faunal analysis and meat procurement: reconstructing the sexual division of labor at shields pueblo,...
TRANSCRIPT

FAUNAL ANALYSIS AND MEAT PROCUREMENT: RECONSTRUCTING THE SEXUAL DIVISION OF LABOR
AT SHIELDS PUEBLO, COLORADO
Tiffany A. Rawlings B.A., University of Texas, Austin, 1994
M.A., University of California, Davis, 1996
DISSERTATION SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF
DOCTOR OF PHILOSOPHY
In the Department
of Archaeology
O Tiffany A. Rawlings 2006
SIMON FRASER UNIVERSITY
Summer 2006
All rights reserved. This work may not be reproduced in whole or in part, by photocopy
or other means, without permission of the author.

APPROVAL
Name: Tiffany A. Rawlings
Degree: Doctor of Philosophy
Title of Dissertation: Faunal Analysis and Meat Procurement: Reconstructing
the Sexual Division of Labor at Shields Pueblo, Colorado
Examining Committee:
Chair: Dr. Ross Jamieson
Dr. Jonathan Driver Senior Supervisor
Dr. A. Catherine D'Andrea Supervisor
Dr. Andrew Duff Supervisor
Dr. John Welch SFU
Dr. Patricia Crown Department of Anthropology University of New Mexico
Date Approved: ADK ?o/(%

SIMON FRASER ' U N M R S I ~ ~ I brary
DECLARATION OF PARTIAL COPYRIGHT LICENCE
The author, whose copyright is declared on the title page of this work, has granted to Simon Fraser University the right to lend this thesis, project or extended essay to users of the Simon Fraser University Library, and to make partial or single copies only for such users or in response to a request from the library of any other university, or other educational institution, on its own behalf or for one of its users.
The author has further granted permission to Simon Fraser University to keep or make a digital copy for use in its circulating collection, and, without changing the content, to translate the thesislproject or extended essays, if technically possible, to any medium or format for the purpose of preservation of the digital work.
The author has further agreed that permission for multiple copying of this work for scholarly purposes may be granted by either the author or the Dean of Graduate Studies.
It is understood that copying or publication of this work for financial gain shall not be allowed without the author's written permission.
Permission for public performance, or limited permission for private scholarly use, of any multimedia materials forming part of this work, may have been granted by the author. This information may be found on the separately catalogued multimedia material and in the signed Partial Copyright Licence.
The original Partial Copyright Licence attesting to these terms, and signed by this author, may be found in the original bound copy of this work, retained in the Simon Fraser University Archive.
Simon Fraser University Library Burnaby, BC, Canada

ABSTRACT
This study investigates the sexual division of meat procurement at Shields
Pueblo, a large aggregated village in the Northern San Juan region of Colorado,
occupied from ca. A.D. 725-1280. This is primarily achieved through analysis of faunal
remains in reference to the environmental, economic, and social factors affecting the
inhabitants of this region from Pueblo I (ca. A.D. 725- 900) until regional depopulation
ca. A.D. 1280. This dissertation supports previous research in the Northern San Juan
region regarding changes to the faunal pattern over time. It is noted that the Shields
Pueblo faunal assemblage is characterized by a decline in artiodactyl frequencies and
an intensification in utilization of lagomorphs and domestic turkeys, starting ca. AD.
1060.
A gendered analysis, using cross-cultural as well as Southwestern ethnographic
data, indicates an interesting pattern in the control/care/production of domestic animals.
Specifically, small household domesticates appear to be the responsibility of the female
head of household. Archaeological evidence of women's production of domestic meat
resources is investigated for Shields Pueblo. It is argued here that as environmental and
social factors changed and large game hunting declined, household-based economies
became more important. As these conditions changed, making large-scale game hunting
increasingly risky, women came to supply much of the community's meat (the majority in
many communities).
In conclusion it is suggested that as environmental conditions declined and the
threat of warfare and violence increased, there was a shift in the organization of labor in
regards to meat procurement. While large game was plentifuI/accessible, men were the
primary suppliers of meat for the community. As domesticated meat resources began to
dominate the pueblo economy, women's control of domestic turkeys allowed them to
attain more prestige -and thus power-- within the household and larger community.
Keywords:
Pueblo Indians-Antiquities
Animal Remains (Archaeology)
Archaeology-Theory-Gender
Social Archaeology
Shields Pueblo (Colorado)
iii

For Ruby, George, and Billie.

Acknowledgements
I would like to acknowledge the following people for all of their support: Dr.
Jonathan Driver for his patience and support (academic and financial) and for giving me
this opportunity to work in the Southwest again (thanks for taking me on) - a truly great
supervisor; Dr. Catherine D'Andrea for her awesome (sometimes painful) editing and for
her friendship and wisdom over the past 8 (and a bit) years - without your advise I would
not have had the courage to change my research topic; Dr. Andrew Duff for his insightful
(and encouraging) comments during the writing process; Dr. Patricia Crown and Dr.
John Welch for their help and advice on revisions (and for giving me some new ideas to
ponder); Crow Canyon Archaeological Center for their support while I worked on their
(never ending) faunal collection; Jude McLellan, Pei Pei Chu, Tracy Rogers, Karyn
Sharp, Jen Ramsay, and Chelsea Dunk for their friendship, and "moron support" through
the course of my degree program; my comrades in the Grad Lab for laughs, beers, and
for putting up with my million boxes of dusty bones; Robyn Banerjee and Lynda Przybyla
for their friendship and for helping me during all of my paperwork and bureaucratic
crises; Austin for being her big red furry self (give mama kisses); and last but not least
Mom and Dad for their expert data entry and for cracking that whip!

TABLE OF CONTENTS .......................................................................................................... APPROVAL ii ...
ABSTRACT ......................................................................................................... 111
DEDICATION ...................................................................................................... iv ..................................................................................... ACKNOWLEDGEMENTS V
TABLE OF CONTENTS ....................................................................................... vi ... ............................................................................................... LIST OF TABLES VIII
............................................................................................... LIST OF FIGURES x
CHAPTER 1 : INTRODUCTION AND RESEARCH DESIGN ................................. 1 ............................................................................................... Background -2
Potential Causal Factors for Culture Change ............................................... 3 Expected Faunal Patterning for Potential Causal Factors ............................ 9 Research Design ...................................................................................... 13
............................................................................... Analytic Design 15 Chapter Summary ..................................................................................... 16
CHAPTER 2: THEORY: FAUNAL REMAINS AND GENDER ............................. 18 ................................................................. Gender and Faunal Remains 1 9
Definition and Brief Review of Research ........................................ 19 .............................................................. Methods of Reconstruction 21
Problems with Ethnographic Analogy in Gender Studies .................. 28 The Sexual Division of Labor in the Ethnographic Record .......................... 30 Chapter Summary ..................................................................................... 39
........................................ CHAPTER 3: PHYSICAL AND CUTLURAL SETTING 42 ........................................................................................ Physical Setting 42
.............................................................................. Flora and Fauna 46 Soils and Geology ........................................................................... 49 Climate .......................................................................................... -49
................................................................................... Paleoclimate 50 .............................................................................. Archaeological Setting 53
............................................................. Archaeological Background 53 Excavation and Collection of Data ............................................................. 63
....................................................................................................... Dating 71 ..................................................................................... Chapter Summary 72
CHAPTER 4: SHIELDS PUEBLO FAUNA .......................................................... 74 ............................................... Methodology: Identification and Recording -74
............................................................................................ Quantification 75 Stable Isotope Analysis ............................................................................. 80
....................................................................... Sampling Procedures 81 ................................................................................... Taxon Frequencies 82
........................................................................................ Mammals 83 ............................................................................................... Birds 87
............................................ Reptiles. Amphibians. and Gastropods 88 .............................................................. Preservation/Taphonomic Biases 88
...................................................................... Natural Accumulation 89 ........................................................................ Canid Accumulation 90
..................................................................... Raptoer Accumulation 92

............................................................. Other Natural Accumulation 92 ........................................................................ Cultural Modification 93
Sullegic Processes (Archaeological Decisions) .............................. 101 ........................................... Post-depositional Destruction of Bone 102
Summary of Taphonomy of Shields Pueblo Faunal Remains ................... 105 ...................................................................... Skeletal Part Frequencies 1 0 6
Shields Pueblo Skeletal Part Frequencies ..................................... 107 ............. Age Estimation: Seasonality and Evidence of Turkey Production 111
.................................................. Change in Taxon Frequency Over Time 114 Artiodactyls Through Time ...................................................... 121 Lagomorphs Through Time ........................................................... 124 Turley and Large Bird Through Time ............................................. 125 Change in Fauna and Human Population ...................................... 126
..................................................... Spatial Patterning of Faunal Remains 130 ................................................................................... Chapter Summary 137
CHAPTER 5: SHIELDS PUEBLO IN A REGIONAL CONTEXT ......................... 140 .................................. Changes in Faunal Patterning at a Regional Scale 140
Pueblo I Sites (A.D. 750-900) ........................................................ 141 .................................................... Pueblo II Sites (A.D. 900-1 150) 1 4 5
.................................................. Pueblo Ill Sites (A.D. 1150-1300) 148 .............................................................................................. Discussion 152
Chapter Summary ................................................................................... 158
CHAPTER 6: GENDERED ANALYSIS OF FAUNAL DATA ............................... 160 Meat Procurement: Hunting .................................................................... 160
................................................................................... Turkey Production 162 .......................................... Spatial Evidence of Turkey Production 163
...................................................................... Turkey and Gender 166 .............................................................................................. Discussion 168
................................................................................... Chapter Summary 176
CHAPTER 7: CONCLUSIONS .......................................................................... 178 Research Expectations ........................................................................... 179
........................................................................... Faunal Variability 179 ...................................................... Hunted versus Tended Fauna -180
Population and the Decline of Large Game .................................... 184 ......................................................................... Potential Causal Factors 186
.................................................................................. Environment 186 ...................................................................... Population Pressure 188
............................................................................... Social Factors 189 Warfare ......................................................................................... 191
.............................................................................................. Discussion 192 ............................................ Evaluation and Suggestions for Research 1 9 5
REFERENCES CITED ..................................................................................... 200 APPENDICES .................................................................................................. 236
Appendix A: Crow Canyon Archaeological Center Zooarchaeological Identification and Recording Standards ............................... 236
Appendix B: Modified Bone Frequncies for a Series of Sites .................... 239
vii

Table 1.
Table 2.
Table 3.
Table 4.
Table 5.
Table 6.
Table 7.
Table 8.
Table 9.
Table 10.
Table 1 1.
Table 12.
Table 13.
Table 14.
Table 15.
Table 16.
Table 17.
Table 18.
Table 19.
Table 20.
LIST OF TABLES
Archaeological Evidence of Warfare
Tasks associated with the procurement and processing of crops
Northern San Juan Biotic Communities
Cultural Trends Summary: Northern San Juan Region and McElmo Dome
Excavation Block Summary: Structure Descriptions and Date Ranges
Relative Frequency of Faunal Remains by Class
Relative Frequency of Mammals
Relative Frequency of Bird Taxa
Frequency of Carnivore Modification- Shields Pueblo
Frequency of Carnivore Modification for Selected Taxa (site Comparisons)
Frequency of Complete and Fragmented Bone - Rodents
Culturally Modified Bone - Cut Marks
Culturally Modified Bone - Spiral Fractures
Frequency of Breakage for Selected Taxa at Shields Pueblo Compared With Other Sites on the Mc Elmo Dome
Culturally Modified Bone - GroundIPolished
Culturally Modified Bone - Burned
Frequency of Weathering Among Faunal Remains, Shields Pueblo 104
Frequency of Weathering Among Faunal Remains (site comparison) 104
Skeletal Part Frequencies - MNE counts by skeletal region for major taxa 108
MAU Values: Lepus (A); Sylvilagus (6); and M. gallopavo (C)
viii

Table 21.
Table 22.
MAU Values: Artiodactyls
Frequency of Weaker Long Bone Ends for Sylvilagus and Cynomys
Epiphyseal Fusion Frequencies (NISP) of Long Bones for Selected Species by FusionIAge Category
Table 23.
Table 24. Relative Frequency of Selected Taxa- Change through time
Table 25.
Table 26.
Taxon Frequency by Sub Period
Observed Frequency (NISP) for Selected Species by Sub Period
Table 27. Expected Frequency (NISP) for Selected Species by Sub Period
Table 28. Standardized Residuals for Selected Species by Sub Period
Table 29.
Table 30.
Table 31.
Rate of Pottery Deposition: Shields Pueblo
Rate of Faunal Deposition (NISP): Shields Pueblo
Frequency of Selected Taxa From Each Excavation Block
Standardized Residuals for Major Groups by Excavation Block
Table 32.
Table 33. Relative Frequency (NISP) of Selected Taxa and Pottery Weights: Structure vs. Non-structure
Northern San Juan Region Pueblo I Sites: Relative Frequencies and lndices
Table 34.
Table 35. Northern San Juan Region Pueblo II Sites: Relative Frequencies and lndices
Table 36. Northern San Juan Region Pueblo Ill Sites: Relative Frequencies and lndices
Table 37.
Table 38.
Rate of Deposition of Eggshell
Isotopic values for samples of jackrabbit (LEP), cottontail (SYL), and turkey (MEG) collagen

LIST OF FIGURES
Figure 1. Map of the Southwestern Region of the United States 43
Figure 2. Map Showing the Location of Shields Pueblo (5MT 3807) 44
Figure 3. McElmo Drainage Area Map 45
Figure 4. Climatic Reconstruction: Southwest U.S. A.D. 600-1400 52
Figure 5. Shields Pueblo 67
Figure 6.
Figure 7.
Figure 8.
Figure 9.
Figure 10.
Figure 1 1.
Figure 12.
Figure 13.
Frequency of Cultural Modification for Selected Species
Relative Frequency of Complete and Fragmented Bone
Frequency of Degree of Fusion for Selected Species By FusionIAge Category Taxon Frequency by Sub Period
Artiodactyl lndices per Sub Period
Lagomorph lndices per Sub Period
Turkey lndices per Sub Period
Relationship between the artiodactyl index and lagomorph index
Figure 14.
Figure 15.
"Corrected" TurkeyILarge Bird lndices
Change in Relative Depositional Rates of Ceramics and Fauna Over Time
Figure 16. Specific Taxonomic lndices and Pottery Deposition Rates Over Time
Figure 17.
Figure 18.
Frequency of Taxa per Excavation Block
Frequency of Taxa and Pottery Weights: Structure vs. Non-structure
Taxonomic Frequencies: Pueblo 1, Northern San Juan Region
Figure 19.
Figure 20. Taxonomic Frequencies: Pueblo 11, Northern San Juan Region
Figure 21. Taxonomic Frequencies Pueblo Ill, Northern San Juan Region

Figure 22. Relative Rate of Deposition of Eggshell per Sub Period Compared to Pottery Deposition
Figure 23 Macaw Boxes
Figure 24. Turkey and Macaw Boxes.

CHAPTER 1
INTRODUCTION AND RESEARCH DESIGN
Recent research on the Ancient Puebloans (formerly known as Anasazi) of the
Four Corners region of the American Southwest has brought to light an intriguing pattern
of population expansion and contraction. Dramatic increases in population and site
aggregation were followed by periods of depopulation and decline (Cordell 1984; Cordell
and Gumerman 1989; Gumerman 1988; Larson and Michaelson 1990; Leonard and
Reed 1993; Varien 1999). This pattern of expansion and contraction is found throughout
the greater American Southwest, and is particularly well defined in the Mesa Verde
region (the Northern San Juan) from AD. 900 to 1300. The Northern San Juan region
was largely depopulated around A.D. 1300.
In order to better understand the cycle of expansion and depopulation in the
Mesa Verde region, Crow Canyon Archaeological Center began a research project
entitled, Communities through Time: Migration, Cooperation, and Conflict. This project
was designed to collect artifactual and ecofactual evidence from residential structures
occupied between AD. 900 and 1300. Shields Pueblo was chosen for study because it
is believed to be a key site in the Goodman Point community. It also has a long
occupation spanning Pueblo I (AD. 725- 800); a period of depopulation (A.D.800 -
1020); and resettlement (AD. 1020) with continuous habitation until the site was
depopulated permanently at approximately A.D. 1280 (Duff and Ryan 2000).
This dissertation presents the faunal data from Shields Pueblo in order to
understand how meat procurement strategies and gendered labor roles changed in
response to environmental and social changes. It brings together several lines of
evidence, such as faunal, ethnographic, warfare theory, and intraregional comparison to
trace social change at Shields Pueblo over the entirety of its inhabitation. Explanation

based on these lines of evidence should contribute a better understanding of changing
labor roles at the regional level.
Background
There is a commonly noted trend in faunal assemblages from sites in the
northern Anasazi area during Pueblo Ill (A.D. 1150-1300) consisting of a distinct drop in
the amount of Artiodactyla (mainly deer) and other large mammalian game compared to
preceding periods (Driver 1997, 2002, Driver et a/. 1999; Muir 1999). During this time
period there was also greater dependence upon domesticated turkey (Meleagris
gallopavo) and upon lagomorphs (particularly Sylvilagus sp.). During Pueblo Ill Munro
(1994) describes a great increase of domestic turkey remains in the archaeological
record indicating intensification of production. There was also a rise in human population
and a general aggregation of habitation sites (a movement from small dispersed
settlements into fewer, larger settlements) (Varien 1999). In addition, researchers have
postulated an increase in evidence of warfare (interpersonal violence) (Upham and Reed
1989; LeBlanc 1999; Lightfoot and Kuckelman 2001), cannibalism (Hurlbut 2001;
Lambert, Billman, and Leonard 2000; Ogilvie and Hilton 2000;Turner and Turner 1999;
White 1992), and larger numbers of defensive sites (Adams 1989; Lightfoot and
Kuckelman 2001). This period of apparent upheaval and change should be
accompanied by changes in social organization. Changes in social organization have
been addressed in regards to settlement patterning and developing social and site
hierarchy. A few studies have specifically focused on labor organization (e.g., Leonard
and Reed 1993). This dissertation takes this a step further by discussing changes in
social organization in relation to changes in the sexual division of labor.

