hat element population genetics in anopheles gambiae s.l. in mozambique
TRANSCRIPT

UNCO
RRECTEDPROO
F
2 hAT element population genetics in Anopheles gambiae s.l. in Mozambique
34 David A. O’Brochta1, Ramanand A. Subramanian1, Jamison Orsetti1, Edward Peckham1,5 Nicole Nolan1, Peter Arensburger2, Peter W. Atkinson2 & Derek J. Charlwood3
6 1Center for Biosystems ResearchUniversity of Maryland Biotechnology Institute, Building 036/Room 5115,7 College Park, MD 20742, USA (Phone: 301-405-7680; Fax: 301-314-9075; E-mail: [email protected] d.edu); 2Department of EntomologyUniversity of California, 900 University Avenue, Riverside, CA 92521-9 0001, USA; 3Danish Bilharziasis Laboratory, 1 Jaegersbor Alle, 2920, Charlottenlund, DK Denmark
10 Received 17 January 2005 Accepted 26 September 2005
11 Key words: Anopheles arabiensis, Anopheles gambiae, Anopheles merus, Hermes, Herves, hobo, transgenic12 mosquitoes, transposable elements
1314 Abstract
15 Herves is a functional Class II transposable element in Anopheles gambiae belonging to the hAT super-16 family of elements. Class II transposable elements are used as gene vectors in this species and are also being17 considered as genetic drive agents for spreading desirable genes through natural populations as part of an18 effort to control malaria transmission. In this study, Herves was investigated in populations of Anophe-19 les gambiae s.s., Anopheles arabiensis and Anopheles merus in Mozambique over a period of 2 years. The20 copy number of Herves within these three species was approximately 5 copies per diploid genome and did21 not differ among species or between years. Based on the insertion-site occupancy–frequency distribution22 and existing models of transposable element dynamics, Herves appears to be transpositionally active23 currently or, at least recently, in all species tested. Ninety-five percent of the individuals within the pop-24 ulations of the three species tested contained intact elements with complete Herves transposase genes and25 this is consistent with the idea that these elements are currently active.26
2728 Introduction
29 Transposable elements are significant components30 of eukaryotic genomes andplay an important role in31 genome evolution (Kidwell, 2002). They have also32 been exploited as functional genomics tools in some33 organismswith themost outstanding example being34 thediverse applications inDrosophila melanogaster,35 including transposable element-mediated gene36 insertion, gene replacement, chromosome rear-37 rangements, gene identification and isolation38 (Ryder & Russell, 2003; Miller & Capy, 2004).39 Similar applications of transposable elements are40 being developed for other insects and some novel41 applications have been discussed in relation to42 mosquitoes. A unique approach to combating43 mosquito-transmitted diseases, in particular ma-
44laria, has been proposed involving transgenic mod-45ifications of entire populations of wild mosquitoes46(Collins et al., 1986;Miller et al., 1987). Transgenic47modifications of particular interest will result in48mosquitoes such as Anopheles gambiae s.s., the49major human malaria vector in Africa, becoming50resistant or ‘‘refractory’’ to Plasmodium infection51and thereby interrupting the obligatory route of52transmission of this parasite (Ito et al., 2002;53Moreira et al., 2002; Kim et al., 2004). Efforts to54develop genetic modification technologies for mos-55quitoes have focused on the manipulation of trans-56posable elements and moderate success has been57achieved (Atkinson, Pinkerton&O’Brochta, 2001).58Modifying entire populations of mosquitoes, how-59ever, will require that the introduced transgenes be60present in essentially all individuals of that popula-
Genetica (2005) 00:1–14 � Springer 2005DOI 10.1007/s10709-005-3535-4
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4
AUTHOR’S
PROOF!
PDF-OUTPUT

UNCO
RRECTEDPROO
F
61 tion (Boete & Koella, 2002). Thus, delivering and62 spreading transmission-blocking transgenes is a63 major challenge and one possible solution involves64 the unprecedented use of transposable elements as65 genetic drive or gene-spreading agents (Ribeiro &66 Kidwell, 1994).67 Based largely on natural history studies of68 transposable elements and to a lesser extent on69 empirical studies, it has been shown that trans-70 posable elements can be invasive. That is, elements71 can be transferred between species and once pres-72 ent in a new host species they can actively trans-73 pose under some conditions. The periodic74 ‘‘escape’’ from one species and ‘‘invasion’’ of75 another by transposable elements has been76 suggested to be of major importance to the77 evolution of transposable elements (Lohe et al.,78 1995; Hartl, Lohe & Lozovskaya, 1997; Jordan &79 McDonald, 1998; Silva & Kidwell, 2000; Pinsker80 et al., 2001). Active transposition can result in an81 increase in the element’s copy number and its82 rapid transmission to other members of the83 species. In some cases, for example the invasion of84 D. melanogaster by P elements from D. willistoni,85 spread can be rapid and complete, involving86 essentially all insects within the species over the87 course of decades (Anxolabehere, Kidwell &88 Periquet, 1988; Anxolabehere et al., 1990). Other89 notable examples of element invasions and spread90 in D. melanogaster involve the hobo and I elements91 (Bucheton et al., 1992; Simmons, 1992). In addi-92 tion, the copia element appears to have been93 horizontally transferred from D. melanogaster to94 D. willistoni (Jordan, Matyunina & McDonald,95 1999), and the Penelope element has recently96 invaded D. virilis (Evgen’ev et al., 2000). Various97 mariner-like elements have been detected that98 appear to have crossed a wide range of phyloge-99 netic boundaries (Robertson, 2002). More recent
100 findings on intraspecific sequence diversity of101 transposable elements have been used by some to102 suggest that there have been frequent horizontal103 transfers of transposable elements to Drosophila104 (Sanchez-Gracia, Maside & Charlesworth, 2005).105 Over time the copy number of individual ele-106 ments within element families are thought to reach107 an equilibrium as a result of the balancing of108 forces that result in an increase in element copy109 number (transposition) against those that reduce110 element abundance within a genome (excision and111 selection) (Langley, Brookfield & Kaplan, 1983;
112Charlesworth & Langley, 1989). In addition,113repressors and epigenetic factors can also evolve to114regulate element movement and, consequently,115copy number. Our understanding of the popula-116tion dynamics of transposable elements is based117largely on the study of Class I transposable ele-118ments and a few Class II elements within natural119populations of D. melanogaster and related species120of Drosophila (Charlesworth & Langley, 1990;121Biemont et al., 1994; Nuzhdin, 1999). Studies of122the population dynamics of transposable elements123in non-drosophilid insects have been relatively few124and these too have focused mainly on Class I125elements (Bensaadi-Merchaermek, Salvado &126Mouches, 1994; Tu & Hill, 1999; Rohr et al., 2002;127Biedler et al., 2003; Biedler & Tu, 2003; Zampici-128nini et al., 2004; della Torre, Tu & Petrarca, 2005)129with a few exceptions in which Class II elements130have been studied (Torti et al., 1994; Grossman131et al., 1999; Feschotte & Mouches, 2000; Shao, Qi132& Tu, 2001).133Given the putative role of transposable ele-134ments in the study of mosquitoes and the interest135in using them as a tool in controlling mosquito136vectoral capacity, it is important to investigate the137population genetics of functional, active trans-138posable elements in An. gambiae and other mos-139quitoes being considered as targets for transgenic140population manipulation. Understanding the de-141gree to which transposable element dynamics in142An. gambiae parallels that described in143D. melanogaster will influence decisions about how144or if transgenes could be spread using transposable145elements. Here, we report the results of a study of146the Herves transposable element in a population of147An. gambiae s.l. in Mozambique. This study is part148of a larger effort to characterize the origins and149distribution of the element within African mos-150quitoes and to elucidate historical and contempo-151rary patterns of movement of the element within152populations. These data are relevant to contem-153porary efforts to evaluate the feasibility of using154transposable elements as transmission-blocking,155gene-spreading agents and to predict the fate of156released elements.157Herves, is a member of the hobo, Ac, Tam3158(hAT) family of Class II transposable elements and159was discovered in and isolated originally from160An. gambiae s.s. (Arensburger et al., 2005). hAT161elements are widespread in nature and are found in162plants, insects and vertebrates including humans
2
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
163 (Kunze & Weil, 2002; Robertson, 2002). Herves164 was discovered during a bioinformatics-based165 search of the An. gambiae genome sequence and166 was isolated from the genome of the laboratory167 strain, RSP (Arensburger et al., 2005). Herves is168 3.7 kb in length with 11 bp imperfect inverted169 terminal repeats (ITR) and a 603 amino acid170 transposase open reading frame. The element is171 most similar to the hopper element from Bactro-172 cera tryoni (family: Tephritidae) (30% identity173 and 49% similarity of the transposase amino acid174 sequences) and more distantly related to the Her-175 mes element from Musca domestica (family:176 Muscidae) (21% identity and 37% similarity).177 Like Hermes, Herves is a functional transposable178 element and an effective gene vector in179 D. melanogaster although its complete host range180 has not been determined (Arensburger et al.,181 2005). In this study, we measured the abundance182 and site-occupancy frequency distribution of183 Herves in natural populations of An. gambiae s.s.,184 An. merus, and An. arabiensis in the southern,185 coastal village of Furvela, Mozambique using a186 PCR-based approach. From these data we esti-187 mated rates of transposition and compared these188 data to similar datasets from other organisms.
