first brazilian carcharodontosaurid and other new theropod dinosaur fossils from the...
TRANSCRIPT

at SciVerse ScienceDirect
Cretaceous Research 40 (2013) 131e142
Contents lists available
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
First Brazilian carcharodontosaurid and other new theropod dinosaur fossils fromthe CampanianeMaastrichtian Presidente Prudente Formation, São Paulo State,southeastern Brazil
Rodrigo P. Fernandes de Azevedo a, Felipe Medeiros Simbras a,b, Miguel Rodrigues Furtado a,Carlos Roberto A. Candeiro c,*, Lílian Paglarelli Bergqvist a
a Laboratório de Macrofósseis, Departamento de Geologia, Instituto de Geociências, Universidade Federal do Rio de Janeiro, Av. Athos da Silveira Ramos, 274, Ilha do Fundão,21941-916 Rio de Janeiro, Brazilb Petrobras, Av. Conselheiro Nébias, 159, Paquetá, 11015-001 Santos, São Paulo, Brazilc Laboratório de Geologia, Curso de Geografia, Faculdade de Ciências Integradas do Pontal, Campus Pontal, Universidade Federal de Uberlândia, Rua 20, 1.600 Bairro Tupã,CEP 38304-402, 38300-132 Ituiutaba, Minas Gerais, Brazil
a r t i c l e i n f o
Article history:Received 8 September 2011Accepted in revised form 15 June 2012Available online 26 July 2012
Keywords:TheropodaCarcharodontosauridaeLate CretaceousPresidente Prudente FormationBrazil
* Corresponding author. Tel.: þ51 34 3268 6118.E-mail address: [email protected] (C.R.A. Ca
0195-6671/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.cretres.2012.06.004
a b s t r a c t
We report new theropod dinosaur material from the Presidente Prudente Formation (CampanianeMaastrichtian), Bauru Group, in southwestern São Paulo state. The material comprises a fragment ofright maxilla of Carcharodontosauridae, an anterior portion of a left ilium of Abelisauroidea anda proximal portion of a right fibula of a coelurosaurian. Previous theropod records from the Bauru Basincomprise Abelisauridae and Tetanurae and in São Paulo state these have been represented by only a rightpremaxilla of an abelisaurid and isolated abelisaurid and carcharodontosaurid teeth. The new materialreported here represents the first theropod remains from the Presidente Prudente Formation, andincludes the first abelisauroid and coelurosaurian postcranial remains from the Bauru Basin in São Paulostate and the first latest Cretaceous carcharodontosaurid known from non-dental remains anywhere.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The Late Cretaceous Brazilian theropod dinosaur fossil recordcomprises two main lineages: Ceratosauria and Tetanurae. Cera-tosaurs are represented by Abelisauroidea, which is one of the mostrepresentative groups of theropods in Brazilian Late Cretaceousdeposits, but at present is still very scarce. Bertini (1996) reportedthe first occurrence of Abelisauridae based on a fragmentary rightpremaxilla and tooth, collected in São Paulo state, in the beds of theBauru Group. Subsequently, Kellner and Campos (2002) describedthe first taxonomically determinate Brazilian abelisauroid, Pycno-nemosaurus nevesi, based on five incomplete teeth, parts of sevencaudal vertebrae, a right pubis, a right tibia, and a distal articulationof the right fibula. These remains were collected from the conti-nental deposits of the Late Cretaceous Parecis Group in the Chapadados Guimarães Graben, Mato Grosso state. Teeth assigned to Abe-lisauridae were also reported from Minas Gerais (Candeiro, 2002,2007; Candeiro et al., 2006a, b; 2012b) and São Paulo (Candeiro
ndeiro).
All rights reserved.
et al., 2004) states, all from the Bauru Group. Novas et al. (2008)identified abelisaurid remains from the Marília Formation of theBauru Group in Minas Gerais. This material comprises a dorsalvertebra, the distal part of a femur, and a pedal phalanx, belongingto different specimens.
Tetanurae include three major groups: Spinosauridae, Carchar-odontosauridae and Coelurosauria. Brazilian spinosaurids are rep-resented by three species. Two of these, Angaturama limai (Kellnerand Campos, 1996) and Irritator challengeri Martill et al., 1996b, arefrom AptianeAlbian levels of the Romualdo Member of the SantanaFormation in the Araripe Basin, which has also yielded isolatedteeth (although it should be noted that A. limai has been regardedas a junior synonym of I. challengeri by Sereno et al., 1998). The thirdspecies, Oxalaia quilombensis Kellner et al., 2011, and several iso-lated teeth (Medeiros, 2006) belong to the AlbianeCenomanianAlcântara Formation of the Itapecuru Group in São Luís-GrajaúBasin, Maranhão state.
Brazilian carcharodontosaurids are represented by teeth fromthe São Luís-Grajaú Basin. These have been Alcântara Formation,Itapecuru Group Lage do Coringa, Maranhão State, northeasternBrazil (Vilas-Bôas et al., 1999; Medeiros, 2001). Another dentalrecord of Carcharodontosauridae comes from the Marília
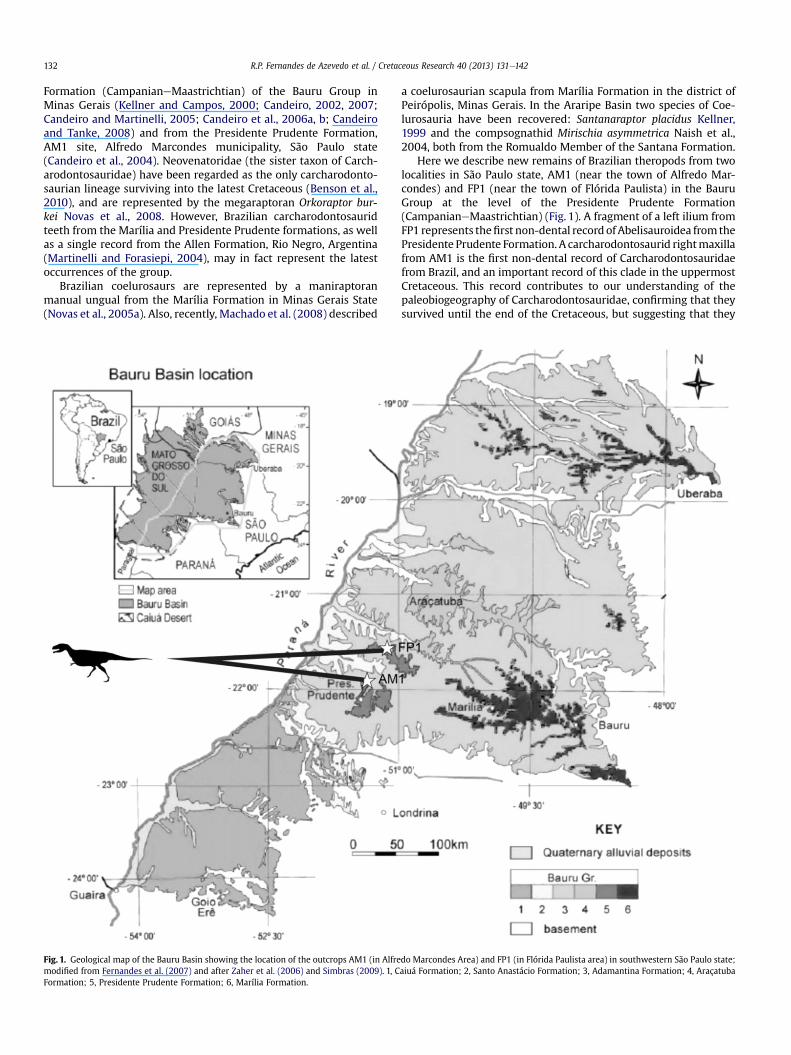
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142132
Formation (CampanianeMaastrichtian) of the Bauru Group inMinas Gerais (Kellner and Campos, 2000; Candeiro, 2002, 2007;Candeiro and Martinelli, 2005; Candeiro et al., 2006a, b; Candeiroand Tanke, 2008) and from the Presidente Prudente Formation,AM1 site, Alfredo Marcondes municipality, São Paulo state(Candeiro et al., 2004). Neovenatoridae (the sister taxon of Carch-arodontosauridae) have been regarded as the only carcharodonto-saurian lineage surviving into the latest Cretaceous (Benson et al.,2010), and are represented by the megaraptoran Orkoraptor bur-kei Novas et al., 2008. However, Brazilian carcharodontosauridteeth from the Marília and Presidente Prudente formations, as wellas a single record from the Allen Formation, Rio Negro, Argentina(Martinelli and Forasiepi, 2004), may in fact represent the latestoccurrences of the group.
Brazilian coelurosaurs are represented by a maniraptoranmanual ungual from the Marília Formation in Minas Gerais State(Novas et al., 2005a). Also, recently, Machado et al. (2008) described
Fig. 1. Geological map of the Bauru Basin showing the location of the outcrops AM1 (in Alfremodified from Fernandes et al. (2007) and after Zaher et al. (2006) and Simbras (2009). 1, CFormation; 5, Presidente Prudente Formation; 6, Marília Formation.
a coelurosaurian scapula from Marília Formation in the district ofPeirópolis, Minas Gerais. In the Araripe Basin two species of Coe-lurosauria have been recovered: Santanaraptor placidus Kellner,1999 and the compsognathid Mirischia asymmetrica Naish et al.,2004, both from the Romualdo Member of the Santana Formation.
