germline passage of mitochondria: quantitative
TRANSCRIPT

Human Reproduction, Vol. 15, (Suppl. 2), pp. 112-128, 2000
Germline passage of mitochondria: quantitativeconsiderations and possible embryological sequelae
Robert P.SJansen1
Sydney IVF and the University of Sydney
'To whom correspondence should be addressed at: Sydney IVF, 4 O'Connell Street,Sydney 2000, Australia. E-mail: [email protected]
Using a semi-quantitative review ofpublished electron micrographs, we haveexplored the passage of mitochondria fromone generation to the next through thecytoplasm of the human female germ cell.We propose a testable hypothesis that themitochondria of the germline are persist-ently 'haploid' (effectively carrying just onemitochondrial chromosome per organelle).For mitochondria, the passage throughgerm cell differentiation, oogenesis, follicleformation and loss could constitute arestriction/amplification/constraint event ofa type previously demonstrated for asexualpurification and refinement of a non-recombining genome. At the restrictionevent (or 'bottleneck') in the human prim-ordial germ cell, which differentiates inembryos after gastrulation, there appear tobe <10 mitochondria per cell. From -100or so such cells, a population of >7X1O6
oogonia and primary oocytes is producedin the fetal ovaries during mid-gestation,with mitochondria numbering up to 10 000per cell, implying a massive amplificationof the mitochondrial genome. A further10-fold or greater increase in mitochondrialnumbers per oocyte occurs during adultfollicular growth and development, asresting primordial follicles develop to pre-
112
ovulatory maturity. So few are the numbersof oocytes that fertilize and successfullycleave to form an embryo of the new genera-tion, that biologists have long suspectedthat a competitive constraint lies behindthe generational completion of this geneticcycle. I propose that maintaining theintegrity of mitochondrial inheritance issuch a strong evolutionary imperative thatfeatures of ovarian follicular formation,function, and loss could be expected tohave been primarily adapted to this specialpurpose. To extend the hypothesis further,the imperative of maintaining mitochon-drial genomic integrity in a populationcould explain why women normally becomesterile a number of years before there isdepletion of ovarian follicles and endocrineovarian failure (i.e. why there is 'anoopause' preceding the menopause).Plausible explanations might also followfor several well-known and puzzling repro-ductive difficulties, including recurrent mis-carriage, unexplained infertility, andpersistent failure of IVF embryos to cleaveor to implant. Current experimentallaboratory manoeuvres that might circum-vent mitochondrial shortcomings (such ascytoplasmic transfusion and karyoplastexchange) are examined and possibleclinical hazards identified.
© European Society of Human Reproduction & Embryology
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
Key words: inheritance/mitochondria/mito-chondrial DNA/oocyte/primordial germ cell
Introduction
A number of important questions concerningovarian function and dysfunction remain inneed of satisfactory explanation: (i) why is itthat, in probably all mammalian species, thecomplement of female gametes is producedearly in the animal's life, before the attainmentof sexual maturity?; (ii) why is there such ahigh rate of attrition of ova in mammals,specifically in humans?; (iii) could there be apurpose for the mitotic frenzy of ovarianfollicular cells during the process of folliclegrowth beyond the secretion of oestradiol andthe release of an oocyte surrounded by mucusthat is sticky for the fimbrial end of theFallopian tube?; (iv) what causes the deteriora-tion of oocyte competence observed in womenin their late 30s, a decade or so before finalfollicular depletion at the menopause?; and(v) might mitochondrial dysfunction be theultimate cause of some disorders of ovulation,unexplained infertility, and recurrent mis-carriage?
Whether a mitochondrial genomic perspec-tive could provide answers to these questionsconstitutes the subject of this paper. The mito-chondrial quantifications have been previouslypublished (Jansen and De Boer, 1998).
Mitochondria during oogenesis
Throughout oogenesis and early embryo-genesis, mitochondria in germ cells differ inappearance from somatic cell mitochondria.Morphologically, except during presumedtimes of high mitochondrial multiplicationclose to the nucleus, mitochondria in femalegerm cells assume unique spherical profiles(Smith and Alcivar, 1993). It is known thatmature mammalian oocytes have about the
same number of mitochondria as they havemitochondrial (mt) DNA circles: they areeffectively haploid, containing mostly just onemtDNA molecule per organelle (Piko andMatsumoto, 1976; Michaels et al, 1982; Pikoand Taylor, 1987). An elongated, typicallysomatic cell mitochondrial morphology (Smithand Alcivar, 1993) and normal replicationpatterns (see Larsson et al, 1998, for con-sequences if replication fails) are establishedtogether, after implantation (see also Shepardet al., 2000).
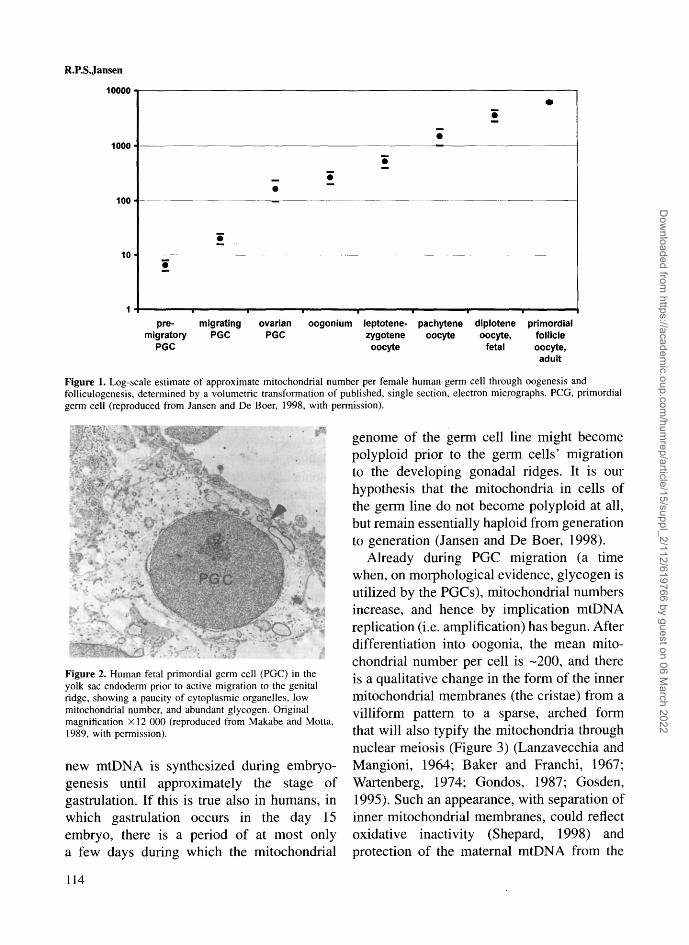
We have used previously published electronmicrographs of the various stages of humanoogenesis to estimate the number of mito-chondria at each stage of human oogenesis(Figure 1) (Jansen and De Boer, 1998). Thefewest mitochondrial profiles per germ cellare seen in the earliest primordial germ cells(PGCs), still in the endoderm of the yolk sacof 3 week embryos, with about four or fivevisible profiles per cell, predicting a mito-chondrial number of <10 (Figure 2). Thisnumber of mitochondria might be the lowestthat occurs in the transmission of cytoplasmfrom one human generation to the next: arestriction event to qualify for the first step inreversing Muller's ratchet (see Jansen, 2000).It cannot be ruled out that an even lowernumber might have occurred in the embryonicgerm cell line at gastrulation, a week earlier.
