five gonadotrophin-releasing hormone receptors in a teleost fish: isolation, tissue distribution and...
TRANSCRIPT

Five gonadotrophin-releasing hormone receptors in a teleostfish: isolation, tissue distribution and phylogeneticrelationships
Natalia Moncaut, Gustavo Somoza1, Deborah M Power and Adelino V M CanárioCentre of Marine Sciences, University of Algarve, Campus de Gambelas, 8005-139 Faro, Portugal
1Laboratorio de Ictiofisiologia y Acuicultura, Instituto de Investigaciones Biotecnologicas-Instituto Tecnologico de Chascomus (IIB-INTECH), Camino de Circunvalacion LagunaKm 6 (B7130IWA), Chascomus, Provincia de Buenos Aires, Argentina.
(Requests for offprints should be addressed to A V M Canário; Email: [email protected] )
Abstract
Gonadotrophin-releasing hormone (GnRH) is the main neurohormone controlling gonadotrophin release in allvertebrates, and in teleost fish also of growth hormone and possibly of other adenohypophyseal hormones. Over 20GnRHs have been identified in vertebrates and protochoordates and shown to bind cognate G-protein couplereceptors (GnRHR). We have searched the puffer fish, Fugu rubripes, genome sequencing database, identified fiveGnRHR genes and proceeded to isolate the corresponding complementary DNAs in European sea bass, Dicentrachuslabrax. Phylogenetic analysis clusters the European sea bass, puffer fish and all other vertebrate receptors into twomain lineages corresponding to the mammalian type I and II receptors. The fish receptors could be subdivided in twoGnRHR1 (A and B) and three GnRHR2 (A, B and C) subtypes. Amino acid sequence identity within receptor subtypesvaries between 70 and 90% but only 50–55% among the two main lineages in fish. All European sea bass receptormRNAs are expressed in the anterior and mid brain, and all but one are expressed in the pituitary gland. There isdifferential expression of the receptors in peripheral tissues related to reproduction (gonads), chemical senses (eyeand olfactory epithelium) and osmoregulation (kidney and gill). This is the first report showing five GnRH receptors ina vertebrate species and the gene expression patterns support the concept that GnRH and GnRHRs play highlydiverse functional roles in the regulation of cellular functions, besides the ‘‘classical’’ role of pituitary function regulation.
Journal of Molecular Endocrinology (2005) 34, 767–779
Introduction
Gonadotrophin-releasing hormone (GnRH) is a deca-peptide synthesized in the preoptic-hypothalamic areaand acts on the pituitary gland where it stimulates thesynthesis and release of gonadotrophic hormones in allvertebrates (Millar et al. 2004), and growth hormone andprolactin in at least some fish species (Marchant et al.1989, Weber et al. 1997). In addition to the preoptic-hypothalamic GnRH, one or two additional GnRHforms are synthesized in extra preoptic-hypothalamicareas within the central nervous system (Lethimonieret al. 2004, Millar et al. 2004). The number of knownGnRH family members has now reached a total of 24,usually named after the species where they were firstisolated (Adams et al. 2003, Millar et al. 2004, Somozaet al. 2002).
GnRHs exert their actions through GnRH receptors(GnRHRs) which are members of the rhodopsin-likeG-protein coupled receptor (GPCR) family. Thesereceptors show three main functional domains: 1) aN-terminal extracellular domain; 2) seven �-helicaltransmembrane (TMs) domains connected by hydro-
philic intra- and extracellular loops and 3) a C-terminalcytoplasmic domain. The extracellular domains andsuperficial regions of the TMs are usually involved inbinding. The TMs are believed to be involved inreceptor configuration. The C-terminal mediates effec-tors binding, propagation of signalling events, desensi-tization and internalization (McArdle et al., 2002, Millaret al., 2004). During the last decade several cDNAs forGnRHR have been cloned, often more than one in thesame species, e.g. three in bullfrog, Rana catesbiana (Wanget al. 2001) and medaka, Oryzias latipes (Okubo et al. 2001,2003) and two in primates (Millar et al. 2001, Neill et al.2001), goldfish, Carassius auratus (Illing et al. 1999) andAfrican catfish, Claria gariepinus (Bogerd et al. 2002). Asa result GnRH receptors have recently been groupedin three main subtypes designated GnRHR1, GnRHR2and GnRHR3, the latter only found in teleost fishes andamphibians (Millar et al. 2004). In addition, duplicatedgenes within these subtypes have been also describedin teleosts, e.g. Oncorhynchus masou (Jodo et al. 2003).The presence of multiple GnRH ligands for severalreceptors suggests complex regulatory mechanisms andraises the question of whether new functions, more
767
Journal of Molecular Endocrinology (2005) 34, 767–7790952–5041/05/034–767 © 2005 Society for Endocrinology Printed in Great Britain
DOI: 10.1677/jme.1.01757Online version via http://www.endocrinology-journals.org

specialization, or both, have been acquired by theduplicated genes.
Data mining of model fish genome sequences hasfacilitated the identification of novel genes and in thispaper we have used the GnRHRs DNA sequencesidentified in the Fugu rubripes genome database to isolatethe corresponding cDNAs in the European sea bass,Dicentrarchus labrax. Like most perciform species, theEuropean sea bass synthesizes three GnRH ligands:sGnRH, sbGnRH and cGnRHII (González-Martínezet al. 2001, 2002, Zmora et al., 2002). However, untilnow only a single GnRHR cDNA from European seabass has been cloned and its expression profile in thebrain and pituitary suggests it may mediate gonado-trophin release (González-Martínez et al. 2004). Inthe current study we report the cloning of four addi-tional European sea bass cDNAs encoding GnRHRs(dlGnRHR), examine their tissue distribution andreassess their classification based on the phylogeneticanalysis of available sequences.
Materials and methods
Animals and tissue sampling
Sexually mature males and females from Europeansea bass were obtained from TIMAR Cultura deÁguas (Livramento, Portugal) and maintained at theRamalhete Experimental Station (University of Algarve,Faro, Portugal) prior to sampling, in through-flowseawater tanks at 17 �2 �C under natural photoperiod.Animal care was in accordance with the ethicalguidelines of the Animal Behaviour Society (ASAB 2003)and national legislation. Fish were killed with anoverdose of 2-phenoxyethanol (1:10000, Sigma–Aldrich,Madrid, Spain), tissues were dissected and immediatelyfrozen in liquid nitrogen and stored at -80 �C until used.
In silico analysis and cloning of dlGnRHRs
The puffer fish (Fugu rubripes) Fugu Consortium GenomeDatabase (http://fugu.hgmp.mrc.ac.uk) was interro-gated with the amino acid sequences of available fishGnRH receptors via the tBLASTn programme using theBlosum62 matrix and an expected value of 10 (Altschulet al. 1997). Scaffolds identified to contain GnRHRsequences were run on the HGMP Nix interface (GWilliams, P Woollard & P Hingamp, unpublished data;http://www.hgmp.mrc.ac.uk/NIX/) providing, as out-put, putative gene organization, coding sequence, aminoacid translation and neighbouring genes. Five differentloci were found with putative open reading frames(ORFs) for GnRHR located in scaffolds FS2243, FS553,FS686, FS3910 and FS1435. The putative codingsequences of Fugu genes together with other availablefish sequences were used to design five primer pairs to
amplify the corresponding cDNAs in European sea bass(Table 1) and the amino acid translations were used inthe phylogenetic analysis.