In this dissertation I will discuss how meat procurement strategies and the
associated labor roles were modified in response to environmental and social changes
at Shields Pueblo. I will attempt to bring together several lines of evidence,
predominantly through gender studies. I argue that men's labor in meat procurement
(namely hunting) declined and was supplemented by women's labor (through the
production of domestic turkey), as men focused their energies on warfare and protection.
Results will be placed in a regional context. The object of my research is to trace these
changes in the northern part of the American Southwest.
Potential Causal Factors for Culture Change
There is a long and varied history of research in the Southwest, covering all
areas of scientific enquiry. Due to a wealth of cultural, environmental, and geophysical
data, the Southwest as a region is well understood. Culture change has been explained
in varying ways depending upon the popularly accepted paradigms of the time. Over the
past two decades three basic kinds of models have been used to explain culture change
in the American Southwest: 1) environmental (drought driven) change (e.g., Dean and
Robinson 1977,l 978), 2) population growth and circumscription (e.g., Rohn 1989; Dean
et a/. 1994), and 3) social circumstances (e.g., Driver 1996, 2002; Leonard and Reed
1993; Lipe l992a; Rohn 1989; Rautman 1993). The first two are typical prime mover
hypotheses arguing that a single physical change in either environment or in population
growth drove a number of other cultural changes. The latter type of argument takes into
consideration a series of factors, mainly in the internal or social realm, that would have
led to a change in archaeological patterning.
The argument that environment is a driving force in culture change has been
popular since the 1970s. The most commonly cited causal factor for regional change is
the "great drought", which was a period of prolonged cool and dry weather that

effectively decreased the amount of productive farmland in the Southwest (Dean and
Robinson 1977, 1978). This dry period began at the end of the 13th century (during the
later part of the Pueblo Ill period) from A.D. I276 to 1299 (Lipe l995:159). The "great
drought" is believed to have been caused or exacerbated by the beginning of the Little
Ice Age (Petersen 1986, 1988, 1992). This was a cooler period that shortened the
growing season, by decreasing the number of frost free days, and interfered with the
summer rainy season, thus decreasing the amount of moisture at the most crucial point
in the maize agricultural season (Petersen 1992). The basic premise of the
environmental argument can be outlined as follows. First, favorable climatic conditions
of the 12th century allowed for an increase in population, which would have created a
greater dependence upon maize agriculture (Dean et a/. 1994; Larson et a/. 1996).
Once the drought began there was a decline in the availability of arable farmable land,
which would have caused a certain amount of population circumscription around
productive locations (Schlanger 1988). Finally, the Anasazi depopulated the Mesa
Verde area and moved into more productive environments with more predictable
resources (Fish et a/. 1994; Gumerman 1994; Schlanger 1988).
There is quite a bit of debate associated with the environmentally driven theory of
culture change and population movement. Some researchers question the severity of
the "great drought ." While individuals such as Petersen argue that the Mesa Verde
Region was completely inhospitable for dry farming (1 986, l988), others have shown
that there were large enough areas of productive land to support local populations (Van
West and Lipe 1992). More recent work has shown that depopulation of the Northern
San Juan Region began before the onset of the so-called "great drought" (Varien et a/.
1996; Lipe and Varien 1999b). In fact, this movement of peoples from the eastern
portion of the Southern Colorado River Basin to the western units began during Pueblo II
(early to mid 12th century) (Lipe and Varien 1999a: 263). Obviously factors other than

simple environmental change affected the movement of peoples and their archaeological
signatures as migrations began before negative climatic change.
Population pressure models have also been used to explain culture change in
the American Southwest. Duff and Wilshusen (2000) argue that there have been
several population shifts (i.e., depopulation in one area and migration into another).
These depopulation episodes occurred between A.D. 375 to 575 and also A.D. 880 -
950 (Duff and Wilshusen 2000:169; Wilshusen 1999; Wilshusen and Ortman 1999).
There were also smaller regional shifts in population. The Dolores region seems to have
been depopulated during the early A.D. 800s (Orcutt et al. 1990:200) and there were
more widespread, short-term depopulation events from A.D. 1070 - 1100 and 11 50-
1 170 (Varien 1999: 191 -1 92, 201). Between these episodes of depopulation were
periods of repopulation. The greatest periods of repopulation occurred between A.D.
575 - 880 and from A.D. 950- 1300 (Duff and Wilshusen 2000: 169). Duff and
Wilshusen (2000) suggest that each of these repopulations may represent settlement of
different cultural groups. This suggests that culture change recognized in the
archaeological record (such as change in ceramic styles) is due to the influx of "new"
cultural groups.
Over-population is often cited as a reason for regional collapse (e.g., the
Northern San Juan Depopulation of 1300). Population is believed to have reached its
peak at the Pueblo II/III transition (Rohn 1989; Dean etal. 1994). Dean et al. estimate
that there were 5,500 people living in Southwestern Colorado and Southeastern Utah at
A.D. 11 50 (1994: Figure 4.1 : 59). Duff and Wilshusen (2000:173) give a much higher
estimate, suggesting a total of 23,224 for Pueblo II and 21,808 for Pueblo Ill for the
Northern San Juan region. This increase in human population is argued, by Duff and
Wilshusen, to have resulted in resource stress and to have caused increasing
competition for highly valued resources, especially large game (Munro 1994). It is

precisely this scarcity of game, which may have forced people in the Northern San Juan
to intensify the production of domesticated animals (i.e. Meleagris gallopavo). While no
one debates the occurrence of regional depopulation ca., A.D. 1300, the rate of
depopulation is unclear. Traditionally, Southwestern archaeologists have argued that
this depopulation event was sudden and dramatic (Gladwin 1957). However, Duff and
Wilshusen (2000) suggest that this depopulation was more gradual, over a number of
generations. This makes sense given the drop in population seen over the span of
Pueblo Ill at many sites (Duff and Wilshusen 2000:189). Regardless of the rate of
depopulation, population pressure -exacerbated by environmental and social factors- is
an important aspect of culture change.
Social explanations are often co-explanatory with demographic change. Social
explanations include changes in social organization and settlement patterning (Driver
1996,2002; Leonard and Reed 1993; Lipe 1992; Rohn 1989; and Rautman 1993).
Recently, archaeologists have begun to examine the development of social hierarchy
and political/economic power as a force in culture change. For example, Hayden
focuses on the control of prestige-linked resources or goods (1 995), the idea being that
archaeologists should be able to recognize a social hierarchy based upon the
distribution of certain rare or "expensive" artifacts. In the Southwest, several burials
contained copper bells, conch and other shell ornaments, cloisonne decorated objects,
macaw skeletons of various ages (indicating trade with Mexico), and local rarities (such
as turquoise and rare pigments) (Lister 1978; Mathien 1981; Reyman 1978). Most
burials contained far more common goods (ceramic pots, bowls, beakers, lithic tools,
bone or shell beads, etcetera). This discrepancy has been argued to be evidence of the
development of social hierarchy and even a connection with the Toltec empire (Lister
1978; Mathien 1981 ; Reyman 1978).

Changes in resource productionlprocurement and the organization of labor may
account for the aggregation of populations into large sites during the mid to late A.D.
1200s. While Leonard and Reed (1993) deal specifically with agriculture, their
hypothesis can explain some of the reasons behind the development of large pueblo
villages. They argue that aggregation was caused by changes to labor organization,
which in turn was attributable to specialization. In order to specialize in the production of
agricultural products, a greater investment in labor is required (i.e., an increase in the
number of laborers). When aggregation no longer becomes economically successful
(due to environmental or cultural factors) or if less labor is required, groups will disperse
into smaller, scattered settlements (Leonard and Reed 1993). Their argument is based
purely upon whether or not certain behaviors are efficient or inefficient given certain
environmental and cultural conditions. This paradigm uses energy expenditures and
energy gain (calories burned in pursuit of an activity and calories gained through the
results of the activity) to explain culture change. In other words, groups will behave in
the most energetically efficient ways. Leonard and Reed focus solely on the decision
making process and how people organize themselves physically. Their approach does
not take into account power structures or belief systems, which often influence groups to
behave in inefficient ways.
A social explanation that has recently attracted attention cites increases in
warfare and violence as an explanatory factor for culture change. This increase in
warfare certainly could explain the appearance of defensive sites and the aggregation of
smaller villages into larger entities (Lightfoot and Kuckelman 2000). Warfare can be
defined as "a state or period of armed hostility existing between politically autonomous
communities, which at such times regard the actions (violent or otherwise) of their
members against opponents as legitimate expressions of the sovereign policy of the
community" (Meggitt l977:lO). For many years anthropologists have been reticent to

use the term "war" in the context of less politically complex groups. Instead, "raiding" has
been used to describe conflict between groups, even those extending over long periods
of time. Keeley (1996) has pointed out that anthropologists have shied away from using
terms such as warfare because they are intent upon portraying aboriginal communities
(past and present) as "noble savages" living in harmony with the world around them.
According to Keeley, this belief completely ignores the fact that warfare in non-state
societies often claims the lives of 20 to 30 percent of the men and 2 to 7 percent of
women (Keeley 1996; Meggitt 1977). These proportions are substantially higher than
those seen in state level societies. Therefore, discounting warfare among less
hierarchically organized groups makes it seem as if these groups did not suffer gravely
from the effects of intergroup violence.
Warfare as a topic of study in the Southwest has primarily focused on proving
that it existed. Wilcox and Haas (1994:211) begin their chapter on warfare by stating,
"we explore the current state of knowledge about this issue, arguing that such processes
did exist and that explanations of the evolution of Southwestern societies must reckon
with them ." Evidence of conflict has been described in the Southwest since the time of
Kidder (1924). Most early research focused on ways to identify warfare in the
archaeological record; for instance, demonstrating that many sites are located in
defensible locations (Hodge 1877; Cushing 1882; Hewitt 1906). Kidder (1924) took this
a step further, by trying to identify the "enemy ." The Athapaskans were Kidder's choice
because they were outsiders to the Pueblo cultures and would have caused aggregation
into larger villages for defense (Kidder 1924). Other infiltrating "outsiders" have also
been named, such as the Numic speaking hunter-gatherers of the Great Basin (Upham
1984), or more mobile Anasazi "hill" peoples raiding the agricultural villages in the river
valleys and drainages (Wilcox and Haas 1994).

Archaeologically, evidence of warfare can be recognized by the type of
architecture present on a site, by specific types of artifacts associated with fighting
(including weapons and defensive tools - such as shields) (Geib 1990; Morris and Burgh
1954), evidence of violent death (Crotty 2001 ; Turner and Turner 1999; White 1992;
Wilcox and Haas 1996), location of sites on the landscape, and by the presence of large
tracts of uninhabited "demilitarized zones" (Farmer 1957; Rohn 1975; Rowlands 1973;
Wilcox and Haas 1996; Winter 1981). Archaeological evidence of warfare is
summarized on the following table (Table 1).
Causes of warfare follow along the same lines as other changes in culture, such
as competition over scarce resources (LeBlanc 1999). Others have argued for the so-
called "bad apple" effect first described by Keeley (1996). Essentially, it only requires
one combative group to begin attacking their neighbors to cause movements of
populations out of harms way and into aggregated or defensible villages. While this
describes how populations might react to violence, it does not necessarily explain the
cause for the initial belligerence. Revenge has also been cited as cause for warfare (for
example, Basso 1971). While revenge certainly explains the continuation of the cycle of
warfare, it does not explain the initial act of conflict.
Expected Faunal Patterning for Potential Causal Factors
If aridification is invoked as a factor of change in the faunal record during Pueblo Ill, then
there should be a decline in large game at all sites within the study area. Additionally,
there should be higher frequency of large game at sites located near permanent water
sources or near particularly productive ecological zones. As far as domesticated turkey
is concerned, the expectation is that its frequency will increase in areas where large
game declines, as turkey is a controllable source of meat (Driver 2002; Muir and Driver
2002). If less beneficial climatic shifts are causal factors in the change of the faunal

Table 1. Archaeological Evidence of Warfare
1 Towers -as defensive beacons (Winter 1981)
Stockades- surrounding farmsteads (Rohn 1975)
surrounding a central plaza (Wilcox and Haas 1996)
Trincheras I hillside retreats- act as "trenches" for keeping out of "enemy fire" while shooting arrows (Wilcox 1989)
features, generally walled (Farmer 1957:250)
shields (Morris and Burgh 1941)
Body Armor- artiodactyls ribs strung in overlapping layers over the torso
Fending Sticks-1 -1.5 meter curved sticks reportedly used to deflect darts (Geib 1990)
Burnedldestroyed sites- such as Salmon Ruin with complete floor contexts and burned bodies (Wilcox and Haas j 996) Rock Art depicting violence- figures holding heads and shields or flayed head skins, bows and arrows (Crotty, 2001 )
Violent injury- blows to the head or face
Dismemberment- violent removal of limbs or the head
Proj.pts. embedded in bone
Scalpinglflaying marks on cranium and face
Headless bodies or isolated heads (indicative of trophy- taking)
Cannibalism- (also associated with perimortem violence and "mistreatment" of -emains
smaller villages located at the access points to larger settlements (Wilcox and Haas j 996) No-Man's Lands - or demilitarized zones- acting as political boundaries between conflicting groups (Rowlands 1973)
pattern, then there should be similar declines in large game for each climatic shift.
If population growth and pressure are causal factors in the change in the faunal
record for Pueblo Ill, a drop in large game and communally hunted prey in areas with the
highest populations and an increase in the frequency of turkey would be expected.

Smaller sites and sites located in sparsely populated areas should then have higher
frequencies of large game because there are fewer people competing for limited game
resources, thus there should be less reliance on turkey
Driver (1996) builds upon Hayden's (1995) prestige and power model to explain
changes in the intersite distribution of faunal remains during Pueblo Ill. Large, regional
centers during this time period have a higher frequency of large game, while smaller
communities have lower frequencies of Artiodactyla or none at all. He argues that large
aggregated communities controlled or owned game resources and that smaller
communities would not have had access to Artiodactyla. This pattern seems to indicate
controlled access to large game. Smaller sites show an intensive reliance on
domesticated turkey as a source of meat, as evidenced by a significant jump in turkey
frequency. Along with this trend there is a decrease in communally hunted jackrabbits,
which seem to be replaced by cottontail rabbits, which can be taken by single individuals
from horticultural fields near the pueblo (Driver 1996, 1997, 2002). If social
differentiation is the cause of the faunal shift in Pueblo Ill, then we should find differential
distribution of highly valued game resources. In this instance, large, powerful social
groups would have higher frequencies of large game (but lower frequencies of less
valuable game such as Lepus). Smaller, outlying sites should then show far lower
frequencies of large game and higher frequencies of lagomorphs and turkeys.
Another social hypothesis that may account for higher frequencies of artiodactyls
in large communities is feasting. Blinman (1989) and Potter (1997, 2000) have been able
to identify the presence of feasting in the archaeological record. Prior to A.D. 1275
feasting has been argued to have been competitive in nature, with larger households
within communities or larger communities vying for prestige or economic power (Blinman
1989; Potter 2000). Competitive feasting, or feasting with the intent of creating or
maintaining social differentiation can be identified in the archaeological record by higher

frequencies of large game, serving vessels, and other prestigeltrade goods concentrated
in certain (generally larger) households. After A.D. 1275, Potter (2000) argues, feasting
became a means of defining and reinforcing group identity. During late Pueblo Ill,
evidence of feasting is typified by a concentration of large game and serving vessels in
religious structures as opposed to habitation rooms. This change in context is believed
to indicate a shift in religious beliefs and social behavior (Potter 2000).
Given the expected faunal patterning described by Driver (1996), it would be
difficult to distinguish between community control over large game resources and
feasting at large communities. Both behaviors would leave the same faunal patterning,
and may, in fact be closely tied together. If larger communities are acting as locations
for solidarity feasts, they might also act as a point of organization for communal hunting
for smaller surrounding sites.
With specialization as a pull to culture change, a specialization in faunal
resources (as noted above) is expected. So, there should be less diversity in species
over-all with a focus on turkey because they are the most dependable meat resource (as
they are raised directly by groups and not as easily influenced by environmental factors)
and the easiest to increase in production through "herd" management (selective
breeding and culling the population to increase meat output). Across the board, turkey
should be the predominant species at sites with all other species (including large game)
playing lesser roles in late Pueblo Ill as hunted game populations fall.
If we consider warfare as a causative factor in culture change in the Southwest,
we expect to see all of the evidence listed in Table 3. If warfare was endemic, then
there should be a change in faunal patterns over time and from site to site. There
should be a decrease in hunted game acquired long distances away from habitation
sites, particularly in sites with smaller populations. If there was a risk of attack, there
would have been a greater need for protection. Smaller sites (with smaller populations)