In their consideration of a numeric restric-tion event in mitochondrial transmission,Michaels et al. (1982) assumed thatembryonic cells prior to primordial germ linedifferentiation would have a general somaticcell (i.e. polyploid) mtDNA copy number perorganelle; they concluded that the reduction(and hence the location of the bottleneck)occurs later in oogenesis. No direct measure-ments of mtDNA copy number have beenmade on PGCs. It should be noted, however,that in species as diverse as the frog (Dawid,1972; El Meziane et al, 1989) and the mouse(Ebert et al, 1988; Piko and Chase, 1993) no
113
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022
R.P.S.Jansen
10000
1000
100
pre-migratory
PGC
migratingPGC
ovarianPGC
oogonium leptotene- pachytene diplotene primordialzygotene oocyte oocyte, follicle
oocyte fetal oocyte,adult
Figure 1. Log-scale estimate of approximate mitochondrial number per female human germ cell through oogenesis andfolliculogenesis, determined by a volumetric transformation of published, single section, electron micrographs. PCG, primordialgerm cell (reproduced from Jansen and De Boer, 1998, with permission).
genome of the germ cell line might becomepolyploid prior to the germ cells' migrationto the developing gonadal ridges. It is ourhypothesis that the mitochondria in cells ofthe germ line do not become polyploid at all,but remain essentially haploid from generationto generation (Jansen and De Boer, 1998).
Already during PGC migration (a timewhen, on morphological evidence, glycogen isutilized by the PGCs), mitochondrial numbersincrease, and hence by implication mtDNAreplication (i.e. amplification) has begun. Afterdifferentiation into oogonia, the mean mito-chondrial number per cell is -200, and thereis a qualitative change in the form of the innermitochondrial membranes (the cristae) from avilliform pattern to a sparse, arched formthat will also typify the mitochondria throughnuclear meiosis (Figure 3) (Lanzavecchia andMangioni, 1964; Baker and Franchi, 1967;Wartenberg, 1974; Gondos, 1987; Gosden,1995). Such an appearance, with separation ofinner mitochondrial membranes, could reflectoxidative inactivity (Shepard, 1998) andprotection of the maternal mtDNA from the
Figure 2. Human fetal primordial germ cell (PGC) in theyolk sac endoderm prior to active migration to the genitalridge, showing a paucity of cytoplasmic organelles, lowmitochondrial number, and abundant glycogen. Originalmagnification X12 000 (reproduced from Makabe and Motta,1989, with permission).
new mtDNA is synthesized during embryo-genesis until approximately the stage ofgastrulation. If this is true also in humans, inwhich gastrulation occurs in the day 15embryo, there is a period of at most onlya few days during which the mitochondrial
114
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
Figure 3. Human fetal pachytene primary oocyte, with an increase in mitochondrial number taking place close to the nucleus,which demonstrates synaptonemal complexes (arrow) indicative of nuclear chromosomal crossover. Original magnificationX5000 (reproduced from Gondos et al., 1986, with permission).
mutagenic stress of reactive oxygen species(Allen, 1996). The replication of germ cellsthat takes place over the next few months offetal development sees an increase in germcell numbers (Baker, 1963) to -7X106 (anincrease of 218 over the yolk sac PGC number),and an implied increase in total germ cellmitochondria to -35 billion (an increase of229 over the assumed yolk sac germ linemitochondrial total). During this explosiveamplification in numbers, mitochondria mostlyretain their spherical profile.
In pachytene of the first meiotic division,when synaptonemal complexes form in thenuclei of the developing oocytes to allowgenetic recombination to take place in thediploid nuclear genome, human mitochondriareplicate while located close to the nucleus(Figure 3) (Lanzavecchia and Mangioni, 1964;Baker and Franchi, 1967; Gondos etal, 1986),giving cross-sections of human pachyteneoocytes the appearance of a necklace of mito-chondria, a 'mitochondrial crown', as it hasbeen called in amphibian oocytes at this stageof germ cell development (Mignotte et al,
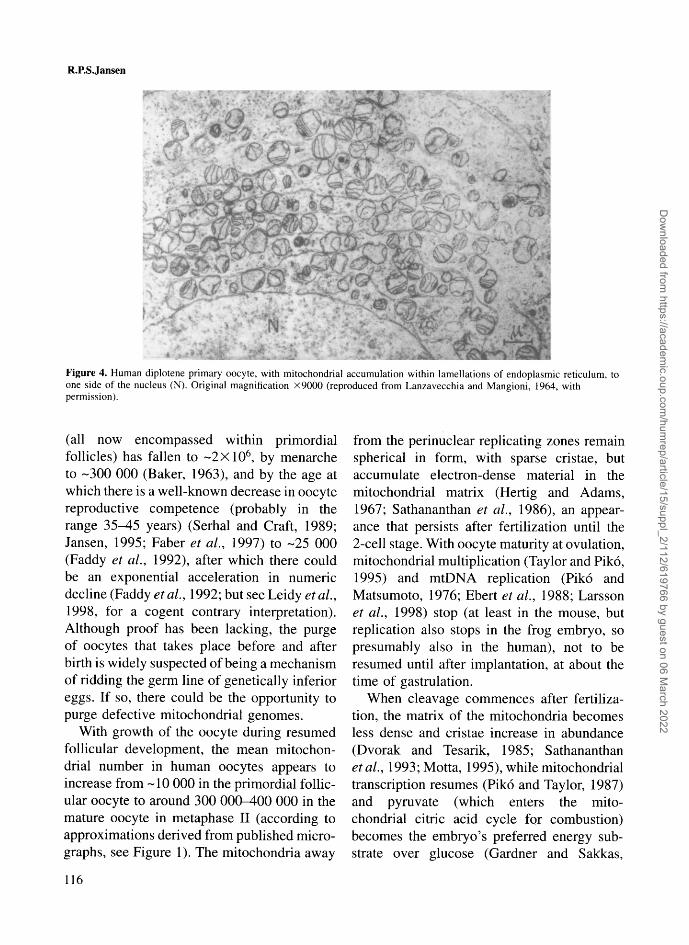
1987). Recent data imply that mitochondrialDNA replication in somatic cells is preferen-tially located close to the nucleus (Davis andClayton, 1996). Later, in diplotene, among thesurviving oocytes that have formed primordialfollicles, the mitochondria are found withinlamellations of endoplasmic reticulum parallelwith and to one side of the nucleus (Figure 4)(Lanzavecchia and Mangioni, 1964). Studiesof Xenopus (frog) pachytene and diploteneoocytes show a similar mitochondrial arrange-ment (Callen et al, 1980; Tourte et al, 1984):incorporation of radioactive thymidine revealsthat mitochondria segregate into two popula-tions at this stage of oogenesis in frogs, withone population close to the nucleus, activelyreplicating mtDNA and building up most ofthe stock of the mitochondria for the full-grown oocyte, while the second population,in which DNA replication has stopped,occupies a 'mitochondrial mass' in the vegetalpole, which constitutes the germ plasm (cyto-plasm destined for the germ cells) in thisspecies.
In humans at birth, the number of oocytes
115
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022
R.P.S.Jansen
Figure 4. Human diplotene primary oocyte, with mitochondrial accumulation within lamellations of endoplasmic reticulum, toone side of the nucleus (N). Original magnification X9000 (reproduced from Lanzavecchia and Mangioni, 1964, withpermission).
(all now encompassed within primordialfollicles) has fallen to -2X106, by menarcheto -300 000 (Baker, 1963), and by the age atwhich there is a well-known decrease in oocytereproductive competence (probably in therange 35^5 years) (Serhal and Craft, 1989;Jansen, 1995; Faber et al, 1997) to -25 000(Faddy et al, 1992), after which there couldbe an exponential acceleration in numericdecline (Faddy etal., 1992; but see Leidy etal.,1998, for a cogent contrary interpretation).Although proof has been lacking, the purgeof oocytes that takes place before and afterbirth is widely suspected of being a mechanismof ridding the germ line of genetically inferioreggs. If so, there could be the opportunity topurge defective mitochondrial genomes.
With growth of the oocyte during resumedfollicular development, the mean mitochon-drial number in human oocytes appears toincrease from -10 000 in the primordial follic-ular oocyte to around 300 000^00 000 in themature oocyte in metaphase II (according toapproximations derived from published micro-graphs, see Figure 1). The mitochondria away
from the perinuclear replicating zones remainspherical in form, with sparse cristae, butaccumulate electron-dense material in themitochondrial matrix (Hertig and Adams,1967; Sathananthan et al, 1986), an appear-ance that persists after fertilization until the2-cell stage. With oocyte maturity at ovulation,mitochondrial multiplication (Taylor and Piko,1995) and mtDNA replication (Piko andMatsumoto, 1976; Ebert et al, 1988; Larssonet al., 1998) stop (at least in the mouse, butreplication also stops in the frog embryo, sopresumably also in the human), not to beresumed until after implantation, at about thetime of gastrulation.
