fine structure of phloematic trypanosomatid–coconut tree interaction
TRANSCRIPT

OTHERS
Fine structure of phloematic trypanosomatid–coconut treeinteraction
Maura da Cunha • Darlı Grativol Keller • Ivan Cunha Bustamante Filho •
Jorge Andre Sacramento de Magalhaes • Valdirene Moreira Gomes •
Wanderley de Souza • Jose Inacio Lacerda Moura • Jose Luiz Bezerra •
Flavio Costa Miguens
Received: 17 February 2009 / Accepted: 16 September 2009 / Published online: 15 December 2009
� The Phytopathological Society of Japan and Springer 2009
Abstract Phytomonas wilt or Hartrot is a fatal disease
of palm (Arecaceae) species including Cocos nucifera
(coconut) and is caused by a phloematic trypanosomatid, a
promastigote parasite that inhabits phloem sieve elements
of disease palms. In the present work, we described the
morphology of the interaction between a phloematic try-
panosomatid (Phytomonas staheli) and C. nucifera. Two
varieties and one ecotype of the adult coconut palm from
northeast and southeast Brazil were analyzed, totaling
34,000 plants. Coconut palm losses due to Hartrot varied
according to the variety or ecotype and geographic area.
Occurrence of Hartrot was insignificant in Rio de Janeiro
state (southeast), but in Bahia state (northeast) losses were
substantial when appropriate cultural practices were not
applied. Symptomatic and healthy palm tissues were ana-
lyzed by light and electron microscopy. Laboratory
diagnoses revealed the twisted promastigote form of phlo-
ematic trypanosomatids in the extracts of shoot apex, leaves,
stems and inflorescence in diseased plants, but not in the
healthy ones. No parasites were found in the roots. Although
the general anatomy of healthy and diseased palms was
similar, callose deposition in the sieve plates was revealed
by histochemistry and immunocytochemistry in the diseased
tissue. Plugging by the P-protein and plastid alterations was
also observed. Our observations strongly suggest that para-
site traffic between sieve elements took place, although their
cell bodies were larger than the sieve pores. Phloematic
trypanosomatid proliferation in the sieve tube elements
might interrupt the transport of phloem or/and consume
plant nutrients. In addition, an association between the
percentage of sieve elements colonized by pathogen in palm
tissues and disease severity was established.
Keywords Phytomonas staheli � Cocos nucifera �Wilt disease � Anatomy and ultrastructure �Histochemistry and immunocytochemistry
Introduction
Phytomonas, a flagellate protozoan belonging to the Try-
panosomatidae family, are heteroxenous promastigotes that
do not synthesize arginase in axenic culture. Their life
cycles take place in insects, as intermediate hosts, and in
plants, as definitive hosts (Camargo 1999; Camargo et al.
1987; Wallace et al. 1992). In 1909, the genus Phytomomas
was created by Donovan (1909) based on Lafont’s
description of protozoa in plants. The genus also has been
further grouped according to the location of the protozoa in
the plant: floricola (flower trypanosomatids), fruticola (fruit
trypanosomatids), laticicola (latex trypanosomatids) or
M. da Cunha � D. G. Keller � I. C. B. Filho �J. A. S. de Magalhaes � F. C. Miguens (&)
Laboratorio de Biologia Celular e Tecidual,
Universidade Estadual do Norte Fluminense Darcy Ribeiro
(UENF), Campos dos Goytacazes, Rio de Janeiro, Brazil
e-mail: [email protected]
V. M. Gomes
Laboratorio de Bioquımica e Fisiologia de Microorganismos,
Universidade Estadual do Norte Fluminense (UENF),
Campos dos Goytacazes, Rio de Janeiro, Brazil
W. de Souza
Laboratorio de Ultraestrutura Celular Hertha Meyer,
Instituto de Biofısica Carlos Chagas Filho, Universidade Federal
do Rio de Janeiro (UFRJ), Rio de Janeiro, Brazil
J. I. L. Moura � J. L. Bezerra
Centro de Pesquisas do Cacau, Comissao Executiva do Plano da
Lavoura Cacaueira (CEPEC/CEPLAC), Itabuna, Bahia, Brazil
123
J Gen Plant Pathol (2010) 76:74–83
DOI 10.1007/s10327-009-0212-y

phloemicola (phloem trypanosomatids) (Vickerman and
Dollet 1992). Phloemicola or phloem-restricted trypano-
somatids (Dollet 2001) have always been associated with
diseases in coffee (Stahel 1931; Vermeulen 1963) and palm
(Camargo 1999; Dollet 2001) species. However, no
symptoms have been detected in laticiferous plants (da
Cunha et al. 2000; Dollet 1984), except in cassava, Man-
ihot esculenta, where they exert a clear pathogenic effect
(Kitajima et al. 1986). Fruticola and floricola flagellates are
never associated with diseases.
Recently, the molecular and serological properties of
phloem trypanosomatids associated with wilts of coconut
and oil palm in Latin America have been determined, and
phloemicola has been proposed to be a well-defined taxon
among all plant trypanosomatids (Dollet 2001; Sturm et al.
2007). Previously, phloem trypanosomatids were described
as a new species, Phytomonas staheli (McGhee and
McGhee 1979). It was found in sieve elements of diseased
coconut and oil palms, causing Hartrot or fatal wilt of
palms (Parthasarthy and van Slobbe 1978; Parthasarthy
et al. 1976) and Marchitez sorpressiva (Dollet and Lopez
1978; Dollet et al. 1977). Attalea funifera (Bezerra et al.
1983), Cocos nucifera (Bezerra and Figueiredo 1982),
Elaeis guineensis (Attias et al. 1987; Oliveira and Bezerra
1982; Oliveira et al. 1987) and Roystonea regia (Attias
et al. 1989) have also been reported to be infected by
phloem trypanosomatids in Brazil. Since 1984, insects of
the genus Lincus have been circumstantially described as a
vector of trypanosomatids in palms in Latin America
(Desmier de Chenon 1984; Louise et al. 1986; Perthuis
et al. 1985). Fatal wilt disease has been transmitted
experimentally by Lincus lobuliger (Hemiptera: Pent-
atomidae) from diseased coconut to healthy oil palm
(Resende et al. 1986). The southeast of Bahia state has
been considered as an endemic area (Moura et al. 2002).