189 Material and methods
190 Collection site
191 An. gambiae s.l. were collected in light traps and192 while resting inside dwellings during November193 2001 and 2003 in the village of Furvela, Mozam-194 bique. Furvela is located 650 km north of Maputo195 and 7 km south of Morrumbene in the province of196 Inhambane, on the main Maputo-Beira highway197 (EN1). Collected female An. gambiae s.l. were198 placed in silica-gel filled tubes and stored at199 ambient temperature until arrival in the laboratory200 at which time they were stored at )80 �C until201 processed.
202 DNA isolation
203 Genomic DNA was isolated from individual mos-204 quitoes using a modified version of a protocol de-205 signed for isolating genomic DNA from individual206 D. melanogaster (Protocol 48 in (Ashburner, 1989).
207Modifications of the protocol included omitting208the phenol/chloroform extraction step and resu-209spending the final nucleic acid pellet in 100 ll of210sterile water.
211Species identification
212Species identification was performed on 1 ll of213genomic DNA solution using the method of Scott,214Brogdon and Collins (1993). Only samples yielding215unambiguous results were used for further analy-216sis. An. gambiae s.s. in many locations has a217complex population structure and extensive chro-218mosomal inversion polymorphisms. This com-219plexity is manifested largely in West Africa. In220East Africa, only the S-form is found characterized221by the 2Rb and 2La polymorphisms. Character-222ization of the form of An. gambiae was unneces-223sary and subsequent analysis is not expected to224confounded by undetected population substruc-225ture (della Torre, Tu & Petrarca, 2005).
226Transposable element display
227Transposable element display is a PCR-based228DNA fingerprinting method that is similar to the229Amplified Fragment Length Polymorphism230(AFLP) method (Vos et al., 1995) however, it231uses a primer specific to the transposable element232of interest and primer specific to the adapters233that are attached to fragments of digested234genomic DNA, to display a subset of genomic235DNA fragments containing the ITR of the ele-236ment and flanking genomic DNA sequences237(Van den Broeck et al., 1998; Casa et al., 2000).238The transposon display method permits the239simultaneous detection of many copies of an240element family in individual insects. The proto-241col used in this study was a modification of that242described by Guimond et al. (2003) and was243designed to display genomic DNA fragments244containing the left ITR of Herves and a variable245amount of flanking genomic DNA. A similar246method was described by Biedler & Tu (2003).247One or two microliters of genomic DNA248(approximately 200 ng) were digested for 2 h in a249volume of 40 ll at 37 �C with 4 units of the250restriction endonucleases MseI using conditions251recommended by the manufacturer (New England252Biolabs). Sixty picomoles of adapters were ligated
3
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
253 to digested genomic DNA by adding 10 ll of the254 appropriate 1� restriction enzyme buffer contain-255 ing 5 mM ATP, 50 mM DTT (dithiothreitol),256 10 lg BSA (bovine serum albumin), 4 units of257 MseI, 1 Weiss unit of T4 DNA ligase and incu-258 bated at 37 �C overnight. The adapters consisted259 of a duplex of oligonucleotides MseIa (5¢-GAC260 GAT GAG TCC TGA G-3¢) and MseIb (5¢-TAC261 TCA GGA CTC AT-3¢). The design of the262 adapters and the restriction/ligation reaction con-263 ditions result in the efficient creation of only264 monomeric MseI-cut genomic DNA fragments265 with terminal adapters.266 Five microliters of the restriction/ligation267 reaction were used as the template in a poly-268 merase chain reaction (‘‘preselective reaction’’)269 performed in a 50 ll reaction volume containing270 1� PCR Buffer II (Applied Biosystems), 0.2 mM271 dNTPs (an equimolar mixture of dATP, dTTP,272 dCTP, dGTP), 2.5 mM MgCl2, 1 unit AmpliT-273 aq� DNA polymerase (Applied Biosystems), and274 24 pmoles of primer MspIa and primer Her-275 vTEDAL1 (5¢-ATT TCG ACG GGT TCC TAC276 C-3¢). HervTEDAL1 is a Herves-specific primer277 that anneals to sequences approximately 150 bp278 from the 5¢ end of the element. The DNA279 polymerase was added as a complex with Taq-280 Start� Antibody (ClonTech) as described by the281 manufacturer for the purposes of ‘‘hot-starting’’282 the reaction. Reactions were preformed in a283 Perkin-Elmer 9600 thermocycler using the fol-284 lowing parameters: 95 �C/3 mins followed by 25285 cycles of 95 �C/15 s, 60 �C/30 s, 72 �C/1.0 min286 and a final cycle of 72 �C/5 min. A second PCR287 was performed (‘‘selective reaction’’) using 5 ll288 of the preselective PCR products as template in289 a 20 ll reaction containing 1� PCR Buffer II,290 0.2 mM dNTPs, 2.5 mM MgCl2, 1 unit Amp-291 liTaq� DNA polymerase (bound to TaqStart�
292 Antibody (ClonTech) as above), 9 pmoles each293 of primers MspIa and Cy5�-labeled HervTE-294 DAL2 (5¢-GTT GAT TAG ATG AAC GTA295 GG-3¢). The Cy5�-labeled primers were purified296 by HPLC prior to their use. HervTEDAL2 an-297 neals to sequences approximately 80 bp from the298 left end of the element. The reaction was initi-299 ated with a single cycle at 95 �C for 3 min300 followed by 5 cycles in which the annealing301 temperature was decreased 1 �C after each cycle302 with the first of these cycles being 95 �C/15 s,303 60 �C/30 s, 72 �C/1.0 min. Following these 5
304cycles of ‘‘touchdown’’ PCR 25 cycles were per-305formed at 95 �C/15 s, 60 �C /30 s, 72 �C/1.0 min306with a final cycle of 72 �C/5 min.307Five microliters of the selective PCR products308were mixed with 5 ll of loading buffer (95%309deionized formamide, 10 mM EDTA) heated to31095 �C for 5 min, cooled quickly on ice and 6 ll311were loaded on a 6% polyacrylamide gel (19:1312acrylamide:bisacrylamide) containing 6.7 M urea313in 1� TBE buffer (90 mM Tris–borate, 2 mM314EDTA). ALFExpress�Sizer�50-500 (Amersham/315Pharmacia) was used as a size standard. Electro-316phoresis was performed at 70W (constant) for3172.5 h at which time the gel was transferred to3183MM filter paper and dried. The dried gel was319scanned on a STORM 860 phosphoimager320(Molecular Dynamics).321Two variations of the method were also devel-322oped thatwere identical to that described above, but323involved the use of a different restriction endonu-324clease and primers. In both cases, the genomicDNA325is digested with HhaI and adapters consisting of a326duplex of HhaIa (5¢-GAT GAG TCC TGA GTA327GC-3¢) and HhaIb (5¢-TAC TCA GGA CTC ATC328AAG-3¢) were added as described above. Pre-329selective PCRs were performed using the primers330HhaIa and HervTEDAL1 or HervTEDAR1 (5¢-331ATG CAT CAC TTA ATA ACA TAA TTA GG-3323¢) depending on whether the right or left ITRs are333to be displayed. Reaction conditions and cycling334parameters are as described above. Selective PCRs335were performed using the primers HhaIa and Cy5�-336labeled HervTEDAL2 or HervTEDAR2 (5¢-GTT337GAT TAG ATG AAC GTA GG-3¢) using the338conditions and cycling parameters described above.
