five novel candida species in insect-associated yeast clades isolated from neuroptera and other...
TRANSCRIPT

Five novel Candida species in insect-associated yeast clades isolated fromNeuroptera and other insects
Nhu H. Nguyen1
Sung-Oui Suh2
Meredith Blackwell3
Department of Biological Sciences, Louisiana StateUniversity, Baton Rouge, Louisiana 70803
Abstract: Ascomycete yeasts are found commonly inthe guts of basidioma-feeding beetles but little isknown about their occurrence in the gut of otherinsects. In this study we isolated 95 yeasts from the gutof adult insects in five neuropteran families (Neu-roptera: Corydalidae, Chrysopidae, Ascalaphidae,Mantispidae and Hemerobiidae) and a roach (Blatto-dea: Blattidae). Based on DNA sequence comparisonsand other taxonomic characteristics, they wereidentified as more than 15 species of Saccharomycetesas well as occasional Cryptococcus-like basidiomyceteyeasts. Yeast species such as Lachancea fermentati,Lachancea thermotolerans and Hanseniaspora vineaewere isolated repeatedly from the gut of three speciesof corydalids, suggesting a close association of thesespecies and their insect hosts. Among the yeastsisolated in this study 12 were identified as five novelCandida species that occurred in three phylogeneti-cally distinct clades. Molecular phylogenetic analysesshowed that Candida chauliodes sp. nov. (NRRL Y-27909T) and Candida corydali sp. nov. (NRRL Y-27910T) were sister taxa in the Candida albicans/Lodderomyces elongisporus clade. Candida dosseyi sp.nov. (NRRL Y-27950T) and Candida blattae sp. nov.(NRRL Y-27698T) were sister taxa in the Candidaintermedia clade. Candida ascalaphidarum sp. nov.(NRRL Y-27908T) fell on a basal branch in a cladecontaining Candida membranifaciens and many otherinsect-associated species. Descriptions of these novelyeast species are provided as well as discussion of theirecology in relation to their insect hosts.
Key words: green lacewings, insect-fungus associ-ations, insect gut yeast
INTRODUCTION
Insects are some of the most successful organisms onearth. They occur in great diversity and high density,and they occupy many niches. The remarkablesuccess of insects has been attributed to their smallsize, reproductive success and exoskeleton modifica-tions, including prevention of desiccation and de-velopment of wings (Siemann et al 1996, Schowalter2000, Dudley 2002). The ecology of insects, includingtheir interactions with symbiotic microorganisms, alsomay contribute to their success. In particular gutmicroorganisms range widely from intracellular bac-teria such as Wolbachia and Buchnera (Werren 1997,Douglas 2003, Dillon and Dillon 2004) to eukaryoticmicrobes inhabiting the gut of New World termitesand wood roaches (Keeling 2002, Kitade 2004). Weare interested in the possibility that yeasts isolatedfrom the guts of insects might produce essentialamino acids, vitamins, enzymes for detoxification,degradation, and fermentation of food resources andeven essential pheromone production (Dowd 1989,1991, Vega and Dowd 2005).
Yeasts have been isolated frequently from the gut orsurface of insects that feed on a variety of materials,including basidiomycete fruiting bodies, woody sub-strates, ephemeral flowers and nectar exudates(Kurtzman 2001, Lachance and Bowles 2002, 2004,Lachance et al 2001a, b, 2005, Rosa et al 2003, Suh etal 2003, 2004a, b, 2005, Teixeira et al 2003, Suh andBlackwell 2004, 2005, Pimentel et al 2005, Nguyen etal 2006). When they were discovered many of theseyeasts represented novel species, some placed inentirely new clades (e.g. Suh et al 2004b, 2006). Mostof the previous isolations, however, have been re-stricted to a small number of specific insect groups,primarily beetles (Coleoptera), various bees andwasps (Hymenoptera), flies (Diptera) and lacewings(Neuroptera). Because of the remarkable diversity ofinsects, we wondered whether yeasts occur regularlyin the gut of other insects. Therefore we attempted toisolate yeasts from the gut and surface of insects in 13orders (48 families), excluding those that feeddirectly on fungi or flowers. Although previously wediscussed some yeasts associated with lacewings (Suhet al 2004a, Nguyen et al 2006) here we focus onadditional isolates from other families of Neuroptera,including those that are associated with other insectsin a more comprehensive study.
Accepted for publication 5 September 2007.
3 Corresponding author. E-mail: [email protected]
1 Current address: Department of Plant and Microbial Biology,University of California, Berkeley, California 94720.2 Current address: American Type Culture Collection, MycologyCollection, 10801 University Boulevard, Manassas, Virginia 20110.
Mycologia, 99(6), 2007, pp. 842–858.# 2007 by The Mycological Society of America, Lawrence, KS 66044-8897
842

MATERIALS AND METHODS
Yeast isolation, culture and identification.—Adult neurop-terans were collected after dark at lights on the sides ofbuildings or in a mercury vapor light trap in Walker andBaton Rouge, Louisiana, and Port Hammock, Florida, in2003, 2004 and 2005, and Barro Colorado Island, Panama,in 2002 (TABLE I). Individual insects were separated in cleancontainers lined with dampened filter paper and held 3 d.Methods for yeast isolation were detailed by Nguyen et al(2006) and Suh et al (2004b). Briefly, starved insects weresubmerged in 95% ethanol 2–3 min to disinfect the surface;this step was followed by a wash with 0.7% saline solution,and the wash liquid was plated on acidified YM agar (DifcoYM broth, 2% plain agar, adjusted to pH 3.5 with HCl) toserve as negative control. Aseptically removed gut contentswere plated separately on acidified YM agar and incubatedat 25 C for 3–4 d. Single yeast colonies were purified at leasttwice and maintained on 2% malt-extract agar (Yarrow1998). An attempt was made to isolate all colonies withdifferent morphologies. Lyophilized holotype specimensand active cultures were deposited in the AgriculturalResearch Service Culture Collection (NRRL) and duplicatecultures at Centraalbureau voor Schimmelcultures (CBS)(TABLE I). Insect vouchers were deposited at the LouisianaState Arthropod Museum (LSAM), Baton Rouge, Louisiana.Morphological and physiological observations were per-formed according to Yarrow (1998) and Barnett et al(2000). Ascospore formation was determined for individualand similar isolates crossed in all combinations on dilute(1:19) V8, half strength cornmeal, and YM agars at 17 and25 C, then observed periodically up to 2 mo.
DNA sequencing and phylogenetic analysis.—Nucleic acidswere extracted and purified with the procedures of Suh andBlackwell (2004) and Suh et al (2004b). The primer setsNS1-SS3E and LS1-LR5 were used for PCR amplification ofthe small subunit (SSU) and large subunit (LSU) rDNA.Purified double-stranded PCR products served as templatesfor sequencing with an ABI PRISMTM BigDye TerminatorCycle sequencing kit v. 3.1. Complete sequences of the SSUrDNA and D1/D2 region of LSU rDNA were obtained withprimers NS1, 18H, SS3E, LS1 and LR3 using an ABI PRISM377 automated DNA sequencer. The sequences from newlyisolated yeasts were compared with sequences acquiredfrom GenBank. These sequences from GenBank were usedin analyses: Candida sp. BG 01-7-24-009A-1-1 (AY242231;AY242343), Candida sp. BG 02-7-19-003A-3-1 (AY520174;AY520304), Candida sp. BG 02-7-21-004G-1-3 (AY520187;AY520317), Candida sp. BCC 7717 (AY228490; AY228491),Candida sp. NRRL Y-27702 (AY640203; AY640205), Candidaalbicans (AB013586; U45776), C. ambrosiae (AY227713;AY013716), C. amphixiae (AY520197; AY520327), C. atlan-tica (AB013527; U45799), C. atmosphaerica (AB013526;U45779), C. arcana (AY242235; AY242347), C. bokatorum(AY426950; AY309798), C. boleticola (AB013532; U45777),C. bolitotheri (AY242142; AY242249), C. cerambycidarum(AY520169; AY520299), C. chrysomelidarum (AY520164;AY520294), C. elateridarum (AY518524; AY518530), C.ergatensis (AB013524; U45746), C. ernobii (AY227714;U94921), C. cylindracea (AB013510; U45823), C. derodonti
(AY242234; AY242346), C. dubliniensis (X99399; U57685),C. fructus (AB018163; U44810), C. fukazawae (AB027756;AY313957), C. fungicola (AB027754; AY313958), C. glabrata(AY198398; U44808), C. haemulonii (AB013572; AY267823),C. homilentoma (AB018166; U45716), C. insectamans(AB013518; U45791), C. insectorum (AB013565; U45791),C. intermedia (AB013571; U44809), C. kruisii (AB013543;U45718), C. lyxosophila (AB013522; U76204), C. maltosa(D14593; U45745), C. membranifaciens (AB013551;U45792), C. mesenterica (AB013552; U45720), C. multi-gemmis (AB013535; U45782), C. nemodendra (AB054546;U70246), C. oleophila (AB013534; U45793), C. oregonensis(AB013545; U44815), C. ovalis (AB054552; U70248), C.palmioleophila (AB013520; U45758), C. parapsilosis(AB013588; U45754), C. peltata (AB054559; U71066), C.picachoensis (AY452053; AY452039), C. pimensis (AY452055;AY452051), C. plutei (AY520388; AY520259), C. pseudoglae-bosa (AB013544; U71072), C. psychrophila (AB013528;U45813), C. pyralidae (AY488127; AY013715), C. quercitrusa(AB013579; U45831), C. rhagii (U63037; U45729), C.sagamina (AB027755; AY313959), C. sake (AB013529;AJ507662), C. shehatae var. insectosa (AB013584; U45773),C. silvicultrix (AB054561; U69879), C. sinolaborantium(AY520198; AY520328), C. sojae (AB013549; U71070), C.sophiae-reginae (AB013542; U45817), C. stellimalicola(AB054573; U84234), C. suecica (AB013554; U45732), C.taliae (AY242145; AY242254), C. tanzawaensis (AY227713;U44811), C. temnochilae (AY242232; AY242344), C. tenuis(AB013516; U45774), C. tropicalis (M55527; U45749), C.tsuchiyae (AB013506; U49064), C. viswanathii (AB013589;U45752), C. carpophila (AY227715; U45707), C. xylopsoci(AY488128; AY013719), Clavispora lusitaniae (M55526;U44817), Debaryomyces hansenii (X58053; U45808), Dekkerabruxellensis (X58052; U45738), Dipodascus tetrasperma(AB000654; U40086), Eremothecium gossypii (AE016820;AE016820), Galactomyces geotrichum (X69842; U40118),Geotrichum clavatum (AB000655; U40112), G. fermentans(AB000651; U40117), G. histeridarum (AY520227;AY520357), Hanseniaspora uvarum (X69844; U84229), H.valbyensis (AY046254; U73596), H. vineae (AY046258;U84224), Issatchenkia orientalis (AB053239; U76347),Kazachstania viticola (AY046234; AF398482), Kluyveromyceslactis (CR382124; CR382124), K. marxianus (X89524;U94924), Kodamaea laetipori (AY520270; AY520398), K.ohmeri (AY520231; AY242300), Lachancea fermentati(X77930; U84239), L. thermotolerans (X89526; U69581),L. waltii (X89527; U69582), Lodderomyces elongisporus(X78600; U45763), Magnusiomyces magnusii (AB000653;U40097), Metschnikowia australis (AB023465;U76526), M.bicuspidata (AB023466; U44822), M. chrysoperlae(AY452054; AY452047), M. corniflorae (AY611607;AY611610), M. noctiluminum (AY611606; AY611609), M.pulcherrima (AB023473; U45736), Nakaseomyces delphensis(X83823; U69576), Naumovia castellii (Z75577; AY545582),Saprochete suaveolens (AB000656; U40119), Pichia anomala(AB054562; AF330115), P. aribbica (AY553853; AY894826),P. holstii (AB018180; U75722), P. karawaiewii (AB018168;U94921), P. membranafaciens (X58055; U75725), P. metha-nolica (AB018181; U75523), P. nakasei (AB053235;U75728), P. nakazawae (AB054279; U45748), P. philogaea
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 843
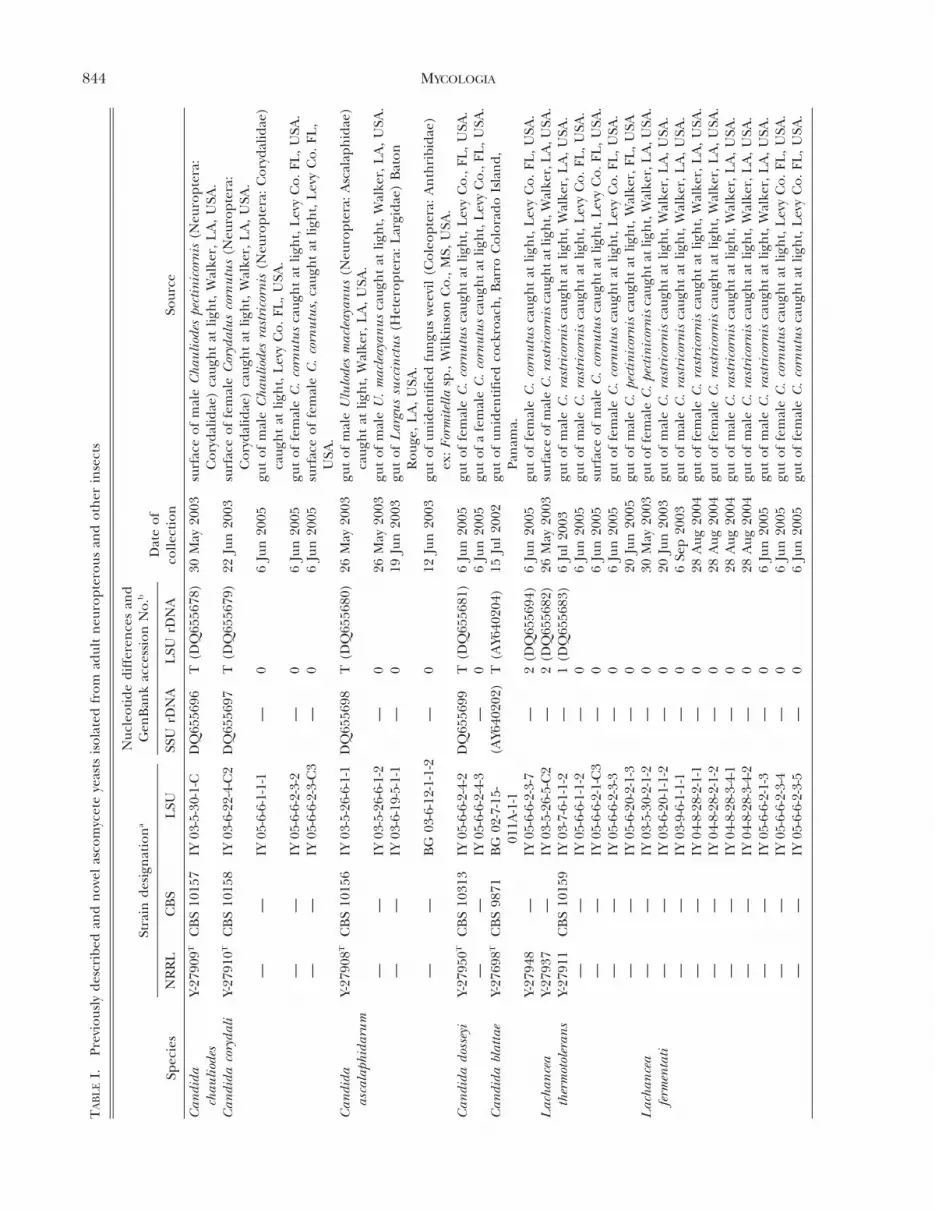
TA
BL
EI.