The most pronounced effects of the wilt syndrome are
water stress and deficiencies in the plant transport systems
(Beckman 1987; Lucas 1998). Callose synthesis and
deposition, lignification, phenolic compounds, and P-pro-
tein accumulation in the sieve plate and pores have been
reported as general plant responses to stress (Beckman
1987; Eschrich 1975; Lucas 1998). Phloem plastids were
also reported to be involved in the defense mechanisms in
plants (Walsh and Melaragno 1981; Zucker 1983). The
mechanisms by which phloematic trypanosomatids causes
diseases in plants are not clear. However, there may be
phloem blockages or changes in phloem transport from the
accumulation of the parasites (Agrios 2005; Dollet 2001).
The first introduction of coconut trees in the western
hemisphere was in Cabo Verde and the Sao Tome Islands
by Vasco da Gama when he was returning from India and
East Africa in 1499 (Harries 1977, 1978). They were first
grown as a plantation crop in the 1840s in Ceylon (Child
1974). In Brazil, coconut was introduced in Sao Vicente
during Portuguese colonial times in the sixteenth century
and several ecotypes were reintroduced during the twenti-
eth century, mainly in the northeast of the country (Ferreira
et al. 1998). In 1996, there were 260,000 ha of coconut
palms in Brazil, and Phytomonas wilt, potential vectors and
susceptible palms shared the same geographic distribution.
Because coconut palm plantations are an important eco-
nomic activity in Brazil, infection of palms by phloematic
trypanosomatids can have serious economic consequences
because they can destroy the palm trees. In this work, we
analyzed the morphological aspects of the protozoa–host
interaction.
Materials and methods
Plant material and study sites
One healthy and five symptomatic 10-year-old coconut
trees, Cocos nucifera (var. Malayan Green Dwarf), and one
10-year-old symptomatic C. nucifera (var. West African
Tall) were collected from the germplasm bank at an
experimental station (Estacao Experimental Lemos Maia)
of the Cacao Research Center (CEPEC/CEPLAC). The
station is located close to the municipality of Una
(15�1800000S, 33�0500000W) in south-eastern Bahia state,
Brazil. The vegetation around the station is typical of the
Atlantic Rain Forest. The soil is clayey. The climate in this
region is warm, tropical humid with relatively high rainfall
and high atmospheric temperatures. The total annual pre-
cipitation was 2,045.8 mm, and the mean annual air tem-
perature was about 28�C in 2004. Twenty-one diseased and
one healthy C. nucifera ecotype Green Dwarf plants were
collected from commercial plantations in Quissama
(22�0602400S, 41�2802000W) and Conceicao de Macabu
(22�0500700S, 41�5200600W), municipalities located in
northern Rio de Janeiro state, Brazil. The vegetation around
the plantations is dominated by grasses. The soil is sandy in
Quissama and clayey in Conceicao de Macabu. The climate
in this region is warm, tropical subhumid with relatively
low rainfall and high atmospheric temperatures. The total
annual precipitation varied between 550 and 850 mm and
the mean annual air temperature was about 26�C in 2004.
Coconut trees were divided into three types according to
morphological symptoms. Palms considered to be in the
last stage of infection were called type 1, which presented
browning of the oldest leaves, usually starting from the
tips, yellowing of younger leaves, blackening of unopened
and open inflorescences, dropping of all nuts and putre-
faction of the shoot apex with a foul odour. In the inter-
mediate infection stage, type 2 plants presented yellowing
of older leaves, browning of unopened and open
J Gen Plant Pathol (2010) 76:74–83 75
123

inflorescences, and nut drop, but no putrefaction of the
shoot apex. Plants of type 3, which had probably been
infected for less than 3 months, presented a gradual yel-
lowing and initial browning of the leaves, and initial
browning of unopened and open inflorescences. The per-
centage of losses from Phytomonas wilt were based on
field and laboratory diagnoses, for each variety or ecotype
and experimental area.
Light and electron microscopy
Extracts from roots, shoot apex, undifferentiated stem,
stem in the first stage of differentiation, leaf primordia,
young leaves, mature leaves, unopened inflorescences,
young and mature opened inflorescences were analyzed
with phase contrast microscopy and scanning electron
microscopy for laboratory diagnoses. Fragments of these
organs were also analyzed with light and electron micros-
copy. To determine the infection level, we counted the
number of infected and uninfected vascular bundles per
unit area (mm2) in 20 areas of stem meristem, stem, basal
petiole, inflorescence and root with the scanning electron
microscope.
For light microscopy, samples were fixed in formalin–
acetic acid–alcohol, dehydrated in ethanol and embedded
in paraffin (58–60�C). Sections (10–12 lm) were stained
with basic fuchsin-astrablau (Roeser 1962). Callose was
stained sky blue with 0.05% aniline blue (Johansen 1940).
Tannins were stained blue-green using 10% aqueous ferric
chloride (Johansen 1940). The samples were observed with
a Zeiss Axioplan photomicroscope (Carl Zeiss AG,
Oberkochen, Germany).
For electron microscopy, samples were fixed in 2.5%
glutaraldehyde, 2.0% formaldehyde in 0.05 M cacodylate
buffer, pH 7.4, for 2 h at room temperature. They were
post-fixed with 1.0% OsO4 in the same buffer. Then,
samples were rinsed in distilled water, stained en bloc in
0.5% aqueous uranyl acetate at room temperature for 2 h,
dehydrated in acetone and embedded in Epoxy resin
(Polybed�812, Polysciences, Inc., Warrington, PA, USA).
Sections were stained with uranyl acetate followed by lead
citrate and observed with a Zeiss EM 900 transmission
electron microscope at 80 kV accelerating voltage (Carl
Zeiss AG). For scanning electron microscopy, after post-
fixation, samples were dehydrated in acetone, dried with a
critical point dryer in CO2, sputter-coated with 20 nm gold
and observed with a Zeiss DSM 962 scanning electron
microscope (Carl Zeiss AG) at 25 kV accelerating voltage.