would be disinclined to allow their strongest warriorslhunters to leave the community
unguarded for long hunting excursions. Conversely, we might expect the frequencies of
large hunted game to remain relatively constant in large regional centers because they
would have large enough populations to allow enough guards to remain behind while
hunters left on expeditions.
It is also possible that the people of the Northern San Juan were acting as
raiders and not simply being attacked by "outsiders". If this is the case, then raiders
should have greater frequencies of large game (or other forms of "booty). Given the
higher frequencies of large game at large sites in the region, it could be possible that
these communities were, in fact, raiding smaller sites in the region. However, given the
examples of mass violence associated with the "abandonment" of particular large sites
(such as Sand Canyon Pueblo and Yellow Jacket Pueblo) this is unlikely. If these larger
sites were the aggressors you would not expect violence at such a scale, nor the
resulting closure of these sites. An interesting study in the frequency of violent injuries in
the La Plata region shows an interesting pattern of multiple healed cranial depression
fractures and poor health among a large portion of young women in the population
(Martin and Atkins 2001). While the reason is unclear, Martin and Atkins (2001) suggest
that this may indicate that these women were war captives brought to the community
from elsewhere, as neither men nor children from contemporary contexts show evidence
of that level of violence and poor treatment. It is possible then, that people from the La
Plata region were raiding other communities in the Northern San Juan.
Research Design
The goal of this dissertation is to reconstruct the gendered division of labor at
Shields Pueblo, Colorado. In order to do this, it is important to distinguish between
tended fauna (small household domesticates) and wild, hunted fauna. This research is

designed to answer a series of questions: 1) what is the archaeological patterning for
hunted versus tended fauna; 2) what is the relationship between hunted and tended
fauna over time (with particular regard to Pueblo 11, Ill, and the Pueblo IV transition); 3)
does this pattern reflect a change in the sexual division of labor; and 4) what are the
possible causes of this change in faunal patterning and cultural behavior? By answering
these questions I hope to establish a series of guidelines for the reconstruction of the
sexual division of labor regarding faunal procurement. Ultimately, my research should
be able to offer some refined hypotheses for the change in faunal patterning seen
throughout the Anasazi Region.
In order to address the changes in meat procurement and their likely causal
factors, there are three basic questions that must be asked:
1. What is the change in species frequencies from Pueblo I to Pueblo Ill?
2. Are there meaningful relationships between hunted and tended prey frequencies
through time? In other words, what is the relationship between frequencies of hunted
and tended fauna?
3. Is there a decrease in large mammals at Shields Pueblo over time, and if so, is this
the result of a decrease in hunting or of short-term residentiallpopulation decline?
Given the general changes to cultural patterns in the Northern San Juan region
specific characteristics are to be expected in the faunal assemblage. First, there should
be a drop in Artiodactyla and communally hunted Lepus and an increase in the
frequency of Meleagris gallopavo and Sylvilagus (in relation to Lepus). Second,
huntedlwild fauna (procured away from the settlement) and tended fauna (kept within
the confines of the sitelhousehold) should have different dietary patterns. Whether an
animal was eating local wild plant foods, or domesticated maize should be clearly
evident by comparing carbon isotope levels. Tended fauna should have carbon isotope
ratios indicative of a maize diet if they are being kept by women in the household;

hunted game (such as jackrabbits) should have C isotope ratios indicative of a diet
consisting of local C3 (or temperate) vegetation (Katzenberg and Kelley 1991).
Cottontails might show a mixed diet of local flora and maize from the fields (Katzenberg
and Kelley 1991). Third, there should be a decline in hunted game in the assemblages
from Pueblo II to Pueblo Ill and further decline or even absence of large game in late
Pueblo Ill. As hunted prey declines, there should be a statistically meaningful increase
in turkey.
Finally, regional demographic studies and changes in ceramic sherd weights for
Shields Pueblo will be used as a proxy of population growth to ensure that drops in
faunal frequencies are not due to declines in population at this site. Previous regional
population estimates indicate that there was an increase in population starting in Pueblo
II, a peak in early Pueblo Ill, and the start of decline in late Pueblo Ill (Lipe 1995,
Schlanger 1988, Varien 1999, 2002). Therefore, a drop in Artiodactyla frequencies in
the Shields Pueblo faunal record should indicate a decline in large game hunting or
availability, and not a decline in population.
Analytic Design
In order to address these research questions I will make comparisons on several
different scales. First it is necessary to address the change in frequencies of hunted
versus tended game over time at Shields Pueblo and contemporary sites of different
sizes. By doing this, a sense of what is happening at Shields Pueblo can be understood
within a larger regional framework. Reconstructing labor in reference to meat
procurement is dependent upon cross-cultural or comparative research. The cross-
cultural survey is intended to sample a wide range of communities in order to identify
any broad trends in the care of domestic animals. The cultures studied here are typically
household based economies, producing crops primarily for their own sustenance with

limited surplus going to market for profit. Both household domesticates and (to a lesser
extent) herd animals are considered in order to illustrate patterns in the division of labor.
Isotopic analysis has also been employed to reconstruct the diet of tended fauna.
This line of analysis is helpful in establishing whether turkeys are fed from household
stores of food or being allowed free forage. This distinction in diet has implications for
the identity of domestic animal caregivers. All of the theories and methods used to
reconstruct the sexual division of labor for Shields Pueblo are described in detail in the
following chapter
Chapter Summary
The purpose of the Shields Pueblo project is to better understand population
expansion and depopulation in the Northern San Juan Region. This cycle of expansion,
contraction, and eventual depopulation of the Northern San Juan region causes certain
changes in the Anasazi cultural pattern, as well as to the natural environment. Typically,
the Northern San Juan faunal assemblages are characterized by a distinctive drop in the
frequency of large game species and an increase in lagomorph and domestic turkey
during the Pueblo Ill period (A.D. 1150-1300). This trend has been variously argued to
have been environmentally driven (by a regional drought); an example of over-hunting
due to population growth and circumscription; a result of change in social organization;
and a result of increasing warfare and violence in the region.
Each of these explanations creates its own expected faunal pattern. If regional
drought is the cause of the drop in large game, then there should be a noticeable decline
in the frequency of large game at all sites in the region. If population pressure is the
causal factor, then there should be a drop in large game in areas with the highest
population densities. If social or site hierarchy is responsible for the faunal trend, then
large game should be more frequent at large sites in the region. Warfare as a causal

factor would result in a decrease of game hunted far away from villages and an increase
in locally hunted and domestic species.
The purpose of this dissertation is to describe the faunal patterning of the Shields
Pueblo assemblage, to relate these changes in species frequencies to changes in labor
organization based upon sex, and ultimately, to test the expected faunal patterns
hypothesized for each possible "causal factor." This will be done by asking the following
research questions:
1) What is the change in species frequencies from Pueblo I through Pueblo Ill?
2) What is the relationship between hunted fauna and tended fauna through time?
3) Is there a decrease in large mammals at Shields Pueblo over time? If so, is this a
result of a decline in hunting large game or the result of a decline in population?
These questions will be answered by comparing the Shields Pueblo fauna at the site
level and at the regional level.

CHAPTER 2
THEORY: FAUNAL REMAINS AND GENDER
Faunal remains are typically used to reconstruct economic and environmental
conditions, particularly subsistence strategies. These often include "menus", or lists of
the animal sources utilized as food (Armelagos 1994) and seasonal information based
upon the species and age structure of the faunal population (Monks 1981). However,
more recently archaeologists have begun to use faunal remains to reconstruct social
patterns. Crabtree (1 990) illustrates how faunal remains can be employed to study trade
relationships between groups, status difference between individuals, and ethnicity.
A more recent volume, Animal Bones, Human Societies (Rowley-Conwy 2000),
provides case studies from around the world that discuss social organization based upon
faunal remains. In the Anasazi region of the American Southwest, faunal remains have
been used to reconstruct inter-site hierarchies, indicating which sites controlled access
to resources, and ritual structures (identifying particular categories of animals with
particular ritual structures) (Driver 1997, 2002; Muir 1999). Some archaeologists have
begun to examine faunal remains from a gendered perspective. Szuter (2000), for
example, describes evidence of male and female activities associated with particular
animals in the American Southwest. Faunal remains have traditionally been associated
with "male" activities (such as hunting), but it is becoming clear that women and
women's labor also greatly affect the faunal record. This chapter reviews the theoretical
approaches used in the interpretation of faunal remains for Shields Pueblo as reflecting
the sexual division of labor.

Gender and Faunal Remains
To date much of the research associated with reconstructing the sexual division
of labor in an archaeological context has focused on artifact use and manufacture
(Brumfiel 1991 ; Gero 1991 ; Nelson 1997: Rice 1991 ), the evolutionary or biological
aspect of gender roles (Mc Brearty and Moniz 1991 ; Zihlman 1989, 1991 ) and spatial
use or patterning (Conkey 1991 ; Hastorf 1991 ; Hendon 1997; Kent 1998; Tringham
1991). A significant portion of the literature consists of critiques of androcentric
research. These critiques call for archaeologists to focus on women and their activities
in the archaeological record. Faunal remains have often been associated with men
(illustrated by much of the research presented in "Man the Hunter" [Lee and DeVore
19681) and are not typically seen as food resources gained through women's labor
(Gilchrist 1999). This is a blatant shortcoming in economic and social archaeology. The
case studies and examples drawn upon in this dissertation show unequivocally that
women were active in meat procurement across many cultures both in the distant and
recent past.
Definition and Brief Review of Research
Division of labor can be understood in different contexts. The term, division of
labor, simply refers to how groups of people assign different tasks to group members.
Categories of labor are divided into levels or into series of often overlapping groupings.
Labor is generally divided among individuals based on sex, age, or status. Undoubtedly
the most basic criterion for division is sex (Durkheim 1964; Halperin 1993; Shapiro 1983;
Shore 1981). Many use the terms division of labor and sexual division of labor
interchangeably. In anthropology the division of labor has traditionally been viewed as a
biological construction (Harris 1971 ; Lee and De Vore 1968). Men were always the

hunters, makers of tools, and inventors in society, while women cared for children and
collected plant materials near camp (Keesing 1975).
Ruth Bunzel (1 938) was the first anthropologist to argue against a biological
basis for labor division. She successfully argued that while men hunted or herded large
animals in some cultures, women often herded large animals in other non-western
cultures. Bunzel also pointed out that women often had a much heavier workload than
their male counterparts.
More recently Bender (1 989) and Cucchiari (1 986) have discussed the differing
influences of biology and culture on the divisions of labor. This research has a distinctly
gynocentric view, meaning that the researchers have focused solely on the activities of
women. Gifford-Gonzalez (1 993) argues that even the "male" activity of hunting is
influenced by women. Decisions about how meat is butchered and which portions are
transported home are often dependent upon the culinary practices of a group. Other
research focuses on cases of women not only influencing, but taking on non-traditional
roles, such as that of "hunter" or "warrior" (Davis-Kimbal 1997; Muller 1985;Shostak
1981 ; Wadley 1998); or discusses cases of men performing "female" tasks (Hays-Gilpen
and Whitley 1998; Joyce 1997). Wadley, in her 1998 study of women in the Stone Age
of South Africa, discusses several cases of women acting as hunters in addition to
gathering plant materials. In Nisa: The Life and Words of a !Kung Woman, Nisa gives
several accounts of snaring or trapping birds and tortoises as well as hunting steenbok
and small kudu (Shostak 1981). Among more socially complex groups there is evidence
of women having participated in warfare. Davis-Kimball (1997) describes a series of
burials in the Sauramatian culture of the Eurasian steppe, dating between 600 to 400
B.C., which contain women buried with large quantities of weapons. One young woman
showed evidence of long hours on horseback (in the form of femoral deformation and

hypertrophied muscle markings). Celtic women were also recorded as having been
warriors as well as leaders of great armies (Muller 1985:95).
In addition to women acting in "male" roles, men have been shown to take on
"feminine" activities and roles. Roscoe (1991) describes the life of a Zuni berdache
(Man-Woman), We'wha, a man who assumed the role of a woman, adopting women's
work and dress. It is fairly widespread among various American Native groups, to
believe that these individuals (sometimes referred to as two spirits) hold special powers
and thus, they are greatly respected within their groups (Joyce and Classen 1997).
Basically, we have come to understand that not only are labor roles influenced by
culture, they are influenced by the definition and assignment of genders within groups
and are highly variable. How groups constitute gender is a historical and cultural
contingency, which may depend upon biological sex, sexual orientation, and
agelreproductive fertility (Conkey and Gero 1991 ; Shapiro 1983). While great strides
have been made in including gendered research in archaeology, two decades of
engendered archaeology has really not advanced our understanding of how societies
function as complete units. Most gender literature is a critique of male-biased hunting
and lithic research and does not always provide workable methods or models for
reconstructing the sexual division of labor. For example, Conkey and Gero (1991);
Conkey and Spector (1 984); Gilchrist (1 999); ; and Wylie (1 992) all provide eloquent
critiques of androcentric archaeology, but do not discuss the methodology used in
gendered archaeology.
Methods of Reconstruction
Reconstructing the sexual division of labor through faunal analysis is not possible
to do with simple taphonomic studies. It relies on the addition of various archaeological
methods to understand social as well as taphonomic contexts. One method employed to

reconstruct labor organization is that of analogy. Analogy is often used as a middle-
range approach to understand how past activities and behaviors are "fossilized" in the
archaeological record. Analogy is seen by some archaeologists as a flawed method of
social reconstruction (Stahl 1992; Wobst 1978). There are two important issues: 1)
whether it is appropriate to reconstruct the behavior of prehistoric peoples based upon
analogies to present day peoples and 2) what are the effects of uniformitarianist thinking
on the interpretations being made (Stahl 1992; Wobst 1978)? Wobst (1978) is the most
vocal opponent of the use of analogy in archaeology. He argues that the ethnographic
record is inherently lacking in essential observations about spatial and temporal factors.
He goes on to argue that observations made by anthropologists are biased by the
constraints of ethnographic field work, and feels that the archaeological record is
superior to the ethnographic one because it is composed of "the precedents and
products of actual behavior, rather than recorded behavior" (Wobst 1978:303). This, of
course leaves the question, "How does one reconstruct past behavior?" While there are
many critics of analogy it has been shown to be highly effective in the reconstruction of
past behaviors. Some researchers have argued that biases or other shortcomings in the
ethnographic record may be overcome through ethnoarchaeological research. Stark
(1993) argues that ethnoarchaeology is critical for both cross-cultural and context
specific archaeological interpretation. However, she does criticize ethnoarchaeologists
for failing to clearly define methodological frameworks in their research. She argues that
in order to produce the kinds of results being sought, methodology must be standardized
to address archaeological questions. Stark suggests that research be more reflexive, or
based upon longer sequences of fieldwork in order to develop "a dialectic between
techniques of field based data collection and generalization in research design over an
extensive period of time" (1 993:lOO). While ethnoarchaeological research is not

possible for all gender research, it is probably one of the most widely used frames of
research in gender archaeology.
Analogies are formed through historic ethnographic accounts, contemporary
ethnoarchaeological research, and to some extent experimentation. Janet Spector
(1 983) was one of the first archaeologists to develop a workable analogical framework
for reconstructing the division of labor. Spector's approach, called the Task
Differentiation Framework, focuses on the material parameters of gender arrangements.
The framework refers to four aspects of task performance: the social, temporal, spatial,
and material realms. A condensed framework of Spector's 1983 Hidatsa study is
presented in Table 2.
Table 2. Tasks associated with the procurement and processing of crops
Garden clearing
Planting
Women of lodge; occasionally old men Women of lodge; assistance from related women from other lodaes
(based upon Spector 1983:86)
Weeding Women of lodge; occasionally assisted by old men
Crop Protection Young girls of lodge, usually two
Gardens (314 mile from summer village) Gardens (314 mile from summer village)
Gardens (314 mile from summer village)
Gardens (314 mile from summer village)
Mornings; Spring and Early summer
Early summer
Dailylall day; late summer- harvest in fall
Hoes, rakes, and digging sticks
Hoes, rakes, wooden bowls
Hoes, rakes
Scarecrows, watching stage, ladder, associated cook hut and cooking equipment