When cleavage commences after fertiliza-tion, the matrix of the mitochondria becomesless dense and cristae increase in abundance(Dvorak and Tesarik, 1985; Sathananthanetal, 1993; Motta, 1995), while mitochondrialtranscription resumes (Piko and Taylor, 1987)and pyruvate (which enters the mito-chondrial citric acid cycle for combustion)becomes the embryo's preferred energy sub-strate over glucose (Gardner and Sakkas,
116
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
1993). This more active appearance of themitochondria is mirrored in the nucleus bythe de-compaction of the nucleolus, indicatingsimultaneous resumption also of nucleartranscription. Thus, it is plausible that thedecreasing density of mitochondrial matrixcontents at this time represents incorporationof previously imported, maternal genome-derived protein components of respiratorychain enzyme complexes, stored in anticipa-tion of mitochondrial transcription and transla-tion of the remaining protein subunits, andincorporation of complete complexes into thenew inner mitochondrial membrane.
Germline mitochondrial DNArecombination?
Just as Chao's viruses recovered genomicfitness faster with a relatively simple form ofrecombination (segment reassortment) addedto their predominant reliance on non-sexualmeans to reverse Muller's ratchet (Chao et al.,1997; Jansen, 2000), so it is plausible thatmitochondria also rely to a degree on bothnon-recombinant and recombinant strategies.The possibility of reciprocal recombinationoccurring between female and male germ cellmitochondria has classically been considereddoubtful (Hauswirth and Laipis, 1985), theevidence against it being that nucleotidesequence differences separated by -3000 bpare consistently coinherited (Olivo et al.,1983). But this evidence contradicts re-combination between interorganismal mtDNAgenomes: it does not argue against intra-organismal recombination. Mitochondria invertebrate oocytes have the enzymaticmachinery to carry out homologous recom-bination, namely excision repair involvingDNA polymerase y (Ryoji et al., 1996) andpossibly mitochondrial DNA ligase (at leastas indicated by its isolation from homogenizedXenopus ovaries) (Pinz and Bogenhagen,1998). Somatic cell mitochondrial protein
extracts are able to catalyse recombination ofplasmid DNA substrates in vitro (Thyagarajanet al., 1996), and it is possible that mitochon-drial deletions in somatic cells occur as aresult of prior illegitimate recombination (seeHowell, 1997, for review).
Embryonic survival and a newgeneration, or failure
The observation that mitochondrial DNA syn-thesis is suspended until gastrulation in speciesas diverse as the frog (El Meziane et al.,1989) and the mouse (Larsson et al, 1998),vertebrate lines that diverged >400 millionyears ago, implies that there could be a univer-sally important biological principle behind thesuspension. Gastrulation is soon followed bysequestration of the first PGCs in the posteriorwall of the yolk sac.
If true also in humans, the morphologicaldata summarized quantitatively in Figure 1places the major site for the mitochondrialpopulation bottleneck at or before the stage ofthe pre-migratory PGC, with a mitochondrialnumber (hence by implication a mtDNA copynumber) of probably <10 per germ-line cell.Depending on how and when mtDNA syn-thesis and replication restarts in the germcell line (presumably from a juxtanuclearmitochondrial subpopulation), it is plausiblethat there might be a further restriction in thenumber of founding mitochondrial chromo-somes, with a minimum in some circumstances(perhaps when the genome is under specialchallenge from one or more mutations) (Bloket al., 1997) of a single founding mitochondrialgenome.
The gap between resumption of transcrip-tion (after the 2-cell stage in the mouse) (Pikoand Taylor, 1987; Taylor and Piko, 1995) andnew mtDNA synthesis (at about the stage ofgastrulation) means that the quantitative aswell as the qualitative endowment of mtDNAspecies to the conceptus will come under strain
117
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

R.P.SJansen
as the extant mitochondria are diluted andpartitioned into multiplying daughter blastom-eres. Recurrent miscarriage can be an earlyfeature of Wilson's disease (preceding clinicalliver disease by many years; Schagen vanLeeuwen et al., 1991), in which a faultycopper-transporting protein gene-productlocated in mitochondria results in intramito-chondrial accumulation of copper, causingoxidative damage to mtDNA (Mansouri et al.,1997). Elucidating the nature of the normalpermissive stimulus for resumption of mtDNAsynthesis (and the requirements for successfulresumption) could yield insights into themolecular aetiology of early pregnancy failure.
Reversing Muller's ratchet: preservingor evolving the mitochondrial genome
For entropy and Muller's ratchet to be not juststopped but reversed (i.e. for there to be animprovement with time in a genome re-producing asexually) it is not enough to havemere winners at the end of a restriction/amplification/constraint cycle. The inheritanceof potentially harmful human mtDNA pointmutations (such as those behind thematernally inherited mitochondrial neuro-muscular dystrophies MERFF, MELAS andLeigh's syndrome) from one generationto the next (Holme et al., 1995) shows thatthe restriction/amplification/constraint cycledemonstrated above, with or without intra-organismal mtDNA recombination should itoccur, is not in itself enough to prevent adeleterious drift of mtDNA along with ageneral decline in cytoplasmic health amongthe individual's descendants and hence amongthe general population (and eventually thespecies). The surviving genome might stillshow a slow degradation over time. There stillneeds to be a way of also preventing lesspathological, but nevertheless deleterious,mutations from becoming fixed.
For free-ranging asexually reproducing
organisms, a fluctuating environment, includ-ing competition with other organisms, will beenough to make sure that degrading genomeswill become extinct during times of environ-mental deterioration, leaving the fittest tosurvive and evolve. However, for the host ofa non-autonomous endosymbiont such as amitochondrion this will not be enough. Thehost species will be in trouble if it is atthe mercy of its parasitic endosymbiont longbefore the endosymbiont itself is threatened.It is therefore probable that the host specieswill have evolved a strategy for putting a stopto mitochondrial genomic degradation moresatisfactory than high adult or even childhoodmortality. I propose that a physiological stateof sterility well before menopause could notonly be mitochondrially based but could con-stitute a strategy for slowing Muller's ratchet.
It has been argued (Lynch et al., 1993) thatweakened mitochondrial genomes should notjust be accepted by the host, that they need tohave their suboptimal effects magnified to theextent that they will be recognized and readilyeliminated from the population; i.e. that 'theextreme longevity of organelle lineages maybe due not to their invulnerability to mutationsand DNA damage, but to their extremesensitivity to such effects'. Experience withthe biochemical and clinical effect of mtDNAmutations in somatic tissues is now consider-able (Brown and Wallace, 1994), if still con-fused by the polyploidal, compensated statusof the somatic cell mitochondrial genome andthe plastic nature of organelle fusion andfission. Nonetheless once a threshold of mito-chondrial mutations is reached, the phenotypicconsequences are typically inexorable anddisastrous. Lynch et al. (1993) list the under-lying molecular reasons for the unmitigatedsequelae: proofreading and excisional repairgenes have been lost; transfer RNA and ribo-somal RNA genes, present in multiple copiesin the nucleus, are single and vulnerable inmitochondria; mutagenic conditions are poten-
118
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
dally harsh and punishing, with excesssuperoxide radicles able to further disrupt thegenotypic and hence phenotypic integrity ofoxidative phosphorylation, generating morefree radicles and a runaway process of acceler-ated mutation ('mutational meltdown'); anddeleted mtDNA circles have a propensity toreplicate faster than full circles, driving themto fixation and to the death of the host.Mitochondrial abnormalities are also highlyleveraged clinically: disease occurs and leadsto death of the organism well before tissuesare critically deprived of ATP and widespreadcell death occurs (E.A.Schon, Department ofNeurology, Columbia University, New York);and mitochondria are intimately associatedwith pathways of apoptosis that lead to pro-grammed cell death in situ (including, in thecase of the ovaries, follicular atresia andoocyte death) (Tilly, 1996; Perez, et al., 1997).