Immunocytochemistry
Antiserum against callose was prepared by immunization
of white New Zealand rabbits with laminarin (polymer of
b-1,3-glucan) from Laminaria digitata (Sigma, St. Louis,
MO, USA). Pre-immune serum was collected before
immunization. For immunocytochemical analysis, samples
were first fixed for 2 h at room temperature in a solution
containing 0.1% glutaraldehyde, and 2.0% formaldehyde in
0.1 M phosphate buffer (PB), pH 7.3, rinsed three times
with 0.1 M PB, pH 7.3, dehydrated in increasing concen-
trations of methanol (30–90%) and processed for LR Gold
(Polysciences, Inc.) embedding. Ultrathin sections were
treated following a previously described protocol (Pain
et al. 1996) with some modifications. A mixture of 10 mM
phosphate-buffered saline (PBS) and 0.15 M NaCl pH 7.5,
containing 1% bovine serum albumin (BSA) was used for
all rinsing steps and for diluting the reagents. Sections were
immunolabelled by immersing grids in drops (40 ll) of
solutions in the following sequence: (1) PBS ? BSA, for
30 min; (2) pre-immune serum in PBS ? BSA, for 20 min;
(3) anti-laminarin serum (1:100, v/v) in PBS ? BSA, for
2 h at room temperature; (4) five changes of buffer, for
10 min each; (5) goat anti-rabbit IgG antibody conjugated
with 5 nm or 10 nm colloidal gold (Sigma) (1:50, v/v) in
PBS ? BSA, for 2 h at room temperature; (6) five changes
of PBS ? BSA, for 10 min each; (7) five changes of
deionized water, for 10 min each. Sections were then air-
dried on a formvar coated grid, stained with uranyl acetate
and lead citrate and observed with a ZEISS 900 transmis-
sion electron microscope. Control sections were prepared
by replacing the primary anti serum with pre-immune
serum.
Results
Adult coconut tree losses due to Phytomonas wilt varied
depending on the variety or ecotype and geographic area.
In Rio de Janeiro state (*32,000 plants in experimental
area), Phytomonas wilt was rare, and the losses of the
Green Dwarf ecotype were \0.5%. In Bahia state at the
EELM, the Malayan Green Dwarf variety (2,000 plants)
had losses of *5% and a substantial loss ([30%) of the
West African Tall variety (1,000 plants in total). When
insecticides that control the vector(s) of the protozoa were
not sprayed, the entire crop of the Green Dwarf ecotype
(12,000 plants) was lost at a commercial farm near Una,
Bahia, Brazil.
Diseased specimens of C. nucifera had a gradient of
morphological symptoms, whereas healthy plants pre-
sented no morphological alterations. The shoot apex
putrefied in 100% of type 1 coconut palms and was related
to secondary infection. This symptom was not common in
type 2 plants and was absent in type 3. Laboratory diag-
noses using light and electron microscopy confirmed
morphological symptoms observed in the field. Phloematic
76 J Gen Plant Pathol (2010) 76:74–83
123

trypanosomatids were observed in extracts of shoot apex,
stems, leaves, and inflorescences from diseased palms at all
stages of infection, but absent in root extracts. Yeasts and
bacteria were observed in type 1 and type 2 palms. Phlo-
ematic trypanosomatids presented twisted promastigote
morphology, between 10 and 18 lm long with 1.5–2.5 lm
diameter, in all organ extracts from diseased plants
(Fig. 1a). Parasites were not observed in organ extracts of
healthy palms (data not shown). In addition, as the disease
progressed, the percentage of infected sieve elements
increased in palm organs (Table 1); in most parts of their
organs, type 1 trees had the highest percentages of infected
sieve elements. However, highly infected sieve elements
were found next to the slightly infected sieve elements
(Figs. 1b, d, 2k).
Anatomically, the vascular bundles of healthy and dis-
eased plants did not differ. In all organs analyzed, bundles
in the differentiated vascular system were randomly dis-
persed in ground parenchyma. Strengthening tissue, form-
ing a bundle sheath with lignified fibers, surrounded the
vascular bundles. The xylem consisted of 1 or 2 elements
of metaxylem and 4–12 of protoxylem (Fig. 1b, c). The
phloem consisted of sieve elements, usually associated
with two companion cells and a variable number of
parenchyma cells (Fig. 1b, d). Tanniferous idioblasts were
observed at parenchyma tissues in healthy and diseased
plants (Fig. 1e).
Xylem and phloem presented a similar ultrastructure
in healthy and infected plants. However, peculiarities
were identified mainly in sieve elements of diseased
plants. Although plastids at the periphery of the sieve
elements were frequent in uninfected palms (Fig. 1f, g),
they were rare in diseased palms (Fig. 1b, d). Phloem
plastids had external and internal membranes, but thy-
lakoid membranes were not identified, and starch grains
were infrequent. Usually, crystalline-like, electron-dense
structures were found in plastid stroma (Fig. 1g). In
longitudinal sections of vascular bundles of severely
diseased type 1 palms, the parasites were also found in
vessel elements in the shoot apex. The abundance of
phloematic trypanosomatids in sieve elements varied
according to disease severity (Fig. 1h–k); the pathogen
occupied a greater volume of the vascular tissue as the
disease became more severe. Colonization level was
always high in stem meristem (palm heart) and undif-
ferentiated organs in the three types of diseased plants
(Fig. 1k).
In sieve elements, phloematic trypanosomatids
appeared as twisted promastigotes with all morphological
features typical of trypanosomatids, including sub-pellic-
ular microtubules, a flagellum attached to the cell body
through specialized junctions, a flagellar pocket, and
sometimes a paraflagellar rod (Fig. 1k). The flagellum had
a basic structure with a 9 ? 2 pattern of axonemal
microtubules. In addition, mitochondrion was usually
normal, but the kinetoplast had different levels of con-
densation (Figs. 1k, 2h).
In the lumen of healthy and diseased plants, filaments
of P-protein were dispersed throughout the sieve ele-
ments, but trapped at sieve pores. Transmission electron
microscopy showed that P-protein filaments were often
distributed close to parasites. Sometimes, parallel fibrils
of P-protein were observed in the sieve pore (Fig. 2a)
and perpendicularly oriented to the cell membrane
(Fig. 2b).