Total RNA was extracted from brain and pituitary ofEuropean sea bass using TRI Reagent (Sigma-Aldrich)according to the manufacturer’s protocol. Five micro-grams of total RNA was retro transcribed using Moloneymurine leukaemia virus (MMLV-RT; Invitrogen,Carlsbad, CA, USA) and oligo(dT)12–18 in a final volumeof 30 µl for 1·5 h at 37 �C. Each PCR was carried outusing 1–2µl cDNA, 2·5 U Taq polymerase (Promega,Biocontech, Lisbon, Portugal) and 1 pmol/µl of eachdifferent pair of primers. Thermocycling parameterswere the same for all primer pairs, except for theannealing temperatures: 4 min at 94 �C, 35 cycles of1 min at 94 �C, 1 min 50–56 �C and 1 min at 72 �C,with 5 min extension at 72 �C. PCR products werevisualized in 1% agarose gel stained with ethidiumbromide and bands of expected size purified using GFXPCR and Gel Band Purification Kit (AmershamBiosciences, Buckinghamshire, UK) and cloned usingthe pGEM-T easy Vector Systems (Promega). Thecloned inserts were used to transform E. coli XL1-Blueand positive colonies were isolated and plasmid DNAextracted using the alkaline lyses method and sequenced(Sistemas Genómicos, Valencia, Spain; Macrogen,Seoul, South Korea).
Construction and screening of cDNA libraries
cDNA libraries from European sea bass brain (pituitaryincluded) and testis were constructed using UNI-ZAPXR Vector (Strategene, Amsterdam, The Netherlands)with reverse-transcribed cDNA obtained from 4–5 µgof poly(A)+ RNA and using UNI-ZAP XR cDNAsynthesis kit (Stratagene) according to the supplier’sinstructions. Library screening was carried out underhigh stringency conditions using the five specificdlGnRHR probes obtained by PCR amplification.Duplicate membranes (Hybond NX- Amersham Bio-sciences, Buckinghamshire, UK) were hybridized witheach of the [�32P]dCTP-labelled dlGnRHR probes(Rediprime Random Labelling kit, Amersham Bio-sciences) overnight at 56 �C in a solution containingEDTA 1 mM, SDS 7% and sodium phosphate 0·25 M(Church–Gilbert hybridization solution; Church &Gilbert 1984). Stringency washes were carried out at56 �C for 5 minutes with 2�SSC, 1�SSC (150 mMNaCl, 15 mM sodium citrate, pH 7·0) and 0·1�SSCcontaining 0·1% SDS.
Tissue distribution of GnRHRs by RT-PCR andSouthern blotting
The distribution pattern of dlGnRHRs expression wasinvestigated by RT-PCR on anterior brain (olfactory
N MONCAUT and others · Novel fish GnRH receptors768
www.endocrinology-journals.orgJournal of Molecular Endocrinology (2005) 34, 767–779

Tab
le1
Prim
erpa
irsus
edfo
rin
itial
PC
Rcl
onin
gof
dlG
nRH
Rs
and
for
RT-
PC
R
Init
ial
clo
nin
gP
rod
uct
size
RT-
PC
RP
rod
uct
size
Gen
edl
GnR
HR
-1A
dlgn
rhR
-F:5
8CG
TG
AT
GC
CT
GT
GG
AT
GC
3834
2dl
gnrh
R-F
342
dlgn
rhR
-R:5
8TT
GTA
GG
CC
GT
TT
C(G
/C)T
(G/C
)CC
A38
dlgn
rhR
-Rdl
GnR
HR
-1B
1gnr
hF2:5
8CC
GC
TG
GC
AG
GA
AA
CT
GT
3818
21g
nrhF
218
21g
nrhR
2:5
8TC
CG
AG
CC
TT
TG
GG
AT
GA
381g
nrhR
2
dlG
nRH
R-2
Adl
gnrh
R-F
:58C
GT
GA
TG
CC
TG
TG
GA
TG
CC
3841
1dl
gnrh
R-F
586
dlgn
rhR
-R:5
8TT
GTA
GG
CC
GT
TT
C(G
/C)T
(G/C
)CC
A38
3gnr
hR-R
:58C
CA
CA
GT
CC
CA
GC
AG
GTA
G38
dlG
nRH
R-2
B4g
nrhR
-F2:5
8CC
CG
AG
AA
CT
TC
AC
CC
AG
3840
64g
nrhR
-F2
490
3gnr
hR-R
:58C
CA
CA
GT
CC
CA
GC
AG
GTA
G38
sqR
1Bre
v:58
TC
TAA
TT
CA
GT
CC
GC
GT
GT
TT
G38
dlG
nRH
R-2
C2g
nrhR
-F:5
8TC
AC
GG
TAC
AG
TG
GT
TG
GC
3835
3R
1Csq
fw3:
58G
CT
GT
GC
TG
TG
GG
CA
GC
CA
AT
G38
587
2gnr
hR-R
:58T
GA
GC
GG
CA
GA
AG
GA
AG
AG
38R
1Csq
rev:
58T
TG
AG
CA
AC
GG
AG
AT
GTA
G38
Novel fish GnRH receptors · N MONCAUT and others 769
www.endocrinology-journals.org Journal of Molecular Endocrinology (2005) 34, 767–779

bulb and telencephalon), middle brain (optic tectum anddiencephalons), posterior brain (cerebellum, medullaoblongata and part of the spinal cord), pituitary gland,eye, olfactory epithelium, gonads, kidney, head kidney,spleen, liver, heart, midgut, gills and muscle. cDNA(40 µl) was prepared from 5 µg total RNA extracted withTRI Reagent and reverse transcribed with MMLV-RTand random hexamers. PCR was performed on 1 µl ofcDNA with specific primers (Table 1) designed to spantwo exon–intron boundaries. Only single RTP-PCRproducts of the size of the expected cDNA wereobtained, indicating the absence of genomic DNAcontamination. Thermocycling conditions were opti-mized for each primer to be on the linear region ofamplification and were 4 min at 94 �C, 27–30 cycles of45 sec at 94 �C, 45 sec at 56–59 �C and 45 sec extensionat 72 �C, with a final 5 min extension at 72 �C. A 540 bpfragment of European sea bass 18S was amplified usingoligonucleotides 18S-fw 5�TCA AGA ACG AAA GTCGGA GG 3� and 18S-rev 5�GGA CAT CTA AGGGCA TCA CA 3� (3 min at 94 �C, 16 cycles of 45 sec at94 �C, 45 sec at 48 �C and 45 sec at 72 �C, followed by5 min extension at 72 �C) and used as an internal controlfor the amount of cDNA per reaction. Each PCRproduct was transferred onto a nylon membrane(Hybond N; Amersham Biosciences) and hybridizedovernight at 64 �C with Church–Gilbert hybridizationsolution with the corresponding probe labelled with[�32P]dCTP by random priming (RediPrime RandomLabelling kit, Amersham Biosciences). Stringency washeswere carried out at 64 �C with 2�SSC, 1�SSC and0·1�SSC containing 0·1% SDS.
Sequence comparison and phylogenetic analysis
The following sequences with accession numbers andabbreviations were used: buffalo, Bubalus bubalis (Bb,CAF21711), cow, Bos taurus, (Bt, NP803480), sheep, Ovisaries (Oa, CAA50978), human, Homo sapiens (R1,AAA35917; R2, Q96P88), Rhesus macaque, Macacamulata (Mm2, AAK52745), Bonnet macaque, Macacaradiata (Mr, AAG43378), green monkey, Aethiops sabeus(As, AAK52746), marmoset, Callitrix jacchus (CjII,AAK60927), horse, Equus caballus (Ec, O18821), dog,Canis canis (Cc, Q9 MZI6), Norwegian rat, Rattusnorvegicus (Rn, NP112300), mouse, Mus musculus (Mm,Q01776), Guinea pig, Cavia porcellus (Cp, Q8CH60),Chicken, Gallus gallus (Gg, NP989984), leopard gecko,Eublepharis macularius (Em, BAD11150), bullfrog, Ranacatesbeiana (RcI AAG42575, RcII AAG42949, RcIIIAAG42574), frog, Rana ridibunda (Rr1 AAP15162, Rr2AAP15163, Rr3 AAP15164), brown frog, Rana dybowskii(Rd1 AAO50198, Rd2 AAO50196, Rd3 AAO50197),African clawed frog, Xenopus laevis, (Xl1 AAF89754,Xl2 AAK49334), rubber eel, Typhlonectes natans (Tn,AAD49750), stripped bass, Morone saxatilis (Ms,
AAF28464), amberjack, Seriola dumerilii (Sd, CAB65407),sea bream, Sparus auratus (Sa, AAS97968), Nile tilapia,Oreochromis niloticus (On1 BAC77241, On2 BAC77240,On3 BAD27389), medaka, Oryzias latipes (Ol1BAB70506, Ol2 BAB70505, Ol3 BAC97833), Africancatfish, Clarias gariepinus (Cg1, BAC97836, Cg2,AAM95605), goldfish, Carassius auratus (CaA,AAD20001, CaB AAD20002), masou salmon, Oncorhyn-chus masou (Oma1, BAC98943, Oma4, BAC98946),rainbow trout, Oncorhynchus mykiss (Om, CAB93351),Japanese eel, Anguilla japonica (Aj, BAB11961).