Here we describe new remains of Brazilian theropods from twolocalities in São Paulo state, AM1 (near the town of Alfredo Mar-condes) and FP1 (near the town of Flórida Paulista) in the BauruGroup at the level of the Presidente Prudente Formation(CampanianeMaastrichtian) (Fig. 1). A fragment of a left ilium fromFP1 represents thefirst non-dental record ofAbelisauroidea from thePresidente Prudente Formation. A carcharodontosaurid rightmaxillafrom AM1 is the first non-dental record of Carcharodontosauridaefrom Brazil, and an important record of this clade in the uppermostCretaceous. This record contributes to our understanding of thepaleobiogeography of Carcharodontosauridae, confirming that theysurvived until the end of the Cretaceous, but suggesting that they
do Marcondes Area) and FP1 (in Flórida Paulista area) in southwestern São Paulo state;aiuá Formation; 2, Santo Anastácio Formation; 3, Adamantina Formation; 4, Araçatuba
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142 133
had a restricted geographic distribution, perhaps limited to SouthAmerica. We also report the left fibula of a basal coelurosaur fromAM1, the first remains of this clade in São Paulo state, expandingits occurrence from the northern and marginal TriânguloMineiro region in the Bauru Basin, Minas Gerais (Machado et al.,2008; Novas et al., 2008; Ghilardi and Fernandes, 2011; Candeiroet al., 2012).
Institutional abbreviations. BMNH, The Natural History Museum,London, England; MCF-PVPH, Museo Carmen Funes, VertebratePaleontology, Plaza Huincul, Neuquén, Argentina; SGM, Ministérede l’Energie et des Mines, Rabat, Níger; UFRJ-DG-R, UniversidadeFederal do Rio de Janeiro, Departamento de Geologia, Répteis.
Systematic nomenclature. We follow the phylogenetic hypothesisof Brusatte and Sereno (2008) and Benson et al. (2010) forAllosauroidea systematics. Allosauroidea includes Carchar-odontosauria, Allosaurus and Sinraptoridae. Carcharodontosauriaincludes Neovenatoridae and Carcharodontosauridae. We followthe phylogenetic hypothesis of Sereno (1999), Holtz (2000) andChoiniere et al. (2010) for Coelurosauria, and Carrano and Sampson(2008) for Ceratosauria.
2. Geology and stratigraphical setting
The Bauru Group comprises an area of 370.000 km2, partiallycovering the Brazilian states of São Paulo, Paraná, Mato Grosso doSul, Minas Gerais and Goiás. This unit was deposited in a depressionthat developed in the central-southern South American Platform,as a result of thermo-mechanical subsidence during the Cretaceousfollowing the break-up of Gondwana and the opening of theAtlantic Ocean (Fernandes and Coimbra, 1996). Southwestern SãoPaulo state encompasses the two areas studied here, the AlfredoMarcondes area close to the municipality of Presidente Prudente,
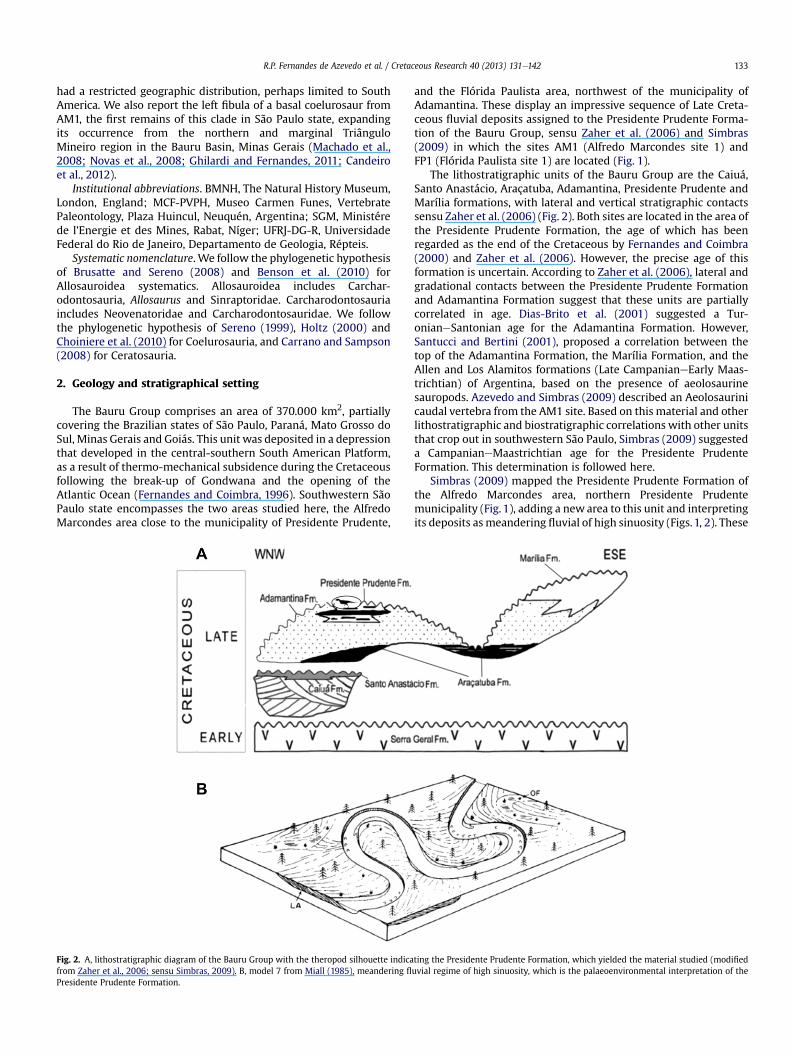
Fig. 2. A, lithostratigraphic diagram of the Bauru Group with the theropod silhouette indicafrom Zaher et al., 2006; sensu Simbras, 2009). B, model 7 from Miall (1985), meandering fl
Presidente Prudente Formation.
and the Flórida Paulista area, northwest of the municipality ofAdamantina. These display an impressive sequence of Late Creta-ceous fluvial deposits assigned to the Presidente Prudente Forma-tion of the Bauru Group, sensu Zaher et al. (2006) and Simbras(2009) in which the sites AM1 (Alfredo Marcondes site 1) andFP1 (Flórida Paulista site 1) are located (Fig. 1).
The lithostratigraphic units of the Bauru Group are the Caiuá,Santo Anastácio, Araçatuba, Adamantina, Presidente Prudente andMarília formations, with lateral and vertical stratigraphic contactssensu Zaher et al. (2006) (Fig. 2). Both sites are located in the area ofthe Presidente Prudente Formation, the age of which has beenregarded as the end of the Cretaceous by Fernandes and Coimbra(2000) and Zaher et al. (2006). However, the precise age of thisformation is uncertain. According to Zaher et al. (2006), lateral andgradational contacts between the Presidente Prudente Formationand Adamantina Formation suggest that these units are partiallycorrelated in age. Dias-Brito et al. (2001) suggested a Tur-onianeSantonian age for the Adamantina Formation. However,Santucci and Bertini (2001), proposed a correlation between thetop of the Adamantina Formation, the Marília Formation, and theAllen and Los Alamitos formations (Late CampanianeEarly Maas-trichtian) of Argentina, based on the presence of aeolosaurinesauropods. Azevedo and Simbras (2009) described an Aeolosaurinicaudal vertebra from the AM1 site. Based on this material and otherlithostratigraphic and biostratigraphic correlations with other unitsthat crop out in southwestern São Paulo, Simbras (2009) suggesteda CampanianeMaastrichtian age for the Presidente PrudenteFormation. This determination is followed here.
Simbras (2009) mapped the Presidente Prudente Formation ofthe Alfredo Marcondes area, northern Presidente Prudentemunicipality (Fig. 1), adding a new area to this unit and interpretingits deposits asmeandering fluvial of high sinuosity (Figs.1, 2). These
ting the Presidente Prudente Formation, which yielded the material studied (modifieduvial regime of high sinuosity, which is the palaeoenvironmental interpretation of the

R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142134
deposits are characterized by the presence of thick mudstone bedsinterbedded with sandstone lenses at the base, which are inter-preted as large overbank fine deposits (Simbras, 2009). Overlyingthese sediments are channelized deposits comprising fining-upward vertical profiles with conglomerates at the base, sand-stones in the middle andmudstones at the top. At the FP1 site thereare deposits similar to the Presidente Prudente Formation at theAM1 site; this region was previously mapped as the same unit byFernandes and Coimbra (2000). The material reported here wasrecovered from a conglomerate with trough cross-bedding at thebase of wide, shallow, meandering channels (Fig. 3).
3. Systematic palaeontology
3.1. Alfredo Marcondes area (AM1 site): Presidente PrudenteFormation
Theropoda Marsh, 1881Tetanurae Gauthier, 1986Allosauroidea Marsh, 1878Carcharodontosauria Benson, Carrano and Brusatte, 2010Carcharodontosauridae Stromer, 1931
Fig. 3. The AM1 and FP1 sections in the Presidente Prudente Formatio
Carcharodontosauridae indetFigs. 4, 5
Material. UFRJ-DG409-R, a fragmentary right maxilla 13 cm long. Ithas five alveoli and one replacement tooth; it lacks part of theanterior, posterior and dorsal portions, as reconstructed in Fig. 4.
