genetic requirements for the single-strand annealing pathway of double-strand break repair in...
TRANSCRIPT

Copyright 0 1996 by the Genetics Society of America
Genetic Requirements for the Single-Strand Annealing Pathway of Double-Strand Break Repair in Saccharomyces cerezkiae
Evgeny L. Ivanov,' Neal Sugawara, Jacqueline Fishman-Lobell' and James E. Haber
Rosenstiel Basic Medical Sciences Research Center and Department of Biology, Brandeis University, Waltham, Massachusetts 02254-91 10
Manuscript received August 25, 1995 Accepted for publication November 14, 1995
ABSTRACT HO endonuclease-induced double-strand breaks (DSBs) within a direct duplication of Eschm'chia coli
lacZ genes are repaired either by gene conversion or by single-strand annealing (SSA), with >80% being SSA. Previously it was demonstrated that the RAD52 gene is required for DSB-induced SSA. In the present study, the effects of other genes belonging to the RAD52 epistasis group were analyzed. We show that RAD51, RAD54, RAD55, and RAD57 genes are not required for SSA irrespective of whether recombination occurred in plasmid or chromosomal DNA. In both plasmid and chromosomal constructs with homologous sequences in direct orientation, the proportion of SSA events over gene conversion was significantly elevated in the mutant strains. However, gene conversion was not affected when the two laczsequences were in inverted orientation. These results suggest that there is a competition between SSA and gene conversion processes that favors SSA in the absence of RAD51, RAD54, RAD55 and RAD57. Mutations in RAD50 and XRS2 genes do not prevent the completion, but markedly retard the kinetics, of DSB repair by both mechanisms in the lacZ direct repeat plasmid, a result resembling the effects of these genes during mating-type ( M A 7 ) switching.
E NVIRONMENTAL stresses induce a wide variety of injuries to DNA, forcing cells to restore the integ-
rity of the genome by DNA repair. Among the different forms of DNA damage, DNA double-strand breaks (DSBs) are considered as the most harmful, because unrepaired DSBs result in the loss of broken chromo- somes, leading to the death of haploid cells or to aneu- ploidy in diploids. Considerable evidence has accumu- lated during the past 20 years that in yeast the major cellular mechanism of DSB repair involves homologous recombination, culminating in the DSB repair model (SZOSTAK et al. 1983). In this model, a DSB or a gap is repaired by a gene conversion event involving broken ends that invade and copy DNA from an intact, homolo- gous donor copy (Figure la).
Further supporting the idea that DSB repair involves homologous recombination is a striking overlap in the genetic requirements for homologous recombination and for DNA repair after treatments known to induce DSBs (e.g. , ionizing radiation). Mutations in R A D S I , RAD52, RAD54, RAD55, and RAD57 genes confer high levels of sensitivity to ionizing radiation; the very same mutations demonstrate defects in homologous recom- bination both in vegetative cells and during meiosis
University, 415 South St., Waltham, MA 022549110. E-mail: [email protected]
02139.
Currespunding author: James E. Haber, Rosenstiel Center, Brandeis
Present address: Transkaryotic Therapies, Inc., Cambridge, MA
' Present address: Center for Neurologic Diseases, Brigham and Women's Hospital, Boston, MA 01801.
Genetics 142 699-704 (March, 1996)
(reviewed in PETES et al. 1991). More recently, the RFAI gene coding for the large subunit of the single-stranded DNA-binding protein RPA was shown to be required for both DSB-induced gene conversion and DNA repair (FIRMENICH et al. 1995).
During the last few years, it has become evident that another pathway exists for DSB repair via homologous recombination. This pathway, called single-strand an- nealing (SSA), relies on the presence of two homolo- gous sequences in direct orientation flanking the site of a DSB (Figure lb) . According to current schemes of SSA, bidirectional 5' to 3' degradation of the DSB ends leads to the exposure of complementary sequences, which then anneal. Subsequent removal of nonhomolo- gous 3'-ended tails followed by ligation completes the process. The outcome of this reaction is the creation of a deletion of one repeat along with the intervening sequence. The mechanism of SSA was first postulated to explain some features of extrachromosomal recombi- nation in mammalian cells (LIN et al. 1984, 1990). We have shown that the SSA process operates in yeast cells as well, representing an alternative, kinetically separa- ble and independently modulated pathway of DSB re- pair (FISHMAN-LOBELL et al. 1992).
SSA is a truly homologous recombination process, in that it displays essentially the same homology require- ments between interacting sequences as does gene con- version ( SUGAWARA and HABER 1992; N. SUGAWARA, un- published results). It is also evident that both SSA and gene conversion strongly depend on the 5' to 3' degra- dation of DSB ends (CAO et al. 1990; WHITE and HABER

694 E. L. Ivanov et al.
/ \
Gene conversion without Gene conversion
crossing-over with crossing-over
/ FIGURE 1.-Two alternative pathways of DSB repair as adapted for the plasmid sub- strate employed in this work. (a) DSB re- pair by gene conversion could theoretically yield both conversions without crossing- over and with crossing-over (SZOSTAK et al. 1983). However, repair of an HO-induced DSB occurring in direct repeats of the lncZ gene was shown to occur without the for- mation of the reciprocal crossover product (RUDIN and HABER 1988; FISHMAN-LOBEI.I. et al. 1992). (b) SSA pathway repairs a DSB leading to a deletion of one of the direct repeats. In this pathway, the small recipro- cal circular Droduct is not exDected to be
0-Q /
Deletion product
formed. For discussion of current schemes of SSA, see IVANOV and HABER (1995).
a b
1990; SUN et a/. 1991; FISHMAN-LOBELL et al. 1992; SU- G A W ~ and HABER 1992). Moreover, the extent of this degradation was shown to be under similar genetic con- trol in both pathways (WHITE and HABER 1990; SUGA- WARA and HABER 1992; IVANOV et al. 1994). At the same time, SSA is clearly different from gene conversion in that it does not involve such molecular steps as strand invasion and the formation and resolution of Holliday structures. Thus one might expect that the genetic re- quirements for SSA would not be identical to those for gene conversion, although some overlap should exist.
At the present time, our knowledge of the genetic control of the SSA process is not complete. Previous results from our lab have demonstrated that two nucleo- tide excision repair genes, R A D 1 and RADIO, are re- quired for the successful completion of the SSA process, being involved in the removal of 3"ended regions of nonhomology that are created during annealing (Fig- ure lb) (FISHMAN-LOBELL and HABER 1992; IVANOV and HABER 1995). However, the formation of these nonho- mologous ends is a characteristic feature of the SSA process and is not expected to occur during recombina- tion by gene convesion involving fully homologous se- quences. In agreement with this assumption, rad1 and rad10 mutants show the wild-type proficiency in inter- chromosomal gene conversion and IMAT switching (SCHIESTL and P m H 1988, 1990; FISHMAN-LOBELL and HABER 1992) and are only marginally sensitive to X-ray and gamma irradiation.