Multisequence alignments of the full-length proteinswere carried out using CLUSTAL X (Thompson et al.1997). Protein sequence similarities between differentforms of GnRHR and between species were calculatedwith GeneDoc (Nicholas et al. 1997) and protein motifswere identified using the Prosite database (Falquet et al.2002). The putative transmembrane domains (TMs)were determined with TMHMM 2·0 (Krogh et al. 2001).Phylogenetic data analysis was restricted to TMs whichwere extracted from all identified genes (TMs 1, 2, 3, 4,5, 6 and 7) and aligned with Clustal X. Parsimoniousconsensus trees of the TMs of all available GnRHR weregenerated using PAUP software (version 4·0, Swofford2002) with the output of Clustal X alignments. Bootstrapvalues were calculated with 1000 replicates to estimatethe robustness of internal branches.
Results
Molecular cloning of dlGnRHRs
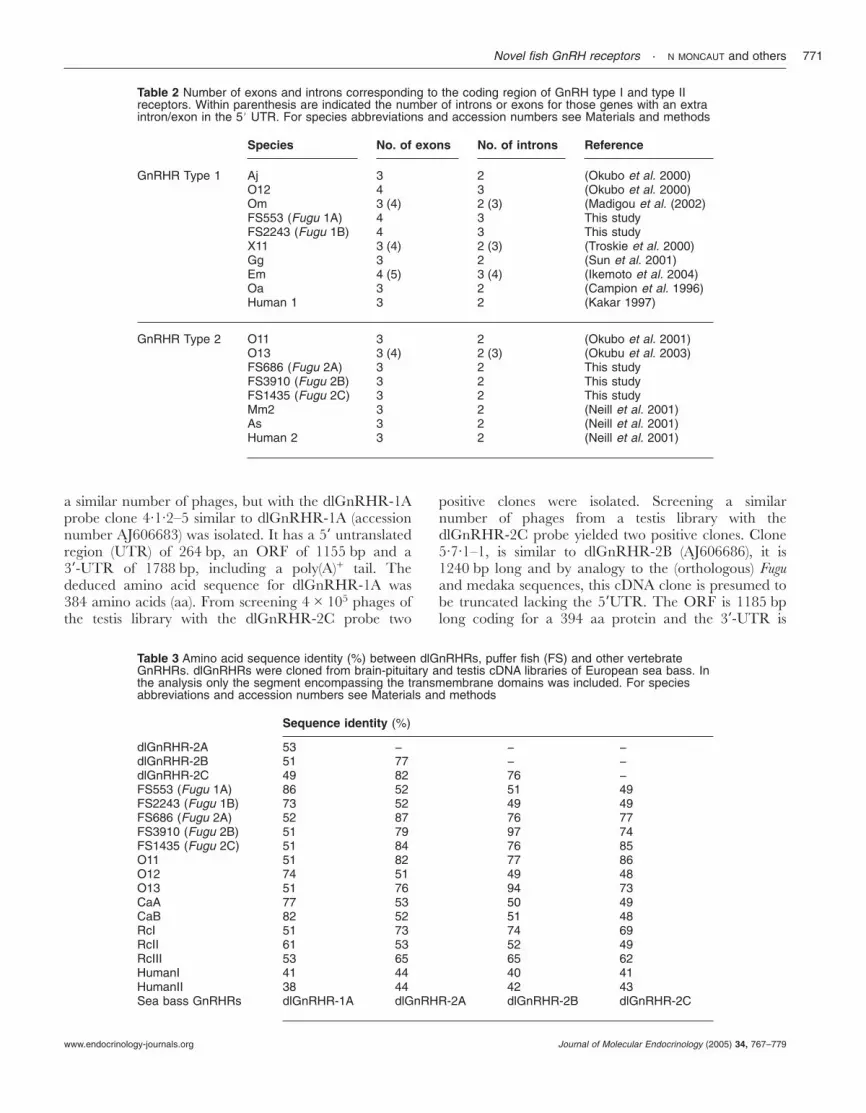
Five different loci with putative open reading frames(ORFs) for GnRHR were identified in scaffolds FS2243,FS553, FS686, FS3910 and FS1435 of the sequencedgenome of the tetraodontiform Fugu rubripes. Analysis ofthe putative GnRHR gene structure of FS2243 andFS553 showed that their coding regions encompassedfour exons and three introns while for FS686, FS3910and FS1435 there were three exons and two introns(Table 2).
RT-PCR using specific primers for each of the fiveGnRH identified in Fugu and the brain and pituitary ofEuropean sea bass led to the amplification of fivedifferent cDNAs showing high sequence homology to thecorresponding Fugu GnRHR sequences (Table 3). Theamplified GnRHRs have been classified as dlGnRHR-1A, -1B, -2A, -2B and -2C on the basis of phylogeneticanalysis (see below). The partial fragments of dlGnRHRisolated by RT-PCR were used as probes to screenbrain-pituitary and testis cDNA libraries in order toisolate the full length receptors. Screening of 4�105
pituitary plus brain phages with the dlGnRHR-2B probeled to the isolation of clone 3·1·2–1 corresponding todlGnRHR-2A (accession number AJ419594 alreadycharacterized, see González-Martínez et al. 2004). From
N MONCAUT and others · Novel fish GnRH receptors770
www.endocrinology-journals.orgJournal of Molecular Endocrinology (2005) 34, 767–779
a similar number of phages, but with the dlGnRHR-1Aprobe clone 4·1·2–5 similar to dlGnRHR-1A (accessionnumber AJ606683) was isolated. It has a 5� untranslatedregion (UTR) of 264 bp, an ORF of 1155 bp and a3�-UTR of 1788 bp, including a poly(A)+ tail. Thededuced amino acid sequence for dlGnRHR-1A was384 amino acids (aa). From screening 4�105 phages ofthe testis library with the dlGnRHR-2C probe two
positive clones were isolated. Screening a similarnumber of phages from a testis library with thedlGnRHR-2C probe yielded two positive clones. Clone5·7·1–1, is similar to dlGnRHR-2B (AJ606686), it is1240 bp long and by analogy to the (orthologous) Fuguand medaka sequences, this cDNA clone is presumed tobe truncated lacking the 5�UTR. The ORF is 1185 bplong coding for a 394 aa protein and the 3�-UTR is
Table 2 Number of exons and introns corresponding to the coding region of GnRH type I and type IIreceptors. Within parenthesis are indicated the number of introns or exons for those genes with an extraintron/exon in the 58 UTR. For species abbreviations and accession numbers see Materials and methods
Species No. of exons No. of introns Reference
GnRHR Type 1 Aj 3 2 (Okubo et al. 2000)O12 4 3 (Okubo et al. 2000)Om 3 (4) 2 (3) (Madigou et al. (2002)FS553 (Fugu 1A) 4 3 This studyFS2243 (Fugu 1B) 4 3 This studyX11 3 (4) 2 (3) (Troskie et al. 2000)Gg 3 2 (Sun et al. 2001)Em 4 (5) 3 (4) (Ikemoto et al. 2004)Oa 3 2 (Campion et al. 1996)Human 1 3 2 (Kakar 1997)
GnRHR Type 2 O11 3 2 (Okubo et al. 2001)O13 3 (4) 2 (3) (Okubu et al. 2003)FS686 (Fugu 2A) 3 2 This studyFS3910 (Fugu 2B) 3 2 This studyFS1435 (Fugu 2C) 3 2 This studyMm2 3 2 (Neill et al. 2001)As 3 2 (Neill et al. 2001)Human 2 3 2 (Neill et al. 2001)
Table 3 Amino acid sequence identity (%) between dlGnRHRs, puffer fish (FS) and other vertebrateGnRHRs. dlGnRHRs were cloned from brain-pituitary and testis cDNA libraries of European sea bass. Inthe analysis only the segment encompassing the transmembrane domains was included. For speciesabbreviations and accession numbers see Materials and methods
Sequence identity (%)
dlGnRHR-2A 53 − − −dlGnRHR-2B 51 77 − −dlGnRHR-2C 49 82 76 −FS553 (Fugu 1A) 86 52 51 49FS2243 (Fugu 1B) 73 52 49 49FS686 (Fugu 2A) 52 87 76 77FS3910 (Fugu 2B) 51 79 97 74FS1435 (Fugu 2C) 51 84 76 85O11 51 82 77 86O12 74 51 49 48O13 51 76 94 73CaA 77 53 50 49CaB 82 52 51 48RcI 51 73 74 69RcII 61 53 52 49RcIII 53 65 65 62HumanI 41 44 40 41HumanII 38 44 42 43Sea bass GnRHRs dlGnRHR-1A dlGnRHR-2A dlGnRHR-2B dlGnRHR-2C
Novel fish GnRH receptors · N MONCAUT and others 771
www.endocrinology-journals.org Journal of Molecular Endocrinology (2005) 34, 767–779

55 bp. Clone 3·2·1–1 (dlGnRHR-2C) is 2677 bp longwith a 282 bp 5�-UTR and an 1138 bp 3�-UTR, andwith a translated protein of 418 amino acids. Until now,only a partial fragment of dlGnRHR-1B (AJ606685) hasbeen cloned (Fig. 1). Hybridization of sea bass genomicDNA in a Southern blot using probes specific for eachreceptor form resulted in a characteristic pattern ofbands, thus confirming that they are encoded bydifferent genes.