Description and comparisons. UFRJ-DG409-R is fragmented, lack-ing the nasal fossa margin, the articular surface for the nasal, theantorbital fossa margin and the jugal process (Fig. 4). On themedial surface, the paradental plates are fused to each other(Fig. 5). According to Coria and Currie (2006), this feature is alsopresent in abelisaurids, Allosaurus fragilis Marsh, 1877, Gigan-otosaurus carolinii Coria and Salgado, 1995, Mapusaurus rosaeCoria and Currie, 2006, Torvosaurus Galton and Jensen, 1976, anddromaeosaurids, but it differs from Marshosaurus Madsen, 1976,Duriavenator hesperis Benson, 2008, Monolophosaurus jiangi Zhaoand Currie, 1993, Piatnitzkysaurus floresi Bonaparte, 1986, Sin-raptor dongi Currie and Zhao, 1993, tyrannosaurids and comp-sognathids (sensu Brusatte and Sereno, 2008). Furthermore, in
n showing the levels from which theropod material was collected.
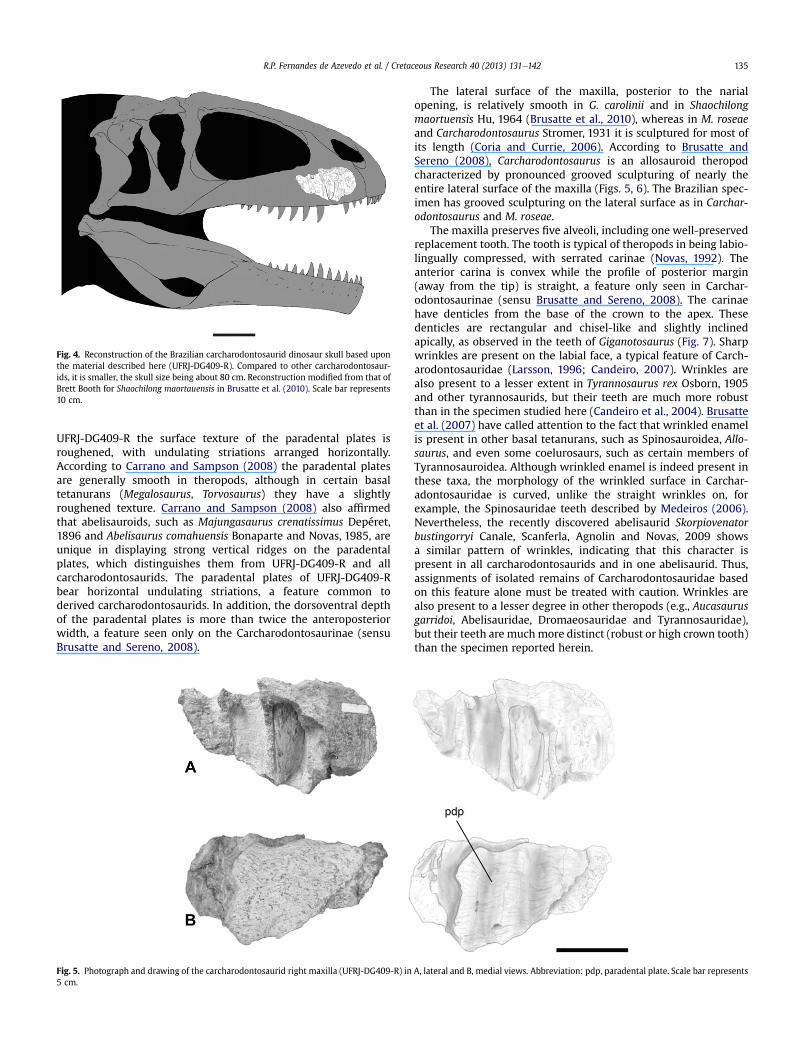
Fig. 4. Reconstruction of the Brazilian carcharodontosaurid dinosaur skull based uponthe material described here (UFRJ-DG409-R). Compared to other carcharodontosaur-ids, it is smaller, the skull size being about 80 cm. Reconstruction modified from that ofBrett Booth for Shaochilong maortauensis in Brusatte et al. (2010). Scale bar represents10 cm.
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142 135
UFRJ-DG409-R the surface texture of the paradental plates isroughened, with undulating striations arranged horizontally.According to Carrano and Sampson (2008) the paradental platesare generally smooth in theropods, although in certain basaltetanurans (Megalosaurus, Torvosaurus) they have a slightlyroughened texture. Carrano and Sampson (2008) also affirmedthat abelisauroids, such as Majungasaurus crenatissimus Depéret,1896 and Abelisaurus comahuensis Bonaparte and Novas, 1985, areunique in displaying strong vertical ridges on the paradentalplates, which distinguishes them from UFRJ-DG409-R and allcarcharodontosaurids. The paradental plates of UFRJ-DG409-Rbear horizontal undulating striations, a feature common toderived carcharodontosaurids. In addition, the dorsoventral depthof the paradental plates is more than twice the anteroposteriorwidth, a feature seen only on the Carcharodontosaurinae (sensuBrusatte and Sereno, 2008).
Fig. 5. Photograph and drawing of the carcharodontosaurid right maxilla (UFRJ-DG409-R) in5 cm.
The lateral surface of the maxilla, posterior to the narialopening, is relatively smooth in G. carolinii and in Shaochilongmaortuensis Hu, 1964 (Brusatte et al., 2010), whereas in M. roseaeand Carcharodontosaurus Stromer, 1931 it is sculptured for most ofits length (Coria and Currie, 2006). According to Brusatte andSereno (2008), Carcharodontosaurus is an allosauroid theropodcharacterized by pronounced grooved sculpturing of nearly theentire lateral surface of the maxilla (Figs. 5, 6). The Brazilian spec-imen has grooved sculpturing on the lateral surface as in Carchar-odontosaurus and M. roseae.
The maxilla preserves five alveoli, including one well-preservedreplacement tooth. The tooth is typical of theropods in being labio-lingually compressed, with serrated carinae (Novas, 1992). Theanterior carina is convex while the profile of posterior margin(away from the tip) is straight, a feature only seen in Carchar-odontosaurinae (sensu Brusatte and Sereno, 2008). The carinaehave denticles from the base of the crown to the apex. Thesedenticles are rectangular and chisel-like and slightly inclinedapically, as observed in the teeth of Giganotosaurus (Fig. 7). Sharpwrinkles are present on the labial face, a typical feature of Carch-arodontosauridae (Larsson, 1996; Candeiro, 2007). Wrinkles arealso present to a lesser extent in Tyrannosaurus rex Osborn, 1905and other tyrannosaurids, but their teeth are much more robustthan in the specimen studied here (Candeiro et al., 2004). Brusatteet al. (2007) have called attention to the fact that wrinkled enamelis present in other basal tetanurans, such as Spinosauroidea, Allo-saurus, and even some coelurosaurs, such as certain members ofTyrannosauroidea. Although wrinkled enamel is indeed present inthese taxa, the morphology of the wrinkled surface in Carchar-adontosauridae is curved, unlike the straight wrinkles on, forexample, the Spinosauridae teeth described by Medeiros (2006).Nevertheless, the recently discovered abelisaurid Skorpiovenatorbustingorryi Canale, Scanferla, Agnolin and Novas, 2009 showsa similar pattern of wrinkles, indicating that this character ispresent in all carcharodontosaurids and in one abelisaurid. Thus,assignments of isolated remains of Carcharodontosauridae basedon this feature alone must be treated with caution. Wrinkles arealso present to a lesser degree in other theropods (e.g., Aucasaurusgarridoi, Abelisauridae, Dromaeosauridae and Tyrannosauridae),but their teeth are muchmore distinct (robust or high crown tooth)than the specimen reported herein.
A, lateral and B, medial views. Abbreviation: pdp, paradental plate. Scale bar represents

Fig. 6. Comparisons between the maxillae of carcharodontosaurids in lateral views. AeD, carcharodontosaurids with vertical grooved sculpturing in the maxillae. A, UFRJ-DG409-R.B, right maxilla of Mapusaurus roseae (from Coria and Currie, 2006). C, left maxilla of Carcharodontosaurus saharicus, neotype, SGM-Din 1 (from Brusatte and Sereno, 2007). D, leftmaxilla of Carcharodontosaurus iguidensis (from Brusatte and Sereno, 2007). EeG, carcharodontosaurids without vertical grooved sculpturing in the maxillae. E, left maxilla ofGiganotosaurus carolinii (Coria and Salgado, 1995). F, left maxilla of Shaochilong maortuensis (from Brusatte et al., 2010). G, right maxilla of Eocarcharia dinops (from Sereno andBrusatte, 2008). Abbreviations: acf, accessory fenestra; af, antorbital fossa; afr, antorbital fossa ridge; ap, ascending process; ar, anterior ramus; jas, jugal articular surface;l, lacrimal; lf, lacrimal foramina; m, maxilla; mf, maxillary fenestra; nas, nasal articular surface; pmf, promaxillary fenestra; pnr, primary neurovascular foramina row; po, post-orbital; q, quadrate; snr, secondary neurovascular foramina row; so, supraorbital; soc, supraoccipital; snr, secondary neurovascular foramina row; vgs, vertical grooved sculpturing.Scale bars represent 5 cm in A, F, 10 cm in B, G, 15 cm in C, D, 100 cm in E.
