz and Gaussian broadeningfactor of 0.19. Proton spectra were acquired with presat-
uration of the water resonance. The instrumental
parameters for the DMX-500 spectrometer were: oper-
ating frequency 36.14 MHz, spectral width 19,841 Hz,
memory size 8 K, acquisition time 0.21 s, number of
transients 2488, pulse angle 90� (30 ls) and relaxation
delay 0.1 s. Spectra were acquired with composite pulse
decoupling of protons. Exponential filtering of 15 Hzwas applied prior to Fourier transformation.
The time-evolution of substrates and products were
followed by acquiring sequential spectra of the reac-
tions. Progress curves were obtained by measuring the
integrals of compounds at different points in time. Max-
imal rates were calculated from good fits to straight lines
(correlation coefficientsP 0.95) of the data for 30 min
for GSSG reduction or 2 h for Mtr reduction. Calibra-tions of the peaks arising from substrates were per-
formed by extrapolating the resonance intensity data
to zero time and assigning to this intensity the appropri-
ate concentration value.
Reduction rates of GSSG or Cys-Cys were measured
employing 1H NMR spectroscopy in lysates or cell-free
extracts suspended in 2H2O:H2O (1:5 v/v), 10 mM KCl,
25 mM NaCl and 50 mM potassium phosphate buffer,pH 7.2. Metronidazole reduction was measured employ-
ing 14N NMR spectroscopy in lysate suspensions in the
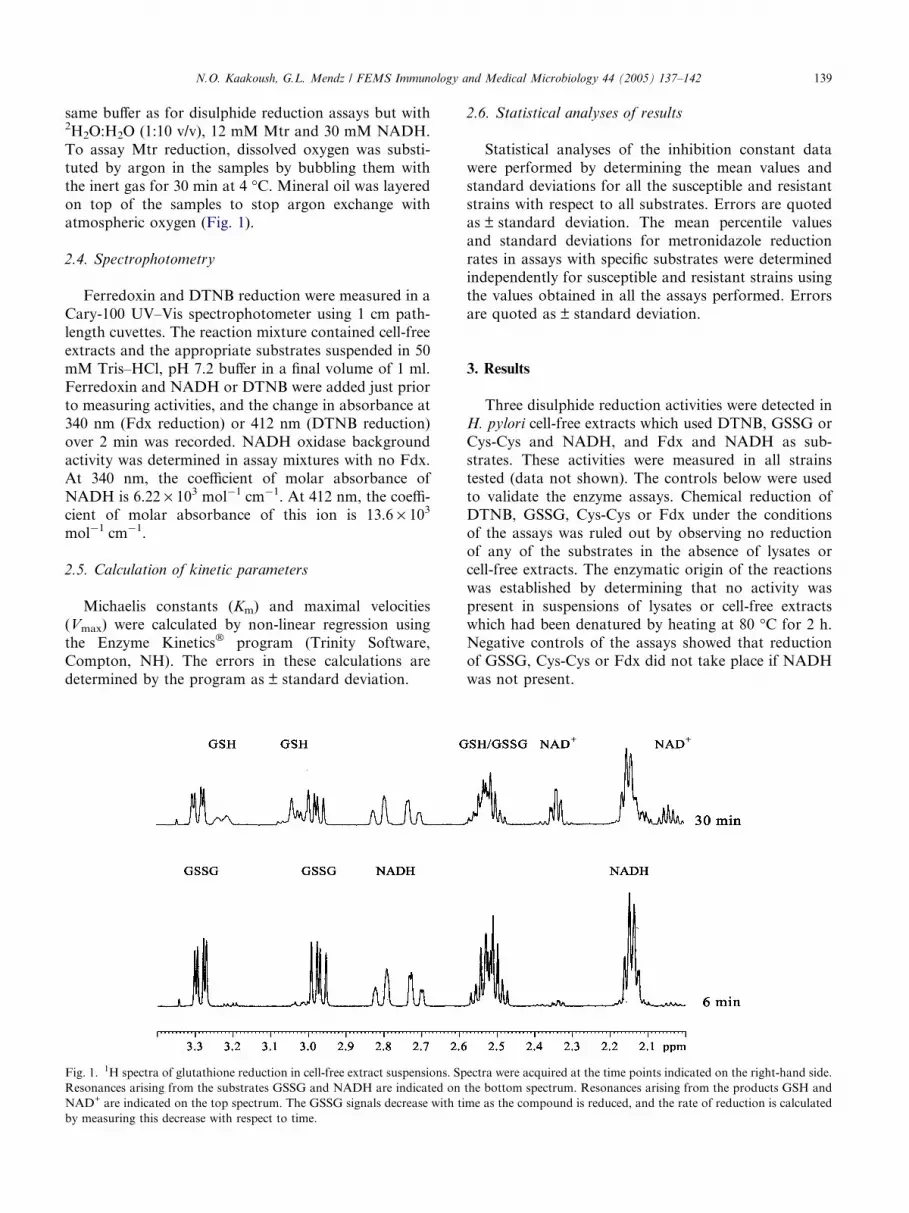
N.O. Kaakoush, G.L. Mendz / FEMS Immunology and Medical Microbiology 44 (2005) 137–142 139
same buffer as for disulphide reduction assays but with2H2O:H2O (1:10 v/v), 12 mM Mtr and 30 mM NADH.
To assay Mtr reduction, dissolved oxygen was substi-
tuted by argon in the samples by bubbling them with
the inert gas for 30 min at 4 �C. Mineral oil was layered
on top of the samples to stop argon exchange withatmospheric oxygen (Fig. 1).
2.4. Spectrophotometry
Ferredoxin and DTNB reduction were measured in a
Cary-100 UV–Vis spectrophotometer using 1 cm path-
length cuvettes. The reaction mixture contained cell-free
extracts and the appropriate substrates suspended in 50mM Tris–HCl, pH 7.2 buffer in a final volume of 1 ml.
Ferredoxin and NADH or DTNB were added just prior