Comparison of the different dlGnRHRs and othervertebrates/teleost
A multisequence alignment of the amino acid sequencesof dlGnRHRs with that of other vertebrates revealed thepresence of conserved features of the class A, rhodopsin-like family of G-protein coupled receptor superfamily,namely the seven transmembrane domains and the N-and C-terminal regions (Fig. 1). Amino acid sequenceidentity among the dlGnRHRs, including the trans-membrane domains but excluding the highly variable N-and C-terminal tails, ranged between 49% and 82%(dlGnRHR-1B not included, see Table 3). Across thevertebrates, sequence similarities between the Europeansea bass receptors ranged between 38% with humanGnRHR2 and 97% with Fugu.
A phylogenetic analysis of the GnRHR transmem-brane domains shows that there are two main receptorlineages encompassing all vertebrates (designated type Iand type II, Fig. 2). Within taxa, gene duplicationsappear to have occurred in the amphibians and inteleost fishes. Considering that each of the clades whichinclude a European sea bass and a Fugu GnRHR isencoded by a separate gene, the lineage which includesthe human GnRHR type 1 appears to contain at leastone teleost specific gene duplication encompassing theCyprinidiformes, Salmoniformes and the modern teleostfishes (Perciformes, Atherinidiformes and Tetraodon-tiformes; Fig. 2). This results in two teleost subtypes.However, the Japanese eel receptor did not fallunambiguously in either of the two teleost subtypes. Inthe lineage which includes human GnRHR2, threeteleost clades can be identified and their tree topologysuggest two gene duplications, with one possibly specificto the modern teleosts. Two clades within this lineageare also present in the amphibians suggesting aduplication event also occurred in this class.
Analysis of the primary structure of the translatedproteins grouped according to the the phylogeneticanalysis showed, as main features, the presence of aC-terminal tail in all receptors except mammalian type I(Fig. 1). These have the shortest protein sequence with328 to 362 aa. In ascending order of size the protein is368–375 aa long from amphibians to birds, 379–390 aain fish subtype 1A, and 350–375 aa in fish subtype 1B(considering only the second Met in rainbow trout), and
380 aa in Japanese eel. In type II receptors theC-terminal tail is longer in all cases in comparison totype I for the same class of animals. The mammalianreceptor ranges from 379–380 aa (excluding humanwhich lacks the N-terminal region). One of theamphibian subtypes has 407 aa in all species studiedand the second 410–424 aa. The fish subtype 2A rangesfrom 412–426 aa (Nile tilapia excluded because itprobably lacks part of the C-terminus). The fish subtype2B from 390–394 aa and the fish subtype C from387–418 aa.
A comparative analysis of conservation of amino acidsthat have been implicated in receptor function withreference to the human type I receptor (Millar et al.2004) show that not all are fully conserved. For thoseamino acids important for the structure of the receptoror of the binding pocket Cys114, Trp164, Cys196, Trp206,Trp280, Trp291 and Pro320 are fully conserved through-out the vertebrates. None of the amino acids implicatedin G-protein coupling in mammalian receptors areconserved in the fish receptors. Amino acids Asp98,Trp101, Asn102, Lys121 and Asn212 implicated in ligandbinding are fully conserved but not Tyr290 (modifiedonly in Nile tilapia) and Asp302 (conserved only in somemammals). Of the amino acids implicated in receptoractivation only Asn87 is not conserved.
Tissue mRNA expression of dlGnRHRs
All dlGnRHRs have a common pattern of medium tohigh abundance in the anterior and middle brain in thetwo females and two males that were analysed (Fig. 3).Of the two GnRHR types (1 and 2), type 1 has thewidest distribution among non reproductive tissues,while type 2 is more restricted to the central nervoussystem. dlGnRHR-1A transcripts are also relativelyabundant in the pituitary, olfactory epithelium, ovaryand eye. Lower levels of expression are detected in theposterior brain region, gills and kidney. Besides itsexpression in brain, olfactory epithelium and kidney,dlGnRHR-1B can be detected in the pituitary, eyes,gills, gut and liver. dlGnRHR-2A has higher expressionin the anterior and midbrain, pituitary and in both ovaryand testis (not shown). Faint expression is found in theolfactory epithelium, eyes and gills. dlGnRHR-2B ismainly expressed along the central nervous system andgonads. Surprisingly, this receptor has no expression inthe pituitary gland. dlGnRHR-2C is also expressed inthe three brain regions, and in the pituitary, eyes andtestis (not shown). Faint expression is found in the ovaryand head kidney.
Discussion
In this study we have used sequence information fromthe model fish Fugu rubripes to isolate five cDNAs for
N MONCAUT and others · Novel fish GnRH receptors772
www.endocrinology-journals.orgJournal of Molecular Endocrinology (2005) 34, 767–779

Figure 1 Multisequence alignment of deduced amino acid sequences of GnRHRs from Europeansea bass, Fugu (FS) and representatives from other fish orders and vertebrate classes. Theputative seven transmembrane domains are indicated. For species abbreviations and accessionnumbers see the Materials and methods section.