Pre
vio
usl
yd
escr
ibed
and
no
vel
asco
myc
ete
yeas
tsis
ola
ted
fro
mad
ult
neu
rop
tero
us
and
oth
erin
sect
s
Spec
ies
Stra
ind
esig
nat
ion
a
Nu
cleo
tid
ed
iffe
ren
ces
and
Gen
Ban
kac
cess
ion
No
.b
Dat
eo
fco
llec
tio
nSo
urc
eN
RR
LC
BS
LSU
SSU
rDN
AL
SUrD
NA
Can
did
ach
au
liod
esY-
2790
9TC
BS
1015
7IY
03-5
-30-
1-C
DQ
6556
96T
(DQ
6556
78)
30M
ay20
03su
rfac
eo
fm
ale
Chau
liod
espec
tin
icor
nis
(Neu
rop
tera
:C
ory
dal
idae
)ca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.C
an
did
aco
rydali
Y-27
910T
CB
S10
158
IY03
-6-2
2-4-
C2
DQ
6556
97T
(DQ
6556
79)
22Ju
n20
03su
rfac
eo
ffe
mal
eC
oryd
alu
sco
rnu
tus
(Neu
rop
tera
:C
ory
dal
idae
)ca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
05-6
-6-1
-1-1
—0
6Ju
n20
05gu
to
fm
ale
Chau
liod
esra
stri
corn
is(N
euro
pte
ra:
Co
ryd
alid
ae)
cau
ght
atli
ght,
Lev
yC
o.
FL
,U
SA.
——
IY05
-6-6
-2-3
-2—
06
Jun
2005
gut
of
fem
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-6-2
-3-C
3—
06
Jun
2005
surf
ace
of
fem
ale
C.
corn
utu
s,ca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.C
an
did
aasc
ala
phid
aru
mY-
2790
8TC
BS
1015
6IY
03-5
-26-
6-1-
1D
Q65
5698
T(D
Q65
5680
)26
May
2003
gut
of
mal
eU
lulo
des
macl
eaya
nu
s(N
euro
pte
ra:
Asc
alap
hid
ae)
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY03
-5-2
6-6-
1-2
—0
26M
ay20
03gu
to
fm
ale
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
03-6
-19-
5-1-
1—
019
Jun
2003
gut
of
Larg
us
succ
inct
us
(Het
ero
pte
ra:
Lar
gid
ae)
Bat
on
Ro
uge
,L
A,
USA
.—
—B
G03
-6-1
2-1-
1-2
—0
12Ju
n20
03gu
to
fu
nid
enti
fied
fun
gus
wee
vil
(Co
leo
pte
ra:
An
thri
bid
ae)
ex:
For
mit
ella
sp.,
Wil
kin
son
Co
.,M
S,U
SA.
Can
did
ados
seyi
Y-27
950T
CB
S10
313
IY05
-6-6
-2-4
-2D
Q65
5699
T(D
Q65
5681
)6
Jun
2005
gut
of
fem
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.,F
L,
USA
.—
—IY
05-6
-6-2
-4-3
—0
6Ju
n20
05gu
to
fa
fem
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.,F
L,
USA
.C
an
did
abl
att
ae
Y-27
698T
CB
S98
71B
G02
-7-1
5-01
1A-1
-1(A
Y640
202)
T(A
Y640
204)
15Ju
l20
02gu
to
fu
nid
enti
fied
cock
roac
h,
Bar
roC
olo
rad
oIs
lan
d,
Pan
ama.
Y-27
948
—IY
05-6
-6-2
-3-7
—2
(DQ
6556
94)
6Ju
n20
05gu
to
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.
FL
,U
SA.
Lach
an
cea
ther
mot
oler
an
sY-
2793
7—
IY03
-5-2
6-5-
C2
—2
(DQ
6556
82)
26M
ay20
03su
rfac
eo
fm
ale
C.
rast
rico
rnis
cau
ght
atli
ght,
Wal
ker,
LA
,USA
.Y-
2791
1C
BS
1015
9IY
03-7
-6-1
-1-2
—1
(DQ
6556
83)
6Ju
l20
03gu
to
fm
ale
C.
rast
rico
rnis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY05
-6-6
-1-1
-2—
06
Jun
2005
gut
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-6-2
-1-C
3—
06
Jun
2005
surf
ace
of
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.
FL
,U
SA.
——
IY05
-6-6
-2-3
-3—
06
Jun
2005
gut
of
fem
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-20-
2-1-
3—
020
Jun
2005
gut
of
mal
eC
.pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
FL
,U
SAL
ach
an
cea
ferm
enta
ti—
—IY
03-5
-30-
2-1-
2—
030
May
2003
gut
of
fem
ale
C.
pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY03
-6-2
0-1-
1-2
—0
20Ju
n20
03gu
to
fm
ale
C.
rast
rico
rnis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY03
-9-6
-1-1
-1—
06
Sep
2003
gut
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
04-8
-28-
2-1-
1—
028
Au
g20
04gu
to
ffe
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
04-8
-28-
2-1-
2—
028
Au
g20
04gu
to
ffe
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
04-8
-28-
3-4-
1—
028
Au
g20
04gu
to
fm
ale
C.
rast
rico
rnis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY04
-8-2
8-3-
4-2
—0
28A
ug
2004
gut
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
05-6
-6-2
-1-3
—0
6Ju
n20
05gu
to
fm
ale
C.
rast
rico
rnis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY05
-6-6
-2-3
-4—
06
Jun
2005
gut
of
fem
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-6-2
-3-5
—0
6Ju
n20
05gu
to
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.
FL
,U
SA.
844 MYCOLOGIA

Spec
ies
Stra
ind
esig
nat
ion
a
Nu
cleo
tid
ed
iffe
ren
ces
and
Gen
Ban
kac
cess
ion
No
.b
Dat
eo
fco
llec
tio
nSo
urc
eN
RR
LC
BS
LSU
SSU
rDN
AL
SUrD
NA
Han
sen
iasp
ora
vin
eae
Y-27
936
—IY
03-5
-26-
5-C
1—
2c(D
Q65
5684
)26
May
2003
surf
ace
of
fem
ale
C.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,U
SA.
——
IY05
-6-6
-2-1
-2—
2c6
Jun
2005
gut
of
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.,
FL
,U
SA.
Y-27
944
—IY
05-6
-6-2
-1-1
—3c
(DQ
6556
85)
6Ju
n20
05gu
to
fm
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-6-2
-2-3
—3c
6Ju
n20
05gu
to
fm
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-6-2
-2-4
—3c
6Ju
n20
05gu
to
fm
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-20-
2-1-
1—
3c20
Jun
2005
gut
of
fem
ale
C.
pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,USA
.—
—IY
05-6
-26-
1-1-
1—
3c26
Jun
2005
gut
of
mal
eC
.pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY05
-6-2
6-1-
1-2
—3c
26Ju
n20
05gu
to
fm
ale
C.
pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
——
IY05
-6-2
6-1-
1-3
—3c
26Ju
n20
05gu
to
fm
ale
C.
pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Can
did
acf
.n
eerl
an
dic
aY-
2793
9—
IY03
-6-5
-15-
2-1
—3
(DQ
6556
86)
5Ju
n20
03gu
to
ffe
mal
eU
.m
acl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,U
SA.
Y-27
940
—IY
03-6
-22-
1-2-
1—
3(D
Q65
5687
)22
Jun
2003
gut
of
un
iden
tifi
edea
rwig
(Der
map
tera
:L
abid
uri
dae
)ca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
03-6
-22-
2-1-
1—
322
Jun
2003
gut
of
un
iden
tifi
edcr
icke
t(O
rth
op
tera
:G
ryll
idae
)ca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
05-6
-10-
1-2-
1—
310
Jun
2005
gut
of
fem
ale
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,L
evy
Co
.FL
,USA
.—
—IY
05-6
-10-
1-2-
2—
310
Jun
2005
gut
of
fem
ale
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,L
evy
Co
.FL
,USA
.—
—IY
05-6
-10-
1-2-
3—
310
Jun
2005
gut
of
fem
ale
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,L
evy
Co
.FL
,USA
.C
an
did
atr
opic
ali
s—
—IY
05-6
-20-
2-1-
2—
020
Jun
2005
gut
of
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.C
an
did
am
embr
an
ifaci
ens
——
IY03
-3-2
7-1-
1—
027
Mar
2003
gut
of
un
iden
tifi
edcr
anef
ly(D
ipte
ra:
Tip
uli
dae
)ca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
03-7
-15-
2-1-
1—
015
July
2003
gut
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
03-8
-14-
1-2-
1—
014
Au
g20
03gu
to
fC
hry
soper
laru
fila
bris
(Neu
rop
tera
:C
hry
sop
idae
)ca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
03-8
-14-
2-3-
1—
014
Au
g20
03gu
to
fu
nid
enti
fied
gree
nla
cew
ing
(Ch
ryso
pid
ae)
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Pic
hia
guil
lier
mon
dii
——
IY04
-8-2
8-3-
1-1
—1c
(DQ
6556
88)
28A
ug
2004
gut
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
05-6
-6-2
-1-C
1—
1c6
Jun
2005
surf
ace
of
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.
