generation of recombinants strains to cellulases production by protoplast fusion between penicillium...
TRANSCRIPT

Gb
AMTa
b
a
ARRA
KPFTPC
1
g�yCdalnro
0d
Enzyme and Microbial Technology 43 (2008) 403–409
Contents lists available at ScienceDirect
Enzyme and Microbial Technology
journa l homepage: www.e lsev ier .com/ locate /emt
eneration of recombinants strains to cellulases production by protoplast fusionetween Penicillium echinulatum and Trichoderma harzianum
ldo José Pinheiro Dillona,∗, Marli Camassolaa, João Antônio Pegas Henriquesa,aria Helena Pelegrinelli Fungarob, Andréia Cristiane Souza Azevedob,
arciso André Ferreira Velhoa, Sergio Echeverrigaray Lagunaa
Instituto de Biotecnologia, Universidade de Caxias do Sul, CP 1130, CEP95100-700 Caxias do Sul, RS, BrazilUniversidade Estadual de Londrina, CP 6001, CEP86051-990 Londrina, PR, Brazil
r t i c l e i n f o
rticle history:eceived 18 February 2008eceived in revised form 23 July 2008ccepted 29 July 2008
eywords:rotoplastusionrichodermaenicillium echinulatumellulases
a b s t r a c t
Nowadays, cellulase importance is growing mainly due to its potential utilization in developing second-generation ethanol technology. Thus, considerable effort has been made to obtain new genotypes withimproved capacity to produce efficient and cost-effective enzyme complexes for cellulose hydrolysis. Themajority of the genetically manipulated strains that produce higher amounts of cellulases come from ofthe wild Trichoderma reesei QM6A. Such as T. reesei cellulolytic mutants, Penicillium echinulatum mutantsalso produce high titles of cellulases (filter paper activities—FPA), and its enzymatic complex effectivelyhydrolyses cellulose. In the present work, we obtained genetic variability with the purpose of enhancingcellulase production by protoplast fusion between P. echinulatum and T. harzianum. The resulting fusants,who were all morphologically like the P. echinulatum parental, were initially characterized by RAPD, andthe results showed that almost all amplified DNA bands showed correlation with P. echinulatum DNAbands. Some of new selected strains presented higher FPA and �-glucosidases activities when comparedto the parental strains. One particular clone – BP2 – showed values of FPA activities around of 2 UI ml−1, insubmerged cultures and all selected strains showed faster and higher secretion of cellulases in solid-statecultures, in comparison to the parental strain of P. echinulatum. Altogether, at the same time that our
results show increased cellulase secretion by P. echinulatum strains, they also show that protoplast fusiontechniques associated with an efficient selection process could be a useful tool in genetic improvementto generate better cellulose-producing strains aimed at economically hydrolyzing cellulose with a viewnol.
gc
tinv
to second-generation etha
. Introduction
Cellulases are enzymatic complex, that comprises exo-�-1,4-lucanases (EC 3.2.1.91), endo-�-1,4-glucanases (EC 3.2.1.4) and-1,4-glucanases (EC 3.2.1.21), that act synergistically in the hydrol-sis of �-1,4-glycosidic bonds present in cellulose polymers.ommercially, these enzymes are mainly used by the textile andetergent industries [1,2]. However, their most significant potential
pplication, which would require high quantities of these enzymes,ies in cellulose saccharification to produce ethanol [3–5]. Sig-ificant efforts have been made to understand the mechanismsegulating the expression of genes encoding cellulases [5–9], tobtain genetic variants in improvement programs involving muta-∗ Corresponding author. Tel.: +55 54 3218 2100; fax: +55 54 3218 2100.E-mail address: [email protected] (A.J.P. Dillon).
emelaivsfr
141-0229/$ – see front matter © 2008 Elsevier Inc. All rights reserved.oi:10.1016/j.enzmictec.2008.07.009
© 2008 Elsevier Inc. All rights reserved.
enesis and selection [10–18], and to obtain genetic transformantsarrying specific markers of interest [1,19,20].
In addition to mutagenesis and genetic transformation, pro-oplast fusion can also be used to achieve genetic alterationsn cellulases secretion in fungi that consists of the alter-ative to inducing gene recombination and generate geneticariation [21–26].
The cellulolytic microorganisms are evolutionary “factories” ofnzymes and a lot of genes, not yet perfectly identified or known,ust operate jointly to achieve an efficient cellulase secretion,
specially in view of the composition and structure of lignocellu-osic materials. From this point of view, the known cellulase genesnd their control system are only a part of all genes, which can be
nvolved with a cellulase secretion. Therefore, all these genes canary in different species or genera of related microorganisms thatecrete cellulases; they could be recombined through protoplastusion and powerful screening techniques would choose the betterecombinant strains of interest.
4 icrobia
iw
asiwips[omceo
mbiptpnsb
gigocrbfsTmi
2
2
tc(mcUs
2
[C0(ga4lf
c(c
TGmpg
fcI(ms
1cm
E[Co1p
2
gs
2
oimwcmr
2
t(umtspSt1Csfapttt
2
aiaf
t
04 A.J.P. Dillon et al. / Enzyme and M
Protoplast fusion is also a very powerful approach for fast geneticmprovement when associated with mutation techniques, as seen
ith tylosin production by Streptomyces fradiae [27].In addition to Trichoderma reesei, Penicillium echinulatum is
mong the microorganisms that show great potential of cellulaseecretion for enzymatically converting biomass into ethanol. Fornstance, mutants of P. echinulatum are able to secrete cellulases
ith filter paper activity (FPA) higher than 2 IU ml−1 when grownn media containing 1% cellulose as carbon source [14,15]. They alsoroduce high yields of cellulases in solid-state fermentation usingugar-cane bagasse (SCB) that reach more than 30 FPU g−1 of SCB28]. Moreover, the FPA and �-glucosidases of P. echinulatum aref interest for cellulose hydrolysis because they show good ther-al stability at 50 ◦C and 55 ◦C, respectively [29], and its cellulase
omplex presents a good proportion of FPA and �-glucosidase forfficiently hydrolyzing cellulose, when compared to the cellulasesf T. reesei [30].
