hb-gam inhibits proliferation and enhances differentiation of neural stem cells
TRANSCRIPT

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 26 (2004) 75–88
HB-GAM inhibits proliferation and enhances differentiation of
neural stem cells$
Anni Hienola,a,* Mari Pekkanen,b Erkki Raulo,a Paivi Vanttola,a and Heikki Rauvalaa,*
aNeuroscience Center, Department of Biosciences and Institute of Biotechnology, University of Helsinki, Helsinki 00014, FinlandbFinnzymes Oy, 02201 Espoo, Finland
Received 15 October 2003; revised 23 January 2004; accepted 30 January 2004
Available online 27 March 2004
Proliferation of neural stem cells in the embryonic cerebral cortex is
regulated by many growth factors and their receptors. Among the key
molecules stimulating stem cell proliferation are FGF-2 and the FGF
receptor-1. This ligand-receptor system is highly dependent on the
surrounding heparan sulfates. We have found that heparin-binding
growth-associated molecule (HB-GAM, also designated as pleiotro-
phin) regulates neural stem cell proliferation in vivo and in vitro.
Deficiency of HB-GAM results in a pronounced, up to 50% increase in
neuronal density in the adult mouse cerebral cortex. This phenotype
arises during cortical neurogenesis, when HB-GAM knockout embryos
display an enhanced proliferation rate as compared to wild-type
embryos. Further, our in vitro studies show that exogenously added
HB-GAM inhibits formation and growth of FGF-2, but not EGF,
stimulated neurospheres, restricts the number of nestin-positive neural
stem cells, and inhibits FGF receptor phosphorylation. We propose that
HB-GAM functions as an endogenous inhibitor of FGF-2 in stem cell
proliferation in the developing cortex.
D 2004 Elsevier Inc. All rights reserved.
Introduction
Early development of the mammalian cerebral cortex can be
roughly divided into two phases: the establishment of the founder
stem cell population in the ventricular epithelium by symmetrical
stem cell divisions, and the neurogenetic interval, when post-mitotic
migrating neurons are born by asymmetric stem cell divisions
(Rakic, 1995; Takahashi et al., 1994). Cell cycle length and the
number of symmetrical and asymmetrical divisions are strictly
regulated. The differentiation rate of born neural stem cells is also
heavily controlled and very minor changes in proliferation kinetics
1044-7431/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.mcn.2004.01.018
$ Supplementary data associated with this article can be found, in the
online version, at doi:10.1016/j.mcn.2004.01.018.
* Corresponding authors. Neuroscience Center, Department of Bio-
sciences and Institute of Biotechnology, University of Helsinki, Viikinkaari
4, PO Box 56, Helsinki 00014, Finland. Fax: +358-9-19157620.
E-mail addresses: [email protected] (A. Hienola),
[email protected] (H. Rauvala).
Available online on ScienceDirect (www.sciencedirect.com.)
can result in astonishing changes in cell number and laminar thick-
ness (Caviness et al., 1995; Rakic, 1995; Takahashi et al., 1997).
The molecular mechanisms regulating the final number and
quality of appearing cortical cells and cell connections are very
complex and poorly understood. Ubiquitous growth factors like
FGFs are known to function in growth and differentiation of cortical
neurons and glia in a very general manner (for examples and
reviews, see Burrows et al., 1997; Junier, 2000; Tropepe et al.,
1999). The concept of stem cell ‘‘niche’’ has become important in
discussions about stem cell regulation. In developing cerebral
cortex, an important part of this niche may be the extracellular
matrix (ECM), which functions as a dynamic local storage for
growth factors and may regulate their binding to cell surfaces.
Heparin-binding growth-associated molecule (HB-GAM, pleio-
trophin, OSF-1) was initially isolated as a neurite outgrowth-
promoting factor from perinatal rat brain (Merenmies and Rauvala,
1990; Rauvala, 1989). It is a basement membrane-associated
protein in embryonic nonneural tissues (Mitsiadis et al., 1995)
and associates with growing axonal pathways in developing brain
(Kinnunen et al., 1999; Rauvala et al., 1994). HB-GAM expression
in the developing cerebral cortex starts at the onset of neurogenesis
(embryonic days 11–12 in mouse), when it is abundant in the
ventricular zone (VZ) of the cortical lobes and the medial gangli-
onic eminence. There it is localized in the radially orientated
processes of neuroepithelial cells (Kinnunen et al., 1998a,b; Rau-
vala et al., 1994; Silos-Santiago et al., 1996). The prominent
expression of HB-GAM in the germinal zones would be compatible
with a role in cortical development, but this possible function
remains unexplored.
HB-GAM consists of two thrombospondin type 1 (TSR)
domains flanked by lysine-rich N- and C-termini that are random
coils in solution (Kilpelainen et al., 2000). The TSR domains are
most likely responsible for binding to heparan sulfates and to the
cell surface, while the role of the unstructured lysine tails is
unknown (Kilpelainen et al., 2000). HB-GAM has a close relative,
midkine (MK), that was isolated and cloned as a retinoic acid-
induced differentiation factor (Matsubara et al., 1990; Nurcombe et
al., 1992). The in vivo functions of HB-GAM are still largely
unsolved, although it is known to affect adult hippocampal plastic-
ity in rodents (Amet et al., 2001; Lauri et al., 1996, 1998; Pavlov et
al., 2002).
A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8876
HB-GAM-induced effects on neurite outgrowth and synaptic
plasticity are mediated at least partially by the heparan sulfate
proteoglycan syndecan-3, also designated as N-syndecan (Carey et
al., 1992; Lauri et al., 1999; Raulo et al., 1994). This receptor
signals further through the cortactin–src-kinase pathway (Kinnu-
nen et al., 1996, 1998a,b), inducing cytoskeletal changes. HB-
GAM has a high affinity to heparan sulfate chains similar to those
which also bind FGF-2. In fact, HB-GAM and FGF-2 compete for
binding to syndecan-3 in vitro (Kinnunen et al., 1996; Raulo et al.,
1994). Both of them also bind to phosphacan, an abundant ECM
molecule in the brain and an extracellular splice variant of another
known receptor for HB-GAM, receptor type protein tyrosine
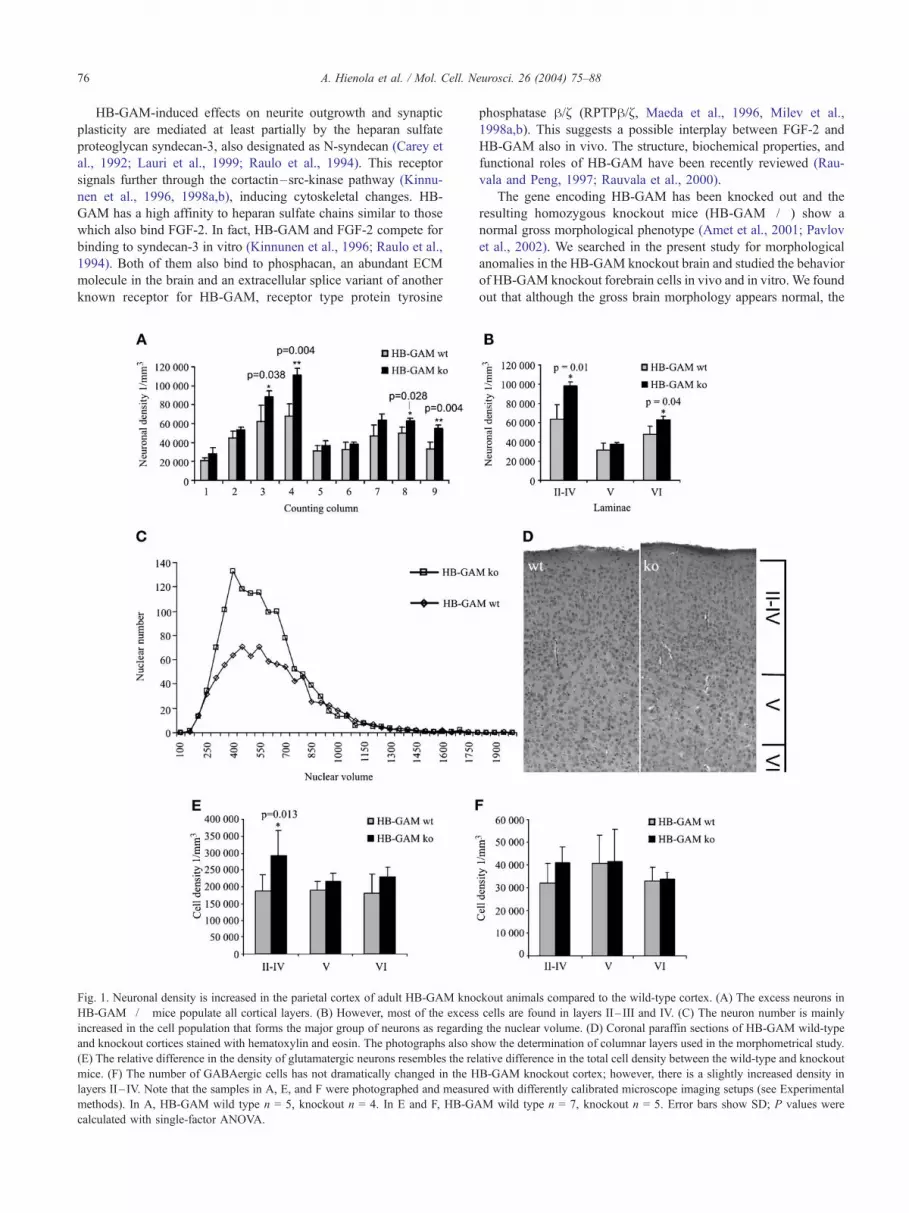
Fig. 1. Neuronal density is increased in the parietal cortex of adult HB-GAM kno
HB-GAM�/� mice populate all cortical layers. (B) However, most of the excess
increased in the cell population that forms the major group of neurons as regardin
and knockout cortices stained with hematoxylin and eosin. The photographs also s
(E) The relative difference in the density of glutamatergic neurons resembles the re
mice. (F) The number of GABAergic cells has not dramatically changed in the H
layers II– IV. Note that the samples in A, E, and F were photographed and measur
methods). In A, HB-GAM wild type n = 5, knockout n = 4. In E and F, HB-GA
calculated with single-factor ANOVA.
phosphatase h/~ (RPTPh/~ , Maeda et al., 1996, Milev et al.,
1998a,b). This suggests a possible interplay between FGF-2 and
HB-GAM also in vivo. The structure, biochemical properties, and
functional roles of HB-GAM have been recently reviewed (Rau-
vala and Peng, 1997; Rauvala et al., 2000).