FL
,U
SA.
——
IY05
-6-6
-2-3
-C2
—1c
6Ju
n20
05su
rfac
eo
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.F
L,U
SA.
——
IY05
-6-6
-2-4
-1—
1c6
Jun
2005
gut
of
fem
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.—
—IY
05-6
-20-
1-2-
1—
1c20
Jun
2005
gut
of
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
05-6
-20-
1-2-
2—
1c20
Jun
2005
gut
of
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
05-6
-20-
1-2-
3—
1c20
Jun
2005
gut
of
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.Y-
2794
9—
IY05
-6-6
-2-3
-C1
—2c
(DQ
6556
89)
6Ju
n20
05su
rfac
eo
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.F
L,U
SA.
Sacc
haro
myc
esce
revis
iae
——
IY03
-5-2
6-5-
1-1
—0
26M
ay20
03gu
to
ffe
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.—
—IY
03-5
-30-
1-1-
1—
030
May
2003
gut
ofm
ale
C.
pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Sacc
haro
myc
esca
rioc
an
us
——
IY05
-6-6
-2-2
-1
—0
6Ju
n20
05gu
to
fm
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.,F
L,
USA
.—
—IY
05-6
-6-2
-2-2
—0
6Ju
n20
05gu
to
fmal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.,
FL
,U
SA.
Tor
ula
spor
adel
bru
ecki
i—
—IY
03-6
-7-6
-1-1
—0
7Ju
n20
03gu
to
ffe
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.
TA
BL
EI.
Co
nti
nu
ed
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 845

Spec
ies
Stra
ind
esig
nat
ion
a
Nu
cleo
tid
ed
iffe
ren
ces
and
Gen
Ban
kac
cess
ion
No
.b
Dat
eo
fco
llec
tio
nSo
urc
eN
RR
LC
BS
LSU
SSU
rDN
AL
SUrD
NA
Can
did
afe
rmen
tati
Y-27
942
—IY
03-7
-6-1
-1-1
—2c
(DQ
6556
90)
6Ju
l20
03gu
to
fm
ale
C.
rast
rico
rnis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Can
did
aqu
erci
tru
saY-
2794
1—
IY03
-6-2
2-4-
1-2
—1c
(DQ
6556
91)
22Ju
n20
03gu
to
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Sacc
haro
myc
opsi
sfe
rmen
tan
s—
—IY
03-6
-20-
1-1-
1—
020
Jun
2003
gut
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.
Kod
am
aea
ohm
eri
——
IY03
-6-2
2-4-
1-1
—0
22Ju
n20
03gu
to
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Pic
hia
gale
ifor
mis
Y-27
938
—IY
03-5
-30-
2-1-
1—
2(D
Q65
5692
)30
May
2003
gut
of
fem
ale
C.
pec
tin
icor
nis
cau
ght
atli
ght,
Wal
ker,
LA
,U
SA.
Am
bros
iozy
ma
mon
ospor
a—
—IY
05-6
-20-
1-1-
1—
1(D
Q65
5695
)20
Jun
2005
gut
of
U.
macl
eaya
nu
sca
ugh
tat
ligh
t,W
alke
r,L
A,
USA
.
Met
schn
ikow
iapu
lcher
rim
aY-
2794
6—
IY05
-6-6
-2-1
-C2
—0
6Ju
n20
05su
rfac
eo
fm
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.F
L,
USA
.
Met
schn
ikow
iach
ryso
per
lae
Y-27
947
—IY
05-6
-6-2
-3-6
—3
(DQ
6556
93)
6Ju
n20
05gu
to
ffe
mal
eC
.co
rnu
tus
cau
ght
atli
ght,
Lev
yC
o.
FL
,U
SA.
Sapro
chet
egi
gas
Y-27
943
CB
S10
312
IY04
-8-2
8-3-
1-C
—4d
28A
ug
2004
surf
ace
of
mal
eC
.ra
stri
corn
isca
ugh
tat
ligh
t,W
alke
r,L
A,U
SA.
Y-27
945
—IY
05-6
-6-2
-1-4
—4d
6Ju
n20
05gu
to
fm
ale
C.
corn
utu
sca
ugh
tat
ligh
t,L
evy
Co
.,F
L,
USA
.
T,
typ
est
rain
;—
,se
qu
ence
no
td
eter
min
ed.
aN
RR
L,
Agr
icu
ltu
ral
Res
earc
hSe
rvic
eC
ult
ure
Co
llec
tio
n,
Nat
ion
alC
ente
rfo
rA
gric
ult
ura
lU
tili
zati
on
Res
earc
h,
Peo
ria,
Illi
no
is;
CB
S,C
entr
aalb
ure
auvo
or
Sch
imm
elcu
ltu
res,
Utr
ech
t,T
he
Net
her
lan
ds;
LSU
,C
oll
ecti
on
of
Gen
etic
Res
ou
rces
,L
SUM
use
um
of
Nat
ura
lSc
ien
ce,
Lo
uis
ian
aSt
ate
Un
iver
sity
,B
ato
nR
ou
ge,
LA
.bD
1/D
2se
qu
ence
so
fL
SUrD
NA
wer
ed
eter
min
edfo
ral
lye
ast
iso
late
s.T
he
seq
uen
ces
wer
eco
mp
ared
wit
hth
ose
of
the
typ
est
rain
of
each
spec
ies.
Nu
cleo
tid
ed
iffe
ren
ces
incl
ud
ega
ps,
inse
rtio
ns,
and
sub
stit
uti
on
s.cT
he
seq
uen
ceco
mp
ared
toth
ety
pe
stra
ino
fth
esp
ecie
s,w
hic
hh
aso
ne
un
det
erm
ined
nu
cleo
tid
e(n
),an
dis
cou
nte
dh
ere
asa
nu
cleo
tid
ed
iffe
ren
ce.
dSe
qu
ence
com
par
edw
ith
ITS
inst
ead
of
the
D1/
D2
regi
on
of
LSU
rDN
A.
TA
BL
EI.
Co
nti
nu
ed846 MYCOLOGIA

(AB054283; U45765), P. scolyti (AB054286; U45788), P.spartinae (AB054292; U45764), P. stipitis (AB054280;U45741), P. triangularis (AB013575; U45796), Saccharomycescariocanus (AY046224; AY130342), S. cerevisiae (SCZ75578;AY048154), Saccharomycodes ludwigii (X69843; U73601),Saturnispora dispora (AB053251; U94937), Schizosaccharomycespombe (X54866; Z19136), Starmera amethionina (AB017897;U75424), Starmerella bombicola (AB013558; U45705), Tetra-pisispora phaffii (AY046245; AY048166), Torulaspora del-brueckii (AB087379; AB087393), Wickerhamiella domercqiae(AB018157; U45847), Williopsis saturnus var. mrakii (Y11318;U94929), Zygosaccharomyces rouxii (X90758; U72163), andZygotorulaspora mrakii (AY046239; U72159).
DNA base pair differences were counted with BLAST 2(Tatusova and Madden 1999) or from a manually alignedsequence database. DNA sequences initially were alignedwith the multi-alignment program Clustal X (Thompson etal 1997) and optimized visually; all missing regions (e.g.beginnings and ends) were excluded from analyses.Maximum parsimony analyses were performed with PAUP4.0b10 (Swofford 2002). Heuristic tree searches wereexecuted with the tree bisection-reconnection branch-swapping algorithm with random sequence analysis. Boot-strap values of the most parsimonious tree were obtainedfrom 1000 replications. The tree was obtained froma combined dataset of SSU and LSU rDNA sequences ofrepresentative species in Saccharomycotina based on theavailability of both gene sequences. Branches containingnew species were separately re-analyzed to include morealigned DNA characters and subsequently superimposed toform a single tree (FIG. 1). Schizosaccharomyces pombe wasdesignated as outgroup taxon for all analyses.
RESULTS
Yeast isolates and new species.—Ninty-five yeast strainswere isolated from species of Neuroptera in fivefamilies. Yeasts were found in the gut of 20 out of 22individual insects in Corydalidae, 12 of 20 individualsin Chrysopidae and 7 of 18 individuals in Ascalaphi-dae. No ascomycete yeasts were isolated from sixindividuals of Mantispidae or four individuals ofHemerobiidae. While the gut contents of a singlecorydalid usually produced more than 100 yeastcolonies on an agar plate, chrysopid and ascalaphidguts produced far fewer colonies.
Rapid identification with the D1/D2 sequencesindicated that 82 isolates were ascomycete yeasts inclass Saccharomycetes and the remaining 13 werebasidiomycete yeasts, mostly identified as Cryptococcusspp. From the sequence comparisons of the D1/D2region the majority of ascomycete yeasts were similarto Lachancea fermentati, L. thermotolerans, Hansenias-pora vineae, Saccharomyces cerevisiae, S. cariocanus,Torulaspora delbrueckii, C. neerlandica, C. membrani-faciens, Pichia guilliermondii, C. fermentati, C. querci-trusa, Saccharomycopsis fermentans, Kodamaea ohmeri,
Pichia galeiformis, C. tropicalis, Ambroziozyma mono-spora, Metschnikowia chrysoperlae and Saprochete gigas(TABLE I). Because these yeasts differed by only 0–2base pairs in the D1/D2 region from the type strainsof each species, they were considered conspecificusing the phenetic standard proposed by Kurtzmanand Robnett (1998). The six isolates near Candidaneerlandica differed by three bp in the D1/D2 regionalong with 10 clear assimilation differences from thetype strain (TABLES I and II). They are being studiedin detail and will not be discussed further in thispaper. Identification of yeasts near Saprochete gigaswas based on the similarity of ITS sequences reportedby Hoog and Smith (2004).
In addition to the previously known yeast taxa 12isolates were identified as five novel species on thebasis of genetic, morphological and physiologicalcharacteristics (TABLES I, II). Some isolates formedhyphae, and all reproduced asexually by budding.Ascospores and asci were not observed in any of thesespecies cultured alone or in crosses, thus placingthem within the limits of Candida, a known poly-phyletic grouping. The new species are described inat least three distinct clades as Candida chauliodes,Candida corydali, Candida ascalaphidarum, Candidadosseyi and Candida blattae.