339Transposase open reading frame detection
340The presence of intact Herves transposase-coding341regions was inferred from the detection of 2100 bp342PCR products using primers that flanked the open343reading frame of the element. One microliter of344genomic DNA was used as template in a 50 ll345reaction containing 1� ThermalAce� (Invitrogen),3460.2 mM dNTPs (an equimolar mixture of dATP,347dTTP, dCTP, dGTP), 2.5 mM MgCl2, 2 units348ThermalAce� DNA polymerase (Invitrogen), and34924 pmoles of primer 1372f (5¢-CCA CAA ATT350GAT CTA CGC TCC-3¢) and primer 3469r351(5¢-GAT GCA TCT ATT ATG ATT AAG GC-3523¢). Amplification reactions were preformed under
4
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
353 the following conditions:95 �C/3 min followed by354 30 cycles of 95 �C/30 s, 48 �C/30 s, 72 �C/3.0 min355 and a final cycle of 72 �C/10 min. Reaction prod-356 ucts were fractionated on a 1% agarose gel.357 Samples that failed to produce a detectable prod-358 uct following one round of PCR were used as359 templates (5 ll) in a second PCR under the same360 conditions described above but with primers 1407f361 (5¢-GATCAAAGGTAACATTAGTCTTG-3¢)362 and 3294r (5¢-CCA TGT TAC AAA TTT TGC363 AAC G-3¢) and rechecked on a 1% agarose gel.364 Samples failing to yield a detectable product after365 the second reaction were assessed for template366 quality by amplifying an actin gene fragment using367 the primers actinf (5¢-ATT AAT GAG AAT CTG368 TGC TAC GTC-3¢) and actinr (5¢-CAT ACG369 ATC AGC AAT ACC TGGG-3¢). Samples failing370 to yield an actin gene fragment were excluded from371 the analysis.
372 Data analysis
373 Images of the scanned transposable element dis-374 play gels were printed and bands were assigned a375 molecular weight based on their mobility relative376 to the size markers using a standard curve devel-377 oped for each gel. All samples of a species collected378 during a single year were analyzed on the same gel,379 permitting a single standard molecular weight380 curve to be used to score each dataset. Each set of381 samples (genomic DNA) was processed for trans-382 posable element display and analyzed three times.383 For all analyses, bands of equal molecular weight384 observed in a species collected during a single year385 were scored as being identical. Bands of differing386 sizes in a species collected during a single year were387 scored as being independent events. These388 assumptions have been tested by isolating, ream-389 plifying and sequencing selected bands and found390 to hold without exception (Guimond et al., 2003).391 Estimations of element activity weremade based392 on the models of Charlesworth and Charlesworth393 (1983) andLangley, Brookfield andKaplan, (1983).394 Charlesworth and Charlesworth (1983) proposed395 that the probability distribution of element fre-396 quencies per site, for any family of elements at397 equilibrium within a population, i.e. with a stable398 copy number, was approximated by a beta distri-399 bution and determined by two parameters, a and b400 [see equation 11, Charlesworth and Charlesworth401 (1983)]. The parameter a (a ¼ 4Neun̂=ðT � n̂))
402reflects the effects of drift and repeated transposi-403tions into a site while b (b ¼ 4Neðvþ sÞ) reflects the404effects of forces that tend to remove elements from a405population (drift, excision and selection)whereNe is406the effective population size, u is the rate of trans-407position, T is the number of possible insertion sites,408n̂ is the copy number at equilibrium, m is the rate of409excision and s is selection against insertions. Be-410cause the copy-number is at equilibrium b also411reflects the forces that tend to add elements to the412population (replicative transposition). Langley,413Brookfield and Kaplan (1983) proposed a related414model but assumed that the possible insertion sites415(T) within a genome were infinite, making a=0 and416resulting in a simplified model with a single417parameter (h).418Parameter estimation methods of Charlesworth419and Charlesworth (1983) and Langley, Kaplan420and Brookfield (1983) 1rely on estimates of element421frequencies and copy numbers per haploid gen-422ome. Transposable element display however,423examines diploid genomes and cannot distinguish424between an element in a heterozygous versus a425homozygous condition. To obtain estimates of426element frequencies and copy numbers per haploid427genome we assumed Hardy–Weinberg equilibrium428and followed Wright et al. (2001) where the mean429number of elements per haploid genome, 1
2 n̂, was:430
1
2n̂ ¼
Xj
1�ffiffiffiffiffiffiffiffiffiffiffiffi1� bj
m
rð4Þ
432432433j corresponds to sites occupied by Herves, bj is the434number of transposable element display bands435observed at site j and m is the number of individ-436uals analyzed in the dataset. Thus, the haploid437copy number was simply the sum of the estimated438haploid element frequencies for all sites. Parame-439ters a and b were estimated as described by Wright440et al. (2001) while h was estimated as described by441Langley, Kapland and Brookfield (1983).
442Results
443TE display
444Transposable element display has been found to be445a reliable and specific method to visualize inser-446tions of mobile elements in insects (Biedler et al.,
5
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
447 2003; Guimond et al., 2003; Zampicinini et al.,448 2004). Although Zampicinini et al. (2004) claimed449 that transposable element display in their hands450 highly underestimated (approximately 10-fold) the451 copy number of the NLRCth1 element of Chir-452 onomus thummi our results suggest that underes-453 timation should be minimal. Because PCR454 extension times were only 1 min genomic DNA455 fragments larger than 1 kb are likely to be under-456 represented, because the rate of extension by Taq457 polymerase is about 1 kb per minute. However,458 very little of the genome of An. gambiae is ex-459 pected to be present on fragments 1 kb or larger460 following digestion with MseI. The AT content of461 the An. gambiae genome is 64.8% and MseI sites462 (TTAA) are therefore expected, on average, every463 91 bases (1/0.3244) (Holt et al., 2002). Conse-464 quently, only 0.001% of the MseI fragments are465 expected to be 1 kb or more in length. Therefore,466 while the transposable element display procedure467 tends to underestimate the number of elements468 present in a genome the problem here seems to be469 minimal. In addition, we found that when TE470 display was performed on An. gambiae using471 methods that permitted left and right ends to be472 displayed following restriction digestion with dif-473 ferent restriction endonucleases, we obtained sim-474 ilar data (data not shown). Therefore, the method475 employed in this study to display Herves elements476 should reflect the complement of elements present477 in the individuals analyzed.
478 Herves is present in An. merus and An. arabiensis
479 The primers used for Herves transposable element480 display was designed using the sequence of the
481element isolated originally from An. gambiae s.s.482(Figure 1). When Herves transposable element483display was attempted with genomic DNA from484An. merus and An. arabiensis the presence of485Herves elements in these species was detected as486indicated by the resulting PCR products487(Figure 2). Re-amplification and DNA sequencing488of 25 elements confirmed the presence of Herves489and demonstrated that the terminal 30 bp of the490elements present in An. merus (n=9) and491An. arabiensis (n=7) were identical to that of the492An. gambiae element (n=9; data not shown).
493Herves copy number and insertion site494polymorphism in An. gambiae s.s., An. arabiensis495and An. merus
496Herves elements were detected in all individuals497sampled from the three species examined in this498study (n=570). Haploid copy numbers did not499differ significantly between years within species or500among species (F1,12=3.0682, P=0.1053; F2,12=5013.0276, P=0.0862, respectively). Individuals con-502tained, on average, 2.5 copies of Herves per hap-503loid genome (Standard Deviation=0.9) (Table 1).504The proportion of elements segregating at low505frequencies was similar among species and506between years, with one exception (Table 2). To507test for a significant difference in the insertion site508distribution between collection years 2�2 contin-509gency tables were constructed for each species510comparing the number of singletons (insertions511observed exactly once in a sample) and the number512of high frequency elements (insertions found in 2513or more individuals in a sample). In An. gambiae514s.s. and An. arabiensis the proportion of singletons
Figure 1. Left end of Herves showing the location and sequence of the left inverted terminal repeat (Left ITR), the sequence of an
attached Mse adapter with the two complimentary oligonucleotides (MseIa and MseIb), the location and sequence of the pre-selec-
tive PCR primer, HervTEDAL1, and the selective PCR primer, HervTEDAL2. The bracket indicates the region of the Herves ele-
ment for which DNA sequence was obtained from selected isolated and re-amplified transposable element display bands.