The gene encoding HB-GAM has been knocked out and the
resulting homozygous knockout mice (HB-GAM�/�) show a
normal gross morphological phenotype (Amet et al., 2001; Pavlov
et al., 2002). We searched in the present study for morphological
anomalies in the HB-GAM knockout brain and studied the behavior
of HB-GAM knockout forebrain cells in vivo and in vitro. We found
out that although the gross brain morphology appears normal, the
ckout animals compared to the wild-type cortex. (A) The excess neurons in
cells are found in layers II– III and IV. (C) The neuron number is mainly
g the nuclear volume. (D) Coronal paraffin sections of HB-GAM wild-type
how the determination of columnar layers used in the morphometrical study.
lative difference in the total cell density between the wild-type and knockout
B-GAM knockout cortex; however, there is a slightly increased density in
ed with differently calibrated microscope imaging setups (see Experimental
M wild type n = 7, knockout n = 5. Error bars show SD; P values were

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–88 77
neuronal number is in fact increased in the adult cerebral cortex of
the HB-GAM knockout mice compared to their wild-type litter-
mates. This phenotype is due to enhanced proliferation of ventric-
ular zone neural stem cells in the early embryonic cortex and
possibly delayed maturation of neurons in the postnatal brain.
Our experiments in vitro show that both recombinant HB-GAM
and its truncated form containing only the TSR domains (di-TSR-
fragment) can inhibit proliferation and induce differentiation in
FGF-2-stimulated neural stem cells. Furthermore, while FGF-2
stimulation of neural stem cells increases the phosphorylation of
FGF receptor, exogenous HB-GAM and its di-TSR-fragment re-
duce this phosphorylation.
Fig. 2. In the perinatal brain, cell density differences are visible between HB-G
thickening of VZ and SVZ in E16 cortex and (B) increased neuronal density in
increased proliferation in E12.5 HB-GAM knockout cortex. (E) The bars show B
Error bars show SD, P values calculated with single-factor ANOVA. Scale bars:
zone; Hi, ventricular zone of hippocampal formation; GE, ganglionic eminence. Ph
taken with 63� objective.
Based on our findings, we postulate that inactivating HB-GAM
in vivo removes an important differentiation-regulating factor from
the developing brain, thus allowing enhanced proliferation of neural
stem cells. This restrictive behavior is likely to be tightly linked to
the proteoglycan environment in the surface of, or surrounding,
neural stem cells. We suggest that HB-GAM specifically inhibits
FGF-2’s function via FGF receptor both in vitro and in vivo.
In this paper, we refer to cells isolated from embryonic mouse
forebrain and cultured as neurospheres as neural stem cells (for
discussion on stem cell or progenitor cell definitions, see Seaberg
and van der Kooy, 2003). Under culture conditions described in this
work, these cells continue self-renewing proliferation and formation
AM knockout and wild-type animals. (A) Lack of HB-GAM results in
the SVZ of P3 pups. (C and D) Six hours of BrdU administration reveals
rdU+ cell densities in E12.5 cortex. HB-GAM�/�, n = 5; wild type, n = 5.
A, B, and D = 100 Am, C = 200 Am. LV, lateral ventricle; VZ, ventricular
otos A, B, and C were taken with 20� objective, A and C are collases. D is
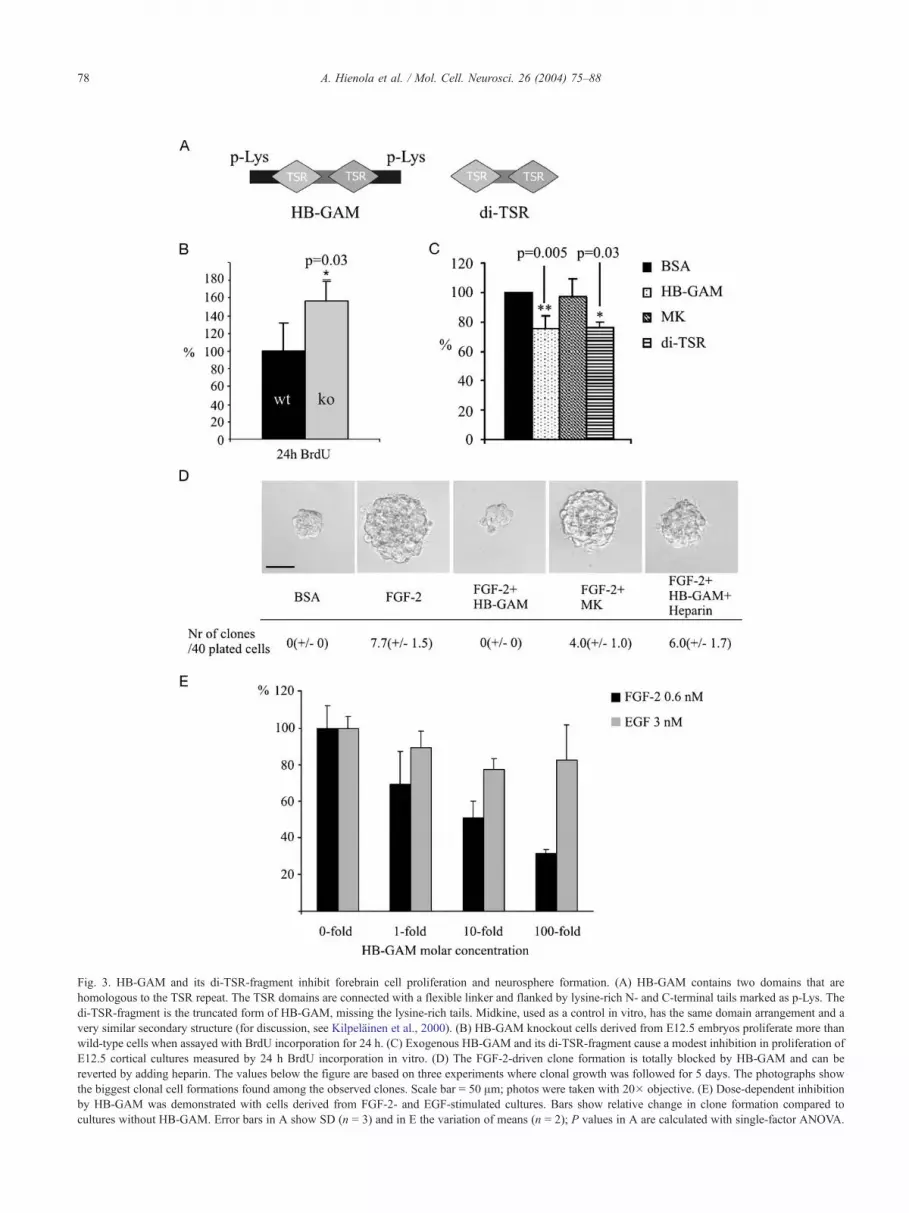
Fig. 3. HB-GAM and its di-TSR-fragment inhibit forebrain cell proliferation and neurosphere formation. (A) HB-GAM contains two domains that are
homologous to the TSR repeat. The TSR domains are connected with a flexible linker and flanked by lysine-rich N- and C-terminal tails marked as p-Lys. The
di-TSR-fragment is the truncated form of HB-GAM, missing the lysine-rich tails. Midkine, used as a control in vitro, has the same domain arrangement and a
very similar secondary structure (for discussion, see Kilpelainen et al., 2000). (B) HB-GAM knockout cells derived from E12.5 embryos proliferate more than
wild-type cells when assayed with BrdU incorporation for 24 h. (C) Exogenous HB-GAM and its di-TSR-fragment cause a modest inhibition in proliferation of
E12.5 cortical cultures measured by 24 h BrdU incorporation in vitro. (D) The FGF-2-driven clone formation is totally blocked by HB-GAM and can be
reverted by adding heparin. The values below the figure are based on three experiments where clonal growth was followed for 5 days. The photographs show
the biggest clonal cell formations found among the observed clones. Scale bar = 50 Am; photos were taken with 20� objective. (E) Dose-dependent inhibition
by HB-GAM was demonstrated with cells derived from FGF-2- and EGF-stimulated cultures. Bars show relative change in clone formation compared to
cultures without HB-GAM. Error bars in A show SD (n = 3) and in E the variation of means (n = 2); P values in A are calculated with single-factor ANOVA.
A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8878

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–88 79
of clonal neurospheres at least for six passages from isolation. The
cells, when plated on laminin, are nestin positive, respond to FGF-2
and EGF, and generate both neurons and glia.
Fig. 4. HB-GAM induces differentiation of neural stem cells. The
maintenance of neurospheres on non-coated glass was determined by
observing the differentiation of the spheres. (a) Shows a neurosphere with
undifferentiated morphology and (b) shows a sphere that has differentiated.
Based on morphological differences, the number of undifferentiated spheres
was counted after 3 days in culture under different conditions: without
FGF-2, with FGF-2, with FGF-2 and HB-GAM, and with FGF-2, HB-
GAM, and heparin. Scale bar = 50 Am; photos were taken with 20�objective. Error bars show SD (n = 3), except for the assay with heparin,
which gave in two independent experiments the percentage values 76.0 and
76.7; P values were calculated with single-factor ANOVA.