While Spector's framework provides ideal guidelines for research development, her
approach is unwieldy and very time consuming, because Spector's framework involves
recording so many lines of data simultaneously. Ideally, all tasks would be recorded, so
that there would be a framework describing the activities and actors in every single
social and economic behavior. In order to build Spector's task framework, an
ethnoarchaeologist would have to spend an extended period of time with the subjects
and would need to have unhindered access to both male and female subjects. Few
archaeologists have followed her approach exactly. Most research uses only certain
aspects of Spector's framework, by focusing on a single activity or a single suite of
activities (such as meat acquisition [Brumbach and Jarvenpa 19971 or craft production
[Nelson et a/. 20021).
Following Spector's framework closely, Brumbach and Jarvenpa (1997) interview
men and women among the present day Chipewyan. Their informants were asked
questions about the social, spatial, temporal, and material dimensions of food
procurement and processing activities. They also used direct observation to take note of
the above aspects of meat acquisition behavior. Brumbach and Jarvenpa outline a
series of systems (e.g., how specific animals are hunted and processed) with specific
sets of patterns allowing archaeologists to reconstruct a range of activities. Among the
Chipewya, both men and women are active in meat procurement, although women tend
not to kill large game as frequently as men. Men and women begin hunting during their
teens. Women give this activity up, or hunt less frequently, when they start to raise a
family. As their children reach maturity, women typically re-enter the hunting scene.
Brumbach and Jarvenpa also make an important point concerning spatial analysis within
habitation sites. As it turns out, women's hunting activities may be easier to identify than
men's given the fact that women tend to go after smaller game, closer to camp, and

which are transported back to camp whole for further processing. Men's hunting of large
game typically takes them far away from habitation areas, with individuals from the
community moving out to the kill site to process the animals (Brumbach and Jarvenpa
1997:30). This is an important distinction that will be addressed below in relation to
care of domestic animals.
Ethnographic analogy and ethnoarchaeology have been used to show that
shellfish are typically collected by women. Classen (1991), in her study of the Shell
Mound Archaic (some 8,000 years B.P.) in the American Southeast, integrated the
ethnographic record of shellfish gathering with burial analysis. She shows that female
burials within the immense shell mounds are far more likely to contain red ochre than
male burials and include more "ceremonial objects." She goes on to argue that this is
evidence of the importance of women in this society as providers of shellfish resources.
Christine Szuter's "Gender and Animals: Hunting Technology, Ritual, and
Subsistence" (2000), follows the changing relationship between gender and animals and
how these changes affect the sexual division of labor. Szuter focuses on the American
Southwest in a larger regional sense. Her research loosely follows Spector's Task
Differentiation Framework in the sense that Szuter attempts to record all the activities
and materials involved in the acts of hunting, other forms of subsistence, and ritual. She
is able to illustrate the vast quantity of activities involved in the preparation, completion,
and end results (i.e., garbage) of these activities in the archaeological record through the
use of ethnographic and historic accounts.
Szuter is able to distinguish between expedient hunting, carried out by men,
women, and children, and more formalized hunting, which may have been dominated by
men or involved the community as a whole. Small animals, birds, reptiles, amphibians,
rodents, and lagomorphs (excluding jackrabbits) were easily caught with traps and
snares near residential areas. People generally procured small game while in the

process of performing other activities such as hunting, horticultural work, or collecting
other wild resources. Because of this, formality and ritual are not associated with this
form of hunting.
Ethnographic evidence from the American Southwest and the Great Basin show
that jackrabbits were hunted communally in large drives (Anell 1969; Lange 1959;
Parsons 1918, 1920, 1921, 1970, 1977; White 1932). This activity may have been
carried out by one or more communities in cooperation and usually consisted of men,
sometimes accompanied by women and even children driving, trapping, and clubbing
jackrabbits in large numbers (Lange 1959; Parsons 1918). Studies of images of small
game trapping on Mimbres bowls (Shaffer et a/. 1996) indicate that men were the sole
trappers of wild birds and other small game.
Large game hunting is very different from the above activities because of the
strong correlation with ritual activity and the prestige of the participants. Hunting of large
game is associated primarily with men. During the Archaic period and Early Pueblo
periods of the Southwestern chronology, hunting probably provided most of the meat
consumed by villagers (Driver 2000). Muir (1999:66-94) notes that hunted animals
(artiodactyls as well as some wild birds) were deposited in patterned or ritualized ways.
Both Szuter (2000) and Muir (1 999) describe the deposition of artiodactyls in ritual
contexts and the display of elements of the skeletons of hunted animals as trophies. In
this context, we can argue that (at least in the Southwest) large game in the
archaeological record is evidence of men's activities and participation within a prestige
or ritual system. Some Mimbres ceramics have also shown men in the act of hunting
large game. A bowl recovered from Old Town, New Mexico shows a pair of hunters in
pursuit of a deer (Darryl Creel, personal communication).
In addition to hunted animals, smaller animals (namely turkeys, macaws, dogs,
and even snakes) were tended by pueblo communities. Szuter argues that these tended

animals were not being used as a food source, but were employed for other purposes
(i.e., dogs for stalking game and ritual burial, turkeys to produce animal products such as
feathers for arrows, and macaws, turkeys, and snakes for ritual purposes) (Hill 2000;
Szuter 2000). Szuter mainly focuses on archaeological sites in the Mimbres and
Hohokam regions of the Southwest, where turkey and other small, tended animals did
not seem to figure prominently as food sources. Because of a lack of historic or
ethnographic accounts of who was caring for these animals, Szuter looks at depictions
of animal tending on Mimbres ceramics and the inclusion of these animals in burial
contexts. The majority of Mimbres pots portray women holding or being associated in
some way with macaws and sometimes turkeys, while men are typically associated with
hunting activities or ritual activities (Munson 2000). Others have noted the connection
between women and birds in both pottery as well as iconography. VanPool and
VanPool (2006) are able to show that particular iconographic designs are used to depict
differences between the sexes. Their study of Casas Grandes effigy pots illustrates that
bird designs, such as the double-headed diamond macaw motif as well as other tutelary
bird images are always associated with female effigy pots (VanPool and VanPool
2006:69).
Other evidence of women's connection with birds can be seen in Puebloan
mythology and associated iconography. The story of Uretsete and Naotsete, sister
deities, tells the story of their battle over peopling the Rio Grande region (Patterson-
Rudolph 1993). The younger sister Uretsete (the winner of this contest) is often
depicted in iconographic images (petroglyphs, sand paintings, kiva murals, etcetera) as
a bird woman, and often specifically as a turkey (Patterson-Rudolph 1993). Part of this
myth is also telling in regards to care of domestic birds. At one point in the myth
Uretsete makes the first corn woman fetish ('lariko') in order to heal her people from an
epidemic, "She wound thongs of deer hide about an ear of corn and placed at its top

feathers of a turkey, which at her order had shaken out its feathers for her" (Patterson-
Rudolph 1993:64). While this is by no means solid evidence of who was tending small
animals within the pueblo, it is a compelling line of evidence providing further support for
the association of women, domesticated birds, and ritual.
Problems with Ethnographic Analogy in Gender Studies
By far the biggest problem with reconstructing gendered behavior is the lack of
ethnographic accounts of women's activities. It is very common in the historical and
anthropological literature for researchers to completely overlook women, by discussing
only male activity or by assuming the reader would simply know what women do in
society. Because so many of the detailed ethnographies of Southwestern Pueblos were
undertaken in the late 19th and early 20th century, women's labor was often subsumed
under a catch-all heading such as "household duties ." Research tended to focus on
very public, highly visible activities (ritual events, celebrations, and communal work
groups). While there were several well known female ethnographers working in the
Southwest (such as Pearl Beaglehole, Florence Hawley Ellis, Elsie Clews Parson,
Matilda Cox Stevenson, and others) there was still a general, biased assumption that
certain aspects of women's work was less interesting because it was associated with
day-to-day living and not with "important" activities such as ritual events. Because of the
lack of ethnographic reports of specific women's activities in the Southwest, it becomes
necessary to look outside of the geographic area to illustrate patterns in the sexual
division of labor as it applies to faunal food sources. In order to employ this kind of
cross-cultural survey, it is necessary to include a wide range of cultures. While broad
survey may not take all cultural or economic differences into consideration, it does allow
one to identify larger patterns of behavior across cultures. In this case, the cultures are
all primarily household-based economies -although some may engage in market

economy to a small extent- and are typically farmers or gardeners. They hunt game
animals (which are associated with prestige) as well as tend domesticated animals.
Tended animals (for the purposes of this study) are those kept within the boundaries of
the household. Animals held in communal pens, and animals kept outside the confines
of the household are also considered as a "contrast" to tended fauna (although it is
interesting to note that many herded animals spend some portion of the day in the realm
of the household for milking, or for overnight protection from predators).
Unfortunately, many ethnographers do not specify which individuals care for
domesticated animals (for example most of the ethnographic reports encountered during
the research phase of this dissertation simply stated that certain domestic animals were
owned or kept by households). In many cases the authors list all of the domestic
animals kept, then go on to discuss male activities and female activities, leaving the
domestic animals out of both discussions.
Predicting how groups divide labor based upon some grand universal schematic
is a difficult (if not impossible) task. However there does seem to be strong patterning in
meat provision cross-culturally. Most discussions about meat provisions for the
household focus on male hunting practices (among Southwestern Puebloan groups for
example: Anell 1969; Beaglehole and Beaglehole 1964; Ellis 1959; Henderson and
Harrington 1914; Lange 1959; Parsons 191 8,1920,1921,1970,1977; Stevenson 1894;
White 1932, 1974a, 1974b). While there are certainly instances of women hunting large
game (Shostak, 1981), it is primarily a male activity associated with ritual and prestige.
The next section in this chapter will show that animals housed in or near the household
(whether domesticated or wild captured) tend to be the responsibility of the female head
of the household. Meat procurement is not a single-sex task, therefore any
reconstruction of the sexual division of labor should first and foremost account for all of
the actors within a labor system.

As with most archaeological research we cannot simply use one line of evidence.
This is especially the case with faunal analysis. Reconstructing social behavior with
faunal remains requires the judicious use of spatial analysis in combination with
ethnographic or experimental research to fully comprehend the social systems and
behaviors at work. As with most archaeological research, analogy must play an
important role in the development of models and the explanation of patterns within
faunal assemblages. However, this enterprise should be undertaken with caution. It is
important to be critical of the ethnographies used as well as to realize that the groups
studied do not represent "fossilized" cultures, but rather living societies who have
changed throughout their own histories and been affected by western or colonial
influences (Stahl 1992; Trigger 1982; Wobst 1978).
While many ethnographic studies are discussed below, it must be noted that
some accounts are rejected. Some sources are considered outdated or contaminated
(either by misinformation or by racist ideology). Either way, these resources are
discussed, but are identified as flawed. This does not mean that the sources are
completely falsified, simply that all of the information given therein may not be accurate,
or may be biased towards a certain perspective. Therefore, these resources should be
viewed with caution.
The Sexual Division of Labor in the Ethnographic Record
There are numerous cultures discussed in this cross-cultural survey who
maintain domestic animals. Communities from around the world are discussed in
relation to their care of particular animals. Care of pigs is discussed first, as this subject
has the most published material. Smaller household domestic animals are discussed
next and compared to care of herd animals. Finally, the pattern of animal care is
outlined and some examples are given of cases where this pattern is not followed.

Melanesian societies, because of their economic behaviors and their strictly
defined gender roles have been the focus of many ethnographic and
ethnoarchaeological studies (i.e., Feil 1984; Hampton 1995; Koch 1970; Oliver 1955;
Rubel and Rosman 1978; Strathern 1971 ; Strathern 1972; Thiessen 2001). These
groups, such as many communities sampled in this research, raise domestic household-
based animals as well as hunt wild animals. Both domestic and wild animals are used
for food and for prestige or religious activities. This area of the world is well known for its
strict rules of behavior concerning men and women's activities (Strathern 1972).
Melanesia is also famous for its keeping and ritualization of domestic pigs. While the
prestige and feasting system is grand in scale among Melanesian groups (such that it
dominates their economic and social spheres), this should not undermine its use as an
analogy for the Anasazi. The key economic activities (such as raising garden crops,
"storage" of these surpluses in the form of domestic animals, and prestige-linked
activities such as hunting and warfare) are similar enough to warrant comparison.
Among most Melanesian groups, each household keeps a separate herd of pigs.
They are typically allowed to forage during the day and are penned or closed up in the
houses in the evenings (Feil 1984; Oliver 1955; Strathern 1971 ; Strathern 1972). In
villages where no domestic pigs are kept, wild piglets are often captured when their
mothers are hunted (Rubel and Rosman 1978: 13). These piglets are then kept and
raised and are used as forms of currency and for ritual such as domestic pigs elsewhere
in Melanesia.
A survey of ethnographic literature of the region provides some insight into day-
to-day activities and how responsibilities are divided between the sexes. In all cases
where specific sex was cited, women were responsible for the care (keeping, herding,
and feeding) of pigs whether wild captive or domesticated (Rubel and Rosman 1978;
Strathern 1971 ; Strathern 1972). While men may hunt wild pig and capture their young,

the care of these animals falls upon the women who take great pride in their charges.
Captured pigs are often named, and addressed by kinship terms, and thus have very
special relationships with the women who keep them (Rubel and Rosman 1978:13).
During ceremonies and other feasting events men act as the formal participants. They
certainly take credit for raising and contributing pigs to feasts, but as Strathern points
out, "they rely on their wives for most of the labor in pig-production" (1971:g). Among
the Hagen of the Western Highlands District of New Guinea, establishing and
maintaining a large herd of pigs is tantamount to increasing a woman's status. Women
undertake all aspects of care, although men will sometimes take the pigs out to forage if
conditions outside the village are dangerous (M. Strathern 1972: 18-1 9).
Pigs are an interesting domesticate in that they can either be allowed freedom for
foraging (as among many Melanesian groups) or they can be penned within or near
living structures (Elmendorf 1976; Feil 198; Nemeth 1998; Oliver 1955; Strathern 1971 ;
Strathern 1972). Groups that keep pigs penned, tend to place the responsibility of care
and feeding upon women. Mayan groups have also been recorded as having kept
domestic pigs (Elmendorf 1976). It is typical for a Mayan woman to hold half a dozen or
so pigs within her household. She is the caregiver and reaps the financial benefits when
her pigs are sold at market (Elmendorf 1976). In the American Southwest, there are very
few references in the ethnographic literature concerning the keeping of pigs by Pueblo
groups (and only one very brief comment on who fed and cared for them), however, pigs
were kept and their care further illustrates women's role in raising domestic fauna.
Schneider and Roberts (1965:70) in Zuni Daily Life fleetingly mention the female head of
the household taking a basket of corn out to feed the pigs "Mo51 comes from NER with
a basket of corn for the hogs."
Small housebound domesticates (animals that are kept within the house itself or
are kept in enclosed coops) are the most obvious animals to be kept by women because

they are most obviously kept within the sphere of the household. In the Andes, guinea
pigs live underfoot in the kitchens and under sleeping platforms. They are kept in the
kitchen for a number of reasons: to keep them warm, to keep them accessible for meals,
and for the traditional belief that guinea pigs need smoke (Morales 1995). Guinea pigs
are raised both for household consumption and for sale in local markets. Traditionally,
guinea pigs (or cuy) were fed wild collected greens and domestic grain by the woman of
the household (Tschopic 1946: 521). In the modern setting, cuy are fed alfalfa as well
as table scraps, potato peels, carrots, gathered grass, and fresh corn cobs (Morales
1995). Cuy may be kept outside of the kitchen. Some individuals build adobe structures
to house their cuy. This is more typical of the recent trend in larger scale cuy production
for sale at markets. Cuy are a very valuable commodity in Peru and Ecuador and even
in New York City (due to the influx of immigrants from South America). Women may
make as much as 100,000 pesos a month (approximately $1 25) from the sale of their
cuy (Morales 1995:28). Many communities have established co-ops, referred to as
"mujer-cuy" (woman- guinea pig) programs. These are moneymaking ventures. Men
often boast of how productive their wives are. Women in these communities are very
proud of their contributions to their families. Many even claim (in jest) that they now
"wear the boots" in the family (Morales 1995:29).
Other small household domesticates include poultry. Poultry (chickens, turkeys,
geese, ducks, guinea fowl, et cetera) are important basic household staples across
many cultures. Among the Maya of the Yucatan and Quintana Roo, chickens and
turkeys are raised for sale and for consumption during ceremonial and feasting
occasions (Kintz 1990; Villa Rojas 1945:262). Women are the sole caregivers for these
animals, and in fact claim ownership of them separately from their husbands (Elmendorf
1976:lOO). Such as guinea pigs, poultry are raised for family consumption and for
market sale. Any money made by women from the sale of domestic animals is theirs

and may go into a household fund or may be kept separately for their own use
(Elmendorf 1976; Villa Rojas 1945). Other Central American cultures (namely the Carib
or Black Carib) kept turkeys as domestic animals. According to historical period
documents, traditionally, labor was clear\y divided between the sexes. Men's primary
occupation was hunting and fishing, while women tended domestic animals and
performed domestic chores, including agricultural labor (Gullick 1976). Even with the
introduction of European plantations, new agricultural products and wage labor in the
mid 1 8th century through to the mid 2oth century, women remained the sole tenders of
domestic animals (turkeys, chickens, and hogs) (Gulllick 1976:60).
The division of labor among the Kanuri of Bornu in Niger is primarily based upon
sex and age (Cohen 1967). Women typically cook, draw water from the well, tend
gardens in the household compound, and keep both chickens and ducks, while men
primarily engage in activities outside of the house compound (i.e., participate in politics,
work in the fields, trade or slaughter cattle, work as laborers, etcetera) (Cohen 1967:75).
Women among the Buganda produce food for the household, growing various garden
crops and by keeping flocks of chickens, along with sheep and goats (Fallers 1964:82).
Many of the African ethnographies encountered in this research are problematic. Childs
(1 949) and Dyson (1 934) are considered to be outdated and racist (Childs was a
missionary and Dyson often used derogatory or racist language in his descriptions).
However, they are briefly discussed here, as I have not been able to locate recent
studies that specifically mention the identity of caregivers of household domestic
animals. These should be considered corrupt sources. Childs (1949), Dyson (2002
[1934]), and Skoggard (2001) state that wives, among the Ovimbundu of Angola,
maintained their own huts, granaries, and kept flocks of chickens within the larger
polygamous household units.

Caring for small household domesticates is looked at as an extension of a
woman's care for her family. Small domesticates are fed the leftovers of the family's
daily fare. This is typical of the daily routine for women in many parts of Eastern and
Southeastern Asia. Women in rural areas of Thailand, Borneo, and Korea begin their
days by preparing breakfast, feeding their families, and then feeding their chickens,
dogs, and pigs with the leftovers (Amyot 1976; Gomes 191 1 ; Sorensen 1988). Because
the women handle and prepare the food for the human household members, they do the
same for the animals.
Other rural communities throughout Europe and the Near East follow the pattern
of women being tied to the household and the care of domestic animals kept therein
(Bringa 1995; Dubisch 1986; Gilliland 1986; Herzfeld 1985; Kremensek 1983; Pavlovic
1973; Walker 1974). In most instances, women are morally expected to remain in the
house or yard surrounding the house and outbuildings. Even in modern times, women
are often expected to maintain traditional behaviors. For example, in small Yugoslavian
towns, there remains a clear division of labor. Men work outside of the household for
wage labor or in agricultural fields. Women are responsible for all household activities
(housework, cooking, gardening, childcare, and tending domestic animals) (Gilliland
1986). This connection to the household is often tied to the notion that women are
somehow polluting to male activities. Being tied to the household controls women and
their connection with the outside world. In rural Greece a "proper" or "moral" woman
spends the majority of her time in the household, or associated with domestic duties
(including tending domestic pigs and chickens) (Dubisch 1986). In Crete, there is even a
linguistic distinction between male-tended animals and female-tended animals. The
term "meat" applies only to flock animals (sheep in this case) tended by men. Chicken is
considered to be a "domestic product" (e.g., associated with the household and women's
work) and, therefore not manly and not "meat" (Herzfeld 1985).