Nevertheless, it has been difficult to obtain new cellulaseutants of P. echinulatum 9A02, although a good methodology has
een applied [15]. Due to this, in the present work, we aimed tosolate genetic variants with improved cellulases secretion. For thisurpose, we induced genetic recombination by bringing togetherhe genomes of P. echinulatum and Trichoderma harzianum throughrotoplast fusion. The T. harzianum strain was isolated as an intesti-al symbiont of the Coleoptera Passalus binominatus erosus and alsoecretes cellulases, appearing at a faster growth than P. echinulatum,ut with a smaller production of it (data not shown).
The genetic recombination does not necessary involve cellulaseenes of both strains; it is more likely that general genes involvedn the growth, if recombined, could produce some new interestingenotypes. Therefore, later after the possible genetic segregationf the heterokaryons, we selected clones that produced faster andlearer halos in the agar plate containing swollen cellulose. Occur-ence of genetic recombinantion in fusion products was evaluatedy Random Amplification of Polymorphic DNA (RAPD), and theusants were studied for FPA and �-glucosidases secretion in bothubmerged fermentation (SF) and in solid-state fermentation (SSF).he new strains obtained showed higher cellulolytic activity in sub-erged cultures and also faster and higher secretion of cellulases
n solid-state cultivations in relation to parental strains.
. Materials and methods
.1. Fungal strains
In this work, we used cellulolytic mutant strains 2427, obtained from the wildype (2HH) of P. echinulatum after UV radiation and selection in agar plate containingellulose for observing hydrolysis haloes [14], and the cellulolytic mutants 5A-AS5AS5) and 5A36, obtained from the wild type (5A) of T. harzianum using the same
ethodologies used with P. echinulatum [31]. Both strains belong to cultures of theollection of the Laboratory of Enzymes and Biomass, Institute of Biotechnology,niversity of Caxias do Sul, Brazil, which after growth at 28 ◦C are maintained onlands of cellulose agar (CA) plate at 4 ◦C and subcultured at 6 months intervals.
.2. Media
All media used were based on 10X concentrated Mandels mineral solution (MS)32] containing (g l−1) KH2PO4, 20; (NH4)2SO4, 3; CO(NH2)2, 3; MgSO4·7H2O, 3;aCl2, 3; FeSO4·7H2O, 0.050; MnSO4·H2O, 0.0156; ZnSO4·7H2O, 0.014; CoCl2·6H2O,.0020. Swollen cellulose gel was produced by placing 5 g of Cellufloc 200 celluloseCelulose e Amido Ltda, Suzano, São Paulo, Brazil), 60 ml of distilled water and 20lass beads (�: 3 mm) in a 500-ml Erlenmeyer flask, which was then sealed andutoclaved at 121 ◦C for 30 min. These flasks were placed in an orbital shaker for8 h at 180 rpm. Later, 140 ml of distilled water was added and the swollen cellu-
ose was stored at 4 ◦C until use. Unless otherwise stated, chemicals were purchasedrom Sigma (São Paulo, SP, Brazil).
To growth and manutention of strains was used the cellulose agar (CA) [15]onsisting of 40% (v/v) of swollen cellulose suspension, 10% (v/v) 10 × MS [32], 0.1%w/v) proteose peptone (Oxoid) and 2% (w/v) agar and to observe halo formation onellulose-containing dishes we used CA medium supplemented with 0.1% (v/v) of
caUaeo
l Technology 43 (2008) 403–409
riton X-100 (TCA) to restrict colony development. When necessary was used theTCA medium, where glucose (0.5–1.5% final concentration) was added to the TCAedium to produced catabolic repression of cellulases. To obtain mycelia to obtain
rotoplast and extracted DNA was used the CLM medium constituted of 1% (w/v) oflucose, 10% (v/v) 10 × MS and 0.1% (w/v) proteose peptone.
In order to select the mutants, media were based on CA containing followingungicides: 12.5 �g ml−1 of benomyl (methyl 1-(butylcarbamoyl)-2-benzimidazole-arbamate) corresponding to the commercial product benlate from DuPont,nc. (BCA) and the medium with cycloheximide contained 50 �g ml−1 of (3-[2-3,5-dimethyl-2-oxocyclohexyl)-2-hydroxyethyl]-glutarimide) (Sigma) (CCA). The
edium with nystatin (NCA) contained 30 U ml−1 of the commercial product myco-tatin of Bristol–Myers Squibb, Inc.
Protoplasts were regenerated in liquid media (PLM) containing 0.6 M of KCl and0% (v/v) of MS [32]. Heterokaryons were selected on CA medium plus 0.6 M of KClontaining either 12.5 �g ml−1 of benomyl and 50 �g ml−1 of cycloheximide (BCCAedium), or 12.5 �g ml−1 of benomyl and 30 U ml−1of nystatin (BNCA medium).
In the submerged growth for cellulases production, fungi were grown in 500 mlrlenmeyer flasks containing 100 ml of liquid media containing 20% (v/v) MS × 1032], 0.2% (w/v) soy meal, 0.1% (v/v) Tween 80 with 1% (w/v) of glucose (LM) orellufloc 200 cellulose (CLM), respectively. The solid-state culture medium consistedf 1 g of wheat bran moistened with 1 ml of 10 × MS [32] and placed in flasks with a2 cm × 3 cm concave base. The flasks were closed with a gauze-covered cotton-woollug and autoclaved at 121 ◦C for 1 h.