Results
Disruption of HB-GAM gene causes an increased neuronal density
in the cerebral cortex
In adult HB-GAM-deficient mice, the gross morphology of the
brain appeared normal as has been previously described (Amet et al.,
2001). When a more detailed stereological analysis of both parietal
and hind limb motor cortex was carried out, an average increase of
39% in neuronal density was found. The increase was detectable
throughout the whole cortical thickness but was most obvious in
layers II– IV (54% increase) and VI (31% increase, Figs. 1A and
1B). The neuronal identity of the counted cells was verified with
anti-GFAP and anti-NF immunostaining. We also calculated the size
distribution curve for the counted andmeasured nuclei of the cortical
cells (Fig. 1C). This showed that the excess cells in the knockout
mice belonged to the size-group that was the most populated also in
the wild-type mice, with nuclear volume ranging from 350 to 750
Am3. The neuronal number in this particular size-group was almost
doubled in the knockout mice in comparison with the wild type. No
increase in the number of the very large pyramidal neurons (>1000
Am3) populating the lamina V could be seen. This may be due to the
relative scarcity of these cells.
We further examined the distribution of different neuronal
subtypes in the cortical laminae. We used GABA and glutamate
receptor 2 immunopositivity for distinguishing between interneur-
ons and pyramidal neurons. We could see an increase in the density
of both of these neuronal types, but the increase in the density of
the GABAergic neurons was less prominent than the density of
glutamatergic neurons. Moreover, the density increase in GABA+
cells was only visible in the high laminae II– IV, whereas the
density of glutamatergic neurons was increased in all layers of the
knockout cortex in approximately the same proportions as the
measured total density (Fig. 1E). The size distribution based on
nuclear volume of the glutamatergic neurons was similar to that
observed in the whole cortical population, meaning that excess
neurons were once again found in the most populated size groups
and only some increase was found in the group of very large
pyramidal neurons populating lamina V (data not shown).
During this more careful histological examination of the adult
cortex, we could not find anything unusual in the structure or
morphology of either neurons or glial cells. Bielschowsky-silver-
staining and immunohistochemistry with anti-neurofilament (NF,
200, 145, and 68 kDa, from Chemicon) and glial fibrillary acidic
protein (GFAP, from Chemicon) antibodies revealed no abnormal-
ities in the distribution of neurites or axonal pathways in the cortex
(not shown). However, based on neurofilament expression, the
neuronal differentiation is delayed in HB-GAM�/� mice (see
Supplementary Fig. 1).
Enhanced cell proliferation in the germinal zones of
HB-GAM�/� embryos
Histology of embryonic (E16) and postnatal (P3) brain sug-
gested increased cell numbers in the cerebral cortex (Figs. 2A and
2B). Immunostaining with an M-phase cell cycle marker, phos-
phorylated histone 3, showed an increase in the density of mitotic
cells in E12 knockout embryos (0.55 F 0.06 � 106 1/mm3, n = 3)
in comparison with wild-type littermate samples (0.32 F 0.04 �106 1/mm3, n = 3). This type of assay, however, only shows the
number of mitotic cells at the time point of sample preparation and
is thus unreliable in estimation of the proliferation rate.
To study more carefully the in vivo stem cell proliferation, we
gave 5-bromo-2V-deoxy-uridine (BrdU) injections to timed-preg-
nant HB-GAM+/� females for 6 h at E12.5. The injection protocol
was based on a study by Takahashi et al., 1996, where the authors
determined the length of the stem cell cycle in the mouse embryonic
VZ at different embryonic ages. The BrdU incorporation time used
in our protocol should result in labeling of all VZ cells undergoing
cell divisions at embryonic day 12.5. A visible increase was found
in the BrdU+ cell density in the HB-GAM knockout ventricular
zone when compared to the wild type (Figs. 2C and 2D). The BrdU+
cell density was on the average 50% higher in the knockouts than in
the wild types (1.84� 106/mm3 F 0.22� 106 and 1.23� 106/mm3
F 0.17 � 106, respectively). The 6-h BrdU administration thus
labeled 60% of the total amount of cells in the knockout and only
38% in the wild-type VZ (Fig. 2E).
HB-GAM and its di-TSR-fragment inhibit proliferation of
embryonic mouse forebrain cells
We compared the amount of BrdU incorporation in the HB-
GAM knockout and wild-type cortical cultures and saw a clear

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8880
increase of 55% in the number of BrdU+ knockout cells (Fig. 3B).
This finding together with the phenotype of the adult HB-GAM
knockout mice raised the question of whether exogenously added
recombinant HB-GAM would induce proliferation arrest and
change the differentiation fates of neural stem cells in vitro. It
was also interesting to see if HB-GAM’s di-TSR-fragment (Fig.
3A) would act in a similar manner as HB-GAM in vitro. E12.5
forebrain cells from HB-GAM knockout embryos were cultured for
24 h in the presence or absence of recombinant HB-GAM, HB-
GAM’s di-TSR-fragment, or midkine, and the amount of cell
proliferation was measured. The proliferation inhibition caused
by recombinant HB-GAM or its di-TSR-fragment within this time
period was less pronounced (30%) than what was expected based
on the knockout phenotype and BrdU incorporation in vivo (Fig.
3C). On the other hand, midkine did not induce any changes at all
as compared to the baseline proliferation (Fig. 3C).
HB-GAM inhibits clone formation in FGF-2 but not in EGF
stimulated neural stem cells
From previous studies, we knew that HB-GAM can compete
with FGF-2 in heparin binding (see Introduction and Discussion)
and that FGF-2 is one of the key factors regulating stem cells in
early VZ. We isolated neural stem cells as neurospheres from
Fig. 5. HB-GAM and its di-TSR-fragment cause growth inhibition on neurospher
grow for 5 days, after which the colonies were photographed and fixed for immu
neurospheres form large discs of cells migrating away from the central neurospher
discs did not spread but stayed compact and sent only few radial processes. When
clearly diminished. Again, adding midkine to the neurosphere solution did not ha
mouse embryos to see if HB-GAM could induce a more pro-
nounced growth arrest in FGF-2-stimulated stem cells than it had
in primary forebrain cells. FGF-2 was used to induce neurosphere
formation and to maintain the stem cells during the assays. We
used EGF-stimulated neurospheres to control the specificity of the
effects of HB-GAM. The concentration of FGF-2 that induced
most efficient clonal growth in our experiments was determined to
be between 1 and 10 ng/ml in a titration assay (data not shown).
Based on this finding and the available literature, we chose the
concentration of FGF-2 to be 10 ng/ml and of EGF 20 ng/ml
(Hulspas et al., 1997).
First we derived neurospheres with FGF-2 as the only stimu-
lating growth factor. The cells were plated in clonal densities in the
presence of FGF-2 and additional HB-GAM or midkine. The FGF-
2-stimulated cells failed to grow any clones in the presence of HB-
GAM after 5 days of observation. With FGF-2 alone, the clonal
growth took place (Fig. 3D). The inhibitory effect of HB-GAM
could be reverted by adding heparin to the culture medium.
Midkine did not display a significant effect on the formation of
spheres in this assay.
Our next experiment was to demonstrate the dose dependency
of HB-GAM-induced proliferation arrest in FGF-2-stimulated stem
cells. We produced neurospheres using both FGF-2 and EGF as
stimulating growth factors in the culture medium. The cells were
e discs grown on laminin. Neurospheres were plated on laminin and let to
nostaining. In the presence of neurosphere medium containing FGF-2, the
e core. When the medium contained no growth factor or just HB-GAM, the
HB-GAM was added to the normal neurosphere medium, the disc size was
ve any effect on disc size or appearance. Scale bar is 100 Am.