6
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4
UNCO
RRECTEDPROO
F515 observed was independent of the year in which the516 observations were made (An. gambiae s.s.: P=517 0.5335 and An. arabiensis: P=0.2002, respec-518 tively). In An. merus, however, this was not the519 case and the proportion of singletons observed was520 not independent of the collection year (P<0.001).521 The 2�2 contingency tables were also used to test522 whether the proportion of singletons observed in523 the two collection years was independent of spe-524 cies. For collections in 2001 the proportion of525 singletons was independent of the species526 (P=0.9446), while this was not the case for col-527 lections in 2003 (P<0.001). Therefore, with the528 exception of An. merus collected in 2003 the pro-529 portion of singletons was the same for all species.530 The transposition activity of Herves in each of531 the species considered in this study was assessed532 assuming the models of Charlesworth and533 Charlesworth (1983) and Langley, Kapland and
534Brookfield (1983). Estimates of a using the model535of Charlesworth and Charlesworth (1983) were536less than 0.01, with few exceptions (data not537shown), suggesting that the number of potential538insertion sites for Herves in the genome of539An. gambiae, arabiensis and merus is very high.540Consequently, the assumption made by Langley,541Kapland & Brookfield (1983) that there are infinite542insertion sites is reasonable in this case, making543the parameters h (Langley, Kapland and544Brookfield 1983) and b (Chalesworth and545Charlesworth, 1983) comparable. Parameter esti-546mations were performed under two conditions. In547An. gambiae s.s. and An. merus there was clear548evidence for at least one very high frequency site549that appeared fixed or nearly fixed (Figures 2 and5503). Such high frequency sites may be occupied by551inactive elements although because these sites were552not shared among the three closely related species
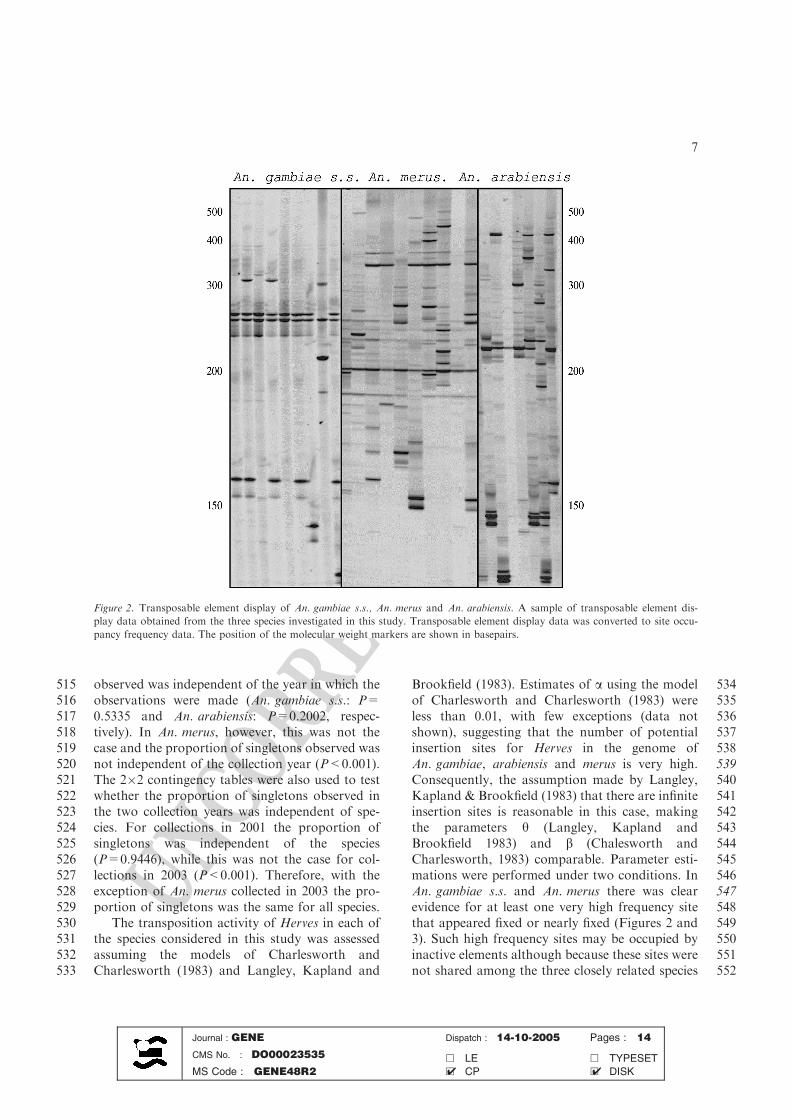
Figure 2. Transposable element display of An. gambiae s.s., An. merus and An. arabiensis. A sample of transposable element dis-
play data obtained from the three species investigated in this study. Transposable element display data was converted to site occu-
pancy frequency data. The position of the molecular weight markers are shown in basepairs.
7
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4
UNCO
RRECTEDPROO
F
553 they are not likely to be ancient fixed sites as were554 described by Wright et al. (2001) in a similar555 analysis of Arabidopsis. Such fixed sites may not be556 contributing to current patterns of population557 dynamics and their affects on parameter estima-558 tion are predictable; stimates of h and b are re-559 duced and the fit of the data to existing models is560 weakened. Parameter estimations of both h and b561 were made on the data including the ‘‘fixed’’ sites
562and then repeated following the removal of the563fixed sites (h*,b*) (Table 1).564To determine if the estimation method (i.e. with565and without the fixed site) resulted in significantly566different parameter estimates we performed a two-567way ANOVA. For each species and parameter (h/
h*, b/b*) the effects of collection date and569parameter estimation method was analyzed. Col-570lection date only contributed significantly to the
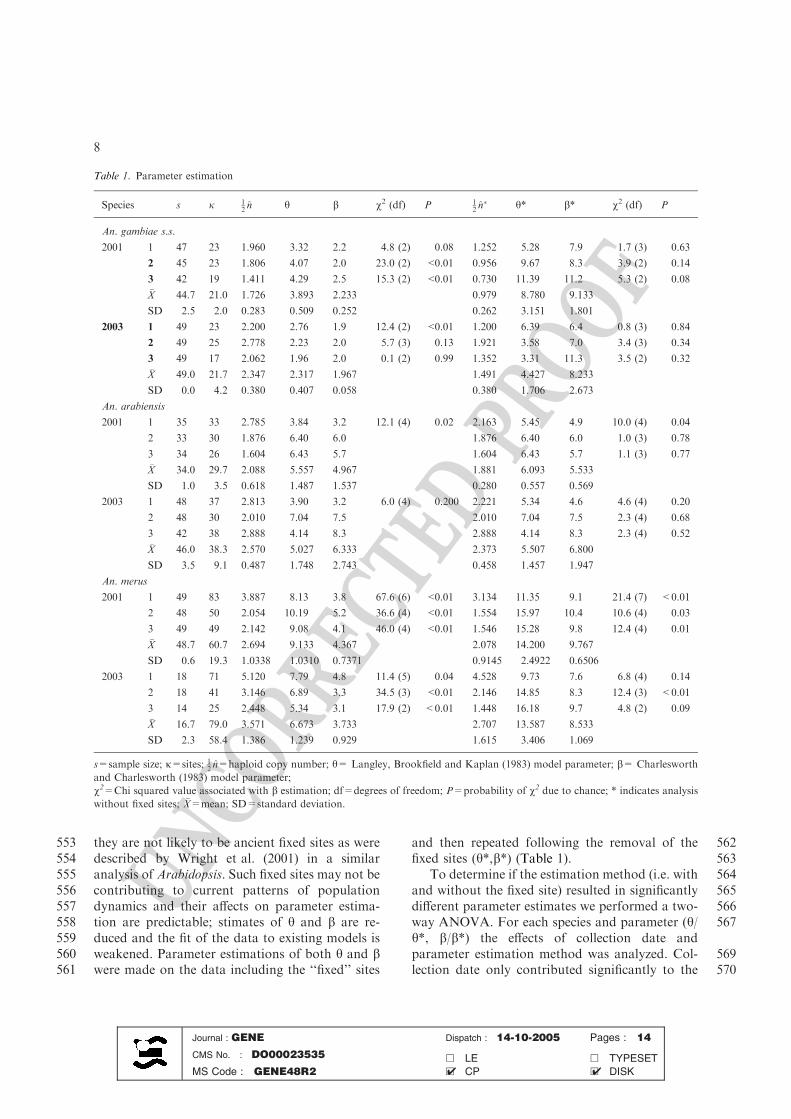
Table 1. Parameter estimation
Species s j 12 n̂ h b v2 (df) P 1
2 n̂� h* b* v2 (df) P
An. gambiae s.s.
