global completeness of the bat fossil record
TRANSCRIPT

ORIGINAL PAPER
Global Completeness of the Bat Fossil Record
Thomas P. Eiting & Gregg F. Gunnell
Published online: 10 July 2009# Springer Science + Business Media, LLC 2009
Abstract Bats are unique among mammals in theiruse of powered flight and their widespread capacityfor laryngeal echolocation. Understanding how andwhen these and other abilities evolved could beimproved by examining the bat fossil record. How-ever, the fossil record of bats is commonly believed tobe very poor. Quantitative analyses of this recordhave rarely been attempted, so it has been difficult togauge just how depauperate the bat fossil recordreally is. A crucial step in analyzing the quality of thefossil record is to be able to accurately estimatecompleteness. Measures of completeness of the fossilrecord have important consequences for our under-standing of evolutionary rates and patterns amongbats. In this study, we applied previously developedstatistical methods of analyzing completeness to thebat fossil record. The main utility of these methodsover others used to study completeness is theirindependence from phylogeny. This phylogenetic-
independence is desirable, given the recent state offlux in the higher-level phylogenetic relationships ofbats. All known fossil bat genera were tabulated at thegeologic stage or sub-epoch level. This binningstrategy allowed an estimate of the extinction ratefor each bat genus per bin. Extinction rate—togetherwith per-genus estimates of preservation probabilityand original temporal distributions—was used tocalculate completeness. At the genus level, the batfossil record is estimated to be 12% complete. Withinthe order, Pteropodidae is missing most of its fossilhistory, while Rhinolophoidea and Vespertilionoideaare missing the least. These results suggest that 88%of bats that existed never left a fossil record, and thatthe fossil record of bats is indeed poor. Much of thetaxonomic and evolutionary history of bats has yet tobe uncovered.
Keywords Chiroptera . Completeness . Fossil record .
Phylogeny-independent
Introduction
Fossils provide the only direct evidence concerningthe diversity of life through time, and as such theyare paramount in studies of evolutionary patterns andrates. Macroevolutionary studies that address suchissues typically employ the relatively densely sam-pled invertebrate fossil record (e.g., Foote and Raup1996; Cheetham and Jackson 1998). However,
J Mammal Evol (2009) 16:151–173DOI 10.1007/s10914-009-9118-x
T. P. Eiting (*)Program in Organismic and Evolutionary Biology,221 Morrill Science Center South,University of Massachusetts,Amherst, MA 01003, USAe-mail: [email protected]
G. F. GunnellMuseum of Paleontology, University of Michigan,Ann Arbor, MI 48109, USAe-mail: [email protected]

several vertebrate groups have well-sampled fossilrecords whose long-term evolutionary histories havebeen thoroughly studied by paleobiologists. Amongfossil mammals, most work that pertains to macro-evolutionary patterns and processes involves eitherlarge-scale analyses of terrestrial land-mammalsthrough the Cenozoic (e.g., Foote and Raup 1996;Alroy 2000), detailed examinations of relativelyshort, densely sampled fossil occurrences (e.g.,Clyde and Gingerich 1994; Alba et al. 2001; Woodet al. 2007), or, less commonly, investigations atscales intermediate between these two (e.g.,MacFadden 1985). In most cases, these studies havebeen restricted to taxonomic groups that have beenstudied extensively and for which considerablediversity or abundance of fossil forms is thought tobe present. Evolutionary patterns in the fossil recordof relatively poorly sampled groups have been onlyrarely examined.
One conspicuous group for which macroevolu-tionary analyses using the fossil record have beenrare is bats (Mammalia: Chiroptera). With morethan 1100 extant species (Simmons 2005), batscomprise an extraordinary taxonomic diversity thatis second only to rodents among living mammals.Phylogenetic investigations of bats have providedsome important insights into the evolutionary historyof the group, including recent efforts reaffirming batmonophyly and uncovering various hypotheses ofmicrochiropteran paraphyly. In principal, a perfectfossil record would illuminate the correct phyloge-netic history of bats; such a record might serve as anindependent test of the correctness of variousphylogenetic hypotheses. However, few studies ofbats have incorporated extinct taxa into phylogeneticanalyses, especially in genus- or species-level anal-yses. Using fossils in systematics will likely becomeless common as molecular-based systematics con-tinues to proliferate (Hermsen and Hendricks 2008).However, depending on the types of hypothesesbeing tested, ignoring fossil taxa that containphylogeny-relevant information not found in extantorganisms may limit our potential to completelyunderstand the evolutionary histories of these groupsand may even produce misleading results in someinstances.
To determine the potential utility of the bat fossilrecord in illuminating patterns of evolution, anunderstanding of its quality must be assessed. How
accurately does the known fossil record reflect thetrue evolutionary history of bats? Two conceptualapproaches have been used to address this questionin other groups of animals. The first is phylogeny-dependent, in which the quality of the fossil recordis assessed for a particular phylogeny. Teeling et al.(2005) presented one type of phylogeny-dependentmethod of assessing the fossil record of bats. Theygenerated Bayesian estimates of divergence dates foreach of the 58 branches (internal and terminal) fromtheir maximum likelihood tree. Teeling et al. thencompared their molecular divergence estimates withthe oldest known fossils from the 58 branches of thetree. They used the differences in ages between themolecular and fossil estimates to calculate a percent-age of missing fossil data per branch. The results oftheir analyses estimated that, on average, 73% of theknown fossil record of 58 lineages of bats wasmissing. Pteropodid bats, long known to have apoorly sampled record, were missing upwards of98% of their fossil history. The four non-pteropodidsuperfamilies were missing between 56% and 86%of their fossil record. Thus, the results of the Teelinget al. (2005) study seem to show that the fossilrecord of bats is quite poor. However, methods likethat presented in Teeling et al. (2005) are heavilydependent upon the lineages included and thetopology of the phylogeny used. Due to recentvolatility in hypotheses of the higher-level phyloge-netic relationships of bats (Springer et al. 2001;Jones et al. 2002, 2005; Eick et al. 2005; Gunnelland Simmons 2005; Teeling et al. 2005; Hermsenand Hendricks 2008), there may be some uncertaintyin estimating the quality of the fossil record of batsusing phylogeny-dependent methods.
Fortunately, the second class of methods forassessing the quality of the fossil record is phyloge-ny-independent. That is, statistical metrics are applieddirectly to the fossil data that quantify the quality ofthe fossil record (Benton 1998; Paul 1998). Themethod developed by Foote and Raup (1996) allowsfor one such quantitative analysis of the fossil recordfor any group independent of phylogeny. By estimat-ing preservation probability, extinction rate, and thestratigraphic duration of taxa, completeness can becalculated deterministically. Completeness is definedas “the proportion of taxa that have left some fossilrecord,” (Foote and Raup 1996; sensu Valentine1989), and it therefore serves as a metric for assessing
152 J Mammal Evol (2009) 16:151–173