Deletion of the RAD52 gene significantly reduces the extent of repair by the SSA mechanism (FISHMAN-LO- BELL et al. 1992; SUGAWARA and HABER 1992). However, it seems that this reduction is not as severe as the effects
of rad52 mutations on gene conversion (see, for exam- ple, RATTRAY and SYMINGTON 1994). Moreover, the requirement for RAD52 during SSA can be overcome by substantially increasing the amount of homology between recombining sequences ( OZENBERGER and ROEDER 1991). The roles for other RAD genes belong- ing to the RAD52 epistasis group in the SSA process are not clear. Mutations in RAD51, RAD54, RAD55, and RAD57genes do not prevent deletion formation during spontaneous mitotic recombination between naturally occurring and artificially constructed direct repeats; on the contrary, deletion formation is stimulated by these mutations (FAN and KLEIN 1994; MCDONALD and ROTHSTEIN 1994; AGUILERA 1995; LIEFSHITZ et al. 1995). However, the major problem with interpreting these data is that SSA is only one of a few possible mechanisms of deletion formation during direct-repeat recombina- tion (reviewed in KLEIN 1995), and the fraction of events being processed through the SSA pathway is un- known. In addition, the identity of the lesion initiating a recombination event (DSB us. single-stranded break) cannot be revealed in any system employing spontane- ous recombination.
To gain more insight into the genetic control of the SSA pathway of DSB repair in yeast, we studied the effects of RAD50, RAD51, RAD52, RAD54, RAD55, RAD57, and XRS2 genes on the repair of a single, HO endonuclease-induced DSB, using two directly repeated lacZgenes of Escherichia coli as a recombination substrate (RUDIN et al. 1989; FISHMAN-LOBEI,I~ and HABER 1992; FISHMAN-LOBELL et al. 1992; IVANOV and HABER 1995). The main advantage of this experimental system for studying the genetic control of SSA is that >80% of

69.5
Psfl ! PSI I a
H~ndlll
Hindlll
conversion without
crossover
PSf I
Psf I
Sma I
- PSf I
Sma I
Psf I Psf I
strand annealing
'- Sma I
' Pst I
Psf I A conversion
without cross over
conversion
- PSf I
Psfl'
pIH6X3 (AIK)l~ssl.:KIIRr\ PI n/. 1992); IL11152, pSM'L0 (S(:blIl.l)
IC4/)55, pSTLll ( L o v ~ n and MORTIMER 1987); RAD57, pSM.51 (S(:l l l l ,D P I n/. 1983); XR72, pE140 (I\ 'AXO\J d n/. 1994).
Strains and media: The strains of Snrrhommy-rs rm-misinr usrtl i n this study are listed i n Table 1. All of these strains arc MATa-inr isogenic derivatives of the previously described strain.JKM40 (I\:\sov and H.ARER 1995); the MATa-inc muta- tion prevents HO cutting. These strains also cary the HO cnclonrtclcase gene under control of thc inducible GAL pro- moter, thc whole construction bring integrated into chromo- somal AlK3 locr~s (SASI)IX.I. and ZAKIAN 1993). For galactose- induction experiments, strains E1534 through El.356 were transfc)rnmed w i t h plasmids pJF6 or pIF5. Strain E1558 and its dcrivativcs bear the /tr%direct repeat integrated into the ufd- 5210~11s on chromosome V ( R ~ W N P/ nl. 1989; see also Figwe h ) made by integrati\ve transformation of EI.534 with plasmid pNR28. The rad mutations were introduced into E1534 and El.5.38 strains by the one-step gene disr~lption/replacement mcthotl (RO~IS'I 'EIN 1983) using the plasmids listed above. For details of strain constructions, see the corresponding ref- erences. All the strain constructions were verified by Southern blot analysis. The lithium acetate method ( I n ) P/ nl. 198.7) was u s e d for yeast transformation.
YEPI) medium and synthetic complete media lacking Ieu- cine (S<>Lcu) o r uracil (SGUra) were described previously (SI II~KMAN P/ nl. 1986). YEPGly and YEPGal were 1 % yeast
/'/ N / . 1983); I W 1 5 4 , pI,s101 (IL\TrR.\Y and ~ W l N ( ; l O N 1995);

696
TABLE 1
Yeast strains
Strain Genotype
E1534
E1548 E1549 E1550 E1551 E1552 E1553 E1556 E1558
E1559 E1560 E1561
MAE-inc adel udd::GAI,::HO leu2-3,
EI534, but rad50::hisG::URA3::hisG E1534, but rud51::URA3 EI534, but ,rad54::UKA3 EI534, but rad55::LEUZ EI534, but rad57::LElD EI534, but xrs2::LEUZ EI534, but md52::LEUZ EI534, but with direct lac% duplication
integrated at u r d - 5 2 EI558, but rad51::IJR43 EI558, but mdjZ::LEU2 EI558, but rad54::URM
~~~~
112 trpl u r d 5 2 Rw'
extract-2% BdctO Peptone media supplemented with 3% (v/v) glycerin or 2% (w/v) galactose, respectively. Synthetic complete medium containing X-Gal was prepared as de- scribed (RUBY et al. 1983). YEPD medium containing 0.015% (v/v) methyl methane sulfonate (MMS) was used to follow the Rad- phenotype. Cultures were incubated at 30", unless otherwise indicated.
Induction of DSBs: Strains to be tested were streaked to single colonies on SCLeu/Ura (for strains bearing pJF6 or pJF5 plasmids) or YEPD (for strains bearing chromosomal integration of Zad) plates. Liquid precultures (5 ml) in the same medium were inoculated from single colonies, grown to midlog phase, and transferred to 50 ml of YEPGly for 8- 10 hr. These cultures were used to inoculate 0.5 liter cultures in YEPGly followed by the growth overnight to a cell density of lo7 cells per ml. At the time 0, an aliquot of cells was removed, while the rest of the culture was harvested and sus- pended in the same volume of warmed YEPGal, and incuba- tion was continued for 30 min. The cells were harvested again and suspended in an equal volume of warmed YEPD, an ali- quot of cells was removed, and incubation resumed with sam- ples ofthe cells being withdrawn at the time points indicated. Appropriate dilutions of cells taken before induction (0 hr) and after induction (0.5 hr) were spread on YEPD plates, grown to colonies, and replica plated on SC-I,eu/Ura plates to measure loss of pJF6 or pJF5 (if applicable) and on X-Gal plates to score for the LacZ phenotype.
In the case of HO induction on plates, 5-ml cultures in SC- L,eu/Ura or YEPD media were grown to saturation, washed twice with water, and equal samples of cells were plated di- rectly on YEPD and YEPGal plates. Colonies were counted after growth for 4-6 days at 23" and for 7-10 days at 30" as cells grew slower on YEPGal plates at 30". After counting, the colonies were analyzed as described above.
Physical analysis of DSB repair: DNA from galactose-in- duced cells was prepared by the glass bead protocol (RUDlN and HABEK 1988), digested with the appropriate restriction enzyme(s), separated on 0.8% neutral agarose gels, and trans- ferred to Biotrans (+) nylon membranes (ICN) in 0.4 kl NaOH. Southern blot hybridization was carried out by the method of CHURCII and GILBERT (1984), using a 1.7-kb Bsu36I-Snd fragment of' the IucZ gene as a probe (FISHMAN- Lom1.1. et al. 1992). '%labeled probes for hybridization were prepared by the random-primer protocol (FEINBEKC; and VO (;EiSTEIN 1984). Densitometry of Southern blot filters was carried out by using a Molecular Dynamics Storage Phos-
phorlmager. The same filters were used to expose Kodak X- OMAT films, these being used to make photographs pre- sented on Figures 3, 5, 6, and 7 .