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142136
Coelurosauria Huene, 1920
Coelurosauria indetFigs. 8, 9
Material. UFRJ-DG 390-R, the proximal portion of a right fibulalacking the proximal articular surface.
Description and comparisons. The proximal part of the fibula(Fig. 8) bears a deep medial fibular fossa. This fossa is ante-roposteriorly symmetrical and twice as long proximodistally as
anteroposteriorly. It also shows parts of a pronounced, anteriorlyoriented iliofibularis tubercle. Bittencourt and Kellner (2009),studying the anatomy and phylogenetic position of the basalsaurischian Staurikosaurus pricei Colbert, 1970, described themedial surface of its fibulae as concave, which is different from thatof UFRJ-DG 390-R. In ceratosaurs such as Ceratosaurus dentisulcatusMadsen and Welles, 2000; Majungasaurus crenatissimus Deperét,1896 and Rajasaurus narmadensis Wilson et al., 2003, and in someTetanurae such as the allosauroid Allosaurus fragilis, the carchar-odontosaurid Mapusaurus roseae, and the sinraptorid Sinraptor
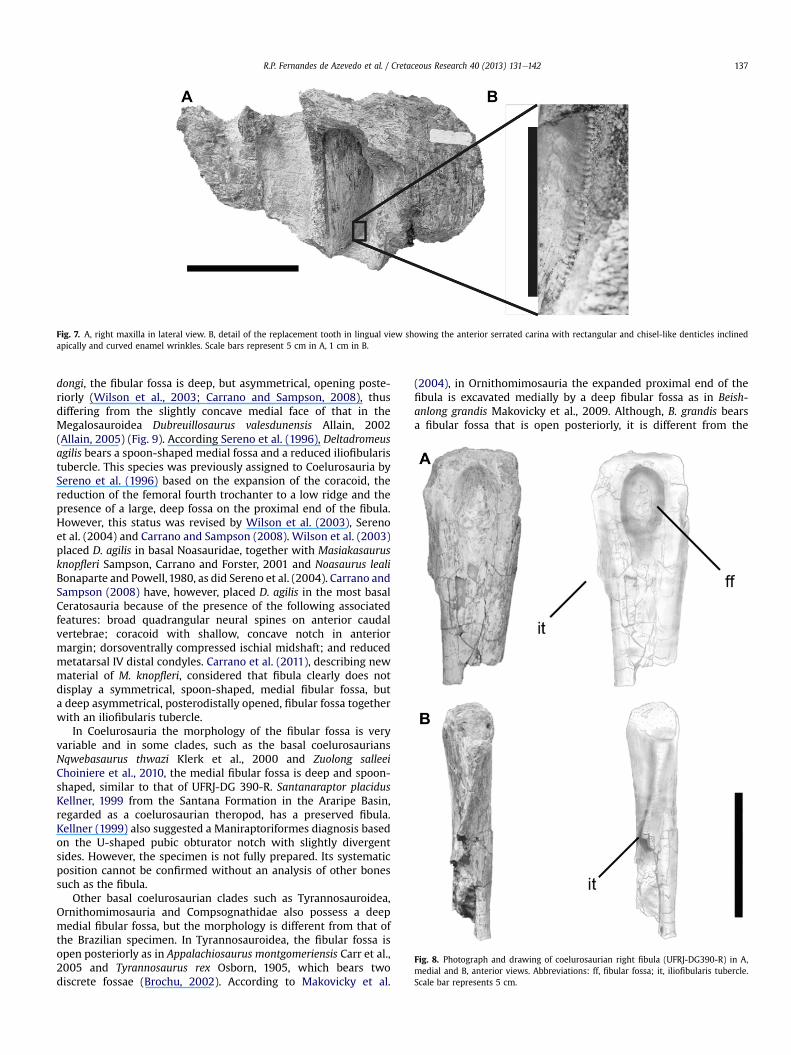
Fig. 7. A, right maxilla in lateral view. B, detail of the replacement tooth in lingual view showing the anterior serrated carina with rectangular and chisel-like denticles inclinedapically and curved enamel wrinkles. Scale bars represent 5 cm in A, 1 cm in B.
Fig. 8. Photograph and drawing of coelurosaurian right fibula (UFRJ-DG390-R) in A,medial and B, anterior views. Abbreviations: ff, fibular fossa; it, iliofibularis tubercle.Scale bar represents 5 cm.
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142 137
dongi, the fibular fossa is deep, but asymmetrical, opening poste-riorly (Wilson et al., 2003; Carrano and Sampson, 2008), thusdiffering from the slightly concave medial face of that in theMegalosauroidea Dubreuillosaurus valesdunensis Allain, 2002(Allain, 2005) (Fig. 9). According Sereno et al. (1996), Deltadromeusagilis bears a spoon-shaped medial fossa and a reduced iliofibularistubercle. This species was previously assigned to Coelurosauria bySereno et al. (1996) based on the expansion of the coracoid, thereduction of the femoral fourth trochanter to a low ridge and thepresence of a large, deep fossa on the proximal end of the fibula.However, this status was revised by Wilson et al. (2003), Serenoet al. (2004) and Carrano and Sampson (2008). Wilson et al. (2003)placed D. agilis in basal Noasauridae, together with Masiakasaurusknopfleri Sampson, Carrano and Forster, 2001 and Noasaurus lealiBonaparte and Powell,1980, as did Sereno et al. (2004). Carrano andSampson (2008) have, however, placed D. agilis in the most basalCeratosauria because of the presence of the following associatedfeatures: broad quadrangular neural spines on anterior caudalvertebrae; coracoid with shallow, concave notch in anteriormargin; dorsoventrally compressed ischial midshaft; and reducedmetatarsal IV distal condyles. Carrano et al. (2011), describing newmaterial of M. knopfleri, considered that fibula clearly does notdisplay a symmetrical, spoon-shaped, medial fibular fossa, buta deep asymmetrical, posterodistally opened, fibular fossa togetherwith an iliofibularis tubercle.
In Coelurosauria the morphology of the fibular fossa is veryvariable and in some clades, such as the basal coelurosauriansNqwebasaurus thwazi Klerk et al., 2000 and Zuolong salleeiChoiniere et al., 2010, the medial fibular fossa is deep and spoon-shaped, similar to that of UFRJ-DG 390-R. Santanaraptor placidusKellner, 1999 from the Santana Formation in the Araripe Basin,regarded as a coelurosaurian theropod, has a preserved fibula.Kellner (1999) also suggested a Maniraptoriformes diagnosis basedon the U-shaped pubic obturator notch with slightly divergentsides. However, the specimen is not fully prepared. Its systematicposition cannot be confirmed without an analysis of other bonessuch as the fibula.
Other basal coelurosaurian clades such as Tyrannosauroidea,Ornithomimosauria and Compsognathidae also possess a deepmedial fibular fossa, but the morphology is different from that ofthe Brazilian specimen. In Tyrannosauroidea, the fibular fossa isopen posteriorly as in Appalachiosaurus montgomeriensis Carr et al.,2005 and Tyrannosaurus rex Osborn, 1905, which bears twodiscrete fossae (Brochu, 2002). According to Makovicky et al.