the quality of the fossil record. In this paper weaddress the quality of the bat fossil record bycalculating completeness using the method presentedin Foote and Raup (1996). We calculate completenessfor Chiroptera as a whole, as well as for the majorhigher-level bat clades. These calculations allow for acomparison of the fossil record for each of thesegroups, as well as an assessment of the quality of thefossil record of bats using a method independent ofphylogeny.
Materials and methods
Data collection
A database of all known fossil bat genera wascompiled from the literature Appendix (1). Generawere tabulated according to their temporal and geo-graphic distribution. Taxonomy followed Simmons(2005), with the exceptions of Perimyotis and Para-strellus, which were considered distinct generafollowing the recommendations of Hoofer et al.(2006). Miniopteridae was considered to be a dis-tinct family, following recommendations of recentanalyses (e.g., Miller-Butterworth et al. 2007).Superfamily membership followed Teeling et al.(2005). Specimens identified as “cf.” (e.g., “cf.Eptesicus indet.”) were considered valid records forrecording presence/absence. Pre-Pleistocene fossilbats were assigned to one of fifteen chronostrati-graphic stages: Ypresian, Lutetian, Bartonian, Pria-bonian, Rupelian, Chattian, Aquitanian, Burdigalian,Langhian, Serravalian, Tortonian, Messinian, Zan-clean, Piacenzian, and Gelasian. The Pleistocene wastabulated as a single bin. Stages and boundary dateswere taken from Gradstein et al. (2004). Correlationof the European Mammal Neogene (MN) systemwas performed using Agustí et al. (2001) andreferences therein. Recent genera were tabulatedand used for comparative purposes (but not for sta-tistical calculations).
Data analysis
Calculations of completeness were carried outaccording to the discrete-time and infinite-windowmethod of Foote and Raup (1996). This methodrequires tabulating the ranges of all taxa (bat genera
in this case) according to discrete time intervals(mostly stages or combinations of two to three stagesfor this study). Our binning strategy attempted toproduce bins of roughly equal durations. Thus, wesummed several smaller bins into fewer large bins inan effort to combat high variance in the duration ofstages in the Cenozoic (older stages tend to be longerthan younger). In particular, we grouped all of themiddle Miocene, all of the late Miocene, and all ofthe Plio-Pleistocene together into three bins (insteadof the eight stages that fall within these timeperiods). The range of a taxon preserved in onlyone time interval is one. Stratigraphic data aretabulated using first and last occurrences, so thatthe range of a taxon equals the number of timeintervals between its first and last occurrences,including the endpoints. Preservation probability R(the probability that a taxon is preserved at leastonce in an interval) was estimated by using thefrequency, f, of taxa with observed ranges of one,two, and three intervals to calculate the range-frequency ratio, FreqRat, as:
R � FreqRat ¼ f ð2Þ2.
f ð1Þf ð3Þ½ �
Estimated preservation probability must be con-sidered with respect to true durations, so calculatingR is insufficient to analyze the quality of the fossilrecord (Foote and Raup 1996; Alba et al. 2001).Therefore, the probability P1(T) that a taxon ispreserved at least once given a true original durationT must be calculated:
P1ðTÞ ¼ 1� 1� Rð ÞT
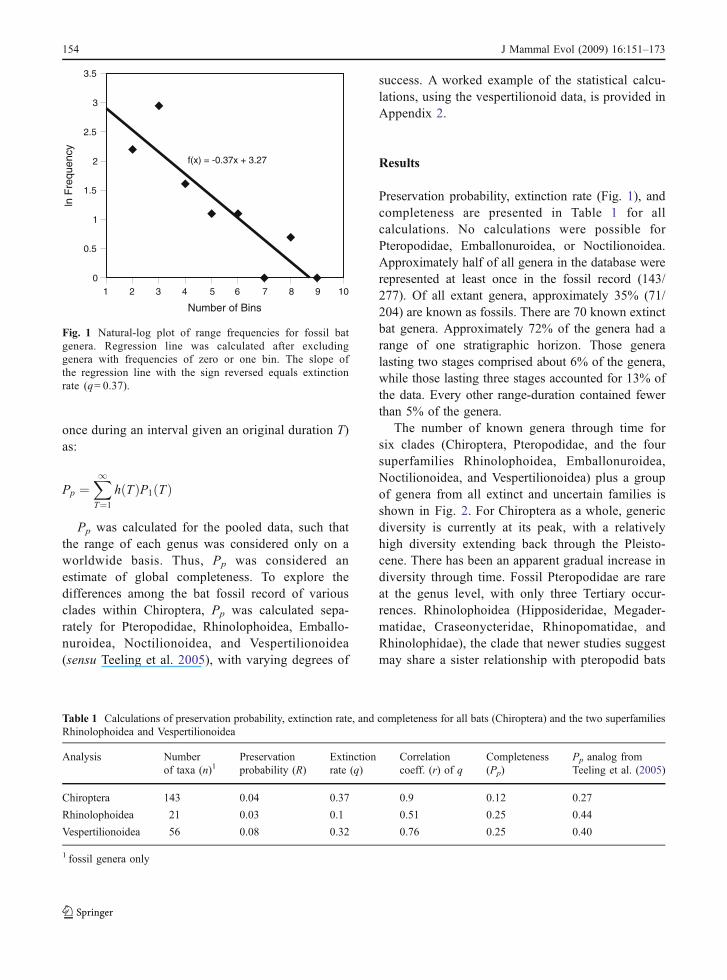
Original durations can be calculated assuming aconstant extinction rate q (and exponential distribu-tion of original durations), where q is calculatedfrom the ln-slope of range-frequencies (omitting taxathat occur in only one bin) as the slope with the signreversed (see Fig. 1). The probability h(T) that theoriginal distribution equals T is calculated as:
hðTÞ ¼ e�q T�1ð Þ � e�qT
Completeness Pp is calculated as the sum, for alloriginal durations (i.e., T=1-∞), of the productbetween h(T) (probability of having original durationT) and P1(T) (probability of being preserved at least
J Mammal Evol (2009) 16:151–173 153
once during an interval given an original duration T)as:
Pp ¼X1T¼1
hðTÞP1ðTÞ
Pp was calculated for the pooled data, such thatthe range of each genus was considered only on aworldwide basis. Thus, Pp was considered anestimate of global completeness. To explore thedifferences among the bat fossil record of variousclades within Chiroptera, Pp was calculated sepa-rately for Pteropodidae, Rhinolophoidea, Emballo-nuroidea, Noctilionoidea, and Vespertilionoidea(sensu Teeling et al. 2005), with varying degrees of
success. A worked example of the statistical calcu-lations, using the vespertilionoid data, is provided inAppendix 2.
Results
Preservation probability, extinction rate (Fig. 1), andcompleteness are presented in Table 1 for allcalculations. No calculations were possible forPteropodidae, Emballonuroidea, or Noctilionoidea.Approximately half of all genera in the database wererepresented at least once in the fossil record (143/277). Of all extant genera, approximately 35% (71/204) are known as fossils. There are 70 known extinctbat genera. Approximately 72% of the genera had arange of one stratigraphic horizon. Those generalasting two stages comprised about 6% of the genera,while those lasting three stages accounted for 13% ofthe data. Every other range-duration contained fewerthan 5% of the genera.
The number of known genera through time forsix clades (Chiroptera, Pteropodidae, and the foursuperfamilies Rhinolophoidea, Emballonuroidea,Noctilionoidea, and Vespertilionoidea) plus a groupof genera from all extinct and uncertain families isshown in Fig. 2. For Chiroptera as a whole, genericdiversity is currently at its peak, with a relativelyhigh diversity extending back through the Pleisto-cene. There has been an apparent gradual increase indiversity through time. Fossil Pteropodidae are rareat the genus level, with only three Tertiary occur-rences. Rhinolophoidea (Hipposideridae, Megader-matidae, Craseonycteridae, Rhinopomatidae, andRhinolophidae), the clade that newer studies suggestmay share a sister relationship with pteropodid bats
1 2 3 4 5 6 7 8 9 100
0.5
1
1.5
2
2.5
3
3.5
f(x) = -0.37x + 3.27
Number of Bins
ln F
requ
ency
Fig. 1 Natural-log plot of range frequencies for fossil batgenera. Regression line was calculated after excludinggenera with frequencies of zero or one bin. The slope ofthe regression line with the sign reversed equals extinctionrate (q=0.37).
Table 1 Calculations of preservation probability, extinction rate, and completeness for all bats (Chiroptera) and the two superfamiliesRhinolophoidea and Vespertilionoidea
Analysis Numberof taxa (n)1
Preservationprobability (R)
Extinctionrate (q)
Correlationcoeff. (r) of q
Completeness(Pp)
Pp analog fromTeeling et al. (2005)
Chiroptera 143 0.04 0.37 0.9 0.12 0.27
Rhinolophoidea 21 0.03 0.1 0.51 0.25 0.44
Vespertilionoidea 56 0.08 0.32 0.76 0.25 0.40
1 fossil genera only
154 J Mammal Evol (2009) 16:151–173
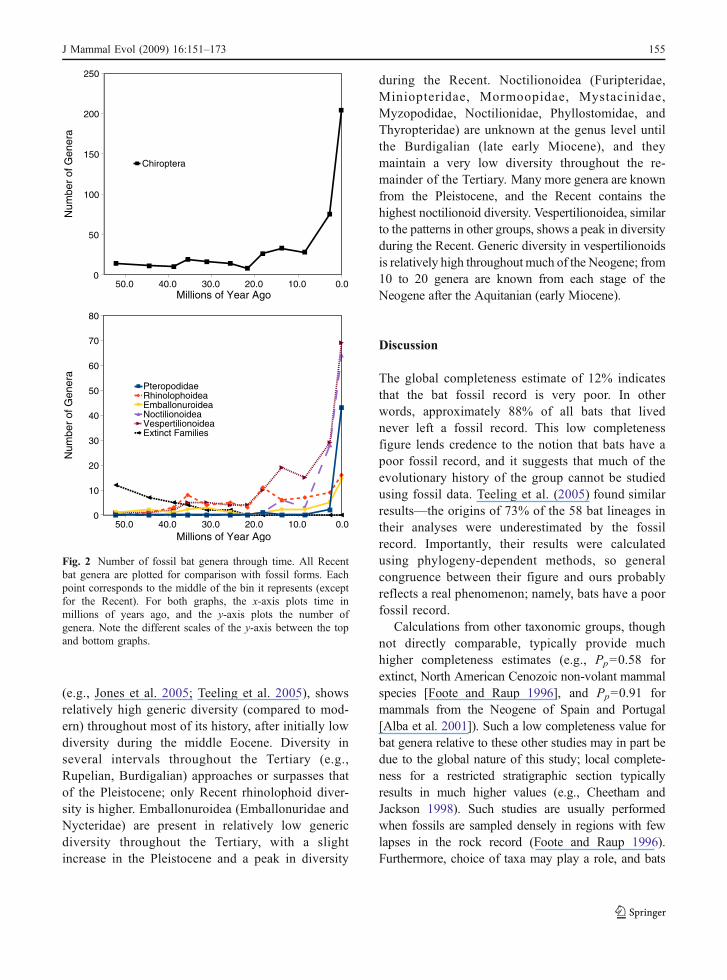
(e.g., Jones et al. 2005; Teeling et al. 2005), showsrelatively high generic diversity (compared to mod-ern) throughout most of its history, after initially lowdiversity during the middle Eocene. Diversity inseveral intervals throughout the Tertiary (e.g.,Rupelian, Burdigalian) approaches or surpasses thatof the Pleistocene; only Recent rhinolophoid diver-sity is higher. Emballonuroidea (Emballonuridae andNycteridae) are present in relatively low genericdiversity throughout the Tertiary, with a slightincrease in the Pleistocene and a peak in diversity
during the Recent. Noctilionoidea (Furipteridae,Miniopteridae, Mormoopidae, Mystacinidae,Myzopodidae, Noctilionidae, Phyllostomidae, andThyropteridae) are unknown at the genus level untilthe Burdigalian (late early Miocene), and theymaintain a very low diversity throughout the re-mainder of the Tertiary. Many more genera are knownfrom the Pleistocene, and the Recent contains thehighest noctilionoid diversity. Vespertilionoidea, similarto the patterns in other groups, shows a peak in diversityduring the Recent. Generic diversity in vespertilionoidsis relatively high throughout much of the Neogene; from10 to 20 genera are known from each stage of theNeogene after the Aquitanian (early Miocene).
Discussion
The global completeness estimate of 12% indicatesthat the bat fossil record is very poor. In otherwords, approximately 88% of all bats that livednever left a fossil record. This low completenessfigure lends credence to the notion that bats have apoor fossil record, and it suggests that much of theevolutionary history of the group cannot be studiedusing fossil data. Teeling et al. (2005) found similarresults—the origins of 73% of the 58 bat lineages intheir analyses were underestimated by the fossilrecord. Importantly, their results were calculatedusing phylogeny-dependent methods, so generalcongruence between their figure and ours probablyreflects a real phenomenon; namely, bats have a poorfossil record.
Calculations from other taxonomic groups, thoughnot directly comparable, typically provide muchhigher completeness estimates (e.g., Pp=0.58 forextinct, North American Cenozoic non-volant mammalspecies [Foote and Raup 1996], and Pp=0.91 formammals from the Neogene of Spain and Portugal[Alba et al. 2001]). Such a low completeness value forbat genera relative to these other studies may in part bedue to the global nature of this study; local complete-ness for a restricted stratigraphic section typicallyresults in much higher values (e.g., Cheetham andJackson 1998). Such studies are usually performedwhen fossils are sampled densely in regions with fewlapses in the rock record (Foote and Raup 1996).Furthermore, choice of taxa may play a role, and bats
0.010.020.030.040.050.00
50
100
150
200
250
Chiroptera
0.010.020.030.040.050.00
10
20
30
40
50
60
70
80
PteropodidaeRhinolophoideaEmballonuroideaNoctilionoideaVespertilionoideaExtinct Families
Millions of Year Ago
Millions of Year Ago
Num
ber
of G
ener
aN
umbe
r of
Gen
era
Fig. 2 Number of fossil bat genera through time. All Recentbat genera are plotted for comparison with fossil forms. Eachpoint corresponds to the middle of the bin it represents (exceptfor the Recent). For both graphs, the x-axis plots time inmillions of years ago, and the y-axis plots the number ofgenera. Note the different scales of the y-axis between the topand bottom graphs.
J Mammal Evol (2009) 16:151–173 155

are probably less easily preserved than othermammals for both anatomical (delicate skeletons)and taphonomic reasons (see discussion below).This effect may be partly responsible for the highproportion of single hits in the data set (72% of allfossil bats occur in just one bin). Foote and Raup(1996) warn that high proportions of single hits mayproduce very high variances in estimates of originaldurations, which produce large errors when calcu-lating completeness.
Fossil bats presumably inhabited environmentssimilar to their modern counterparts, which typicallyare not well sampled in the fossil record. Of thecommon bat habitats, caves offer some of the bestconditions for preservation, and many fossil bats areknown from cave and karst deposits (Kowalski 1995).For example, Tertiary bats from the Mediterraneanbasin typically come from karst deposits (Sigé andLegendre 1983) and many bats from the MioceneRiversleigh Site of Australia apparently come fromcave-fill deposits as well (Hand and Archer 2005).Other than caves, many of the best preserved fossilbats come from sedimentary deposits of extinct lakes(e.g., Jepsen 1966; Habersetzer and Storch 1987;Gunnell et al. 2003; Simmons et al. 2008); lacustrineenvironments provide some of the best conditions forfossilization in the terrestrial realm. Unfortunately,most bats visit lakes only briefly, so their potential forfossilization in these environments usually is verylow.
A rather striking result is the general matchbetween the superfamily-level results from Teelinget al. (2005) with those from this study (Fig. 2),though Teeling et al. found consistently highercompleteness figures for each clade (Table 1). Therank-order from most to least percent missing fromTeeling et al. (2005) is: Pteropodidae, Noctilionoi-dea, Emballonuroidea, Rhinolophoidea, and Vesper-tilionoidea. In the present study, completeness wascalculated for Rhinolophoidea and Vespertilionoidea(in addition to all Chiroptera). Calculations were notpossible for Pteropodidae, Noctilionoidea, or Embal-lonuroidea because of too little data. As Table 1shows, the calculations of extinction rate (q) yieldedPearson coefficients of correlation (between thenumber of stages and the natural log of thefrequency of genus occurrences) of 0.51 for Rhino-lophoidea and 0.76 for Vespertilionoidea. The highercorrelation for Vespertilionoidea may mean that the
extinction rate is more reliable as compared toRhinolophoidea. Furthermore, because the extinctionrate is used to calculate completeness, the complete-ness of Vespertilionoidea may be more robust than itis for Rhinolophoidea.
It is important to note that having too little datato calculate completeness for Pteropodidae, Nocti-lionoidea, and Emballonuroidea does not, by itself,mean that the fossil records for these three groupsare woefully incomplete. Our binning strategy mayhave been too aggressive in lumping together intothe same bin bats that were found in differentstratigraphic levels. If these three clades weregeologically rather young (middle or late Miocene,for example), then there would not be enoughbinned intervals to generate a sufficient statisticalsample to calculate completeness. However, noneof these three clades are young, as shown by theirfossil ages (Appendix 1, Fig. 2) and by phylogeneticanalyses (Teeling et al. 2005; Gunnell and Simmons2005). Examining the pattern of fossil occurrencesthrough time (Fig. 2) provides some insight into thequality of the record for each of these groups. Whencompared to their modern diversity, Pteropodidaeand Noctilionoidea seem to have the worst records,while Rhinolophoidea and Vespertilionoidea havethe best. If we compare groups of similar moderngeneric diversity, it seems that Vespertilionoidea hasa better record than Noctilionoidea, as does Rhino-lophoidea when compared to Emballonuroidea.Completeness values for Rhinolophoidea and Ves-pertilionoidea were twice that of Chiroptera as awhole (25% versus 12%). So, it is reasonable toexpect that groups other than the rhinolophoids andvespertilionoids would have completeness valuessubstantially lower than 12% (though parsing outcompleteness values for each of these other groupswas not possible in the present study).
Considering the apparent dual sister-group rela-tionships between Noctilionoidea and Vespertilio-noidea on the one hand, and Rhinolophoidea andPteropodidae on the other (e.g., Jones et al. 2005),the apparent disparity in the quality of their fossilrecords cannot be attributed to factors affecting allbats. In part, the time available for diversificationmay have differentially affected clades, consideringthat the generic-rich noctilionoid family Phyllosto-midae may be much younger than either Molossidaeor Vespertilionidae that together comprise most of
156 J Mammal Evol (2009) 16:151–173