RESULTS
Analysis of DSB repair by the SSA mechanism on a plasmid substrate: We first analyzed the ability of mu- tants belonging to the epistasis group RAD52 to repair a single, HO endonuclease-induced DSB on plasmid DNA. This DSB must be repaired before the onset of' the next round of DNA replication; otherwise, the plas- mid would be lost. Thus the level of plasmid loss/reten- tion in galactose-induced cells can be used as a measure of the efficiency of DNA repair. Data presented in Table 2 show that rad51, rad54, md55, and rud57 mutant strains displayed essentially the wild-type level of plas- mid retention. Moreover, after induction >BO% of all the cells became LacZ+, demonstrating successful com- pletion of the repair event and arguing against the pos- sibility that the high levels of plasmid retention in mu- tant cells were due to the inefficient cleavage of plasmid DNA. Additionally, in agreement with our previous re- sults (FISHMAN-L,OKFLI. et al. 1992), a pronounced plas- mid loss was seen in a md52 strain implying that HO induction was high in this particular genetic back- ground.
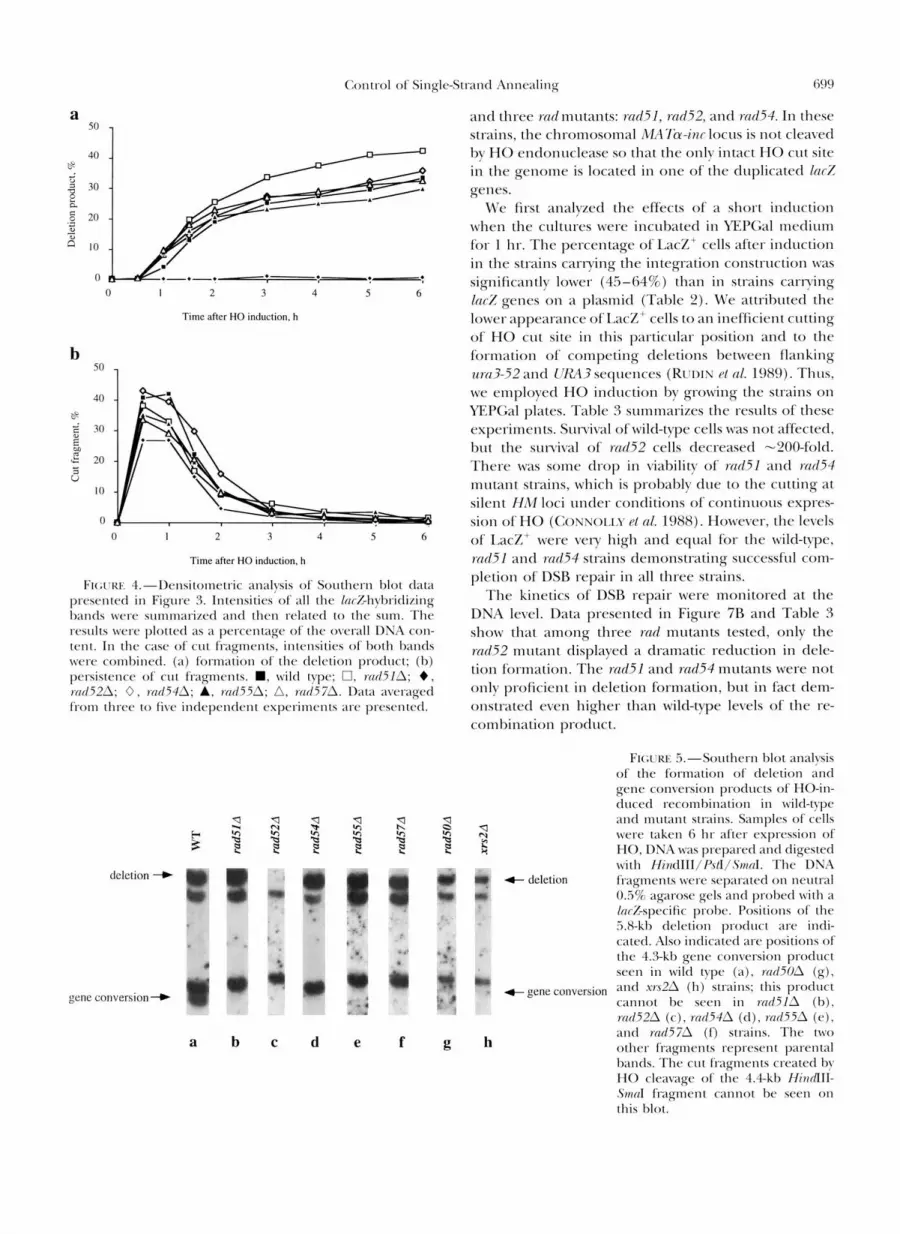
The kinetics of DSB repair of the plasmid substrate were also analyzed by Southern blot hybridization using DNA samples prepared from HO-induced cells. In a wild-type strain (Figure Sa), expression of the HO endo- nuclease led to the cleavage of plasmid DNA as evi- denced by the appearance of cut fragments. These dis- appeared rapidly as the deletion product accumulated. The formation of the deletion product was severely de- creased in a rad52strain (Figure 3c); however, deletions were formed in rad51 (Figure 3b), rad54 (Figure 3d), and rad55 and rad57 (data not shown) mutant strains. A densitometric analysis (Figure 4) of the Southern blot data of Figure 3 shows that the kinetics of appearance and the amount of the deletion product formed were quite similar in the wild-type and all the rcId strains, except rad52 (Figure 4a and Table 2 ) . In Figure 4b, we also analyzed the kinetics of formation and disappear- ance of cut fragments that reflect the kinetics of pro- cessing of the DSB ends. The levels of DSB induction and the rate of their subsequent degradation were es- sentially similar for wild-type and the rad strains. We conclude that RAD51, RAD54, KAII55, and M I 5 7 genes are not required for DSB repair by the SSA mech- anism on plasmid DNA. We note, however, that the efficiency of cutting seen in the md52 strain (530%) does not correspond quantitatively to the high level of plasmid loss (>go%). It could be that the amount of cut fragments seen in the rad52 strain is underestimated due to more extensive 5' to 3' degradation of one strand as observed previously in rad52 mutant cells (WHITE and HABEK 1990; SL~GAWARA and HXUER 1992).

Control of Single-Strand Annealing 697
TABLE 2
HO-induced recombination and plasmid loss in wild-type and mutant strains
Cells containing pJF6 after Fraction of LacZ+ Relative efficiency of Fraction of gene Strains HO induction ( % I n cells (%)” deletion formation‘ conversion ( % )
Wild type r a d l 5 A rad52A rad54A rad55A rad5 7 A
rad50A xrs 2A
84 83 4
68 70 72
26 21
93 90
0 80 83 81
65 73
1 1.26 0.02 1.07 0.79 0.96
0.39 0.45
17 0.2
ND 0.5 0.3 0.4
7.4 5.5
ND, not detectable. “Fraction of Leu+/Ura+ colonies among all the colonies grown on YEPD plates after HO induction. Before induction, these
values varied from 91% for the rad50A strain to 98% for the rad54A strains. Colonies combined from two to three independent ex eriments (n = 150 to 250) were routinely analyzed for each strain.