Sieve plates of the sieve elements were in the lateral
and terminal walls. Diameters of the pores in sieve plates
varied between 0.5 and 1.0 lm. Infected sieve elements
frequently presented fibrils in their lumen (Fig. 2a). Cal-
lose was deposited in sieve elements of healthy and dis-
eased plants. In healthy palms, callose deposition was
slightly stained with aniline blue as seen by light
microscopy (Fig. 2c) but was strongly stained in diseased
plants (Fig. 2d). With transmission electron microscopy,
callose appeared as an electron-lucent structure (Fig. 2a).
Fig. 1 Microghaphs of Cocos nucifera. a Extract of stem apical
meristem of var. Malayan Green Dwarf. Phloematic trypanosomatids
(solid arrow) presenting twisted cellular bodies. Starch grains (openarrows) and cellular debris (arrowheads) are in background. SEM.
b Longitudinal section of leaf primordia from var. West African Tall.
Five sieve elements (square) having different levels of colonization.
Vascular parenchyma (VP) and sclerenchyma (star) in formation
presenting typical anatomy. LM. c Transversal section of stem
meristem from var. Malayan Green Dwarf. Infected vascular bundle
(ellipse) of basal petiole from palm type 2 immature leaf. Vascular
bundle showing small parenchyma small cells, xylem (open arrow)
and phloem (white arrow). SEM. d Longitudinal section of primordial
leaf from ecotype Green Dwarf (type 2). The vascular bundle has five
infected sieve tube elements (asterisk). Note callose deposition in
sieve plate (open arrow) and companion cells (cc). LM. e Longitu-
dinal section of stem meristem from ecotype Green Dwarf (type 2).
A tanniferous idioblast with electron-dense tannin accumulation in
vacuole. TEM. f Transversal section of stem meristem from healthy
ecotype Green Dwarf. The vascular bundle has companion cells (cc)
and sieve elements. Phloem plastid (arrow) in sieve element. TEM.
g High magnification of sieve element from (f). Phloem plastid close
to cell wall has external and internal membranes, but no distinct
thylakoid membrane. Note crystal-like structures in stroma. TEM.
h Longitudinal section of basal petiole from ecotype Green Dwarf
(type 1). Phloematic trypanosomatids are highly colonizing the sieve
elements. SEM. i Longitudinal section of basal petiole from ecotype
Green Dwarf (type 2). Contiguous sieve element with intermediate
level of trypanosomatid colonization. SEM. j Longitudinal section of
basal petiole from ecotype Green Dwarf (type 3). Low colonization
by trypanosomatids is evident in sieve elements of type 3-plants.
SEM. k Transverse section of stem meristem from var. Malayan
Green Dwarf (type 3). Highly colonized sieve elements have a lateral
sieve plate with pores (black arrow). Phloematic trypanosomatids
have typical trypanosomatid morphology. Condensed kinetoplast
(black/white arrow) is the most evident structure. TEM. Scale bars in
a, b, d-f, h-j 10 lm; c 250 lm; g, k 5 lm
c
J Gen Plant Pathol (2010) 76:74–83 77
123

More callose was deposited in infected sieve elements
(Fig. 2a, d, g–l) than in healthy plants. Callose deposition
appeared to be related to disease severity (Table 1).
Immunolabelling strongly suggested that electron-lucent
areas of the sieve plate resulted from callose deposition
(Fig. 2e, f).
Despite the fact that pathogen cell bodies were larger
than sieve pores, several images strongly suggested that the
78 J Gen Plant Pathol (2010) 76:74–83
123
parasite moved between contiguous sieve tube elements
(Fig. 2g–l). Phloematic trypanosomatids were also found in
the sieve pores. Cell bodies spanning both sides of the sieve
pores were also identified (Fig. 2i–l). Apparently, sub-
pellicular microtubules (Fig. 2i) and the cytoplasm
(Fig. 2l) of the parasite cells were rearranged during tran-
sit. In addition, in lateral sieve plates (Fig. 2m) and end
sieve plates (Fig. 2n), parasite cell bodies were preferen-
tially oriented perpendicularly to sieve plates, and their
flagella were difficult to visualize, while those on free
protozoa were easily identified.
Discussion
Losses have been reported from phloematic trypanoso-
matids or P. staheli, mainly in coconut and oil palm in
Latin America (McCoy and Martinez-Lopes 1982; Ohler
1999; Waters 1978). Southeast Bahia state (northeast
Brazil) has been cited as endemic for Phytomonas wilt in
palms; there is an overlap of etiological agent, vectors and
host distribution, and a constant low rate of this disease is
expected (Moura et al. 2002). Yellow Dwarf varieties may
be more resistant than Green Dwarf varieties (Bezerra and
Figueiredo 1982). In the present study, devastation on a
commercial farm in Una was related to ecotype suscepti-
bility or deficient integrated disease management, and an
epidemic affecting the West African Tall variety was due
to crop management. In Rio de Janeiro state, fatal wilt
levels were insignificant, and the vector was not identified.
However, vector identification is fundamental for disease
control. Lincus lobuliger (Hemiptera: Pentatomidae) has
been implicated in protozoan transmission (Resende et al.