Novel fish GnRH receptors · N MONCAUT and others 773
www.endocrinology-journals.org Journal of Molecular Endocrinology (2005) 34, 767–779
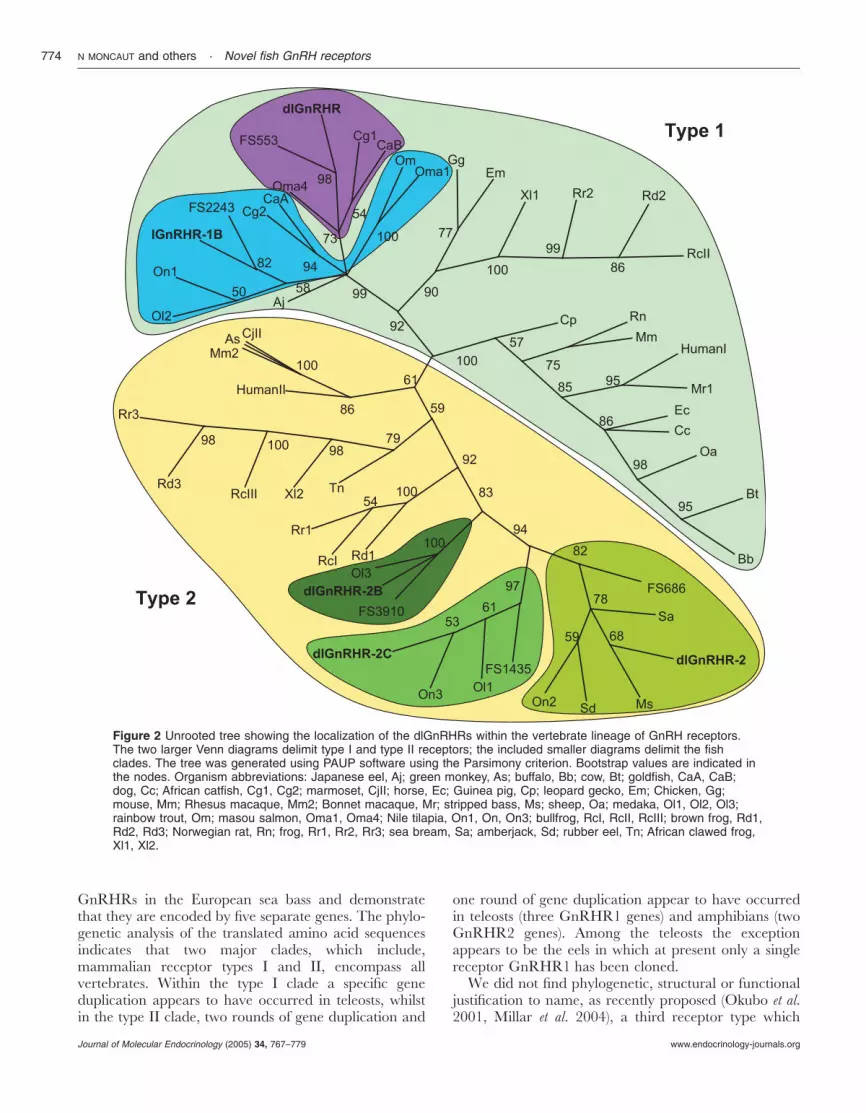
GnRHRs in the European sea bass and demonstratethat they are encoded by five separate genes. The phylo-genetic analysis of the translated amino acid sequencesindicates that two major clades, which include,mammalian receptor types I and II, encompass allvertebrates. Within the type I clade a specific geneduplication appears to have occurred in teleosts, whilstin the type II clade, two rounds of gene duplication and
one round of gene duplication appear to have occurredin teleosts (three GnRHR1 genes) and amphibians (twoGnRHR2 genes). Among the teleosts the exceptionappears to be the eels in which at present only a singlereceptor GnRHR1 has been cloned.
We did not find phylogenetic, structural or functionaljustification to name, as recently proposed (Okubo et al.2001, Millar et al. 2004), a third receptor type which
Figure 2 Unrooted tree showing the localization of the dlGnRHRs within the vertebrate lineage of GnRH receptors.The two larger Venn diagrams delimit type I and type II receptors; the included smaller diagrams delimit the fishclades. The tree was generated using PAUP software using the Parsimony criterion. Bootstrap values are indicated inthe nodes. Organism abbreviations: Japanese eel, Aj; green monkey, As; buffalo, Bb; cow, Bt; goldfish, CaA, CaB;dog, Cc; African catfish, Cg1, Cg2; marmoset, CjII; horse, Ec; Guinea pig, Cp; leopard gecko, Em; Chicken, Gg;mouse, Mm; Rhesus macaque, Mm2; Bonnet macaque, Mr; stripped bass, Ms; sheep, Oa; medaka, Ol1, Ol2, Ol3;rainbow trout, Om; masou salmon, Oma1, Oma4; Nile tilapia, On1, On, On3; bullfrog, RcI, RcII, RcIII; brown frog, Rd1,Rd2, Rd3; Norwegian rat, Rn; frog, Rr1, Rr2, Rr3; sea bream, Sa; amberjack, Sd; rubber eel, Tn; African clawed frog,Xl1, Xl2.
N MONCAUT and others · Novel fish GnRH receptors774
www.endocrinology-journals.orgJournal of Molecular Endocrinology (2005) 34, 767–779

would split mammalian type II and the closestamphibian clade from the other amphibian and,following our designation, fish type II receptors. If athird receptor type were designated the resultingproposed clades would split through a phylogeneticdivision separating higher from lower vertebrates.Analysis of gene organization corresponding to thecoding region of the cDNAs (Table 2) shows that in bothGnRHR1 and GnRHR2 types, genes with three exonsand two introns and genes with four exons and threeintrons (four and five, respectively in gecko) are present,the latter in some but not all fishes (Madigou et al. 2000,Okubo et al. 2000, 2001, 2003) and Xenopus (Troskie et al.2000). In the genes with more than three exons, the 1st
exon appears to have acquired an intron (Okubo et al.2003). Overall, this indicates that gene organizationper se is not diagnostic to classify GnRH receptor types.The suggestion of three receptor types has been partlyfuelled by the fact that ligand evolution appears to haveled to three main clades, one of which consisting onlyof teleost fish (Millar et al. 2004). However, despiteextensive studies of different GPCR families and theirreceptors there is so far no clear evidence supportingthe notion that ligand evolution coincides with receptorevolution. Furthermore, similar observations of fishspecific duplications have occurred for other genes infish, including nuclear receptors, developmental genes,enzymes, etc. (Amores et al. 1998, Tchoudakova &
Callard 1998, Robinson-Rechavi et al. 2001). It appearsthat in fishes a specific round of genome duplication withgene loss has occurred (Jaillon et al. 2004) and it ispossible that segmentary genomic duplications may haveresulted in the larger number of GnRHRs, at least insome fish species. Recent segmentary genome duplica-tions have also been proposed to have occurred inhuman (5–6% euchromatin; She et al. 2004) and mousegenomes (2%; Bailey et al. 2004). The most parsimoniousconclusion is therefore that in vertebrates the GnRHreceptor family is composed of two receptor types andthat specific duplications have occurred early in theevolution of amphibians and ray-finned fishes (possiblymore than once). In the following discussion GnRHRnomenclature in fish will be adapted to the Europeansea bass notation which is based on the topology of thephylogenetic tree (Fig. 2).
Amino acid sequence comparisons of the TMs of theGnRHRs extracted from the Fugu rubripes genome andfrom the sea bass cDNAs had highest similarities ofbetween 77% and over 80% between receptorsbelonging to the same major clade (type I or type II)while across clades sequence similarity was low, between51 and 55%.
The analyses of amino acids that have beenimplicated in receptor function show that the highestconservation is present in those associated with ligandbinding. An exception is Asp302 (numbering in reference
Figure 3 Expression of dlGnRHRs in central and peripheral tissues detected by RT-PCR andSouthern blotting. Amplification of rRNA 18s was used to standardize the quantity of mRNA. M,muscle; G, gills; L, liver; Gt, gut; H, heart; S, spleen; K, kidney; HK, head kidney; O, ovary; AB,anterior brain; MB, middle brain; PB, post brain; P, pituitary; E, eye; OE, olfactory epithelium.
Novel fish GnRH receptors · N MONCAUT and others 775
www.endocrinology-journals.org Journal of Molecular Endocrinology (2005) 34, 767–779

to human GnRHR1) which is often replaced by Gluanother negatively charged amino acid in GnRHR1receptors. The acidic residue in the mammalian receptoris proposed to induce a high affinity conformation ofmammalian GnRH that allows it to interact with a finalbinding pocket (Fromme et al. 2001). However, an acidicamino acid at position 302 does not appear to berequired for high affinity binding to GnRH II (Millaret al. 2004). This is consistent with the lack of conserva-tion of Asp302 in GnRHR2 receptors and being replacedby small or hydrophobic amino acids: Pro in mammalsand some amphibians, Gln in some amphibians or Hisin all fishes. Although GnRHR2 in different speciesgenerally bind GnRH II, but not mammalian GnRH,with high affinity (Millar et al. 2004), the results of ligandbinding studies do not show a clear pattern of associa-tion between putative GnRH ligands and GnRHRstypes and subtypes (e.g. Illing et al. 1999, Millar et al.2001, Neill et al. 2001, Okubo et al. 2001, Wang et al.2001, Bogerd et al. 2002).