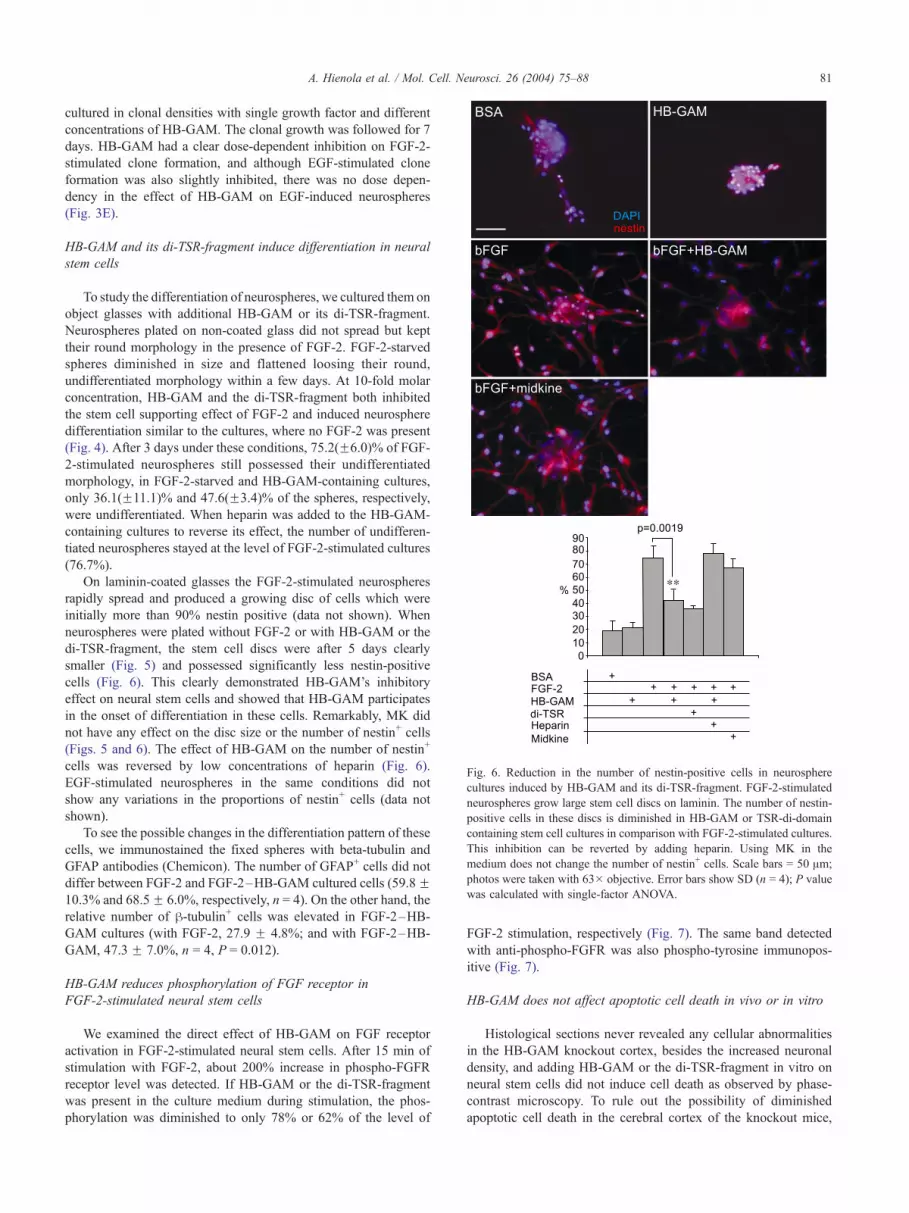
Fig. 6. Reduction in the number of nestin-positive cells in neurosphere
cultures induced by HB-GAM and its di-TSR-fragment. FGF-2-stimulated
neurospheres grow large stem cell discs on laminin. The number of nestin-
positive cells in these discs is diminished in HB-GAM or TSR-di-domain
containing stem cell cultures in comparison with FGF-2-stimulated cultures.
This inhibition can be reverted by adding heparin. Using MK in the
medium does not change the number of nestin+ cells. Scale bars = 50 Am;
photos were taken with 63� objective. Error bars show SD (n = 4); P value
was calculated with single-factor ANOVA.
A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–88 81
cultured in clonal densities with single growth factor and different
concentrations of HB-GAM. The clonal growth was followed for 7
days. HB-GAM had a clear dose-dependent inhibition on FGF-2-
stimulated clone formation, and although EGF-stimulated clone
formation was also slightly inhibited, there was no dose depen-
dency in the effect of HB-GAM on EGF-induced neurospheres
(Fig. 3E).
HB-GAM and its di-TSR-fragment induce differentiation in neural
stem cells
To study the differentiation of neurospheres, we cultured them on
object glasses with additional HB-GAM or its di-TSR-fragment.
Neurospheres plated on non-coated glass did not spread but kept
their round morphology in the presence of FGF-2. FGF-2-starved
spheres diminished in size and flattened loosing their round,
undifferentiated morphology within a few days. At 10-fold molar
concentration, HB-GAM and the di-TSR-fragment both inhibited
the stem cell supporting effect of FGF-2 and induced neurosphere
differentiation similar to the cultures, where no FGF-2 was present
(Fig. 4). After 3 days under these conditions, 75.2(F6.0)% of FGF-
2-stimulated neurospheres still possessed their undifferentiated
morphology, in FGF-2-starved and HB-GAM-containing cultures,
only 36.1(F11.1)% and 47.6(F3.4)% of the spheres, respectively,
were undifferentiated. When heparin was added to the HB-GAM-
containing cultures to reverse its effect, the number of undifferen-
tiated neurospheres stayed at the level of FGF-2-stimulated cultures
(76.7%).
On laminin-coated glasses the FGF-2-stimulated neurospheres
rapidly spread and produced a growing disc of cells which were
initially more than 90% nestin positive (data not shown). When
neurospheres were plated without FGF-2 or with HB-GAM or the
di-TSR-fragment, the stem cell discs were after 5 days clearly
smaller (Fig. 5) and possessed significantly less nestin-positive
cells (Fig. 6). This clearly demonstrated HB-GAM’s inhibitory
effect on neural stem cells and showed that HB-GAM participates
in the onset of differentiation in these cells. Remarkably, MK did
not have any effect on the disc size or the number of nestin+ cells
(Figs. 5 and 6). The effect of HB-GAM on the number of nestin+
cells was reversed by low concentrations of heparin (Fig. 6).
EGF-stimulated neurospheres in the same conditions did not
show any variations in the proportions of nestin+ cells (data not
shown).
To see the possible changes in the differentiation pattern of these
cells, we immunostained the fixed spheres with beta-tubulin and
GFAP antibodies (Chemicon). The number of GFAP+ cells did not
differ between FGF-2 and FGF-2–HB-GAM cultured cells (59.8F10.3% and 68.5F 6.0%, respectively, n = 4). On the other hand, the
relative number of h-tubulin+ cells was elevated in FGF-2–HB-
GAM cultures (with FGF-2, 27.9 F 4.8%; and with FGF-2–HB-
GAM, 47.3 F 7.0%, n = 4, P = 0.012).
HB-GAM reduces phosphorylation of FGF receptor in
FGF-2-stimulated neural stem cells
We examined the direct effect of HB-GAM on FGF receptor
activation in FGF-2-stimulated neural stem cells. After 15 min of
stimulation with FGF-2, about 200% increase in phospho-FGFR
receptor level was detected. If HB-GAM or the di-TSR-fragment
was present in the culture medium during stimulation, the phos-
phorylation was diminished to only 78% or 62% of the level of
FGF-2 stimulation, respectively (Fig. 7). The same band detected
with anti-phospho-FGFR was also phospho-tyrosine immunopos-
itive (Fig. 7).
HB-GAM does not affect apoptotic cell death in vivo or in vitro
Histological sections never revealed any cellular abnormalities
in the HB-GAM knockout cortex, besides the increased neuronal
density, and adding HB-GAM or the di-TSR-fragment in vitro on
neural stem cells did not induce cell death as observed by phase-
contrast microscopy. To rule out the possibility of diminished
apoptotic cell death in the cerebral cortex of the knockout mice,

Fig. 7. HB-GAM inhibits phosphorylation of the FGF receptor. Neural stem
cells plated on laminin were first deprived of growth factor and then
stimulated with FGF-2 for 15 min. In total cell lysates the a-phospho-FGFR
antibody detected an increased signal in FGF-2-stimulated cells as
compared to nonstimulated controls. HB-GAM and di-TSR-fragment
prevented the increase. a-phospho-tyrosine immunoblotting detected a
band of the same size (120 kDa) as a-phospho-FGFR and showed a similar
variation in the level of phosphorylation. a-FGFR1 immunoblotting was
used to verify the constant amount of FGFR1 in all samples. Error bars
show SD (n = 3); P values were calculated with paired t test for means.
A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8882
we stained E12, E16, and P10 brain sections and embryonic
primary neurons in culture with anti-active caspase-3 antibody.
Activation of caspase-3 is considered to be an irreversible phase in
the complex protein activation cascade that leads to programmed
cell death (Grutter, 2000). Seven to 10 sample sections spanning the
whole cortical hemispheres from four animals per genotype were
averaged. At E12, 3.6 F 1.2 and 3.5 F 1.2 apoptotic figures per
section were detected in wild-type and knockout animals, respec-
tively. The corresponding figures at E16 were 3.4 F 0.3 and 3.4 F0.5, and at P10 13.1 F 2.9 and 12.0 F 2.1 in wild-type and
knockout animals. Similarly, in vitro apoptotic activity during HB-
GAM treatment stayed the same as in control cultures: 16.9F 1.3%
of HB-GAM-treated and 14.5F 5.7% of untreated cells were active
caspase-3 positive.
Discussion
In this study, we show that depleting HB-GAM in mice results in
increased neuronal density in the cerebral cortex. The increase in the
laminae II– IV can exceed 50%, which we think is a very significant
change. The magnitude of this phenotype indicates an important
regulatory role for HB-GAM in cortical development. We also show
that exogenous recombinant HB-GAM and its heparin-binding di-
TSR-fragment can inhibit neural stem cell proliferation and neuro-
sphere formation in vitro, and induce differentiation towards a
neuronal phenotype. We also show that this inhibition is rather
specific for FGF-2-responsive neural stem cells because a similar
inhibition was not seen in EGF-derived neural stem cells. In
addition, FGF-2-induced FGF receptor phosphorylation detected
in stem cell lysates is reduced by exogenous HB-GAM and its di-
TSR-fragment.
We suggest that the mechanism of inhibition is likely due to
competition between HB-GAM and FGF-2 in binding to proteo-
glycan co-receptor(s) required for FGF-2-induced proliferation. We
base this claim on several findings presented both in this paper and
in studies by others: first, the knockout phenotype described in the
present study is essentially a reverse situation of the phenotype
found in the FGF-2 knockout (Dono et al., 1998; Raballo et al.,
2000). In the FGF-2 knockout, a decreased neuronal density has
been found essentially in the same layers where we have found an
increased neuronal density in the present study.
Second, the present paper demonstrates that HB-GAM and its
heparin–heparan sulfate binding di-TSR-fragment are able to in-
hibit neural stem cell proliferation and suppress FGF-2-driven
neurosphere formation to the level seen during growth factor
deprivation. The effect on neural stem cell growth is clearly
dependent on heparan sulfate proteoglycans because the growth
arrest can be reversed with low concentrations of heparin. Further,
no such inhibition can be seen in the EGF-stimulated neural stem cell
cultures.