(2004), in Ornithomimosauria the expanded proximal end of thefibula is excavated medially by a deep fibular fossa as in Beish-anlong grandis Makovicky et al., 2009. Although, B. grandis bearsa fibular fossa that is open posteriorly, it is different from the
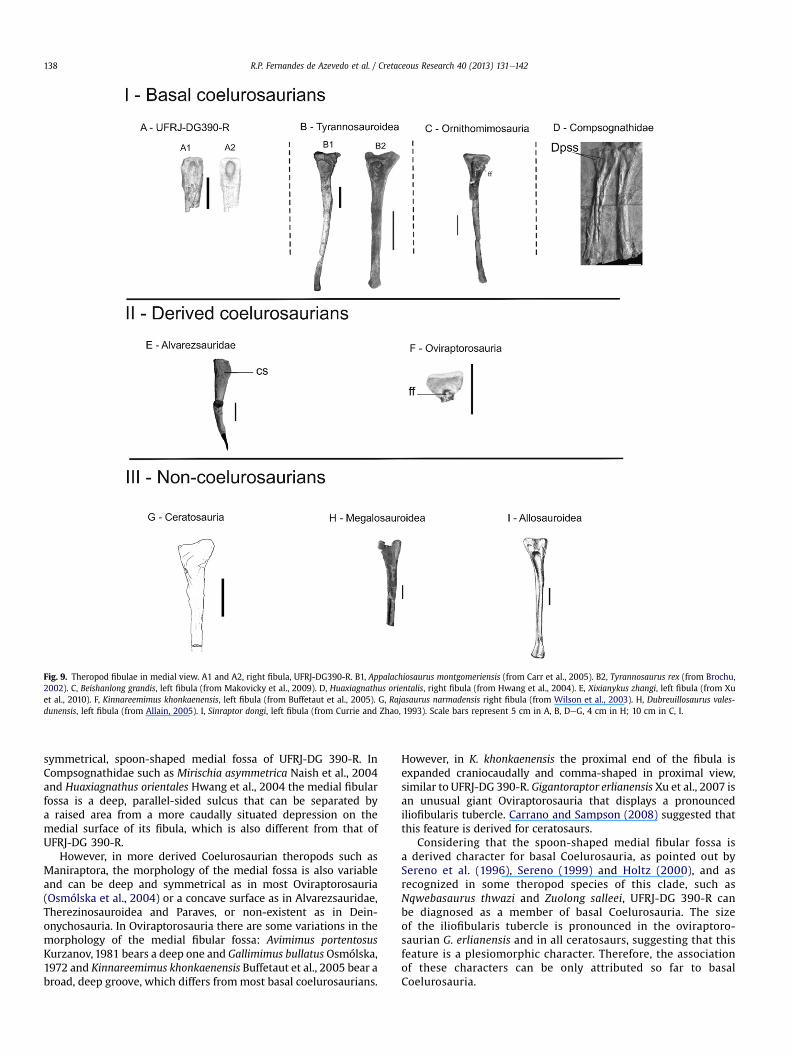
Fig. 9. Theropod fibulae in medial view. A1 and A2, right fibula, UFRJ-DG390-R. B1, Appalachiosaurus montgomeriensis (from Carr et al., 2005). B2, Tyrannosaurus rex (from Brochu,2002). C, Beishanlong grandis, left fibula (from Makovicky et al., 2009). D, Huaxiagnathus orientalis, right fibula (from Hwang et al., 2004). E, Xixianykus zhangi, left fibula (from Xuet al., 2010). F, Kinnareemimus khonkaenensis, left fibula (from Buffetaut et al., 2005). G, Rajasaurus narmadensis right fibula (from Wilson et al., 2003). H, Dubreuillosaurus vales-dunensis, left fibula (from Allain, 2005). I, Sinraptor dongi, left fibula (from Currie and Zhao, 1993). Scale bars represent 5 cm in A, B, DeG, 4 cm in H; 10 cm in C, I.
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142138
symmetrical, spoon-shaped medial fossa of UFRJ-DG 390-R. InCompsognathidae such as Mirischia asymmetrica Naish et al., 2004and Huaxiagnathus orientales Hwang et al., 2004 the medial fibularfossa is a deep, parallel-sided sulcus that can be separated bya raised area from a more caudally situated depression on themedial surface of its fibula, which is also different from that ofUFRJ-DG 390-R.
However, in more derived Coelurosaurian theropods such asManiraptora, the morphology of the medial fossa is also variableand can be deep and symmetrical as in most Oviraptorosauria(Osmólska et al., 2004) or a concave surface as in Alvarezsauridae,Therezinosauroidea and Paraves, or non-existent as in Dein-onychosauria. In Oviraptorosauria there are some variations in themorphology of the medial fibular fossa: Avimimus portentosusKurzanov, 1981 bears a deep one and Gallimimus bullatus Osmólska,1972 and Kinnareemimus khonkaenensis Buffetaut et al., 2005 bear abroad, deep groove, which differs frommost basal coelurosaurians.
However, in K. khonkaenensis the proximal end of the fibula isexpanded craniocaudally and comma-shaped in proximal view,similar to UFRJ-DG 390-R. Gigantoraptor erlianensis Xu et al., 2007 isan unusual giant Oviraptorosauria that displays a pronouncediliofibularis tubercle. Carrano and Sampson (2008) suggested thatthis feature is derived for ceratosaurs.
Considering that the spoon-shaped medial fibular fossa isa derived character for basal Coelurosauria, as pointed out bySereno et al. (1996), Sereno (1999) and Holtz (2000), and asrecognized in some theropod species of this clade, such asNqwebasaurus thwazi and Zuolong salleei, UFRJ-DG 390-R canbe diagnosed as a member of basal Coelurosauria. The sizeof the iliofibularis tubercle is pronounced in the oviraptoro-saurian G. erlianensis and in all ceratosaurs, suggesting that thisfeature is a plesiomorphic character. Therefore, the associationof these characters can be only attributed so far to basalCoelurosauria.
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142 139
3.2. Flórida Paulista area (FP1 site): Presidente Prudente Formation
Theropoda Marsh, 1881Ceratosauria Marsh, 1884Abelisauroidea Bonaparte and Novas, 1985
Abelisauroidea indet.Figs. 10, 11
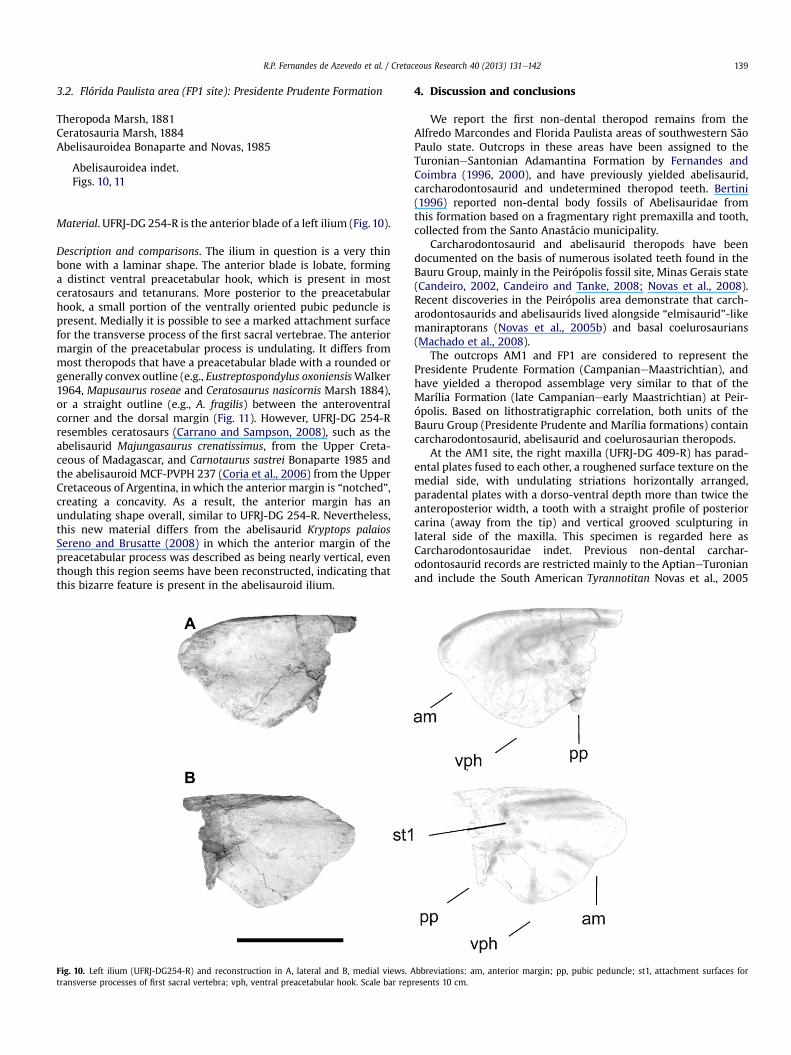
Material. UFRJ-DG 254-R is the anterior blade of a left ilium (Fig. 10).
Description and comparisons. The ilium in question is a very thinbone with a laminar shape. The anterior blade is lobate, forminga distinct ventral preacetabular hook, which is present in mostceratosaurs and tetanurans. More posterior to the preacetabularhook, a small portion of the ventrally oriented pubic peduncle ispresent. Medially it is possible to see a marked attachment surfacefor the transverse process of the first sacral vertebrae. The anteriormargin of the preacetabular process is undulating. It differs frommost theropods that have a preacetabular blade with a rounded orgenerally convex outline (e.g., Eustreptospondylus oxoniensisWalker1964, Mapusaurus roseae and Ceratosaurus nasicornis Marsh 1884),or a straight outline (e.g., A. fragilis) between the anteroventralcorner and the dorsal margin (Fig. 11). However, UFRJ-DG 254-Rresembles ceratosaurs (Carrano and Sampson, 2008), such as theabelisaurid Majungasaurus crenatissimus, from the Upper Creta-ceous of Madagascar, and Carnotaurus sastrei Bonaparte 1985 andthe abelisauroid MCF-PVPH 237 (Coria et al., 2006) from the UpperCretaceous of Argentina, in which the anterior margin is “notched”,creating a concavity. As a result, the anterior margin has anundulating shape overall, similar to UFRJ-DG 254-R. Nevertheless,this new material differs from the abelisaurid Kryptops palaiosSereno and Brusatte (2008) in which the anterior margin of thepreacetabular process was described as being nearly vertical, eventhough this region seems have been reconstructed, indicating thatthis bizarre feature is present in the abelisauroid ilium.