to measuring activities, and the change in absorbance at
340 nm (Fdx reduction) or 412 nm (DTNB reduction)
over 2 min was recorded. NADH oxidase background
activity was determined in assay mixtures with no Fdx.
At 340 nm, the coefficient of molar absorbance of
NADH is 6.22 · 103 mol�1 cm�1. At 412 nm, the coeffi-cient of molar absorbance of this ion is 13.6 · 103
mol�1 cm�1.
2.5. Calculation of kinetic parameters
Michaelis constants (Km) and maximal velocities
(Vmax) were calculated by non-linear regression using
the Enzyme Kinetics� program (Trinity Software,Compton, NH). The errors in these calculations are
determined by the program as ± standard deviation.
Fig. 1. 1H spectra of glutathione reduction in cell-free extract suspensions. Sp
Resonances arising from the substrates GSSG and NADH are indicated on
NAD+ are indicated on the top spectrum. The GSSG signals decrease with ti
by measuring this decrease with respect to time.
2.6. Statistical analyses of results
Statistical analyses of the inhibition constant data
were performed by determining the mean values and
standard deviations for all the susceptible and resistant
strains with respect to all substrates. Errors are quotedas ± standard deviation. The mean percentile values
and standard deviations for metronidazole reduction
rates in assays with specific substrates were determined
independently for susceptible and resistant strains using
the values obtained in all the assays performed. Errors
are quoted as ± standard deviation.
3. Results
Three disulphide reduction activities were detected in
H. pylori cell-free extracts which used DTNB, GSSG or
Cys-Cys and NADH, and Fdx and NADH as sub-
strates. These activities were measured in all strains
tested (data not shown). The controls below were used
to validate the enzyme assays. Chemical reduction ofDTNB, GSSG, Cys-Cys or Fdx under the conditions
of the assays was ruled out by observing no reduction
of any of the substrates in the absence of lysates or
cell-free extracts. The enzymatic origin of the reactions
was established by determining that no activity was
present in suspensions of lysates or cell-free extracts
which had been denatured by heating at 80 �C for 2 h.