If these leveraged processes operate amonghaploid mitochondria of a germ line they couldbe just as tough in effect, if different in detail.In the case of the germ line, the ultimate testof the mitochondria for the new generationwould be during the time between the onsetof cleavage and successful implantation andgastrulation, when sequestration of a newgerm line occurs. The mechanism for this finaltest of mitochondrial genomic integrity couldbe as simple as the constitutional enforcementof a delay in new mtDNA synthesis. Thiswould ensure the demise of a conceptus thatdid not possess a sufficient number of normalmitochondrial genomic species. I furtherexamine below what the expected clinicalconsequences of such an evolutionary devicecould be expected to be.
On a population level it is apparent thatthe mitochondrial genome does graduallydeteriorate, with replacement of conservedsequences by more or less silent poly-morphisms among the coding genes moreprevalent within species than between species(Nachman, 1998), including the different
hominoids (Hasegawa et al., 1998). Muller'sratchet 'clicks', despite the opposing safe-guards discussed above. It has been estimated,however, that the resultant reduction of fitnessfrom the accumulation of mutations is notlikely to imperil many species on time scalesof much less than a million years (Lynchand Blanchard, 1998), so this ultra-long termphenomenon can be ignored clinically.
In the shorter term of children or grand-children, an improvement in mitochondrialgenome quality ought to be possible fromone generation to the next: the 'clock' ofmitochondrial degradation that every adultanimal accrues with age is 'reset' with everynew generation.
Folliculogenesis and competitionbetween follicles and oocytes
Of ~7 X106 germ cells that form in the ovaries(see above), at most a few hundred willovulate. In the span of 4 weeks (every ovariancycle during the reproductive years), tens orhundreds of follicles start their growth fromthe resting primordial state. Yet in most cir-cumstances just one follicle each month pre-sents its oocyte to be fertilized. Theobserver could reasonably suspect that he orshe is witnessing a competition: waves offolliculogenesis and waves of atresia, with avery small number of winners (Krakauer andMira, 1999).
Yet this is essentially the extent of theevidence that competition between ovarianfollicles and between oocytes actually takesplace. The evidence is circumstantial. Thereis still no compelling theory to explain eventhe initiation of a particular follicle's growthand development, let alone an indication ofwhat it is that distinguishes follicles that canmake it through to ovulation from those thatcannot. The endocrine tide of fluctuating levelsof follicle stimulating hormone and the once-a-month event of a LH surge is only the test
119
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

R.P.SJansen
for the follicles: it does not explain the basisof differentiation between the candidates.
Ovarian folliculogenesis (and the oppor-tunity for follicular atresia) is phylogeneticallyancient, with junctional complexes betweenfollicle cells and oocytes being found indeveloping follicles not only in all vertebrateclasses, but also in the echinoderms (such asthe sea urchins), which diverged from thechordate line >600 million years ago(Beijnink, et al, 1984). So it could be thatwhatever it is that distinguishes somefollicles over others will also be biochemicallyancient and fundamental. The ability of foll-icles to express the means of generatingenergy, perhaps in the form of ATP, ought to bea strong contender. Follicles win by growingefficiently. They grow through the mitosis oftheir constituent granulosa cells. Meanwhilethe oocyte nucleus rests in prophase ofmeiosis I, as it has since fetal life. Its mitotictest (should it return to diploidy through fertil-ization) comes after the follicle is spent, mani-festing as cleavage, blastulation, gastrulation,and so on. Could the mitotic competenceof the granulosa cells somehow be a directindicator of what the oocyte's genomicintegrity, what its metabolic strength, and whatits mitotic potential will be after syngamy?So inextricably linked is the fate of an oocyteto the fate of its follicle that it would bereasonable to put the hypothesis that it is themetabolic potential of the oocytes (i.e. thegenomic integrity of oocyte mitochondria) thatis being tested by a metabolic competitionbetween follicles.
At this stage it is speculation, but forillustrative purposes there are plausiblemechanisms linking oocyte and follicle cellmetabolic competence, mechanisms thatwould confer further purpose upon theintimate cytoplasmic contact granulosa cellprocesses have with oocytic cytoplasm viagap junctions permeable to small solublemolecules (Buccione et al, 1990; Grazul-
Bilska et al., 1997). Such junctional contact isabsolutely required for oocyte growth (Eppig,1979), and mediates the transfer of substancesinhibitory to meiosis from granulosa cells tothe oocyte (Tsafriri et al, 1982), but alsotransmits from ooplasm to adjacent granulosacells soluble substances that cause differ-entiation of the developing antral follicle (Liand Mather, 1997) and, later, of cumulus cells(Eppig et al, 1997). The oocyte's ATP contentcorrelates significantly with the developmentalpotential of the embryo after fertilizationin vitro (Van Blerkom et al, 1995), but it isuncertain whether the ATP is generated by theoocyte, by the granulosa (follicles with poorlycompetent oocytes are hypoxic) (Van Blerkomet al, 1997), or by both, as gap junctions arefreely permeable to nucleotides, including theintracellular signalling nucleotide cyclic AMP(Lawrence et al, 1978). At this point, it mayat least be accepted that metabolic trafficbetween granulosa cell cytoplasm and oocytecytoplasm will be two-way.
If the mitochondrial mutational load of atleast some ovulated oocytes turns out to be asignificant constraint on the development ofmany superovulated and recovered oocytes inclinical IVF programmes (Keefe et al, 1995),then the ability of the mitochondrial genometo restore itself to homoplasmic normality forvirtually every newborn (Koehler et al, 1991)(if this assumption is true: data are limited)looks like a remarkable physiological achieve-ment. I suggested above that physiologicalsterility well before functional follicular deple-tion at the menopause, the phenomenon wehave called the oopause (Jansen and De Boer,1998), could be an essential component ofthe required survival strategy of the haploidmitochondrial genome, and could be basedon the simple (and evolutionarily conserved)device of postponement of new mtDNAsynthesis in the new host organism until aftergastrulation. Too few normal mtDNA species
120
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
in the fertilized oocyte and the ovum wouldbe permanently blighted.
As an end-point, however, female sterilitywill not be a favoured option for survivalof a species if it happens too readily. Inevolutionary terms, one would suppose thatthis certainty would surely have causedimmense pressure to have devolved back onthe processes of oogenesis and folliculo-genesis, oocyte competition, and competitionbetween follicles to correct genetic inferiority.I propose that the imperative of conserving themitochondrial genome could be a fundamentalreason why these structures and this systemdeveloped.
The oopause and other enigmaticmanifestations of reproductive
dysfunction
The stringent bottleneck through which themitochondrial genome passes, and by whichmitochondrial DNA is refined and fixed fromone generation to the next, could thus be thesame bottleneck that permits only relativelyyoung eggs with healthy cytoplasm to have ahigh chance of implanting successfully afterfertilization in vivo or in vitro (Jansen, 1995;Faber et al., 1997). At what age can sterilityfrom such a mechanism be expected?
The age at which that the oopausal transitioncan become important, at least commerciallyin the United States, is as early as -33 years,after which donated eggs begin to produceprogressively lower rates of pregnacy, thustending to disqualify recruitment as a paiddonor in US donor egg programmes (Faberet al., 1997). A decrease in fecundability isseen earlier than this, being evident after theage of 30 years, but it is accelerated after 35and it falls sharply after 40. From 42-44years, depending partly on the quality ofthe laboratory, pregnancy with IVF becomesextremely rare (although natural successfulconception is still reported among 1 in 10
women at 45 in societies where contraceptivesare minimal (Lorimer, cited by Guttmacher,1956). The average age at which physiologicalsterility occurs in women can be estimatedfrom populations of women with a low naturalrate of environmentally induced sterility andwho do not practise contraception, such asthe North American Hutterite communities.Hutterite women in the Dakotas, Montana,Alberta and Manitoba have a median age atmarriage of 22 and a total primary sterilityrate of only 3.4%; they conceive an averageof 10.4 times in a reproductive lifetime(Guttmacher, 1956), and their median age atlast conception is -40.5 years (Tietze, 1957),which is 10 years earlier than the medianage at menopause among Western women(Walsh, 1978).