Among herding societies, there is some variation concerning the sex and age of
the herders. Some herding societies consider the activity of taking animals into the bush
to be exclusively male. Beaglehole (1 937) notes that herding was considered a male
activity among the Hopi of the American Southwest (despite what Fewkes reports in
1890 [cited in Cushing et a/. 19221). Beaglehole also notes that care of "chickens"
(which may be a term indicating all poultry) comes under the jurisdiction of women
(1 937: 18-1 9). Beaglehole is yet another source that must be viewed with caution. The
information given is considered to be corrupt or inaccurate on many accounts. Again, it
is included because it is the only Southwestern source that directly links women with the
care of poultry. In other groups, it is often children or young women without children
that are sent to watch the flocks. The Tarahumara of Chihuahua and the Sierra Madres
often rely of the labor of children and unmarried women to watch the herds of goats and
sheep (Bennett 1935; Kennedy 1978; Pennington 1963). It is clear, however, that when
herding away from home is dangerous (either because of terrain or animallhuman
predators) men will take over this activity (Kennedy 1978). The age and sex of the
herders depends upon the availability of labor within the household and the concept of
danger outside the household. Despite the fact that females may be involved with
herding under certain conditions, day-to-day care of domestic animals seems to be
exclusively performed by women among the Tarahumara. Among the Ayamara of
Bolivia, women and children perform herding as well as daily husbandry (Tschopik
1946). Domestic animals are housed near the households for protection, thus daily
feeding and care falls to the women of the household. The rural communities of the
Jebala Mountains of Morocco keep cattle and donkeys as domesticates. While men
generally use these domesticated animals as beasts of burden (for threshing and
plowing), women and young girls are responsible for their care (Moreno-Garcia
2004:328). Other herding societies, such as Bosnian Muslims, follow this same pattern

(Bringa 1995; Lockwood 1975). There is a split between female and male tasks
associated with herd animals; men traditionally herded animals and women cared for
them while they were stabled and were responsible for milking and cheese making
(Bringa 1995).
Most modern agrarian communities, whether they herd or keep animals in owned
pastures, keep multiple species of domesticated animals. Even the Saami (or Lapps),
known for keeping large herds of reindeer, keep smaller domesticates as well. While,
there is some fluidity concerning which sex participates in any given activity, it is more
common for women to handle household duties (including raising sheep, goats, cows,
chickens and other fowl, and horses) while men are away from the premises with the
reindeer herds (Anderson 1978). In the Kragujavac region of Sumdaija, Serbia there is
a very strong tradition of male herding and hunting and female tending of small
domesticates. Keeping chickens, geese, ducks, and turkeys is done exclusively by
women. Poultry are kept in the courtyard of the household and are fed mixtures of water
and flour, cooked greens, and often wet cornbread (Pavlovic 1973:22). Cattle, sheep,
and goats are herded by men and young adolescent males. Pigs may also be herded to
forage in the woods during the day (by young boys), but at night are penned and cared
for by the woman of the house (Pavlovic 1973:56). It should also be noted that hunting
is still a common practice among rural Serbs. While hunting is ancillary to domestic
animal production, it is an extremely important activity for men and can greatly affect
their position in the community (Pavlovic 1973). This is also the case among various
Asian groups such as the Toradja of Indonesia (Adriani and Kruijit 1951), the Khasis of
India (Mc Cormack 1964), the preindustrial Okinawan of Japan (Glacken 1953), the Sea
Dyak of Borneo (Gomes 191 I) , the Monguors of China (Schram 1954), and Korean
farmers (Sorensen 1988). The Khasis of India, for example, keep both small and large
animals. Men keep the cattle and goats and hunt wild game, while the women keep

chickens and ducks (Mc Cormack 1964). The Toradja of the Central Celebes of
Indonesia do the same. Men keep buffalo in addition to hunting as a sign of prestige,
while women keep chickens and pigs (Adriani and Kruijt 1951).
In all cases, the household and many of the things associated with it are
considered the realm of the woman. This is nicely exemplified in how the croft (or
household) is divided in rural Scotland. The croft (the house and all of the land owned or
farmed by the family) is split into male and female domains (Walker 1974). The
woman's domain includes the interior of the house, the front yard, the barn and the yard
(where peat is stored and where the chickens are housed). Women care for the
domestic animals (including chickens, cats, dogs, and dairy cows) while they reside in
female space (Walker 1974:61). The male domain includes the area that surrounds the
croft (the agricultural fields, peat bog, and pastures). Men perform all of the duties that
take place in this realm of the household (including cutting and hauling peat,
maintenance of the buildings, and herding and caring for cattle and sheep while they are
out to pasture) (Walker 1974:61).
There is a very clear pattern in all of this. Any animal that is kept in the
household for meat or secondary products comes under the care of the female
household head. This is particularly evident when domestic animals are fed kitchen
scraps (or food prepared especially for them) by the female head of the household.
There are some exceptions to this pattern, particularly with the care of birds. When birds
are not being kept for food they may come under the care of men. In the American
Southwest, eagles are captured and cared for exclusively by men (Beaglehole and
Beaglehole 1964; Ellis 1959;Lange 1959; Parsons 1970, 1977; Stevenson 1894). In this
instance the birds (raised for their feathers exclusively) are used in ritual. There is also a
certain amount of prestige gained by men who are able to capture eagles, as it is
considered a dangerous activity (Parsons 1920). Other instances of male care of birds

can be found among the Tiv and the Bemba of Africa. In these cases cocks and pigeons
are kept as signs of prestige (Bohannan 1968, Richards 1939). Such as the Puebloan
examples, these birds are reared for religious sacrifice (and in the case of the Tiv for
Cock fighting) (Bohannan 1968). In these cases, however, household domestic animals
(for consumption or household related ritual) were always cared for by women. Care of
herd animals is often split between males and females. Males, in most cases, tend the
animals while they are outside of the household, where as women take over feeding and
extraction of secondary products (e.g., milk) while the animal is housed within the
household. There is a very distinctive split between female spheres of influence (the
household) and male spheres of influence (the outside world) in every single case
discussed above. The particulars of ownership (i.e., owned by men, women, or the
household in general) may differ, however it is clear that women are responsible for the
care of household domestic animals. Given the strength of this pattern among the
surveyed cultures, it is reasonable for analogue characteristics (discussed on pages 28
and 29).
Chapter Summary
The purpose of this chapter is to review the theoretical approaches to gender
research and to suggest a method for the reconstruction of the sexual division of labor.
Much of the research that has been done concerning gender has focused on a critique
of male-biased research. Many archaeologists associate faunal remains in the
archaeological record with male activities. This has begun to change over the last
decade with many researchers offering evidence of women performing what had
previously been defined as "male" tasks (e.g., hunting, participating in warfare, etcetera).
The difficulty of gendered research lies with the methods of reconstruction. The
use of analogy is the typical method used to interpret archaeological remains. This

method, while controversial, has been employed by researchers such as Spector (1983),
Brumback and Jarvenpa (1997), Classen (1991), and Szuter (2000). The most serious
problem with using analogy to reconstruct the gendered behavior of the Anasazi is the
lack of accounts of women's activities in Southwestern ethnographies. Thus, it is
necessary to use cross-cultural analysis. This can be undertaken by choosing analogue
cultures with similar economic or social characteristics. All of the cultures used in this
study practice primarily household-level production, farm or garden, all hunt wild game
or fish (to varying degrees), as well as keeping domestic animals.
Groups that are considered for cross-cultural analysis in this dissertation include
peoples from around the world: some African groups (the Ovimbundu, the Kanuri of
Bornu, the Tiv, the Bemba, and rural communities in the Jebala Mountains of Morocco);
Asian peoples from India, China, Japan, Korea, Thailand, Borneo, Indonesia, and
Melanesia; "traditional" European societies including groups from rural Greece,
Yugoslavia, Serbia, Bosnian Muslims, Lapps, and even Scottish crofters; and New World
peoples including Mayan groups from the Yucatan and Quintana Roo, the Tarahumara
from Chihuahua and the Sierra Madres, Andean communities, Caribs, and Puebloan
groups from the Southwest.
Among all of these groups there is a basic pattern in the care of animals.
Animals kept within the household (either for meat or for their secondary products) are
the responsibility of the female head of household. Additionally, household domestic
animals are often fed surplus household stores (such as maize, rice, sweet potatoes, or
other food scraps). Care of herd animals is more variable. Males tend to care for herd
animals if they are kept far away from the home (especially if these areas are considered
dangerous). Women may care for herd animals if they are herded near to home or while
they are housed within the confines of the household for feeding or milking. Given this

pattern, therefore, it is likely that the domestic turkey kept by the Anasazi also came
under the care of women whether they were kept for ritual purposes or eaten as food.

CHAPTER 3
PHYSICAL AND CULTURAL SETTING
The Ancient Puebloan region has a long history of human occupation and its
cultural and environmental histories are well understood due to the large number of
archaeological sites, excellent preservation, and access to highly accurate dating
techniques. Shields Pueblo is located in the northern San Juan (or southern Colorado)
River Basin (Figures 1 and 2). This region is defined by a series of drainages, running
from the northeast to the southwest that feed the San Juan River. There are seven key
drainages within the region: Monument-McElmo, Dolores, Ute, Mesa VerdeIMancos, La
Plata, Animas, and the Upper San Juan-Piedra drainages. Shields Pueblo is located on
private land immediately north of Goodman Point Monument (Figure 3) in the
Monument-McElmo drainage, on the mesa top above the head of Goodman Canyon.
The elevation of the Monument-McElmo drainage ranges from approximately 2300
meters in the northeastern-most corner to 1525 meters in the southwestern corner along
Yellowjacket Creek, with Shields Pueblo sitting at just over 2000 meters above sea level
(USGS 2005).
The Monument-McElmo drainage system is composed of a series of small and
medium-sized canyons. Most of the streams running through the arroyos are
ephemeral. Of these canyons, only the Dolores River Canyon, along the northeastern
corner of the Monument-McElmo drainage, is a permanent water source. There are
some fresh water sources in the form of springs and seeps located at canyon heads
(Connolly 1992).

Figure 1. Map of the Southwestern Region of the United States

Figure 2. Map Showing the Location of Shields Pueblo (5MT 3807) (Courtesy of Crow Canyon Archaeological Center 2000).

Figure 3. McElmo Drainage Area Map (Courtesy of Crow Canyon Archaeological Center).

Flora and Fauna
There are seven separate biotic communities represented in the Northern
San Juan drainage system today (Table 3). The Pinyon -Juniper woodland and
the Sagebrush-Saltbrush biotic communities dominate the Monument-McElmo
drainage, but there are pockets of grassland along the western Utah border and
a very small pocket of Gambel oak scrubland in the foothills of the Durango area
which was likely more widely spread prehistorically. Approximately 81.7% of the
Monument-McElmo drainage land area is comprised of the pinyon-juniper
woodland (Adams and Petersen 1999:Table 2-1 :I 8). Shields Pueblo, because of
its mesa top location sits on the boundary between the pinyon-juniper woodland
and the sage-brush saltbrush biotic community, providing inhabitants of this site
with access to all of the economically important resources available in both
zones. The pinyon-juniper community is dominated by juniper (Juniperus spp.)
and pinyon pine (Pinus edulis), which seem to share dominance in this region.
While pinyon pine has been used as a food resource in other areas (in the Great
Basin), it does not seem to produce dependable quantities, as they can be
patchy in availability. Adams and Petersen (1999:16) argue that grasses and
shrubby plants found in the grasslands seem to be much more economically
important archaeologically for this region both as an actual food source and as
an attractive locale for large game species. This is a productive environment for
large artiodactyls (namely elk [Cervus elaphus] and mule deer [Odocoileus
hemionus]) especially during the winter months (Adams and Petersen 1999).
The sagebrush-saltbrush biotic community comprises approximately
15.1% of the total land area for the Monument Mc-Elmo region. Species diversity
is lower in this community with big Sagebrush (Artemisia tridentate) as the

Table 3. Northern San Juan Biotic Communities
Grass lands r- Scrubland
1 Pine-Douglas-Fir
Spruce-Fir-Forest
Alpine Tundra r (Adams and Petersen
Above 3500-3800 m.
rabbit brush, winterfat, prickly pear and cholla, ricegrass, grama, and dropseed Grama, ricegrass, Junegrass, dropseed, wild rose, sumac, Yucca (and all of the above shrubs and cacti) Juniper, pinyon ricegrass, western wheatgrass, dropseed, Junegrass sage brush, yuccas, serviceberry, mountain mahogany, and various cacti Gambel oak, and the coniferous trees, mountain mahogany, serviceberry, choke- cherry, hackberry, wild rose, currant, and sumac Pines, Douglas-fir, true firs, poiderosa pine, aspen, various grasses and fruit- bearing shrubs Willow, alder, maple, limber pine, bristlecone, Douglas-fir, ponderosa, dwarf iunipers, various herbaceous plants, surrant, and ~lderberrv LOW growing woody shrubs, herbaceous Aants, mosses, ichens
antelope, desert bighorn sheep
Small mammals and birds
Elk, mule deer, small game
Mule deer, wild turkey
Mule deer, elk, grey ~ o l f , small mammals, i r ds
Elk, mule deer
3ighorn sheep, med. nammals

dominant species. There are a few other species of shrubs (such as saltbrush
[Atriplex], rabbitbrush [Chyrsothamnus], and winterfat [Ceratoides]) Grasses
(e.g., ricegrass/needlegrass [Stipa], western wheatgrass [Agropyron smithii],
dropseed grass [Sporobulus], and Junegrass [Koeleria]) again, are the
economically important plant species, but prickly pear (Opuntia spp.) may have
also been used by native peoples. Small mammals, such as lagomorphs and
sciuridae are common in this environment along with coyote (Canis latrans).
Large artiodactyls are rare in this community, but Adams and Petersen argue
that antelope (Antilocapra) and desert bighorn (Ovis canadensis )would have
been available during prehistoric times (1999).
The grasslands are found in the southwestern portion of the Monument
Mc-Elmo drainage and comprise only about 1.6% of the land area. Typically,
grasslands would have been open plains of mixed grasses controlled by periodic
wild fires. Historic and modern farming and grazing practices have decreased
the numbers of wild fires, which has allowed bushes and shrubby species to
infiltrate the grasslands causing them to shrink in size (Brown 1982). The
grasses found in the grasslands include: grama (Bouteloua), rice grass (Stipa ),
Junegrass (Koeleria ), and dropseed grass (Sporobolus). Infiltrating shrubs
include: saltbrush (Atriplex), Sagebrush (Artemisia spp.), winterfat (Ceratoides
spp.), wild rose (Rosa spp.), yucca (Yucca spp.), and rabbitbrush
(Chyrsothamnus spp). Within the study area, juniper has begun to invade the
grasslands. A wide variety of mammals (such as antelope [Antilocapra] and
jackrabbit [Lepus] ) and various birds are available in this biotic community
(Adams and Petersen 1999).
The Gambel oak biotic community makes up a scant 1.6% of the total
Monument-McElmo drainage landscape. Oak scrub tends to be found on south

facing slopes. This community is dominated by Gambel oak (Quercus gambelio,
but there is no indication that acorns were harvested prehistorically (Adams and
Petersen 1999). Other scrubby trees such as: mountain mahogany
(Cercocarpus), serviceberry (Amelanchier), chokecherry (Prunus spp.), and
hackberry, elderberry, current, and sumac (Celtis, Sambucus, Ribes, and Rhus
respectively) are also common. This is a particularly important habitat for mule
deer (Odocoileus hemionus) and wild turkey (Meleagris gallopavo) and would
have been economically important for hunting (Adams and Petersen 1999).
Soils and Geology
Soils are typically rocky and may be poorly developed at lower
elevations, but tend to be thicker and better suited for agriculture at higher
elevations along the southern and southwestern slopes of the McElmo Creek
tributaries (Adams and Petersen 1999). The soils are comprised of weathered
sandstones and shales, along with mixed eolian materials from the San Juan
Basin. According to the Soil Conservation Service, the eolian upland soils and
alluvial soils of the lowland floodplains are particularly good for dry farming
(Adams and Petersen 1999).
Climate
The Northern San Juan Region is considered to be a cold middle
latitude, semiarid environment and follows the typical "Southwestern" pattern of
biseasonal rainfall. Precipitation typically falls as snow in the winter and as
heavy, unpredictable thunderstorms during the summer rainy season. According
to Dean (1996) this has been the prevalent weather pattern since A.D. 966,
interrupted during the great drought from A.D. 1250 to 1450. This pattern of

precipitation is necessary for planting crops (in the spring) and maturing crops (in
the summer months) (Adams and Petersen 1999). The Monument-McElmo
region receives between 25.4 to 50.8 cm of precipitation per year (Western
Regional Climate Center 2005). The average totals for rainfall at the Cortez
weather station (the closest station to Shields Pueblo) is 32.9 cm (for the years
1929 through 2004) (Western Regional Climate Center 2005). This area is quite
dry and only slightly above the minimum requirement of 30.48 cm of annual
rainfall required for growing maize (Hack 1942:23). Maize also requires a certain
number of mild days for proper growth and maturation. Siemer (1 977) argues
that maize requires a minimum of 2,500 growing degree-days in order to produce
a reasonable crop. Corn Growing Degree Days are calculated by adding one
unit for every degree above 60 degrees Fahrenheit on an average daily basis.
Cortez has a reported Corn Growing Degree Days measurement of 3160, placing
Shields Pueblo well within the required minimum (Adams and Petersen 1999:25).
Maize also requires a certain number of frost-free days. Varieties of maize
grown prehistorically in the Upper San Juan region generally require at least 120
frost-free days for proper development (Adams and Petersen 1999:26). The
Cortez weather station reports an average of 120 frost-free days, falling right at
the minimum (Adams and Petersen 1999:26). Shields Pueblo, therefore (at least
according to modern climatic records) falls nicely within all of the minimum
precipitation and temperature requirements needed for successfully growing
maize with dry farming techniques.
Paleoclimate
Paleoclimate has been an important topic of research in the Southwest
since the development of the tree-ring chronologies in 1929 (Douglas 1929,