Negative controls of the assays showed that reductionof GSSG, Cys-Cys or Fdx did not take place if NADH
was not present.
ectra were acquired at the time points indicated on the right-hand side.
the bottom spectrum. Resonances arising from the products GSH and
me as the compound is reduced, and the rate of reduction is calculated

140 N.O. Kaakoush, G.L. Mendz / FEMS Immunology and Medical Microbiology 44 (2005) 137–142
Matched pairs of Mtr susceptible and resistant strains
were employed to investigate the relationships between
disulphide reduction and Mtr reduction. The kinetic
parameters of DTNB, GSSG and Fdx reduction for
the 10593/2 matched pairs of isolates are given in Table
1. The Km and Vmax of DTNB reduction in the resistantstrain were smaller than in the susceptible strain. No sig-
nificant differences were observed in the kinetic parame-
ters of GSSG reduction for the pair of susceptible and
resistant isolates. Ferredoxin reduction was observed
in the susceptible isolate but was absent in the resistant
one (Table 1). In the other matched pairs of isolates,
Fdx reduction was observed in the resistant strain but
the Km of Fdx reduction in the resistant isolates weresignificantly smaller than in the susceptible
counterparts.
The effects of Mtr on disulphide reduction were inves-
tigated by measuring the rates of reduction in the pres-
ence of different concentrations of Mtr. The three
disulphide reduction activities were inhibited by Mtr.
The mode of inhibition was determined by measuring
the kinetic parameters of the reductions with and with-out 0.5 mM Mtr. In the presence of Mtr, larger Km
and similar Vmax values were measured, indicating that
the inhibition of these activities by Mtr was competitive.
At concentrations well below the Km of the sub-
strates, the inhibition constant (Ki) can be calculated
from the expression
v0=v ¼ 1þ I=K i;
where �v0� and �v� are the uninhibited and inhibited rates
of enzyme activity, respectively, and �I� is the concentra-tion of inhibitor which results in a �v� rate of reaction
[14]. The Ki values for the inhibition of GSSG and
Cys-Cys reduction activities of the matched pairs are gi-
ven in Table 2. Larger Ki values were observed in resis-tant strains than in their susceptible counterparts,
suggesting stronger effects of Mtr on the latter. Similar
observations were made for the other matched pairs of
strains. Statistical analyses of these results were per-
formed and the Ki mean value of the susceptible strains
for both substrates, 1.2 ± 0.3 mM, was significantly
Table 1
Disulphide reduction activities in H. pylori 10593/2 Mtr-susceptible and resi
Isolate Substrate
10593/2 Susceptible DTNB
GSSG:NADH
Fdx:NADH
10593/2 Resistant DTNB
GSSG:NADH
Fdx:NADH
DTNB concentrations ranged from 10 lM to 1 mM; GSSG concentration
NADH concentrations were 0.5 mM for DTNB assays, 50 mM for GSSG ass
13 rates. Errors were calculated using the Enzyme Kinetics program from n
lower than the mean value for the resistant strains,
3.8 ± 1.1 mM.
Metronidazole reduction was measured in H. pylori
lysate suspensions employing 14N NMR spectroscopy.
The rate of Mtr reduction in the 10593/2 Mtr-resistant
isolate was significantly lower (p < 0.02) than in the sus-ceptible isolate (Fig. 2), correlating with the phenotype
of the isolates. Similar results were obtained for the
RIG 117 and SS1 matched pairs of strains (data not
shown). The potential roles of disulphide reductases in
Mtr reduction were investigated by performing substrate
competition experiments. Metronidazole reduction rates
for the susceptible strains were inhibited in the presence
of the substrates DTNB (p < 0.03), GSSG (p < 0.02) orFdx (p < 0.02). The reduction rates for the resistant
strains were also inhibited in the presence of the sub-
strates DTNB (p < 0.03), GSSG (p < 0.03) or Fdx
(p < 0.03). The results for the 10593/2 matched pair of
isolates are shown in Fig. 2. Similar data were obtained
for Mtr-Fdx competition experiments for the other
matched pairs of strains.