.3. Maintenance of strains and inocula preparation
All strains were grown and maintained on cellulose agar (CA) [15]. Cultures wererown for up to 7 days at 28 ◦C until conidia formation, and used to inoculate theubmerged fermentation (SF) or solid-state fermentation (SSF) or stored at 4 ◦C.
.4. Obtaining genetic markers
Spores obtained from 7-day colonies grown in cellulose agar (CA) were used tobtain 10 ml suspension of 2 × 108 conidia ml−1. This conidia suspension disposedn open Petri dish was exposed to ultraviolet radiation (UV 254 nm) using a low
ercury pressure Tohawalite—G15T8 germicidal lamp, 15 cm away from the source,ith periodic agitation (∼40 s) for the amount of time necessary to kill 95% of the
ells [15]. Volumes of 0.1 ml of the treated conidia suspension were spread in the CAedium containing benomyl, cycloheximide, or nystatin to isolate clones containing
esistance to these substances.
.5. Protoplast preparation and fusion
To obtain protoplasts, we used 0.2 g of wet mycelia obtained from liquid cul-ures grown after filtration in the LM medium during 24 h (T. harzianum) or 48 hP. echinulatum), being the 0.2 g of mycelia obtained by filtration of liquid culturesing Whatman no. 1 Filter Paper and washed one time with distilled water. Theycelia was immediately treated for 3–4 h at 35 ◦C with 5 ml of Novozym 234® solu-
ion (5 mg/5 ml of KCl 0.6 M in a citrate buffer 0.05 M, pH 4.8) or 5 ml of Glucanex®
olution (50 mg/5 ml of KCl 0.6 M solution in a citrate buffer 0.05 M, pH 4.8). Proto-last concentration was estimated with use of the hemacytometer and microscope.uspensions containing equal amounts of protoplast (approximately 1 × 106 pro-oplasts) from both strains were gently centrifuged at 3000 rpm, and the added ofml of the fusant solution (30% of polypropynoglycol 4000 in a 0.05-M solution ofaCl2 and glycine 0.025 M, pH 8) and maintained for 10 min at 28 ◦C. The suspen-ions were then washed with KCl 0.6 M solution and gently centrifuged at 1000 × gor 20 min. The resulting pellets were re-suspended in regeneration medium (PLM),nd kept under 60 rpm orbital agitation at 28 ◦C for 24 h. During this period, proto-last germination was followed by observation under the optical microscope. Later,he suspension was spread on selective Petri dishes containing BCCA or BNCA selec-ive media. The dishes were kept at 28 ◦C for 7 days and the developed colonies wereransferred to new selective plate dishes, 1–7 times.
.6. RAPD Analysis
For DNA extraction, mycelia were obtained from 4-day old cultures (at 28 ◦Ct 180 rpm) grown in 500 ml Erlenmeyer flasks containing 100 ml of CLM mediumnoculated with 107 conidia. The mycelia were filtered in Whatman no. 1 filter papernd washed three times with distilled water, and either used directly or stored inreezers at −80 ◦C until needed.
For genomic DNA extraction and RAPD procedures we followed standard pro-ocols as previously described by Ferreira and Grattapaglia [33]. For RAPD, we used
ommercial primers belonging to the OPERON kit, OPB1-15 and OPX 1-20 kits. Themplification was performed on a Techgene thermal cycler (Techne, Princeton, NJ,SA) by 40 cycles with the following sequence: 92 ◦C (45 s); 40 ◦C (1 min and 30 s)nd 72 ◦C (5 min) after 92 ◦C for 4 min. Amplification products were separated bylectrophoresis in 1.5% agarose gel, Tris-borate buffer (0.09 M) and a constant voltagef 100 V, using a Hoefer HE-99 horizontal submarine unit (Hoefer Instruments, San
icrobial Technology 43 (2008) 403–409 405
Fi
2
tcS
btcadbEaoowWa
2
uu
ru
2
pb
3
3
lcTtBAhan
itAsates
wtfobpwB
Fa
thGBf
3b
natory events induced by the formation of heterokaryons, wecompared the amplification profiles of parental strains and fusantsby random amplified polymorphic DNA analysis (RAPD). Over-all, the patterns of amplification obtained with different sets ofprimers were similar between the P. echinulatum parent and fusants
A.J.P. Dillon et al. / Enzyme and M
rancisco, CA, USA) followed by staining with ethidium bromide and visualizationn a UV transilluminator.
.7. Enzyme production in submerged and solid-state cultures
The submerged cultivation was carried out in 500 ml Erlenmeyer flasks con-aining 100 ml of PM media inoculated with conidial suspension to provide a finaloncentration of 1 × 105 conidia per ml and agitated (180 rpm), at 28 ◦C, for 8 days.amples of 2 ml were taken on days 4, 6 and 8 and analyzed for enzymatic activity.
The solid-state culture was performed in flasks with a 12 cm × 3 cm concavease; the flasks were closed with a gauze-covered cotton wool plug. Each flask con-ained 1 g of mass of production media and 1 ml basal salt solution (MS). In thisondition, oxygen is not limited and there is no rise in temperature. The flasks wereutoclaved at 121 ◦C for 20 min. Each flask was then inoculated with sufficient coni-ial suspension to produce a final concentration of 1 × 106 conidia per gram of wheatran. The flasks were incubated at 28 ◦C with a relative humidity of 90% for 5 days.xperiments were carried out with three replicates for each medium compositionnd for each incubation time. To extract the enzymes after incubation, the contentsf each flask were separately added to a 125-ml Erlenmeyer flask containing 10 mlf distilled water, the pH was measured, and 17 ml of 0.05 M citrate buffer (pH 4.8)ere added, mixed, incubated under agitation for 30 min at 4 ◦C, and filtered, usinghatman no. 1 filter paper. The filtrate was stored at 4 ◦C until assay for enzymes
ctivities, as described below.