Fig. 10. Left ilium (UFRJ-DG254-R) and reconstruction in A, lateral and B, medial views. Atransverse processes of first sacral vertebra; vph, ventral preacetabular hook. Scale bar rep
4. Discussion and conclusions
We report the first non-dental theropod remains from theAlfredo Marcondes and Florida Paulista areas of southwestern SãoPaulo state. Outcrops in these areas have been assigned to theTuronianeSantonian Adamantina Formation by Fernandes andCoimbra (1996, 2000), and have previously yielded abelisaurid,carcharodontosaurid and undetermined theropod teeth. Bertini(1996) reported non-dental body fossils of Abelisauridae fromthis formation based on a fragmentary right premaxilla and tooth,collected from the Santo Anastácio municipality.
Carcharodontosaurid and abelisaurid theropods have beendocumented on the basis of numerous isolated teeth found in theBauru Group, mainly in the Peirópolis fossil site, Minas Gerais state(Candeiro, 2002, Candeiro and Tanke, 2008; Novas et al., 2008).Recent discoveries in the Peirópolis area demonstrate that carch-arodontosaurids and abelisaurids lived alongside “elmisaurid”-likemaniraptorans (Novas et al., 2005b) and basal coelurosaurians(Machado et al., 2008).
The outcrops AM1 and FP1 are considered to represent thePresidente Prudente Formation (CampanianeMaastrichtian), andhave yielded a theropod assemblage very similar to that of theMarília Formation (late Campanianeearly Maastrichtian) at Peir-ópolis. Based on lithostratigraphic correlation, both units of theBauru Group (Presidente Prudente and Marília formations) containcarcharodontosaurid, abelisaurid and coelurosaurian theropods.
At the AM1 site, the right maxilla (UFRJ-DG 409-R) has parad-ental plates fused to each other, a roughened surface texture on themedial side, with undulating striations horizontally arranged,paradental plates with a dorso-ventral depth more than twice theanteroposterior width, a tooth with a straight profile of posteriorcarina (away from the tip) and vertical grooved sculpturing inlateral side of the maxilla. This specimen is regarded here asCarcharodontosauridae indet. Previous non-dental carchar-odontosaurid records are restricted mainly to the AptianeTuronianand include the South American Tyrannotitan Novas et al., 2005
bbreviations: am, anterior margin; pp, pubic peduncle; st1, attachment surfaces forresents 10 cm.
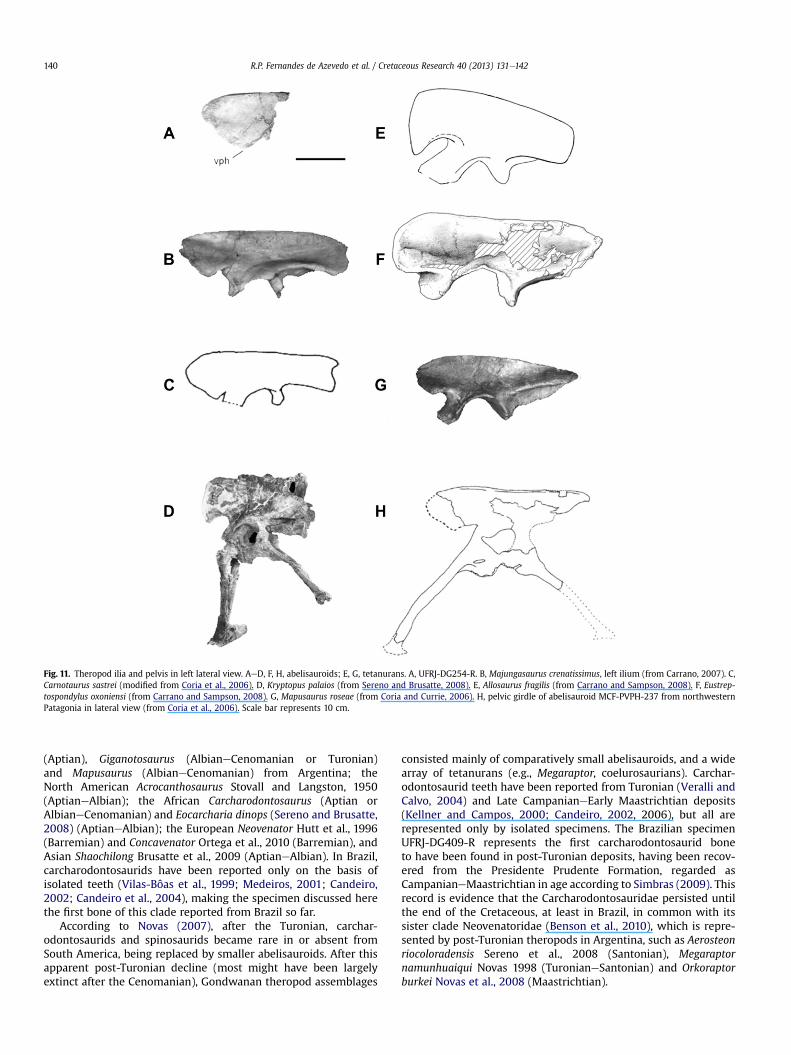
Fig. 11. Theropod ilia and pelvis in left lateral view. AeD, F, H, abelisauroids; E, G, tetanurans. A, UFRJ-DG254-R. B, Majungasaurus crenatissimus, left ilium (from Carrano, 2007). C,Carnotaurus sastrei (modified from Coria et al., 2006). D, Kryptopus palaios (from Sereno and Brusatte, 2008). E, Allosaurus fragilis (from Carrano and Sampson, 2008). F, Eustrep-tospondylus oxoniensi (from Carrano and Sampson, 2008). G, Mapusaurus roseae (from Coria and Currie, 2006). H, pelvic girdle of abelisauroid MCF-PVPH-237 from northwesternPatagonia in lateral view (from Coria et al., 2006). Scale bar represents 10 cm.
R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142140
(Aptian), Giganotosaurus (AlbianeCenomanian or Turonian)and Mapusaurus (AlbianeCenomanian) from Argentina; theNorth American Acrocanthosaurus Stovall and Langston, 1950(AptianeAlbian); the African Carcharodontosaurus (Aptian orAlbianeCenomanian) and Eocarcharia dinops (Sereno and Brusatte,2008) (AptianeAlbian); the European Neovenator Hutt et al., 1996(Barremian) and Concavenator Ortega et al., 2010 (Barremian), andAsian Shaochilong Brusatte et al., 2009 (AptianeAlbian). In Brazil,carcharodontosaurids have been reported only on the basis ofisolated teeth (Vilas-Bôas et al., 1999; Medeiros, 2001; Candeiro,2002; Candeiro et al., 2004), making the specimen discussed herethe first bone of this clade reported from Brazil so far.
According to Novas (2007), after the Turonian, carchar-odontosaurids and spinosaurids became rare in or absent fromSouth America, being replaced by smaller abelisauroids. After thisapparent post-Turonian decline (most might have been largelyextinct after the Cenomanian), Gondwanan theropod assemblages
consisted mainly of comparatively small abelisauroids, and a widearray of tetanurans (e.g., Megaraptor, coelurosaurians). Carchar-odontosaurid teeth have been reported from Turonian (Veralli andCalvo, 2004) and Late CampanianeEarly Maastrichtian deposits(Kellner and Campos, 2000; Candeiro, 2002, 2006), but all arerepresented only by isolated specimens. The Brazilian specimenUFRJ-DG409-R represents the first carcharodontosaurid boneto have been found in post-Turonian deposits, having been recov-ered from the Presidente Prudente Formation, regarded asCampanianeMaastrichtian in age according to Simbras (2009). Thisrecord is evidence that the Carcharodontosauridae persisted untilthe end of the Cretaceous, at least in Brazil, in common with itssister clade Neovenatoridae (Benson et al., 2010), which is repre-sented by post-Turonian theropods in Argentina, such as Aerosteonriocoloradensis Sereno et al., 2008 (Santonian), Megaraptornamunhuaiqui Novas 1998 (TuronianeSantonian) and Orkoraptorburkei Novas et al., 2008 (Maastrichtian).

R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142 141
The fibula (UFRJ-DG 390-R) is assigned to basal Coelurosauriabecause of the deep, spoon-shaped medial fossa, only observed inthis group in the South-African Nqwebasaurus thwazi and theChinese Zuolong salleei. The other theropod clades, Ceratosauria,basal Tetanurae and most derived Coelurosauria, do not havea medial fossa of the fibula of this morphology. Even in the largeCoelurosauria group, this morphology of the medial fossa of thefibula together the presence of the iliofibularis tubercle only occursin the two basal taxa noted above.
The anterior margin of the preacetabular process of the abeli-sauroid ilium (UFRJ-DG 254-R) from the FP1 site is undulating asseen inmost ceratosaurs more derived than Ceratosaurus, includingNoasauridae and Abelisauridae, forming the taxon Abelisauroidea(Carrano and Sampson, 2008). This morphology could be a featureof Abelisauridae. However, no noasaurid preserves this region ofthe ilium, so its presence cannot be assessed (Carrano andSampson, 2008). Hence, despite the strong similarity betweenUFRJ-DG 254-R and the abelisaurids (e.g., Majungasaurus andCarnotaurus) and even though the Abelisauridae are the mostrepresentative of theropods in South American Late Cretaceousdeposits (Novas, 2007) including the Bauru Basin (Novas et al.,2008), we classify this bone as an Abelisauroidea.