Third, HB-GAM and the di-TSR-fragment also reduce FGF
receptor phosphorylation in FGF-2-stimulated neural stem cells. In
addition, HB-GAM and its di-TSR-fragment reduce the number of
nestin-immunopositive cells in neural stem cell cultures, indicating
the start of differentiation of these cells. This differentiation is
directed towards the neuronal phenotype, indicated by h-tubulinexpression. Specificity of the effect is suggested by the unexpected
finding that the closest HB-GAM homologue midkine that binds to
heparin–heparan sulfates as strongly as HB-GAM does not inhibit
proliferation of the stem cells or diminish the number of nestin-
expressing cells. Interestingly, HB-GAM has been previously
linked to cell differentiation at a more general level in limb bud
development and chondrogenesis involving FGF-2 (Dreyfus et al.,
1998; Szabat and Rauvala, 1996).
Fourth, HB-GAM binds strongly to heparin and co-purifies with
FGF-2 in salt gradient elution from heparin-Sepharose (Rauvala,
1989). Naturally occurring heparan sulfates contain a wide variety
of carbohydrate epitopes binding growth factors and matrix mole-
cules; the HB-GAM-binding carbohydrate epitope overlaps with
that binding FGF-2 (Kinnunen et al., 1996). In addition, previous in
vitro results suggest competition in the binding of HB-GAM and
FGF-2 to heparin–heparan sulfate (Raulo et al., 1994; Szabat and
Rauvala, 1996). In particular, HB-GAM and FGF-2 display a
similar binding to brain-derived N-syndecan and are competitive
in direct binding assays (Raulo et al., 1994).
Finally, HB-GAM is expressed in the rodent brain up to 100-
fold molar excess as compared to FGF-2 (Rauvala, 1989). From
this viewpoint and considering high local expression of HB-GAM
in the germinal zones at the stage when FGF-2 is an essential
proliferation-enhancing factor (Rauvala et al., 1994), it seems
obvious that the expression level of HB-GAM in early develop-

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–88 83
ing brain should be high enough to brake FGF-driven prolifera-
tion. Our finding that the density of both GABAergic and
glutamatergic neurons is increased in the knockout mice supports
the idea about HB-GAM’s general inhibitory function in the
germinal areas of the early brain. These two distinct types of
cortical neurons have separate sources—glutamatergic pyramidal
neurons are born in the VZ underlying their future cortical
column, to where they will migrate radially, and GABAergic
interneurons arise from the ventral parts of the cortex, migrating
tangentially to their target areas. However, the HB-GAM muta-
tion has a more prominent effect on the glutamatergic population,
suggesting that in vivo HB-GAM has more impact on the VZ
stem cells during neurogenesis.
The proliferation phenotype that we have described in this
study could reflect either a shorter stem cell cycle or a decreased
number of cells escaping the stem cell cycle, or a combination of
these two. Our methods cannot, unfortunately, distinguish be-
tween these two alternatives. The number of cells in the M-phase
at E12 was clearly elevated in HB-GAM knockout embryos, and
labeling the whole proliferating cell population with BrdU
revealed an increase of similar magnitude. In both of these
assays, the results could be due to an increase in the total number
of proliferating cells or to a decrease in the cell cycle length,
which increases the probability of an M-phase cell to be captured
by the assay. A more sophisticated method involving prolonged
labeling of the proliferating populations with two separate mitotic
markers (Caviness et al., 1999) should be used to determine the
Fig. 8. Function of HB-GAM in neural stem cell proliferation arrest and in
proteoglycans regulates dynamically the amount of glycosaminoglycan (GAG)-b
growth factors, including FGF-2. FGF-2 binding to its receptor FGFR1 is regulat
proteoglycans like phosphacan and RPTPh/~ . The activity of FGFR1 keeps the ne
stem cells, it is able to displace FGF-2 from the proteoglycan co-receptor(s) and
binding to N-syndecan and RPTPh/~ . Cell signaling upon HB-GAM binding to the
src-kinase pathway (Kinnunen et al., 1998a,b; Maeda and Noda, 1998). ECM, ex
cell cycle length. Because FGF-2 is known to simultaneously
decrease the G1 duration in stem cells and increase the proportion
of proliferative divisions (Lukaszewicz et al., 2002) and HB-
GAM inhibits FGF-2, it seems reasonable that HB-GAM may
affect the same parameters as FGF-2.
Our observations speak for the importance of glycosamino-
glycan (GAG) epitopes in the regulation of neural stem cells. It
has been shown that several proteins associated with neural
growth bind brain-derived GAGs (Herndon et al., 1999). This
raises the questions of whether there are any differences in the
functional specificity between different GAG epitopes and what
the carrier proteins for these GAGs could be. Evidence exists for
the distinction between FGF-2 and FGF-1 (aFGF) binding hep-
aran sulfate epitopes in developing murine brain (Chipperfield et
al., 2002; Joseph et al., 1996; Nurcombe et al., 1993). Syndecan
family members and perlecan are known to interact with FGF-2
and modify its function in several tissue types (Nugent and Iozzo,
2000; Tumova et al., 2000; Whitelock et al., 1996). Also,
chondroitin sulfate proteoglycans like phosphacan have been
shown to enhance growth factor function (Milev et al., 1998b).
The extracellular matrix protein phosphacan and its transmem-
brane form RPTPh/~ (receptor-type protein tyrosine phosphatase
h/~) bind HB-GAM, too, and phosphacan can reverse the
inhibitory effect of HB-GAM in neural stem cells in vitro
(Hienola and Rauvala, unpublished results). Interestingly, low
concentrations of heparin inhibit HB-GAM binding to phospha-
can (Maeda et al., 1996), suggesting a similarity in HB-GAM
neuronal differentiation and migration. A population of transmembrane
inding molecules at the cell surface. Among these molecules are several
ed strongly at least by syndecans and possibly by some chondroitin sulfate
ural stem cells in a self-renewing cell cycle. If HB-GAM is present near the
can thus inhibit FGF-2 activity. HB-GAM also induces cell signaling via
transmembrane proteoglycans is mediated, at least in part, by the cortactin–
tracellular matrix.

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8884
binding carbohydrate epitope(s) found in heparin–heparan sulfate
and phosphacan. Mouse mutants have been created at least for
the HB-GAM receptors syndecan-3 (Kaksonen et al., 2002;
Reizes et al., 2001) and RPTPh/~ (Harroch et al., 2000), and
for perlecan (Arikawa-Hirasawa et al., 1999; Hart et al., 2001).
They offer opportunities to study more closely the specific roles
of these molecules in the germinal zones of the brain.
A wide number of proteins containing TSR domains are
expressed in the developing nervous system in addition to HB-
GAM and midkine, including F-spondin, semaphorins F and G,
and thrombospondins (for review, see Adams and Tucker, 2000).
For example, TSR domains closely resembling those of HB-
GAM are found in F-spondin (Klar et al., 1992; Tzarfaty-Majar et
al., 2001). Very recent studies have suggested that the TSR
sequence motif defines a similar h-structure in different proteins
of the family and that the TSR domains mediate interactions with
heparin–heparan sulfate and the cell surface (Kilpelainen et al.,
2000; Tan et al., 2002). The present results suggest that the TSR
domains of HB-GAM specifically restrict proliferation of neural
stem cells because midkine did not display this effect. Further
studies are however warranted for understanding whether other
TSR proteins play a role in stem cell regulation in neural or
nonneural tissues.
HB-GAM promotes neurite outgrowth and migration through
binding to N-syndecan (syndecan-3) and RPTPh/~ (Imai et al.,
1998; Kinnunen et al., 1996; Maeda and Noda, 1998). The
migration-promoting effect of HB-GAM in neuronal cells
depends on signaling through the cortactin–Src-kinase pathway
(Kinnunen et al., 1998a,b). These findings suggest that HB-GAM
may have a role in the radial migration of new neurons. The
current histological analysis of HB-GAM-deficient mice supports
this view. First, HB-GAM mRNA colocalizes with RC-2 immu-
noreactivity in embryonic cortical slices (Hienola and Rauvala,
unpublished results). Second, in the prenatal phases of cortical
histogenesis, we can detect most of the excess cells in HB-GAM
knockouts in the lower layers of the cortex, for example, the SVZ
at E16, but in the adult animals, the most dramatic density
differences can be seen in the relatively high layers, namely in
II– III and IV. In addition, about 10 days after birth, we can still
see in the cortices of HB-GAM knockouts radially oriented cell
stripes that are no longer detectable in the wild-type animals and
resemble late migrating, clonally related cells (Hienola and
Rauvala, unpublished results). It therefore appears that radial
migration is delayed in the HB-GAM knockout brain. In addition,
we find it interesting that the excess GABAergic neurons in the
HB-GAM knockout cortex are localized almost exclusively in the
higher laminae II– IV. The tangential migration of interneurons
ends in an event when the cells make a decision to migrate
radially to their final destination (Nadarajah et al., 2003). Why an
excess of these neurons should end up in the upper layers in HB-
GAM knockout remains to be solved. We hope to shed more
light on these issues in a separate study characterizing the
migration phenotype in these animals.
This study reveals an important endogenous inhibitor for the very
basic stem cell regulating growth factor, FGF-2. The regulation is
most likely due to competition between FGF-2 and HB-GAM in
binding the specific GAG epitopes at the cell surfaces and in the
extracellular matrix. Fig. 8 summarizes our current view, according
to which HB-GAM binding to cell surface proteoglycans links
proliferation arrest to the start of differentiation and migration of
developing neural cells.
Experimental methods
Cell culture reagents and proteins
All cell culture experiments were done in Dulbecco’s modified
Eagle’s medium (DMEM) with additional penicillin–streptomycin
and L-glutamine. Either bovine serum albumin (BSA, 10 mg/ml) or
fetal calf serum (FCS, 10%) was added to the medium. In neuro-
sphere medium B27 cell culture supplement (Gibco) was used
together with human recombinant FGF-2 (10 ng/ml or approxi-
mately 0.6 nM, Promega) or with FGF-2 and human recombinant
EGF (20 ng/ml or approximately 3 nM, Promega).