.8. Enzymatic assay
The enzymatic activity was analyzed on filter paper (filter paper activity—FPA),sing the procedure of Mandels et al. [34] and the �-glucosidase activity was dosedsing salicin as the substrate, according to Chahal [35].
One unit of both FPA or �-glucosidase activity was defined as the capacity toelease 1 �mol of reducing sugar per minute per milliliter of crude or diluted filtratender assay condition.
.9. Statistical analysis
The results were statistically analyzed using analysis of variance with the Tukeyost-test for a p < 0.05, or the genetic variants were compared with the parental oney the t test for a p < 0.05 using the PrismGraphPad program.
. Results
.1. Fusants between P. echinulatum and T. harzianum
To improve the efficiency of protoplast fusion between P. echinu-atum and T. harzianum, we isolated strains resistant to the benomyl,iclohexamide, or nystatin through UV irradiation mutagenesis.hree clones were selected and used to generate protoplasts forhe fusion experiments, a benomyl-resistant P. echinulatum, namedEN3, a cicloheximide and a nystatin-resistant T. harzianum, namedS5ch3 and 5A36N, respectively. These resistance-bearing clonesydrolyze swollen cellulose on a Petri dish and secrete cellulasesnd �-glucosidases at similar levels as their parental strains (dataot presented).
A total of 17 colonies resistant to both benomyl and cyclohex-mide were isolated in 10 protoplast fusion experiments betweenhe P. echinulatum BEN3 (benomyl-resistant) and the T. harzianumS5ch3 (ciclohexamide-resistant). Predominantly, all 17 cloneshowed phenotypic characteristics of P. echinulatum and lost thebility to grow in the presence of cyclohexamide after five rounds ofransfer. Two of these clones, namely PFBC10 and PFBC14, showedarly formation of hydrolytic halos in GTCA medium, and wereelected for further enzymatic activity studies.
Three additional clones resistant to both benomyl and nystatinere selected in 5 protoplast fusion experiments using BEN3 and
he T. harzianum 5A36N (nystatin-resistant). Similar to the otherusion products, these colonies showed phenotypic characteristics
f P. echinulatum and lost their ability to grow in the presence ofenomyl and nystatin after 4–6 rounds of transfer. One fusant inarticular, BP2, showed earlier formation of cellulolytic halo (Fig. 1)hen compared to the parental strain, BEN3, and two other fusants,P4 and BP6 (Fig. 1). We have also observed further variability inFt
ig. 1. Characteristics of the colonies of fusants between Penicillium echinulatumnd Trichoderma harzianum.
he quality and the time required for the formation of hydrolyticalos when conidia from BP4 and BP6 strains were inoculated intoTCA medium (Fig. 2). Two of these segregated colonies (BP6S1 andP6S2), showed clearer halos (Fig. 2 arrows), and were selected for
urther enzymatic studies.
.2. Characterization of the clones obtained by protoplast fusiony RAPD
To obtain molecular evidence of the occurrence of recombi-
ig. 2. Variability in the clones from the BP6 fusants between Penicillium echinula-um and Trichoderma harzianum.
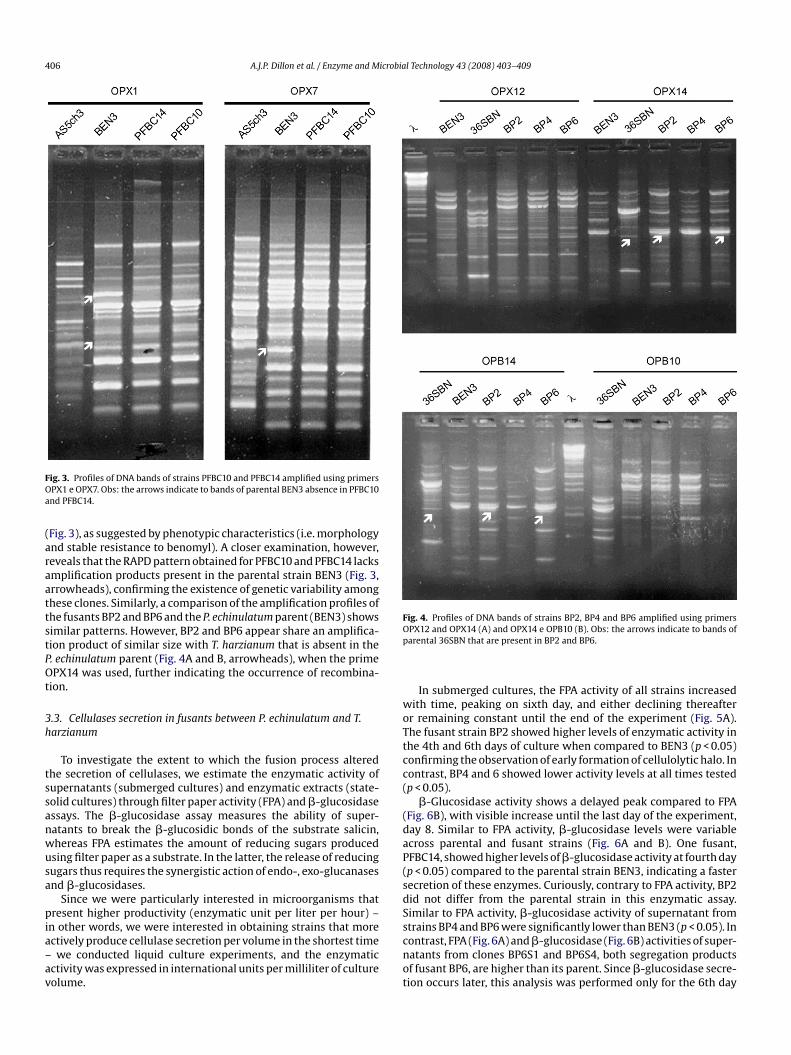
406 A.J.P. Dillon et al. / Enzyme and Microbial Technology 43 (2008) 403–409
FOa
(araattstPOt
3h
tssanwusa
pia–av
Fig. 4. Profiles of DNA bands of strains BP2, BP4 and BP6 amplified using primersOp
woTtcc(
(daP(sdS
ig. 3. Profiles of DNA bands of strains PFBC10 and PFBC14 amplified using primersPX1 e OPX7. Obs: the arrows indicate to bands of parental BEN3 absence in PFBC10nd PFBC14.