Thus, we report here a new theropod record from the São Paulostate, with new postcranial and cranial remains. The relative scar-city of theropods contrasts with the large number of titanosauridsauropods (Arid and Vizotto, 1971; Kellner and Azevedo, 1999;Candeiro et al., 2004; Santucci and Bertini, 2006; Bittencourt andKellner, 2011) and crocodylomorph bones (Carvalho and Bertini,1999; Campos et al., 2001; Carvalho et al., 2005, 2007; Nobre andCarvalho, 2006; Marinho and Carvalho, 2009). The fibula is thefirst coelurosaurian bone record from São Paulo state, indicatingthat this group has a wider geographical distribution in the BauruGroup of the Uberaba municipality, Minas Gerais state and insouthwestern São Paulo state. The maxilla is the first Brazilianrecord of a carcharodontosaurid bone and the first record of thisgroup at the end of the Cretaceous anywhere. It suggests that thepaleogeographical distribution of this clade at the end of theCretaceous was restricted to Brazil and possibly to Gondwana.
Acknowledgments
We thank Agustín Guillermo Martinelli (Argentina) for helpwith revising this paper and for discussion. The manuscript wassubstantially improved by Drs. Roger Benson (University of Cam-bridge) and DavidMartill (University of Portsmouth).We also thankthe Universidade Federal do Rio de Janeiro (UFRJ) for financialsupport of field work in Alfredo Marcondes and Flórida Paulistacounties, southwestern São Paulo state. R. Candeiro and L. Berqgvistwere supported by Conselho Nacional de Desenvolvimento Cien-tífico e Tecnológico (CNPq)/Bolsista de Produtividade em Pesquisafellowships.
References
Allain, R., 2005. The postcranial anatomy of the Megalosaur Dubreuillosaurusvalesdunensis (Dinosauria Theropoda) from the Middle Jurassic of Normandy,France. Journal of Vertebrate Paleontology 25, 850e858.
Arid, F.M., Vizotto, L.D., 1971. Antarctosaurus brasiliensis, um novo saurópodo doCretáceo Superior do sul do Brasil. 25� Congresso Brasileiro de Geologia(São Paulo), Boletim de Resumos Expandidos, pp. 297e305.
Azevedo, R.P.F., Simbras, F.M., 2009. New titanosaur sauropod remains from theBauru Group (Late Cretaceous) of the Alfredo Marcondes Region, São PauloState. Libro de Resúmenes. 24a Jornadas Argentinas de Paleontología de Ver-tebrados (San Rafael), p. 15.
Benson, R.B.J., Carrano, M.T., Brusatte, S.L., 2010. A new clade of archaic large-bodiedpredatory dinosaurs (Theropoda: Allosauroidea) that survived to the latestMesozoic. Naturwissenschaften 97, 71e78.
Bertini, R.J., 1996. Evidências de Abelisauridae (Carnosauria: Saurischia) do Neo-cretáceo da Bacia do Paraná. 4� Simpósio sobre o Cretáceo do Brasil (Rio Claro),Boletim de Resumos Expandidos, pp. 267e271.
Bittencourt, J.S., Kellner, A.W.A., 2009. The anatomy and phylogenetic position ofthe Triassic dinosaur Staurikosaurus pricei Colbert, 1970. Zootaxa 2079, 1e56.
Brochu, C.A., 2002. Osteology of Tyrannosaurs rex: insights from nearly completeskeleton and high-resolution computed tomographic analysis of the skull.Journal of Vertebrate Paleontology 22, 1e138.
Brusatte, S.L., Sereno, P.C., 2008. Phylogeny of Allosauroidea (Dinosauria: Ther-opoda): comparative analysis and resolution. Journal of Systematic Palae-ontology 6, 155e182.
Brusatte, S.L., Benson, R.B.J., Carr, T.D., Williamson, T.E., Sereno, P.C., 2007. Thesystematic utility of theropod enamel wrinkles. Journal of Vertebrate Paleon-tology 27, 1052e1056.
Brusatte, S.L., Chure, D.J., Benson, R.B.J., Xu, X., 2010. The osteology of Shaochilongmaortuensis, a carcharodontosaurid (Dinosauria: Theropoda) from the LateCretaceous of Asia. Zootaxa 2334, 1e46.
Campos, D.A., Suarez, J.M., Riff, D., Kellner, A.W.A., 2001. Short note on a newBaurusuchidae (Crocodyliformes, Metasuchia) from the Upper Cretaceous ofBrazil. Boletim do Museu Nacional, Nova Série Geologia 57, 1e7.
Candeiro, C.R.A., 2002. Dentes de Theropoda da Formação Marília (Santonia-noeMaastrichtiano), Bacia Bauru, Região de Peirópolis, Uberaba, Minas Gerais,Brazil. Unpublished Master’s thesis, Universidade Federal do Rio de Janeiro,Brasil, 136 pp.
Candeiro, C.R.A. 2007. Padrões morfológicos dos dentes de Abelisauroideae Carcharodontosauridae (Theropoda, Dinosauria) do Cretáceo da América doSul. Programa de Pós-Graduação em Geologia, Universidade Federal do Rio deJaneiro, Doctorate dissertation, 205 pp.
Candeiro, C.R.A., Martinelli, A.G., 2005. Abelisauroidea and Carcharodontosauridae(Theropoda, Dinosauria) in the Cretaceous of South America. Paleogeographicaland Geocronological implications. Sociedade & Natureza, Uberlândia 17 (33),5e19.
Candeiro, C.R.A., Tanke, D.H., 2008. A pathological Late Cretaceous carchar-odontosaurid tooth from Minas Gerais, Brazil. Bulletin of Geosciences 83,351e354.
Candeiro, C.R.A., Abranches, C.T., Abrantes, E.A., Avilla, L.S., Martins, V.C.,Moreira, A.L., Torres, S.R., Bergqvist, L.P., 2004. Dinosaur remains from westernSão Paulo state, Brazil (Bauru Basin, Adamantina Formation, Upper Cretaceous).Journal of South American Earth Sciences 18, 1e10.
Candeiro, C.R.A., Cau, A., Fanti, F., Nava, W., Novas, F.E., 2012a. First evidence of anunenlagiid (Dinosauria, Theropoda, Maniraptora) from the Bauru Group, Brazil.Cretaceous Research 37, 223e226.
Candeiro, C.R.A., Currie, P.J., Bergqvist, L.P., 2012b. Theropod teeth from MaríliaFormation (late Maastrichtian) at the Paleontological Site of Peirópolis in MinasGerais State, Brazil. Revista Brasileira de Geociências 42, 323e330.
Candeiro, C.R.A., Martinelli, A.G., Avilla, L.S., Rich, T.H., 2006b. Tetrapods from theUpper Cretaceous (TuronianeMaastrichtian) Bauru Group of Brazil: a reap-praisal. Cretaceous Research 27, 923e946.
Candeiro, C.R.A., Santos, A.R., Rich, T.H., Marinho, T.S., Oliveira, E.C., 2006a. Verte-brate fossils from the Adamantina Formation (Late Cretaceous), Prata paleon-tological district, Minas Gerais State, Brazil. Geobios 39, 319e327.
Carrano, M.T., Sampson, S.D., 2008. The phylogeny of Ceratosauria (Dinosauria:Theropoda). Journal of Systematic Palaeontology 6, 183e236.
Carrano, M.T., Loewen, M.A., Sertich, J.J.W., 2011. New materials of Masiakasaurusknopfleri Sampson, Carrano, and Forster, 2001, and implications for themorphology of the Noasauridae (Theropoda: Ceratosauria). SmithsonianContributions to Paleobiology 95, 1e53.
Carvalho, I.S., Bertini, R.J., 1999. Mariliasuchus, um novo Crocodylomorpha (Noto-suchia) do Cretáceo da Bacia Bauru, Brasil. Revista Geologia Colombiana 24,83e105.
Carvalho, I.S., Campos, A.C.A., Nobre, P.H., 2005. Baurusuchus salgadoensis, a newCrocodylomorpha from the Bauru Basin (Cretaceous), Brazil. GondwanaResearch 8, 11e30.
Carvalho, I.S., Vasconcellos, F.M., Tavares, S.A.S., 2007. Montealtosuchus arruda-camposi, a new peirosaurid crocodile (Mesoeucrocodylia) from the Late Creta-ceous Adamantina Formation of Brazil. Zootaxa 1607, 35e46.
Choiniere, J.N., Clark, J.M., Forster, C.A., Xu, X., 2010. A basal coelurosaur (Dino-sauria: Theropoda) from the Late Jurassic (Oxfordian) of the ShishugouFormation in Wucaiwan, People’s Republic of China. Journal of VertebratePaleontology 30, 1773e1796.
Coria, R.A., Currie, P.J., 2006. A new carcharodontosaurid (Dinosauria, Theropoda)from the Upper Cretaceous of Argentina. Geodiversitas 28, 71e118.
Coria, R.A., Currie, P.J., Carabajal, A.P., 2006. A new abelisauroid theropod fromnorthwestern Patagonia. Canadian Journal of Earth Sciences 43, 1283e1289.