1935). In addition to the tree-ring chronology, pollen, plant macrofossils, and
packrat middens have also been used in environmental reconstruction
(Betancourt 1984; Betancourt and Briggard 1985; Betancourt et al. 1990; Carrara
et al. 1991 ; Davis 1996; Hevly 1988; Jennings 1980; Maher 1961 ; Petersen
1985, 1988; Van Pelt 1978; Wright et al. 1973). While these methods cannot
provide actual amounts of precipitation, they do provide a proxy for how well
trees and plants did during the growing season.
Climate has been a key factor in explaining culture change in the
Southwest, and as such there is a plethora of research on the subject (Ahlstrom
et al. 1995; Dean 1996; Dean et al. 1994; Euler 1988; Hevly 1981 ; Hevly et al.
1979; Hughs and Diaz 1994; Petersen 1985,1988, 1994; Schlanger 1988;
Schoenwetter 1966), therefore this discussion will focus on the time periods that
most pertain to this dissertation (Pueblo Periods I through Ill). From A.D. 750 to
1300 there were two major climatic patterns, the Medieval Warm period (A.D.
800 - 1200) and the Little Ice Age (A.D. 1250-1850) (Petersen 1988, 1994).
These trends are illustrated in Figure 4. Within these major episodes, the
paleoclimate was characterized by a series of oscillations (Hevly 1981 ; Hevly et
al. 1979; Petersen 1985; Schoenwetter 1966).
While there were small oscillations, the overall pattern of the Medieval
warm period was that it was characterized by warmer, wetter weather (Adams
and Petersen 1999). Petersen (1985) and Hughes and Diaz (1994) argue that
the Medieval warm period coincides with the development and growth of the
Puebloan system. Longer growing periods and increased precipitation allowed
the Anasazi to move into areas that were previously unsuited for dry farming
(Schlanger 1988). This period of beneficial weather

Climatic Reconstruction: Southwest U.S. A.D. 600-1400
Years A.D.
Figure 4. Climatic Reconstruction: Southwest U.S. A.D. 600-1400 (after Davis 1996: Figure 12-5).
lasted for some 400 years, until approximately A.D. 1250 (when the weather
patter shifted dramatically).
The Little Ice Age was characterized by cooler summer temperatures
(decreasing the growing period of corn) and by less predictable and different
patterns of precipitation (Adams and Petersen 1999). Aside from the cooler
temperatures and dryer weather, the Little Ice Age witnessed a change in global
atmospheric patterns (Kreuts et a/. 1997). During this time period, summer rains
came later in the season, were concentrated further south, and did not last as
long as the present-day "monsoon" cycle (Adams and Petersen 1999). This
change in temperature and precipitation is argued to have led to the population

depopulation (i.e., emigrations) of the Northern San Juan region (Ahlstrom et al.
1995; Dean 1996; Dean et al. 1994; Euler 1988).
Archaeological Setting
The archaeological setting of the Anasazi of the San Juan Region is well
understood. Many archaeologists have published reviews and accounts of the
culture history of the Anasazi (Cordell 1984; Cordell and Gumerman 1989; Lipe
1999; Minnis and Redman 1990; Rohn 1989; Varien 1999; Wilshusen 1999). In
this section, I will not discuss the cultural setting in detail. Instead, I will provide a
brief summary of the archaeological history for the Northern San Juan River
Basin. The major archaeological trends for each time period are summarized in
Table 4.
Archaeological Background
There are six major cultural periods in the Northern San Juan:
Basketmaker II and Ill, Pueblo I, 11, Ill, and IV (or depopulation period). l will
outline general trends for Basketmaker II through Pueblo I and present the
evidence of social change for Pueblo II and Ill by discussing settlement pattern
and population, diet, and evidence of warfare in more detail.
Basketmaker 11 (1000 B.C. - A.D. 500) was the period when nomadic and
semi-sedentary hunting and gathering was replaced by sedentary maize
horticulture supplemented by large game hunting (Lipe 1999: 1 59-1 63).
Basketmaker II period sites are rare, with only about 150 reported for the upper
San Juan Basin (Lipe 1999: 152). Basketmaker II is defined by Kidder as an
"agricultural, atlatl-using, non-pottery-making" period (1927:490). Artifact types

I , b .G 3 o - .- 2 ' 0 a,.!&? a," a, 3 .;; a - 0 g 0.1 g m = 3 a, L u !?!.suu a u 0 b2.5 $ Cr = m ~ a , ~ m o

u a, 4- ([I -
.i! 2 c 0 ([I a 3 a, -Ju
). fn $ 5 2 0. .- '5 ([I - 0 5 € 2 6 5 s z s N LYS a, 0 7
C 0 . - C ([I - 3 a 0 a

called "horseshoe" shaped) metates; single and two-handed manos; expanding-stem
drills; spindle drills; stone pipes; and woven and coiled basketry and sandals (Lipe
1999).
There are several types of Basketmaker II sites. Residents typically constructed
one or more circular or oval shallow pithouses with stone slab entries. House floors are
generally characterized by a central hearth, a series of postholes, sub floor storage
cysts, and one or more stone metates (Eddy 1972; Hammack and Walkenhorst 1991 ;
Morris and Burgh 1954). Communities settled in areas suitable for both agriculture and
hunting and gathering wild resources. Early Basketmaker II sites seem to be more
aggregated with "villages" of up to 11 houses (Eddy1972:lg). Late Basketmaker II sites
became more dispersed. At sites, such as Cedar Mesa, pit structures are dispersed
over a 1.5 km area, with only one or two houses clustered together (Dohm 1994:271-
272). Also common during this period are rock shelters yielding beautifully preserved
sandals and basketry. Limited use sites include camps and lithic procurement areas.
These sites may have been used for a single purpose, but were likely used repeatedly
(Matson 1991 : 82-89).
During Basketmaker II, the Anasazi people began to incorporate domesticated
maize, but were still somewhat reliant upon wild resources. Chisholm and Matson
(1994: 248) have shown that Basketmaker II protein intake consisted of 60 to 84 percent
maize. In addition to maize, cucurbits were grown and several wild plant species were
gathered (grass seeds, pinyon pine nuts, and various weedy plant species) (Lipe 1999).
Faunal remains show a dependence upon deer species, lagomorphs, bighorn sheep,
and wild Merriam's turkey (Lipe 1999).
Basketmaker Ill (AD. 500 -750) is not simply a continuation of the previous
period. The Anasazi began to manufacture ceramics; more often than not, represented
by undecorated brown, grey, and white wares. Dart points were replaced by arrow

points, and the horseshoe shaped metate and cobble mano combination was replaced
by the trough form metate and rectangular mano (Plog 1979). Some of the most
beautiful early woven and coiled sandals and basketry were produced during this time
period.
Architecture during Basketmaker Ill became very distinctive, with styles changing
nearly every generation (Hewitt 1983). Basically, early Basketmaker Ill houses retained
many of the characteristics of the previous period (i.e., circular shallow depressions); but
by the early A.D. 700s, habitation structures were excavated 20 to 30 cm deeper (with
circular, subterranean antechambers disappearing at the end of the period) and showed
evidence of changes to roof construction and the general layout of house floor
(Wilshusen 1999a). In addition to the floor features of Basketmaker II houses,
Basketmaker Ill houses were constructed with large central hearths with deflectors,
storage bins lined in stone slabs, and wing walls (Wilshusen 1999a: 174). Population in
the Northern San Juan region remained low until late Basketmaker Ill when it effectively
doubled in the Monument-McElmo drainage (Wilshusen 1999a: 188-1 91). This is
evidenced by an overall increase in Basketmaker Ill sites and by a change in community
organization. Communities were typically made up of 5-9 households grouped in a
larger multihousehold site such as a neighborhood (Wilshusen 1999a). Several
Basketmaker Ill sites were surrounded by stockades (the Knobby Knee Stockade,
Gilliland, Palote Azul, Cloud Blower, Crooked Man, and Rabbit sites). The stockades
were made up of a series of 2 to 3 meter tall posts set into the ground every 10- 30 cm
with woven twigs and branches filling the spaces between (Wilshusen 1999a: 180)
These are all large hamlets located in the Monument-McElmo drainage. It is possible
that these stockades are evidence of an increased need for families to protect
themselves from outsiders, but more research needs to be done in this area.

Basketmaker Ill is characterized by an increased dependence upon cultivated
foods (as evidenced by increasingly larger and more formalized storage structures).
The increasing importance of agriculture is also supported by the appearance of field
houses. These were located near agricultural fields away from the main habitation sites
ostensibly providing shelter for workers while guarding their fields (Wilshusen 1999a).
During Pueblo I (A.D. 750 - 900) the three major regional cultures of the greater
Southwest (Anasazi, Hohokam, and Mogollon) developed. Pueblo I peoples of the
Northern San Juan began to increase their ceramic production. During this period,
ceramics are primarily represented by plain grey ware, but some were decorated with
banding around the necks of jars. Piedra Black-on-white shows up in late period sites
(Wilshusen and Blinman 1992; Wilson and Blinman 1993). The bow and arrow was
introduced just prior to Pueblo I. Pueblo I projectile points are characterized by arrow
points with large tangs and contracted stems (Bradley 1988).
Surface architecture consisting of masonry unit pueblos of habitation and storage
rooms with an associated kiva and midden also appeared in the Northern San Juan at
this time (Gumerman and Dean 1989). Communities built seven distinctive site types
during the Pueblo I period. Villages consist of a grouping of 15 to 20 houses (with at
least 50 rooms); large hamlets of 3 to 7 houses; small hamlets with 1 or 2 houses; large,
public architecture (including great kivas); field houses; and non residential artifact
scatters (Wilshusen 1999b).
Populations began to increase dramatically in the western drainages during the
late A.D. 700s, leading to the first of a series of population movements out of the
northern San Juan into northwestern New Mexico (Wilshusen and Ortman 1999). As
populations began to grow and to shift into aggregated villages, the Anasazi responded
by intensifying maize production (as evidenced by an increase in the number of grain
storage structures) and by cementing exchange relationships (as evidenced by ceramic

exchange) (Kohler and Van West 1996). Animal resources would probably have
become more scarce at this time due to increasing human populations. While turkeys
were not present in large numbers, Munro (1994: 149) argues that this was when they
became domesticated. Domestication is demonstrated through the presence of non-
osteological remains (such as large quantities of eggshell, gizzard stones,
accumulations of turkey droppings, and pens or enclosures) and osteological remains
such as the presence of all age categories of birds (Munro 1994: 94). At this juncture in
time turkey seems to been used primarily for ritual purposes (evidenced through turkey
burials in kivas or in human burials) and for utilitarian purposes (collecting feathers for
blanket manufacture, cordage, and fletching for arrows) ( Hill 2000; Mc Kusik 1980).
Pueblo II (A.D. 900 to about 1150) is characterized by two major trends. First,
we see the development of highly localized stylistic traditions, a so-called parochial
movement. In the Northern San Juan region there are specific artifact styles associated
with each major drainage area with neck-banded, black-on-white, black-on-red, and
corrugated vessels common (Cordell and Gumerman, 1989). Projectile points are
typically small, narrow-stemmed, corner-notched arrow points (Bradley 1988).
Architectural styles are more varied. Some communities in the Mesa Verde region
began to build masonry architecture, while other communities to the northwest
constructed surface structures of jacal (Morris 1991: 659-661).
Secondly, there is yet another surge in the development of large aggregated
habitation sites and complex social organization. Prior to A.D 1050, Pueblo II sites
commonly consisted of unit pueblos with a kiva, one or two surface rooms and an
underground milling room (Lipe and Varien 1999a). Community structure began to
change after A.D. 1050 with the development of the Chaco Canyon complex, which
dominated the Anasazi cultural area. The Chacoan system consisted of the main cultural
center (Chaco Canyon) and dozens of Chacoan great houses ranging from 25 to 55

rooms surrounded by smaller satellite communities, many linked by road systems.
Populations were extremely concentrated within an eightykilometer region around
Chaco Canyon. The Northern San Juan really became an extension of this system
during the latter half of Pueblo II. Some have asked how Chaco was able to influence
such a large number of communities. Sebastian (1992) suggests that residents of
Chaco were able to produce large surpluses of food and used thesle surpluses to create
relationships of indebtedness with others within their community and in surrounding
communities during difficult years. This system escalated as successful farmers began
to build large public architecture (Kantner 1996). In the A.D. 1000s Chaco grew
substantially, with residents showing evidence of high sociopolitical rank (Sebastian
l992:l2O-l26).
The first evidence of warfare and possible cannibalism can be dated to the
Basketmaker II period. While there is earlier evidence of violence, it does not seem to
escalate until Pueblo II. Turner and Turner (1999) argue that Chacoan elites used public
violence as a means of social control. There is documented evidence at thirteen
different sites throughout the Anasazi area for violence and cannibalism (Billman et a/.
2000).
During Pueblo II faunal food sources were similar to that of the preceding time
periods. Artiodactyla (especially deer) are common in faunal assemblages as are other
communally hunted animals. At about A.D. 900, turkey shifted in importance from a
primarily ritual use to that of a food resource, and thus increases in frequency in
archaeological assemblages (Munro 1994).
Pueblo Ill (A.D. 1150 to about 1300) is believed to be the period of the most
intense habitation at Shields Pueblo. This period was a time of extreme instability in the
Southwest. There was a new wave of depopulation (e.g., relocation to other areas) and
the establishment of new regional centers. In early Pueblo Ill there was a trend toward

increased residential aggregation. A number of large sites were established in the Mesa
Verde and Monument-McElmo drainages (Lipe and Varien 1999b). There was an
overall population shift into the western drainages of the Northern San Juan, and from
A.D. 1180-1 190, a surge in population. Habitation sites tended to be loosely clustered
on mesa tops associated with productive agricultural land. Community centers were
made up of great kivas or remodeled Chacoan great houses, and semi-aggregated
clusters of habitation rooms.
Late Pueblo Ill saw some drastic changes to settlement patterns. Both location
and human population levels changed notably. During the later portion of the Pueblo Ill
period many residential sites moved from mesa tops to canyon rims or the talus slopes
below (e.g., Sand Canyon Pueblo, Seven Towers site, Yellow Jacket Pueblo)
(Kuckelman 1997, Lipe and Varien 1999). Villages and subsidiary settlements also
became more tightly aggregated on the landscape with the majority of the population
living in larger villages located along the western drainages.
Additionally, there were major changes in subsistence activities both in terms of
intensification and in dominant species utilized. Agricultural production was intensified,
evidenced by increased manipulation and control of water resources, with reservoirs,
check dams, and other water capture devises (Lipe and Varien 1999; Rohn 1963, 1977;
Winter 1978). Driver and Muir note an interesting change in faunal remains during
Pueblo Ill times in the Northern San Juan area (Driver 1997, 2002; Muir and Driver
2001). There was a shift in the location of large mammal remains (particularly
Artiodactyla) at the site level. Comparisons of site faunal assemblages show higher
frequencies of large game animals in large aggregated communities, while small site
assemblages (from seemingly "richer" resource areas) have far fewer artiodactyl
remains, but higher frequencies of rabbits and turkey (Driver 1997, 2002; Muir and
Driver 2001). Different explanations for this trend have been suggested and are

discussed in Chapter 1 as potential causal factors, including climatic change, population
growth and pressure, and changes to social and site hierarchy.
In the later part of Pueblo Ill evidence of social unrest appeared. There was an
increase in defensible site locations, defensive architecture, and a general increase in
evidence of interpersonal violence (Lightfoot and Kuckelman 2000). LeBlanc argues
that a cycle of warfare began in the northern Southwest around A.D. 1200. This
violence was widespread and there are numerous examples of mass graves, destruction
and depopulation of villages where the dead were simply left exposed and not formally
buried, and even evidence of cannibalism (Ahlstrom et a/. 1995; Haas and Creamer
1996; LeBlanc 1999; Turner and Turner 1999; White 1992). The last half of Pueblo Ill
was likely a very difficult period for the Ancient Puebloans. Even if violence was not
constant (but sporadic) the fear of attack and the necessity for constant vigilance would
have been a difficult burden.
The final period, Pueb1o.W (A.D. 1300 to 1540) has also been referred to as the
period of depopulation. What is defined as Pueblo IV culture began to appear in some
communities as early A.D. 1250 (Adams and Duff 2004:3). Basically, Pueblo IV culture
is identified by the presence of particular pottery types (White Mountain Red Ware, Zuni
Glaze Ware, Salado Polychrome, and others) (Carlson 1970) and by the appearance of
the Katsina Cult iconography present on ceramics and rock art (Adams 1991). The
Northern San Juan region was depopulated by Anasazi groups and there was a general
reorganization of settlements along the Little Colorado River, Rio Grande, and the
historic pueblos (Hopi, Zuni, Acoma) (Wilshusen and Towner 1999). Non-Anasazi
groups began to move into the Northern San Juan. The Ute began to move into
northwestern Colorado by A.D. 1300-1400 and were in the Northern San Juan region by
I626 (Janetski 1994; Schroeder 1965). Athabascan groups (the Navajo and Apache)