Fig. 3), as suggested by phenotypic characteristics (i.e. morphologynd stable resistance to benomyl). A closer examination, however,eveals that the RAPD pattern obtained for PFBC10 and PFBC14 lacksmplification products present in the parental strain BEN3 (Fig. 3,rrowheads), confirming the existence of genetic variability amonghese clones. Similarly, a comparison of the amplification profiles ofhe fusants BP2 and BP6 and the P. echinulatum parent (BEN3) showsimilar patterns. However, BP2 and BP6 appear share an amplifica-ion product of similar size with T. harzianum that is absent in the. echinulatum parent (Fig. 4A and B, arrowheads), when the primePX14 was used, further indicating the occurrence of recombina-
ion.
.3. Cellulases secretion in fusants between P. echinulatum and T.arzianum
To investigate the extent to which the fusion process alteredhe secretion of cellulases, we estimate the enzymatic activity ofupernatants (submerged cultures) and enzymatic extracts (state-olid cultures) through filter paper activity (FPA) and �-glucosidasessays. The �-glucosidase assay measures the ability of super-atants to break the �-glucosidic bonds of the substrate salicin,hereas FPA estimates the amount of reducing sugars producedsing filter paper as a substrate. In the latter, the release of reducingugars thus requires the synergistic action of endo-, exo-glucanasesnd �-glucosidases.
Since we were particularly interested in microorganisms thatresent higher productivity (enzymatic unit per liter per hour) –
n other words, we were interested in obtaining strains that morectively produce cellulase secretion per volume in the shortest timewe conducted liquid culture experiments, and the enzymatic
ctivity was expressed in international units per milliliter of cultureolume.
scnot
PX12 and OPX14 (A) and OPX14 e OPB10 (B). Obs: the arrows indicate to bands ofarental 36SBN that are present in BP2 and BP6.
In submerged cultures, the FPA activity of all strains increasedith time, peaking on sixth day, and either declining thereafter
r remaining constant until the end of the experiment (Fig. 5A).he fusant strain BP2 showed higher levels of enzymatic activity inhe 4th and 6th days of culture when compared to BEN3 (p < 0.05)onfirming the observation of early formation of cellulolytic halo. Inontrast, BP4 and 6 showed lower activity levels at all times testedp < 0.05).
�-Glucosidase activity shows a delayed peak compared to FPAFig. 6B), with visible increase until the last day of the experiment,ay 8. Similar to FPA activity, �-glucosidase levels were variablecross parental and fusant strains (Fig. 6A and B). One fusant,FBC14, showed higher levels of �-glucosidase activity at fourth dayp < 0.05) compared to the parental strain BEN3, indicating a fasterecretion of these enzymes. Curiously, contrary to FPA activity, BP2id not differ from the parental strain in this enzymatic assay.imilar to FPA activity, �-glucosidase activity of supernatant fromtrains BP4 and BP6 were significantly lower than BEN3 (p < 0.05). In
ontrast, FPA (Fig. 6A) and �-glucosidase (Fig. 6B) activities of super-atants from clones BP6S1 and BP6S4, both segregation productsf fusant BP6, are higher than its parent. Since �-glucosidase secre-ion occurs later, this analysis was performed only for the 6th day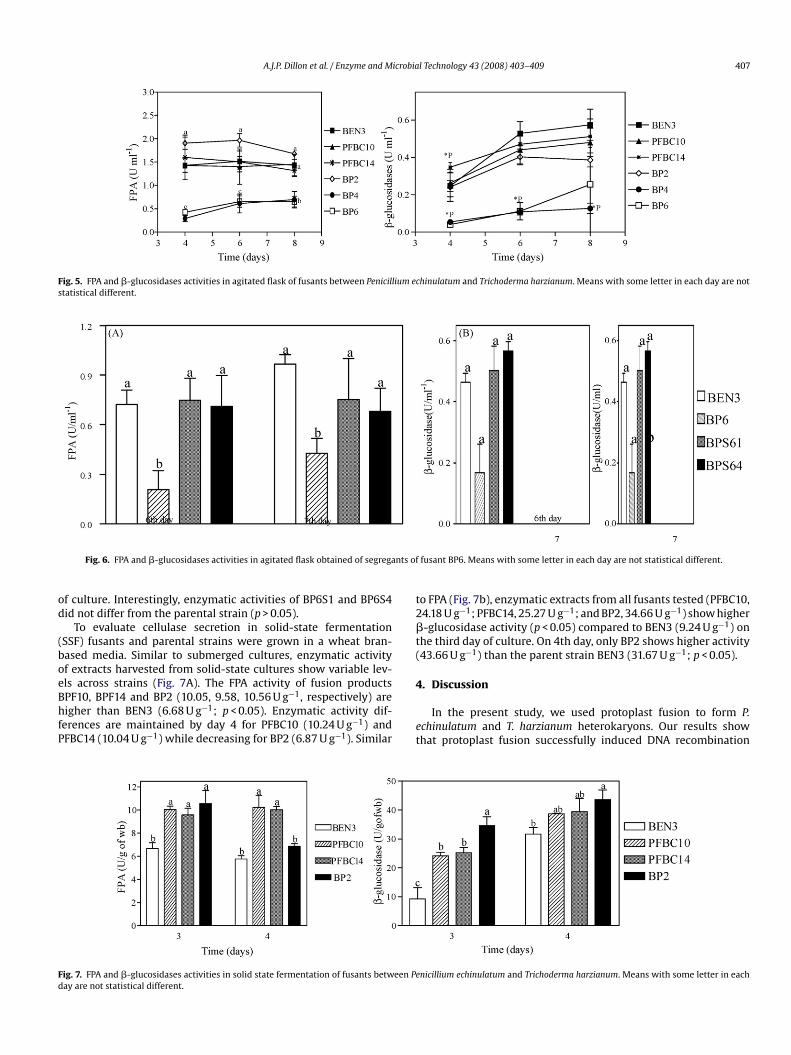
A.J.P. Dillon et al. / Enzyme and Microbial Technology 43 (2008) 403–409 407
Fig. 5. FPA and �-glucosidases activities in agitated flask of fusants between Penicillium echinulatum and Trichoderma harzianum. Means with some letter in each day are notstatistical different.