TAXONOMY
Candida chauliodes N.H. Nguyen, S.-O. Suh et M.Blackwell sp. nov. FIGS. 2–3In medio liquido dextrosum et peptonum et extractum
levidenis continente post 7 d ad 25 C cellulae vegetativaeellipsoideae et subglobosae aut globosae (3–6 3 3–7 mm),singulae vel racemae. Hyphae verae et pseudohyphae nonfiunt. Cultura in agaro extramalti et faecis continente post7 d ad 25 C, cremea et butyrosa; superfices teres. In agarofarina Zeae maydis confecto post 7 d ad 25 C, hyphae veraefiunt sed pseudohyphae non fiunt. Ascosporae non fiunt.Glucosum, galactosum et sucrosum fermentantur. Malto-sum, a-methyl-D-glucosidum, a,a-trehalosum, melibiosum,lactosum, cellobiosum, melezitosum, raffinosum, inulinum,amylum solubile et D-xylosum non fermentantur. Gluco-sum, galactosum, L-sorbosum, D-glucosaminum, D-xylosum,L-arabinosum, trehalosum, glycerolum, ribitolum, xylito-lum, D-glucitolum, D-mannitolum, D-glucono-1, 5-lactonum,2-keto-D-gluconatum, D-gluconatum, acidum succinicum,acidum citricum et ethanolum assimilantur. D-ribosum, D-arabinosum, L-rhamnosum, sucrosum, maltosum, a-methyl-D-glucosidum, cellobiosum, salicinum, arbutinum, melibio-sum, lactosum, raffinosum, melezitosum, inulinum, amylumsolubile, erythritolum, L-arabinitolum, galactitolum, inosi-tolum, D-glucuronatum, DL-acidum lacticum, methanolum,1, 2-propanodiolum, 2, 3-butanodiolum, acidum quinicumet D-glucaratum non assimilantur. Ethylaminum, L-lysinumet cadaverinum assimilantur. Kalium nitratum, natriumnitritum, creatinum, creatininum, glucosaminum, imidazo-
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 847
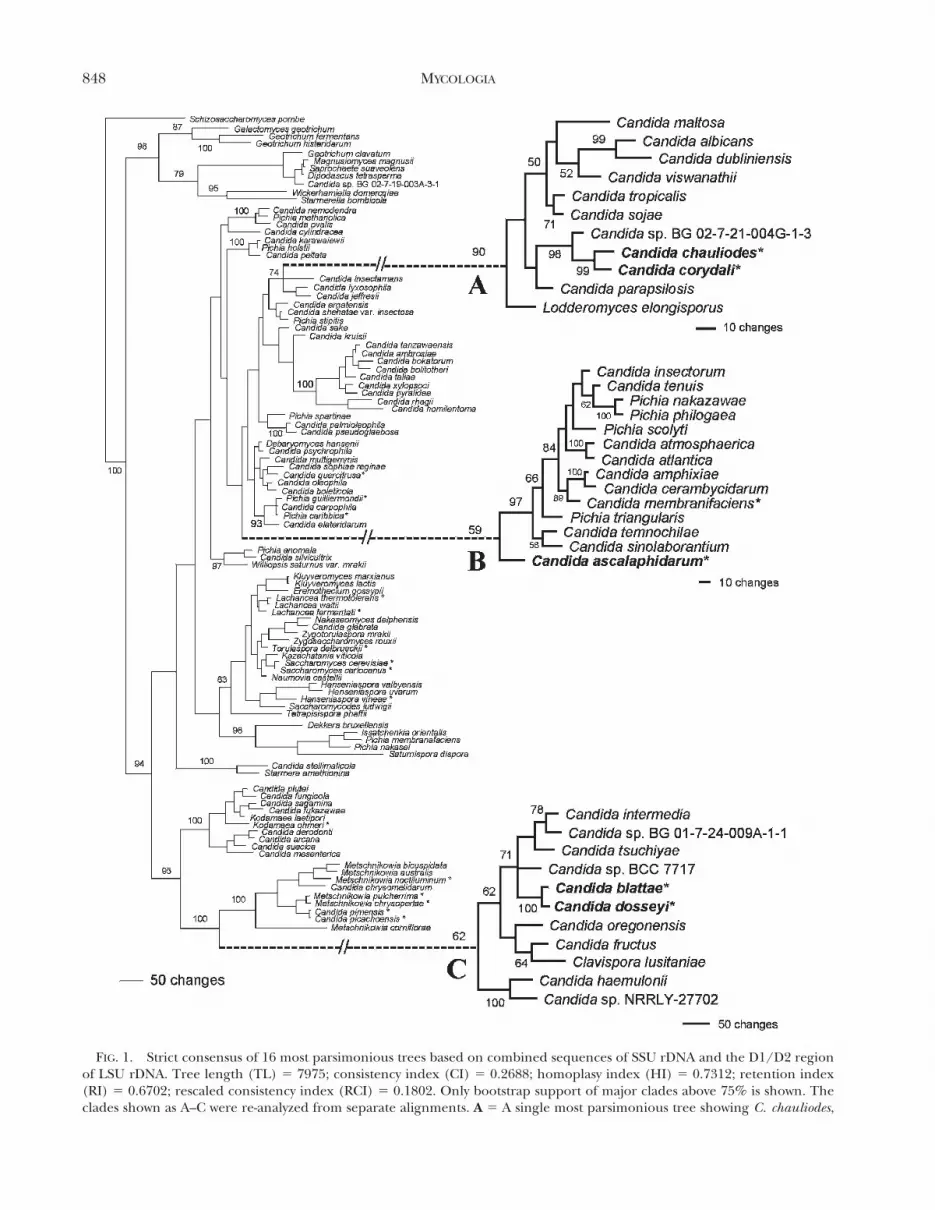
FIG. 1. Strict consensus of 16 most parsimonious trees based on combined sequences of SSU rDNA and the D1/D2 regionof LSU rDNA. Tree length (TL) 5 7975; consistency index (CI) 5 0.2688; homoplasy index (HI) 5 0.7312; retention index(RI) 5 0.6702; rescaled consistency index (RCI) 5 0.1802. Only bootstrap support of major clades above 75% is shown. Theclades shown as A–C were re-analyzed from separate alignments. A 5 A single most parsimonious tree showing C. chauliodes,
848 MYCOLOGIA

lum et D-tryptophanum non assimilantur. Amylum nonformatur. Biotinum externum ad crescentiam necessariumest. Augmentum in 30 C, at non 35 C. In medio 10 mg mL2l
cycloheximido addito crestcit, nec 100 mg mL2l.
HOLOTYPUS. NRRL Y-27909T (5CBS 10157)designat stirpem typicum. Isolata superficio neurop-terum Chauliodes pectinicornis, Walker, LivingstonParish, Louisiana, USA, depositata in CollectioneCulturarum ARS (NRRL), Peoria, Illinois, USA.
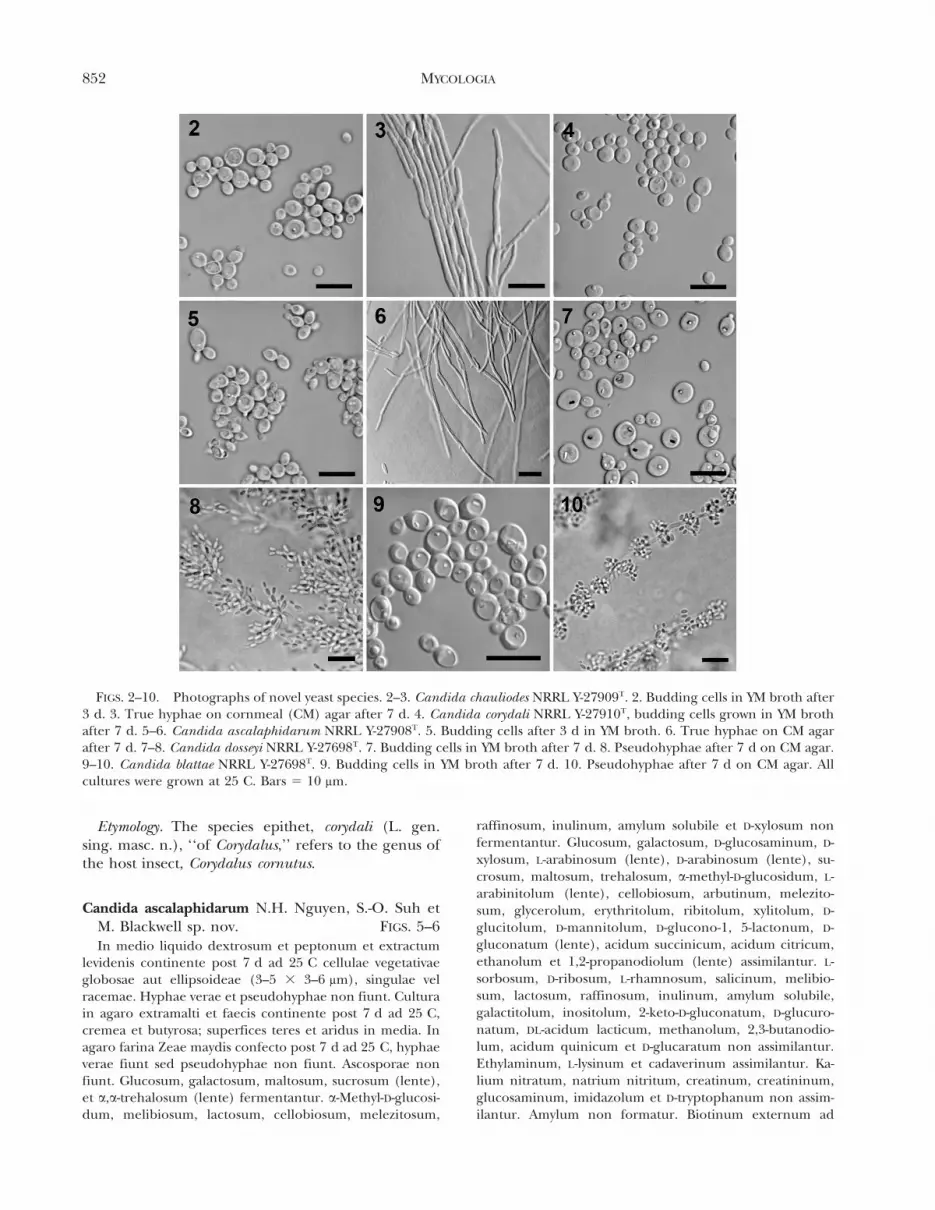
After 7 d growth in YM broth at 25 C cells aremostly globose to some ellipsoidal (3–6 3 3–7 mm),and occur singly, in pairs, or clusters (FIG. 2). Truehyphae and pseudohyphae are not present. After 7 don YM agar at 25 C, colony surface is smooth, cream,butyrous with an undulate margin. After 7 d onDalmau plate culture on cornmeal agar at 25 C truehyphae are present (FIG. 3) but pseudohyphae arenot present. Aerobic growth is off white and thecolony has a smooth surface. Colony margin issmooth with occasional filamentous growth. Asci arenot produced under the conditions tested. (SeeTABLE II for a summary of physiological character-istics.)
Type strain. NRRL Y-27909T (5 CBS 10157) ispreserved as a lyophilized preparation in the Agricul-tural Research Service Culture Collection (NRRL),Peoria, Illinois, USA. The type strain, represented bya single isolate, was taken from the surface of a malefishfly, Chauliodes pectinicornis, at night under lights inWalker, Livingston Parish, Louisiana, USA.
Etymology. The species epithet, chauliodes (L. plu.fem. adj.), refers to the genus of the host insect,Chauliodes pectinicornis.
Candida corydali N.H. Nguyen, S.-O. Suh et M.Blackwell sp. nov. FIG. 4In medio liquido dextrosum et peptonum et extractum
levidenis continente post 7 d ad 25 C cellulae vegetativaeellipsoidae aut globosae et subglobosae (3–7 3 3–8 mm),singulae, binae et racemae; pseudohyphae non fiunt.Cultura in agaro extramalti et faecis continente post 7 dad 25 C, cremea et butyrosa, superfices teres. In agaro farinaZeae maydis confecto post 7 d ad 25 C, pseudohyphae ethyphae verae non fiunt. Ascosporae non fiunt. Glucosum,galactosum (lente), maltosum (infirme), a,a-trehalosum
(lente) fermentantur. a-Methyl-D-glucosidum, sucrosum,melibiosum, lactosum, cellobiosum, melezitosum, raffino-sum, inulinum, amylum solubile et D-xylosum non fermen-tantur. Glucosum, galactosum, L-sorbosum, D-glucosami-num, D-ribosum (lente), D-xylosum (lente), L-arabinosum,sucrosum, maltosum, trehalosum, a-methyl-D-glucosidum,salicinum, arbutinum, melezitosum, glycerolum, ribitolum,xylitolum, L-arabinitolum (infirme), D-glucitolum, D-manni-tolum, D-glucono-1, 5-lactonum, 2-keto- D-gluconatum, D-gluconatum, DL-acidum lacticum (infirme), acidum succi-nicum, acidum citricum et ethanolum assimilantur. D-arabinosum, L-rhamnosum, cellobiosum, melibiosum, lac-tosum, raffinosum, inulinum, amylum solubile, erythrito-lum, galactitolum, inositolum, D-glucuronatum, methano-lum, 1,2-propanodiolum, 2,3-butanodiolum, acidumquinicum et D-glucaratum non assimilantur. Ethylaminum,L-lysinum et cadaverinum assimilantur. Kalium nitratum,natrium nitritum, creatinum, creatininum, glucosaminum,imidazolum et D-tryptophanum non assimilantur. Amylumnon formatur. Biotinum externum ad crescentiam neces-sarium est. Augmentum in 30 C, at non 35 C. In medio10 mg mL2l cycloheximido addito infirme crestcit, nec100 mg mL2l.
HOLOTYPUS. NRRL Y-27910T (5CBS 10158)designat stirpem typicum. Isolata in superficio neu-ropterum Corydalus cornutus, Walker, LivingstonParish, Louisiana, USA, depositata in CollectioneCulturarum ARS (NRRL), Peoria, Illinois, USA.