4. Discussion
H. pylori is one of the most prevalent bacterial infec-
tions in humans. The bacterium is susceptible to several
antibiotics including metronidazole, but it readily devel-
ops resistance in vivo and in vitro on exposure to the
drug. The prevalence of Mtr-resistant isolates varies be-tween 10% and 80% for different parts of the world [15].
It was discovered that Mtr resistance in H. pylori can
result from the loss of activity of RdxA, an oxygen
insensitive NADPH nitroreductase. This finding pro-
voked renewed interest to achieve a full understanding
of the causes of resistance to this drug [4]. Later, the
finding of frameshift mutations in the gene frxA in resis-
tant strains suggested a role for the NAD(P)H flavinoxidoreductase FrxA, in Mtr resistance in H. pylori
[6,16]. Investigations of matched susceptible and resis-
tant pairs showed the existence of unchanged rdxA in
susceptible and resistant isolates, and of different rdxA
stant cells
Km Vmax (nmol mg�1 min�1)
45 ± 3 lM 17 ± 2
2.3 ± 0.2 mM 129 ± 4
3.0 ± 0.4 lg ml�1 10 ± 1
17 ± 2 lM 12 ± 1
2.8 ± 0.2 mM 166 ± 2
No reduction No reduction
s from 0.5 to 70 mM; and Fdx concentrations from 0 to 80 lg ml�1.
ays, and 0.2 mM for Fdx assays. Kinetic fits were performed using 10–
on-linear regression fits to the data.

Table 2
Metronidazole inhibition constants (Ki) of disulphide reductase
activities for Mtr-susceptible and Mtr-resistant strains
Strain Ki (mM)
Substrates Susceptible Resistant
SS1 GSSG 1.1 ± 0.2 5.5 ± 0.8
10593/2 GSSG 1.6 ± 0.1 2.8 ± 0.3
Cys-Cys 1.3 ± 0.2 3.9 ± 0.5
10827/6 Cys-Cys 0.7 ± 0.1 2.7 ± 0.4
RIG117 Cys-Cys 1.2 ± 0.2 4.1 ± 0.6
Enzyme activities were measured in lysates suspended in potassium
phosphate or TrisHCl buffer for the GSSG and Fdx assays, respec-
tively. Five rates were used for each inhibition plot. Errors were
determined from the best-fitted line in the inhibition plot.
Fig. 2. Metronidazole reduction activities of H. pylori 10593/2
susceptible and resistant lysates from cells grown under microaerobic
conditions. Lysates were suspended in phosphate buffer and subjected
to argon treatment for 30 min. Initial substrate concentrations were 12
mM Mtr and 30 mM NADH. DTNB, GSSG and Fdx were added in
concentrations of 15 lM, 4 mM and 0.17 mg ml�1 for the susceptible
lysates and 30 lM, 6 mM and 0.17 mg ml�1 for the resistant lysates.
Errors were calculated from the straight line fitting of the values used
to determine the Mtr reduction rates.
N.O. Kaakoush, G.L. Mendz / FEMS Immunology and Medical Microbiology 44 (2005) 137–142 141
in susceptible strains [5,13,16–21]. Similarly, it was dem-
onstrated that unchanged frxA are found in susceptible
and resistant isolates [20–23]. Thus, both RdxA and
FrxA are involved in resistance to Mtr by H. pylori,but the available data suggest that there are other fac-
tors contributing to the resistant phenotype. For exam-
ple, other enzymes capable of reducing metronidazole,
mechanisms which modulate the expression of rdxA
and/or frxA, etc.