2001 1 47 23 1.960 3.32 2.2 4.8 (2) 0.08 1.252 5.28 7.9 1.7 (3) 0.63
2 45 23 1.806 4.07 2.0 23.0 (2) <0.01 0.956 9.67 8.3 3.9 (2) 0.14
3 42 19 1.411 4.29 2.5 15.3 (2) <0.01 0.730 11.39 11.2 5.3 (2) 0.08
�X 44.7 21.0 1.726 3.893 2.233 0.979 8.780 9.133
SD 2.5 2.0 0.283 0.509 0.252 0.262 3.151 1.801
2003 1 49 23 2.200 2.76 1.9 12.4 (2) <0.01 1.200 6.39 6.4 0.8 (3) 0.84
2 49 25 2.778 2.23 2.0 5.7 (3) 0.13 1.921 3.58 7.0 3.4 (3) 0.34
3 49 17 2.062 1.96 2.0 0.1 (2) 0.99 1.352 3.31 11.3 3.5 (2) 0.32
�X 49.0 21.7 2.347 2.317 1.967 1.491 4.427 8.233
SD 0.0 4.2 0.380 0.407 0.058 0.380 1.706 2.673
An. arabiensis
2001 1 35 33 2.785 3.84 3.2 12.1 (4) 0.02 2.163 5.45 4.9 10.0 (4) 0.04
2 33 30 1.876 6.40 6.0 1.876 6.40 6.0 1.0 (3) 0.78
3 34 26 1.604 6.43 5.7 1.604 6.43 5.7 1.1 (3) 0.77
�X 34.0 29.7 2.088 5.557 4.967 1.881 6.093 5.533
SD 1.0 3.5 0.618 1.487 1.537 0.280 0.557 0.569
2003 1 48 37 2.813 3.90 3.2 6.0 (4) 0.200 2.221 5.34 4.6 4.6 (4) 0.20
2 48 30 2.010 7.04 7.5 2.010 7.04 7.5 2.3 (4) 0.68
3 42 38 2.888 4.14 8.3 2.888 4.14 8.3 2.3 (4) 0.52
�X 46.0 38.3 2.570 5.027 6.333 2.373 5.507 6.800
SD 3.5 9.1 0.487 1.748 2.743 0.458 1.457 1.947
An. merus
2001 1 49 83 3.887 8.13 3.8 67.6 (6) <0.01 3.134 11.35 9.1 21.4 (7) <0.01
2 48 50 2.054 10.19 5.2 36.6 (4) <0.01 1.554 15.97 10.4 10.6 (4) 0.03
3 49 49 2.142 9.08 4.1 46.0 (4) <0.01 1.546 15.28 9.8 12.4 (4) 0.01
�X 48.7 60.7 2.694 9.133 4.367 2.078 14.200 9.767
SD 0.6 19.3 1.0338 1.0310 0.7371 0.9145 2.4922 0.6506
2003 1 18 71 5.120 7.79 4.8 11.4 (5) 0.04 4.528 9.73 7.6 6.8 (4) 0.14
2 18 41 3.146 6.89 3.3 34.5 (3) <0.01 2.146 14.85 8.3 12.4 (3) <0.01
3 14 25 2.448 5.34 3.1 17.9 (2) <0.01 1.448 16.18 9.7 4.8 (2) 0.09
�X 16.7 79.0 3.571 6.673 3.733 2.707 13.587 8.533
SD 2.3 58.4 1.386 1.239 0.929 1.615 3.406 1.069
s=sample size; j=sites; 12 n̂=haploid copy number; h= Langley, Brookfield and Kaplan (1983) model parameter; b= Charlesworth
and Charlesworth (1983) model parameter;
v2=Chi squared value associated with b estimation; df=degrees of freedom; P=probability of v2 due to chance; * indicates analysis
without fixed sites; �X=mean; SD=standard deviation.
8
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
571 observed variation between h and h* in An. gam-572 biae s.s. (F1,8=7.9555, P=0.0225). This was also573 the case with Bonferroni and Tukey post-hoc tests574 with a=0.05. Parameter estimation methods (i.e.575 including or excluding the fixed site) contributed576 significantly to the observed variation between h577 and h*, and b and b* in An. gambiae s.s. and578 An. merus. Bonferroni and Tukey post-hoc tests579 (with a=0.05) also showed that excluding the fixed580 site resulted in significantly higher parameter esti-581 mates. This was not the case for An. arabiensis. In582 this species, neither collection time nor estimation583 method contributed significantly to the observed584 variation between h and h* (F1,8=0.4859,585 P=0.5055; F1,8=0.4028, P=0.5434), and b and b*586 (F1,8=1.486, P=0.2576; F1,8=0.2288, P=0.6452).587 The parameters b, b*,h and h* were also ana-588 lyzed using two-way ANOVA to determine if there589 were interaction effects between the species and590 collection dates. Only in the case of h was there a591 significant effect of collection date on the observed592 variation (b, F1,12=0.058, P=0.815; b*,593 F1,12=0.140, P=0.715; h, F1,12=7.549, P=0.018;594 h*, F1,12=2.797, P=0.120, respectively). Variation595 in parameter estimates was significant among596 species: b, F2,12=10.017, P=0.003; b*,597 F2,12=5.770, P=0.018; h, F2,12=25.067, P<0.001598 and h*, F2,12=21.638, P<0.001. Bonferroni and599 Tukey post-hoc tests (with a=0.05) showed sig-600 nificant differences between the b estimates of601 An. gambiae and An. arabiensis, the b* estimates602 of An. arabiensis and An. merus, the h estimates of
603all three species and the h* estimates of An. gam-604biae and An. merus, and An. arabiensis and An.605merus.
606Herves open reading frames are present in607An. gambiae s.s., An. arabiensis and An. merus
608The frequency of individuals containing complete609transposase open reading frames was estimated610using a PCR-based method. Using a sequential,611nested PCR strategy individuals were identified612that contained approximately 2 kb of internal613Herves sequences encompassing the open reading614frame. Approximately 96% of the individuals615tested (n=216) displayed evidence of an intact616open reading frame based on the presence of ele-617ments that did not contain large deletions in that618region of the element (Table 3, Figure 4). Only 2619of the 216 individuals tested for the presence of620open reading frames had evidence of internally621deleted elements.
622Discussion
623The results reported here show that An. gambiae624s.s., An. arabiensis and An. merus in Mozambique625contain the Herves element and preliminary626transposable element display data from An. melas627and An. quadriannulatus indicate that these species628also contain the element (R. A. Subramanian and629D. A. O’Brochta, unpublished) and thus Herves630appears to be present in all members of the species631complex (data for An. bwambae is unavailable).632These distribution data are consistent with the idea633that Herves was present in this lineage prior to the634divergence of An. gambiae s.s., arabiensis and635merus and probably prior to the divergence of the636species complex. Other observations, however, are637somewhat difficult to reconcile with the idea that638this element is an old component of these ge-639nomes. For example, DNA sequence analysis of640the 30 terminal nucleotides of 25 elements sampled641from 3 species did not reveal any sequence poly-642morphisms. Of the 216 mosquitoes that were643sampled only 2 showed any evidence for the644presence of internally deleted elements. This is645somewhat surprising since the accumulation of646internally deleted elements through the process of647interrupted templated gap repair appears to be a648hallmark of Class II elements as illustrated by P,
Table 2. Insertion site data
Ia II III
1b >1c 1 >1 1 >1
An. gambiae
2001 7 16 12 11 4 15
2003 9 14 7 18 4 13
An. arabiensis
2001 5 28 13 17 11 15
2003 8 29 8 22 9 29
An. merus
2001 28 55 17 33 17 32
2003 37 34 26 15 17 8
aRoman numerals: replicate analyses.bSingletons: sites found in exactly one individual.cNon-singletons: sites in more than one individual.