After 7 d growth in YM broth at 25 C cells areglobose to ellipsoidal (3–7 3 3–8 mm) and occur singly,in pairs, or in small clusters (FIG. 4). Neither pseudo-hyphae nor true hyphae are present. After 7 d on YMagar at 25 C colonies are beige, butyrous with a smoothsurface and margin. After 7 d on Dalmau plate cultureon cornmeal agar at 25 C pseudohyphae and truehyphae are not present. Aerobic growth is off whitewith an undulate colony margin. Ascospores are notproduced under the conditions tested. (See TABLE IIfor a summary of physiological characteristics.)
Type strain. NRRL Y-27910T (5CBS 10158) is pre-served as a lyophilized preparation in the AgriculturalResearch Service Culture Collection (NRRL), Peoria,Illinois, USA. The type strain was isolated from thesurface of a female Corydalus cornutus (Neuroptera:Corydalidae), at night under lights in Walker, Living-ston Parish, Louisiana, USA.
r
C. corydalis and their relatives. TL 5 488; consistency index 5 0.7992; HI 5 0.2008; RI 5 0.5739; RCI 5 0.4587. B 5 A singlemost parsimonious tree showing C. ascalaphidarum and its closest known relatives. TL 5 628; CI 5 0.7245; HI 5 0.2755; RI 5
0.5564; RCI 5 0.4031. C 5 Consensus of two most parsimonious trees showing C. dosseyi, C. blattae and their relatives. TL 5
962; CI 5 0.6850; HI 5 0.3150; RI 5 0.4856; RCI 5 0.3326. Numbers on branches of clades A–C indicate bootstrap supportvalues above 50% derived from 1000 replicates with parsimony analysis. Schizosaccharomyces pombe was designated as outgrouptaxon for all analyses. Asterisks (*) mark yeasts isolated from neuropterans in this study. Dashed lines connect magnified treesto their locations on the base tree.
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 849
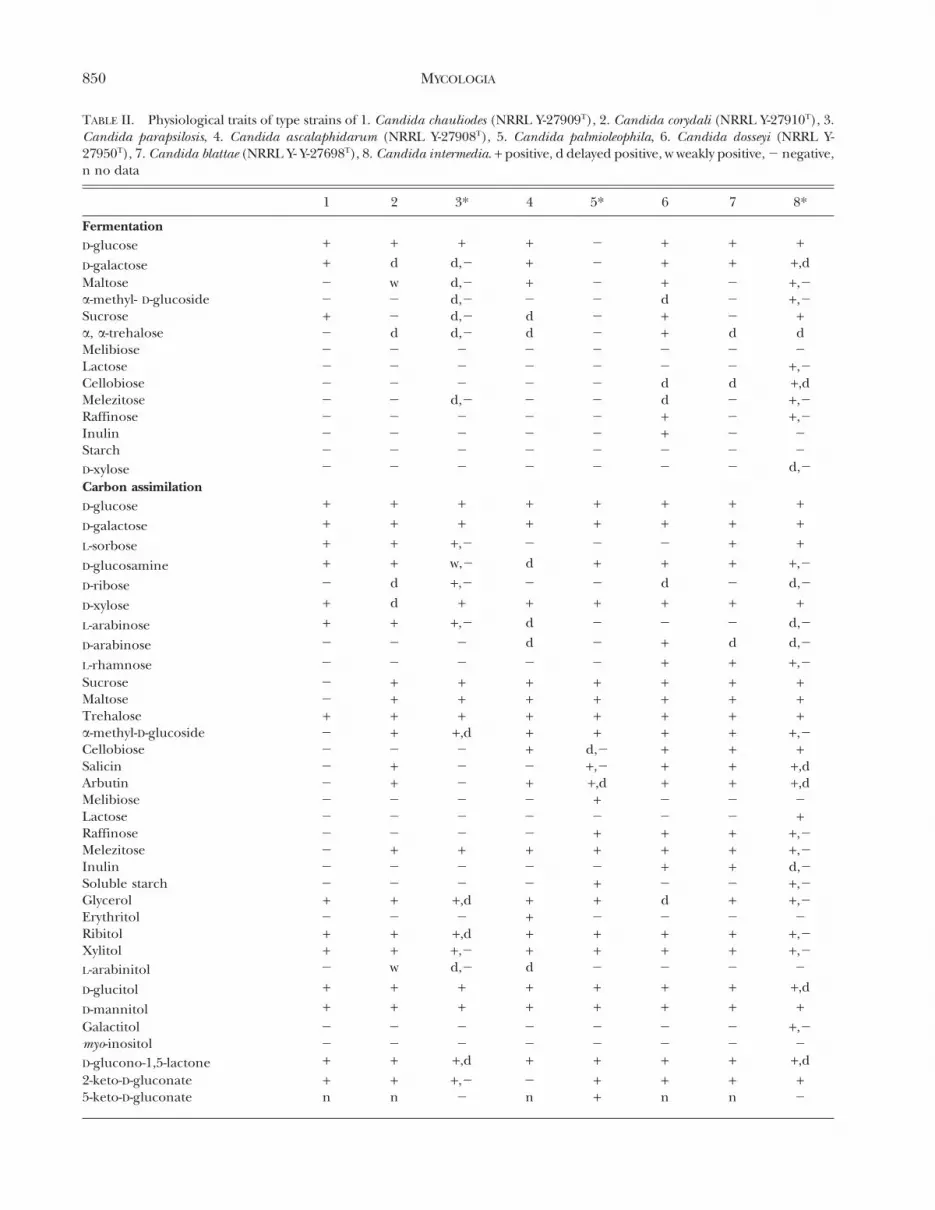
TABLE II. Physiological traits of type strains of 1. Candida chauliodes (NRRL Y-27909T), 2. Candida corydali (NRRL Y-27910T), 3.Candida parapsilosis, 4. Candida ascalaphidarum (NRRL Y-27908T), 5. Candida palmioleophila, 6. Candida dosseyi (NRRL Y-27950T), 7. Candida blattae (NRRL Y- Y-27698T), 8. Candida intermedia. + positive, d delayed positive, w weakly positive, 2 negative,n no data
1 2 3* 4 5* 6 7 8*
Fermentation
D-glucose + + + + 2 + + +D-galactose + d d,2 + 2 + + +,d
Maltose 2 w d,2 + 2 + 2 +,2a-methyl- D-glucoside 2 2 d,2 2 2 d 2 +,2Sucrose + 2 d,2 d 2 + 2 +a, a-trehalose 2 d d,2 d 2 + d dMelibiose 2 2 2 2 2 2 2 2
Lactose 2 2 2 2 2 2 2 +,2Cellobiose 2 2 2 2 2 d d +,dMelezitose 2 2 d,2 2 2 d 2 +,2Raffinose 2 2 2 2 2 + 2 +,2Inulin 2 2 2 2 2 + 2 2
Starch 2 2 2 2 2 2 2 2
D-xylose 2 2 2 2 2 2 2 d,2
Carbon assimilation
D-glucose + + + + + + + +D-galactose + + + + + + + +L-sorbose + + +,2 2 2 2 + +D-glucosamine + + w,2 d + + + +,2
D-ribose 2 d +,2 2 2 d 2 d,2
D-xylose + d + + + + + +L-arabinose + + +,2 d 2 2 2 d,2
D-arabinose 2 2 2 d 2 + d d,2
L-rhamnose 2 2 2 2 2 + + +,2
Sucrose 2 + + + + + + +Maltose 2 + + + + + + +Trehalose + + + + + + + +a-methyl-D-glucoside 2 + +,d + + + + +,2Cellobiose 2 2 2 + d,2 + + +Salicin 2 + 2 2 +,2 + + +,dArbutin 2 + 2 + +,d + + +,dMelibiose 2 2 2 2 + 2 2 2
Lactose 2 2 2 2 2 2 2 +Raffinose 2 2 2 2 + + + +,2Melezitose 2 + + + + + + +,2Inulin 2 2 2 2 2 + + d,2Soluble starch 2 2 2 2 + 2 2 +,2Glycerol + + +,d + + d + +,2Erythritol 2 2 2 + 2 2 2 2
Ribitol + + +,d + + + + +,2Xylitol + + +,2 + + + + +,2
L-arabinitol 2 w d,2 d 2 2 2 2
D-glucitol + + + + + + + +,d
D-mannitol + + + + + + + +Galactitol 2 2 2 2 2 2 2 +,2myo-inositol 2 2 2 2 2 2 2 2
D-glucono-1,5-lactone + + +,d + + + + +,d
2-keto-D-gluconate + + +,2 2 + + + +5-keto-D-gluconate n n 2 n + n n 2
850 MYCOLOGIA
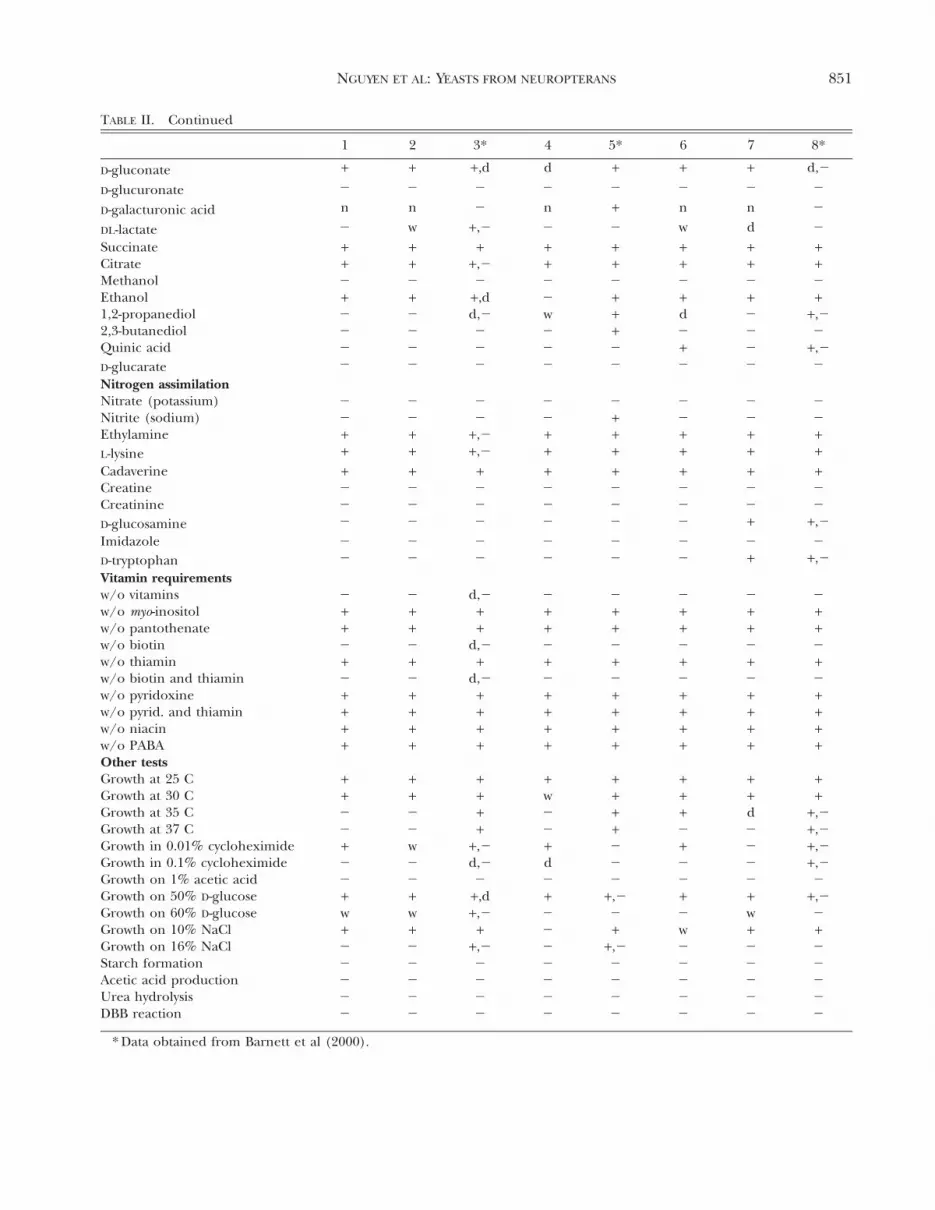
1 2 3* 4 5* 6 7 8*
D-gluconate + + +,d d + + + d,2
D-glucuronate 2 2 2 2 2 2 2 2
D-galacturonic acid n n 2 n + n n 2
DL-lactate 2 w +,2 2 2 w d 2
Succinate + + + + + + + +Citrate + + +,2 + + + + +Methanol 2 2 2 2 2 2 2 2
Ethanol + + +,d 2 + + + +1,2-propanediol 2 2 d,2 w + d 2 +,22,3-butanediol 2 2 2 2 + 2 2 2
Quinic acid 2 2 2 2 2 + 2 +,2
D-glucarate 2 2 2 2 2 2 2 2
Nitrogen assimilationNitrate (potassium) 2 2 2 2 2 2 2 2
Nitrite (sodium) 2 2 2 2 + 2 2 2
Ethylamine + + +,2 + + + + +L-lysine + + +,2 + + + + +Cadaverine + + + + + + + +Creatine 2 2 2 2 2 2 2 2
Creatinine 2 2 2 2 2 2 2 2
D-glucosamine 2 2 2 2 2 2 + +,2
Imidazole 2 2 2 2 2 2 2 2
D-tryptophan 2 2 2 2 2 2 + +,2
Vitamin requirementsw/o vitamins 2 2 d,2 2 2 2 2 2
w/o myo-inositol + + + + + + + +w/o pantothenate + + + + + + + +w/o biotin 2 2 d,2 2 2 2 2 2
w/o thiamin + + + + + + + +w/o biotin and thiamin 2 2 d,2 2 2 2 2 2
w/o pyridoxine + + + + + + + +w/o pyrid. and thiamin + + + + + + + +w/o niacin + + + + + + + +w/o PABA + + + + + + + +Other testsGrowth at 25 C + + + + + + + +Growth at 30 C + + + w + + + +Growth at 35 C 2 2 + 2 + + d +,2Growth at 37 C 2 2 + 2 + 2 2 +,2Growth in 0.01% cycloheximide + w +,2 + 2 + 2 +,2Growth in 0.1% cycloheximide 2 2 d,2 d 2 2 2 +,2Growth on 1% acetic acid 2 2 2 2 2 2 2 2
Growth on 50% D-glucose + + +,d + +,2 + + +,2Growth on 60% D-glucose w w +,2 2 2 2 w 2
Growth on 10% NaCl + + + 2 + w + +Growth on 16% NaCl 2 2 +,2 2 +,2 2 2 2
Starch formation 2 2 2 2 2 2 2 2
Acetic acid production 2 2 2 2 2 2 2 2
Urea hydrolysis 2 2 2 2 2 2 2 2
DBB reaction 2 2 2 2 2 2 2 2
* Data obtained from Barnett et al (2000).