Several studies on H. pylori demonstrated that the
intracellular oxygen status and redox potential play a
role in metronidazole resistance [7–10]. The observa-tions that resistance to Mtr can be overcome by expos-
ing H. pylori to short periods on anaerobiosis [7,8],
and that susceptible strains have greater cytosolic
NADH-oxidase activities that their respective resistant
mutants [7,9], suggested that the oxygen status of the cell
is a major contributor to susceptibility of H. pylori the
Mtr. Also, changes in the activities pyruvate oxidore-
ductase, a-ketoglutarate oxidoreductase, and pyru-
vate:flavodoxin oxidoreductase were observed in theresistant phenotype [10], implicating oxidoreductases
in the resistance of H. pylori to Mtr.
Disulphide reductases catalyse oxireduction reactions
which contribute to the redox balance of the cell. They
have been shown to be virulence and invasive factors,
and involved in drug resistance in different pathogenic
bacteria [24–26]. A study with matched pairs ofH. pylori
Mtr-susceptible strains and Mtr-resistant mutants dem-onstrated that susceptible strains have higher levels of
disulphide reduction, and that the total disulphide
reduction activity of the cell is modulated by Mtr [10].
Evidence for a role of disulphide reductases in the sus-
ceptibility of H. pylori to Mtr was provided by the find-
ing that the alkyl hydroperoxide reductase activity of
Mtr-susceptible strains was absent in their Mtr-resistant
counterparts [12,27]. Thus, it became important toinvestigate the role of specific disulphide reduction activ-
ities in H. pylori resistance to Mtr.
Three disulphide reduction activities which use
DTNB, GSSG or Cys-Cys and NADH, or Fdx and
NADH as substrates, were identified and characterised
in H. pylori (Table 1). Metronidazole inhibited disul-
phide reduction competitively in each of the three activ-
ities, and the measured Ki of Mtr for the reduction ofGSSG or Cys-Cys indicated that the effects of Mtr were
stronger in susceptible strains than in resistant ones (Ta-
ble 2). The data demonstrated that Mtr modulated di-
rectly the disulphide reductases and suggested a role
for these reductases in Mtr reduction.
Confirmation of this role was found by examining the
involvement of disulphide reductases in Mtr reduction.
The presence of the substrates DTNB, GSSG or Fdx,inhibited Mtr reduction in situ, indicating that these
disulphide reductases competed with Mtr as acceptors
in redox reactions, and suggesting that they participated
in the reduction of Mtr. Together with previous findings
these results provided evidence that disulphide reduc-
tases play a role in the activation of Mtr, and thus, in
the susceptibility of H. pylori to this antibiotic.
The role of these reductase activities in the resistantphenotype needs to be investigated further, as well as
putative molecular mechanisms relating these disulphide
reductases to the activities of RdxA and FrxA, in partic-
ular the effects of the intracellular redox potential on the
expression of the latter two enzymes. Metronidazole
resistance is found in other pathogenic microaerophilic
organisms besides H. pylori. It will be important to
ascertain whether disulphide reductase activities alsocontribute to the development of Mtr resistance in, for
instance, Campylobacter spp. and various parasites.

142 N.O. Kaakoush, G.L. Mendz / FEMS Immunology and Medical Microbiology 44 (2005) 137–142
Acknowledgement
This work was made possible by the support of the
Australian Research Council.
References
[1] Scarpignato, C. (2004) Towards the ideal regimen for Helicobac-
ter pylori eradication: the search continues. Dig. Liver Dis. 36,
243–247.
[2] Land, K.M. and Johnson, P.J. (1999) Molecular basis of
metronidazole resistance in pathogenic bacteria and protozoa.
Drug Resist. Updat. 2, 289–294.