Except for FGF-2 and EGF, the other recombinant proteins
were self-produced. Baculovirus expression system was used for
recombinant HB-GAM (Raulo et al., 1992). Recombinant midkine
and HB-GAM’s di-TSR-fragment production in Escherichia coli
and their purification were carried out as described previously
(Kilpelainen et al., 2000). Low-molecular weight heparin (Sigma)
was used at low concentrations (100 ng/ml) to specifically block
HB-GAM binding to heparan sulfates in cell cultures.
Animals and organs
The HB-GAM male knockout mice in 129/SvEv background
were crossbred with 129/SvEv females and the inbred mutant
mouse line was established by sibling breeding. In all of our
studies, littermate wild-type controls were used. Genotyping and
sample preparation for histology of adult brain samples have been
described before (Amet et al., 2001).
Brain samples from P3 and P10 pups were collected by
decapitating the pups and dissecting the brains out on a petri dish
kept on ice. For histological stainings, the brains were cut in two
parts separating the cerebellum from the rostral part of the brain.
The samples were fixed in 4% PFA and 0.5% glutaraldehyde in PBS
overnight.
HB-GAM knockout embryos for histological staining and for
isolation of cortical cells were collected from timed-pregnant
females. The pregnant mice were killed with CO2 and by cervical
dislocation, and the uteri were dissected out and placed in sterile
PBS. Embryos were cut out and either the placenta or the leftover
torso of each embryo was used for genotyping.
Material for cortical and neurosphere cultures was collected
either from HB-GAM mutant or outbred NMRI mouse embryos
aged E12.5–E14.5. The frontal cortical lobes were separated and
transferred to cell culture medium with 10% FCS. The lobes were
dissociated and homogenized by passing the cell suspension
through 20 G needles attached to a sterile syringe several times.
After this, the FCS medium was changed and the cells were left to
recover for 30 min. Cells were collected and washed twice in BSA
medium. When mixed litters of HB-GAM knockout and wild-type
embryos were used, all embryos were handled separately.
Optical dissecting—selector method
Adult brain samples were sectioned as described before (Amet et
al., 2001). From each sectioned sample, a random series was
collected and stained. The sections were photographed with oil-
immersion (Olympus AX50, 60� objective). The random sample
consisted of about 40 sections/brain and the sample area covered
about 2 mm of the cortex in rostral–caudal direction. The sample
volume was located and chosen based on the size and shape of the

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–88 85
dentate gyrus in hippocampal formation. Photocollases of parietal
cortices were divided into nine counting layers (Fig. 1D). Neurons
were identified during counting from hematoxylin-eosin or Nissl-
stained sections according to their nuclear morphology. In the case
of E12.5 embryos, the whole embryo was sectioned in the coronal
direction and the stereological sample volume was chosen from the
rostral tip of the brain to the caudal edge of lateral ventricles. In the
case of embryonic brain samples, the random sample consisted of
20 sections.
The photographs were measured with an image processing
software ImagePro. The optical quantification was based on
selector-method (McMillan and Sorensen, 1992) that allows one
to ignore the sample thickness (Everall et al., 1997). In selector-
method, every morphologically identified neuronal nucleus (based
on size and presence of Nissl bodies) found inside the square-
shaped probe (approximately 500 Am2) or hitting its inclusion lines
was counted and its radius was measured. The relative area (Irel)
occupied by all measured objects on the counting area was then
calculated. The shape of the nuclei was assumed to be sphere-like
and the volume of the objects was calculated based on that
assumption. The average volume (Vmean) of all counted objects
in the counting field was then used with the Irel to calculate the
neuronal density (ND = Irel / Vmean) as a number per cubic
millimeter. The observers were not aware of the genotype of
photographed and measured samples at any point of the analysis.
The measured data were afterwards grouped according to geno-
types for statistical evaluation. Single-factor analysis of variance
(ANOVA) was used to determine the significance of difference
between HB-GAM wild-type and knockout samples.
Identification of the subtypes of cortical neurons
Cortical samples consisting of 5-Am-thick sections from HB-
GAM wild-type and knockouts were stained for GABA and
glutamate receptor 2 immunopositive cells to distinguish between
the populations of interneurons and glutamatergic, pyramidal
neurons, respectively. The mouse anti-GABA (1 Ag/ml, Chemicon)
and mouse anti-GluR2 (1 Ag/ml, Zymed) antibodies were diluted in
PBS with 0.3% Triton-X and 2% BSA and the sections were
incubated in the primary antibody solution overnight in +4jC. Thesections were then incubated with biotinylated anti-mouse IgG
(Zymed) overnight in +4jC. DAB colorimetry was used for
immunodetection. The sections were photographed with oil im-
mersion (Zeiss AxioPlan 2, 63� objective). The average density of
GABA+ and GluR2+ cells in the cortical laminae II– IV, V, and VI
was estimated as described above.
Proliferation in primary cortical cultures
Cortical cultures from HB-GAM knockout and wild-type em-
bryos were plated on polystyrene 96-well microplates, and cells
were plated at 150000/ml density, that is, 15000 per well. HB-
GAM, di-TSR-fragment, or MK was added on knockout cells in the
BSA-medium at 6-nM concentration. BSA medium alone was used
as a control for base level proliferation. Cell proliferation was
measuredwith BrdUCell Proliferation Kit (Boehringer-Mannheim).
Neurosphere assays
Neurospheres were derived from NMRI mouse embryos. Cor-
tical cells were collected at E14 as described above. After recovery
period, the cells were plated on non-coated petri dishes in neuro-
sphere medium. The next day the cells were collected, triturated,
and cultured at 150000/ml cell density in the neurosphere medium.
The grown neurospheres were used in assays after two or three
passages (about a week in culture).
Clonal neurosphere formation
We derived neurospheres from primary forebrain cultures by
using either FGF-2 only or both FGF-2 and EGF as growth factors
in the medium. Spheres derived from FGF-2-stimulated cultures
were used directly in the assays after three passages in culture. The
spheres were triturated and plated on nontreated 96-well plates at a
density of 2–10 cells/well. The clonal growth was followed for 5
days in the presence of FGF-2 (0.6 nM), HB-GAM (6 nM), FGF-2,
and HB-GAM, FGF-2, HB-GAM, and heparin (100 ng/ml), and
FGF-2 and midkine (6 nM), and the number of generated clones
per 40 plated cells was counted. Dose-dependent inhibition by HB-
GAM in clonal growth was studied in cultures derived from
cultures with combined growth factors. The cells were cultured
with both FGF-2 and EGF for three passages. After this the
cultures were split in half and subcultures were continued in the
presence of either FGF-2 or EGF only. The single-growth factor
cultures were continued for extra two passages, and the cells were
triturated and plated as described above. The molar concentration
of HB-GAM varied from 0- to 100-fold with respect to the growth
factor concentration. The minimum number of cells was 100 in
each condition, and the clone formation was followed for 7 days.
Maintenance and differentiation
The experiments were carried on object glasses with eight
separate wells (1 cm2/well, Costar). Forty to 80 neurospheres per
well were plated in neurosphere medium containing FGF-2. The
absolute number of spheres per well was counted after plating.
Spheres were cultured on non-coated object glasses for 3 days in
neurosphere medium without FGF-2, with FGF-2 (0.6 nM), with
FGF-2 and HB-GAM (6 nM), or di-TSR-fragment (6 nM). Heparin
was added to block HB-GAM activity in the solution (100 ng/ml).
Neurospheres remaining morphologically undifferentiated were
counted every day (Fig. 4).
A similar assay was carried on laminin-coated (10 Ag/ml) object
glasses in the presence of either FGF-2 or EGF. In this assay also the
effect of MK (6 nM) in growth factor-stimulated cells was studied.
After 5 days in culture the spreading neurospheres were photo-
graphed. The glasses were fixed with 4% PFA in PBS for 1 h and
stained with anti-nestin (DSHB, University of Iowa, US), anti-h-tubulin (Zymed), and anti-GFAP (Chemicon) antibodies according
to the recommendations from the manufacturers. FITC and TRITC-
conjugated secondary antibodies (Molecular Probes) were used for
detection with fluorescence microscopy.
BrdU labeling and immunohistochemistry in vivo
BrdU injections were given intraperitoneally to timed-pregnant
females 12 days after detection of vaginal plug (embryonic day
12.5). BrdU (50 Ag/g, Sigma) in PBS was injected three times in 2-
h intervals (cumulative dose of 150 Ag/g), and the embryos were
collected 6 h after the first injection. Four wild-type and four
knockout embryos from both age groups were fixed with 4% PFA
in PBS overnight and embedded in paraffin. Thin sections (4 Am)

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8886
were cut, hydrated, and incubated overnight with anti-BrdU ready-
to-use solution (Amersham, UK). Biotinylated secondary antibody
was detected with peroxidase–DAB reaction. Sections were photo-
graphed and BrdU-positive cells were counted with the selector-
method (see above).
Immunostaining for M-phase cells
We used antibodies against phosphorylated histone 3 (rabbit
IgG, Upstate Biotechnology, US) as an M-phase marker to reveal
the number of mitotic cells in E12 embryos at the moment of
preparation. Paraffin sections (5 Am thick) were incubated in 0.5
Ag/ml antibody in PBS with 0.3% Triton-X and 2% BSA
overnight. Biotinylated secondary IgG was used with peroxidase
and DAB for colorimetric immunodetection. The sections were
photographed (Zeiss Axioplan 2, 63� oil objective) and the
density of phosphohistone 3-positive cells was estimated as
described above.