TABLE II. Continued
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 851
Etymology. The species epithet, corydali (L. gen.sing. masc. n.), ‘‘of Corydalus,’’ refers to the genus ofthe host insect, Corydalus cornutus.
Candida ascalaphidarum N.H. Nguyen, S.-O. Suh etM. Blackwell sp. nov. FIGS. 5–6In medio liquido dextrosum et peptonum et extractum
levidenis continente post 7 d ad 25 C cellulae vegetativaeglobosae aut ellipsoideae (3–5 3 3–6 mm), singulae velracemae. Hyphae verae et pseudohyphae non fiunt. Culturain agaro extramalti et faecis continente post 7 d ad 25 C,cremea et butyrosa; superfices teres et aridus in media. Inagaro farina Zeae maydis confecto post 7 d ad 25 C, hyphaeverae fiunt sed pseudohyphae non fiunt. Ascosporae nonfiunt. Glucosum, galactosum, maltosum, sucrosum (lente),et a,a-trehalosum (lente) fermentantur. a-Methyl-D-glucosi-dum, melibiosum, lactosum, cellobiosum, melezitosum,
raffinosum, inulinum, amylum solubile et D-xylosum nonfermentantur. Glucosum, galactosum, D-glucosaminum, D-xylosum, L-arabinosum (lente), D-arabinosum (lente), su-crosum, maltosum, trehalosum, a-methyl-D-glucosidum, L-arabinitolum (lente), cellobiosum, arbutinum, melezito-sum, glycerolum, erythritolum, ribitolum, xylitolum, D-glucitolum, D-mannitolum, D-glucono-1, 5-lactonum, D-gluconatum (lente), acidum succinicum, acidum citricum,ethanolum et 1,2-propanodiolum (lente) assimilantur. L-sorbosum, D-ribosum, L-rhamnosum, salicinum, melibio-sum, lactosum, raffinosum, inulinum, amylum solubile,galactitolum, inositolum, 2-keto-D-gluconatum, D-glucuro-natum, DL-acidum lacticum, methanolum, 2,3-butanodio-lum, acidum quinicum et D-glucaratum non assimilantur.Ethylaminum, L-lysinum et cadaverinum assimilantur. Ka-lium nitratum, natrium nitritum, creatinum, creatininum,glucosaminum, imidazolum et D-tryptophanum non assim-ilantur. Amylum non formatur. Biotinum externum ad
FIGS. 2–10. Photographs of novel yeast species. 2–3. Candida chauliodes NRRL Y-27909T. 2. Budding cells in YM broth after3 d. 3. True hyphae on cornmeal (CM) agar after 7 d. 4. Candida corydali NRRL Y-27910T, budding cells grown in YM brothafter 7 d. 5–6. Candida ascalaphidarum NRRL Y-27908T. 5. Budding cells after 3 d in YM broth. 6. True hyphae on CM agarafter 7 d. 7–8. Candida dosseyi NRRL Y-27698T. 7. Budding cells in YM broth after 7 d. 8. Pseudohyphae after 7 d on CM agar.9–10. Candida blattae NRRL Y-27698T. 9. Budding cells in YM broth after 7 d. 10. Pseudohyphae after 7 d on CM agar. Allcultures were grown at 25 C. Bars 5 10 mm.
852 MYCOLOGIA

crescentiam necessarium est. Augmentum in 35 C, at non40 C. In medio 10 mg mL2l cycloheximido addito crestcit,nec 100 mg mL2l.
HOLOTYPUS. NRRL Y-27908T (5 CBS 10156),designat stirpem typicum. Isolata in ile neuropter-orum Ululodes macleayanus, Walker, Livingston Par-ish, Louisiana, USA, depositata in Collectione Cultur-arum ARS (NRRL), Peoria, Illinois, USA.
After 7 d growth in YM broth at 25 C cells aremostly ellipsoidal to oval with some globose (3–5 3 3–6 mm), occurring singly, in pairs, or clusters (FIG. 5).True hyphae and pseudohyphae are not present.After 7 d on YM agar at 25 C, colony surface appearsin two layers, the outer is smooth, cream and butyrouswhile the inner layer is white, slightly folded and dry.After 7 d on Dalmau plate culture on cornmeal agarat 25 C true hyphae are present (FIG. 6) butpseudohyphae are not present. Aerobic growth is offwhite and smooth. Colony margin is smooth withoccasional filamentous growth. Asci are not producedunder the conditions tested. (See TABLE II fora summary of physiological characteristics.)
Type strain. NRRL Y-27908T (5 CBS 10156) ispreserved as a lyophilized preparation in the Agricul-tural Research Service Culture Collection (NRRL),Peoria, Illinois, USA. The type strain was isolated fromthe gut of a male owlfly, Ululodes macleayanus, 2003 byN.H. Nguyen, at night under lights in Walker,Livingston Parish, Louisiana, USA.
Etymology. The species epithet, ascalaphidarum (L.gen. fem. n.), ‘‘of Ascalaphidae,’’ refers to the familyof the host insect.
Candida dosseyi N.H. Nguyen, S.-O. Suh et M.Blackwell sp. nov. FIGS. 7–8In medio liquido dextrosum et peptonum et extractum
levidenis continente post 7 d ad 25 C cellulae vegetativaeellipsoidae, globosae et subglobosae (3–7 3 5–7 mm),singulae, binae et racemae parvae. Hyphae verae etpseudohyphae non fiunt. Cultura in agaro extramalti etfaecis continente post 7 d ad 25 C, pineceus candidus,butyrosa, superfices teres. In agaro farina Zeae maydisconfecto post 7 d ad 25 C, margina ciliata, superfices teres.Pseudohyphae hyphae fiunt sed hyphae verae non fiunt.Ascosporae non fiunt. Glucosum, galactosum, maltosum a-methyl-D-glucosidum (lente), sucrosum, a,a-trehalosum,cellobiosum (lente), melezitosum (lente), raffinosum etinulinum fermentantur. Melibiosum, lactosum, amylumsolubile et D-xylosum non fermentantur. Glucosum, galac-tosum, D-glucosaminum, D-ribosum (lente), D-xylosum, D-arabinosum, L-rhamnosum, sucrosum, maltosum, trehalo-sum, a-methyl-D-glucosidum, cellobiosum, salicinum, arbu-tinum, raffinosum, melezitosum, inulinum, glycerolum(lente), ribitolum, xylitolum, D-glucitolum, D-mannitolum,D-glucono-1, 5-lactonum, 2-keto-D-gluconatum, D-glucona-tum, DL-acidum lacticum (infirme), acidum succinicum,
acidum citricum, ethanolum, 1,2-propanodiolum (lente), etacidum quinicum assimilantur. L-sorbosum, L-arabinitolum,melibiosum, lactosum, amylum solubile, erythritolum, L-arabinosum, galactitolum, inositolum, D-glucuronatum,methanolum, 2,3-butanodiolum et D-glucaratum non assim-ilantur. Ethylaminum, L-lysinum et cadaverinum assimilan-tur. Kalium nitratum, natrium nitritum, creatinum, creati-ninum, glucosaminum, imidazolum et D-tryptophanum nonassimilantur. Biotinum externum ad crescentiam necessar-ium est. Amylum non formatur. Diazonium caerulian B nonrespondens. Augmentum in 35 C at non 40 C. In medio10 mg mL2l cycloheximido addito crestcit, nec 100 mg mL2l.
HOLOTYPUS. NRRL Y-27950T (5CBS 10313),designat stirpem typicum. Isolata in ile neuropter-orum Corydalus cornutus, 2005, leg. A. Dossey, PortHammock, Levy County, Florida, USA, et isolata abN.H. Nguyen, et depositata in Collectione Cultur-arum ARS (NRRL), Peoria, Illinois, USA.
After 7 d growth in YM broth at 25 C cells areglobose to ellipsoidal (3–7 3 5–7 mm), and occursingly, in pairs, or in small clusters (FIG. 7). Pseudo-hyphae are present but not true hyphae. After 7 d onYM agar at 25 C, colonies are beige and slightlypinkish, butyrous with smooth surface and an un-dulate margin. After 7 d on Dalmau plate culture oncornmeal agar at 25 C pseudohyphae are present(FIG. 8) but true hyphae are not present. Aerobicgrowth is off white with a filamentous colony margin.Ascospores are not produced under the conditionstested. (See TABLE II for a summary of physiologicalcharacteristics.)
Type strain. NRRL Y-27950T (5CBS 10313) is pre-served as a lyophilized preparation in the AgriculturalResearch Service Culture Collection (NRRL), Peoria,Illinois, USA. The type strain was isolated from the gutof a female Corydalus cornutus caught at night underlights in Port Hammock, Levy County, Florida, USA.
Etymology. The species epithet, dosseyi (L. gen.masc. adj.), honors Aaron Dossey, who providedFlorida insect specimens sampled in this study.
Candida blattae N.H. Nguyen, S.O. Suh et M. Black-well sp. nov. FIGS. 9–10In medio liquido dextrosum et peptonum et extractum
levidenis continente post 7 d ad 25 C cellulae vegetativaeglobosae, ellipsoidae, aut subglobosae (3–5 3 4–6 mm),singulae, racemae, et catenae. Pseudohyphae fiunt sed hyphaeverae non fiunt. Cultura in agaro extramalti et faeciscontinente post 7 d ad 25 C, alba, butyrosa, superfices teres.In agaro farina Zeae maydis confecto post 7 d ad 25 C,margina ciliata, superfices teres. Hyphae vera et pseudohyphaehyphae fiunt. Ascosporae non fiunt. Glucosum, galactosum(lente), maltosum (lente), sucrosum (lente), a,a-trehalosum,cellobiosum (lente), melezitosum (lente), raffinosum etinulinum fermentantur. a-Methyl-D-glucosidum, melibiosum,lactosum, amylum solubile et D-xylosum non fermentantur.