the vespertilionoid diversity. But even in the Mio-cene—likely after the origin of the first phyllostomid(Jones et al. 2005)—the vespertilionoid recordcontains much more diversity than the noctilionoidrecord. So, the disparity in the fossil records may inpart be a real phenomenon that can be explained byat least three non-exclusive factors. First, it probablyreflects a collector bias, as vespertilionoids andrhinolophoids constitute a disproportionate share ofthe bat fossils known from Europe and NorthAmerica, both of which have a rather good samplingof fossil forms (Gunnell and Simmons 2005; Hulvaet al. 2007). An increased sampling effort throughoutthe Old and New World tropics may diminish theproblem of a depauperate bat fossil record, at least tosome degree. Second, differences in the quality ofthe fossil record between bat clades could be due todifferent taphonomic effects. Pteropodids and mostnoctilionoids today inhabit tropical regions in whichenvironmental conditions offer little hope of becom-ing fossils. For temperate bats, dominated byvespertilionoids, a large percentage inhabits cavesand faces fewer taphonomic impediments to fossil-ization immediately after death. Finally, the disparityin the fossil record of bat clades could be due to apoor rock record. In this case, it may be that diverseassemblages of poorly represented bat clades werefossilized, but these fossils have been buried ordestroyed as a result of subsequent geologic pro-cesses. Different bat clades could have experiencedhigh diversity at different points in time, and some ofthese time periods may have been preferentially lostin the rock record.
While numerous questions cannot be directlyaddressed with this study (e.g., paleobiogeographicpatterns), the results confirm the notion that thegenus-level fossil record of bats is poor. As such, itis unclear how incorporating extinct bat genera intophylogenetic analyses would really enhance theresulting phylogenetic hypotheses, though simula-tions indicate that, even in the face of very highdegrees of incompleteness, incorporating fossil datacan improve phylogeny estimation (Fox et al.1999). Some studies have shown a fairly high degreeof consistency between phylogenetic analyses thatdo and do not incorporate extinct forms and theirstratigraphic positions (e.g., Norell and Novacek1992); others have demonstrated that fossils play acritical role in phylogeny estimation (e.g., Gauthier
et al. 1988). Unfortunately, fossil bats often arerepresented by little more than isolated teeth or, atbest, mandibular or maxillary fragments (Czaplewskiet al. 2008). Though numerous parsimony-informa-tive characters are found in the skull and dentition,many diagnostic characters are found in other partsof the skeleton (e.g., Gunnell et al. 2003; Simmonset al. 2008), and an absence of sufficient materialmay preclude definitive taxonomic or characterassignment. Many important early bats cannot beassigned to genera (or even higher level groups) withany certainty (e.g., Archer 1978; Ducrocq et al.1993; Tejedor et al. 2005). Such specimens havebeen omitted from this study, as we await morediagnostic material. However, any estimate of thecompleteness of the fossil record is predicated on theassumption that taxonomic diagnoses have beenmade correctly, so this issue may not preferentiallybias our results.
Conclusions
This study marks the first phylogeny-independentestimate of the global completeness of the fossilrecord of bats. The genus-level bat fossil record isestimated to be 12% complete. This figure agreesbroadly with phylogeny-derived completeness esti-mates by Teeling et al. (2005) in confirming thenotion that the bat fossil record is poor. Such resultsindicate that much of the taxonomic and morphologicdiversity of bats remains to be uncovered. Despite thedepauperate fossil record, important steps in theevolutionary history of bats are recorded in fossils,such as the apparent evolution of flight before high-frequency echolocation in Eocene bats (Simmons etal. 2008). Future work should examine Tertiarydeposits from historically under-sampled regions toincrease our knowledge of fossil bats and thusimprove our understanding of this remarkable groupof mammals.
Acknowledgements We thank N. Czaplewski, I. Horáček, J.Hutcheon, G. Morgan, K. Samonds, and N. Simmons fordiscussions or providing reprints. We appreciate commentsfrom J. Wible and from an anonymous reviewer that greatlyimproved the quality of this manuscript. TE thanks G. Smithand D. Nelson, University of Michigan, for logistical supportand the Society of Vertebrate Paleontology for funds to attendthe 2007 annual meeting.
J Mammal Evol (2009) 16:151–173 157
Appendix 1
Tab
le2
Com
pilatio
nof
batfossilg
eneraused
inthisstud
y.Tem
poralrecords
ofbatg
eneraweretabu
latedas
present(1)
orabsent
(0)in
each
discretetim
einterval.S
ources,sho
wn
belowthetable,record
thefirstandlastoccurrencesof
each
genu
s(excluding
Recent),p
lusadditio
nalinterveningoccurrenceswhenavailable(non-exh
austive).S
econ
dary
sources
wereused
incaseswhere
prim
arysourceswereun
available.
Stratigraph
icbinabbreviatio
nsas
follo
ws:Eo=Eocene,Olig
o=Olig
ocene,
Mio
=Miocene,P-P
=Plio
cene
throug
hPleistocene,R
ec=Recent;Ypr
=Ypresian,
Lut
=Lutetian,
Bart=
Bartonian,P
ria=Priabon
ian,
Rup
=Rup
elian,
Chat=
Chattian,A
qui=
Aqu
itanian,B
urd=Burdigalian,
MMio
=MiddleMiocene,LMio
=LateMiocene.Ma=Millions
ofYears
Ago
.Records
forRecentbatswereno
tused
instatistical
calculations
Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Emballo
nuroidea
?Emballo
nuridae
†Epp
sinycteris
10
00
00
00
00
00
63
Emballo
nuroidea
Emballo
nuridae
†Dho
farella
00
01
10
00
00
00
44,118
Emballo
nuroidea
Emballo
nuridae
†Tachypteron
01
00
00
00
00
00
127
Emballo
nuroidea
Emballo
nuridae
†Vespertiliavus
01
11
11
00
00
00
6,35
,82
,114,
139
Emballo
nuroidea
Emballo
nuridae
Balan
tiopteryx
00
00
00
00
00
11
34
Emballo
nuroidea
Emballo
nuridae
Centron
ycteris
00
00
00
00
00
01
Emballo
nuroidea
Emballo
nuridae
Coleura
00
00
00
00
00
11
141
Emballo
nuroidea
Emballo
nuridae
Cormura
00
00
00
00
00
01
Emballo
nuroidea
Emballo
nuridae
Cyttarops
00
00
00
00
00
01
Emballo
nuroidea
Emballo
nuridae
Diclid
urus
00
00
00
00
11
11
18
Emballo
nuroidea
Emballo
nuridae
Emba
llonu
ra0
00
00
00
00
00
1
Emballo
nuroidea
Emballo
nuridae
Mosia
00
00
00
00
00
01
Emballo
nuroidea
Emballo
nuridae
Perop
teryx
00
00
00
00
00
11
24
Emballo
nuroidea
Emballo
nuridae
Rhyncho
nycteris
00
00
00
00
00
01
Emballo
nuroidea
Emballo
nuridae
Saccolaimus
00
00
00
00
00
11
13,14
1
Emballo
nuroidea
Emballo
nuridae
Saccop
teryx
00
00
00
00
00
01
Emballo
nuroidea
Emballo
nuridae
Taph
ozou
s0
00
00
00
11
11
111,76
Emballo
nuroidea
Nycteridae
†Chiba
nycteris
00
00
10
00
00
00
118
Emballo
nuroidea
Nycteridae
Nycteris
00
00
00
00
00
11
9,92
Extinct
?Hassianycteridae
†Cam
baya
10
00
00
00
00
00
124
Extinct
†Archaeony
cteridae
†Archa
eonycteris
11
10
00
00
00
00
97,10
3,12
0,12
4
Extinct
†Archaeony
cteridae
†Protonycteris
10
00
00
00
00
00
124
Appendix
1
158 J Mammal Evol (2009) 16:151–173

Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Extinct
†Hassianycteridae
†Hassian
ycteris
11
00
00
00
00
00
122,
123
Extinct
†Icarony
cteridae
†Icaronycteris
10
00
00
00
00
00
68,97
,10
3
Extinct
†Incertaecedis
†Ageina
10
00
00
00
00
00
103
Extinct
†Incertaecedis
†Alastorius
00
00
01
00
00
00
42,12
9
Extinct
†Incertaecedis
†Archa
eopterop
us0
00
01
00
00
00
042
,83
Extinct
†Incertaecedis
†Australon
ycteris
10
00
00
00
00
00
57
Extinct
†Incertaecedis
†Cha
dron
ycteris
00
01
00
00
00
00
91
Extinct
†Incertaecedis
†Jaegeria
10
00
00
00
00
00
124
Extinct
†Mixop
terygidae
†Carcinipteryx
01
11
00
00
00
00
81
Extinct
†Mixop
terygidae
†Mixop
teryx
00
01
11
00
00
00
81
Extinct
†Ony
chon
ycteridae
†Onychon
ycteris
10
00
00
00
00
00
121
Extinct
†Palaeochiropteryg
idae
†Cecilion
ycteris
01
00
00
00
00
00
59
Extinct
†Palaeochiropteryg
idae
†Lap
ichiropteryx
00
10
00
00
00
00
133
Extinct
†Palaeochiropteryg
idae
†Matthesia
01
00
00
00
00
00
116
Extinct
†Palaeochiropteryg
idae
†Microchirop
teryx
10
00
00
00
00
00
124
Extinct
†Palaeochiropteryg
idae
†Palaeochiropteryx
11
10
00
00
00
00
99,10
6
Extinct
† Tanzany
cteridae
†Tan
zanycteris
01
00
00
00
00
00
43
Noctiliono
idea
Furipteridae
Amorph
ochilus
00
00
00
00
00
11
86
Noctiliono
idea
Furipteridae
Furipterus
00
00
00
00
00
11
24
Noctiliono
idea
Mormoo
pidae
Mormoo
ps0
00
00
00
00
01
187
,119
Noctiliono
idea
Mormoo
pidae
Pterono
tus
00
00
00
00
00
11
24
Noctiliono
idea
Mystacinidae
†Icarops
00
00
00
01
10
00
56
Noctiliono
idea
Mystacinidae
Mystacina
00
00
00
00
00
11
10,14
6
Noctiliono
idea
Myzop
odidae
Myzop
oda
00
00
00
00
00
11
10
Noctiliono
idea
Noctilionidae
Noctilio
00
00
00
00
11
11
18,22
,31
Noctiliono
idea
Phy
llostom
idae
†Noton
ycteris
00
00
00
00
10
00
31,10
5
Noctiliono
idea
Phy
llostom
idae
†Palynephyllu
m0
00
00
00
01
00
031
Noctiliono
idea
Phy
llostom
idae
Ametrida
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Ano
ura
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Ardop
s0
00
00
00
00
00
1
Noctiliono
idea
Phy
llostom
idae
Ariteus
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Artibeus
00
00
00
00
00
11
24,34
Noctiliono
idea
Phy
llostom
idae
Brachyphylla
00
00
00
00
00
11
84,119
Noctiliono
idea
Phy
llostom
idae
Carollia
00
00
00
00
00
11
24
Noctiliono
idea
Phy
llostom
idae
Centurio
00
00
00
00
00
01
J Mammal Evol (2009) 16:151–173 159
Tab
le2
(con
tinued)
Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Noctiliono
idea
Phy
llostom
idae
Chiroderm
a0
00
00
00
00
01
124
Noctiliono
idea
Phy
llostom
idae
Cho
eron
iscus
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Cho
eron
ycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Chrotop
terus
00
00
00
00
00
11
24
Noctiliono
idea
Phy
llostom
idae
Desmod
us0
00
00
00
00
01
124
,26
,89
Noctiliono
idea
Phy
llostom
idae
Diaem
us0
00
00
00
00
00
1
Noctiliono
idea
Phy
llostom
idae
Diphylla
00
00
00
00
00
11
24
Noctiliono
idea
Phy
llostom
idae
Ectop
hylla
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Enchisthenes
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Eroph
ylla
00
00
00
00
00
11
87,119
Noctiliono
idea
Phy
llostom
idae
Glossop
haga
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Glyph
onycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Hylon
ycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Lam
pron
ycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Leptonycteris
00
00
00
00
00
11
34
Noctiliono
idea
Phy
llostom
idae
Licho
nycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Lionycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Lon
chop
hylla
00
00
00
00
00
11
24
Noctiliono
idea
Phy
llostom
idae
Lon
chorhina
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Lop
hostom
a0
00
00
00
00
01
130
Noctiliono
idea
Phy
llostom
idae
Macroph
yllum
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Macrotus
00
00
00
00
00
11
87,119
Noctiliono
idea
Phy
llostom
idae
Mesop
hylla
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Micronycteris
00
00
00
00
00
11
24,30
Noctiliono
idea
Phy
llostom
idae
Mimon
00
00
00
00
00
11
24
Noctiliono
idea
Phy
llostom
idae
Mon
ophyllu
s0
00
00
00
00
01
187
,119
Noctiliono
idea
Phy
llostom
idae
Muson
ycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Neonycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Phyllo
derm
a0
00
00
00
00
00
1
Noctiliono
idea
Phy
llostom
idae
Phyllo
nycteris
00
00
00
00
00
11
87,119
160 J Mammal Evol (2009) 16:151–173

Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Noctiliono
idea
Phy
llostom
idae
Phyllo
ps0
00
00
00
00
01
113
0
Noctiliono
idea
Phy
llostom
idae
Phyllo
stom
us0
00
00
00
00
01
124
Noctiliono
idea
Phy
llostom
idae
Platalin
a0
00
00
00
00
00
1
Noctiliono
idea
Phy
llostom
idae
Platyrrhinu
s0
00
00
00
00
01
124
Noctiliono
idea
Phy
llostom
idae
Pygod
erma
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Rhino
phylla
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Scleronycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Spha
eron
ycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Stenod
erma
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Sturnira
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Tona
tia0
00
00
00
01
11
118
,24
Noctiliono
idea
Phy
llostom
idae
Tracho
ps0
00
00
00
00
01
130
Noctiliono
idea
Phy
llostom
idae
Trinycteris
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Uroderm
a0
00
00
00
00
00
1
Noctiliono
idea
Phy
llostom
idae
Vampyressa
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Vampyrodes
00
00
00
00
00
01
Noctiliono
idea
Phy
llostom
idae
Vampyrum
00
00
00
00
00
01
Noctiliono
idea
Thy
ropteridae
Thyroptera
00
00
00
00
11
11
18
Pteropo
didae
Pteropo
didae
†Propo
tto0
00
00
00
10
00
011,14
0
Pteropo
didae
Pteropo
didae
Acerodo
n0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Aetha
lops
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Alio
nycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Aproteles
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Balionycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Casinycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Chirona
x0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Cynop
terus
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Dob
sonia
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Dyacopterus
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Eidolon
00
00
00
00
00
11
141
Pteropo
didae
Pteropo
didae
Eon
ycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Epo
mop
horus
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Epo
mop
s0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Hap
lonycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Harpyionycteris
00
00
00
00
00
01
J Mammal Evol (2009) 16:151–173 161
Tab
le2
(con
tinued)
Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Pteropo
didae
Pteropo
didae
Hypsign
athu
s0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Latidens
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Lissonycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Macroglossus
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Megaerops
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Megalog
lossus
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Melon
ycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Micropterop
us0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Myonycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Nan
onycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Neopteryx
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Notop
teris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Nyctim
ene
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Otopterop
us0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Paran
yctim
ene
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Penthetor
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Plerotes
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Pteno
chirus
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Pteralopex
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Pteropu
s0
00
00
00
00
00
1
Pteropo
didae
Pteropo
didae
Rou
settu
s0
00
00
00
00
01
110
4
Pteropo
didae
Pteropo
didae
Scoton
ycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Spha
erias
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Styloctenium
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Syconycteris
00
00
00
00
00
01
Pteropo
didae
Pteropo
didae
Tho
opterus
00
00
00
00
00
01
Rhino
loph
oidea
Craseon
ycteridae
Craseon
ycteris
00
00
00
00
00
01
Rhino
loph
oidea
Hippo
sideridae
†Archerops
00
00
00
01
00
00
55
Rhino
loph
oidea
Hippo
sideridae
†Brevipa
latus
00
00
00
01
00
00
54,55
Rhino
loph
oidea
Hippo
sideridae
†Miophyllorhina
00
00
00
01
00
00
50
162 J Mammal Evol (2009) 16:151–173

Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Rhino
loph
oidea
Hippo
sideridae
†Palaeop
hyllo
phora
00
11
11
00
00
00
35
Rhino
loph
oidea
Hippo
sideridae
†Parap
hyllo
phora
00
01
00
00
00
00
101
Rhino
loph
oidea
Hippo
sideridae
†Riversleigh
a0
00
00
00
10
00
052
Rhino
loph
oidea
Hippo
sideridae
†Vaylatsia
00
01
11
00
00
00
55,114,
120,
139
Rhino
loph
oidea
Hippo
sideridae
†Xenorhino
s0
00
00
00
10
00
053
Rhino
loph
oidea
Hippo
sideridae
Antho
ps0
00
00
00
00
00
1
Rhino
loph
oidea
Hippo
sideridae
Asellia
00
00
00
01
11
11
37,74
,77
,12
6
Rhino
loph
oidea
Hippo
sideridae
Aselliscus
00
00
00
00
00
01
Rhino
loph
oidea
Hippo
sideridae
Cloeotis
00
00
00
00
00
01
Rhino
loph
oidea
Hippo
sideridae
Coelops
00
00
00
00
00
01
Rhino
loph
oidea
Hippo
sideridae
Hippo
sideros
00
11
11
11
11
11
4,11,35
,47
,49
,51
,55
,69
,77
,10
4,10
8,110,
117,
118,
126,
141,
151,
152
Rhino
loph
oidea
Hippo
sideridae
Paracoelops
00
00
00
00
00
01
Rhino
loph
oidea
Hippo
sideridae
Rhino
nicteris
00
00
00
01
11
11
51
Rhino
loph
oidea
Hippo
sideridae
Triaenop
s0
00
00
00
00
01
110
4
Rhino
loph
oidea
?Megadermatidae
†Necroman
tis0
11
10
00
00
00
082
Rhino
loph
oidea
Megadermatidae
†Sah
arad
erma
00
01
00
00
00
00
44
Rhino
loph
oidea
Megadermatidae
Cardiod
erma
00
00
00
00
00
11
10
Rhino
loph
oidea
Megadermatidae
Lavia
00
00
00
00
00
01
Rhino
loph
oidea
Megadermatidae
Macroderm
a0
00
00
00
01
11
145
,48
,90
Rhino
loph
oidea
Megadermatidae
Megad
erma
00
00
01
11
11
11
37,39
,10
8,10
9,110,
112,
126,
135,
151,
152
Rhino
loph
oidea
Rhino
loph
idae
†Palaeon
ycteris
00
00
00
01
00
00
44
Rhino
loph
oidea
Rhino
loph
idae
Rhino
loph
us0
00
11
11
11
11
11,
3,4,
35,37
,60
,69
,71
,73
,74
,93
,10
7,10
8,10
9,12
6,13
4,13
7,15
1,15
2
Rhino
loph
oidea
Rhino
pomatidae
†Qarun
ycteris
00
01
00
00
00
00
44
Rhino
loph
oidea
Rhino
pomatidae
Rhino
poma
00
00
00
00
01
11
67
Vespertilion
oidea
†Philisidae
†Dizzya
10
00
00
00
00
00
115
Vespertilion
oidea
†Philisidae
†Philisis
00
00
10
00
00
00
118
Vespertilion
oidea
†Philisidae
†Scotoph
ilisis
00
00
00
01
00
00
66,14
2
Vespertilion
oidea
†Philisidae
†Vam
pyravus
00
00
10
00
00
00
113,
120
Vespertilion
oidea
†Philisidae
†Witw
atia
00
01
00
00
00
00
44
J Mammal Evol (2009) 16:151–173 163
Tab
le2
(con
tinued)
Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Vespertilion
oidea
Miniopteridae
Miniopterus
00
00
00
00
11
11
1,4,
60,71
,93
,10
7,12
6,14
5,15
2
Vespertilion
oidea
Molossidae
†Cuvierimop
s0
00
11
10
00
00
042
,78
Vespertilion
oidea
Molossidae
†Petramop
s0
00
00
00
01
00
046
Vespertilion
oidea
Molossidae
†Potam
ops
00
00
00
00
10
00
18,23
Vespertilion
oidea
Molossidae
†Wallia
00
11
00
00
00
00
28,12
8
Vespertilion
oidea
Molossidae
Cha
erepho
n0
00
00
00
00
00
1
Vespertilion
oidea
Molossidae
Cheirom
eles
00
00
00
00
00
01
Vespertilion
oidea
Molossidae
Cynom
ops
00
00
00
00
00
01
Vespertilion
oidea
Molossidae
Eum
ops
00
00
00
00
11
11
18,21
,23
,24
,10
1
Vespertilion
oidea
Molossidae
Molossops
00
00
00
00
00
01
Vespertilion
oidea
Molossidae
Molossus
00
00
00
00
00
11
24
Vespertilion
oidea
Molossidae
Mop
s0
00
00
00
00
00
1
Vespertilion
oidea
Molossidae
Mormop
terus
00
00
00
11
11
11
18,23
,78
,12
6
Vespertilion
oidea
Molossidae
Myopterus
00
00
00
00
00
01
Vespertilion
oidea
Molossidae
Nyctin
omop
s0
00
00
00
00
01
122
,24
Vespertilion
oidea
Molossidae
Otomop
s0
00
00
00
00
00
1
Vespertilion
oidea
Molossidae
Platymop
s0
00
00
00
00
00
1
Vespertilion
oidea
Molossidae
Promop
s0
00
00
00
00
01
124
Vespertilion
oidea
Molossidae
Saurom
ys0
00
00
00
00
00
1
Vespertilion
oidea
Molossidae
Tada
rida
00
01
11
11
11
11
1,2,
24,29
,35
,78
,87
,88
,10
7,10
9,12
6
Vespertilion
oidea
Molossidae
Tomop
eas
00
00
00
00
00
01
Vespertilion
oidea
Natalidae
†Hon
rovits
10
00
00
00
00
00
7
Vespertilion
oidea
Natalidae
†Primon
atalus
00
00
00
01
00
00
87
Vespertilion
oidea
Natalidae
Chilona
talus
00
00
00
00
00
01
Vespertilion
oidea
Natalidae
Natalus
00
00
00
00
00
11
24,87
Vespertilion
oidea
Natalidae
Nyctiellu
s0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
†Ancenycteris
00
00
00
00
10
00
131
Vespertilion
oidea
Vespertilion
idae
†Anzan
ycteris
00
00
00
00
00
10
15,28
,14
3
164 J Mammal Evol (2009) 16:151–173

Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Vespertilion
oidea
Vespertilion
idae
†Cha
mtwaria
00
00
00
01
00
00
11
Vespertilion
oidea
Vespertilion
idae
†Han
akia
00
00
01
11
00
00
65
Vespertilion
oidea
Vespertilion
idae
†Karstala
00
00
00
01
00
00
25
Vespertilion
oidea
Vespertilion
idae
†Kho
nsun
ycteris
00
01
00
00
00
00
44
Vespertilion
oidea
Vespertilion
idae
†Miomyotis
00
00
00
01
00
00
75
Vespertilion
oidea
Vespertilion
idae
†Miostrellu
s0
00
00
00
01
00
065
,12
6
Vespertilion
oidea
Vespertilion
idae
†Olig
omyotis
00
00
10
00
00
00
5,38
Vespertilion
oidea
Vespertilion
idae
†Paleptesicus
00
00
00
00
11
00
126,
149,
150,
152
Vespertilion
oidea
Vespertilion
idae
†Pleistomyotis
00
00
00
00
00
10
148
Vespertilion
oidea
Vespertilion
idae
†Plio
nycteris
00
00
00
00
00
10
80
Vespertilion
oidea
Vespertilion
idae
†Potam
onycteris
00
00
00
00
10
00
20
Vespertilion
oidea
Vespertilion
idae
†Quinetia
00
00
10
00
00
00
65,96
Vespertilion
oidea
Vespertilion
idae
†Sam
onycteris
00
00
00
00
01
00
101,
126
Vespertilion
oidea
Vespertilion
idae
†Sha
nwan
gia
00
00
00
00
10
00
147
Vespertilion
oidea
Vespertilion
idae
†Simon
ycteris
00
00
00
00
00
10
20,12
5
Vespertilion
oidea
Vespertilion
idae
†Stehlinia
01
11
11
00
00
00
35,82
,10
0,111
Vespertilion
oidea
Vespertilion
idae
†Sua
ptenos
00
00
00
01
00
00
75
Vespertilion
oidea
Vespertilion
idae
†Sub
myotodo
n0
00
00
00
01
00
015
2
Vespertilion
oidea
Vespertilion
idae
Antrozous
00
00
00
00
11
11
28,33
,13
2
Vespertilion
oidea
Vespertilion
idae
Arielulus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Barba
stella
00
00
00
00
00
11
72,10
7
Vespertilion
oidea
Vespertilion
idae
Bau
erus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Cha
linolob
us0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Cistugo
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Corynorhinu
s0
00
00
00
00
11
158
,13
8,13
9
Vespertilion
oidea
Vespertilion
idae
Eptesicus
00
00
00
01
11
11
3,14
,21
,24
,30
,32
,33
,61
,70
,72
,79
,87
,92
,95
,10
7,13
0,13
7,15
1,15
2
Vespertilion
oidea
Vespertilion
idae
Eud
erma
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Eud
iscopu
s0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Falsistrellu
s0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Glaucon
ycteris
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Glischropu
s0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Harpiocepha
lus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Hesperoptenus
00
00
00
00
00
01
J Mammal Evol (2009) 16:151–173 165
Tab
le2
(con
tinued)
Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Vespertilion
oidea
Vespertilion
idae
Histio
tus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Hypsugo
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Ia0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Idionycteris
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Kerivou
la0
00
00
00
00
01
116
,64
Vespertilion
oidea
Vespertilion
idae
Laeph
otis
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Lasionycteris
00
00
00
00
00
11
32
Vespertilion
oidea
Vespertilion
idae
Lasiurus
00
00
00
00
01
11
21,27
,32
,41
,62
,13
2
Vespertilion
oidea
Vespertilion
idae
Mimetillus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Murina
00
00
00
00
00
11
72,94
Vespertilion
oidea
Vespertilion
idae
Myotis
00
00
01
11
11
11
1,3,
5,17
,19
,24
,27
,35
,40
,58
,60
,69
,71
,72
,73
,93
,94
,95
,10
1,10
7,110,
126,
136,
137,
144,
151,
152
Vespertilion
oidea
Vespertilion
idae
Neoromicia
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Nyctalus
00
00
01
11
11
11
65,10
7
Vespertilion
oidea
Vespertilion
idae
Nycticeino
ps0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Nycticeius
00
00
00
00
00
11
12
Vespertilion
oidea
Vespertilion
idae
Nyctoph
ilus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Otonycteris
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Parastrellus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Perimyotis
00
00
00
00
00
11
85,98
Vespertilion
oidea
Vespertilion
idae
Pha
rotis
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Philetor
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Pho
niscus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Pipistrellus
00
00
00
00
01
11
12,95
,10
7,10
9,13
7
Vespertilion
oidea
Vespertilion
idae
Plecotus
00
00
00
00
11
11
60,70
,72
,73
,85
,95
,10
7,13
9,15
2
Vespertilion
oidea
Vespertilion
idae
Rho
geesa
00
00
00
00
00
11
30
166 J Mammal Evol (2009) 16:151–173

Sup
erfamily
Fam
ilyGenus
Eo
Olig
oMio
P-P
Rec
Sou
rce
Ypr
Lut
Bart
Pria
Rup
Chat
Aqu
iBurd
MMio
LMio
P-P
Rec
Ma:
55.8
48.6
40.4
37.2
33.9
28.4
23.0
20.4
16.0
11.6
5.3
0.01
2
Vespertilion
oidea
Vespertilion
idae
Scoteana
x0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Scotoecus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Scotom
anes
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Scotop
hilus
00
00
00
00
11
11
9,36
Vespertilion
oidea
Vespertilion
idae
Scotorepens
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Scotozou
s0
00
00
00
00
00
1
Vespertilion
oidea
Vespertilion
idae
Tylonicteris
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Vespad
elus
00
00
00
00
00
01
Vespertilion
oidea
Vespertilion
idae
Vespertilio
00
00
00
00
11
11
37,10
2
1:Agu
ilaretal.1
984;
2:Arroy
o-Cabralesetal.2
002;
3:Avery
1998
;4:
Avery
2003
;5:
Bailey20
04;6:
Bargh
oorn
1977;7:
Beard
etal.1
992;
8:BiochroM’9719
97;9:
Black
and
Krishtalka19
86;10:Butler19
78;11:ButlerandHop
woo
d19
57;12:ButlerandGreenwoo
d19
65;13:Butler19
84;14:Cahn19
39;15:Cassiliano
1999;16
:Čermák
etal.20
07;
17:Cho
ateandHall19
67;18:Cozzuol
2006
;19:Czaplew
ski19
87;20:Czaplew
ksi19
91;21
:Czaplew
ski19
93;22
:Czaplew
ski19
96;23
:Czaplew
ski19
97;24
:Czaplew
ksiand
Cartelle
1998
;25
:Czaplew
skiandMorgan20
00;26
:Czaplew
skiandPeachey
2003
;27:Czaplew
skiet
al.19
99;28:Czaplew
skiet
al.20
08;29
:Czaplew
skiet
al.20
03a;
30:
Czaplew
skietal.2
005;
31:Czaplew
skietal.2
003b;32:Dalqu
est19
78;33:Dalqu
est19
83;34
:Dalqu
estandRoth19
70;35
:deBon
isetal.1
973;
36:Eng
esser19
72;37
:Eng
esser
andZiegler
1996
;38:
Galbreath
1962
;39:
Ginsburg19
63;4
0:God
awa19
93;4
1:Grady
andOlson
2006
;42:
Gun
nellandSim
mon
s20
05;4
3:Gun
nelletal.2
003;
44:G
unnelletal.
2008
;45
:Hand19
85;46:Hand19
90;47:Hand19
93;48
:Hand19
96;49:Hand19
97a;50
:Hand19
97b;
51:Hand19
97c;52
:Hand19
98a;53:Hand19
98b;
54:HandandArcher
2005
;55:
HandandKirsch20
03;5
6:Handetal.1
998;
57:Handetal.1
994;
58:Handley
1959;59:Heller19
35;6
0:Heller19
36;61:Hendey19
81;6
2:Hibbard
1950
;63:
Hoo
ker
1996
;64:
Horáček
1986
;65:
Horáček
2001
;66:
Horáček
etal.200
6;67:H
ulva
etal.2
007;
68:Jepsen19
66;6
9:Jinetal.2
000;
70:K
ormos
1930
;71:
Kow
alski1
956;
72:K
owalski
1962
a;73:Kow
alski19
62b;
74:Lavocat
1961;75:Law
rence19
43;76:Legendre19
80;77:Legendre19
82;78
:Legendre19
85;79:Lem
onandChu
rcher19
61;80
:Lindsay
and
Jacobs
1985
;81
:Maitreet
al.20
08;82:Marandatet
al.19
93;83
:Meschinelli19
03;84:Morgan19
89;85:Morgan19
91;86:MorganandCzaplew
ski19
99;87
:Morganand
Czaplew
ski2
003;
88:M
organandRidgeway
1987
;89:
Morganetal.1
988;
90:M
yersetal.2
001;
91:O
strand
er19
83;9
2:Pocock19
87;9
3:Pop
ov20
04;9
4:Qiu
andStorch20
00;
95:Q
iuetal.1
985;
96:Q
uinet1
965;
97:R
anaetal.2
005;
98:R
ay19
67;9
9:Revilliod19
17;1
00:R
evilliod19
19;1
01:R
evilliod19
22;1
02:R
ossina
etal.2
006;
103:
Russelletal.
1973
;10
4:Sam
onds
2007
;10
5:Savage19
51;10
6:SavageandRussell19
83;10
7:Sevilla19
89;10
8:Sevilla19
90;10
9:Sevilla19
91;110:
Sigé19
68;111:
Sigé19
74;112:
Sigé
1976
;113:S
igé19
85;114:S
igé19
90;115:S
igé19
91;116:S
igéandRussell19
80;117:S
igéetal.1
982;
118:
Sigéetal.1
994;
119:
Silv
a-Taboada
1974
;120:S
immon
sandGeisler
1998
;12
1:Sim
mon
set
al.20
08;12
2:Smith
andStorch19
81;12
3:Smith
andRussell19
92;12
4:Smith
etal.20
07;12
5:Stirton19
31;12
6:Storch19
99;12
7:Storchet
al.20
02;
128:
Storer19
84;12
9:Strand19
28;13
0:SuárezandDíaz-Franco20
03;1
31:S
uttonandGenow
ays19
74;13
2:ThewissenandSmith
1987;13
3:Ton
g19
97;1
34:Top
ál19
63;13
5:Top
ál19
74;13
6:Top
ál19
83;13
7:Top
ál19
85;13
8:Top
ál19
89a;
139:
Top
ál19
89b;
140:
Walker19
69;14
1:Wesselm
ann19
84;14
2:Wesselset
al.20
03;14
3:White
1969
;14
4:Wilson
1968;14
5:Wołoszyn19
86;14
6:WorthyandHoldaway
1994;14
7:Yang19
77;14
8:Yoo
net
al.19
84;14
9:Zapfe
1950
;15
0:Zapfe
1970
;15
1:Ziegler
2003
;15
2:Ziegler
1994
J Mammal Evol (2009) 16:151–173 167