[3] Debets-Ossenkopp, Y.J., Pot, R.G.J., Van Westerloo, D.J.,
Goodwin, A., Vandenbroucke-Grauls, C.M.J.E., Berg, D., Hoff-
man, P.S. and Kusters, J.G. (1999) Insertion of mini-IS605 and
deletion of adjacent sequences in the nitroreductase (rdxA) gene
cause metronidazole resistance in Helicobacter pylori
NCTC11637. Antimicrob. Agents Chemother. 43, 2657–2662.
[4] Goodwin, A., Kersulyte, D., Sisson, G., Veldhuyzen van Zanten,
S.J.O., Berg, D.E. and Hoffman, P.S. (1998) Metronidazole
resistance in Helicobacter pylori is due to null mutations in a gene
(rdxA) that encodes an oxygen-insensitive NADPH nitroreduc-
tase. Mol. Microbiol. 28, 383–393.
[5] Kwon, D.H., El-Zaatari, F.A.K., Kato, M., Osato, M.S., Reddy,
R., Yamaoka, Y. and Graham, D.Y. (2000) Analysis of rdxA and
involvement of additional genes encoding NAD(P)H flavin
oxidoreductase (FrxA) and ferrodoxin-like protein (FdxB) in
metronidazole resistance in Helicobacter pylori. Antimicrob.
Agents Chemother. 44, 2133–2142.
[6] Kwon, D.H., Kato, M., El-Zaatari, F.A.K., Osato, M.S. and
Graham, D.Y. (2000) Frame-shift mutations in NAD(P)H flavin
oxidoreductase encoding gene (frxA) from metronidazole resis-
tant Helicobacter pylori ATCC43505 and its involvement in
metronidazole resistance. FEMS Microbiol. Lett. 118, 197–202.
[7] Smith, M.A., Jorgensen, M.A., Mendz, G.L. and Hazell, S.L.
(1998) Metronidazole resistance and microaerophily in Campylo-
bacter species. Arch. Microbiol. 170, 279–284.
[8] Smith, M.A. and Edwards, D.I. (1995) Redox potential and
oxygen concentration as factors in the susceptibility of Helico-
bacter pylori to nitroheterocyclic drugs. J. Antimicrob. Chemo-
ther. 35, 751–764.
[9] Smith, M.A. and Edwards, D.I. (1997) Oxygen scavenging,
NADH-oxidase and metronidazole resistance in Helicobacter
pylori. J. Antimicrob. Chemother. 39, 347–353.
[10] Mendz, G.L. and Trend, M.A. (2001) Intracellular redox status
and antibiotic resistance in enterogastric microaerophilic bacteria:
evidence for the �scavenging of oxygen� hypothesis. Redox Rep. 6,
179–181.
[11] Hoffman, P.S., Goodwin, A., Johnson, J., Magee, K. and
Veldhuyzen van Zanten, S.J.O. (1996) Metabolic activities of
metronidazole-sensitive and -resistant strains of Helicobacter
pylori : repression of pyruvate oxidoreductase and expression of
isocitrate lyase activity correlate with resistance. J. Bacteriol. 178,
4822–4829.
[12] Trend, M.A., Jorgensen, M.A., Hazell, S.L. and Mendz, G.L.
(2001) Oxidases and reductases are involved in metronidazole
sensitivity in Helicobacter pylori. Int. J. Biochem. Cell Biol. 33,
143–153.
[13] Jorgensen, M.A., Trend, M.A., Hazell, S.L. and Mendz, G.L.
(2001) Potential involvement of several nitroreductases in metro-
nidazole resistance in Helicobacter pylori. Arch. Biochem. Bio-
phys. 392, 180–191.
[14] Schulz, A.R. (1994) Enzyme Kinetics, from Diastase to Multi-
enzyme Systems p. 246. Cambridge University Press, New York.
[15] Alarcon, T., Domingo, D. and Lopez-Brea, M. (1999) Antibiotic
resistance problems with Helicobacter pylori. Int. J. Antimicrob.