The traditional explanation for decliningoocyte health and embryonic potential withfemale age has been an increasing prevalenceof chromosomal aneuploidy. Does thisexplanation challenge a mitochondrialexplanation? Many authors (see Schon et al.,2000) speculate that it might not.
First, it should be remembered that althoughthe prevalence of aneuploidy rises amongconceptions with age, this prevalence alwaysfalls far short of 100%, irrespective of howmuch fertility falls as female age rises. Evenat the age of 45 years, when the risk ofmiscarriage is >50% of conceptions, a largeminority of miscarriages have a normal karyo-type, and no other diagnosable cause can befound. Second, aneuploidy itself results fromdefective chromosomal-spindle attachment orfunction: mitochondria normally aggregatearound the metaphase spindle in mouseoocytes, implying a local need for ATP produc-tion in spindle formation and molecular motorfunction (Van Blerkom and Runner, 1984; Liand Fan, 1998); oxidative stress has beencausally linked to meiotic nondisjunction(Tarin et al., 1998); defective spindles inpublished electron micrographs of human
121
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

R.P.S.Jansen
oocytes have been noted to have compara-tively few mitochondria in their vicinity(Sathananthan et al., 1986); and a mutationthat prevents mitochondrial binding to meioticspindles in the fruit-fly Drosophila leads tosterility (Basu and Li, 1998). In other words,it might be that mitochondrial dysfunctionin oocytes is more fundamental than, andpossibly even causes, aneuploidy.
From clinical observations (R.P.S.Jansen,unpublished), a woman at the age of theoopausal transition can have repeated karyo-typically normal miscarriages despite success-fully having had several children at an earlierage with the same partner, the final outcomeby the late-30s to mid-40s being otherwiseunexplained infertility. Larsson et al. (1998)showed that transgenic mouse embryos homo-zygous for disruption of mitochondrial tran-scription factor A (a nuclear gene productessential for initiation of mitochondrial DNAreplication—see Clayton, 2000) and trans-ferred to normal mothers will proceed throughimplantation and gastrulation, before ATPproduction fails and embryonic resorptionoccurs. Gastrulation is the point at whichdilution of the oocyte's original mitochondrialcomplement falls below that required tosustain further development (see above). Anequivalent embryonic mitochondrial failure inhumans would be expected to present at orbefore an equivalent time, with failure of theembryonic mass to proceed, namely a blightedovum (the classical term for an anembryonicpregnancy sac). Looking for mitochondrialgenetic causes for sporadic and recurrent mis-carriages at any age could be a fruitful areafor research.
A genomic insufficiency of mtDNA in theoocyte might account for progressively severereproductive dysfunction at any stage beforenew mitochondria are due to be produced,both with natural conception and with IVF,and without necessarily incurring aneuploidy(Lower et al., 1991). Starting with early
122
miscarriages, biochemical pregnancies (sub-clinical miscarriages) could be expected tofollow increasingly severe quantitative orqualitative mitochondrial insufficiency, in turnfollowed by unexplained implantation failure(unexplained infertility or failure to conceivewith IVF/embryo transfer), then slow orarrested cleavage in vitro, fertilization failure,and finally a failure even of ordered pre-ovulatory follicular growth, if follicular cellmitosis can indeed be linked to oocyte health.
In summary, as well as determining whatcould be called an oopause a decade or sobefore menopause, I propose that therecould be a mitochondrial basis behind a num-ber of previously inexplicable reproductivedifficulties, such as unexplained infertility andrecurrent miscarriage. Furthermore, poorlyselected oocytes from superovulated ovariesor imperfect culture conditions in IVF labora-tories (adding an iatrogenic component tothe damage delivered by imperfect oocytemitochondria) could explain the discrepancythat whereas 10% of last conceptions canoccur at > 45 years in natural circumstances(Tietze, 1957), non-donor oocyte IVF concep-tions become exceedingly rare after 42 or 43years. Preliminary quantification of mtDNAfrom superovulated secondary oocytes inwomen of different ages indicate that therecould be a trend towards lower counts withincreasing age (De Boer et al., 1999). Ifso, this could be a widespread phenomenonamong female vertebrates, as a similar age-related decline in oocyte mtDNA has beendescribed in frogs (Callen et al., 1980).
Physiological prevention of pregnancy priorto oocyte depletion could be the ultimateconstraint that prevents degraded mito-chondrial genomes entering or persisting inthe reproducing population. According to thishypothesis, the mitochondrial genome, notexposed to fluctuating external environmentsfor natural selection, is instead subjected to adifferent yet tough test of compliance: the
L
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
need for there to be numerically sufficientnormally functioning mtDNA circles in anoocyte for effective survival of the zygote for2 weeks before new mtDNA synthesis ispermitted. If normal mitochondrial DNAforms are too few in number to outstrip thedilution that divides a fixed number of formsinto the multiplying cells of the conceptus,the conceptus ought to stop developing andpregnancy will then fail. If ageing oocytescarry fewer non-degraded mitochondrialgenomes, this means that the device of post-poning mitochondrial DNA replication untilgastrulation has occurred could substantiallylimit intergenerational mitochondrial genomicdeterioration, albeit at the cost of sterility wellbefore final depletion of follicles and oocytes.For abnormal mitochondrial genomes that areable to slip through whatever selection processis going on (such as those with certain singlepoint mutations), a fitness decline with reducedreproductive capacity in the new host organismis predictable, ranging clinically from the mildto the catastrophically severe (Holme et al,1995; Ferlin et al, 1997). Both of theseclinical phenomena, mitochondrially depend-ent physiological sterility and the spectre ofmitochondrial disease among the offspring,could turn out to be important in the manage-ment of subfertility and sterility in olderwomen.
Implications for therapeutic IVF
What might be done therapeutically for womenfacing physiological but unanticipated sterilityfrom a mitochondrially determined oopausaltransition? Aside from whole egg donation bya younger woman, there are two obviousstrategies for overcoming ooplasmic ageing, atleast while fertilization and normal pronuclearformation in vitro is still possible. There hasalready been clinical experience with both, soit is worth extending the theoretical considera-tions derived above to these manoeuvres.
Cytoplasmic replacement by karyoplastexchange
Replacing the bulk of the mitochondrialgenome could be achieved by swapping thepronuclei of a fertilized oocyte from 'old'cytoplasm to the enucleated fertilized oocyteof a donor (the micromanipulations of thekaryoplasts involved have been shown to beworkable in animal studies; Smith and Alcivar,1993) or by exchanging the nucleus of agerminal vesicle stage oocyte prior to matura-tion. These could be options worth exploring,not just for women in the oopausal transitionbut also for women with clinically importantmtDNA mutations. A number of studies thathighlight practical problems have been per-formed.
First, ooplasmic mitochondria aggregatearound pronuclei about to undergo syngamy(Van Blerkom and Runner, 1984), so it isvirtually impossible not to include a substantialpopulation of mitochondria with the takingof a karyoplast from a pronuclear zygote,removing useful mitochondria from therecipient cytoplasm (the donated oocyte) andtransferring questionable mitochondria alongwith the infertile couple's pronuclei into thenormal remaining recipient cytoplasm. Studiesin mice have indicated that such karyoplast-associated mitochondria might be favoured,even during cleavage (Meirelles and Smith,1998).
Second, there are data implicating interfer-ence with essential nuclear-mitochondrialdialogue to explain decreased cytoplasmicperformance and increased failure to formblastocysts among reconstructed zygotes madefrom interspecific crosses among mice (Smithand Alcivar, 1993; Nagao et al, 1998). Thedata are difficult to interpret. There are alsoindications that there could be considerabletolerance in nuclear-mitochondrial dialogue inearly embryos, even across species (Dominkoet al, 1999). Intraspecific transfer of calf
123
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

R.P.S.Jansen
embryo nuclei from separated early morulablastomeres into enucleated secondary oocyteshas been associated with high variability inbody size among offspring (Gartner et al,1998) - perhaps an unsurprising consequence,given the greatly different state of nuclear andmitochondrial transcription between these twostages. Whether difficulties in nuclear-mito-chondrial communication will accompanyintraspecific, possibly intrafamilial exchangesof comparably staged human pronuclearembryos or 2-cell embryos awaits furtherresearch.