Activation of FGF receptor in neural stem cells
Neural stem cells grown in FGF-2 neurosphere medium were
triturated and plated on laminin for 2 days. The cells were deprived
in growth factor-free medium for 24 h. Cells were stimulated with
FGF-2 (0.6 nM) for 15 min after which the cells were washed once
with cold PBS and lysed in cold RIPA buffer (1% nonidet P40,
0.5% Na-deoxycholate, and 0.1% SDS in PBS) containing protease
inhibitors PMSF (1 mM) and aprotinin (1 Ag/ml), and Na-o-
vanadate (1 mM) as a phosphatase inhibitor. HB-GAM (6 nM)
and di-TSR-fragment (6 nM) were used together with FGF-2 to
examine their effect on receptor activation. Cell lysates were
homogenized with a 20-G needle and a syringe, and the lysates
were boiled for 5 min with SDS containing gel-loading buffer. The
samples were loaded in polyacrylamide gel and transferred to
nitrocellulose membrane. The relative amount of FGF receptor
phosphorylation was determined by immunoblotting the samples
with anti-phospho-FGFR (Cell Signaling Technologies), and the
equality of sample loads was verified by stripping the membrane
and blotting it with anti-FGFR1 antibody (Cell Signaling Technol-
ogies). The blots were visualized with ECL (Amersham Pharma-
cia) on X-ray film, and the relative optical densities were measured
digitally from the scanned images (Quantity One, BioRad). Sam-
ples were also blotted with anti-phospho-tyrosine antibody (Sigma)
to verify changes in the phosphorylation level.
Caspase-3 immunostaining
Polyclonal anti-caspase-3 antibodies (Zymed) were diluted in
buffer with 0.3% Triton-X, applied on paraffin sections or on
methanol-fixed cultured cells on object glasses overnight, and
detected with biotinylated secondary antibodies and peroxidase–
DAB reaction.
Acknowledgments
The authors wish to thank Erja Huttu, Seija Lehto, and Eeva-
Liisa Saarikalle for their excellent technical assistance, and doctors
Sarka Tumova, Marjo Salminen, and Richard Margolis for their
valuable comments, critics, and suggestions. This work has been
supported by the Academy of Finland and the Technical Research
Centre of Finland (Programme of Molecular Neurobiology), and the
Sigrid Juselius Foundation.
References
Adams, J.C., Tucker, R.P., 2000. The thrombospondin type 1 repeat (TSR)
superfamily: diverse proteins with related roles in neuronal develop-
ment. Dev. Dyn. 218, 280–299.
Amet, L.E., Lauri, S.E., Hienola, A., Croll, S.D., Lu, Y., Levorse, J.M.,
Prabhakaran, B., Taira, T., Rauvala, H., Vogt, T.F., 2001. Enhanced
hippocampal long-term potentiation in mice lacking heparin-binding
growth-associated molecule. Mol. Cell. Neurosci. 17, 1014–1024.
Arikawa-Hirasawa, E., Watanabe, H., Takami, H., Hassell, J.R., Yamada,
Y., 1999. Perlecan is essential for cartilage and cephalic development.
Nat. Genet. 23, 354–358.
Burrows, R.C., Wancio, D., Levitt, P., Lillien, L., 1997. Response diversity
and the timing of progenitor cell maturation are regulated by develop-
mental changes in EGFR expression in the cortex. Neuron 19, 251–267.
Carey, D.J., Evans, D.M., Stahl, R.C., Asundi, V.K., Conner, K.J., Garbes,
P., Cizmeci-Smith, G., 1992. Molecular cloning and characterization of
N-syndecan, a novel transmembrane heparan sulfate proteoglycan.
J. Cell Biol. 117, 191–201.
Caviness, V.S.J., Takahashi, T., Nowakowski, R.S., 1995. Numbers, time
and neocortical neuronogenesis: a general developmental and evolu-
tionary model. Trends Neurosci. 18, 379–383.
Caviness Jr., V.S., Takahashi, T., Nowakowski, R.S., 1999. The G1 restric-
tion point as critical regulator of neocortical neuronogenesis. Neuro-
chem. Res. 24, 497–506.
Chipperfield, H., Bedi, K.S., Cool, S.M., Nurcombe, V., 2002. Heparan
sulfates isolated from adult neural progenitor cells can direct phenotypic
maturation. Int. J. Dev. Biol. 46, 661–670.
Dono, R., Texido, G., Dussel, R., Ehmke, H., Zeller, R., 1998. Impaired
cerebral cortex development and blood pressure regulation in FGF-2-
deficient mice. EMBO J. 17, 4213–4225.
Dreyfus, J., Brunet-de Carvalho, N., Duprez, D., Raulais, D., Vigny, M.,
1998. HB-GAM/pleiotrophin but not RIHB/midkine enhances chondro-
genesis in micromass culture. Exp. Cell Res. 241, 171–180.
Everall, I.P., DeTeresa, R., Terry, R., Masliah, E., 1997. Comparison of two
quantitative methods for the evaluation of neuronal number in the
frontal cortex in Alzheimer disease. J. Neuropathol. Exp. Neurol. 56,
1202–1206.
Grutter, M.G., 2000. Caspases: key players in programmed cell death. Curr.
Opin. Struct. Biol. 10, 649–655.
Harroch, S., Palmeri, M., Rosenbluth, J., Custer, A., Okigaki, M., Shrager,
P., Blum, M., Buxbaum, J.D., Schlessinger, J., 2000. No obvious ab-
normality in mice deficient in receptor protein tyrosine phosphatase
beta. Mol. Cell. Biol. 20, 7706–7715.
Hart, M., Li, L., Tokunaga, T., Lindsey, J.R., Hassell, J.R., Snow, A.D.,
Fukuchi, K., 2001. Overproduction of perlecan core protein in cultured
cells and transgenic mice. J. Pathol. 194, 262–269.
Herndon, M.E., Stipp, C.S., Lander, A.D., 1999. Interactions of neural
glycosaminoglycans and proteoglycans with protein ligands: assess-
ment of selectivity, heterogeneity and the participation of core proteins
in binding. Glycobiology 9, 143–155.
Hulspas, R., Tiarks, C., Reilly, J., Hsieh, C.-C., Recht, L., Quesenberry,
P.J., 1997. In vitro cell density-dependent clonal growth of EGF-respon-
sive murine neural progenitor cells under serum-free conditions. Exp.
Neurol. 148, 147–156.
Imai, S., Kaksonen, M., Raulo, E., Kinnunen, T., Fages, C., Meng, X.,
Lakso, M., Rauvala, H., 1998. Osteoblast recruitment and bone forma-
tion enhanced by cell matrix-associated heparin-binding growth-associ-
ated molecule (HB-GAM). J. Cell Biol. 143, 1113–1128.
Joseph, S.J., Ford, M.D., Barth, C., Portbury, S., Bartlett, P.F., Nurcombe,
V., Greferath, U., 1996. A proteoglycan that activates fibroblast growth
factors during early neuronal development is a perlecan variant. Devel-
opment 122, 3443–3452.

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–88 87
Junier, M.P., 2000. What role(s) for TGFalpha in the central nervous sys-
tem? Prog. Neurobiol. 62, 443–473.
Kaksonen, M., Pavlov, I., Voikar, V., Lauri, S.E., Hienola, A., Riekki, R.,
Lakso, M., Taira, T., Rauvala, H., 2002. Syndecan-3 deficient mice
exhibit enhanced LTP and impaired hippocampus-dependent memory.
Mol. Cell. Neurosci. 21, 158–172.
Kilpelainen, I., Kaksonen, M., Avikainen, H., Fath, M., Linhardt, R.J.,
Raulo, E., Rauvala, H., 2000. Heparin-binding growth-associated mol-
ecule contains two heparin-binding beta-sheet domains that are homol-
ogous to the thrombospondin type I repeat. J. Biol. Chem. 275,
13564–13570.
Kinnunen, T., Raulo, E., Nolo, R., Maccarana, M., Lindahl, U., Rauvala,
H., 1996. Neurite outgrowth in brain neurons induced by heparin-bind-
ing growth-associated molecule (HB-GAM) depends on the specific
interaction of HB-GAM with heparan sulfate at the cell surface. J. Biol.
Chem. 271, 2243–2248.
Kinnunen, A., Kinnunen, T., Kaksonen, M., Nolo, R., Panula, P., Rauvala,
H., 1998a. N-syndecan and HB-GAM (heparin-binding growth-associ-
ated molecule) associate with the early axonal tracts in the rat brain.
Eur. J. Neurosci. 10, 635–648.
Kinnunen, T., Kaksonen, M., Saarinen, J., Kalkkinen, N., Peng, H.B.,
Rauvala, H., 1998b. Cortactin–Src kinase signaling pathway is in-
volved in N-syndecan-dependent neurite outgrowth. J. Biol. Chem.
273, 10702–10708.
Kinnunen, A., Niemi, M., Kinnunen, T., Kaksonen, M., Nolo, R., Rauvala,
H., 1999. Heparan sulphate and HB-GAM (heparin-binding growth-
associated molecule) in the development of the thalamocortical pathway
of rat brain. Eur. J. Neurosci. 11, 491–502.
Klar, A., Baldassare, M., Jessell, T.M., 1992. F-spondin: a gene expressed
at high levels in the floor plate encodes a secreted protein that promotes
neural cell adhesion and neurite extension. Cell 69, 95–110.
Lauri, S.E., Taira, T., Kaila, K., Rauvala, H., 1996. Activity-induced en-
hancement of HB-GAM expression in rat hippocampal slices. Neuro-
Report 7, 1670–1674.
Lauri, S.E., Rauvala, H., Kaila, K., Taira, T., 1998. Effect of heparin-binding
growth-associated molecule (HB-GAM) on synaptic transmission and
early LTP in rat hippocampal slices. Eur. J. Neurosci. 10, 188–194.