1986), and vectors of phloematic trypanosomatids have
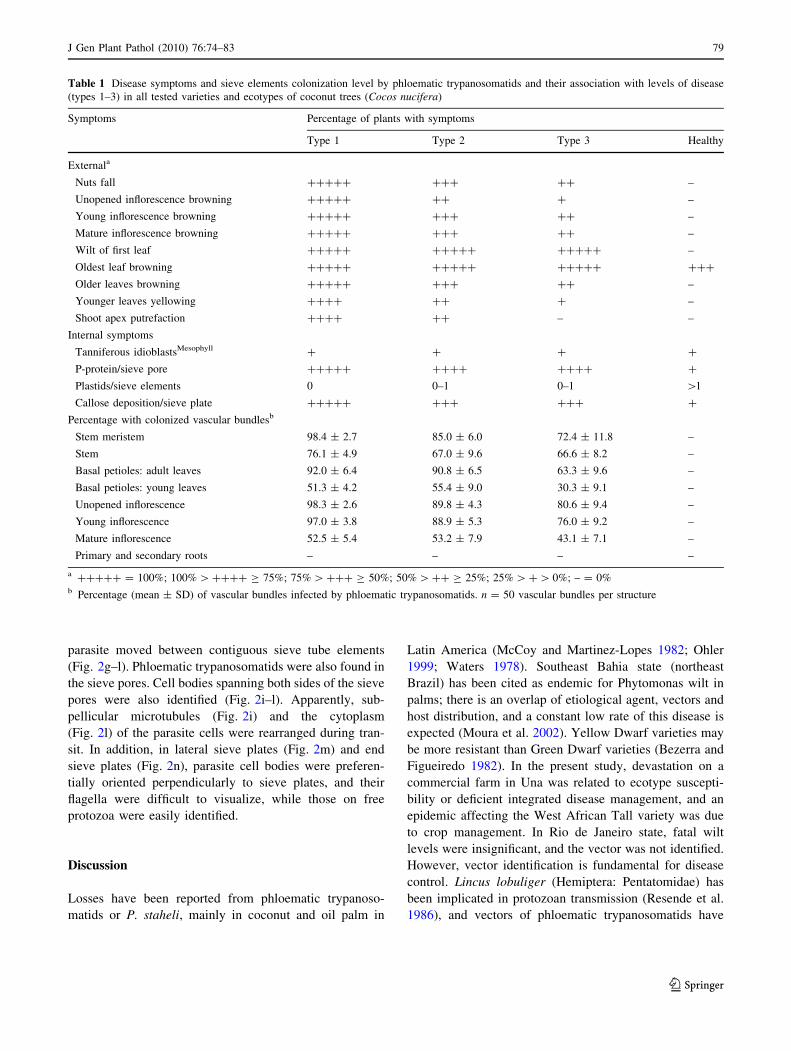
Table 1 Disease symptoms and sieve elements colonization level by phloematic trypanosomatids and their association with levels of disease
(types 1–3) in all tested varieties and ecotypes of coconut trees (Cocos nucifera)
Symptoms Percentage of plants with symptoms
Type 1 Type 2 Type 3 Healthy
Externala
Nuts fall ????? ??? ?? –
Unopened inflorescence browning ????? ?? ? –
Young inflorescence browning ????? ??? ?? –
Mature inflorescence browning ????? ??? ?? –
Wilt of first leaf ????? ????? ????? –
Oldest leaf browning ????? ????? ????? ???
Older leaves browning ????? ??? ?? –
Younger leaves yellowing ???? ?? ? –
Shoot apex putrefaction ???? ?? – –
Internal symptoms
Tanniferous idioblastsMesophyll ? ? ? ?
P-protein/sieve pore ????? ???? ???? ?
Plastids/sieve elements 0 0–1 0–1 [1
Callose deposition/sieve plate ????? ??? ??? ?
Percentage with colonized vascular bundlesb
Stem meristem 98.4 ± 2.7 85.0 ± 6.0 72.4 ± 11.8 –
Stem 76.1 ± 4.9 67.0 ± 9.6 66.6 ± 8.2 –
Basal petioles: adult leaves 92.0 ± 6.4 90.8 ± 6.5 63.3 ± 9.6 –
Basal petioles: young leaves 51.3 ± 4.2 55.4 ± 9.0 30.3 ± 9.1 –
Unopened inflorescence 98.3 ± 2.6 89.8 ± 4.3 80.6 ± 9.4 –
Young inflorescence 97.0 ± 3.8 88.9 ± 5.3 76.0 ± 9.2 –
Mature inflorescence 52.5 ± 5.4 53.2 ± 7.9 43.1 ± 7.1 –
Primary and secondary roots – – – –
a ????? = 100%; 100% [ ???? C 75%; 75% [ ??? C 50%; 50% [ ?? C 25%; 25% [ ? [ 0%; – = 0%b Percentage (mean ± SD) of vascular bundles infected by phloematic trypanosomatids. n = 50 vascular bundles per structure
J Gen Plant Pathol (2010) 76:74–83 79
123

been identified for the genera Lincus and Oclerus
(Camargo 1999; Dollet 2001).
Pathogenic species of the Trypanosomatidae have been
found in sieve elements of diseased coconut (Parthasarthy
and van Slobbe 1978; Parthasarthy et al. 1976), oil palms
(Dollet and Lopez 1978; Dollet et al. 1977) and other
plants from the Arecaceae family (Camargo 1999). Results
presented here establish a clear relationship between dis-
ease development and host morphological alterations and
also show the anatomy and ultrastructure of healthy and
diseased palms.
Phloem colonization is an important step in the pro-
gression of Phytomonas wilt in palms (Camargo 1999;
Dollet 1984; Wallace et al. 1992). The presence of the
parasite on both sides of sieve pores suggest that the larger
protozoa may pass through the smaller sieve pore by
deformation of flexible cell bodies; however, this question
about transport across the pore remains open. McGhee and
Cosgrove (1980) based on microscopic observations of the
swimming of several Trypanosomatidae genera show that
these flagella are capable of a considerable variety of
movements. Bending activity produces a sinusoidal wave,
which is not quite planar, so that the cell rotates at a low
frequency; the wave normally propagates from tip to base,
but when the flagellum touches an obstruction anywhere
along its length, the direction of propagation may be
reversed by changing the direction of movement. In the
present study, we found parasite flagella plugging sieve
pores of diseased tissue contrary to present knowledge of
the Trypanosomatidae species movement and flagellum
physiology.
The anatomy of healthy and diseased palms was very
similar, suggesting that trypanosomatids never caused a
destructive effect against host plant cells. Callose depo-
sition was the main difference. Callose is a linear b-1,3-
glucan with some 1,6- branches, and as a glucosidic
non-cellulosic polysaccharide, it differs from cellulose in
the type of linkage between the glucose residues
(Hopkins 1995). Usually, callose is detected by its
specific staining with aniline blue (Hong et al. 2001) and
by immunocytochemistry (Northcote et al. 1989). Callose
has been localized in the cell plate, plasmodesmata, root
hair, cotton-seed hair, and spiral thickenings even in
tracheids of healthy plants (Stone and Clarke 1992).