9
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4
UNCO
RRECTEDPROO
F
649hobo, and mariner elements in insects and Ds ele-650ments in plants. So at this point we have a very651unclear understanding of the evolutionary history652of Herves in Anopheles. A more extensive sampling653and analysis of appropriate DNA sequences are654required in order to resolve the question of whe-655therHerves is an ancient or modern addition to the656genomes of the species examined.657The copy numbers of Herves in the three spe-658cies tested were not significantly different from
y g p
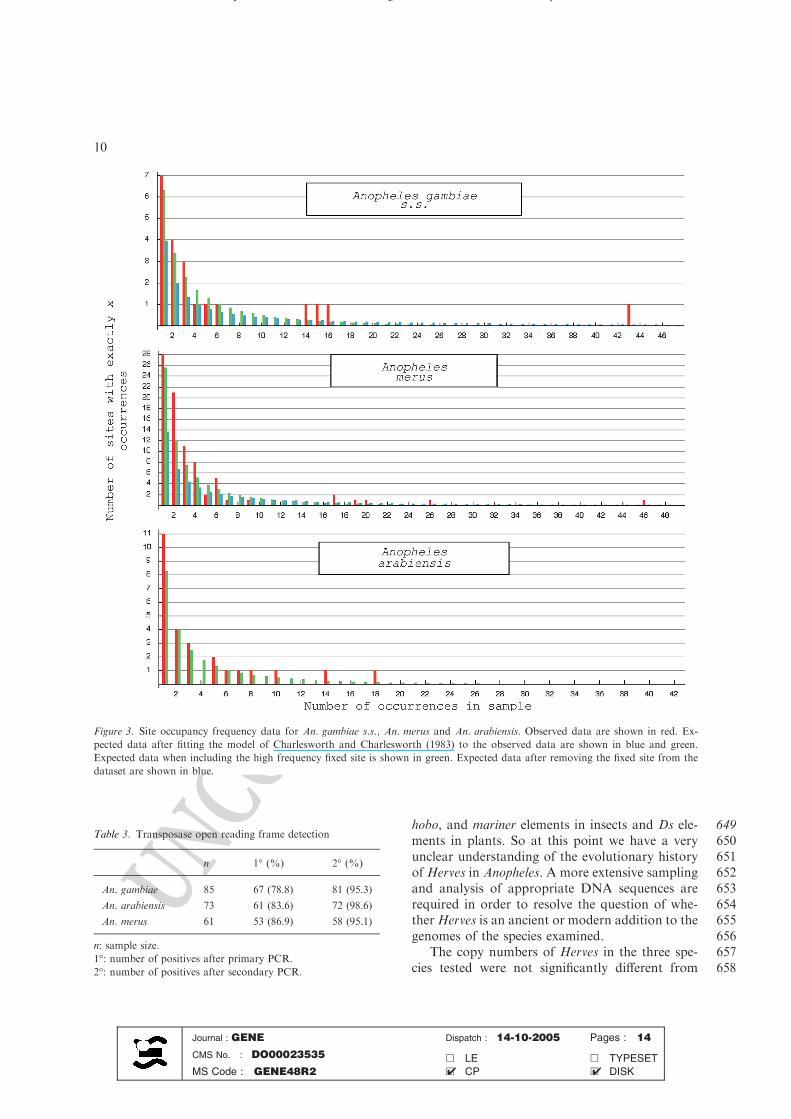
Figure 3. Site occupancy frequency data for An. gambiae s.s., An. merus and An. arabiensis. Observed data are shown in red. Ex-
pected data after fitting the model of Charlesworth and Charlesworth (1983) to the observed data are shown in blue and green.
Expected data when including the high frequency fixed site is shown in green. Expected data after removing the fixed site from the
dataset are shown in blue.
Table 3. Transposase open reading frame detection
n 1� (%) 2� (%)
An. gambiae 85 67 (78.8) 81 (95.3)
An. arabiensis 73 61 (83.6) 72 (98.6)
An. merus 61 53 (86.9) 58 (95.1)
n: sample size.
1�: number of positives after primary PCR.
2�: number of positives after secondary PCR.
10
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F659 each other. When the high frequency fixed cites in660 An. gambiae and arabiensis were included in the661 calculations the overall mean copy number per662 mosquito was 5.0 with a standard deviation of 0.9.663 Furthermore, of the 570 individuals analyzed in664 this study none lacked the element entirely.665 Therefore, Herves appears to be maintained in666 An. gambiae, arabiensis and merus at low copy667 number levels relative to well-studied insect ele-668 ments such as P elements in D. melanogaster,669 which are present in copy numbers up to 30–40 per670 genome (Engels, 1989). The low copy number of671 Herves in Anopheles is similar to what has been672 observed of Bari 1 in D. melanogaster, Hermes in673 Musca domestica (D. O’Brochta, unpublished674 observations) and of various hAT elements in675 plants (Kunze & Weil, 2002). In contrast to this,676 however, is hobo, a related hAT element found in677 D. melanogaster, which can be found at relatively678 high copy numbers in those strains of the species679 that contain the element. Blackman et al. (1989)680 found that intact elements were present at copy681 numbers from 2 to 10 while there can be 30–75682 copies of internally deleted hobo elements per683 genome. Additional studies of Herves in popula-684 tions from other geographic locations will be685 needed to determine the extent to which copy686 number varies.687 The observed occupancy frequency profiles of688 transposable elements have been used to estimate
689the magnitude of the forces that tend to remove690elements from populations (e.g. excision and691selection) (Charlesworth & Langley, 1990). Since692the elements are assumed to be at equilibrium the693forces removing the elements are counterbalanced694by equal forces that result in new copies of the695element being added to the genome (e.g. replicative696transposition). Consequently, observed occupancy697frequency profiles can be used to estimate the698transposition rate relative to the effective popula-699tion size (Charlesworth & Charlesworth, 1983;700Langley, Brookfield & Kaplan, 1983). The701parameters b and h are approximately equal to7024Nem, where m is the replicative transposition rate.703These models have been applied to the study of704retrotranposable elements in D. melanogaster705(Leigh-Brown & Moss, 1987; Charlesworth &706Langley, 1990; Charlesworth, Lapid & Canada,7071992; Biemont et al., 1994; Nuzhdin, 1995; Lab-708rador, del Carmen Seleme & Fontdevila, 1998;709Nuzhdin, 1999) and more recently to similar ele-710ments in vertebrates (Neafsey, Blumenstiel &711Hartl, 2004). Class II elements have only occa-712sionally been analyzed with respect to site-occu-713pancy frequency (Biemont et al., 1994; Wright714et al., 2001). In the present study, values of the715parameters b and h were much greater that one,716indicating a significant deviation from neutral717expectations where the observed distributions are718the consequence largely of genetic drift (Table 1).
Figure 4. Open reading frame detection assay. Examples of the types of results obtained with this assay are shown. Lane 1 shows a
product of the expected size after one round of PCR using a template with a sequenced open reading frame. Lane 2 shows a prod-
uct of the expected size after one round of PCR using an individual’s genomic DNA as a template. Lane 3 shows the results after
one round of PCR using an individual’s genomic DNA as a template. Lane 4 shows the results after two rounds of PCR using the
same template as used in lane 3. The nested PCR product is expected to be 210 bp shorter than the primary product if there are
no other internal deletions. Lane 5 and 6 are the PCR products obtained after two rounds of PCR using genomic DNA samples
from individuals with internally deleted elements. Molecular weight markers are shown in kilobase pairs.
11
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
719 Thus, it appears that the Herves element is actively720 transposing in the three species tested. This con-721 clusion holds whether the presumptive fixed sites722 were included or removed from the analysis.723 Effective population size has been estimated for724 populations of An. gambiae s.s. and An. arabiensis725 in east Africa and are in the range of 0.5�104
726 (Lehmann et al., 1998). Similar estimates have727 been made in west-African populations (Taylor728 et al., 1993). If these estimates of Ne are approxi-729 mately correct then the transposition rates of730 Herves are expected to be in the range of 10–3 to731 10)4. These are rates that are consistent with esti-732 mated rates of movement of other elements in wild733 populations (Eggleston, Johnson-Schlitz & Engels,734 1988; Harada, Yukuhiro & Mukai, 1990; Nuzhdin735 & Mackay, 1995; Dominguez & Albornoz, 1996).736 In further support of the conclusion that Herves is737 actively transposing in wild populations of738 An. gambiae is the observation that most individ-739 uals within the populations studied contained ele-740 ments with evidence for complete transposase741 coding regions.742 The data presented here suggests that the743 models of Charlesworth and Charlesworth (1983)744 and Langley, Brookfield and Kaplan, (1983)745 appear to be appropriate for Herves in An. gam-746 biae under some conditions, based on the goodness747 of fit of the observed and expected site-frequency748 distributions (v2 and P in Table 1). When high749 frequency fixed sites were included in the analysis750 the fit of the Charlesworth and Charlesworth751 (1983) model to the observed data was generally752 poor. Removal of the fixed sites from the analysis753 greatly improved the fit of the model to the754 observed data from An. gambiae s.s. and An. ara-755 biensis. The model fit the An. merus data poorly in756 most cases (Table 1).757 Herves presents an opportunity to explore the758 dynamics of transposable elements in an insect of759 significance to public health. It is likely to provide760 insight into how active, autonomous Class II761 transposable elements such as piggyBac, mariner,762 Minos and Hermes, elements currently used as763 mosquito gene vectors and being considered as764 genetic drive agents, will behave following their765 introduction into Anopheles gambiae. It will also766 provide opportunities to assess models of trans-767 posable element population dynamics developed768 largely from the study of elements in D. melanog-769 aster.
770Acknowledgements
771The work at the UMBI has been supported by a772grant from the National Institutes of Health,773GM48102. The work at the UCR was supported774by a grant from the National Institutes of775Health, AI45741. We would like to acknowledge776the valuable comments of two reviewers.
777References
778Anxolabehere, D., K. Hu, D. Nouaud & G. Periquet, 1990. PM
779system:A survey ofDrosophila melanogaster strains from the
780People’s Republic of China. Genet. Sel. Evol. 22: 175–188.