nts of
od
(boeBhfP
t2�t(
Fd
Fig. 6. FPA and �-glucosidases activities in agitated flask obtained of segrega
f culture. Interestingly, enzymatic activities of BP6S1 and BP6S4id not differ from the parental strain (p > 0.05).
To evaluate cellulase secretion in solid-state fermentationSSF) fusants and parental strains were grown in a wheat bran-ased media. Similar to submerged cultures, enzymatic activityf extracts harvested from solid-state cultures show variable lev-
ls across strains (Fig. 7A). The FPA activity of fusion productsPF10, BPF14 and BP2 (10.05, 9.58, 10.56 U g−1, respectively) areigher than BEN3 (6.68 U g−1; p < 0.05). Enzymatic activity dif-erences are maintained by day 4 for PFBC10 (10.24 U g−1) andFBC14 (10.04 U g−1) while decreasing for BP2 (6.87 U g−1). Similar
4
et
ig. 7. FPA and �-glucosidases activities in solid state fermentation of fusants between Peay are not statistical different.
fusant BP6. Means with some letter in each day are not statistical different.
o FPA (Fig. 7b), enzymatic extracts from all fusants tested (PFBC10,4.18 U g−1; PFBC14, 25.27 U g−1; and BP2, 34.66 U g−1) show higher-glucosidase activity (p < 0.05) compared to BEN3 (9.24 U g−1) on
he third day of culture. On 4th day, only BP2 shows higher activity43.66 U g−1) than the parent strain BEN3 (31.67 U g−1; p < 0.05).
. Discussion
In the present study, we used protoplast fusion to form P.chinulatum and T. harzianum heterokaryons. Our results showhat protoplast fusion successfully induced DNA recombination
nicillium echinulatum and Trichoderma harzianum. Means with some letter in each

4 icrobia
bvs
tiatbeiat
bobcaatlaior
eiaisti
nbriai
ftiiMcFtF�sspf
ttoityeh
tsto
pot[to(
cssd1awloa
g(3ldos
5
sttctFslcfistt
R
08 A.J.P. Dillon et al. / Enzyme and M
etween these two species and that the resulting fusants showariable capacity to secrete cellulases in both submerged and semi-olid cultures.
Because none of the individual strains used carried two resis-ance markers, the colonies recovered in the presence of the twonhibitory substances likely originated from heterokaryons. Thelternate possibility of that the colonies may have arisen dueo a mutation to a second resistance marker is highly unlikely,ecause (i) we did not observe any colony growth in controlledxperiments containing only protoplasts from one species, and (ii)solated clones lost their capacity to grow in selective medium afterfew transfers, suggesting fast chromosomal segregation leading
o somatic cell haploidization common to heterokaryons [23].Our data indicate that protoplast fusion induced DNA recom-
ination in fusants. Overall, the RAPD analysis confirmed thebservations that fusants are similar to P. echinulatum as suggestedy phenotypic characteristic. Nonetheless, apparently an amplifi-ation product of similar size present in the T. harzianum, which isbsent in the P. echinulatum parent was found in two fusants (BP2nd BP6), indicating the possible existence of a processes involvingransfer of DNA segments between the T. harzianum and P. echinu-atum genomes. The distinct RAPD profiles, may be interpreted asn indication of rearrangements occurring in the primer anneal-ng regions that prevent or allow the amplification, as previouslybserved [36] for protoplast fusion between strains of Penicilliumoqueforti and between Volvoriella volvacea and V. bombycina [37].
Our major interest in using protoplast fusion was to first gen-rate genetic variability, because we had been having problemsn obtaining new cellulase mutants from P. echinulatum, althoughn efficient methodology has been applied [15]. The variabil-ty observed in FPA and �-glucosidase activities in the fusanttrains confirm observations that protoplast fusion is an impor-ant approach to produce new strains with different properties ofnterest to cellulase production [21–26].
Some changes occurred in the P. echinulatum, although we wereot able to identify, which specific events were responsible forringing about the genetic variability as seen in most previousesults that used the methodology of protoplast fusion. In addition,t has been shown recently that the association between mutationnd protoplast fusion is very important to obtain fast improvementsn tylosin production from Streptomyces fradiae [27].