Clinical experiments involving germinalvesicle-stage nuclear transfers in humans havenot proven successful to date (Zhang et al,1998), but the main difficulties could lie withthe in-vitro maturation these oocytes haverequired.
Cytoplasmic transfusion or cytoplasttransplantation
Without its having been stated explicitly, mito-chondrial supplementation could be the reasonbehind the birth of infants after transfer ofanucleate donor oocyte cytoplasm to recipientsecondary oocytes reported in a group of olderinfertile women (Cohen et al, 1997, 1998).Women who were aged >35 years weretreated; women with a record of good numbersof eggs retrieved, but with multiple unsuccess-ful embryo transfers after IVF: the embryoswere deemed poor on morphological criteria.Cohen et al. found that normal fertilizationwas higher after direct injection of 7-14% ofooplasmic volume at intracytoplasmic sperminsertion (which produced a 63% fertiliza-tion rate, improved embryo morphology andseveral pregnancies) than after electrofusionof donor ooplasts 40-90 min before ICSI(which produced a 23% fertilization rate, pro-nuclear anomalies, no correction of embryomorphology, and no pregnancies). At least 10
delivered or ongoing pregnancies have nowbeen reported (see Barritt et al., 2000).
Nuclear and mtDNA fingerprinting profilesof the donor and recipient of such cytoplasmictransfusion for oocytes were studied at amnio-centesis in the first ongoing oocytoplasmictransfusion pregnancy, later delivered at term,and no donor profiles for the donor genome(nuclear or mitochondrial) were detected byPCR and sequencing (Cohen et al., 1997;1998). The infant was clearly the daughterof her parents' nuclear genomes. Donor andrecipient mtDNA differed at positions ntl6219and ntl6311 (part of the small, non-coding,D-loop region of the genome, where poly-morphisms are frequent) and an examinationwas undertaken that excluded any significantsurvival of donated mtDNA. It would appearfrom these limited data that the donated mito-chondria were perhaps sufficient to assistembryonic survival beyond implantation andgastrulation but, at least in this case, then didnot take part in the subsequent resumption ofmitochondrial replication.
It is plausible that children born after cyto-plasmic transfusion of secondary oocytes willbe born with a mitochondrial genome of alevel of fitness that would normally not becompatible with survival. This need not bebecause of inadvertent injection of mutantmtDNA, but rather would represent theeffects of permitted replication of a suboptimalhost oocyte mtDNA complement throughtemporary augmentation by donated mito-chondria. There could be a life-long risk ofprematurely developing mitochondrial diseaseas age-related mtDNA mutations accumulate(Wallace, 1992; Cortopassi and Arnheim,1993; Shigenaga et al, 1994; Brown andWallace, 1994) on a compromised initial mito-chondrial genotype. These concerns aretheoretical, but the technique is so novel, andso potentially powerful, that they need tobe considered. Among male children with amitochondrial genome of suboptimal fitness,
124
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
infertility is more likely than average (Frankand Hurst, 1996), and female children mightplausibly themselves undergo their ownoopausal transition early (see Medvedev, 1981,for a pre-mtDNA account of thefundamentals involved: in other species thereis some evidence of improvement in female-line fertility for subsequent generations ifreproduction occurs at an earlier age).
Careful follow-up of the children who resultfrom ooplasmic transfusion will be importantand a detailed analysis of their mitochondrialgenomes should be performed.
Conclusion: testable hypotheses
The evolutionary need for faithful inheritanceof mtDNA, an asexually reproducinggenome, is proposed to be the imperativebehind well-known physiological and patho-logical phenomena in mammalian reproduc-tion, particularly oogenesis, folliculogenesis,apparent competition between follicles, andfemale age-related sterility well beforemenopause.
Other authors have observed that mtDNAcopy numbers in secondary oocytes could beonly slightly greater than the mitochondrialorganelle number and that mtDNA replicationin secondary oocytes that are fertilized issuspended for a significant time into embry-onic development. These observations requirefurther testing, but if they are essentiallycorrect then it is reasonable to postulate:(i) that the female germ-line mitochondrialgenome is persistently haploid: that is, thatduring all stages of embryogenesis andoogenesis the effective number of mtDNAmolecules in female germ line cells willapproximate the number of mitochondrialorganelles, and (ii) that the greatest restrictionin mtDNA copy number between generations('the bottleneck') will occur between the timeof resumption of mtDNA replication in theembryo and the commencement of migration
of the primitive germ cell from the yolksac. It has also been postulated (iii) thatcompetition between ovarian follicles ulti-mately reflects the relative mitochondrialgenomic integrity of the contained oocyte;and (iv) that age-related embryonic failure(including miscarriage and sterility well beforemenopause in humans - the oopause) couldbe a consequence of a numerical shortageof qualitatively adequate mtDNA moleculesamong ovulated oocytes.
ReferencesAllen, J.F. (1996) Separate sexes and the mitochondrial
theory of ageing. J. Theor. Biol, 180, 135-140.Baker, T.G. (1963) A quantitative and cytological study
of germ cells in human ovaries. Proc. R. Soc. Lond.B, 158, 417^33.
Baker, T.G. and Franchi, L.L. (1967) The fine structureof oogonia and oocytes in human ovaries. J. Cell Sci.,2, 213-224.
Barritt, J.A., Brenner, C.A., Willadsen, S. et al. (2000)Spontaneous and artificial changes in ooplasmicmitochondria. Hum. Reprod., 15 (Suppl. 2), 207-217.
Basu, J. and Li, Z. (1998) The Des-1 protein, required forcentral spindle assembly and cytokinesis, is associatedwith mitochondria along the meiotic spindle apparatusand with the contractile ring during male meiosis inDrosophila melanogaster. Mol. Gen. Genet., 259,664-673.
Beijnink, F.B., Walker, C.W. and Voogt, P.A. (1984)An ultrastructural study of relationships between theovarian hemal system, follicle cells and primaryoocytes in the sea star, Asterias rubens. Implicationsfor oocyte nutrition. Cell Tiss. Res., 238, 339-347.
Blok, R.B., Gook, D.A., Thorburn, D.R. and Dahl,H.H.M. (1997) Skewed segregation of the mtDNA nt8993 (T—>G) mutation in human oocytes. Am. J. Hum.Genet, 60, 1495-1501.
Brown, M.D. and Wallace, D.C. (1994) Molecular basisof mitochondrial disease. J. Bioenerg. Biomembr., 26,273-289.
Buccione, R.,Schroeder, A.C. and Eppig, J.J. (1990)Interactions between somatic cells and germ cellsthroughout mammalian oogenesis. Biol. Reprod., 43,543-547.
Callen, J.C., Dennebouy, N. and Mounolou, J.C. (1980)Development of the mitochondrial mass andaccumulation of mtDNA in previtellogenic stages ofXenopus laevis oocytes. J. Cell Sci., 41, 307-320.
Chao, L., Tran, T.T. and Tran, T.T. (1997) The advantageof sex in the RNA virus phi6. Genetics, 147, 953-959.
125
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

R.P.SJansen
Clayton, D.A. (2000) Transcription and replication ofthe mitochondrial genome. Hum. Reprod., 15 (Suppl.2), 11-17.
Cohen, J., Scott, R., Schimmel, T. et al. (1997) Birthof infant after transfer of anucleate donor oocytecytoplasm into recipient eggs. Lancet, 350, 186-187.
Cohen, J., Scott, R., Alikani, M. et al. (1998) Ooplasmictransfer in mature human oocytes. Mol. Hum. Reprod.,4, 269-280.
Cortopassi, G. and Arnheim, N. (1993) Accumulationof mitochondrial DNA mutation in normal aging brainand muscle. In DiMauro, S. and Wallace, D.C. (eds),Mitochondrial DNA in Human Pathology. RavenPress, New York, USA, pp. 125-136.
Davis, A.F. and Clayton, D.A. (1996) In situ localizationof mitochondrial DNA replication in intactmammalian cells. J. Cell BioL, 135, 883-893.