781Anxolabehere, D., M.G. Kidwell & G. Periquet, 1988. Molec-
782ular characteristics of diverse populations are consistent
783with the hypothesis of a recent invasion of Drosophila mel-
784anogaster by mobile P elements. Mol. Biol. Evol. 5:
785252–269.
786Arensburger, P., J. Orsetti, Y.-J. Kim, C. Aluvihare & D.A.
787O’Brochta, 2005. A new active transposable element,
788Herves, from the African malaria mosquito Anophe-
789les gambiae. Genetics 169: 697–708.
790Ashburner, M., 1989. Drosophila: A Laboratory Manual. Cold
791Spring Harbor Laboratory Press, Cold Spring Harbor.
792Atkinson, P.W., A.C. Pinkerton & D.A. O’Brochta, 2001.
793Genetic transformation systems in insects. Ann. Rev.
794Entomol. 46: 317–346.
795Bensaadi-Merchaermek, N., J.C. Salvado & C. Mouches, 1994.
796Mosquito transposable elements. Genetica 93: 139–148.
797Biedler, J., Y. Qi, D. Holligan, A. della Torre, S. Wessler & Z.
798Tu, 2003. Transposable element, (TE) display and rapid
799detection of TE insertion polymorphism in the Anophe-
800les gambiae species complex. Insect Mol. Biol. 12: 211–216.
801Biedler, J. & Z. Tu, 2003. Non-LTR retrotransposons in the
802African malaria mosquito Anopheles gambiae: Unprece-
803dented diversity and evidence of recent invasion. Mol. Biol.
804Evol. 20: 1811–1825.
805Biemont, C., F. Lemeunier, M.P. Garcia Guerreiro, J.F.
806Brookfield, C. Gautier, S. Aulard & E.G. Pasyukova,
8071994. Population dynamics of the copia, mdg1, mdg3, gypsy
808and P transposable elements in a natural population of
809Drosophila melanogaster. Genet. Res. 63: 197–212.
810Blackman, R.K., M. Macy, D. Koehler, R. Grimaila & W.M.
811Gelbart, 1989. Identification of a fully-functional hobo
812transposable element and its use for germ-line transforma-
813tion of Drosophila. EMBO J. 8: 211–217.
814Boete, C. & J.C. Koella, 2002. A theoretical approach to
815predicting the success of genetic manipulation of malaria
816mosquitoes in malaria control. Malaria J. 25: 1–3.
817Bucheton, A., C. Vaury, M.C. Chaboissier, P. Abad, A.
818Pelisson & M. Simonelig, 1992. I elements and the
819Drosophila genome. Genetica 86: 175–190.
820Casa, A.M., C. Brouwer, A. Nagel, L. Wang, Q. Zhang, S.
821Kresovich & S.R. Wessler, 2000. The MITE family Heart-
822breaker, (Hbr): Molecular markers in maize. Proc. Natl.
823Acad. Sci. USA 97: 10083–10089.
12
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
824 Charlesworth, B. & D. Charlesworth, 1983. The population
825 dynamics of transposable elements. Genet. Res. 42: 1–27.
826 Charlesworth, B. & C.H. Langley, 1989. The population
827 genetics of Drosophila transposable elements. Annu. Rev.
828 Genet. 23: 251–287.
829 Charlesworth, B. & C.H. Langley, 1990. Population genetics of
830 transposable elements in Drosophila, pp. 150–177 in Evo-
831 lution at the Molecular Level, edited by R.K. Selander,
832 A.G. Clark &T.S. Whittam. Sinauer Associates Inc, Sun-
833 derland.
834 Charlesworth, B., A. Lapid & D. Canada, 1992. Distribution of
835 transposable elements within and between chromosomes in
836 a population of Drosophila melanogaster II. Inferences on
837 the nature of selection against elements. Genet. Res. 60:
838 115–130.
839 Collins, F.H., R.K. Sakai, K.D. Vernick, S. Paskewitz, D.C.
840 Seeley, L.H. Miller, W.E. Collins, C. CC & R.W. Gwadz,
841 1986. Genetic selection of a Plasmodium-refractory strain of
842 the malaria vector Anopheles gambiae. Science 234:
843 607–610.
844 della Torre, A., Z. Tu & V. Petrarca, 2005. On the distribution
845 and genetic differentiation of Anopheles gambiae s.s. molec-
846 ular forms. Insect Biochem. Mol. Biol. 35: 755–769.
847 Dominguez, A. & J. Albornoz, 1996. Rates of movement of
848 transposable elements in Drosophila melanogaster. Mol.
849 Gen. Genet. 251: 130–138.
850 Eggleston, W.B., D.M. Johnson-Schlitz & W. Engels, 1988.
851 P-M hybrid dygenesis does not mobilize other transposable
852 element families in D. melanogaster. Nature 331: 368–370.
853 Engels, W.R., 1989. P elements in Drosophila melanogaster, pp.
854 439–484, in Mobile DNA, edited by D.E. Berg & M.M.
855 How. American Society for Microbiology, Washington DC.
856 Evgen’ev, M., H. Zelentsova, L. Mnjoian, H. Poluecotova &
857 M.G. Kidwell, 2000. Invasion of Drosophila virilis by the
858 Penelope transposable element. Chromosoma 109: 350–357.
859 Feschotte, C. & C. Mouches, 2000. Recent amplification of
860 miniature inverted-repeat transposable elements in the
861 vector mosquito Culex pipiens: characterization of the
862 Mimo family. Gene 250: 1009–1116.
863 Grossman, G.L., A.J. Cornel, C.S. Rafferty, H.M. Robertson &
864 F.H. Collins, 1999. Tsessebe, Topi and Tiang: three distinct
865 Tc1–like transposable elements in the malaria vector,
866 Anopheles gambiae. Genetica 105: 69–80.
867 Guimond, N., D.K. Bideshi, A.C. Pinkerton, P.W. Atkinson &
868 D.A. O’Brochta, 2003. Patterns of Hermes Transposition in
869 Drosophila melanogaster. Mol. Gen. Genet. 268: 779–790.
870 Harada, K., K. Yukuhiro & T. Mukai, 1990. Transposition
871 rates of movable genetic elements in Drosophila melanog-
872 aster. Proc. Natl. Acad. Sci. USA 87: 3248–3252.
873 Hartl, D.L., A.R. Lohe & E.R. Lozovskaya, 1997. Modern
874 thoughts on an ancyent marinere: function, evolution,
875 regulation. Annu. Rev. Genet. 31: 337–358.
876 Holt, R.A., G.M. Subramanian, A. Halpern, G.G. Sutton, R.
877 Charlab, D.R. Nusskern, P. Wincker, A.G. Clark, J.M.
878 Ribeiro, R. Wides, S.L. Salzberg, B. Loftus, M. Yandell,
879 W.H. Majoros, D.B. Rusch, Z. Lai, C.L. Kraft, J.F. Abril,
880 V. Anthouard, P. Arensburger, P.W. Atkinson, H. Baden,
881 V. de Berardinis, D. Baldwin, V. Benes, J. Biedler, C. Blass,
882 R. Bolanos, D. Boscus, M. Barnstead, S. Cai, A. Center, K.
883 Chatuverdi, G.K. Christophides, M.A. Chrystal, M. Clamp,
884 A. Cravchik, V. Curwen, A. Dana, A. Delcher, I. Dew, C.A.
885Evans, M. Flanigan, A. Grundschober-Freimoser, L. Frie-
886dli, Z. Gu, P. Guan, R. Guigo, M.E. Hillenmeyer, S.L.
887Hladun, J.R. Hogan, Y.S. Hong, J. Hoover, O. Jaillon, Z.
888Ke, C. Kodira, E. Kokoza, A. Koutsos, I. Letunic, A.
889Levitsky, Y. Liang, J.J. Lin, N.F. Lobo, J.R. Lopez, J.A.
890Malek, T.C. McIntosh, S. Meister, J. Miller, C. Mobarry, E.
891Mongin, S.D. Murphy, D.A. O’Brochta, C. Pfannkoch, R.
892Qi, M.A. Regier, K. Remington, H. Shao, M.V. Sharakh-
893ova, C.D. Sitter, J. Shetty, T.J. Smith, R. Strong, J. Sun, D.
894Thomasova, L.Q. Ton, P. Topalis, Z. Tu, M.F. Unger, B.
895Walenz, A. Wang, J. Wang, M. Wang, X. Wang, K.J.
896Woodford, J.R. Wortman, M. Wu, A. Yao, E.M. Zdobnov,
897H. Zhang & Q. Zhao, 2002. The genome sequence of the
898malaria mosquito Anopheles gambiae. Science 298: 129–149.