Agents 12, 19–26.
[16] Tankovic, J., Lamarque, D., Delchier, J.C., Soussy, C.J., Labigne,
A. and Jenks, P.J. (2000) Frequent association between alteration
of the rdxA gene and metronidazole resistance in French and
North African isolates of Helicobacter pylori. Antimicrob. Agents
Chemother. 44, 608–613.
[17] Jenks, P.J., Ferrero, R.L. and Labigne, A. (1999) The role
of the rdxA gene in the evolution of metronidazole
resistance in Helicobacter pylori. J. Antimicrob. Chemother.
43, 753–758.
[18] Solca, N.M., Bernasconi, M.V. and Piffaretti, J.C. (2000) Mech-
anism of metronidazole resistance in Helicobacter pylori: com-
parison of the rdxA gene sequences of 30 strains. Antimicrob.
Agents Chemother. 44, 2207–2210.
[19] Paul, R., Postius, S., Melchers, K. and Schafer, K.P. (2001)
Mutations of the Helicobacter pylori genes rdxA and pbp1 cause
resistance against metronidazole and amoxicillin. Antimicrob.
Agents Chemother. 45, 962–965.
[20] Kwon, D.H., Hulten, K., Kato, M., Kim, J.J., Lee, M., El-
Zaatari, F.A., Osato, M.S. and Graham, D.Y. (2001) DNA
sequence analysis of rdxA and frxA from 12 pairs of metronida-
zole-sensitive and -resistant clinical Helicobacter pylori isolates.
Antimicrob. Agents Chemother. 45, 2609–2615.
[21] Jeong, J.Y., Mukhopadhyay, A.K., Dailidiene, D., Wang, Y.,
Velapatino, B., Gilman, R.H., Parkinson, A.J., Nair, G.B., Wong,
B.C., Lam, S.K., Mistry, R., Segal, I., Yuan, Y., Gao, H.,
Alarcon, T., Brea, M.L., Ito, Y., Kersulyte, D., Lee, H.K., Gong,
Y., Goodwin, A., Hoffman, P.S. and Berg, D.E. (2000) Sequential
inactivation of rdxA (HP0954) and frxA (HP0642) nitroreductase
genes causes moderate and high-level metronidazole resistance in
Helicobacter pylori. J. Bacteriol. 182, 5082–5090.
[22] Jeong, J.Y. and Berg, D.E. (2000) Mouse colonizing Helicobacter
pylori SS1 is unusually susceptible to metronidazole due to two
complementary reductase activities. Antimicrob. Agents Chemo-
ther. 44, 3127–3132.
[23] Jeong, J.Y., Mukhopadhyay, A.K., Akada, J.K., Dailidiene, D.,
Hoffman, P.S. and Berg, D.E. (2001) Roles of FrxA and RdxA
nitroreductases of Helicobacter pylori in susceptibility and resis-
tance to metronidazole. J. Bacteriol. 183, 5155–5162.
[24] Watarai, M., Tobe, T., Yoshikawa, M. and Sasakawa, C. (1995)
Disulfide oxidoreductase activity of Shigella flexneri is required
for release of Ipa proteins and invasion of epithelial cells. Proc.
Natl. Acad. Sci. USA 92, 4927–4931.
[25] Yu, J. and Kroll, J.S. (1999) DsbA: a protein folding catalyst
contributing to bacterial virulence. Microbes. Infect. 1, 1221–
1228.
[26] Jaeger, T., Budde, H., Flohe, L., Menge, U., Singh, M., Trujillo,
M. and Radi, R. (2004) Multiple thioredoxin-mediated routes to
detoxify hydroperoxides in Mycobacterium tuberculosis. Arch.
Biochem. Biophys. 423, 182–191.
[27] Mendz, G.L. and Megraud, F. (2002) Is the molecular basis of
metronidazole resistance in microaerophilic organisms under-
stood? Trends Microbiol. 10, 370–375.