Dias-Brito, D., Musacchio, E.A., Castro, J.C., Maranhão, M.S.A.S., Suárez, J.M.,Rodrigues, R., 2001. Grupo Bauru: uma Unidade Continental do Cretáceo noBrasil e Concepções baseadas em dados micropaleontológicos, isotópicos eestratigráficos. Revue de Paléobiologie 20, 245e304.
Fernandes, L.A., Coimbra, A.M., 1996. Estratigrafia y Ambientes Deposicionales de LaCuenca Bauru (Cretácico Superior, Brasil). Acta Geologica Hispanica 30, 11e30.
Fernandes, L.A., Coimbra, A.M., 2000. Revisão Estratigráfica da Parte Oriental daBacia Bauru (Neocretáceo). Revista Brasileira de Geociências 30, 717e728.
Fernandes, L.A., Castro, A.B., Basilici, G., 2007. Seismites in continental sand seadeposits of the Late Cretaceous Caiuá Desert, Bauru Basin, Brazil. SedimentaryGeology 199, 51e54.

R.P. Fernandes de Azevedo et al. / Cretaceous Research 40 (2013) 131e142142
Ghilardi, A.M., Fernandes, M.A., 2011. Dentes de Theropoda da Formação Ada-mantina (Cretáceo Superior, Bacia Bauru) da região do município de Ibirá, SãoPaulo, Brasil. In: Calvo, J., Porfiri, J., Gonzalez-Riga, B., Santos, D. (Eds.), 2011.Paleontología y Dinosaurios desde América Latina, first ed., vol. 1. EDIUNC,Mendoza, pp. 115e123.
Holtz T.R., Jr., 2000. Tyrannosauroidea. In: Weishampel, D.B., Dodson, P.,Osmólska, H. (Eds.), The Dinosauria. University of California Press, Berkeley, 5.pp. 111e136.
Kellner, A.W.A., 1999. Short note on a new dinosaur (Theropoda, Coelurosauria)from the Santana Formation (Romualdo Member, Albian), Northeastern Brazil.Boletim do Museu Nacional, Nova Série Geologia 49, 1e8.
Kellner, A.W.A., Azevedo, S.A.K., 1999. A new sauropod dinosaur (Titanosauria) fromthe Late Cretaceous of Brazil. In: National Science Museum Monographs, Tokyo,1. 111e142.
Kellner, A.W.A., Campos, D.A., 1996. First Early Cretaceous theropod dinosaur fromBrazil with comments on Spinosauridae. Neues Jahrbuch für Geologie undPaläontologie, Abhandlungen 199, 151e166.
Kellner, A.W.A., Campos, D.A., 2000. Brief review of dinosaur studies and perspec-tives in Brazil. Anais da Academia Brasileira de Ciências 72, 509e538.
Kellner, A.W.A., Campos, D.A., 2002. On a theropod dinosaur (Abelisauria) from thecontinental Cretaceous of Brazil. Arquivos do Museu Nacional 60, 163e170.
Larsson, H.C.E., 1996. Cranial morphology of the African theropod Carchar-odontosaurus saharicus (Allosauroidea). Journal of Vertebrate Paleontology 16(3), 47A.
Machado, E.B., Campos, D.A., Kellner, A.W.A., 2008. On a Theropoda scapula (UpperCretaceous) from the Marília Formation, Bauru Group, Brazil. PaläontologischeZeitischrift 82, 308e313.
Makovicky, P.J., Kobayashi, Y., Currie, P.J., 2004. Ornithomimosauria. In:Weishampel, D.B., Dodson, P., Osmólska, H. (Eds.), The Dinosauria 2. Universityof California Press, Berkeley, CA, pp. 111e136.
Marinho, T.S., Carvalho, I.S., 2009. An armadillo-like sphagesaurid crocodyliformfrom the Late Cretaceous of Brazil. Journal of South American Earth Sciences 27,36e41.
Martinelli, A.G., Forasiepi, A.M., 2004. Late Cretaceous vertebrates from the Bajo deSanta Rosa (Allen Formation), Rio Negro, Argentina, with the description ofa new sauropod dinosaur (Titanosauridae). Revista del Museo Argentino deCiencias Naturales 6, 257e305.
Medeiros, M.A., 2001. A Laje do Coringa (Ilha do Cajual, Bacia de São Luís, Baía deSão Marcos, MA): conteúdo fossilífero, bioestratinomia, diagênese e implicaçõesna paleobiogeografia do Mesocretáceo do Nordeste Brasileiro. Instituto deGeociências da Universidade Federal do Rio Grande do Sul, Porto Alegre,Doctorate dissertation, 107 pp.
Medeiros, M.A., 2006. Large theropod teeth from the Eocenomanian of northeasternBrazil and the occurrence of Spinosauridae. Revista Brasileira de Paleontologia9, 333e338.
Miall, A.D., 1985. Architectural-element analysis: a new method of facies analysisapplied to fluvial deposits. Earth-Science Reviews 22, 261e308.
Nobre, P.H., Carvalho, I.S., 2006. Adamantinasuchus navae: A new GondwananCrocodylomorpha (Mesoeucrocodylia) from the Late Cretaceous of Brazil.Gondwana Research 10, 370e378.
Novas, F.E., 1992. La Evolucíon de los dinosaurios carnívoros. In: II Curso de Pale-ontología em Cuenca. Instituto de Valdés, Ayuntamiento de Cuenca, Valdés,Actas, 92 pp.
Novas, F.E., 2007. Evolution of South American Dinosaurs. In: Carvalho, I.S.,Cassab, R.C.T., Schwanke, C., Carvalho, M.A., Fernandes, A.C.S., Rodrigues, M.A.C.,Carvalho, M.S.S., Arai, M., Oliveira, M.E.Q. (Eds.), 2007. Paleontologia: Cenáriosde Vida, 1. Interciência, Rio de Janeiro, pp. 341e361.
Novas, F.E., Carvalho, I.S., Ribeiro, L.C.B., Méndez, A.H., 2008. First abelisaurid boneremains from the Maastrichtian Marília Formation, Bauru Basin, Brazil. Creta-ceous Research 29, 625e635.
Novas, F.E., Ribeiro, L.C.B., Carvalho, I.S., 2005a. Maniraptoran theropod ungual fromthe Marília Formation (Upper Cretaceous), Brazil. Revista Museo ArgentinoCiencias Naturales 7, 31e36.
Novas, F.E., Valais, S., de Vickers-Rich, P., Rich, T., 2005b. A large Cretaceoustheropod from Patagonia, Argentina, and the evolution of carcharodontosaur-ids. Naturwissenschaften 92, 226e230.
Osmólska, H., Currie, P.J., Barsbold, R., 2004. Oviraptosauria. In: Weishampel, D.B.,Dodson, P., Osmólska, H. (Eds.), The Dinosauria. University of California Press,Berkeley, pp. 165e183.
Santucci, R.M., Bertini, R.J., 2001. Distribuição paleogeográfica e biocronológica dosTitanosauros (Saurischia, Sauropoda) do Grupo Bauru, Cretáceo Superior doSudeste Brasileiro. Revista Brasileira de Geociências 31, 307e314.
Santucci, R.M., Bertini, R.J., 2006. A new titanosaur from western São Paulo state,Upper Cretaceous Bauru Group, south-east Brazil. Palaeontology 49, 59e66.
Sereno, P.C., 1999. The evolution of dinosaurs. Science 284, 2137e2147.Sereno, P.C., Dutheil, D.B., Iarochene, M., Larsson, H.C.E., Lyon, G.H., Magwenw, P.,
Sidor, C.A., Varrichio, D.J., Wilson, J.A., 1996. Predatory dinosaurs from theSahara and Late Cretaceous faunal differentiation. Science 272, 986e991.
Sereno, P.C., Brusatte, S.L., 2008. Basal abelisaurid and carcharodontosaurid thero-pods from the Lower Cretaceous Elrhaz Formation of Niger. Acta Palae-ontologica Polonica 53, 15e46.
Simbras, F.M., 2009. Análise Estratigráfica do Intervalo Superior do Grupo Bauru(Cretáceo Superior) na região de Alfredo Marcondes-SP. UndergraduateMonograph, Departamento de Geografia, Universidade Federal do Rio deJaneiro, 34 pp.
Veralli, C., Calvo, J.O., 2004. Dientes de terópodos carcharodontosáuridos del Tur-oniano superioreConiaciano inferior del Neuquén, Patagonia, Argentina. Ame-ghiniana 41, 1e4.
Vilas-Bôas, I., Carvalho, I.S., Medeiros, M.A., Pontes, H., 1999. Dentes de Char-carodontosaurus (Dinosauria, Tyranosauridae) do Cenomaniano, Bacia de SãoLuis (Norte do Brasil). Anais da Academia Brasileira de Ciências 71 (4-I),846e847.
Wilson, J.A., Sereno, P.C., Srivastava, S., Bhatt, D.K., Khosla, A., Sahni, A., 2003. A newabelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous,Maastrichtian) of India. In: Contributions from the Museum of Paleontology,University of Michigan, 31. 1e42.
Zaher, H., Pol, D., Carvalho, A.B., Riccomini, C., Campos, D.A., Nava, W., 2006.Redescription of the cranial morphology of Mariliasuchus amarali, and itsphylogenetic affinities (Crocodyliformes, Notosuchia). American MuseumNovitates 3512, 1e40.