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 853

Glucosum, galactosum, L-sorbosum, D-glucosaminum, D-xylosum, D-arabinosum (lente), L-rhamnosum, sucrosum,maltosum, trehalosum, a-methyl-D-glucosidum, cellobio-sum, salicinum, arbutinum, raffinosum, melezitosum, in-ulinum, glycerolum, ribitolum, xylitolum, D-glucitolum, D-mannitolum, D-glucono-1, 5-lactonum, 2-keto-D-glucona-tum, D-gluconatum, DL-acidum lacticum (infirme), acidumsuccinicum, acidum citricum, ethanolum, 1,2-propanodio-lum (infirme) et acidum quinicum assimilantur. D-ribosum,L-arabinitolum, melibiosum, lactosum, amylum solubile,erythritolum, L-arabinosum, galactitolum, inositolum, D-glucuronatum, methanolum, 2,3-butanodiolum et D-glucar-atum non assimilantur. Ethylaminum, L-lysinum et cadaver-inum, glucosaminum et D-tryptophanum assimilantur.Kalium nitratum, natrium nitritum, creatinum, creatininumet imidazolum non assimilantur. Biotinum externum adcrescentiam necessarium est. Amylum non formatur. Di-azonium caerulian B non respondens. Augmentum lente in35 C at non 40 C. In medio 10 mg mL2l cycloheximidoaddito crestcit, nec 100 mg mL2l.
HOLOTYPUS. Y-27698T (5 CBS 9871) designat stirpemtypicum. Isolata in ile blattae, 2002, at Barro ColoradoIsland, Panama, et depositata in Collectione CulturarumARS (NRRL), Peoria, Illinois, USA.
After 7 d growth in YM broth at 25 C cells areglobose to ellipsoidal (3–5 3 4–6 mm), and occursingly, in pairs or in small clusters and chains (FIG. 9).Pseudohyphae are present (FIG. 10). After 7 d on YMagar at 25 C, colonies are off white, butyrous withsmooth surface and an undulate margin. After 7 d onDalmau plate culture on cornmeal agar at 25 C bothpseudohyphae (FIG. 10) and true hyphae are present.Aerobic growth is off white with a filamentous colonymargin. Ascospores are not produced under theconditions tested. (See TABLE II for a summary ofphysiological characteristics.)
Type strain. Y-27698T (5 CBS 9871) is preserved asa lyophilized preparation in the Agricultural ResearchService Culture Collection (NRRL), Peoria, Illinois,USA. The type strain was isolated from the gut of anunidentified cockroach at Barro Colorado Island,Panama.
Etymology. The species epithet, blattae (L. gen. fem.n.), is from the Greek word blatta, ‘‘roach,’’ the sourceof the type strain of this species.
Isolates of new species.—Candida chauliodes wasrepresented by a single isolate (NRRL Y-27909) whileothers were isolated on at least two occasions(TABLE I). There was no D1/D2 sequence variationamong the strains in each new species except in C.blattae, in which the strains differed at two base pairs(TABLE I). BLAST with the D1/D2 sequences showedthat C. chauliodes and C. corydali were most similar tosequences of Candida sp. ST-18, isolated from Thai-land and Candida sp. BG 02-7-21-004G-1-3 fromPanama in GenBank. Candida ascalaphidarum
showed the highest similarity to C. palmioleophila inthe D1/D2 sequence, while C. dosseyi and C. blattaewere most closely related to Candida sp. ST-211, alsoisolated from Thailand. However the new specieswere clearly distinguished from the most similarsequences by nine or more base pairs.
DISCUSSION
Phylogeny of the new yeast species.—A consensus of 16most parsimonious trees based on combined SSU andLSU rDNA indicates that both novel and previouslydescribed yeasts associated with neuropteran insectsdo not form a monophyletic group (FIG. 1). Thenovel species were placed within three clades (A–C),only one of which has relatively high bootstrapsupport of 90% (clade A). Within the C. albicansclade (A), C. chauliodes and C. corydali were sister taxaand their grouping was well supported by bootstrapanalysis (99%). The two species were stronglysupported as members of a monophyletic group withCandida sp. ST-18 and Candida sp. BG 02-7-21-004G-1-3 based on analysis of D1/D2 sequences (data notshown). Sequence comparisons also indicated thatthe two novel species are most closely related toCandida sp. ST-18.
The phylogenetic position of Candida ascalaphi-darum is unclear and no closely related sister taxonwas identified. Although it was placed basal to theCandida membranifaciens clade (B), the position wasnot supported statistically. Furthermore C. palmioleo-phila was the sequence most similar to C. ascalaphi-darum, differing by 27 bp in the D1/D2 region butnot grouping together in the phylogenetic tree(FIG. 1). Candida dosseyi and Candida blattae wereplaced as sister species within the Candida intermediaclade (C) with strong bootstrap support. The twonovel taxa formed a monophyletic group withCandida sp. ST-211 with 100% bootstrap supportbased on D1/D2 sequences (tree not shown).
The assimilation and fermentation capabilities ofthe yeasts described here are generally different frommany of the yeasts associated with mushroom orwood-ingesting beetles (Suh et al 2003, 2004b).Physiological profiles of the novel species also varysubstantially from the closest described species andfrom each other (TABLE II), and these traits may beuseful in designing taxonomic keys to species.However DNA sequence data provide a faster andmore effective way to identify an unknown yeast.
Insect hosts and yeasts.—The order Neuropteraincludes primitive holometabolous insects that pos-sess membranous yet strongly veined wings. Speciesfrom five of the more common neuropteran families
854 MYCOLOGIA

present in the southeastern USA were chosen for thisstudy. These include the green lacewings (Chrysopi-dae), which are well known in biological controlstudies, fishflies and dobsonflies (Corydalidae), owl-flies (Ascalaphidae), mantisflies (Mantispidae) andbrown lacewings (Hemerobiidae). As adults Coryda-lidae and Chrysopidae feed mostly on varioussaprobic substances while Ascalaphidae, Mantispidaeand Hemerobiidae are predaceous. In contrast thelarvae of all five families are predators (Tauber 1991).
Corydalidae. Many of the yeasts isolated in this studywere associated with the family Corydalidae, of whichwe sampled adults of three species, Corydalus cornutus,Chauliodes rastricornis and Chauliodes pectinicornis(TABLE I). The larva of the dobsonfly, Corydaluscornutus, is a well known, large insect of aquaticsystems. It spends its larval stages in the water of rocky-bottomed streams where it preys on larvae of blackflies(Diptera: Simuliidae) and net spinning caddisflies(Trichoptera: Hydropsychidae) (Stewart et al 1973). Incontrast the adult has not received extensive study andits ecology is poorly known. The two Chauliodes speciesalso have biological characteristics similar to Corydaluscornutus. The feeding habits of these short-lived adultshave been reported only from laboratory experimentswhere they lap up water or sweet liquid substances.Adults of all three species have been reported to visittraps baited with a mixture of bananas, brown sugar,molasses and beer (Parfin 1952).
Representatives of the four novel yeast speciesdescribed here were isolated from the three adultcorydalids mentioned above, although not consistent-ly (TABLE I). Six species within the ‘‘Saccharomycescomplex’’ were found in the gut of these corydalids.Interestingly, three of these six species (L. thermo-tolerans, L. fermentati and H. vineae) were isolatedrepeatedly from several localities throughout thethree years of the study (TABLE I).
In general L. thermotolerans and L. fermentati havebeen isolated from sweet, fermenting substratesaround the world. Conversely Hanseniaspora vineaealso has a widespread distribution but is generallyknown from soil and spoiled food (Barnett et al2000). The repeated isolation of these yeasts suggeststhat they are closely associated with the corydalidscollected in this study. It is interesting to note thatthese yeasts have worldwide distributions, perhapsdue to insects dispersing them widely (Lachance et al2001b), inoculating yeast spawn to fresh saccharinesubstrates and spoiling them (Stratford et al 2002).Insects such as Drosophila and small beetles probablyare the first to inoculate fresh substrates, but thecorydalid vectors likely contribute to dispersion of theyeasts.
Isolation of Saccharomyces cerevisiae from corydalids
is of interest. Although renowned as an industrialyeast, little is known about its occurrence in nature.Literature reports of natural occurrences of theseyeasts are sparse and they are not often isolated, evenin complex communities containing many otherspecies. For example Sniegowski et al (2002) found10 isolates out of 79 with enrichment cultures forSaccharomyces species associated with oak trees andsurrounding soil. Morais et al (1992) found threeisolates out of 962 while sampling yeasts that occurwith Drosophila communities in Brazil. Naumov et al(1998) found only two isolates from slime fluxes ofoak. Similarly we obtained only two isolates withina 4 d period in the gut of two different corydalidhosts. The frequency of isolation based on theprevious studies mentioned above and our currentdata indicates that the species does not occurabundantly in natural habitats. The inconsistentisolation of this species and others that were foundonly once or twice in this study (TABLE I) suggests thatthey may not have been closely associated withneuropteran insects but perhaps were only transientsacquired during feeding.
Observations of the gut morphology in all threespecies of corydalids from which yeasts were isolatedshowed five finger-like projections (diverticulae) thatapparently have not been reported previously. Thesediverticulae occur in the foregut region behind thecrop. Such pouches in the gut of insects, includingchrysopids, often serve for temporary storage of food(Tang and Ward 1998) in which yeasts, bacteria orother obligate microorganisms reside (Bismanis 1976,Stamopoulos and Tzanetakis 1988, Marchini et al2002, Woolfolk et al 2004). However we did notdetermine whether yeasts were present in thepouches, but if they are localized there this mayaccount for the consistent associations observedbetween corydalid neuropterans and certain yeasts.
Chrysopidae. Green lacewing larvae are predaceouson small insects, including aphids, but as adults theyfeed on sweet substances such as nectar, insecthoneydew and pollen (Hagen et al 1970). Yeasts havenot been isolated from larvae, but they often occur inthe gut of adults. Yeasts in the genus Metschnikowiawere found to be associated with lacewings (e.g.Metschnikowia noctiluminum and Metschnikowia chry-soperlae) (Woolfolk and Inglis 2003, Suh et al 2004a,Nguyen et al 2006). Their related anamorphs, Candidapicachoensis and Candida pimensis, were describedfrom green lacewing gut isolates collected in Arizonaand subsequently found in Louisiana (Suh et al 2004a,Nguyen et al 2006). Metschnikowia chrysoperlae wasfound only in Arizona and not in Louisiana. Woolfolkand Inglis (2003) reported M. pulcherrima-like yeastsin the gut of Chrysoperla rufilabris, and Suh et al
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 855

(2004a) classified the species as M. chrysoperlae basedon sequence comparisons and ascospore formation. Inthis study M. chrysoperlae and M. pulcherrima also wereisolated from Corydalus cornutus collected fromFlorida, although each yeast species was collectedrespectively only once from the gut and the bodysurface of different insect individuals (TABLE I). Thesedata taken together indicate that the association ofgreen lacewings and yeasts in the Metschnikowia cladeis probably widespread across the southern USA.
Unlike the corydalid gut there have been severalreports of adult lacewing gut ultrastructure showingmany yeast cells within diverticulae, which take upconsiderable space within the body cavity (Woolfolket al 2004). Interestingly it was noted that lacewingsthat fed on sugary substances have a much largertracheal trunk around the diverticulum, a modifica-tion suggested as necessary to satisfy the oxygendemand of resident yeasts (Canard et al 1990, Gibsonand Hunter 2005). Direct observation of yeast cells,isolation in culture, and gut structural modificationsindicate that yeasts make up some of the underap-preciated normal biota of the adult green lacewinggut.
Other insects. Other neuropteran insects sampledincluding owlflies, mantisflies and brown lacewings(Ascalaphidae, Mantispidae and Hemerobiidae respec-tively) are predaceous in both larval and the adultstages. Few yeasts were recovered from these insects,and these were mostly basidiomycete yeasts, whichmight be environmental contaminants. Generally pre-daceous insects we have sampled previously (e.g. tigerbeetles, carabid beetles and reduviid bugs) did notcontain gut yeasts, and this trend continued in thisstudy. For example yeasts were not isolated from theguts of predaceous lacewing larvae but they werepresent in the guts of adults that feed on plantmaterials. The yeasts isolated from owlflies were foundin the gut of other insects (TABLE I), suggesting thatthese might be contaminants resulting from predationon insects. In addition the gut of adult owlflies did notcontain any well defined pouches or projections such asthose found in the corydalids and chrysopids. We haveobserved a similar trend in fungus-feeding beetles, inwhich the absence of specialized gut pouches some-times corresponds with a lack of yeast/insect specificity.Such observations deserve additional study.