Appendix 2
Statistical calculations for Vespertilionoidea, follow-ing the method of Foote and Raup (1996).
Using the data for all vespertilionoid bats(Appendix 1), we first computed the total numberof temporal bins in which each genus occurs. Thereare 11 stratigraphic bins (excluding the Recent),beginning with the Ypresian (early Eocene) andending with the Plio-Pleistocene. Of the 56 vesperti-lionoid genera that occur as fossils, 38 occur in onlyone bin, five range through two bins, eight occur inthree bins, one occurs in four bins, two occur in fivebins, and one each occurs in six (Myotis) and eight(Tadarida) bins. No vespertilionoid bats lastedexactly seven bins, or in any more than eight bins.
To estimate preservation probability, R, we calcu-late the FreqRat (which estimates R) following theformula:
R � FreqRat ¼ f ð2Þ2.
f ð1Þf ð3Þ½ �
where f(x) is the frequency of genera with observedrange-durations of x bins. For Vespertilionoidea,R � FreqRat ¼ 52
�38ð Þð8Þ½ � ¼ 0:082.
To account for differential original temporal dis-tributions of bat genera, R must be scaled according tooriginal durations. Estimated preservation probabilityis used to calculate an estimate of the probabilityP1(T) that a taxon is preserved at least once given atrue original duration T. The probability that a genuswith original duration T is not at all preserved equals(1-R)T. Thus, the probability that it is preserved atleast once equals:
P1ðTÞ ¼ 1� 1� Rð ÞT
For example, to calculate the probability that a genusis preserved given a true original duration of three bins,the equation became P1ð3Þ ¼ 1� 1� 0:082ð Þ3¼0:0227. This same procedure was used for all possiblevalues of T (i.e., T=1-∞, though in this and subsequentcalculations it is usually sufficient to take out to justseveral hundred).
We next estimated extinction rate, q, which was tobe used to compute original genus durations. We usedlinear regression on the ln-transformed frequencies ofgenera with a range of two or more bins. The slope ofthe linear regression line equals q (the extinction rate)with the sign reversed. Next, original durations were
calculated, assuming a constant extinction rate, q, andan exponential distribution of original durations. qwas calculated from the ln-slope of range-frequencies(omitting taxa that occur in only one bin) as the slopewith the sign reversed. For Vespertilionoidea, the ln-slope of range-frequency distributions is −0.3226, sothe extinction rate q equals 0.3226.
Using our estimate for q, we calculated the prob-ability, h(T), that the original distribution equaled T as:
hðTÞ ¼ e�q T�1ð Þ � e�qT
Using T of 3 again, we see that hð3Þ ¼e� 0:3226ð Þ 3�1ð Þ� e� :3226ð Þð3Þ ¼ 0:145. Again, this proce-dure was calculated for all values of T.
Finally, to calculate completeness, we computed thesum, for all original durations, T, of the product betweenh(T) (probability of having original duration T) andP1(T) (probability of being preserved at least onceduring an interval given an original duration T) as:
Pp ¼X1T¼1
hðTÞP1ðTÞ
As T increased, the completeness for that number ofbins decreases rapidly, so this summation was to onlybe taken to several hundred rows in a spreadsheet.Doing this for Vespertilionoidea to 900 rows gave acompleteness of 24.5% (rounded and included inTable 1). Summing the completeness calculations upto only 50 rows produced the same overall complete-ness value (to eight digits), indicating that it issufficient to compute the summation using a finite setof values for T that is substantially less than 1000.
References
Aguilar J-P, Brandy LD, Thaler L (1984) Les rongeurs deSalobreña (sud de l’Espagne) et le probleme de lamigration Messinienne. Paléobiol Cont 14:3–17
Agustí J, Cabrera L, Garcés M, Krijgsman W, Oms O, Parés JM(2001) A calibrated mammal scale for the Neogene ofWestern Europe. State of the Art. Earth-Sci Rev 52:247–260
Alba DM, Agustí J, Moyà-Solà S (2001) Completeness of themammalian fossil record in the Iberian Neogene. Paleobi-ology 27:79–83
Alroy J (2000) New methods for quantifying macroevolution-ary patterns and processes. Paleobiology 26:707–733
Archer M (1978) Australia’s oldest bat, a possible rhinolophid.Proc Roy Soc Queensl 89:23–24
168 J Mammal Evol (2009) 16:151–173

Arroyo-Cabrales J, Gregorin R, Schlitter DA, Walker A (2002)The oldest African molossid bat cranium (Chiroptera:Molossidae). J Vertebr Paleontol 22:380–387
Avery DM (1998) An assessment of the lower Pleistocenemicromammalian fauna from Swartkrans Members 1–3,Gauteng, South Africa. Géobios 31:393–414
Avery DM (2003) Early and middle Pleistocene environmentsand hominid biogeography; micromammalian evidencefrom Kabwe, Twin Rivers and Mumbwa Caves in centralZambia. Palaeogeogr Palaeoclimatol Palaeoecol 189:55–69
Bailey BE (2004) Biostratigraphy and biochronology of earlyArikareean through late Hemingfordian small mammalfaunas from the Nebraska panhandle and adjacent areas.Paludicola 4:81–113
Barghoorn SF (1977) New material of Vespertiliavus Schlosser(Mammalia, Chiroptera) and suggested relationships ofemballonurid bats based on cranial morphology. Am MusNovitates 2618:1–29
Beard KC, Sigé B, Krishtalka L (1992) A primitive vesperti-lionoid bat from the early Eocene of central Wyoming. CR Acad Sci Paris, sér. 2a 314:735–741
Benton MJ (1998) The quality of the fossil record of thevertebrates. In: Donovan SK, Paul CRC (eds) TheAdequacy of the Fossil Record. John Wiley & Sons,Chichester, pp 269–303
BiochroM’97 (1997) Biochronologie mammalienne du Cén-ozoïque en Europe at domaines reliés. Synthèses ettableaux de corrélations. In: Aguilar JP, Legendre S, andMichaux J (eds) Actes du Congrès BiochroM’97. MémTrav E P H E Inst, Montpellier, France, vol. 21:769–805
Black CC, Krishtalka L (1986) Rodents, bats, and insectivoresfrom the Plio-Pleistocene sediment to the East of LakeTurkana, Kenya. Nat Hist Mus LA Co, Contrib Sci 372:1–15
Butler PM (1978) Insectivora and Chiroptera. In: Maglio VJ,Cooke HBS (eds) Evolution of African Mammals.Harvard University Press, Cambridge, pp 56–68
Butler PM (1984) Macroscelidea, Insectivora and Chiropterafrom the Miocene of East Africa. Palaeovertebrata14:117–200
Butler PM, Greenwood M (1965) Insectivora and Chiroptera.In: Leakey LSB (ed) Olduvai Gorge 1951–1961. Vol. 1.Fauna and Background. Cambridge University Press,Cambridge, pp 13–15
Butler PM, Hopwood AT (1957) Insectivora and Chiropterafrom the Miocene rocks of Kenya colony. Fossil MammalsAfr 13:1–35
Cahn AR (1939) Pleistocene fossils from a cave in AndersonCounty, Tennessee. J Mammal 20:248–250
Cassiliano ML (1999) Biostratigraphy of Blancan and Irving-tonian mammals in the Fish Creek-Vallecito Creek section,southern California, and a review of the Blancan-Irvingto-nian boundary. J Vertebr Paleontol 19:169–186
Čermák S, Wagner J, Fejfar O, Horáček I (2007) New Pliocenelocalities with micromammals from the Czech Republic: apreliminary report. Fossil Rec 10:60–68
Cheetham AJ, Jackson JBC (1998) The fossil record ofcheilostome Bryozoa in the Neogene and Quaternary oftropical America: adequacy for phylogenetic and evolu-tionary studies. In: Donovan SK, Paul CRC (eds) The
Adequacy of the Fossil Record. John Wiley & Sons,Chichester, pp 227–242
Choate JR, Hall ER (1967) Two new species of bats, genusMyotis, from a Pleistocene deposit in Texas. Am Mid Nat78:531–534
Clyde WC, Gingerich PD (1994) Rates of evolution in thedentition of early Eocene Cantius: comparison of size andshape. Paleobiology 20:506–522
Cozzuol MA (2006) The Acre vertebrate fauna: age, diversity,and geography. J So Amer Earth Sci 21:185–203
Czaplewski NJ (1987) Middle Blancan vertebrate assemblagefrom the Verde Formation, Arizona. Contrib Geol, UnivWyoming 25:133–155
Czaplewksi NJ (1991) Miocene bats from the lower ValentineFormation of northeastern Nebraska. J Mammal 72:715–722
Czaplewski NJ (1993) Late Tertiary bats (Mammalia, Chirop-tera) from the southwestern United States. Southwest Nat38:111–118
Czaplewski NJ (1996) Opossums (Didelphidae) and bats(Noctilionidae and Molossidae) from the late Miocene ofthe Amazon basin. J Mammal 77:84–94
Czaplewski NJ (1997) Chiroptera. In: Kay RF, Madden RH,Cifelli RL, Flynn JJ (eds) Vertebrate Paleontology in theNeotropics: The Miocene fauna of La Venta, Colombia.Smithsonian Institution Press, Washington, pp 410–431
Czaplewski NJ, Bailey BE, Corner RG (1999) Tertiary bats(Mammalia: Chiroptera) from northern Nebraska. TransNebr Acad Sci 25:83–93
Czaplewksi NJ, Cartelle C (1998) Pleistocene bats from cavedeposits in Bahia, Brazil. J Mammal 79:784–803
Czaplewski NJ, Morgan GS (2000) A new vespertilionid bat(Mammalia: Chiroptera) from the early Miocene (Hemi-ngfordian) of Florida, USA. J Vertebr Paleontol 20:736–742
Czaplewski NJ, Morgan GS, McLeod SA (2008) Chiroptera.In: Janis CM, Gunnell GF, Uhen MD (eds) Evolution ofTertiary mammals of North America. Volume 2. Smallmammals, xenarthrans, and marine mammals. CambridgeUniversity Press, Cambridge, pp 174–197
Czaplewski NJ, Morgan GS, Naeher T (2003a) Molossid batsfrom the late Tertiary of Florida with a review of theTertiary Molossidae of North America. Acta Chiropterolog5:61–74
Czaplewski NJ, Takai M, Naeher TM, Shigehara N, SetoguchiT (2003b) Additional bats from the Middle Miocene LaVenta Fauna of Colombia. Rev Acad Colomb Cienc27:263–282
Czaplewski NJ, Peachey WD (2003) Late Pleistocene bats fromArkenstone Cave, Arizona. Southwest Nat 48:597–609
Czaplewski NJ, Rincón AD, Morgan GS (2005) Fossil bat(Mammalia: Chiroptera) remains from Inciarte Tar Pit,Sierra de Perijá, Venezuela. Caribb J Sci 41:768–781
Dalquest WW (1978) Early Blancan mammals of the BeckRanch Local Fauna of Texas. J Mammal 59:269–298
Dalquest WW (1983) Mammals of the Coffee Ranch LocalFauna Hemphillian of Texas. Tex Mem Mus, Pearce-Sellards Ser 38:1–41
Dalquest WW, Roth E (1970) Late Pleistocene mammals froma cave in Tamaulipas, Mexico. Southwest Nat 15:217–230
de Bonis L, Crochet J-Y, Rage J-C, Sigé B, Sudre J, Vianey-Liaud M (1973) Nouvelles faunes de vertébrés oligocènes
J Mammal Evol (2009) 16:151–173 169