moved into the Four Corners region of the Southwest later during the historic period, by
about A.D. 1750 (Wilshusen and Towner 1999).
Excavation and Collection of Data
The Shields Pueblo research project is part of Crow Canyon Archaeological
Center's "Communities Through Time: Migration, Cooperation, and Conflict" regional
project. This research project was established to look at the cycle of development and
depopulation of pueblo communities in the Mesa Verde Region of southwestern
Colorado (Duff and Ryan 2000). Crow Canyon Archaeolological Center has focused on
the Pueblo II and Ill periods (A.D. 900 - 1300) because of dynamic changes in site size
and population. Shields Pueblo has occupational deposits ranging from as early as A.D.
700 to 900 all the way through to the end of Pueblo Ill when the site was depopulated.
This is important to this research as the deposits are sequential at the same site and
provide an opportunity to understand change over time.
The earliest known excavations at Shields Pueblo took place in the mid 1900s
(Hayes and Chappell 1962). Artifacts collected during these excavations are housed as
part of the Chappell collection at the Anasazi Heritage Center. Colorado Mountain
College held a field school at Shields Pueblo during the summers of 1975, 1976, and
1977 (Bagwell 1975, 1976, 1977). Five of Shields' kivas were excavated along with one
roomblock, and various cultural deposits. Crow Canyon conducted a survey of the
Goodman Point and Sand Canyon pueblos in the 1980s, recording segments of Shields
Pueblo at that time (Adler 1990).
Preliminary archaeological survey of Shields Pueblo was conducted in 1996 in
preparation for excavation the following summer. The faunal assemblage used for this
project was collected by Crow Canyon Archaeological Center over a four-year period
starting in April of 1997 and ending in November 2000 (Duff and Ryan 2000). The

purpose of the excavation was to collect general information on large-scale changes
over time from Pueblo II to Ill, and to gather information for intersite comparisons. For
this reason, Crow Canyon did not focus on uncovering complete floor contexts. They
randomly sampled the major artifact concentrations and ran trenches through the center
of any structures that were discovered (Duff and Ryan 2000:4).
During survey, eighteen high-density concentrations of cultural remains were
identified and mapped (Duff and Ryan 2000). The locations of these concentrations
were then used to plan a research strategy. Each high-density area was systematically
surface collected and was excavated using randomly selected 1 x 1 m. units. If
structures were discovered, they were excavated using judgmental sampling (Duff and
Ryan 2000).
The first field season was devoted to sampling the southern half of the site and to
a remote sensing survey to help identify possible subsurface structures. Subterranean
structures were tested and excavated in the 1998, 1999, and 2000 seasons (Duff and
Ryan 2000). Crow Canyon tested some 35 structures by the end of the 1999 season
(Duff and Ryan 2000: 4). The final season was devoted to surface collection and
excavation of previously uninvestigated areas and continuation of judgmental testing of
subsurface anomalies discovered during subsurface survey. Trenches were dug across
structures to ascertain their boundaries and to establish their depositional history. A
number of 2 x 2 m. units were excavated to the southeast of known structures in order to
locate midden deposits (Duff and Ryan 2000). Shields Pueblo is located on property
that was historically farmed. As such, a plow zone of historically disturbed sediments
blankets the site. This plow zone varies in depth from 20 to 40 cm (Duff 2005 personal
communication). While much of the surface architecture was destroyed, the
subterranean deposits were fairly undisturbed and ranged in depth from 1 to 2.5 m. deep
(Duff and Ryan 2000.). The overall shape of Shields Pueblo is northerly oriented

horseshoe (or u-shape). It is unclear if Shields was a continuous series of room blocks
(along the lines of Sand Canyon Pueblo) or a series of separate roomblocks with
associated kivas and pit structures, as the site has been mostly destroyed by historic
farming. However, the dating of the structures in the excavation blocks seems to
indicate the latter. Excavation blocks are described in Table 5 (where T= Trashlmidden
fill; B= burned) and then each block is summarized below. Figure 5 shows all of the
excavation locales at Shields Pueblo and a basic site layout.
Table 5. Excavation Block Summary: Structure Descriptions and Date Ranges
104l~asonrv surface room - CMC I ILPII I I 1 10I~arthen pit structure I ~EPI 1725-800 1
121 1Masonry surface room 1 ~LPII 122 123 124 136l~arthen-edged pit structure NIE
Masonry Kiva Masonry Kiva built within STR 122 Earthen mealing room
I ~LPII
149 150 151
200
X
Subterranean room NIE NIE
221 222 223 224
X X
LPll
205
208 213
Masonry Kiva Masonry Kiva Masonry Kiva Masonry Kiva
LPll EPIII-LPIII LPll
1125
X
Masonry pit room
Masonry Kiva Post abandonment structure built in STR 208 fill
X
X X
Plll
LPlll LPlll
1225-1 260
LPlll EPlll LPlll LPlll
1248
1256 1255

I I 2251~asonry Kiva with post abandonment wall I IX ILPIII I I
I 2341~iva with earthen-edges and stone pilasters 1x1 ILPII 1 229 Postabandonment stone circle in fill of
STR225
237 241
LPlll
400
Earthen pit structure Masonry Kiva - post-abandonment walls in fill
405 406
243 401
407 408
409
41 0
X
Masonry Kiva Masonrv Kiva
1 100
Kiva- CMC Post-abandonment circle over STR 405
Post-abandonment circle in fill of STR 405 Masonry Kiva - CMC
Post-abandonment circle in fill of STR 408 - CMC
Kiva - CMC
1200
1 13001 13071Earthen pit structure built within STR 1310 I IMPII
X
X X
1 102 1 106 1 108
12091~arthen it structure
LPl l LPlll
LPlll
LPlll
EPlll
1 1 13 1 1 14 1205 1206
I ~EPIII I
LPll LPlll
1131
LPlll LPlll
1212
Slab-lined pit Masonry Kiva Pit structure
1308 131 5
1250
1250 1229
Masonry Kiva Masonry Kiva Masonry Kiva Partial masonrv Kiva
131 61~asonry Kiva
X X
Earthen-edged and bench pit structure Masonrv Kiva
1400
X X
1 IEPIII
141 1 141 2
EPlll EPlll EPlll
X
1212 131 8 1402 1408
1413
1414
T= trashlmidden fill; B= burned (*extensively burned); CMC= excavated by Colorado Mountain College; NIE= not excavated (Shields Pueblo Provenience Database; Duff and Ryan 1999,2000)
1199 1224
EPlll EPlll EPlll EPlll
Post-abandonment structure over STR 408 Post-abandonment structure over STR 1402
1500
1205 1203
X
Earthen pit structure Masonry Kiva Masonry Kiva
Masonry pit room connected to STR 1408 by tunnel Earthen it structure
LPl l l LPlll
141 6
1504 1505
MPll LPlll
X X
-
X
1251
LPlll
EPlll Earthen pit structure
Masonry Kiva Masonrv Kiva
EPI LPlll LPlll
1258 1255
X
X
EPlll
EPlll EPlll 1155


Block 100 consists of a series of structures, nonstructural features, and backhoe
trenches. There are 19 structures in the 100 Block including: four masonry surface
rooms and one wattle and daub surface room (dating to Late Pueblo II), six earthen pit
structures, three masonry kivas, and three structures that were not excavated (from
CCAC Shields Pueblo Provenience Database). Other prehistoric features include two
undated burials, and 22 samples from various contexts (CCAC Shields Pueblo
Provenience Database). Block 100 seems to have two distinctive temporal occupations,
the first during early Pueblo I (A.D. 770s), and the second during late Pueblo II (early to
mid A.D. 1100s) (Duff and Ryan 1999). Block 100 is the highest part of Shields Pueblo.
The masonry roomblock is associated with a prehistoric road running to Casa Negra,
and has certain architectural similarities to Chacoan great houses (e.g., thin, tabular
walls, and large room sizes) thus suggesting an association with the Chacoan system
(Duff and Ryan 1999). However, there does not appear to be a great kiva associated
with this roomblock (Duff and Ryan 1999). Further investigations during the 2000 field
season did not uncover any structures befitting a great kiva, leaving the question of
whether Shields Pueblo had a Chacoan great house unresolved (Duff and Ryan 2000).
Block 200 contains 13 structures (8 full masonry kivas, 2 partial masonry pit
structures, one earthen pit structure, two post-depopulation structures built within the fill
of earlier structures (Shields Pueblo Provenience Database). Block 200 has tree-ring
and ceramics dates indicating an occupation from the late A.D. 1000s to depopulation in
the late 1200s (Duff and Ryan 1999).
Block 300 does not contain any archaeological features or structures, but does
contain deposits of pottery (dating between A.D. 1000 - 1 180).
Block 400 was excavated by Colorado Mountain College in 1976 and 1977 (Duff
and Ryan 2000). Crow Canyon did not excavate any of the structures identified by CMC

in this block, but did open four 1 x 1 m. units and a backhoe trench (ibid). The trench
exposed a unit with three separate structural occupations. Structure 405, a burned
masonry kiva with a sipapu, is the oldest portion of this architectural structure dating to
the Late Pueblo Ill subperiod. On top of this lies structure 401, a circular masonry post-
depopulation structure and 407, a circular masonry structure built on top of post
depopulation fill (Duff and Ryan 2000). Duff and Ryan (1999) suggest a period of use
for this block between A.D. 1000 and 1180, with post-depopulation structures dating into
the A.D. 1200s.
One arbitrary 1 x 1 m unit was excavated by Crow Canyon in block 500,
uncovering a single pit feature (Duff and Ryan 1999). Previous excavations (prior to
1975) uncovered a pit structure, however no records are available for this feature (Duff
and Ryan 2000). Duff and Ryan (1999) indicate that block 500 was used between late
A.D. 1000 into the mid 1 100s.
No features have been uncovered in block 600 (Duff and Ryan 1999). Two
possible masonry-lined pit structures were excavated before 1975, again with no
available excavation records (Adler 1990, Duff and Ryan 1999). This block has been
dated through its ceramic assemblage from A.D. 1 100 - 1300 (Duff and Ryan 1999).
Block 700 contains the remnants of a historic homestead (Ward 1997). No
prehistoric archaeological components were found (Duff and Ryan 1999).
Portions of a structure, a full masonry kiva, were excavated in block 800 (Duff
and Ryan 1999). After depopulation, a wall was constructed across the kiva to close off
the southern recess (ibid.). The ceramic assemblage indicates use during early Pueblo
Ill (Shields Pueblo Provenience Database).
Blocks 900 and 1000 have no prehistoric deposits. No archaeological features
were uncovered in block 900, while block 1000 contains only a single pit structure. This
pit structure was originally thought to be a candidate for a great kiva, but had been

excavated in the historic period and no prehistoric deposits were uncovered (Duff and
Ryan 1999).
Blocks 1100 and 1200 have a small number of prehistoric components. Four
structures were excavated in block 1100 (three full masonry kivas and one-slab-lined pit)
all dating from early Pueblo Ill (Shields Pueblo Provenience Database). Only three
structures were uncovered in block 1200 (two masonry kivas and one unusually shaped
earthen pit structure, 1209) (Shields Pueblo Provenience Database). Duff and Ryan
(2000) describe this structure as bell-shaped. While it has both a prepared roof and
floor, its function is not known (Duff and Ryan 2000).
Block 1300 contains five structures (three earthen pit structures and two masonry
kivas) (Duff and Ryan 1999). Three midden deposits were sampled in addition to the
structures (Shields Pueblo Provenience Database). Block 1300 was used primarily in
mid pueblo II (when Shields Pueblo was re-inhabited), however, one pit structure (1 31 8)
dates to early Pueblo I and structure 1315 to late Pueblo Ill (ibid.).
A total of seven prehistoric structures were excavated in block 1400 (two
masonry-lined kivas, two post depopulation circular masonry structures built within the
depressions of the previous kivas, a masonry-lined subterranean room, one masonry
and earthen pit structure, and one completely earthen pit structure) (Duff and Ryan
1999). The masonry kivas (both burned) yielded tree-ring dates of A.D. 1250 (Shields
Pueblo Provenience Database). Both the earthen pit structure and semi-masonry pit
structure were used earlier in the late A.D. 1000s through the late A.D. 1100s (Duff and
Ryan 1999).
Two masonry kivas were uncovered in block1 500, one intensively burned, the
other unburned (Shields Pueblo Provenience Database). Tree-ring dating places the
burned kiva at A.D. 1155.

Blocks 1600, 1700, 1800, and 1900 contain no structures or archaeological
features and were found to be sterile under the plow zone (Duff and Ryan 2000).
Dating
Initial dating was based upon tree ring dates from structures at Shields Pueblo.
The earliest dates start in the A.D. 770s during early Pueblo I period (Duff and Ryan
2001:2). Block 100 was the primary area of habitation during this time (with a single
possible early Pueblo I structure in block 1300). A number of faunal remains (some 429
elements) were recovered from Shields Pueblo dating from between A.D. 725 and 800.
Based upon the dates assigned to study units, there is an extensive period of
"depopulation" at this site. There are no faunal remains dating from A.D. 800 to 1020.
During the middle of Pueblo II (A.D. 1020-1060), habitation seems to resume, with
approximately 620 faunal elements dating to this period. Mid-Pueblo II occupation was
limited to the 1300 block. The heaviest occupation of the site seems to be between A.D.
1060 and 1225 (starting in late Pueblo II times through late Pueblo Ill). Structures and
features dating to this period can be found throughout the site, although late Pueblo II
structures dominate the 100 block and early Pueblo Ill dominate in block 1100. There
are no late Pueblo Ill structures in the 100 block, however, most blocks have some late
components. Some 5,218 faunal elements have been recovered from this time period.
After A.D. 1225, population drops again (dated faunal elements falling to about 4,627).
Adler and Varien (1994) argue that A.D. 1225 is when the community center shifts from
Shields Pueblo, moving to Goodman Point Pueblo. While not completely depopulated
(with some 20 habitation structures still occupied in the mid A.D. 1200s) it is unclear
whether this represented a continuation of habitation of Shields Pueblo or a short-term
resettlement (Duff and Ryan 2000).

Chapter Summary
Shields Pueblo is located on private land just north of Goodman Point Monument
in the McElmo drainage (McElmo Dome) of the Northern San Juan River Basin. Shields
Pueblo sits at an elevation just above 2000 m at the head of the Goodman Creek
Canyon.
There are seven biotic communities in the Northern San Juan Drainage: the
sagebrush-saltbrush, grasslands, pinyon-juniper woodland, gambrel oak scrubland,
pine-Douglas-fir forest, spruce-fir-forest, and the alpine tundra. Shields Pueblo lies on
the boundary between the sagebrush-saltbrush community and the pinyon-juniper
woodland, which covers approximately 80% of the McElmo Dome region. This biotic
community provides a wide array of floral and faunal resources (including elk, mule deer,
and small game). Soils in the area are typically rocky and thin. However, higher
elevation along the southern and southwestern slopes of the McElmo Creek tributaries
are better developed and good for dry farming.
Today, the climate of the Northern San Juan region is "cold" and semiarid.
Precipitation generally falls as snow during the winter months and as heavy
unpredictable thunderstorms during the summer rainy season. This has been the
prevalent weather pattern since A.D. 966. This bi-seasonal precipitation pattern is
particularly beneficial for maize horticulture. Various paleoclimatic data illustrate major
changes in temperature and rainfall over time, which would have influenced the
availability of resources and the extent of maize agriculture, thus influencing population
size and location on the landscape.
Archaeologically, the Anasazi are well understood. There are six major cultural
periods covered in this chapter: Basketmaker 11 (1000 B.C.-A.D. 500), Basketmaker I l l
(A.D. 500-750), Pueblo I (A.D. 750-900), Pueblo II (A.D. 900-1 150), and Pueblo Ill (A.D.
11 50-1300). Each of these periods is characterized by distinctive cultural traits. Shields

Pueblo contains deposits from Pueblo I through to the depopulation of the site during
late Pueblo Ill. The site consists of 18 high-density concentrations of cultural remains
(used to establish excavation blocks 100 through 1800) organized into a horseshoe.
Structures in each block are described in Table 5. There were two periods of occupation
separated by a period of "depopulation ." The earliest dates indicate initial habitation
beginning around A.D. 725. There was a hiatus in occupation from A.D. 800-1020.
Shields Pueblo was re-occupied around A.D. 1020 and was thereafter, continuously
occupied until the site was depopulated around A.D. 1280.