’Fraction of blue colonies on X-Gal plates among Leu+/Ura+ colonies. ‘ Densitometric analysis of Southern blot data shown in Figure 3. Intensities of all laczhybridizing bands seen at the 6 hr time
point were integrated. The intensity of the deletion product band was normalized to the resulting sum and expressed as a fraction of overall DNA content, the value being 33.7 2 3.3% (mean 2 SD) for the wild-type strain. The fractions of the deletion product in mutant strains were related to that of the wild-type strain. Presented are data averaged from two to five measurements for each strain. ‘ Densitometric analysis of Southern blot data shown in Figure 5. Intensities of lacZ-hybridizing bands corresponding to the
recombination product (deletions plus gene conversions) were integrated from Phosphorimager analysis and the percentage representing gene conversion events was calculated. Bands with as little as 0.5% of total DNA can be detected by this approach.
It is also possible that in the rad52 strain, expression of HO is not completely shut off after transferring the cells into glucose-containing medium resulting in additional cutting of plasmid DNA. In any case, successful recom- bination to produce Lac+ recombinants was nearly com- pletely blocked in rad52 mutant strains.
In plasmid pJF6, only -80% of HO-induced DSBs are repaired by SSA with the rest being repaired by gene conversion (FISHMAN-LOBELL et al. 1992). We de- cided to test whether the distribution of deletion us. gene conversion events is different in the rad strains. By using a triple HindIII/PstI/SmaI restriction digest of DNA prepared from galactose-induced cells, it is possi- ble to follow the formation of both recombination prod- ucts simultaneously. The results are presented in Figure 5. A densitometric analysis of these data showed that in the wild-type strain (Figure 5a) the whole recombina- tion product (deletion plus gene conversion) repre- sented almost 60% of the total ZacZhybridizing signal, ofwhich 83% were deletion and 17% were gene conver- sion events (Table 2). As expected, no visible recombi- nation products were seen in the rad52 strain (Figure 5c). In the rad51, rad54, rad55, and rad57strains (Figure 5, b and d-f, respectively), although the total recombina- tion product was formed at even higher than wild-type level (60-70% of the total lacZ-hybridizing signal), al- most no gene conversion product was formed. More precisely, according to densitometric analysis, the effi- ciency of gene conversion was 30-80 times lower in rad51, rad54, rad55, and rad57 strains compared with the wild type (Table 2).
In a parallel series of experiments, we analyzed the structure of recombinant plasmids from individual LacZ+ colonies grown on YEPD plates after induction. Each colony contained a single LacZhybridizing band, either the 7-kb band corresponding to the deletion product or the - 11-kb band corresponding to the gene conversion event (see Figures 2 and 3). The results of this analysis were in general agreement with the time- course experiments. Among 38 LacZ+ clones derived from the wild-type strain, 31 (82%) were deletions and seven (18%) were gene convertants. For 39 lacZ+ clones derived from the rad57 mutant, these figures were 38 (97%) and one (3%), respectively. All 20 LacZ+ clones analyzed for the rad51 mutant were deletions. Thus the proportion of gene convertants was greatly affected by rad51, rad54, rad55, and rad57 mutations within direct repeats of lacZ genes.
This result was rather unexpected because previously we showed that rad51, rad54, rad55, rad57 mutants are proficient in HO-induced gene conversion between two MAT genes held on a plasmid (SUGAWARA et al. 1995). To test whether the absence of gene conversion in these rad mutants was an intrinsic property of lad-containing plasmids, we used plasmid pJF5 in which the two lacZ genes are in inverted rather than in direct configura- tion. As shown in Figure 2b, an HO-induced DSB is repaired in this system exclusively by gene conversion events half of which are expected to be accompanied by crossing-over (RUDIN et al. 1989). The gene conversion events accompanied by crossing-over can be detected by Southern blot analysis of DNA prepared from galactose-

I. ."I .I -. . "- ., I - . . -. -. - -
F l c ; t ' w J."Southcrn 1 ) l o t ;In;ll!sis of the kinetics o f dclcrion formation i n Ivilcl-type and n I I t t ; 1 1 1 1 stlxins. Expression o l ' thc MO cndonuclcasc \ \ x initiated hy t h r addition of ga1;~tosr a t t i n l r 0 . A t the time points intlicatctl. s;~~nplcs ol'cclls ~ v c r c t;tkct~, ;Ind DNA \vas p r q m r c d and tligcstctl w i t h A / I . The DNA li-agnlcnts \vert srparatetl o n ncrltl-;d 0.8% ;~garosc gels ;Ind p r o l x d w i t h ;I /w%-spc*cilic prol,c ;IS tlcscrihrtl in JI:YII;.KL\I.S .\SI) \ I~:IIIOI)S . Positions o f ' thc 1l.l-kl1 duplication I'I-;Ignwnt (dl), 8 . 7 and 2.441) CIII 1'I;Ignwnts (el], and the i.0-kh tlclction protlIlct (tlcl) arc intlicatrtl. Thc other prod~tct o f ~~cconIl~in;ltio~l. ;I gc~lc convcl-sion cvent. c;Innot he srcn on these Sorlthcrn I,lots tlrlc t o ;I small tliffcrcnrc i n sizes o l ' the gene convcv.sion and the p;Ircnral IxInds. )\ DX)\ I x I n d a t the position of 3.4 kt) seen on sonic ol' t h r blots is probably due. t o nonspccilic hyhidimtion I~cr;~usr i t is s(*c11 i n nonintlucctl cc*lls. (a) \vi l t l typv; (1)) r d 5 1 A ; ( e ) rcrd52A; ((1) t d54A; ( e ) r d 5 0 A : (I) st:t2A.
induced colls containing pJF.5. The results of'this analy- sis are shown in Figure 6. I n the wild-typc strain (Figure ha), l m d s chanlctcristic o f ' the crossover product started t o appear a s early a s 30-60 m i n after induction o f a DSB. I n m / 5 1 (Figure (ib). r d 5 - l (Figure 6tl) and r(rd55and rod57 (data n o t shown) strains, the formation of recombination product was indisting~~ishal,Ic f i - o m that o f ' wild type both i n kinetics and i n thc fil1iil
amount. RccomI,in;ltion was still alxrnt in the t d 5 2 strain (Figure 6c).
All ofthc ahovc-lnentioIlcd experiments were carried out a t 30". Howc~cr, i t \vas sho\vn that i n t d 5 5 and ,nd57 mutants defects i n DNA repair and reconllina- tion are exaggerated by inculx~tion at a lower tclnpcx1- ture ( L o \ w - r and M ~ K I I \ I I : . I < 1987; H;\\:c; r / c d . 199.5; R,YI-IXAY and SYSIISC~I~OS 1995). A s the salnc r d 5 5 iI11tl
md57 mutant strains show complete tleficicnc\f i n MAT switching when tested at 30" (SL.(~~\\V;\R.\ r / n / . 1993), we f e l t i t unlikely t h a t the proficicncy of the r d 5 5 ;Ind r d 5 7 mutants i n /ur% recombination was c l w t o the higher tcmpcl-attlrc. Howcvcr, to r d e o u t this possilil- ity completcly, w e cat-rietl o u t HO induction on plates containing galactosc a t 23" or 30" iltltl then plasmid loss
was estirnatctl. N o incl-casc i n loss o f either pJF6 or pJF3 a t 2:3" \vas s w n i n t h r r d 5 5 and m d 5 i mutants, tior i n thc wild-type, t d 5 1 or r d 5 4 mutants (data n o t s h o w ) .