Dawid, I.B. (1972) Cytoplasmic DNA. In Biggers, J.D.and Schuetz, A.W. (eds), Oogenesis. University ParkPress, Baltimore, USA, pp. 215-226.
De Boer, K.A., Jansen, R.P.S., Leigh, D. and Mortimer,D. (1999) Quantification of mtDNA copy number inthe human secondary oocyte. [Abstr. no. 0-165]. Hum.Reprod., 14 (Abstract Book), 91-92.
Dominko,T., Mitalipova,M., Haley,B. et al. (1999)Bovine oocyte cytoplasm supports development ofembryos produced by nuclear transfer of somatic cellnuclei from various mammalian species. BioL Reprod.,60, 1496-1502.
Dvorak, M. and Tesarik, J. (1985) Differentiation ofmitochondria in the human preimplantation embryogrown in vitro. Scr. Med. (Brno), 58, 161-170.
Ebert, K.M., Liem, H. and Hecht, N.B. (1988)Mitochondrial DNA in the mouse preimplantationembryo. J. Reprod. Fertil, 82, 145-149.
El Meziane, A., Callen, J.-C. and Mounolou, J.-C. (1989)Mitochondrial gene expression during Xenopus laevisdevelopment: a molecular study. EMBO J, 8, 1649-1655.
Eppig, J.J. (1979) A comparison between oocyte growthin coculture with granulosa cells and oocytes withgranulosa cell-oocyte junctional contact maintainedin vitro. J. Exp. ZooL, 209, 345-353.
Eppig, J.J., Chesnel, R, Hirao, Y. et al. (1997) Oocytecontrol of granulosa cell development: how and why.Hum. Reprod., 12 (Natl. Suppl.) JBFS, 2, 127-132.
Faber, B.M., Mercan, R., Hamacher, P. et al. (1997) Theimpact of an egg donor's age and her prior fertilityon recipient pregnancy outcome. Fertil. Steril., 68,370-372.
Faddy, M.J., Gosden, R.G., Gougeon, A. et al. (1992)Accelerated disappearance of ovarian follicles in mid-life: Implications for forecasting menopause. Hum.Reprod., 7, 1342-1346.
Ferlin, T., Landrieu, P., Rambaud, C. et al. (1997)Segregation of the G8993 mutant mitochondrial DNA
through generations and embryonic tissues in a familyat risk of Leigh syndrome. J. Pediatn, 131, 447-449.
Frank, S.A. and Hurst, L.D. (1996) Mitochondria andmale disease. Nature, 383, 224-224.
Gardner, D.K. and Sakkas, D. (1993) Mouse embryocleavage, metabolism and viability: Role of mediumcomposition. Hum. Reprod., 8, 288-295.
Gartner, K., Bondioli, K., Hill, K. and Rapp, K. (1998)High variability of body sizes within nucleus-transfer-clones of calves: artifacts or a biological feature?Reprod. Dom. Anim., 33, 67-75.
Gondos, B. (1987) Comparative studies of normal andneoplastic ovarian germ cells: 1. Ultrastructure ofoonia and intercellular bridges in the fetal ovary. Int.J. Gynecol. Pathol., 6, 114-123.Gondos, B., Westergaard, L. and Byskov, A.G. (1986)Initiation of oogenesis in the human fetal ovary:ultrastructural and squash preparation study. Am. J.Obstet. Gynecol, 155, 189-195.
Gosden, R.G. (1995) Ovulation 1: oocyte developmentthroughout life. In Grudzinskas, J.G. and Yovich, J.L.(eds), Gametes - the Oocyte. Cambridge UniversityPress, Cambridge, UK, pp. 119-149.
Grazul-Bilska, A.T., Reynolds, L.P. and Redmer, D.A.(1997) Gap junctions in the ovaries. BioL Reprod.,57, 947-957.
Guttmacher, A.F. (1956) Factors affecting normalexpectancy of conception. J. Am. Med. Assoc, 161,855-860.
Hasegawa, M., Cao, Y. and Yang, Z.H. (1998)Preponderance of slightly deleterious polymorphismin mitochondrial DNA: Nonsynonymous/synonymousrate ratio is much higher within species than betweenspecies. Mol. BioL Evol., 15, 1499-1505.
Hauswirth, W.W. and Laipis, P.J. (1985) Transmissiongenetics of mammalian mitochondria: a molecularmodel and experimental evidence. In Quagliariello,E., Slater, E.C., Palmieri, F. et al. (eds), Achievementsand Perspectives of Mitochondrial Research, vol.II, Biogenesis. Elsevier Science, Amsterdam, TheNetherlands, pp. 49-59.
Hertig, A.T. and Adams, E.C. (1967) Studies on thehuman oocyte and follicle. I. Ultrastructural andhistochemical observations on the primordial folliclestage. J. Cell BioL, 34, 647-675.
Holme, E., Tulinius, M.H., Larsson, N.-G. and Oldfors,A. (1995) Inheritance and expression of mitochondrialDNA point mutations. Biochim. Biophys. Acta, 1271,249-252.
Howell, N. (1997) mtDNA recombination: What doin vitro data mean. Am. J. Hum. Genet., 61, 19-22.
Jansen, R.P.S. (1995) Older ovaries: ageing andreproduction. Med. J. Aust., 162, 623-624.
Jansen, R.P.S. (2000) The origin and persistence of themitochondrial genome. Hum. Reprod., 15 (Suppl. 2),1-10.
126
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

Germline transmission of mitochondria
Jansen, R.P.S. and De Boer, K.A. (1998) The bottleneck:mitochondrial imperatives in oogenesis and ovarianfollicular fate. Mol. Cell. Endocrinol., 145, 81-88.
Keefe, D.L., Niven-Fairchild, T., Powell, S. andBuradagunta, S. (1995) Mitochondrial deoxy-ribonucleic acid deletions in oocytes and reproductiveaging in women. Fertil. Steril, 64, 577-583.
Koehler, CM., Lindberg, G.L., Brown, D.R. et al. (1991)Replacement of bovine mitochondrial DNA by asequence variant within one generation. Genetics, 129,247-255.
Krakauer, D.C. and Mira, A. (1999) Mitochondria andgerm cell death. Nature, 400, 125-126.
Lanzavecchia, G. and Mangioni, C. (1964) Etude de lastructure et des constituants du follicule humain dansl'ovaire fetal. I. - Le follicule primordial. J.Microscopie, 3, 447—4-64.
Larsson, N.-G., Tulinius, M.H., Holme, E. et al. (1992)Segregation and manifestations of the mtDNAtRNA(Lys) A—>G(8344) mutation of myoclonusepilepsy and ragged-red fibers (MERFF) syndrome.Am. J. Hum. Genet., 51, 1212
Larsson, N.-G., Wang, J., Wihelmsson, H. et al. (1998)Mitochondrial transcription factor A is necessary formtDNA maintenance and embryogenesis in mice.Nature Genet., 18, 231-236.
Lawrence, T.S., Beers, W.H. and Gilula, N.B. (1978)Transmission of hormonal stimulation by cell-to-cellcommunication. Nature, 272, 501-506.
Leidy, L.E., Godfrey, L.R. and Sutherland, M.R. (1998)Is follicular atresia biphasic? Fertil. Steril, 70, 851-859.
Li, C.J. and Fan, B.Q. (1998) Changes in the 3-dimensional distribution of mitochondria duringmeiotic divisions of mouse oocytes. Theriogenology,48, 33-41.
Li, R. and Mather, J.P. (1997) Lindane, an inhibitor ofgap junction formation, abolishes oocyte directedfollicle organizing activity in vitro. Endocrinology,138, 4477-4480.
Lower, A.M., Mulcahy, M.T. and Yovich, J.L. (1991)Chromosome abnormalities detected in chorionicvillus biopsies of failing pregnancies in a subfertilepopulation. Br. J. Obstet. Gynaecoi, 98, 1228-1233.
Lynch, M. and Blanchard, J.L. (1998) Deleteriousmutation accumulation in organelle genomes.Genetica, 103, 29-39.
Lynch, M., Burger, R., Butcher, D. and Gabriel, W.(1993) The mutational meltdown in asexualpopulations. J. Hered., 84, 339-344.