des phosphorites du Quercy Bull Mus Natn Hist Nat,Paris, 3e série 174:105–113
Ducrocq S, Jaeger J-J, Sigé B (1993) Un mégachiroptère dansl’eocène supérieur de Thaïlande—incidence dans ladiscussion phylogénique du groupe. N Jb Geol PaläontMh 9:561–575
Eick GN, Jacobs DS, Matthee CA (2005) A nuclear DNAphylogenetic perspective on the evolution of echolocationand historical biogeography of extant bats (Chiroptera).Mol Biol Evol 22:1869–1886
Engesser B (1972) Die obermiozäne Säugetier Fauna vonAnwil (Baselland). Tätigkeitsberichte NaturforschGesellsch, Baselland 28:37–363
Engesser B, Ziegler R (1996) Didelphids, insectivores, andchiropterans from the later Miocene of France, CentalEurope, Greece, and Turkey. In: Bernor RL, Fahlbusch V,Mittman H-W (eds) The Evolution of Western EurasianNeogene Mammal Faunas. Columbia University Press,New York, pp 157–167
Foote M, Raup DM (1996) Fossil preservation and thestratigraphic ranges of taxa. Paleobiology 22:121–140
Fox DL, Fisher DC, Leighton LR (1999) Reconstructingphylogeny with and without temporal data. Science284:1816–1819
Galbreath EC (1962) A new myotid bat from the middleOligocene of northeastern Colorado. Trans Kansas AcadSci 65:448–451
Gauthier JA, Kluge AG, Rowe T (1988) Amniote phylogenyand the importance of fossils. Cladistics 4:105–209
Ginsburg L (1963) Les mammifères fossiles récoltés à Sansanau cours du XIXe siècle. Bull Soc Geol France, 7e série5:3–15
Godawa J (1993) Pliocene bats of the genus Myotis (Mamma-lia: Chiroptera) from Podlesice (Poland) and Osztramos 9and 13 (Hungary). Acta Zool Cracov 36:241–250
Gradstein FM, Ogg JG, Smith AG (eds) (2004) A GeologicTimescale 2004. Cambridge University Press, Cambridge
Grady FV, Olson SL (2006) Fossil bats from Quaternarydeposits on Bermuda (Chiroptera: Vespertilionidae). JMammal 87:148–152
Gunnell GF, Jacobs BF, Herendeen PS, Head JJ, Kowalski E,Msuya C, Mizambwa FA, Harrison T, Habersetzer J,Storch G (2003) Oldest placental mammal from sub-Saharan Africa: Eocene microbat from Tanzania—evi-dence for early evolution of sophisticated echolocation.Palaeontol Electron 5(3):10
Gunnell GF, Simmons NB (2005) Fossil evidence and theorigin of bats. J Mammal Evol 12:209–246
Gunnell GF, Simons EL, Seiffert ER (2008) New bats(Mammalia: Chiroptera) from the late Eocene and earlyOligocene, Fayum Depression, Egypt. J Vertebr Paleontol28:1–11
Habersetzer J, Storch G (1987) Klassifikation und funktionelleFlügelmorphologie paläogener Fledermäuse (Mammalia,Chiroptera). Cour Forschung Senckenberg 91:11–150
Hand SJ (1985) New Miocene megadermatids (Chiroptera:Megadermatidae) from Australia with comments onmegadermatid phylogenetics. Aust Mammal 8:5–43
Hand SJ (1990) First Tertiary molossid (Microchiroptera:Molossidae) from Australia: its phylogenetic and biogeo-graphic implications. Mem Queensl Mus 28:175–192
Hand SJ (1993) First skull of a species of Hipposideros(Brachipposideros) (Microchiroptera: Hipposideridae),from Australian Miocene sediments. Mem Queensl Mus33:179–192
Hand SJ (1996) New Miocene and Pliocene megadermatids(Mammalia, Microchiroptera) from Australia, with com-ments on broader aspects of megadermatid evolution.Géobios 29:365–377
Hand S (1997a) Hipposideros bernardsigei, a new hipposiderid(Mammalia: Microchiroptera) from the Australian Mio-cene and a reconsideration of the monophyly of relatedspecies groups. Münchner Geowiss Abh 34:73–92
Hand SJ (1997b) Miophyllorhina riversleighensis gen. et sp.nov., a Miocene leaf-nosed bat (Microchiroptera: Hippo-sideridae) from Riversleigh, Queensland. Mem QueenslMus 41:351–354
Hand SJ (1997c) New Miocene leaf-nosed bats (Microchir-optera: Hipposideridae) from Riversleigh, northwesternQueensland. Mem Queensl Mus 41:335–349
Hand SJ (1998a) Riversleigha williamsi gen. et sp. nov., a largeMiocene hipposiderid (Microchiroptera) from Riversleigh,Queensland. Alcheringa 22:259–276
Hand SJ (1998b) Xenorhinos, a new genus of Old World leaf-nosed bats (Microchiroptera: Hipposideridae) from theAustralian Miocene. J Vertebr Paleontol 18:430–439
Hand SJ, Archer M (2005) A new hipposiderid genus (Micro-chiroptera) from an early Miocene bat community inAustralia. Palaeontology 48:371–383
Hand SJ, Kirsch JAW (2003) Archerops, a new annectenthipposiderid genus (Mammalia: Microchiroptera) from theAustralian Miocene. J Paleontol 77:1139–1151
Hand SJ, Murray P, Megirian D, Archer M, Godthelp H (1998)Mystacinid bats (Microchiroptera) from the AustralianTertiary. J Paleontol 72:538–545
Hand S, Novacek M, Godthelp H, Archer M (1994) FirstEocene bat from Australia. J Vertebr Paleontol 14:375–381
Handley CO (1959) A Revision of Bats of the Genera Eudermaand Plecotus. Smithsonian Institution Press, Washington
Heller F (1935) Fledermäuse aus der Eozänen Braunkohle desGeisel tales bei Halle a. S. Nov Acta Leopold Neue Folge2:301–314
Heller F (1936) Eine oberplicäne Wirbeltierfauna aus Rhein-hessen. N Jb Mineral, Geol, Paläont B 76:99–160
Hendey QB (1981) Paleoecology of the late Tertiary fossiloccurrences in “E” Quarry, Langebaanweg, South Africa,and a reinterpretation of their geological context. Ann SAfr Mus 84:1–104
Hermsen EJ, Hendricks JR (2008) W(h)ither fossils? Studyingmorphological character evolution in the age of molecularsequences. Ann Mo Bot Gard 95:72–100
Hibbard CW (1950) Mammals from the Rexroad Formationfrom Fox Canyon, Kansas. Contr Mus Paleont, Univ Mich8:113–192
Hoofer SR, van den Bussche RA, Horáček I (2006) Genericstatus of the American pipistrelles (Vespertilionidae) withdescription of a new genus. J Mammal 87:981–992
Hooker JJ (1996) A primitive emballonurid bat (Chiroptera,Mammalia) from the earliest Eocene of England. Palae-overtebrata 25:287–300
Horáček I (1986) Kerivoula (Mammalia, Chiroptera), fossil inEurope? Acta Univ Carolinae-Geol, Špinar 2:213–222
170 J Mammal Evol (2009) 16:151–173

Horáček I (2001) On the early history of vespertilionid bats inEurope: the lower Miocene record from the BohemianMassif. Lynx 32:123–154
Horáček I, Fejfar O, Hulva P (2006) A new genus ofvespertilionid bat from early Miocene of Jebel Zelten,Libya, with comments on Scotophilus and early history ofvespertilionid bats (Chiroptera). Lynx 37:131–150
Hulva P, Horáček I, Benda P (2007) Molecules, morphometricsand new fossils provide an integrated view of theevolutionary history of Rhinopomatidae (Mammalia:Chiroptera). BMC Evol Biol 7:165, 15 pp
Jepsen GL (1966) Early Eocene bat from Wyoming. Science154:1333–1339
Jin C-Z, Dong W, Liu J-Y, Wei G-B, Xu Q-Q, Zheng J-J,Zheng L-T, Han L-G, Wang F-Z (2000) A preliminarystudy on the early Pleistocene deposits and the mammalianfauna from the Renzi Cave, Fanchang, Anhui, China. ActaAnthropol Sinica 19(supp):235–245
Jones KE, Bininda-Emonds ORP, Gittleman JL (2005) Bats,clocks, and rocks: diversification patterns in Chiroptera.Evolution 59:2243–2255
Jones KE, Purvis A, MacLarnon A, Bininda-Emonds ORP,Simmons NB (2002) A phylogenetic supertree of the bats(Mammalia: Chiroptera). Biol Rev 77:223–259
Kormos T (1930) Diagnosen neuer Säugetiere aus der ober-pliozänen Fauna des Somlyóberges bei Püspökkfürdő.Ann Mus National Hungarici 27:237–246
Kowalski K (1956) Insectivores, bats and rodents from theearly Pleistocene bone breccia of Podlesice near Kroczyce(Poland). Acta Palaeontol Pol 1:331–393
Kowalski K (1962a) Bats of the early Pleistocene fromKoneprusy (Czechoslovakia). Acta Zool Cracov 7:145–156
Kowalski K (1962b) Fauna of bats from the Pliocene of Wężein Poland. Acta Zool Cracov 7:39–51
Kowalski K (1995) Taphonomy of bats (Chiroptera). Géobios18:251–256
Lavocat R (1961) Le gisement de vertébrés Miocènes de BeniMellal (Maroc): étude systematique de la faune demammiféres. Notes Mém Serv Mines Carte Géol Maroc155:29–144
Lawrence B (1943) Miocene bat remains from Florida, withnotes on the generic characters of the humerus of bats. JMammal 24:356–369
Legendre S (1980) Un chiroptère emballonuridé dans lenéogène d’Europe occidentale; considerations paléobio-géographiques. Géobios 13:839–847
Legendre S (1982) Hipposideridae (Mammalia: Chiroptera)from the Mediterranean middle and late Neogene, andevolution of the genera Hipposideros and Asellia. JVertebr Paleontol 2:372–385
Legendre S (1985) Molossidés (Mammalia, Chiroptera) cén-ozoïques de l’Ancien et du Nouveau Monde; statutsystématique; integration phylogénique des données. NJb Geol Paläont Abh 170:205–227
Lemon RRH, Churcher CS (1961) Pleistocene geology andpaleontology of the Talara Region, northwest Peru. Am JSci 259:410–429
Lindsay EH, Jacobs LL (1985) Pliocene small mammals fromChihuahua. Paleontol Mex 51:1–45
MacFadden BJ (1985) Patterns of phylogeny and rates ofevolution in fossil horses: hipparions from the Miocene
and Pliocene of North America. Paleobiology 11:245–257
Maitre E, Sigé B, Escarguel G (2008) A new family of bats inthe Paleogene of Europe: systematics and implications forthe origin of emballonurids and rhinolophoids. N Jb GeolPaläont Abh 250:199–216
Marandat B, Crochet J-Y, Godinot M, Hartenberger J-L,Legendre S, Rémy JA, Sigé B, Sudre J, Vianey-Liaud M(1993) Une nouvelle faune à mammifères d’âge éocène(Lutétien supérieur) dans les phosphorites du Quercy.Géobios 26:617–623
Meschinelli L (1903) Un nuovo chirottero fossile (Archae-opteropus transiens Mesch.) delle Ligniti di Monteviale.Atti Reale Ist Veneto Sci, Lett, Arti 62:1329–1344
Miller-Butterworth CM, Murphy WJ, O’Brien SJ, Jacobs DS,Springer MS, Teeling EC (2007) A family matter: conclu-sive resolution of the taxonomic position of the long-fingered bats, Miniopterus. Mol Biol Evol 24:1553–1561
Morgan GS (1989) Fossil Chiroptera and Rodentia from theBahamas, and the historical biogeography of the Baha-mian mammal fauna. In: Woods CA (ed) Biogeography ofthe West Indies. Sandhill Crane, Gainesville, pp 685–740
Morgan GS (1991) Neotropical Chiroptera from the Plioceneand Pleistocene of Florida. Bull Am Mus Nat Hist206:176–213
Morgan GS, Czaplewski NJ (1999) First fossil record ofAmorphochilus schnablii (Chiroptera: Furipteridae), fromthe late Quaternary of Peru. Acta Chiropterolog 1:75–79
Morgan GS, Czaplewski NJ (2003) A new bat (Chiroptera:Natalidae) from the early Miocene of Florida, withcomments on natalid phylogeny. J Mammal 84:729–752
Morgan GS, Linares OJ, Ray CE (1988) New species of fossilvampire bats (Mammalia: Chiroptera: Desmodontidae)from Florida and Venezuela. Proc Biol Soc Wash101:912–928
Morgan GS, Ridgeway RB (1987) Late Pliocene (late Blancan)vertebrates from the St. Petersburg Times site, PinellasCounty, Florida, with a brief review of Florida Blancanfaunas. Papers Florida Paleontol 1:1–22
Myers T, Crosby K, Archer M, Tyler M (2001) The EncoreLocal Fauna, a late Miocene assemblage from Riversleigh,northwestern Queensland. Mem Assoc Austr Palaeontol25:147–154
Norell MA, Novacek MJ (1992) The fossil record andevolution: comparing cladistic and paleontologic evidencefor vertebrate history. Science 255:1690–1693
Ostrander GE (1983) New early Oligocene (Chadronian)mammals from the Raben Ranch Local Fauna, northwestNebraska. J Paleontol 57:128–139
Paul CRC (1998) Adequacy, completeness and the fossil record.In: Donovan SK, Paul CRC (eds) The Adequacy of theFossil Record. John Wiley & Sons, Chichester, pp 1–22
Pocock TN (1987) Plio-Pleistocene fossil mammalian micro-fauna of southern Africa—a preliminary report includingdescription of two new fossil muroid genera (Mammalia:Rodentia). Paleontol Afr 26:69–91
Popov VV (2004) Pliocene small mammals (Mammalia, Lip-otyphla, Chiroptera, Lagomorpha, Rodentia) from Muse-lievo (north Bulgaria). Geodiversitas 26:403–491
Qiu Z, Storch G (2000) The early Pliocene micromammalianfauna of Bilike, Inner Mongolia, China (Mammalia: Lip-
J Mammal Evol (2009) 16:151–173 171