For those amino acids important for the structure ofthe receptor or of the binding pocket, Cys114, Trp164,Cys196, Trp206, Trp280, Trp291 and Pro320, are fullyconserved throughout the vertebrates. Of interest isthe fact that of the two pairs of cysteines which wouldenable disulfide bond formation in the extracellulardomain (Millar et al. 2004), only Cys114–Cys196 isconserved in all receptors, while Cys14–Cys200 are onlypresent simultaneously in mammalian GnRH type Ireceptors. These differences will modify the receptorstertiary structure and add further complexity in trying todeduce the contribution of structural elements for ligandbinding.
Analysis of mRNA expression of the dlGnRHRsmakes it evident that the action of GnRHs is not limitedto the central nervous system and the pituitary gland.dlGnRHRs are also expressed in the peripheral tissuesrelated to the senses, reproduction and homeostasis: theeyes, olfactory epithelium, gonads, kidney, gut, liver, andgills. The role of GnRHs in peripheral tissues remains tobe established.
All the dlGnRHRs are expressed in the brain,especially in the anterior and mid brain which includethe olfactory bulb, telencephalon to diencephalon andthe optic tectum. An analysis of co-localization in thebrain of the three European sea bass GnRH forms(González-Martínez et al. 2002) and the five receptors,may provide an insight on ligand specificity andfunction. Binding sites have also been described indifferent pituitary cells in pejerrey, Odontesthes bonariensis(Stefano et al. 1999). Of the European sea bass receptors,dlGnRHR-2A appears to be expressed only in LH-gonadotrophs and the level correlates with thereproductive cycle (González-Martínez et al. 2004).However, in teleost fishes, hypothalamic GnRHneurones innervate the adenohypophysis to control the
release not only of gonadotrophins (Somoza et al. 2002)but also of growth hormone (Marchant et al. 1989, Liet al. 2002), prolactin (Weber et al. 1997), somatolactin(Kakizawa et al. 1997) and thyroid stimulating hormone(Roy et al. 2000). In the goldfish GnRHR-1A and -1Bhave an overlapping expression and are present mainlyin gonadotropes and to a lesser extent in somesomatotrophs (Illing et al. 1999). With heterologousantisera in Nile tilapia – immunoreactive GnRHR-1Alocalized to FSH and LH cells and GnRH-1B toprolactin FSH- and LH-containing cells and as smallclusters scattered along the periphery of the parsintermedia of the pituitary (Parhar et al. 2002). Alsoheterologous antisera to GnRHR-2A and -2C stainedGH-containing cells of the Nile tilapia pituitary (Parharet al. 2002). The fact that dlGnRHR-2B is not expressedin the pituitary supports the hypothesis of GnRHpeptides possibly having specialized functions not relatedto gonadotrophin release.
Nevertheless, all European sea bass receptors, apartfrom dlGnRHR-1B, were strongly expressed in thegonads (i.e. testis and/or ovary). High levels ofGnRHRs expression have also been detected in thegonads of other teleosts (African catfish, Bogerd et al.2002; goldfish, Illing et al. 1999; rainbow trout, Madigouet al. 2000) and in mammals (Dong et al. 1996, Kakar& Jennes, 1995). This has also been confirmed bybinding studies (Pati & Habibi 1993), suggesting adirect effect of GnRH on sex steroid synthesis andgametogenesis.
dlGnRHR-2C and dlGnRHR-1A had relativelyhigher expression in the eye, an observation that has alsobeen made in rainbow trout (GnRHR-1B) and Japaneseeel, Anguilla japonica (GnRHR1) (Madigou et al. 2000,Okubo et al. 2000). Furthermore, GnRH immunoreac-tive fibers have been detected in the retina of platyfish,Xyphophorus maculatus (Munz et al. 1981), goldfish (Kahet al. 1986) and pejerrey (Miranda et al. 2003) and in theolfactory systems of several teleost species, including theAmerican eel, Anguilla rostrata (Grober et al. 1987) andmasu salmon (Kudo et al. 1994). Fish olfactory receptorsare also highly sensitive to GnRH (Andersen & Doving1991). Immunoreactive GnRH somata have beenidentified along the rostro-caudal extent of the olfactorynerve, and clustered within the medial component of theolfactory nerve as it arises from the olfactory epithelium(Nevitt et al. 1995). Since at least some cells in this clusterproject to the retina, possibly being part of a terminalnerve ganglion, it has been suggested that perhapsGnRH facilitates visual and olfactory perception duringsexual interactions (Nevitt et al. 1995). A role forolfactory stimuli in the regulation of GnRH secretionhas been suggested in sea lamprey, Petromyzon marinus,based on the overlap of olfactory- and GnRH-containing fibres from prolarval stages to metamorphosis(Tobet et al. 1996). It has been recently demonstrated
N MONCAUT and others · Novel fish GnRH receptors776
www.endocrinology-journals.orgJournal of Molecular Endocrinology (2005) 34, 767–779

using GnRH-receptor antagonist that GnRH secreted bymammalian olfactory cells promotes the differentiationand migration of the olfactory sensory lineage cells thatare committed to become GnRH neurons (Romanelliet al. 2004). The expression of both dlGnRHR-2A anddlGnRHR-2B in the olfactory rosettes suggest that theyare likely mediators of these processes.
Of particular interest is the expression of GnRHRs intissues related to osmoregulation. We show for the firsttime GnRHR expression in the gills (dlGnRHR-1A, -1Band -2A) and kidney (dlGnRHR-2A and dlGnRHR-2B).Few species have been also shown to express GnRHpeptide. Teleost (Haplochromis burtoni) kidney express atleast one form of GnRH (White & Fernald 1998) andhuman kidneys express two GnRH forms (Kakar &Jennes 1995). Since GnRH has never been associatedwith osmoregulation, a possible explanation for thepresence of GnRH and its receptor in kidney is that thistissue contains a high concentration of mast cells whichexpress GnRH in high levels (Khalil et al. 2003,Marchetti et al. 1996, Rissman 1996).
In conclusion, the existence of five different GnRHreceptors to three GnRH forms in this model ofvertebrate species indicates a complex interplay occursbetween ligands and receptors in the regulation ofpituitary and hormone secretion. The fact that GnRHRsshow a wide and different pattern of tissue distribution,is suggestive that, in addition to regulating the secretionof gonadotrophins from the anterior pituitary, GnRHand its receptor also play a role in the regulation of arange of cellular functions in an autocrine or paracrinemanner (Millar 2003). However, when considering thevarious species from different phylogenetic positions, inwhich GnRHRs expression has been studied, it is notpossible as yet to identify a specific pattern of expressioncharacteristic of each receptor type or subtype. Gener-ally speaking the common feature is the expression of thevarious receptors in the central nervous system, but thedistribution in other tissues is more variable. Thissuggests that in any one species (or higher taxonomicgroup) the GnRHRs receptors are generally involved insome ancestral functions, e.g. those related to reproduc-tion, but at a peripheral level specialization of functionshave also evolved independently.
Acknowledgements
This work received the support from a Portugal–Argentina Science and Technology Cooperation Agree-ment, SECYT-GRICES 472 Program PO/PA02-BI/001. N P M was in receipt of a fellowship covered bythe European Social Fund and National funds underPortuguese National Science Foundation (FCT)POCTI/SFRH/BD/6083/2001.
References
Adams BA, Tello JA, Erchegyi J, Warby C, Hong DJ, AkinsanyaKO, Mackie GO, Vale W, Rivier JE & Sherwood NM 2003 Sixnovel gonadotropin-releasing hormones are encoded as triplets oneach of two genes in the protochordate, Ciona intestinalis.Endocrinology 144 1907–1919.