Several generalizations can be made based on ourfindings: (i) The novel species described here werenot isolated in high frequency, and they probably arenot specifically associated with their hosts; (ii) threespecies of yeasts from the ‘‘Saccharomyces complex’’were isolated frequently from the three insect speciessampled in the family Corydalidae; (iii) greenlacewing adults seem to be connected more often
with species of Metschnikowia than with other yeasts;and (iv) adult predaceous insects such as owlflies,mantisflies and brown lacewings were not closelyassociated with yeasts.
ACKNOWLEDGMENTS
We thank Michael Ferro for identification of the owlflies. Weacknowledge Ms Glenda Newman, principal, North CorbinElementary School, Walker, Louisiana, for permission tocollect at the school. GenBank and the NRRL and CBScollections of fungal cultures are gratefully acknowledged.This research was supported by the National ScienceFoundation, Biodiversity Surveys and Inventories Program(DEB-0072741 and DEB-0417180), including REU supple-ments. The REU supplements and a Howard HughesMedical Institute grant through the Undergraduate Bi-ological Sciences Education Program to Louisiana StateUniversity provided support for NHN. We also acknowledgethe use of the DNA sequencing facility supported by NSFMulti-user Equipment Grant (DBI-0400797) to Robb Brum-field, and the assistance of Nannette Crochet for sequencing.
LITERATURE CITED
Barnett JA, Payne RW, Yarrow D. 2000. Yeasts: characteristicsand identification. 3rd ed. Cambridge: CambridgeUniversity Press. 1139 p.
Bismanis JE. 1976. Endosymbionts of Sitodrepa panicea. CanJ Microbiol 22:1415–1424.
Canard M, Kokubu K, Duelli P. 1990. Tracheal trunkssupplying air to the foregut and feeding habits in adultsof European green lacewing species (Insecta: Neurop-tera: Chrysopidae). In: Mansell MW, Aspock H, eds.Advances in Neuropterology. Proceedings of the 3rdInternational Symposium on Neuropterology. Pretoria,Republic of South Africa: Dept. Agricultural Develop-ment. p 277–286.
de Hoog GS, Smith MT. 2004. Ribosomal gene phylogenyand species delimitation in Geotrichum and its tele-omorphs. Stud Mycol 50:489–515.
Dillon RJ, Dillon VM. 2004. The Gut Bacteria of Insects:nonpathogenic interactions. Annu Rev Entomol 49:71–92.
Douglas AE. 2003. Buchnera bacteria and other symbionts ofaphids. In: Bourtzis K, Miller TA, eds. Insect Symbiosis.Boca Raton: CRC Press. p 23–38.
Dowd PF. 1989. In situ production of hydrolytic detoxifyingenzymes by symbiotic yeasts in the cigarette beetle(Coleoptera: Anobiidae). J Econ Entomol 82:396–400.
———. 1991. Symbiont-mediated detoxification in insectherbivoures. In: Barbosa P, Krischik VA, Jones CG, eds.Microbial Mediation of Plant-herbivore Interactions.New York: John Wiley & Sons. p 411–440.
Dudley R. 2002. The Biomechanics of Insect Flight: form,function, evolution. Princeton: Princeton UniversityPress. 476 p. 6 pl.
Gibson CM, Hunter MS. 2005. Reconsideration of the role
856 MYCOLOGIA

of yeasts associated with Chrysoperla green lacewings.Biol Control 32:57–64.
Hagen KS, Tassan RL, Sawall EF. 1970. Some ecophysiolog-ical relationships between certain Chrysopa, honeydewsand yeasts. Boll Lab Ent Ag Portici 28:113–134.
Keeling JP. 2002. Molecular phylogenetic position ofTrichomitopsis termopsidis (Parabasalia) and evidencefor the Trichomitopsiinae. Europ J Protistol 38:279–286.
Kitade O. 2004. Comparison of symbiotic flagellate faunaebetween termites and a wood-feeding cockroach of thegenus Cryptocercus. Microbes Environ 19:215–220.
Kurtzman CP. 2001. Six new anamorphic ascomycetousyeasts near Candida tanzawaensis. FEMS Yeast Res 1:177–185.
———, Robnett CJ. 1998. Identification and phylogeny ofascomycetous yeasts from analysis of nuclear largesubunit (26S) ribosomal DNA partial sequences.Antonie van Leeuwenhoek 73:331–371.
Lachance M-A, Bowles JM. 2002. Metschnikowia arizonensisand Metschnikowia dekortorum, two new large-sporedyeast species associated with floricolous beetles. FEMSYeast Res 2:81–86.
———, ———. 2004. Metschnikowia similis sp. nov. andMetschnikowia colocasiae sp. nov., two ascomycetousyeasts isolated from Conotelus spp. (Coleoptera: Niti-dulidae) in Costa Rica. Stud Mycol 50:69–76.
———, ———, Kwon S, Marinoni G, Starmer WT, JanzenDH. 2001a. Metschnikowia lochheadii and Metschniko-wia drosophilae, two new yeast species isolated frominsects associated with flowers. Can J Microbiol 47:103–109.
———, Ewing CP, Bowles JM, Starmer WT. 2005. Metschni-kowia hamakuensis sp. nov, Metschnikowia kamakouanasp. nov., and Metschnikowia mauinuiana sp. nov., threeendemic yeasts from Hawaiian nitidulid beetles. Int JSyst Evol Microbiol 55:1369–1377.
———, Starmer WT, Rosa CA, Bowles JM, Barker JS, JanzenDH. 2001b. Biogeography of the yeasts of ephemeralflowers and their insects. FEMS Yeast Res 1:1–8.
Marchini D, Rosetto M, Dallai R, Marri L. 2002. Bacteriaassociated with the oesophageal bulb of the medflyCeratitis capitata (Diptera: Tephritidae). Curr Micro-biol 44:120–124.
Morais PB, Hagler AN, Rosa CA, Mendonca-Hagler LC,Klaczko LB. 1992. Yeasts associated with Drosophila intropical forests of Rio de Janeiro, Brazil. Can JMicrobiol 38:1150–1155.
Naumov GI, Naumova ES, Sniegowski PD. 1998. Saccharo-myces paradoxus and Saccharomyces cerevisiae areassociated with exudates of North American oaks.Can J Microbiol 44:1045–1050.
Nguyen NH, Suh S-O, Erbil CK, Blackwell M. 2006.Metschnikowia noctiluminum sp. nov., Metschnikowiacorniflorae sp. nov., and Candida chrysomelidarum sp.nov., isolated from green lacewings and beetles. MycolRes 110:346–356.
Parfin IS. 1952. The Megaloptera and Neuroptera ofMinnesota. Am Midl Nat 47:421–434.
Pimentel MR, Antonini Y, Martins RP, Lachance MA, Rosa
CA. 2005. Candida riodocensis and Candida cellae, twonew yeast species from the Starmerella clade associatedwith solitary bees in the Atlantic rain forest of Brazil.FEMS Yeast Res 5:875–879.
Rosa CA, Lachance MA, Silva JO, Teixeira AC, Marini MM,Antonini Y, Martins RP. 2003. Yeast communitiesassociated with stingless bees. FEMS Yeast Res 4:271–275.
Schowalter DT. 2000. Insect Ecology: an ecosystem ap-proach. San Diego: Academic Press. 496 p.
Siemann E, Tilman D, Haarstad J. 1996. Insect speciesdiversity, abundance and body size relationships.Nature 380:704–706.
Sniegowski PD, Dombrowski PG, Fingerman E. 2002.Saccharomyces cerevisiae and Saccharomyces paradoxuscoexist in a natural woodland site in North Americaand display different levels of reproductive isolationfrom European conspecifics. FEMS Yeast Res 1:299–306.
Stamopoulos DC, Tzanetakis NM. 1988. Bacterial floraisolated from the oesophageal bulb of the olive fruitfly Dacus oleae (Gmelin). Entomol Hell 6:43–48.
Stewart KW, Friday GP, Rhame RE. 1973. Food habits ofhellgrammite larvae, Corydalus cornutus (Megaloptera:Corydalidae), in the Brazos River, Texas. Ann EntomolSoc Am 66:959–963.
Stratford M, Bond CJ, James SA, Roberts N, Steels H. 2002.Candida davenportii sp. nov., a potential soft-drinksspoilage yeast isolated from a wasp. Int J Syst EvolMicrobiol 52:1369–1375.
Suh S-O, Blackwell M. 2005. The beetle gut as a habitat fornew species of yeasts. In: Vega FE, Blackwell M, eds.Insect Fungal Associations: ecology and evolution. NewYork: Oxford University Press. p 244–256.
———, ———. 2004. Three new beetle-associated yeasts inthe Pichia guilliermondii clade. FEMS Yeast Res 5:87–95.
———, Gibson CM, Blackwell M. 2004a. Metschnikowiachrysoperlae sp. nov., Candida picachoensis sp. nov. andCandida pimensis sp. nov., isolated from the greenlacewings Chrysoperla comanche and Chrysoperla carnea(Neuroptera: Chrysopidae). Int J Syst Evol Microbiol54:1883–1890.
———, Marshall CJ, McHugh JV, Blackwell M. 2003. Woodingestion by passalid beetles in the presence of xylose-fermenting gut yeasts. Mol Ecol 12:3137–3145.
———, McHugh JV, Blackwell M. 2004b. Expansion of theCandida tanzawaensis yeast clade: 16 new Candidaspecies from basidiocarp-feeding beetles. Int J Syst EvolMicrobiol 54:2409–2429.
———, ———, Pollock DD, Blackwell M. 2005. The beetlegut: a hyperdiverse source of novel yeasts. Mycol Res109:261–265.
———, Nguyen NH, Blackwell M. 2006. A yeast clade nearCandida kruisii uncovered: nine novel Candida speciesassociated with basidioma-feeding beetles. Mycol Res110:1379–1394.
Swofford DL. 2002. PAUP*: phylogenetic analysis usingparsimony (*and other methods), version 4.0b10.Sunderland, Massachusetts: Sinauer Associates.
Tang Y, Ward RD. 1998. Sugar feeding and fluid destination
NGUYEN ET AL: YEASTS FROM NEUROPTERANS 857

control in the phlebotomine sandfly Lutzomyia longipalpis(Diptera: Psychodidae). Med Vet Entomol 12:13–19.
Tatusova TA, Madden TL. 1999. BLAST 2 sequences, a newtool for comparing protein and nucleotide sequences.FEMS Microbiol Lett 174:247–250.
Tauber CA. 1991. Order Neuroptera. In: Stehr FW, ed.Immature Insects. Vol. II. Iowa: Kendall/Hunt. p 126–143.
Teixeira AC, Marini MM, Nicoli JR, Antonini Y, Martins RP,Lachance MA, Rosa CA. 2003. Starmerella meliponi-norum sp. nov., a novel ascomycetous yeast speciesassociated with stingless bees. Int J Syst Evol Microbiol53:339–343.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, HigginsDG. 1997. The Clustal X Windows interface: flexiblestrategies for multiple sequence alignment aided byquality analysis tool. Nucleic Acids Res 25:4876–4882.
Vega FE, Dowd PF. 2005. The role of yeasts as insect
endosymbionts. In: Vega FE, Blackwell M, eds. Insect-Fungal Associations: ecology and evolution. New York:Oxford University Press. p 211–243.
Werren JH. 1997. Biology of Wolbachia. Ann Rev Entomol42:587–609.
Woolfolk SW, Inglis GD. 2003. Microorganisms associatedwith field-collected Chrysoperla rufilabris (Neuroptera:Chrysopidae) adults with emphasis on yeast symbionts.Biol Control 29:155–168.
———, Cohen AC, Inglis GD. 2004. Morphology of thealimentary canal of Chrysoperla rufilabris (Neuroptera:Chrysopidae) adults in relation to microbial symbionts.Ann Entomol Soc Am 97:796–808.
Yarrow D. 1998. Methods for the isolation, maintenance andidentification of yeasts. In: Kurtzman CP, Fell JW, eds.The yeasts, a taxonomic study. 4th ed. Amsterdam:Elsevier. p 77–100.
858 MYCOLOGIA