However, mature sieve-plate pores in monocotyledons
have little or no callose (Behnke 1971; Walsh and Evert
1975). The synthesis of callose can also be induced by
wounding, infection, and physiological stress (Kauss
1996; Stone and Clarke 1992). Callose is also deposited
in an apposition layer against the cell wall in diseased
tomato (Beckman et al. 1982). We found different
degrees of callose production during phloematic trypan-
osomatids infection. It is possible that a callosic apposi-
tion may seal the sieve plates, and that occlusions by
trypanosomatids and callose accumulation could func-
tionally disturb phloem transport in phloematic trypano-
somatid-infected coconut.
The sieve tubes of the phloem contain proteinaceous
structures, collectively called P-proteins (phloem pro-
teins), which accumulate in differentiating sieve ele-
ments and persist in translocating sieve elements. The
functional role of P-proteins remain unresolved. How-
ever, it seems likely that polymerized and unpolymer-
ized P-proteins exist in dynamic equilibrium within sieve
elements, where the concentration of each is responsive
to physiological changes within the vascular system
(Golecki et al. 1999). The P-protein is deposited initially
as ultrastructurally distinct polymorphous or crystalline
bodies during sieve element differentiation (reviewed in
Cronshaw 1975; Cronshaw and Sabnis 1990; Sabnis and
Sabnis 1995). Disruption of sieve elements during
wounding results in the accumulation of P-protein fila-
ments at the sieve plate, ostensibly blocking transloca-
tion via P-protein plugs (Hopkins 1995). In C. nucifera,
we identified two morphological kinds of P-like protein
in the sieve tube elements, and their accumulation at the
sieve plates was probably due to the infection by phlo-
ematic trypanosomatids and/or related vector feeding
that wounded sieve elements. P-protein plugs probably
affect long-distance movements of macromolecules in
phloem and impact the physiology of diseased coconut
trees.
In addition, phloem plastids have been reported to
explode in immediate response to a pressure release in the
sieve elements, which in turn frees substances that react
with the P-protein body material (Knoblauch and van Bel
1998). Starch and protein granules from broken plastids
could contribute to the plugging of the sieve tube elements
(Walsh and Melaragno 1981). Although the function of
plastid inclusions in monocotyledons is not clear, there is
evidence that they are involved in plugging mechanisms
(Eleftheriou 1990). Phloem plastids were highly visible in
uninfected vascular bundles of C. nucifera, but not in
infected ones. Our observations suggest that plastids frag-
ments in trypanosomatids-infected sieve elements of the
vascular bundle, also contribute P-protein that plugs the
phloem vessels in the presence of the protozoa. Injury to
the sieve elements by vectors could result in the same local
response.
In conclusion, our observations show that in symp-
tomatic coconut palms, at least 30% of sieve elements
of the phloem contain phloematic trypanosomatids or
80 J Gen Plant Pathol (2010) 76:74–83
123

P. staheli. In the terminal stage of disease, about 98% of
vascular bundles contain protozoa. In contrast, the path-
ogen was present in the vessel elements of the xylem only
in terminal stage of fatal wilt. These data strongly suggest
that the long-distance translocation of molecules in the
phloem is affected by the presence of phloematic try-
panosomatids, partially due to the plant response to the
infection.
J Gen Plant Pathol (2010) 76:74–83 81
123

Ackowledgements This research was supported by CNPq (Conse-
lho Nacional de Desenvolvimento Cientıfico e Tecnologico), FINEP
(Financiadora de Estudos e Projetos), FENORTE (Fundacao Estadual
do Norte Fluminense) and PRONEX (Programa de Nucleos de
Excelencia).
References
Agrios GN (2005) Plant pathology, 5th edn. Academic Press, San
Diego
Attias M, Bezerra JL, de Oliveira DP, de Souza W (1987)
Ultrastructure of Phytomonas staheli in diseased coconut (Cocosnucifera) and oil palm (Elaeis guineesis). J Submicrosc Cytol
19:93–100
Attias M, Resende MLV, Bezerra JL, de Souza W (1989) Occurrence
of Phytomonas staheli in Roystonea regia (royal palm). Microsc
Eletron Biol Cell 13:183–190
Beckman CH (1987) The nature of wilt disease of plants. APS Press,
St. Paul
Beckman CH, Mueller WC, Tessier BJ, Harrison NA (1982)
Recognition and callose in response to vascular infection in
fusarium wilt-resistant or susceptible tomato plants. Physiol
Plant Pathol 20:1–10
Behnke HD (1971) The contents of the sieve plate pores in
Aristolochia. J Ultrastruct Res 36:493–498
Bezerra JL, Figueiredo JM (1982) Ocorrencia de Phytomonas staheliMcGheee & McGhee em coqueiro (Cocos nucifera L) no Estado
da Bahia, Brasil (in Portuguese with English summary).
Fitopatol Brasil 7:139–143
Bezerra JL, Oliveira DP, Carvalho J (1983) Phytomonas staheli em
piacaveira (Attalea funifera Mart) na Bahia, Brazil (in
Portuguese). CEPLAC/CEPEC, Informe Tecnico 226. Bahia
Camargo EP (1999) Phytomonas and other Trypanosomatid parasites
of plants and fruit. Adv Parasitol 42:29–112
Camargo EP, Silva S, Roitman I, de Souza W, Jankevicius JV, Dollet
M (1987) Enzymes of ornithine-arginine metabolism in trypan-
osomatids of the genus Phytomonas. J Protozool 34:439–441
Child R (1974) Coconuts. Longmans, London
Cronshaw J (1975) P-proteins. In: Aronoff S, Dainty J, Gorham PR,
Srivastava LM, Swanson CA (eds) Phloem transport. Plenum
Press, New York, pp 79–147
Cronshaw J, Sabnis DD (1990) Phloem proteins. In: Behnke HD,
Sjolund RD (eds) Sieve elements, comparative structure, induc-
tion and development. Springer, Berlin, pp 257–283
da Cunha M, Gomes VM, Xavier-Filho J, Attias M, de Souza W,
Miguens FC (2000) The laticifer system of Chamaesycethymifolia. A closed host environment for plant trypanosomatids.