Altschul SF, Madden TL, Schaffer AA, Zhang JH, Zhang Z, MillerW & Lipman DJ 1997 Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic AcidsResearch 25 3389–3402.
Amores A, Force A, Yan YL, Joly L, Amemiya C, Fritz A, Ho RK,Langeland J, Prince V, Wang YL et al. 1998 Zebrafish hoxclusters and vertebrate genome evolution. Science 282 1711–1714.
Andersen O & Doving KB 1991 Gonadotropin-releasing hormone(GnRH): A novel olfactory stimulant in fish. Neuroreport 2 458–460.
ASAB 2003 Guidelines for the treatment of animals in behaviouralresearch and teaching. Animal Behaviour 65 249–255.
Bailey JA, Church DM, Ventura M, Rocchi M & Eichler EE 2004Analysis of segmental duplications and genome assembly in themouse. Genome Research 14 789–801.
Bogerd J, Bas Diepenbroek W, Hund E, van Oosterhaut F, TevesAC, Leurs R & Blomenröhr M 2002 Two gonadotropin-realisinghormone receptor in the African catfish: No differences in ligandselectivity, but differences in tissue distribution. Endocrinology 1434673–4682.
Campion CE, Turzillo AM & Clay CM 1996 The gene encodingthe ovine gonadotropin-releasing hormone (GnRH) receptor:cloning and initial characterization. Gene 170 277–280.
Church GM & Gilbert W 1984 Genomic sequencing. PNAS 811991–1995.
Dong KW, Duval P, Zeng Z, Gordon K, Williams RF, Hodgen GD,Jones G, Kerdelhue B & Roberts JL 1996 Multiple transcriptionstart sites for the GnRH gene in rhesus and cynomolgus monkeys:a non-human primate model for studying GnRH gene regulation.Molecular and Cellular Endocrinology 117 121–130.
Falquet L, Pagni M, Bucher P, Hulo N, Sigrist CJ, Hofmann K &Bairoch A 2002 The PROSITE database, its status in 2002.Nucleic Acids Research 30 235–238.
Fromme BJ, Katz AA, Roeske RW, Millar RP & Flanagan CA 2001Role of aspartate7·32(302) of the human gonadotropin-releasinghormone receptor in stabilizing a high-affinity ligandconformation. Molecular Pharmacology 60 1280–1287.
González-Martínez D, Madigou T, Zmora N, Anglade I, Zanuy S,Zohar Y, Elizur A, Muñoz-Cueto A & Kah O 2001 Differentialexpression of three different prepro-GnRH (gonadotrophin-releasing hormone) messengers in the brain of the European seabass (Dicentrarchus labrax). Journal of Comparative Neurology 429144–155.
González-Martínez D, Zmora N, Mañanos E, Saligaut D, Zanuy S,Zohar J, Elizur A, Kah O & Muñoz-Cueto JA 2002Immunohistochemical localization of three different prepro-GnRHs in the brain and pituitary of the European sea bass(Dicentrarchus labrax) using antibodies to the corresponding GnRH-associated peptides. Journal of Comparative Neurology 446 95–113.
González-Martínez D, Madigou T, Mananos-Sanchez E,Cerdá-Reverter JM, Zanuy S, Kah O & Muñoz-Cueto JA 2004Cloning and expression of gonadotrophin-releasing hormonereceptor in the brain and pituitary of the European sea bass: an insitu hybridization study. Biology of Reproduction 70 1380–1391.
Grober MS, Bass AH, Burd G, Marchaterre MA, Segil N, Scholz K& Hodgson T 1987 The nervus terminalis ganglion in Anguillarostrata: an immunocytochemical and HRP histochemical analysis.Brain Research 436 148–152.
Ikemoto T, Enomoto M & Park MK 2004 Identification andcharacterization of a reptilian GnRH receptor from the leopardgecko. Molecular and Cellular Endocrinology 214 137–147.
Novel fish GnRH receptors · N MONCAUT and others 777
www.endocrinology-journals.org Journal of Molecular Endocrinology (2005) 34, 767–779

Illing N, Troskie BE, Nahorniak C, Hapgood JP, Peter R & MillarRP 1999 Two gonadotropin-releasing hormone receptor subtypeswith distinct ligand selectivity and differential distribution in brainand pituitary in the Goldfish (Carassius auratus). PNAS 962526–2531.
Jaillon O, Aury JM, Brunet F, Petit JL, Stange-Thomann N,Mauceli E, Bouneau L, Fischer C, Ozouf-Costaz C, Bernot A etal. 2004 Genome duplication in the teleost fish Tetraodon nigroviridisreveals the early vertebrate proto-karyotype. Nature 431 946–957.
Jodo A, Ando H & Urano A 2003 Five different types of GnRHreceptor gene are expressed in the brain of masu salmon(Oncorhynchus masou). Zoological Science 20 1117–1125.
Kah O, Breton B, Dulka JG, Nunez-Rodriguez J, Peter RE,Corrigan A, Rivier JE & Vale WW 1986 A reinvestigation of theGn-RH (gonadotrophin-releasing hormone) systems in the goldfishbrain using antibodies to salmon Gn-RH. Cell and Tissue Research244 327–337.
Kakar SS 1997 Molecular structure of the human gonadotropin-releasing hormone receptor gene. European Journal of Endocrinology137 183–192.
Kakar SS & Jennes L 1995 Expression of gonadotropin-releasinghormone and gonadotropin-releasing hormone receptor mRNAsin various non-reproductive human tissues. Cancer Letters 98 57–62.
Kakizawa S, Kaneko T & Hirano T 1997 Effects of hypothalamicfactors on somatolactin secretion from the organ-cultured pituitaryof rainbow trout. General and Comparative Endocrinology 105 71–78.
Khalil MH, Silverman AJ & Silver R 2003 Mast cells in the ratbrain synthesize gonadotropin-releasing hormone. Journal ofNeurobiology 56 113–124.
Krogh A, Larsson B, von Heijne G & Sonnhammer ELL 2001Predicting transmembrane protein topology with a hidden Markovmodel: Application to complete genomes. Journal of MolecularBiology 305 567–580.
Kudo H, Ueda H, Kawamura H, Aida K & Yamauchi K 1994Ultrastructural demonstration of salmon-type gonadotropin-releasing hormone (sGnRH) in the olfactory system of masusalmon (Oncorhynchus masou). Neuroscience Letters 166 187–190.
Lethimonier C, Madigou T, Muñoz-Cueto JA, Lareyre JJ & Kah O2004 Evolutionary aspects of GnRHs, GnRH neuronal systemsand GnRH receptors in teleost fish. General and ComparativeEndocrinology 135 1–16.
Li WS, Lin HR & Wong AOL 2002 Effects of gonadotropin-releasing hormone on growth hormone secretion and geneexpression in common carp pituitary. Comparative Biochemistry andPhysiology B-Biochemistry and Molecular Biology 132 335–341.
Madigou T, Mananos-Sanchez E, Hulshof S, Anglade I, Zanuy S &Kah O 2000 Cloning, tissue distribution, and central expression ofthe gonadotropin-releasing hormone receptor in the rainbow trout(Oncorhynchus mykiss). Biology of Reproduction 63 1857–1866.
Madigou T, Uzbekova S, Lareyre JJ & Kah O 2002 Two messengerRNA isoforms of the gonadotrophin-releasing hormone receptor,generated by alternative splicing and/or promoter usage, aredifferentially expressed in rainbow trout gonads duringgametogenesis. Molecular Reproduction and Development 63 151–160.
Marchant TA, Chang JP, Nahorniak CS & Peter RE 1989 Evidencethat gonadotropin-releasing hormone also functions as a growthhormone-releasing factor in the goldfish. Endocrinology 1242509–2518.
Marchetti B, Gallo F, Farinella Z, Romeo C & Morale MC 1996Luteinizing hormone-releasing hormone (LHRH) receptors in theneuroendocrine-immune network - Biochemical bases andimplications for reproductive physiopathology. Basis for CancerManagement 784 209–236.
McArdle CA, Franklin J, Green L & Hislop JN 2002 Signalling,cycling and desensitisation of gonadotroping releasing hormonereceptors. Journal of Endocrinology 173 1–11.