I n summaly, 1LW51, 1bW54, 1&\1)55, m d I M D 5 i genes arc n o t required for DSB 1-epair by t h e SSA mech- anism when duplicated /rrr%gencs arc i n direct orienta- tion. The very same genes arc not rc!quiretl Ihr HO- intltlcetl gene conveI-sion bctwrcn / r r% sequences in inverted orientation. However, i n the corresponding mutants, gene conversion is sevcrc.ly I-educed whcn the recombining /ur% sequences are i n direct orientation and SSA is ;I competing proccw.
Analysis of DSB repair by the SSA mechanism on a chromosome: Prcxviously, we have shown that rud51. r d 5 4 , r d 5 5 , and r d 5 7 mutants :~re able t o p e r f o m AIA'I's\vitching 0 1 1 a plasmid, btlt not on the chromo- some (SL,(;.\\\,.\K\ P/ {//. 1995). Thus we tlecitlrd t o test whether these genes arc reqrlircd for DSB repair by the SSA mechanism when rcwmhining sequences consti- tutc ;I part of a chromosome. A dil-ect tlllplication of /rr%genes used hefore was integrated into chromosome l f a s tlescribetl i n \IATIN..\I.S A N I ) \~~rlIo[)s . Repair o f ' HO-induced DSBs was analyzrd for ;I wild-type strain
a 1
0 I 2 3 4 5 6
Time after HO induction, h
b 1
40
E-u E 30
2 20
r; 10
0
genc conversion- U a
4 P e
8
b c d e
and three mdmutants: md51, r(m52, and rc1d54. In these strains, the chromosomal MA'l'cu-int locus is not cleaved by HO endonuclease s o that the only intact HO cut site in the genome is located in one of the duplicated lor% genes.
We first analyzed the efrects of a short induction when the cdtures were incubated in YEPGal medium for 1 hr. The percentage of LacZ' cells after induction in the strains carrying the integration construction was significantly lower (45-64%) than i n strains carrying Irc% genes on a plasmid (Table 2). We attributed the lower appearance o f L a d ' cells to an inefficient cutting o f HO cut site in this particular position and to the formation of competing deletions between flanking zrm3-52 and LU?A?sequences ( R L ~ I S Pt nl. 1989). Thus, we employed HO induction by growing the strains on YEPGal plates. Table 3 summarizes the results of these experiments. Sunrival of wild-type cells was not affected, but the sunival of md52 cells decreased -200-fold. There was some drop in viability o f md51 and rc1d54 mutant strains, which is probably due t o the cutting at silent H M loci under conditions of continuous expres- sion of HO ( C O N N ~ I . I S Pt crl. 1988). However, the levels of LacZ' were very high and equal for the wild-type, rod51 and rr(d54 strains demonstrating successful com- pletion of DSR repair in all three strains.
The kinetics of DSR repair were monitored at the DNA level. Data presented in Figure 7B and Table 3 show that among three rod mutanb tested, only the md52 mutant displayed a dramatic reduction in dele- tion formation. The md51 and rod54 mutants were not only proficient in deletion formation, but in fact dem- onstrated even higher than wild-type levels of the re- combination product.
b + deletion - k
+gene conversion
h
F I G ~ I R E 5.-Sortthern blot analysis of thc formation of deletion and gene conversion products of HO-in- tluced recombin;ltion in wild-type and mutant strains. Samples of cclls werc taken 6 hr after expression of HO. DNA was prepared and digested with H ~ ~ ~ ~ l I l l / ~ ' ~ ~ l / ~ ~ ~ ~ ~ a I . Thc DNA fragments were separated o n neutral 0.3% agarose gels and probed with a lac%-sprcific probc. Positions o f the :i.X-kb deletion product arc indi- cated. Also indicated are positions o f the 4.3-kb genc conversion product seen in wild type (a), mr15OA (g), and xrs2A (11) strains; this product cannot be scen i n md51A (b) , rnd52A ( c ) , r ~ d 5 4 A (d), rd55A ( e ) , and r(rd57A ( f ) strains. The two other fragments represent parental hands. The cut fragments created by
S n d fragment cannot he scen on this blot.
HO cle>1\.21ge of the 4.4-kh Hi?~dlIl-

700 E. L,. Ivanov P/ n/.
Hnurs Hours
""
00.51 1.5 2 3 4 5 6 " " .
00.5 1 1 . 5 2 3 4 5 6 - Pf d"-- e f c o I
L
v .- cf
a. wild type
Hours
~ 4-co + co
b. rad5lA Hours
0 0.5 1 1.5 2 3 4 5 6
0 0.5 I 1.5 2 3 4 5 6
1 ~- - ' g-co
I I p(' co
cf
cf
c. rad52A d. rad54A
M'e then wanted to know whether the distribution of recombination products (deletions vs. gene conver- sion) resembled that for plasmid substrate. Analysis of thc structure of individual LacZ' colonies showed that 37 of 37 clones derived from md51 strain and 40 of 40 clones drrivrd from mI54 strain were deletions; no genc conwrtants were found among these two sets of I x % + colonies. Among 40 LncZ' clones derived from the wild-typc strain, five (13%) were gene convertants and 33 (87%) were deletions, which is close to the distri-
TABLE 3
HO-induced recombination within direct lacZ duplication integrated into chromosome V
Pf
cf
cf
4- co 4- co Pf
Pf
cf
cf
FIGI'RF. 6."Southern blot analysis of the ki- netics of HO-induced recombination in wild- type and mutant strains using plasmid pJF.5 as ;I
substrate. The timecourse experiment w a s car- ried out and the DNA was prepared and analyzed as described in the legend t o Figure 3. Positions of 7.4- and 6.0-kb parental fragments (pl), 4.4- and 3.0-kb cut fragment., ( c Q , and i .O- and 6.5- kb crossing-over products (co) arc shown. The i.Rkb hand corresponding to gene conversion without crossing over cannot be resolved on these Southern blots. A DNA ktnd at the posi- tion of -3.4 kb is probably due to nonspecific hybridization. (a) wild type; (h) rrrd5lA; (c) md52A; ( d ) md54A.
bution seen for the plasmid substrate. We conclnde from these data that as for the plasmid pJF6 recombina- tion substrate, RAD51 and RAD54 genes are not re- quired to carry out SSA on a chromosome, but in the corresponding mutants, HO-induced gene conversion behveen directly repeated k c % genes is severely re- pressed or being prevailed by SSA.
Mutations in RAD50 and XRS2 genes decrease the efficiency of SSA on a plasmid: Although RAD50 and XRS2 genes belong to the IWI52 epistasis group, these two genes have some properties making them very dif- ferent from other members of this group [for detailed discussion of this point, see IVANO\' et nl. (1994)]. Al- though md50A and m 2 A deletion alleles do not pre- vent the completion of MATswitching, the kinetics of this process in mutant cells are markedly retarded. Mo- lecular analysis of this phenomenon demonstrated that the slower kinetics of MAT switching in md50A and xrs2A mutants is at least partly due to the decreased 5' to 3' degradation of the ends of HO-induced DSBs (SLIGIWARA and HARER 1992; IVANOY P/ (11. 1994). As the SSA process is critically dependent on 5' to 3' degra- dation of DSB ends (Figure 1 b), we analyzed the effects of rnd50A and m.s2A mutations on the efficiency and kinetics of DSB repair in the pJF6 plasmid system.