Lauri, S.E., Kaukinen, S., Kinnunen, T., Ylinen, A., Imai, S., Kaila, K.,
Taira, T., Rauvala, H., 1999. Regulatory role and molecular interactions
of a cell-surface heparan sulfate proteoglycan (N-syndecan) in hippo-
campal long-term potentiation. J. Neurosci. 19, 1226–1235.
Lukaszewicz, A., Savatier, P., Cortay, V., Kennedy, H., Dehay, C., 2002.
Contrasting effects of basic fibroblast growth factor and neurotrophin 3
on cell cycle kinetics of mouse cortical stem cells. J. Neurosci. 22,
6610–6622.
Maeda, N., Noda, M., 1998. Involvement of receptor-like protein tyrosine
phosphatase zeta/RPTPbeta and its ligand pleiotrophin/heparin-binding
growth-associated molecule (HB-GAM) in neuronal migration. J. Cell
Biol. 142, 203–216.
Maeda, N., Nishiwaki, T., Shintani, T., Hamanaka, H., Noda, M., 1996.
6B4 proteoglycan/phosphacan, an extracellular variant of receptor-like
protein-tyrosine phosphatase zeta/RPTPbeta, binds pleiotrophin/hepa-
rin-binding growth-associated molecule (HB-GAM). J. Biol. Chem.
271, 21446–21452.
Matsubara, S., Tomomura, M., Kadomatsu, K., Muramatsu, T., 1990.
Structure of a retinoic acid-responsive gene, MK, which is transiently
activated during the differentiation of embryonal carcinoma cells and
the mid-gestation period of mouse embryogenesis. J. Biol. Chem. 265,
9441–9443.
McMillan, A.M., Sorensen, F.B., 1992. The efficient and unbiased estima-
tion of nuclear size variability using the ‘selector’. J. Microsc. 165,
433–437.
Merenmies, J., Rauvala, H., 1990. Molecular cloning of the 18-kDa
growth-associated protein of developing brain. J. Biol. Chem. 265,
16721–16724.
Milev, P., Chiba, A., Haring, M., Rauvala, H., Schachner, M., Ranscht, B.,
Margolis, R.K., Margolis, R.U., 1998a. High affinity binding and over-
lapping localization of neurocan and phosphacan/protein-tyrosine phos-
phatase-zeta/beta with tenascin-R, amphoterin, and the heparin-binding
growth-associated molecule. J. Biol. Chem. 273, 6998–7005.
Milev, P., Monnerie, H., Popp, S., Margolis, R.K., Margolis, R.U., 1998b.
The core protein of the chondroitin sulfate proteoglycan phosphacan is a
high-affinity ligand of fibroblast growth factor-2 and potentiates its
mitogenic activity. J. Biol. Chem. 273, 21439–21442.
Mitsiadis, T.A., Salmivirta, M., Muramatsu, T., Muramatsu, H., Rauvala,
H., Lehtonen, E., Jalkanen, M., Thesleff, I., 1995. Expression of the
heparin-binding cytokines, midkine (MK) and HB-GAM (pleiotro-
phin) is associated with epithelial–mesenchymal interactions during
fetal development and organogenesis. Development 121, 37–51.
Nadarajah, B., Alifragis, P., Wong, R.O., Parnavelas, J.G., 2003. Neuronal
migration in the developing cerebral cortex: observations based on real-
time imaging. Cereb. Cortex 13, 607–611.
Nugent, M.A., Iozzo, R.V., 2000. Fibroblast growth factor-2. Int. J. Bio-
chem. Cell Biol. 32, 115–120.
Nurcombe, V., Fraser, N., Herlaar, E., Heath, J.K., 1992. MK: a pluripo-
tential embryonic stem-cell-derived neuroregulatory factor. Develop-
ment 116, 1175–1183.
Nurcombe, V., Ford, M.D., Wildschut, J.A., Bartlett, P.F., 1993. Develop-
mental regulation of neural response to FGF-1 and FGF-2 by heparan
sulfate proteoglycan. Science 260, 103–106.
Pavlov, I., Voikar, V., Kaksonen, M., Lauri, S.E., Hienola, A., Taira, T.,
Rauvala, H., 2002. Role of heparin-binding growth-associated molecule
(HB-GAM) in hippocampal LTP and spatial learning revealed by stud-
ies on overexpressing and knockout mice. Mol. Cell. Neurosci. 20,
330–342.
Raballo, R., Rhee, J., Lyn-Cook, R., Leckman, J.F., Schwartz, M.L., Vac-
carino, F.M., 2000. Basic fibroblast growth factor (Fgf2) is necessary
for cell proliferation and neurogenesis in the developing cerebral cortex.
J. Neurosci. 20, 5012–5023.
Rakic, P., 1995. A small step for the cell, a giant leap for mankind: a
hypothesis of neocortical expansion during evolution. Trends Neurosci.
18, 383–388.
Raulo, E., Julkunen, I., Merenmies, J., Pihlaskari, R., Rauvala, H., 1992.
Secretion and biological activities of heparin-binding growth-associat-
ed molecule. Neurite outgrowth-promoting and mitogenic actions of
the recombinant and tissue-derived protein. J. Biol. Chem. 267,
11408–11416.
Raulo, E., Chernousov, M.A., Carey, D.J., Nolo, R., Rauvala, H., 1994.
Isolation of a neuronal cell surface receptor of heparin binding growth-
associated molecule (HB-GAM). Identification as N-syndecan (synde-
can-3). J. Biol. Chem. 269, 12999–13004.
Rauvala, H., 1989. An 18-kd heparin-binding protein of developing brain
that is distinct from fibroblast growth factors. EMBO J. 8, 2933–2941.
Rauvala, H., Peng, H.B., 1997. HB-GAM (heparin-binding growth-associ-
ated molecule) and heparin-type glycans in the development and plas-
ticity of neuron-target contacts. Prog. Neurobiol. 52, 127–144.
Rauvala, H., Vanhala, A., Castren, E., Nolo, R., Raulo, E., Merenmies, J.,
Panula, P., 1994. Expression of HB-GAM (heparin-binding growth-
associated molecules) in the pathways of developing axonal processes
in vivo and neurite outgrowth in vitro induced by HB-GAM. Brain Res.
Dev. Brain Res. 79, 157–176.
Rauvala, H., Huttunen, H.J., Fages, C., Kaksonen, M., Kinnunen, T., Imai,
S., Raulo, E., Kilpelainen, I., 2000. Heparin-binding proteins HB-GAM
(pleiotrophin) and amphoterin in the regulation of cell motility. Matrix
Biol. 19, 377–387.
Reizes, O., Lincecum, J., Wang, Z., Goldberger, O., Huang, L., Kaksonen,
M., Ahima, R., Hinkes, M.T., Barsh, G.S., Rauvala, H., Bernfield, M.,
2001. Transgenic expression of syndecan-1 uncovers a physiological
control of feeding behavior by syndecan-3. Cell 106, 105–116.
Seaberg, R.M., van der Kooy, D., 2003. Stem and progenitor cells: the
premature desertion of rigorous definitions. Trends Neurosci. 26,
125–131.
Silos-Santiago, I., Yeh, H.J., Gurrieri, M.A., Guillerman, R.P., Li, Y.S.,
Wolf, J., Snider, W., Deuel, T.F., 1996. Localization of pleiotrophin

A. Hienola et al. / Mol. Cell. Neurosci. 26 (2004) 75–8888
and its mRNA in subpopulations of neurons and their corresponding
axonal tracts suggests important roles in neural–glial interactions dur-
ing development and in maturity. J. Neurobiol. 31, 283–296.
Szabat, E., Rauvala, H., 1996. Role of HB-GAM (heparin-binding growth-
associated molecule) in proliferation arrest in cells of the developing rat
limb and its expression in the differentiating neuromuscular system.
Dev. Biol. 178, 77–89.
Takahashi, T., Nowakowski, R.S., Caviness, V.S.J., 1994. Mode of cell
proliferation in the developing mouse neocortex. Proc. Natl. Acad.
Sci. U. S. A. 91, 375–379.
Takahashi, T., Nowakowski, R.S., Caviness, V.S.J., 1996. The leaving of Q
fraction of the murine cerebral proliferative epithelium: a general model
of neocortical neuronogenesis. J. Neurosci. 16, 6183–6196.
Takahashi, T., Nowakowski, R.S., Caviness, V.S.J., 1997. The mathematics
of neocortical neuronogenesis. Dev. Neurosci. 19, 17–22.
Tan, K., Duquette, M., Liu, J., Dong, Y., Zhang, R., Joachimiak, A., Law-
ler, J., Wang, J., 2002. Crystal structure of the TSP-1 type 1 repeats: a
novel layered fold and its biological implication. J. Cell Biol. 159,
373–382.
Tropepe, V., Sibilia, M., Ciruna, B.G., Rossant, J., Wagner, E.F., van der
Kooy, D., 1999. Distinct neural stem cells proliferate in response to
EGF and FGF in the developing mouse telencephalon. Dev. Biol. 208,
166–188.
Tumova, S., Woods, A., Couchman, J.R., 2000. Heparan sulfate proteogly-
cans on the cell surface: versatile coordinators of cellular functions. Int.
J. Biochem. Cell Biol. 32, 269–288.
Tzarfaty-Majar, V., Lopez-Alemany, R., Feinstein, Y., Gombau, L., Gold-
shmidt, O., Soriano, E., Munoz-Canoves, P., Klar, A., 2001. Plasmin-
mediated release of the guidance molecule F-spondin from the extra-
cellular matrix. J. Biol. Chem. 276, 28233–28241.
Whitelock, J.M., Murdoch, A.D., Iozzo, R.V., Underwood, P.A., 1996. The
degradation of human endothelial cell-derived perlecan and release of
bound basic fibroblast growth factor by stromelysin, collagenase, plas-
min, and heparanases. J. Biol. Chem. 271, 10079–10086.