Biocell 24:123–132
Desmier de Chenon R (1984) Recherches sur le genre Lincus,
Hemiptera: Pentatomidae Discocephalinae, et son role eventuel
dans la transmission de la Marchitez du palmier a huile et du
Hart–Rot du cocotier (in French with English summary).
Oleagineux 39:1–6
Dollet M (1984) Plant diseases caused by flagellate protozoa
(Phytomonas). Ann Rev Phytopathol 22:115–132
Dollet M (2001) Phloem-restricted trypanosomatids form a clearly
characterised monophyletic group among trypanosomatids iso-
lated from plants. Int J Parasitol 31:459–467
Dollet M, Lopez G (1978) Etude sur l’association de protozoaries
flagelles a la Marchitez Sorpressiva du Palmier a Huile en
Amerique Sud (in French with English summary). Oleagineux
33:209–217
Dollet M, Giannotti J, Ollagnier M (1977) Observation de protozo-
aires flagelles dans les tubes cribles de Palmiers a Huile malades
(in French with English summary). CR Sci Acad Sci (Serie D)
284:643–645
Donovan C (1909) Kala-azar in Madras, specially with regard to its
connexion with dog and the bug (Chonorrhinus). Lancet
177:1195–1196
Eleftheriou EP (1990) Sieve elements. In: Behnke HD, Sjolund RD
(eds) Monocotyledons. Springer, New York, pp 60–91
Eschrich W (1975) Sealing systems in phloem. In: Zimmermann MH,
Milburn JA (eds) Encyclopædia of plant physiology. Springer,
Berlin, pp 3–56
Ferreira JMS, Warwick DRN, Siqueira LA (1998) Cultura do
coqueiro no Brasil (in Portuguese). EMBRAPA-SPI, Aracaju
Fig. 2 Microghaphs of Cocos nucifera. a Longitudinal section of
primordial leaf from ecotype Green Dwarf (type 2). Contiguous sieve
elements showing terminal sieve area and phloematic trypanosomat-
ids (arrows). In cell plate, cell wall (cw), callose apposition (c) and
fibril P-like protein in pore lumen (pl) can be identified. TEM.
b Transverse section of stem meristem from var. Malayan Green
Dwarf (type 3). The sieve element has crystal-like P-protein adhering
to the cell membrane (arrow). TEM. c Longitudinal section of stem
meristem from healthy ecotype Green Dwarf. Sieve plate (openarrow) has a slender callose deposit. Aniline blue. LM. d Longitu-
dinal section of stem meristem from var. Malayan Green Dwarf (type
3). Thick callose apposition in sieve plate (open arrows). Aniline
blue. LM. e Immunocytochemical analysis of callose. Control
reaction was carried out replacing primary anti-serum by pre-immune
serum. Compare with callose immunolocalization (f). Transverse
section of stem meristem from ecotype Green Dwarf (type 2). Note
sieve element cell wall (cw), callose apposition (c) and sieve pore
lumen (pl) without gold particles. TEM. f Immunocytochemical
analysis of callose. Transverse section of stem meristem from ecotype
Green Dwarf (type 2). Callose localization using antisera against
laminarin (b-1,3-glucan polymer). Anti-rabbit IgG antibody conju-
gated with 10 nm colloidal gold labeled mainly callose apposition (c)
in sieve element cell wall. Note sieve element cell wall (cw) and sieve
pore lumen (pl) without gold particles. TEM. g Transverse section of
sieve plate from ecotype Green Dwarf (type 2). Pathogen flagella
(open arrows) can be observed in two sieve pores lumen. Note callose
(c) apposition surrounding sieve pore (pl). CW cell wall. TEM. h High
magnification of (g). Parasite flagellum in sieve pore lumen shows a
typical 9 ? 2 pattern. Note callose (c) apposition surrounding sieve
pores and cell wall (cw). TEM. i Longitudinal section of sieve pore in
a sieve plate from ecotype Green Dwarf (type 2). Pathogen cell body
spanning both sides of the sieve pore. Note a large number of sub-
pellicular microtubules (black/white arrow). Callose (c) apposition
surrounding sieve pore. TEM. j Longitudinal section of sieve plate
(black arrow) from Green Dwarf (type 1). Pathogen cell bodies
spanning sieve pore lumen (open arrows). Note callose apposition
surrounding sieve pores. TEM. k Highly colonized sieve element of
stem meristem from Malayan Green Dwarf (type 3). Pathogen cell
body (rectangle) appears to have been moving through sieve pore
lumen. Note callose apposition surrounding sieve pores. TEM.
l Longitudinal section of stem meristem from var. Malayan Green
Dwarf (type 3). Close-up of rectangle indicated in (k). Pathogen cell
body in sieve pore lumen with numerous cytoplasmic vesicles and
mitochondrion. Note callose (c) apposition surrounding sieve pores
and cell wall (cw). TEM. m Transverse section of sieve element close
to sieve plate from Green Dwarf (type 1). Twisted pathogen cell
bodies (arrows) are evident in the sieve element. SEM. n Longitu-
dinal section of sieve elements from ecotype Green Dwarf (type 2).
Lateral view of terminal sieve plate. Twisted pathogen cell bodies are
preferentially oriented perpendicularly on both sides of sieve plate
(white arrow) where flagella were difficult to visualize; flagella of
free protozoa were easily identified (open arrows). SEM. Scale barsin a, h, i, l 1 lm; b, g 2 lm; c, d, k, n 10 lm; e, f 0.5 lm; j, m 5 lm
b
82 J Gen Plant Pathol (2010) 76:74–83
123

Golecki B, Schulz A, Thompson GA (1999) Translocation of
structural P-proteins in the phloem. Plant Cell 11:127–140
Harries HC (1977) The Cape Verde region (1499–1549): the key to
coconut culture in the Western Hemisphere? Turrialba 27:227–
231
Harries HC (1978) The evolution, dissemination and classification of
Cocos nucifera L. Bot Rev 44:265–320
Hong Z, Delauney AJ, Verma DPS (2001) A cell plate-specific
callose synthase and its interaction with phragmoplastin. Plant
Cell 13:755–768
Hopkins WG (1995) Introduction to plant physiology. Wiley, New
York
Johansen D (1940) Plant microtechnique. McGraw-Hill, New York
Kauss H (1996) Callose synthesis. In: Smallwood M, Knox JP,
Bowles DJ (eds) Membranes: specialized functions in plants.