Considering the importance of protoplast fusion as a methodor obtaining variability, it is possible that future works will be ableo interpret general results occurring in protoplast fusion. Accord-ngly, in the present work, three of the isolated strains showedmproved cellulolytic activity when compared to parental strains.
ore specifically, BP2 showed higher FPA activity in submergedultures, while BP2, PFBC10, and PFBC14 presented higher yield toPA and �-glucosidase activities in solid-state cultures. It is impor-ant to note that phenotype alterations were not always positive.usant BP6, for example, showed lower enzymatic values, FPA, and-glucosidase when compared to the parental strain. Interestingly,egregation products obtained from this strain, BP6S1 and BP6S4,howed normal cellulases secretion that did not differ from thearental strain, suggesting that genetic instability can generate new
avorable combinations.Moreover, FPA and �-glucosidases of P. echinulatum show good
hermal stability at 50 and 55 ◦C, respectively. This FPA characteris-ic is important if these enzymes would be applied for the defuzzingf cellulosic fibers to reduce the piling tendency of fabrics dur-
ng domestic washing and the stonewashing of denim garmentso obtain the fashionable ‘worn look’ of jeans [29]. Further, andet more important, is the fact that in the enzymatic complex of P.chinulatum there is a good FPA:�-glucosidases ratio to efficientlyydrolyze cellulose [30]. In addition, the secretion of P. echinula-l Technology 43 (2008) 403–409
um enzymes contains all the enzymes of the cellulose complex andecretes xylanases (data not published). It is a valuable characteris-ic for its application in processes such as the enzymatic hydrolysisf cellulose and lignocellulose to produce glucose syrup [28,30].
The 2427 strain of P. echinulatum used as parental strain in theresent work produces cellulases at a yield of nearly 150 FPU mg−1
f cellulose, values that are higher or similar to those obtained withhe famous mutant RUT C30 of T. reesei under the same conditions14]. However, more important is that the BP2 strain, obtained inhe present work through protoplast fusion, presented an increasef 25% in FPA yield, with values of nearly 200 FPU mg−1 of celluloseFig. 5) in relation to the 2427 strain.
The SSF is a very important fermentative process to reduce theost of cellulase production for the saccharification of cellulose,ince lignocellulosic residues can be used as substrate [35]. Fig. 7hows that both fusion products of BP2, PFBC10 and PFBC14 pro-uce more cellulases per gram of wet wheat bran (wwb) (nearly0 FPU g−1 of wwb) than the 2427 (parental) strain, which presentsSSF yield of approximately 6–7 FPU g−1 wwb. Considering thatheat bran has 23% of moisture and containing only 20% of cel-
ulose, the new strains presented over 13 FPU g−1 of dry matter orver 65 FPU g−1 of cellulose, yields that can be considered higher ort least similar to those obtained with other cellulolytic strains [28].
Another important fact in this work is the higher �-lucosidase yield presented on the fourth day of SSF by BP243.66 FPU g−1 wwb), while the parent strain BEN3 showed1.67 FPU g−1 wwb (p < 0.05). The suitable characteristics of the cel-ulases complex in P. echinulatum to hydrolyze cellulose are possiblyue to the high �-glucosidase activities [30]. This can also bebserved in the BEN 3 cellulases and more enhanced in the BP2train generated by protoplast fusion.
. Conclusion
In the present study, we induced genetic variability for cellulaseecretion in strains of P. echinulatum and T. harzianum through pro-oplast fusion. The resulting fusants, morphologically more similaro the parental P. echinulatum, showed variable capacity to secreteellulases in both submerged and solid-state cultures. More impor-antly, some of these fusants showed higher enzymatic titles ofPA e �-glucosidases, in comparison to the P. echinulatum parentaltrain, which secrets cellulases at yield of nearly 130 FPU g−1 of cel-ulose, similar to the mutant strains of T reesei under the sameonditions. The significance of these results is mainly due to theact that mutants of P. echinulatum are very important microorgan-sms for secreting cellulases to hydrolyze cellulose with a view toecond-generation ethanol. Altogether, our results show that pro-oplast fusion can be a useful tool in genetic improvement programso generated better cellulolytic strains.
eferences
[1] Miettinen-Oinonen A, Suominen P. Production of Trichoderma reesei. Endoglu-canases and use of the new cellulase preparations in producing thestonewashed effect on denim fabric. Appl Environ Microbiol 2002;68:3956–64.
[2] Anish R, Suto M, Tomida F. Application of cellulases from an alkalother-mophilic Thermomonospora sp. in biopolishing of denims. Biotechnol Bioeng2007;96:48–56.
[3] Gusakov AV, Salanovich TN, Antonov AI, Ustinov BB, Okunev ON, Burlingame R,et al. Design of highly efficient cellulase mixtures for enzymatic hydrolysis ofcellulose. Biotechnol Bioeng 2007;97:1028–38.
[4] Yinbo Q, Zhu M, Liu K, Bao X, Lin J. Studies on cellulosic ethanol production for
sustainable supply of liquid fuel in China. J Biotechnol 2006;1:1235–40.[5] Himmel ME, Ruth MF, Wyman CE. Cellulase for commodity products from cel-lulosic biomass. Curr Opin Biotechnol 1999;10:358–64.
[6] Aro N, Saloheimo A, Ilmén M, Penttilã M. ACEII, a novel transcriptional activatorinvolved in regulation of cellulase and xylanase genes of Trichoderma reesei. JBiol Chem 2001;276:24309–14.

icrobia
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
A.J.P. Dillon et al. / Enzyme and M
[7] Henrique-Silva F, El-Gogary S, Carle-Urioste E, Matheucci JR, Crivellaro O, El-Dorry H. Two regulatory regions controlling basal cellulose induced expressionof the gene encoding cellobiohydrolase I of Trichoderma reesei are adjacent toits TATA box. Biochem Biophys Res Commun 1996;228:229–37.
[8] Saloheimo A, Aro N, Ilmén M, Penttilã M. Isolation of ace1 gene encoding aCys2-His2 transcription factor involved in regulation of activity of the cellulasepromoter cbh1 of Trichoderma reesei. J Biol Chem 2000;275:5817–25.
[9] Torigoi E, Henrique-Silva F, Escobar-Vera J, Carle-Urioste C, Crivellaro O, El-Dorry H, et al. Mutants of Trichoderma reesei are defective in cellulose induction,but not basal expression of cellulase-encoding genes. Gene 1996;173:199–203.