Data in Table 2 show that HO-induced plasmid loss was significantly increased in rad50 and xn2 mutants, implying that repair of HO-induced DSBs was affected. However, the fraction of L a d ' cells after induction is not significantly lower in the mutant strains compared

Control of Single-Strand Annealing 70 1
A
B
ura3-52 HOcs
LEU2 uRA3 I I ”
LACZ LACZ P P
ura3-52 LEU2 uRA3 P . I -
LA CZ P P
ura3-52 LEU2 uRA3 F.- ”
LACZ LACZ P P
Hours 0 1 6
Hours Hours 0 1 6 0 1 6
C-u df @ del cf - del
7 :: y7”-
df cf del
Deletion
Gene conversion
Hours 0 1 6 nrr?lnm:
~r*y df cf
.=“ del I
FIGURE 7.-Repair of an HO-induced DSB within the direct duplication of larZ genes integrated into the ura3-52 locus on chromosome V. (A) schematic representation of the structures of integration and its recomhinational derivatives. The positions of two relevant Pstl sites (P) are indicated. The sizes of pa- rental and HO-induced AtI-fragments hybridizing to the 1arZ prohe nsed are the same as for the plas- mid pJFG suhstrate (Figure 2a). (B) Southern blot analysis of deletion formation in wild-type and mu- tant strains. The timecourse experiment was carried out and the DNA was prepared and analyzed as de- scribed in the legend to Figure 3, except that only three time points (0, 1 and G hr) were analyzed. Positions of the 11.1-kh duplication fragment (do, 8.7- and 2.4-kh cut fragment5 (cf), and the 7.0-kh deletion product (del) are indicated. The other product of recombination, a gene conversion event, cannot he seen on these Southern blots (a) wild type; (h) md5ZA; (c) md52A; (d) rud54A.
cf cf
cf cf
a. Wild type b. radSlA c. radS2A d. mdS4A
with the wild-type and proficient rad mutants indicating successful completion of recombination events. One ex- planation for this apparent discrepancy could be that rad50 and xr.32 mutants do not prevent but markedly retard recombinational repair of HO-induced DSBs be- tween direct repeats of l a d genes, as was shown to be the case for MATswitching (IVANOV d al. 1994). If so, a significant fraction of DSBcontaining plasmids would remain unrepaired at the next round of DNA replica- tion leading to increased plasmid loss. At the same time among recovered plasmids, completion of recombina- tion should have the high proportion of LacZf cells seen in wild-tvpe cells.
Analysis of the repair events at the DNA level supports the obsewation that the efficiency of DSB repair within the 1acZ duplication was substantially reduced in rad50 (Figure Se) and m32 (Figure Sf) mutants. By 6 hr after induction, only 40-45% of the wild-type level of dele- tion production was seen in the mutant strains (Figure 8a). Characteristically, the amount of persistent cut fragment was dramatically increased in mutant cells (Figure 8b). Disappearance of the cut fragments re- flects the degradation of DSB ends. Thus accumulation of this intermediate in mutant cells support5 the idea
that the extent of 5’ to 3’ degradation of DSBs is de- creased in rad50 and ms2 cells.
We then wanted to know whether both deletion and gene conversion formation were affected to the same extent by rad50 and ms2 mutations. Data presented in Figure 5, g and h, respectively, and in Table 2 show that the efficiency of gene conversion was decreased two- to threefold in rad50 and ms2 strains compared with wild type, according to a densitometric analysis of Southern blot data. To exclude the possibility that there was a bias in survival of LacZ’ colonies bearing a dele- tion or a gene conversion plasmid derivative, we ana- lyzed individual LacZf clones derived from the us2 strain. For 20 clones analyzed, two (10%) were gene convertants and 18 (90%) were deletions, which is close to the results derived from timecourse experiments. We conclude that rad5OA and xrs2A deletion mutations decreased HO-induced recombination in directly re- peated lac% genes, both deletion and gene conversion events being affected approximately to the same extent.
DISCUSSION
Genes belonging to the epistasis group RAD52 share many common effects on DNA repair and recombina-

702 E. L. Ivanov rt nl.
a 40 1
0 1 2 3 4 5 6
Time after HO induction, h
b 60
10
T" 0 1 2 3 4 5 6
Time after HO induction, h
FIGURE 8.-Densitometric analysis of Southern blot data on the formation of deletions in rad50A (Figure 3e) and xrs2A (Figure 30 strains. The analysis was carried out as de- scribed in the legend to Figure 4. The data for the wild-type strain are the same as in Figure 4. ., wild type; 0, rad50A; + , xrs2A. The data for the rnd50A and xrs2A strains are from four independent experiments for each strain.
tion [for review, see PETES et al. (1991)l. Moreover, genetic and biochemical data suggest that the corre- sponding proteins interact and even form a multipro- tein complex (SHINOHARA et al. 1992; MILNE and WEAVER 1993; HAYS et al. 1995). However, in last few years, it has become evident that different genes play different roles in DNA metabolism. For example, null mutations in RAD52 cause more severe defects in spon- taneous mitotic recombination than do null mutations in RALl51, RAD54, RAD55, and RAD57 (RATTRAY and SYMINCTON 1994, 1995). In the case of HO-induced DSBs, the situation is even more complicated. Muta- tions in each of these five genes completely block, at some early step, MATswitching on a chromosome, how- ever, only RAD52 is required for this gene conversion event when the donor sequence is simultaneously not silenced and located on a plasmid (SU(;AWARA et al. 1995). In this communication, we extend this finding to show that only the RAD52 gene is required for gene conversion on a plasmid containing an inverted repeat of lacZ genes whereas the RAD51, RAD54, RAD55 and RALl57genes are not (Figure 6) . An attractive interpre- tation of these data is that only the RAD52 gene is re- quired for recombination per se, whereas the other RAD genes are only needed to gain access to otherwise inac- cessible regions of chromatin (SUCAWARA et nl. 1995). Despite the finding that the Rad51 protein has a strand- exchange activity (SUNG 1994), our data suggest that Rad51 may play a different role in vivo. One would
expect more general defects in recombination in rad51 mutants if this strand-exchange activity were the major cellular one.
The main result of this work is that KAL)51, RALl54, W 5 5 , and RAD57genes are also not required for HO- induced DSB repair by the SSA mechanism. SSA, either on a plasmid substrate (Figures 3 and 4 and Table 2) or on a chromosome (Figure 7 and Table 3 ) , is not affected by null mutations in these genes. It is important to note that SSA is greatly repressed, both on a plasmid and on a chromosome, by a rad52 mutation demonstra- ting again the difference between RAD52 and other genes of the RAD52 epistasis group with respect to HO- induced recombination.