899Ito, J., A. Ghosh, L.A. Moreira, E.A. Wimmer & M. Jacobs-
900Lorena, 2002. Transgenic anopheline mosquitoes impaired
901in transmission of a malaria parasite. Nature 417: 452–455.
902Jordan, I.K., L.V. Matyunina & J.F. McDonald, 1999.
903Evidence for the recent horizontal transfer of long terminal
904repeat retrotransposon. PNAS 96: 12621–12625.
905Jordan, I.K. & J.F. McDonald, 1998. Evolution of the copia
906retrotransposon in Drosophila melanogaster species sub-
907group. Mol. Biol. Evol. 15: 1160–1171.
908Kidwell, M.G., 2002. Transposable elements and the evolution
909of genome size in eukaryotes. Genetica 115: 49–63.
910Kim, W., H. Koo, A.M. Richman, D. Seeley, J. Vizioli, A.D.
911Klocko & D.A. O’Brochta, 2004. Ectopic expression of a
912cecropin transgene in the human malaria vector mosquito
913Anopheles gambiae, (Diptera: Culicidae): Effects on suscep-
914tibility to Plasmodium. J. Med. Entomol. 41: 447–455.
915Kunze, R. & C.F. Weil, 2002. The hAT and CACTA
916superfamilies of plant transposons, pp. 565–610 in Mobile
917DNA II, edited by N.L. Craig, R. Craige, M. Gellert
918& A.M. Lambowitz. American Society of Microbiology,
919Washington, DC.
920Labrador, M., M. del Carmen Seleme & A. Fontdevila, 1998.
921The evolutionary history of Drosophila buzzatii. XXXIV.
922The distribution of the retrotransposon Osvaldo in original
923and colonizing populations. Mol. Biol. Evol. 15: 1532–
9241547.
925Langley, C.H., J.F.Y. Brookfield & N. Kaplan, 1983. Trans-
926posable elements in mendelian populations. I. A theory.
927Genetics 104: 457–471.
928Lehmann, T., W.A. Hawley, H. Grebert & F.H. Collins, 1998.
929The effective population size of Anopheles gambiae in
930Kenya: Implications for population structure. Mol. Biol.
931Evol. 15: 264–276.
932Leigh-Brown, A.J. & J.E. Moss, 1987. Transposition of the I
933element and copia in natural populations of Drosophila mel-
934anogaster. Genet. Res. 49: 231–237.
935Lohe, A.R., E.N. Moriyama, D.A. Lindholm & D.L. Hartl,
9361995. Horizontal transmission, vertical inactivation, and
937stochasitc loss of mariner-like transposable elements. Mol.
938Biol. Evol. 12: 62–72.
939Miller, J.M. & P. Capy, 2004. Mobile Genetic Elements:
940Protocols and Genomic Applications. Humana Press,
941Totowa.
942Miller, L.H., R.K. Sakai, P. Romans, R.W. Gwadz, P. Kantoff
943& H.G. Coon, 1987. Stable integration and expression of a
944bacterial gene in the mosquito Anopheles gambiae. Science
945237: 779–781.
13
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4

UNCO
RRECTEDPROO
F
946 Moreira, L.A., J. Ito, A. Ghosh, M. Devenport, H. Zieler, E.G.
947 Abraham, A. Crisanti, T. Nolan, F. Catteruccia & M.
948 Jacobs-Lorena, 2002. Bee venom phospholipase inhibits
949 marlaria parasite development in transgenic mosquitoes. J.
950 Biol. Chem. 25: 40839–40843.
951 Neafsey, D.E., J.P. Blumenstiel & D.L. Hartl, 2004. Different
952 regulatory mechanisms underlie similar transposable ele-
953 ment profiles in pufferfish and fruitflies. Mol. Biol. Evol3 .
954 Nuzhdin, S.V., 1995. The distribution of transposable elements
955 on X chromosomes from a natural population of Drosoph-
956 ila simulans. Genet. Res. 66: 159–166.
957 Nuzhdin, S.V., 1999. Sure facts, speculations, and open
958 questions about the evolution of transposable element copy
959 number. Genetica 107: 129–137.
960 Nuzhdin, S.V. & T.G. Mackay, 1995. The genomic rate of
961 transposable element movement in Drosophila melanogas-
962 ter. Mol. Biol. Evol. 12: 180–181.
963 Pinsker, W., E. Haring, S. Hagemann & W.J. Miller, 2001. The
964 evolutionary life history of P transposons: from horizontal
965 invaders to domesticated neogenes. Chromosoma 110: 149–
966 158.
967 Ribeiro, J.M.C. & M.G. Kidwell, 1994. Transposable elements
968 as population drive mechanisms: specification of critical
969 parameter values. J. Med. Entomol. 31: 10.
970 Robertson, H.M., 2002. Evolution of DNA transposons in
971 eukaryotes, pp. 1093–1110 in Mobile DNA II, edited by
972 N.L. Craig, R. Craige, M. Gellert & A.M. Lambowitz.
973 ASM Press, Washington DC.
974 Rohr, C.J.B., H. Ranson, X. Wang & N.J. Besansky, 2002.
975 Structure and evolution of mtanga, a retrotransposon
976 actively expressed on the Y chromosome of the African
977 malaria vector Anopheles gambiae. Mol. Biol. Evol. 19:
978 149–162.
979 Ryder, E. & S. Russell, 2003. Transposable elements as tools for
980 genomics and genetics in Drosophila. Brief. Funct. Geno-
981 mic. Proteomic. 2: 57–71.
982 Sanchez-Gracia, A., X. Maside & B. Charlesworth, 2005. High
983 rate of horizontal transfer of transposable elements in
984 Drosophila. Trends Genet. 21: 200–203.
985 Scott, J.A., W.G. Brogdon & F.H. Collins, 1993. Identification
986 of single specimens of the Anopheles gambiae complex by
987polymerase chain reaction. Am. J. Trop. Med. Hyg. 49:
988520–529.
989Shao, H., Y. Qi & Z. Tu, 2001. MsqTc3, a Tc3–like transposon
990in the yellow fever mosquito Aedes aegypti. Insect Mol.
991Biol. 10: 421–425.
992Silva, J.C. & M.G. Kidwell, 2000. Horizontal transfer and
993selection in the evolution of P elements. Mol. Biol. Evol. 17:
9941542–1547.
995Simmons, G., 1992. Horizontal transfer of hobo transposable
996elements within the Drosophila melanogaster species com-
997plex: Evidence from DNA sequencing. Mol. Biol. Evol. 9:
9981050–1060.
999Taylor, C.E., Y.T. Toure, M. Coluzzi & V. Petrarca, 1993.
1000Effective population size and persistence of Anopheles ara-
1001biensis during the dry season in west Africa. Med. Vet.
1002Entomol. 7: 351–357.
1003Torti, C., A.R. Malacrida, G. Yannopoulos, C. Louis & G.
1004Gasperi, 1994. Hybrid dysgenesis-like phenomena in the
1005medfly, Ceratitis capitata, (Diptera, Tephritidae). J. Hered.
100685: 92–99.
1007Tu, Z. & J.J. Hill, 1999. Mosqul, a novel family of mosquito
1008retrotransposons distantly related to the Drosophila I
1009factors, may consist of elements of more than one origin.
1010Mol. Biol. Evol. 16: 1675–1686.
1011Van den Broeck, D., T. Maes, M. Sauer, H. Zethof, P. De
1012Keukeleire, M. D’Hauw, M. Van Montagu & T. Gerats,
10131998. Transposon display identifies individual transposable
1014elements in high copy number lines. Plant J. 13: 121–129.
1015Vos, P., R. Hogers, M. Bleeker, M. Reijans, T. van de Lee, M.
1016Hornes, A. Frijters, J. Pot, J. Peleman, M. Kuiper & M.
1017Zabeau, 1995. AFLP: a new technique for DNA finger-
1018printing. Nuc. Acid Res. 23: 4407–4414.
1019Wright, S.I., Q.H. Le, D.J. Schoen & T.E. Bureau, 2001.
1020Population dynamics of an Ac-like transposable element in
1021self- and cross-pollinating Arabidopsis. Genetics 158: 1279–
10221288.
1023Zampicinini, G., A. Blinov, P. Cervella, V. Guryev & G. Sella,
10242004. Insertional polymorphism of a non-LTR mobile
1025element, (NLRCth1) in European populations of Chirono-
1026mus riparius, (Diptera, Chironomidae) as detected by
1027transposon insertion display. Genome 47: 1154–1163.
1028
14
Journal : GENE Dispatch : 14-10-2005 Pages : 14
CMS No. : DO00023535 h LE h TYPESET
MS Code : GENE48R2 h CP h DISK4 4