10] Anwar MN, Suto M, Tomida F. Cellulase for commodity products from cellulosicbiomass. Appl Microbiol Biotechnol 1996;45:684–7.
11] Bailey MJ, Nevalainen KMH. Induction, isolation and testing of stable Tri-choderma reesei mutant with improved production of solubilizing cellulase.Enzyme Microb Technol 1981;3:153–7.
12] Brown JA, Falconer DJ, Wood TM. Isolation and properties of mutants of thefungus Penicillium pinophillum with enhanced cellulase and �-glucosidase pro-duction. Enzyme Microb Technol 1987;9:47–52.
13] Chand P, Aruna A, Maqsood AM, Rao LV. Novel mutation method for increasedcellulase production. J Appl Microbiol 2005;98:318–23.
14] Dillon AJP, Paesi-Toresan S, Barp LB. Isolation of cellulase-producingmutants from Penicillium sp. strains denominated 3MUV3424. Rev Bras Gen1992;15:491–8.
15] Dillon AJP, Zorgi C, Camassola M, Henriques JAP. Use of 2-deoxyglucose in liquidmedia for the selection of mutant strains of Penicillium echinulatum produc-ing increased cellulase and �-glucosidase activities. Appl Microbiol Biotechnol2006;70:740–6.
16] Gadgil NJ, Daginawala HL, Chakrabarti T, Khanna P. Enhanced cellulase produc-tion by a mutant of Trichoderma reesei. Enzyme Microb Technol 1995;17:942–6.
17] Joglekar AV, Karanth NG. Studies on cellulase production by amutant—Penicillium funiculosum UV-49. Biotechnol Bioeng 1984;26:1079–84.
18] Montenecourt BS, Eveleigh DE. Preparation of mutants of Trichoderma ree-sei with enhanced cellulase production. Appl Environ Microbiol 1977;34:777–82.
19] Barnett CC, Berka RM, Fowler T. Cloning and amplification of the gene encodingan extracellular �-glucosidase from Trichoderma reesei: Evidence for improvedrates of saccharification of cellulosic substrates. Biotechnology 1991;9:562–
6.20] Haakana H, Miettinem-Oinonen A, Joutsjoki V, Mäntylä A, Suominen P,Vehmaanperä J. Cloning of cellulase genes from Melanocarpus albomyces andtheir efficient expression in Trichoderma reesei. Enzyme Microbiol Technol2004;34:159–67.
21] Ferenczy L, Kevey F, Zsolt J. Fusion of fungal protoplast. Nature 1974;248:793–4.
[
[
l Technology 43 (2008) 403–409 409
22] Joseph IN, Vallin C, Bravo C, Bugallo F, Marques C. Intraespecific fusionbetween mutants of the cellulolytic strain Trichoderma hamatum. Ciênc Biol1986;16:3–10.
23] Manczinger L, Ferenczy L. Somatic cell fusion of Trichoderma reesei resulting innew genetic combination. Appl Microbiol Biotechnol 1985;22:72–6.
24] Meza V, Moreno P, Tengerdy RP, Gutierrez-Correa M. Transfer of a benomylresistance marker by heat-inactivated Trichoderma reesei protoplast. BiotechnolLett 1995;17:827–32.
25] Prabavathy VR, Mathivanan N, Sagadevan E, Murugesan K, Latithakumari D.Intra-strain fusion enhances carboxymethyl cellulase activity in Trichodermareesei. Enzyme Microb Technol 2006;38:719–23.
26] Toyama H, Shinmy A, Okada H. Protoplast formation from conidia of Tricho-derma reesei by cell wall lytic enzymes of strain of Trichoderma viride. J FermenTechnol 1983;61:409–11.
27] Zhang Y-X, Perry K, Vinci VA, Powell K, Stemmer WPC, Cardayre SB.Genome shuffing leads to rapid phenotypic improvement in bacteria. Nature2002;415:644–6.
28] Camassola M, Dillon AJP. Production of cellulases and hemicellulases by Peni-cillium echinulatum grown on pretreated sugar cane bagasse and wheat bran insolid-state fermentation. J Appl Microbiol 2007;103:2196–204.
29] Camassola M, Bittencourt LR, Sehnem NT, Andreaus J, Dillon AJP. Characteriza-tion of the cellulase complex of Penicillium echinulatum. Biocatal Biotransform2004;22:391–6.
30] Martins LF, Kolling D, Camassola M, Dillon AJP, Ramos LP. Comparisonbetween Penicillium echinulatum and Trichoderma reesei cellulases in rela-tion to their activity against various cellulosic substrates. Bioresour Technol2008;99:1417–24.
31] Toss E, Meneghetti LR, Dillon AJP. Melhoramento genético na linhagem deTrichoderma sp. B72.36S. In: 41◦ Congresso Brasileiro de Genética. Anais, Cax-ambu:1995;230.
32] Mandels M, Reese ET. Induction of cellulase in Trichoderma viride as influencedby carbon source and metals. J Bacteriol 1957;73:269–78.
33] Ferreira ME, Grattapaglia D. Introducão ao uso de marcadores moleculares emanálise genética. 1st ed. Brasília: EMBRAPA-CENARGEN; 1995.
34] Mandels M, Andreotti R, Roche C. Measurement of saccharifying cellulase.Biotechnol Bioeng Symp 1976;6:21–33.
35] Chahal DS. Solid-state fermentation with Trichoderma reesei for cellulase pro-duction. Appl Environ Microbiol 1985;49:205–10.
36] Durand N, Reymond P, Fevre M. Randomly amplified polymorphic DNA recom-bination following an induced parasexual cycle in Penicillium roqueforti. CurrGenet 1993;24:417–20.
37] Zhao JS, Chang T. Interspecific hybridization between Volvoriella volvacea andV. bombycina by PEG-induced protoplast fusion. World J Microb Biotechnol1997;13:145–51.