The absence of requirements for RAD51, W 5 4 , W 5 5 , and RAD57genes in the SSA pathway is compli- cated, however, by the negative effects of corresponding mutations on the gene conversion process in the same direct-duplication system. For the chromosomal sub- strate these effects could be explained by the require- ments for these KAL, genes to promote recombination in the context of chromatin (SLJGAWARA et nl. 1995). At the same time, it is difficult to understand why gene conversion is repressed on plasmid DNA (Figure 5 and Table 2) given the essentially normal levels of gene conversion between lac% sequences in inverted orienta- tion. We explain this apparent contradiction in the fol- lowing way. As was shown before (FISHMAN-LOBELL d
al. 1992), in the pJF6 plasmid system, deletion and gene conversion events represent two alternative, competing and independently modulated outcomes of repair of an HO-induced DSB. Factors decreasing the efficiency of deletion formation (for example, increasing the dis- tance between two recombining sequences) increase the fraction of gene conversion; conversely, factors pre- venting gene conversion increase the proportion of de- letions (FISHMAN-LOBELL et al. 1992). We propose that in rad51, rad54, rad55, and rad57mutant cells, the gene conversion process on a plasmid has a modest defect. This defect cannot be seen when the recombining se- quences are in inverted orientation (as in the case of plasmid pJF5) because gene conversion is the only way to repair an HO-induced DSB. However, in the direct repeat configuration, this defect is apparent, due to the presence of an alternative, highly efficient SSA pathway that competes with gene conversion. In agreement with this view are results obtained with plasmid pNR43 that contains a 4.4kb insert between the direct repeats lead- ing to a higher proportion of gene conversion in wild- type cells (FISHMAN-LOBELL. el al. 1992). In a rad51 mu- tant cells, this also leads to the appearance of some gene conversion product, albeit with slower kinetics and at a lower level compared with wild type ( J . FISHMAN- LOBELL, unpublished results).
Alternatively, one can imagine that gene conversion in ,rad51, rad54, md55, and md57cells is unaffected but instead the efficiency of the SSA process is increased

Control of Single-Strand Annealing 703
due, for example, to higher levels of single-stranded DNA formation as seen in the case of MAT switching (SUGAWARA et al. 1995). This idea, however, does not find support when analyzing the kinetics of DSB repair on plasmid DNA. Indeed, if in these rad cells DSB ends were degraded and processed into the next recombina- tional intermediate faster than in wild-type strains, this would cause an apparent decrease in the amount of cut fragments on Southern blots (Figure 3 ) . However, a densitometric analysis of the Southern blot data (Figure 4) does not suggest that the stability of DSB ends is changed in these mutants.
In any case, our results show that in rad51, rad54, rad55, and rad57 mutants DSB-induced direct-repeat recombination proceeds almost exclusively via the SSA mechanism. This could serve as a basis to explain increased deletion formation seen in these mutants during spontaneous mitotic recombination involving directly repeated sequences (FAN and KLEIN 1994; MG DONALD and ROTHSTEIN 1994; AGUILERA 1995; LIEF% HITZ et al. 1995).
In this communication, we report that null mutations in RAD50 and XRS2 genes affect the efficiency of SSA leading to increased plasmid loss. Our results confirm our previous observations that rud50A and xrs2A muta- tions decrease the extent of 5’ to 3’ degradation of the ends of HO-induced DSBs (SUGAWARA and HABER 1992; IVANOV et al. 1994). The observed decrease in the effi- ciency of deletion formation, however, is only two- to threefold (Table 2). This could explain why during spontaneous mitotic direct-repeat recombination, rad50 mutants show little or no effect (MCDONALD and ROTHSTEIN 1994).
The effects of rad50 and xrs2 mutations on MAT switching suggest that the rate of 5’ to 3‘ degradation of DSB ends is a rate-limiting factor for this specialized gene conversion event (IVANOV et al. 1994). In lac2 di- rect-duplication substrates, rad50and xrs2mutations de- crease deletion and gene conversion events to approxi- mately the same extent (Figure 5 and Table 2). This argues that degradation of HO-induced DSB ends is a rate-limiting factor for gene conversion not only during MAT switching but in other recombination systems as well.
So far, only a handful of genes are known to signifi- cantly affect the SSA process. These include the RAD52 (FISHMAN-LOBELL et ul. 1992; SUGAWARA and HABER 1992), RADl, and RAD10 (FISHMAN-LOBELL and HABER 1992; IVANOV and HABER 1995) genes. Recently, a muta- tion in the CDCl gene was described that most likely affects the SSA pathway of HO-induced intrachromoso- mal recombination (HALBROOK and HOEKSTRA 1994). At the same time, many genes known to be essential for “classical” homologous recombination were shown here not to be required for SSA. This means that the genetic requirements of SSA are different from those of gene conversion. Thus additional genes controlling
SSA remain to be discovered. To this point, we have recently undertaken a genetic screen to isolate mutants defective in SSA. Among the mutations isolated, some are neither UV or MMS sensitive, making them differ- ent from mutations in the above-mentioned genes (S. SATO, A. MALKOVA and J. E. WER, unpublished re- sults). We hope that characterization of these mutations will allow us to identify new genes responsible for SSA in yeast and to use them as probes to detect their human counterparts.
We are grateful to A. MALKOVA for continuous discussion of this work. This work was supported by National Institutes of Health Grant GM-20056 to J.E.H.
LITERATURE CITED
ABOUSSEKHRA,A., R. CHANET,A.ADJIRI and F. FABRE, 1992 Semidomi- nant suppressors of Srs2 helicase mutations of Saccharomyces cere- visiaemap in the RAD51 gene, whose sequence predicts a protein with similarities to procaryotic RecA proteins. Mol. Cell. Biol.
AGUILERA, A,, 1995 Genetic evidence for different RAD52depen- dent intrachromosomal recombination pathways in Saccharomyces cerevisiae. Curr. Genet. 27: 298-305.
ALANI, E., S. SUBBIAH and N. KLECKNER, 1989 The yeast RAD50 gene encodes a predicted 153kd protein containing a purine nucleotide binding domain and two large heptad-repeat regions. Genetics 122: 47-57.
CAo, L., E. AIANI and N. KLECXNER, 1990 A pathway for generation and processing of double-strand breaks during meiotic recombi- nation in S. cerevisiae. Cell 61: 1089-1101.
CHURCH, G. M. and W. GILBERT, 1984 Genomic sequencing. Proc. Natl. Acad. Sci. USA 81: 1991-1995.
CONNOLLY, B., C. I . WHITE and J. E. HABER, 1988 Physical monitor-
Biol. 8: 2342-2349. ing of mating type switching in Saccharomyces cereuisiae. Mol. Cell.
FAN, H., and H. L. KLEIN, 1994 Characterization of mutations that suppress the temperature-sensitive growth of the h p 1 D mutant of Saccharomyces cermisiae. Genetics 137: 945-956.
FEINBERG, A. P., and B. VOGELSTEIN, 1984 A technique for radiola- beling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 137: 266-267.
FIRMENICH, A. A,, M. ELW%ARNANZ and P. BERG, 1995 A novel allele of Saccharomyces cerevisiae REA1 that is deficient in recombination and repair and suppressible by RAD52 Mol. Cell. Biol. 1 5 1620- 1631.
FISHMAN-LOBEIL, J., and J. E. HABER, 1992 Removal of nonhomolo- gous DNA ends in double-strand break recombination: the role of the yeast ultraviolet repair gene R A D I . Science 258: 480-484.