Bios Scientific Publishers, Guildford, pp 77–92
Kitajima EW, Vainstein MH, Silveira JSM (1986) Flagellate proto-
zoan associated with poor development of the root system of
cassava within Espirito Santo State. Phytopathology 76:638–642
Knoblauch M, van Bel AJE (1998) Sieve tubes in action. Plant Cell
10:35–50
Louise C, Dollet M, Mariau D (1986) Recherches sur le Hartrot du
cocotier, maladie a Phytomonas (Trypanosomatidae) et sur son
vecteur Lincus sp (Pentatomidae) en Guyane (in French with
English summary). Oleagineux 10:437–449
Lucas JA (1998) Plant pathology and plant pathogens. Blackwell,
Oxford
McCoy RE, Martinez-Lopes G (1982) Phytomonas staheli associated
with coconut and oil palm disease in Colombia. Plant Dis
66:675–677
McGhee RB, Cosgrove WB (1980) Biology and physiology of the
lower Trypanosomatidae. Microbiol Rev 44:140–173
McGhee RB, McGhee AH (1979) Biology and structure of Phyto-monas staheli sp. n. a trypanosomatid located in sieve tubes of
coconut and oil palms. J Protozool 26:348–351
Moura JIL, Sgrillo RB, Miguens FC (2002) Manejo Integrado das
Principais Pragas do Coqueiro. In: Poltronieri LS, Trindade DR
(eds) Manejo Integrado das Principais Pragas e Doencas de
Cultivos Amazonicos (in Portuguese). EMBRAPA Informacao e
Tecnologia, Belem, pp 67–90
Northcote DH, Davey R, Lay J (1989) Use of antisera to localise
callose, xylan and arabinogalactan in the cell-plate, primary and
secondary walls of plant cells. Planta 178:353–366
Ohler JG (1999) Modern coconut management. Palm cultivation and
products. FAO, UK
Oliveira DP, Bezerra JL (1982) Ocorrencia de ‘‘marchitez sorpresiva’’
do dendezeiro no Estado da Bahia, Brasil (in Portuguese with
English summary). Theobroma 12:107–108
Oliveira DP, Bezerra JL, de Souza W, Attias M, Resende MLV
(1987) Progressos no estudo de Phytomonas staheli McGhee &
McGheee em dende na Bahia, Brasil (in Portuguese with English
summary). Theobroma 17:38–45
Pain NA, Green JR, Jones GL, O’Connell RJ (1996) Composition and
organization of extracellular matrices around germ tubes and
appressoria of Colletotrichum lindemuthianum. Protoplasma
190:119–130
Parthasarthy MV, van Slobbe WG (1978) Hartrot or Fatal Wilt of
palms. I. Coconuts (Cocos nucifera). Principes 22:3–14
Parthasarthy MV, van Slobbe WG, Soudant C (1976) Trypanosomatid
flagellate in the phloem of diseased coconut palms. Science
192:1346–1348
Perthuis B, Desmier de Chenon R, Merland E (1985) Mise en
evidence din vecteur de la Marchitez Sorpresiva du palmier a
huile, la punaise Lincus lethifer Dolling (Hemiptera: Pentatom-
idae, Discocephalinae) (in French with English summary).
Oleagineux 40:473–475
Resende MLV, Borges CEL, Bezerra JL, de Oliveira DP (1986)
Transmissao da Murcha de Phytomonas a Coqueiros e Dend-
ezeiros por Lincus lobuliger (Hemiptera, Pentatomidae).
Resultados Preliminares (in Portuguese with English summary).
Theobroma 16:148–154
Roeser KR (1962) Die Nadel der Schwarzkiefer-Massenprodukt und
Kunstwert der Naturen (in German with English summary).
Mikrokosmos 61:31–36
Sabnis DD, Sabnis HM (1995) Phloem proteins: structure, biochem-
istry and function. In: Iqbal M (ed) The cambial derivatives.
Gebruder Borntraeger, Berlin, pp 271–292
Stahel G (1931) Sur kenntnis der sierbrohrenkranheit (Phloemkrose)
des kaffeebaums in Surinam. I. Mikroskopiche untersuchungen
und infektionsversuche (in German). Phytopathology (Z)
4:65–82
Stone BA, Clarke AE (1992) Chemistry and physiology of higher
plant 1, 3-b-glucans (callose). In: Stone BA, Clarke AE (eds)
Chemistry and biology of (1, 3)-b-glucans. La Trobe University
Press, Bundoora, pp 365–429
Sturm NR, Dollet M, Lukes J, Campbell DA (2007) Rational sub-
division of plant trypanosomes (Phytomonas spp.) based on
minicircle conserved region analysis. Infect Genet Evol 7:570–
576
Vermeulen H (1963) A wilt of Coffea liberica in Surinam and is
association with a flagellate, Phytomonas leptovasorum Stahel.
J Protozool 10:216–222
Vickerman K, Dollet M (1992) Report on the second Phytomonasworkshop. Oleagineux 47:594–600
Wallace FC, Roitman I, Camargo EP (1992) Trypanosomatids of
plants. In: Kreier JP, Baker JR (eds) Parasitic protozoa.
Academic Press, London, pp 55–85
Walsh MA, Evert RF (1975) Ultrastructure of metaphloem sieve
elements in Zea mays. Protoplasma 83:365–388
Walsh MA, Melaragno JE (1981) Structural evidence for plastid
inclusions as a possible ‘sealing’ mechanism in the phloem of
monocotyledons. J Exp Bot 32:311–320
Waters H (1978) A wilt disease of coconuts from Trinidad associated
with Phytomonas sp., a sieve tube restricted protozoan flagellate.
Ann Appl Biol 90:239–302
Zucker WV (1983) Tannins: does structure determine function? An
ecological perspective. Am Nat 121:335–365
J Gen Plant Pathol (2010) 76:74–83 83
123