Makabe, S. and Motta, P.M. (1989) Migration of humangerm cells and their relationship with the developingovary: ultrastructural aspects. In Motta, P.M. (ed.),Developments in Ultrastructure of Reproduction. AlanR.Liss, New York, USA, pp. 41-54.
Mansouri, A., Gaou, I., Fromenty, B. et al. (1997)Premature oxidative aging of hepatic mitochondrial
DNA in Wilson's disease. Gastroenterology, 113,599-605.
Medvedev, Z.A. (1981) On the immortality of the germline: genetic and biochemical mechanisms. A review.Mech. Ageing Develop., 17, 331-359.
Meirelles, F.V. and Smith, L.C. (1998) Mitochondrialgenotype segregation during preimplantationdevelopment in mouse heteroplasmic embryos.Genetics, 148, 877-883.
Michaels, G.S., Hauswirth, W.W. and Laipis, P.J. (1982)Mitochondrial DNA copy number in bovine oocytesand somatic cells. Dev. Biol., 94, 246-251.
Mignotte, F., Tourte, M. and Mounolou, J.C. (1987)Segregation of mitochondria in the cytoplasm ofXenopus vitellogenic oocytes. Biol. Cell, 60, 97-102.
Motta, P.M. (1995) Ultrastructure of human reproductionfrom folliculogenesis to early embryo development.A review. Ital. J. Anat. Embryol, 100, 9-52.
Nachman, M.W. (1998) Deleterious mutations in animalmitochondrial DNA. Genetica, 103, 61-69.
Nagao, Y, Totsuka, Y, Atomi, Y. etal. (1998) Decreasedphysical performance of congenic mice with mismatchbetween the nuclear and the mitochondrial genome.Genes Genet. Syst., 73, 21-27.
Olivo, P.D., Van de Walle M.J., Laipis, P.J. andHauswirth, W.W. (1983) Nucleotide sequenceevidence for rapid genotypic shifts in the bovinemitochondrial DNA D-loop. Nature, 306, 400-402.
Perez, G.I., Knudson, CM., Leykin, L. et al. (1997)Apoptosis-associated signaling pathways are requiredfor chemotherapy-mediated female germ celldestruction. Nature Med., 3, 1228-1232
Piko, L. and Chase, D.G. (1993) Role of themitochondrial genome during early development inmice. Effects of ethidium bromide andchloramphenicol. J. Cell Biol., 58, 357-378.
Piko, L. and Matsumoto, L. (1976) Number ofmitochondria and some properties of mitochondrialDNA in the mouse egg. Dev. Biol., 49, 1-10.
Piko, L. and Taylor, K.D. (1987) Amounts ofmitochondrial DNA and abundance of somemitochondrial gene transcripts in early mouseembryos. Dev. Biol., 123, 354-374.
Pinz, K.G. and Bogenhagen, D.F. (1998) Efficient repairof abasic sites in DNA by mitochondrial enzymes.Mol. Cell. Biol., 18, 1257-1265.
Ryoji, M., Katayama, H., Fusamae, H. et al. (1996)Repair of DNA damage in a mitochondrial lysateof Xenopus laevis oocytes. Nucleic Acids Res., 15,4057-W62.
Sathananthan, A.H., Trounson, A.O. and Wood, C.(1986) Atlas of Fine Structure of Human SpermPenetration, Eggs and Embryos Cultured In Vitro.Praeger, New York, pp. 109-109.
Sathananthan, H., Ng, S.-C, Bongso, A. et al. (1993)Visual Atlas of Early Human Development for Assisted
127
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022

R.P.S.Jansen
Reproductive Technology. National UniversityHospital Department of Obstetrics and Gynaecology,Singapore.
Schagen van Leeuwen, J.H., Christiaens, G.C. andHoogenraad, T.U. (1991) Recurrent abortion and thediagnosis of Wilson's disease. Obstet. Gynecol., 78,547-549.
Schon, E.A., Kim, S.H., Ferreira, J.C. et al. (2000)Chromosomal nondisjuntion in human oocytes: isthere a mitochondrial connection? Hum. Reprod., 15(Suppl. 2), 160-172.
Serhal, P.F. and Craft, I.L. (1989) Oocyte donation in61 patients. Lancet, i, 1185-1187.
Shepard, T.H., Muffley, L.A., Smith, L.T. (1998)Ultrastructural study of mitochondria and their cristaein embryonic rats and primate (N-nemistrina) Anat.Rec, 3, 383-392.
Shepard, T.H., Muffley, L.A., Smith, L.T. (2000)Mitochondrial ultrastructure in embryos afterimplantation. Hum. Reprod., 15 (Suppl. 2), 218-228.
Shigenaga, M.K., Hagen, T.M. and Ames, B.N. (1994)Oxidative damage and mitochondrial decay in aging.Proc. NatlAcad. Sci. USA, 91, 10771-10778.
Smith, L.C. and Alcivar, A.A. (1993) Cytoplasmicinheritance and its effects on development andperformance. J. Reprod. Fertii, 48 (Suppl.), 31-43.
Tarfn, J.J., Vendrell, F.J., Ten, J. and Cano, A. (1998)Antioxidant therapy counteracts the disturbing effectsof diamide and maternal ageing on meiotic divisionand chromosomal segregation in mouse oocytes. Mol.Hum. Reprod, 4, 281-288.
Taylor, K.D. and Piko, L. (1995) Mitochondrialbiogenesis in early mouse embryos: expression of themRNAs for subunits IV, Vb,and VIIc of cytochromec oxidase and subunit 9 (PI) of H(+)-ATP synthase.Mol. Reprod. Dev., 40, 29-35.
Thyagarajan, B., Padua, R.A. and Campbell, C. (1996)Mammalian mitochondria possess homologous DNArecombination activity. J. Biol. Chem., 271, 27536-27543.
Tietze, C. (1957) Reproductive span and rate ofreproduction among Hutterite women. Fertii. Sterii,8, 89-97.
Tilly, J.L. (1996) Apoptosis and ovarian function. Rev.Reprod, 1, 162-172.
Tourte, M., Mignotte, F. and Mounolou, J.-C. (1984)Heterogeneous distribution and replication activity ofmitochondria in Xenopus laevis oocytes. Eur. J. CellBiol, 34, 171-178.
Tsafriri, A., Dekel, N. and Bar-Ami, S. (1982) The roleof maturation inhibitor in follicular regulation ofoocyte maturation. J. Reprod. Fertii., 64, 541-551.
Van Blerkom, J. and Runner, M.N. (1984) Mitochondrialreorganization during resumption of arrested meiosisin the mouse oocyte. Am. J. Anat., Ill, 335-355.
Van Blerkom, J., Davis, P.W. and Lee, J. (1995) ATPcontent of human oocytes and developmental potential
and outcome after in-vitro fertilization and embryotransfer. Hum. Reprod., 10, 415^24.
Van Blerkom, J., Antczak, M. and Schrader, R. (1997)The developmental potential of the human oocyte isrelated to the dissolved oxygen content of follicularfluid: Association with vascular endothelial growthfactor levels and perifollicular blood flowcharacteristics. Hum. Reprod., 12, 1047-1055.
Wallace, D.C. (1992) Mitochondrial genetics: a paradigmfor aging and degenerative diseases. Science, 256,628-632.Walsh, R.J. (1978) The age of the menopause ofAustralian women. Med. J. Aust., 2, 181-181.
Wartenberg, H. (1974) Spermatogenese-Oogenese: eincyto-morphologischer Vergleich. Verh. Anat. Ges., 68,63-92.
Zhang, J., Wang, C.-W, Krey, L. et al. (1998) In vitromaturation (IVM) of human preovulatory oocytesreconstructed by germinal vesicle (GV) transfer. InKempers, R.D., Cohen, J., Haney, A.F. and Younger,J.B. (eds), Fertility and Reproductive Medicine.Elsevier Science, Amsterdam, The Netherlands,pp. 629-635.
128
Dow
nloaded from https://academ
ic.oup.com/hum
rep/article/15/suppl_2/112/619766 by guest on 06 March 2022