FISHMAN-LOBEIL, J., N. RUDIN andJ. E. HABER, 1992 Two alternative pathways of double-strand break repair that are kinetically sepa- rable and independently modulated. Mol. Cell. Biol. 12: 1292- 1303.
HALBROOK, J., and M. F. HOEKSTRA, 1994 Mutations in the Saccharo- myces cereuisiae CDCl gene affect double-strand-break-induced in- trachromosomal recombination. Mol. Cell. Biol. 14: 8037-8050.
HAYS, S. L., A. FIRMENICH and P. BERG, 1995 Complex formation in yeast double-strand break repair: participation of Rad51, Rad52, Rad55, and Rad57 proteins. Proc. Natl. Acad. Sci. USA 92: 6925- 6929.
ITO, H.,Y. FUKADA, K. MURATA and A. KIMURA, 1983 Transformation of intact yeast cells treated with alkali cations. J. Bacteriol. 153: 163-168.
IVANOV, E. L., and J. E. HABER, 1995 RADl and RADIO, but not other excision repair genes, are required for double-strand break-induced recombination in Saccharomyces cereuisiae. Mol. Cell. Biol. 15: 2245-2251.
IVANOV, E. L., N. SUGAWARA, C. I . WHITE, F. FABRE and J. E. HABER, 1994 Mutations in XRSL, and RAD50 delay but do not prevent mating-type switching in Saccharomyces cmeuisiae. Mol. Cell. Biol. 1 4 3414-3425.
12: 3224-3234.

704 E. L. Ivanov et al.
KLEIN, H. L., 1995 Genetic control of intrachromosomal recombina- tion. BioEssays 17: 147-159.
LIEFSHITZ, B., A. P..UU(ET, R. MAYA and M. KUPIEC, 1995 The role of DNA repair genes in recombination between repeated sequences in yeast. Genetics 140: 1199-1211.
LIN, F.-L., K. SPERLE and N. STERNBERG, 1984 Model for homolo- gous recombination during transfer of DNA into mouse L cells: role for DNA ends in the recombination process. Mol. Cell. Biol. 4: 1020-1034.
LIN, F.-L., K. SPERI.E and N. STERNBERG, 1990 Intramolecular recom- bination between DNAs introduced into mouse L cells by a non- concervative pathway that leads to crossover products. Mol. Cell. Biol. 10: 103-112.
Lovt-rr, S. T., and R. K. MORTIMER, 1987 Characterization of null mutants of the RAD55 gene of Saccharomyces cereuisim effects of temperature, osmotic strength and mating type. Genetics 116: 547-553.
MCDONALD, J. P., and R. ROTHSTEIN, 1994 Unrepaired heterc- duplex DNA in Saccharomyces cereuisiae is decreased in R A D 1 RAD52-independent recombination. Genetics 137: 393-405.
MII.NE, G. T., and D. T. WEAVER, 1993 Dominant negative alleles of RAD52 reveal a DNA repair/recombination complex including Rad51 and Rad52. Genes Dev. 7: 1755-1765.
OZENBERGER, B., and G. S. ROEDER, 1991 A unique pathway of dou- ble-strand break repair operates in tandemly repeated genes. Mol. Cell. Biol. 11: 1222-1231.
PETES, T. D., R. E. MAI.ONE and L. S. SYMINGTON, 1991 Recombina- tion in yeast, pp. 407-521 in The Molecular and Cellular Biology of the Yeast Saccharomyres, edited by J. BROACH, J. PRINGI.E and E.
JONES. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. RhTrwx, A. J., and L. S. SYMINGTON, 1994 Use of a chromosomal
inverted repeat to demonstrate that the RAD51 and RAD52genes of Saccharomyres cermisiae have different roles in mitotic recombi- nation. Genetics 138: 587-595.
RATTRAY, A. J., and L. S. SYMINCXON, 1995 Multiple pathways for homologous recombination in Saccharomyces cereuisiae. Genetics 139: 45-56.
ROTHSTEIN, R., 1983 One step gene disruption in yeast. Methods Enzymol. 101: 202-211.
RUBY, S. W., J. W. SXOSTAK and A. W. MURRAY, 1983 Cloning regu- lated yeast genes from a pool of ZacZ fusions. Methods Enzymol. 101: 253-269.
KLTIIN, N., and J . E. HABER, 1988 Efficient repair of HO-induced chromosomal breaks in Saccharomyces cereuisiae by recombination
between flanking homologous sequences. Mol. Cell. Biol. 8:
RUDIN, N., E. SUGARMAN and J. E. WER, 1989 Genetic and physical analysis of double-strand break repair and recombination in Sac- charomyces cereuisiae. Genetics 122: 519-534.
SANDELL, L. L., and V. A. ZAIUAN, 1993 Loss of a yeast telomere: arrest, recovery, and chromosome loss. Cell 75: 729-739.
SCHIESTL, R. H., and S. PRAKASH, 1988 R A D I , an excision repair gene of Saccharomyces cereuisiae, is also involved in recombination. Mol. Cell. Biol. 8: 3619-3626.
SCHIESTL, R. H., and S. PRAKASH, 1990 RADIO, an excision repair gene of Saccharomyces cereuisiae, is involved in the RAD1 pathway of mitotic recombination. Mol. Cell. Biol. 10: 2485-2491.
SCHII.D, D., I. L. CALDERON, C. R. CONTOPOUI.OU and R. K MORTI- MER, 1983 Cloning of yeast recombination repair genes and evidence that several are nonessential genes, pp. 417-427 in Cellular Responces to DNA Damage, edited by E. C. FRIEDBERG and B. A. BRIDGES. Alan R. Liss, New York.
SHERMAN, F., G. R. FINK^^^ J. B. HICKS, 1986 Methods zn Yeast &et- ics: Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
SHINOHARA, A,, H. OGAWA and T. OGAWA, 1992 Rad51 protein in- volved in repair and recombination in S. cereuisiae is a RecA-like protein. Cell 6 9 457-470.
SUGAWARA, N., and J. E. HABER, 1992 Characterization of double- strand break induced recombination: homology requirements and single-stranded DNA formation. Mol. Cell. Biol. 12: 563- 575.
SUGAWARA, N., E. L. IVANOV, J. FISHMAN-LOBELL, B. L. RAY, X. Wu et al., 1995 DNA structuredependent requirements for yeast RAD genes in gene conversion. Nature 373: 84-86.
SUN, H., D. TRECO and J. W. SZOSTAK, 1991 Extensive 3"overhang- ing, single-stranded DNA associated with meiosis-specific double- strand breaks at the ARM recombination initiation site. Cell 6 4 1155-1161.
SUNG, P., 1994 Catalysis of ATP-dependent homologous DNA pair- ing and strand exchange by yeast RAD51 protein. Science 265: 1241-1243.
SZOSTAK, J. W., T. L. ORR-WEAVER, R. J. ROTHSTEIN and F. W. STAHI., 1983 The double-strand-break repair model for recombination. Cell 33: 25-35.
WHITE, C. I., and J. E. HABER, 1990 Intermediates of recombination
J. 9: 663-673. during mating type switching in Saccharomyces cereuisiae. EMBO
3918-3928.
Communicating editor: S. JINK~ROBERTSON