Millar RP 2003 GnRH II and type II GnRH receptors. Trends inEndocrinology and Metabolism 14 35–43.
Millar R, Lowe S, Conklin D, Pawson A, Maudsley S, Troskie B,Ott T, Millar M, Lincoln G, Sellar R et al. 2001 A novelmammalian receptor for the evolutionarily conserved type IIGnRH. PNAS 98 9636–9641.
Millar RP, Lu ZL, Pawson AJ, Flanagan CA, Morgan K &Maudsley SR 2004 Gonadotropin-releasing hormone receptors.Endocrine Reviews 25 235–275.
Miranda LA, Strobl-Mazzulla PH, Strussmann CA, Parhar I &Somoza GM 2003 Gonadotropin-releasing hormone neuronaldevelopment during the sensitive period of temperature sexdetermination in the pejerrey fish, Odontesthes bonariensis. General andComparative Endocrinology 132 444–453.
Munz H, Stumpf WE & Jennes L 1981 LHRH systems in brain ofplatyfish. Brain Research 221 1–13.
Neill JD, Duck LW, Sellers JC & Musgrove LC 2001 Agonadotropin-releasing hormone (GnRH) receptor specific forGnRH II in primates. Biochemical and Biophysical ResearchCommunications 282 1012–1018.
Nevitt GA, Grober MS, Marchaterre MA & Bass AH 1995GnRH-like immunoreactivity in the peripheral olfactory systemand forebrain of Atlantic salmon (Salmo salar): A reassessmentat multiple life-history stages. Brain Behavior and Evolution 45350–358.
Nicholas KB, Nicholas HB Jr & Deerfield II DW 1997 GeneDoc:Analysis and visualization of genetic variation. Embnet News 4 1–4.
Okubo K, Suetake H, Usami T & Aida K 2000 Molecular cloningand tissue-specific expression of a gonadotropin-releasing hormonereceptor in the Japanese eel. General and Comparative Endocrinology119 181–192.
Okubo K, Nagata S, Ko R, Kataoka H, Yoshiura Y, Mitani H,Kondo M, Naruse K, Shima A & Aida K 2001 Identification andcharacterization of two distinct GnRH receptor subtypes in ateleost, the Medaka Oryzias latipes. Endocrinology 142 4729–4739.
Okubo K, Ishii S, Ishida J, Mitani H, Naruse K, Kondo M, ShimaA, Tanaka M, Asakawa S, Shimizu N et al. 2003 A novel thirdgonadotropin-releasing hormone receptor in the medaka Oryziaslatipes: evolutionary and functional implications. Gene 314121–131.
Parhar IS, Soga T, Sakuma Y & Millar RP 2002 Spatio–temporalexpression of gonadotropin releasing hormone receptor subtypesin gonadotropes, somatotropes and lactotropes in the cichlid fish.Journal of Neuroendocrinology 14 657–665.
Pati D & Habibi H 1993 Characterization of gonadotropin-releasinghormone receptors in goldfish ovary: variation during folliculardevelopment. American Journal of Physiology 264 R227–R234.
Rissman EF 1996 Behavioral regulation of gonadotropin-releasinghormone. Biology of Reproduction 54 413–419.
Robinson-Rechavi M, Marchand O, Escriva H, Bardet PL, Zelus D,Hughes S & Laudet V 2001 Euteleost fish genomes arecharacterized by expansion of gene families. Genome Research 11781–788.
Romanelli RG, Barni T, Maggi M, Luconi M, Failli P, Pezzatini A,Pelo E, Torricelli F, Crescioli C, Ferruzzi P et al. 2004 Expressionand function of gonadotropin-releasing hormone (GnRH) receptorin human olfactory GnRH-secreting neurons: an autocrine GnRHloop underlies neuronal migration. Journal of Biological Chemistry279 117–126.
Roy P, Datta M, Dasgupta S & Bhattacharya S 2000 Gonadotropin-releasing hormone stimulates thyroid ativity in a feshwater murrel,Channa gachua (Ham.), and carps, Catla catla (Ham.) and Cirrhinusmrigala (Ham.). General and Comparative Endocrinology 117 456–463.
She X, Jiang Z, Clark RA, Liu G, Cheng Z, Tuzun E, Church DM,Sutton G, Halpern AL & Eichler EE 2004 Shotgun sequenceassembly and recent segmental duplications within the humangenome. Nature 431 927–930.
Somoza GM, Miranda LA, Strobl-Mazzulla P & Guilgur LG 2002Gonadotropin-releasing hormone (GnRH): From fish tomammalian brains. Cellular and Molecular Neurobiology 22 589–609.
N MONCAUT and others · Novel fish GnRH receptors778
www.endocrinology-journals.orgJournal of Molecular Endocrinology (2005) 34, 767–779

Stefano AV, Vissio PG, Paz DA, Somoza GM, Maggese MC &Barrantes GE 1999 Co-localization of GnRH binding sites withgonadotropin-, somatotropin-, somatolactin-, and prolactin-expressing pituitary cells of the pejerrey, Odontesthes bonariensis, invitro. General and Comparative Endocrinology 116 133–139.
Sun Y-M, Flanagan CA, Illing N, Ott TR, Sellar R, Fromme BJ,Hapgood J, Sharp P, Sealfon SC & Millar RP 2001 A chickengonadotropin-releasing hormone receptor that confers agonistactivity to mammalian antagonists. Identification of D-Lys6 in theligand and extracellular loop two of the receptor as determinants.Journal of Biological Chemistry 276 7754–7761.
Swofford DL 2002 PAUP*. Phylogenetic Analysis Using Parsimony(*and other methods), CD-ROM. Sunderland, Massachusetts:Sinauer Associates.
Tchoudakova A & Callard GV 1998 Identification of multipleCYP19 genes encoding different cytochrome P450 aromataseisozymes in brain and ovary. Endocrinology 139 2179–2189.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F & Higgins DG1997 The CLUSTAL X windows interface: flexible strategies formultiple sequence alignment aided by quality analysis tools. NucleicAcids Research 25 4876–4882.
Tobet SA, Chickering TW & Sower SA 1996 Relationship ofgonadotropin-releasing hormone (GnRH) neurons to the olfactorysystem in developing lamprey (Petromyzon marinus). Journal ofComparative Neurology 376 97–111.
Troskie BE, Hapgood JP, Millar RP & Illing N 2000Complementary deoxyribonucleic acid cloning, gene expression,and ligand selectivity of a novel gonadotropin-releasing hormonereceptor expressed in the pituitary and midbrain of Xenopus laevis.Endocrinology 141 1764–1771.
Wang L, Bogerd J, Choi HS, Seong JY, Soh JM, Chun SY,Blomenröhr M, Troskie BE, Millar RP, Yu WH et al. 2001 Threedistinct types of GnRH receptor characterized in the bullfrog.PNAS 98 361–366.
Weber GM, Powell JFF, Park M, Fischer WH, Craig AG, Rivier JE,Nanakorn U, Parhar IS, Ngamvongchon S, Grau EG et al. 1997Evidence that gonadotropin-releasing hormone (GnRH) functionsas a prolactin-releasing factor in a teleost fish (Oreochromismossambicus) and primary structures for three native GnRHmolecules. Journal of Endocrinology 155 121–132.
White RB & Fernald RD 1998 Ontogeny of gonadotropin-releasinghormone (GnRH) gene expression reveals a distinct origin forGnRH-containing neurons in the midbrain. General and ComparativeEndocrinology 112 322–329.
Zmora N, González-Martinez D, Muñoz-Cueto JA, Madigou T,Mañanos-Sanchez E, Zanuy-Doste S, Zohar Y, Kah O & ElizurA 2002 The GnRH system in the European sea bass (Dicentrarchuslabrax). Journal of Endocrinology 172 105–116.
Received 11 February 2005Accepted 15 February 2005
Novel fish GnRH receptors · N MONCAUT and others 779
www.endocrinology-journals.org Journal of Molecular Endocrinology (2005) 34, 767–779