otyphla, Chiroptera, Rodentia, Lagomorpha). SenckenbergLeth 80:173–229
Qiu Z, Han D, Qi G, Yufen L (1985) A preliminary report on amicromammalian assemblage from the hominoid localityof Lufeng Co. Yunnan Province. Acta Anthropol Sinica4:13–32
Quinet G (1965) Myotis misonnei n. sp. chiroptere de l’Oligo-cene de Hoogbutsel. Bull Inst R Sci Nat Belg 41:1–11
Rana RS, Singh H, Sahni A, Rose KD, Saraswati PK (2005)Early Eocene chiropterans from a new mammalianassemblage (Vastan Lignite Mine, Gujarat, Western Pen-insular Margin): oldest known bats from Asia. J PalaeontolSoc India 50:93–100
Ray CE (1967) Pleistocene mammals from Ladds, BartowCounty, Georgia. Bull Georgia Acad Sci 25:120–150
Revilliod P (1917) Fledermäuse aus der Braunkohle vonMessel bei Darmstadt. Abh Grossherz-hess Geol Land-esanst 7:161–201
Revilliod P (1919) L’état actuel de nos connaissances sur leschiroptères fossiles (note préliminaire). C R Soc Sci PhysNat Genève 36:93–96
Revilliod P (1922) Contribution a l’étude des chiropters desterrains tertiares. Troisième partie et fin. Mém Soc PaléontSuisse 45:133–195
Rossina VV, Kruskop SV, Tesakov AS, Titov VV (2006) Thefirst record of Late Miocene Bat from European Russia.Acta Zool Cracov 49A:125–133
Russell DE, Louis P, Savage DE (1973) Chiroptera andDermoptera of the French early Eocene. Univ Cal PubGeol Sci 95:1–57
Samonds KE (2007) Late Pleistocene bat fossils from AnjohibeCave, northwestern Madagascar. Acta Chiropterolog 9:39–65
Savage DE (1951) A Miocene phyllostomatid bat fromColombia, South America. Univ Cal Bull Dep Geol Sci28:357–366
Savage DE, Russell DE (1983) Mammalian Paleofaunas of theWorld. Addison-Wesley, Reading, Massachusetts
Sevilla P (1989) Quaternary fauna of bats in Spain: paleoeco-logic and biogeographic interest. In: Hanák V, Horáček I,Gaisler J (eds) European Bat Research 1987. CharlesUniversity Press, Prague, pp 349–355
Sevilla P (1990) Rhinolophoidea (Chiroptera, Mammalia) fromthe upper Oligocene of Carrascosa del Campo (CentralSpain). Géobios 23:173–188
Sevilla P (1991) Murcielagos fosiles de España. In: Benzal J, dePaz O (eds) Los Murciélagos de España y Portugal. Icona,Madrid, pp 21–36
Sigé B (1968) Les chiroptères du miocène inférieur deBouzigues. I.-Étude Systématique. Palaeovertebrata 1:65–133
Sigé B (1974) Données nouvelles sur le genre Stehlinia(Vespertilionoidea, Chiroptera) du Paléogène d’Europe.Palaeovertebrata 6:253–272
Sigé B (1976) Les Megadermatidae (Chiroptera, Mammalia)Miocènes de Béni Mellal, Maroc. Géol Médit 3:71–86
Sigé B (1985) Les chiroptères oligocènes du Fayum, Egypte.Geol Palaeontol 19:161–189
Sigé B (1990) Nouveaux chiroptères de l’oligocène moyen desphosphorites du Quercy, France. C R Acad Sci Paris310:1131–1137
Sigé B (1991) Rhinolophoidea et Vespertilionoidea (Chirop-tera) du Chambi (eocène inférieur de Tunisie). Aspectsbiostratigraphique, biogéographique et paléoécologique del’origine des chiroptères modernes. N Jb Geol Paläont Abh182:355–376
Sigé B, Hand S, Archer M (1982) An Australian MioceneBrachipposideros (Mammalia, Chiroptera) related to Mio-cene representatives from France. Palaeovertebrata12:149–172
Sigé B, Legendre S (1983) L’historie des peuplements dechiroptères du bassin Méditerranéen: l’apport comparé desremplissages karstiques et des dépôts fluvio-lacustres.Mém Biospéol 10:209–225
Sigé B, Russell DE (1980) Compléments sur les chiroptères del’Eocène moyen d’Europe. Les genres Palaeochiropteryxand Cecilionycteris. Palaeovertebrata, Mém Jubil R Lavo-cat, pp 91–126
Sigé B, Thomas H, Sen S, Gheerbrant E, Roger J, Al-SulaimaniZ (1994) Les chiropters de Taqah (oligocène inférieur,Sultanat d’Oman). Premier inventaire systématique.Münchner Geowiss Abh 26:35–48
Silva-Taboada G (1974) Fossil Chiroptera from cave deposits incentral Cuba, with descriptions of two new species (generaPteronotus and Mormoops) and the first West Indianrecord of Mormoops megalophylla. Acta Zool Cracov19:33–73
Simmons NB (2005) Order Chiroptera. In: Wilson DE, ReederDM (eds) Mammal Species of the World: a Taxonomicand Geographic Reference, 3rd edn. The Johns HopkinsUniversity Press, Baltimore, pp 312–529
Simmons NB, Geisler JH (1998) Phylogenetic relationships ofIcaronycteris, Archaeonycteris, Hassianycteris, and Palaeo-chiropteryx to extant bat lineages, with comments on theevolution of echolocation and foraging strategies inMicrochiroptera. Bull Am Mus Nat Hist 235:1–182
Simmons NB, Seymour KL, Habersetzer J, Gunnell GF (2008)Primitive early Eocene bat from Wyoming and theevolution of flight and echolocation. Nature 451:818–822
Smith JD, Storch G (1981) New middle Eocene bats from“Grube Messel” near Darmstadt, W-Germany (Mammalia:Chiroptera). Senckenberg Biol 61:153–167
Smith R, Russell DE (1992) Mammifères (Marsupialia,Chiroptera) de l’Yprésien de la Belgique. Bull Inst RoySci Nat Belg, Sci Terre 62:223–227
Smith T, Rana RS, Missiaen P, Rose KD, Sahni A, Singh H,Singh L (2007) High bat (Chiroptera) diversity in the earlyEocene of India. Naturwissenschaften 94:1003–1009
Springer MS, Teeling EC, Madsen O, Stanhope MJ, de JongWW (2001) Integrated fossil and molecular data recon-struct bat echolocation. Proc Natl Acad Sci USA 98:6241–6246
Stirton RA (1931) A new genus of the family Vespertilionidaefrom the San Pedro Pliocene of Arizona. Univ California,Bull Dep Geol Sci 20:27–30
Storch G (1999) Order Chiroptera. In: Rossner GE, Heissig K(eds) The Miocene Land Mammals of Europe. Verlag DrFriedrich Pfeil, Munich, Germany, pp 81–90
Storch G, Sigé B, Habersetzer J (2002) Tachypteron franzeni n.gen., n. sp., earliest emaballonurid bat from the middleEocene of Messel (Mammalia, Chiroptera). Paläontol Zeit76:189–199
172 J Mammal Evol (2009) 16:151–173

Storer JE (1984) Mammals of the Swift Current Creek LocalFauna (Eocene; Uintan), Saskatchewan. Nat Hist Contrib,Mus Nat Hist, Regina 7:1–158
Strand E (1928) Miscellanea nomenclatorica zoologica etpalaeontologica. Arch Naturgesch 92:30–75
Suárez W, Díaz-Franco S (2003) A new fossil bat (Chiroptera:Phyllostomidae) from a Quaternary cave deposit in Cuba.Caribb J Sci 39:371–377
Sutton JF, Genoways HH (1974) A new vespertilionine batfrom the Barstovian deposits of Montana. Occ Papers,Mus Tx Tech Univ 20:1–8
Teeling EC, Springer MS, Madsen O, Bates P, O’Brien SJ,Murphy WJ (2005) A molecular phylogeny for batsilluminates biogeography and the fossil record. Science307:580–584
Tejedor MF, Czaplewski NJ, Goin FJ, Aragón E (2005) Theoldest record of South American bats. J Vertebr Paleontol25:990–993
Thewissen JGM, Smith GR (1987) Vespertilionid bats (Chi-roptera, Mammalia) from the Pliocene of Idaho. ContribMus Paleont, Univ Mich 27:237–245
Tong Y-S (1997) Middle Eocene small mammals fromLiguanqiao Basin of Henan Province and Yuanqu Basinof Shanxi Province, central China. Paleontol Sinica C26:1–256
Topál G (1963) Description of a new bat, Rhinolophusmacrorhinus sp. n. from the lower Pleistocene of Hungary.Vertebr Hung 5:219–227
Topál G (1974) The first record of Megaderma in Hungary(Pliocene sediments of Osztramos, Locality 10). VertebrHung 15:95–104
Topál G (1983) New and rare fossil mouse-eared bats from themiddle Pliocene of Hungary (Mammalia, Chiroptera).Fragm Mineral Palaeontol 11:43–54
Topál G (1985) Bats from the lowermost Pleistocene locality 15at Beremend, Hungary (Mammalia, Chiroptera). FragmMineral Palaeontol 12:51–57
Topál G (1989a) New Tertiary plecotines from Hungary(Mammalia, Chiroptera). In: Hanák V, Horáček I, GaislerJ (eds) European Bat Research 1987. Charles UniversityPress, Prague, pp 77–86
Topál G (1989b) Tertiary and early Quaternary remains ofCorynorhinus and Plecotus from Hungary (Mammalia,Chiroptera). Vertebr Hung 23:33–55
Valentine JW (1989) How good was the fossil record? Cluesfrom the California Pleistocene. Paleobiology 15:83–94
Walker A (1969) True affinities of Propotto leakeyi Simpson1967. Nature 223:647–648
Wesselmann HB (1984) The Omo Micromammals. Systematicsand Paleoecology of Early Man Sites from Ethiopia.Karger, New York
Wessels W, Fejfar O, Peláez-Campomanes P, van der MeulenA, de Bruijn H (2003) Miocene small mammals fromJebel Zelten, Libya. Coloq Paleont 1(suppl):699–715
White JA (1969) Late Cenozoic bats (subfamily Nyctophyli-nae) from the Anza-Borrego Desert of California. MiscPubl, Univ Kansas Mus Nat Hist 51:275–282
Wilson RL (1968) Systematics and faunal analysis of a lowerPliocene vertebrate assemblage from Trego County,Kansas. Contrib Mus Paleontol, Univ Mich 22:75–126
Wołoszyn BW (1986) A new species of long-winged batMiniopterus tao sp. n. (Mammalia: Chiroptera) fromLocality 1 at Choukoutien, China. Acta Universit Carolin-Geol, Spinar 2:205–211
Wood AR, Zelditch ML, Rountrey AN, Eiting TP, Sheets HD,Gingerich PD (2007) Multivariate stasis in the dentalmorphology of the Paleocene-Eocene condylarth Ectocion.Paleobiology 33:248–260
Worthy TH, Holdaway RN (1994) Quaternary fossil faunasfrom caves in Takaka valley and on Takaka Hill, northwestNelson, South Island, New Zealand. J Roy Soc NewZealand 24:297–391
Yang J (1977) On some Salientia and Chiroptera fromShanwang, Linqu, Shandong Vertebr Palasiatica 15:76–80
Yoon MH, Kuramoto T, Uchida TA (1984) Studies of middlePleistocene bats including Pleistomyotis gen. et sp. nov.and two new extinct Myotis species from the Akiyoshi-daiPlateau. Bull Akiyoshi-dai Mus Nat Hist 19:15–26
Zapfe H (1950) Die fauna der Mioznäen Spaltenflülung vonNeudorf an der March (ČSSR). Chiroptera. Sitzungber-ichte Akad Wissenschaft Wien, Abt 1 159:51–64
Zapfe H (1970) Paleptesicus nom. nov. fur. “Paraptesicus”(Chiroptera) aus der Mioznäen Spaltenflülung von Neudorfan der March (ČSSR). Sitzungsberichte Math-Naturwissen-schaft Klasse, Österreich Akad Wissenschaft 6:93–94
Ziegler R (1994) Die Chiroptera (Mammalia) aus demUntermiozän von Stubersheim 3 (Baden-Württemberg).Münchner Geowiss Abh 26:97–116
Ziegler R (2003) Bats (Chiroptera, Mammalia) from middleMiocene karstic fissure fillings of Petersbuch near Eich-stätt, Southern Franconian Alb (Bavaria). Géobios36:447–490
J Mammal Evol (2009) 16:151–173 173