CHAPTER 4
SHIELDS PUEBLO FAUNA
This chapter presents Shields Pueblo's faunal assemblage. This section will
begin by discussing the methods used to record and analyze the faunal data. The
faunal data will then be described in terms of relative abundance and an overview of the
natural and cultural taphonomic processes that have affected the assemblage will be
given. The temporal change in the general frequencies of common taxa will be
discussed in reference to specific sub assemblages. This chapter concludes with a
discussion of spatial and contextual data based upon the percent composition of
assemblages.
The faunal assemblage presented in this chapter includes the remains collected
during the field seasons from 1997 to 2000. All remains were analyzed, no sampling of
the collection was done.
Methodology: ldentification and Recording
This research is predominantly observational, measuring the relative frequencies
of various taxa among the collected faunal assemblage. ldentification was primarily
done using the comparative faunal collection at Simon Fraser University. Small rodents,
birds, and some carnivores were identified at the Burke Museum. A standardized
method of data recording using codes and procedures established by Driver (1999) was
used (see Appendix A). Each fragment was coded for twelve different items of
information (taxon, element, part, side, proximal or anterior fusion, distal or posterior
fusion, proximal or anterior break, distal or posterior break, modification, length, cortical
thickness, and number of fragments for each element). These categories were
designed to provide information on species, age categories, preservation and other

taphonomic processes affecting the bone, and any evidence of human modification. All
fragments were recorded by hand on data sheets and were entered into Excel for ease
of quantification and analysis of data. The data will be available online through Crow
Canyon Archaeological Center in the Shields Pueblo Site Report. All faunal remains as
well as all categories of artifacts are curated at the Anasazi Heritage Center in Dolores,
Colorado and are available for further study.
Quantification
The faunal assemblage was quantified by measuring the relative frequencies of
each taxon to establish the faunal patterning at Shields Pueblo. This was done by
establishing the number of identified specimen (NISP), which counts the total number of
specimens assigned to a particular taxon (Grayson 1984). From this we can derive the
relative abundance of taxa. While NISP has various criticisms (discussed below), it is
the most commonly used method of quantification in archaeology. This method does
not measure the actual number of individual animals present in the sample, but it is the
base line for most quantitative methods, analyses, and indices. From NISP other
measures of abundance can be derived, such as the minimum number of individuals in a
taxon, the size of the death population (i.e., the number of animals killed) (Fieller and
Turner 1982), and bone weight for each taxon, which can then be used to estimate meat
weight (White 1953). This method can further be used to calculate basic species
indices (e.g., artiodactyl index, lagomorph index, and turkey index), thus describing how
different species relate to one another within an assemblage.
That being said, it is important to note that there are several problems with using
NISP. The most serious problem with this method is that it treats each fragment as a
separate specimen, not as a part of a larger whole, thus resulting in the over-
representation of taxa with large numbers of identifiable skeletal elements (Gilbert et a/.

1982; Grayson 1979). In addition, it does not take into account fragmentation of the
elements (Chase and Hagman l987), differential preservation (Brain 1969; Koch 1990;
Lyman l982), differential representation of elements associated with butchering patterns
or the schlep effect (Binford 1978, 1981). All these short-comings, however can be dealt
with by applying skeletal part analyses (discussed later in this chapter).
Another method used to quantify faunal remains is MNI or minimum number of
individuals (White 1953). To use this method in its simplest form, the most abundant
skeletal element for each separate taxon is sorted into left and right sides. The highest
count (of either left or right) indicates the minimum number of individuals represented in
the sample. Unpaired elements (such as the first cervical vertebra) may also be used to
calculate MNI. The danger of this method is that it assumes that the left and right
elements are pairs (which may or may not be the case). MNI is also problematic
because there are multiple methods for calculating the number of individuals within a
sample. One method used by zooarchaeologists is to simply count the number of
specimens and divide by the number of occurrences in the body (Klein and Cruz-Uribe
1984). Another method uses the procedure of matching. Archaeologists have devised
methods of estimating MNI by separating elements by age (based on teeth or fusion of
epiphyses), the length of an element, or by the sex of an animal (Bokonyi 1970; Chaplin
1971). Left and right sides are matched together based upon these criteria. Not only are
paired elements matched, separate elements (such as a radius and a femur) may be
shown to come from the same individual (Klein and Cruz-Uribe 1984). Sorting elements
into lefts and rights and matching elements can yield very different MNls. If, for
example, a sample contains 12 proximal femora, 8 left and 4 right, an MNI based upon
sorting by side would yield an estimate of 8 individuals. If the same sample is matched
by age category and three of the rights are not in the same age category as the lefts,
then the MNI is increased to 11 individuals. Matching based upon bone size is also

problematic, mainly because most adults in the same species will not vary significantly in
size from one another (O'Connor 2000). Therefore, it is best to match based upon some
discrete trait (such as the fusion of epiphyses). Paired bones fuse at the same time
(unless there is some underlying trauma to the bone), so that a left unfused distal radius
and a fused right distal radius are unlikely to come from the same individual.
Minimum number of individuals, such as NISP, has certain inherent problems.
The most troublesome is related to the problem of aggregation, in other words, how the
sample is separated or grouped for analysis (Grayson 1984). The way a sample is
clustered (by stratigraphic unit, arbitrary levels, etc.) controls the outcome of the MNI
calculation. Thus, the same faunal collection can produce very different MNI values
(Grayson 1979; Ringrose l993:126-128). For example, if two turkeys had been eaten
and disposed of in three separate locations, MNI might be estimated for each context,
thus inflating the total MNI for that taxon. In addition, problems with aggregation may
also inflate the importance of rare species (Klein and Cruz-Uribe 1984). A rare species,
such as Haliaetus leucocephalus (the bald eagle) may be used and disposed of in
different contexts for religious purposes (e.g., the wings may be present in one location,
while the hindlimbs and talons are deposited elsewhere) thereby increasing or doubling
the MNI if calculated separately for each context. Deciding how to aggregate the sample
is also problematic, as there is no standardized method (Grayson 1984). Once the MNI
of a sample is calculated this number cannot be used for further analyses; MNI is not an
absolute number, but an estimate (Plug and Plug 1990). If you have an MNI of 32, this
translates to "at least 32" not "definitely 32 ." 0 ' Conner (2000:60-61) describes this
problem as follows:
As 'more than twenty' cannot be added to 'more than ten' in any sensible way, it follows that MNI values derived from any one assemblage cannot be combined with those from any other. The problem of aggregation is thus simply resolved: the data cannot be arithmetically aggregated, so any aggregation of sedimentary units requires physical

aggregation of the bone samples and complete re-analysis. Furthermore, conversion of MNI estimates within a sample to percentage data, to facilitate comparison with another sample, is mathematical nonsense as it involves the summation of a series of minima.
Yet another problem with MNI lies with identification of taxa. In many cases
elements are not identifiable to species level. In these instances, elements may only be
identified in basic taxonomic categories. For example, some elements may be identified
as Canidae, while others may be specifically identified as Canis latrans or Canis
familiaris. Calculating MNI does not allow for non-specific identifications, while NlSP
allows the researcher to combine categories. Thus, nonspecific categories are not
included in the quantification. Similarly, MNI does not allow the researcher to add
different species together to calculate the total within that order, while NlSP does. For
example, an assemblage may contain Cervus elaphus, Odocoileus hemionus, and
Antilocapra americana. All these species belong to the larger Artiodactyla order. NlSP
allows the researcher to simply add all of the species counts together to find the total
number of artiodactyls. Using MNI, the researcher must throw out the MNI results for
each species category and recalculate it for the entire generic group.
Another method of quantification has been used to calculate original meat weight
(White 1953). This method multiplies MNI with the amount of meat on a carcass, thus
addressing the disparity between different sized species. Ten lagomorphs are clearly
not as large in meat weight as one Cervus elaphus. This technique removes the focus
from the amount of elements and places it on the overall amount of meat available for
each species, however it suffers from the same problems as MNI because the
calculation is based upon MNI. Others have weighed the bones from each species,
arguing that the relationship of bone weight to meat weight is directly correlated (Klein
and Cruz-Uribe 1984). However, this is very problematic because it ignores the fact that
many larger bodied game animals are field dressed and only certain portions of the

animal are carried back to habitation sites. Given differential transportation of elements
and differential preservation of elements, large dense bones might be more common
than less dense bones (Casteel 1978; Chaplin 1971). Additionally, Casteel (1 978) has
shown that the positive correlation between bone weight and meat weight does not hold
true for all species. In pigs the reverse is true: as the meat weight increases, bone
weight decreases (Casteel 1978).
Given all of the problems with simply counting elements or trying to estimate
species numbers with pairing, matching, and meat weight, one is left to choose between
intrinsically flawed methods. Additionally, Grayson argues that MNI and NlSP are tightly
correlated, so that any information gained by calculating MNI is already to be found in
the NlSP (1984: 62-63). MNI has other flaws: 1) there is no standardized method for
calculating MNI, thus making it difficult to compare data from different sites; 2) MNI may
change drastically depending on how a sample is aggregated; 3) MNI does not take
generic identifications into account; and 4) MNI does not allow the researcher to "total"
species into a more generic group. Thus, NISP, by its very nature is standardized
allowing researchers to compare data sets from different sites and can only be
calculated by counting each fragment. Calculating NlSP does not encounter the same
problems with sample aggregation, as does MNI, because it treats each fragment as a
separate specimen (Grayson 1979, 1984). Using NlSP allows the researcher to utilize
generic categories for specimens that cannot be identified to the level of species. This
method also allows the data to be further analyzed for skeletal part frequencies, thus
correcting the problem of under or over representing certain elements (Grayson 1984).
Finally, NlSP allows for the easy addition of species to calculate the total within an order.
Keeping all of this in mind, NlSP is arguably the better choice for the purposes of this
study because it allows more freedom in analysis and comparison.

Stable Isotope Analysis
Stable isotope analysis measures the ratios of certain stable isotopes (such as
I3c to I2c or I5N to I4N) in bone collagen (Ambrose 1993; Chisholm 1983; Matson and
Chisholm 1991; Schoeninger and Moore 1992; Van der Metwe 1982). Stable isotope
analysis is employed to reconstruct prehistoric diets in animals and in humans. The
ratios of stable carbon isotopes I3c and I2c are used to distinguish between temperate
grasses, trees and shrubs (or C3 plants) and tropical and savanna grasses (or C4
plants) (Van der Metwe 1982). Plants that fix carbon dioxide into three carbon
molecules (C3) incorporate less I3c into their tissue than plants that fix carbon dioxide
into four carbon molecules (C4) (Van der Merwe 1982). This ratio of I3c to I2c is passed
through the food chain and becomes fixed in the bone tissue of the organisms eating the
plants. In the American Southwest both C3 and C4 plants are present. Native flora
(with the exception of Opuntia, or prickly pear cactus) are C3 plants, while the introduced
plant Zea mays (corn) is a C4 plant. By measuring the ratio of I3c to I2c isotopes in
turkey and lagomorphs in the Shields Pueblo faunal sample, it will be possible to assess
whether turkeys are being tended by being fed table scraps or from household stores of
corn, or if they are being left to run loose to forage for themselves. In lagomorphs, the
I3c to '*C ratio will give us some idea as to where the Lepus and Sylvilagus are coming
from. Are they being hunted outside of the boundaries of the site or are they being
trapped in the cultivated fields?
There are a series of wild, local plants that are also C4 plants. These include:
saltbrush (Atriplex), amaranthus (Amaranthus), and members of the purselane
family(Portu1aca ) (Katzenberg and Kelley 1991). Crassulacean acid metabolism (CAM)
plants native to the Southwest include various cacti, such as cactus pear (Opuntia sp.)
and hedgehog cactus (Echinocereus). While CAM plants are not strictly C4 plants, they
do produce similar ratios in carbon isotope studies (Bergfeld, Bergmann, and von

Sengbusch 2006). It is possible that domestic birds may have eaten wild C4 or CAM
plants as they foraged. Therefore, it is important to look at nitrogen levels in the Shields
Pueblo Fauna as an additional indicator of how free-range these birds were. It is also
possible that domestic turkey were fed wild, gathered plants as part of their diet. In this
case, it is not possible to tell the difference between wild C4 plants and maize.
However, given the availability of maize in the household, it is more likely that turkey
were fed from domestic grain stores.
The ratio of 15N isotopes to I4N isotopes increase as they move up the food chain
from plants to animals. A low ratio of I5N to 14N indicates an agricultural or plant food
oriented diet (Schoeninger and Moore 1992; Sponheimer et a/. 2003). Organisms that
depend on meat (or blood or milk) have higher 15N values than those dependent upon
plants (Ambrose1993). For this project, nitrogen values are used to ascertain if turkeys
were eating primarily plant food (fed by hand) or were allowed to forage and eat
invertebrates. This could provide some insight into whether they were left to roam or
were penned in some manner, thus limiting people's access to wild foods.
Sampling Procedures
Samples for stable isotope analysis were chosen by species (Meleagris
gallopavo, Lepus sp. and Sylvilagus sp.). The number of samples that could be taken
was constrained by budget. A total of 18 dated individuals were removed from the
collection. Single elements from a total of ten turkeys were removed from the collection
(two from Middle Pueblo II and Late Pueblo II, and three each from Early Pueblo Ill and
Late Pueblo Ill). One element from each time period was sampled for both jackrabbit
and cottontail rabbit. Taxon samples for each time period under consideration were
chosen randomly and came from an equal number of structure and non-structure
contexts (to try and avoid bias).

The samples were prepared by Cheryl Takahashi from the Simon Fraser
University Department of Archaeology Archaeometry Lab. First the sample bone
surfaces were brushed lightly with a wire brush to remove surface contamination.
Further possible contamination was removed by milling (removing a thin portion of the
exterior) with a dremel tool fitted with a carbide burr. Each bone was then drilled with a
carbide bur to produce clean shavings. A 20 mg portion of each sample underwent
standard collagen extraction. Each sample was soaked in weak acid to de-mineralize.
The insoluble protein matrix was then soaked in even weaker acid under low heat for
several hours. The resulting solution was ultra-filtered and freeze-dried in order to
isolate and concentrate high molecular weight protein. A portion of each sample was
then sent to Iso-Analytical for stable isotope analysis. The results of these analyses will
be discussed below.
Taxon Frequencies
Shields Pueblo yielded an extraordinarily large number of faunal specimens. In
fact, it is one of the largest assemblages from the Northern San Juan Region. I
analyzed a total of 40,952 specimens. Of these fragments 18,770 (46%) are identifiable
to element and are thus assigned to a taxon. Table 6 presents the relative frequency of
faunal remains by class. Mammals are the dominant class with 13,241 total fragments.
Birds are the second largest class represented by 5,406 fragments. Amphibians and
Reptiles are the least abundant totaling only 8 fragments for amphibians and 96 for
reptiles. Gastropoda, in the form of land snail, is represented by 6 shell fragments.

Table 6. Relative Frequency of Faunal Remains by Class
Mammalia
A ves I I I
Mammals
Birds
Amphibia
Gastropoda
Total Identified
Total Unidentified
Mammals
A total of 47 taxa were identified in the sample (including miscellaneous
taxonomic assignments such as "medium mammal"). Table 7 presents these data,
providing the NlSP and relative frequency of a specific taxon among the mammals as
well as among all identified taxa. Lagomorphs are the most common taxon among the
mammals with a NlSP of 9,813 individual fragments or 74% of all mammal remains.
There are three categories of lagomorphs (or rabbits): a general category, Lagomorpha
(used when a specimen can not be distinguished at a genus or species level); Lepus sp.
(jackrabbits and hares), and Sylvilagus sp. (cottontail rabbits). Elements categorized as
Lagomorpha were generally fragmented or broken in such a way as to make specific
assignment impossible. In southwestern Colorado there were at least seven different
species of lagomorphs present (Ochotona princeps, Sylvilagus sp., Sylvilagus nuttallii,
Sylvilagus audubonii, Lepus sp., Lepus americanus, and Lepus californicus)(Driver
1999:7).
Grand Total
13241
5406
Amphibians
Land snail
40952
70.5%
28.8%
8
6
18770
22182
0.0%
0.0%
100.0%

Table 7. Relative Frequency of Mammals
(~agomorpha (~agomorpha lrabbits and hares 174 10.56% (0.40% J
Oder lnsectivora
l ~ e p u s sp. hackrabbit or hare 11 685 (1 2.70% 18.98% I
Taxan .. lnsectivora
Rodentia
- '
cornman man Name " '
insectivores
Sciuridae Spermophilus sp. Cynomys sp. Thomomys sp.
Sylvilagus sp. Rodentia
Muridae Peromyscus sp. Neotoma sp.
I Ismall rodent lwoodrat or smaller 167 10.50% 10.36% I
NISP 1
squirrels ground squirrels prairie dog pocket gopher
Neotoma cinerea Microtus sp. Erethizon dorsatum Castor canadensis
]carnivora l~arnivora lcarnivore 118 10.13% 10.09% 1
cottontail rodents
deer mice, voles, etc. mice wood rat
?4 of Mam- 0.008%
113 1 730 480
bushy-tailed wood rat vole porcupine beaver
%dl Taxa 0.00%
8054 157
6 43 202
Canidae Canis sp. Canis latrans
1 Gulo luscus lwolverine 11 10.00% 10.00% 1
0.85% 0.00% 5.50% 3.62%
2 3 22 31
Canis lupus Canis familiaris Vulpes sp. Vulpes vulpes Mustela sp. Mustela frenata
62.80% 1.20%
0.60% 0.00% 3.90% 2.56%
0.05% 0.32% 1.50%
dogs, wolves dog, wolf, coyote coyote
I 1 ~ y n x rufus ]bobcat 117 (0.13% 10.09% J
42.90% 0.84%
0.03% 0.23% 1.07%
0.01 % 0.02% 0.17% 0.23%
wolf domestic dog red or kit fox red fox weasel long-tailed weasel
Taxidea taxus Felidae Felis concolor
Ismall carnivore lsmaller than fox 119 10.10% 10.1 0% I
0.01 % 0.02% 0.12% 0.1 7%
72 207 5
(medium carnivore (fox size or larger 17 10.05% 10.04%
2 129 4 2 1 1
badger cats mountain lion
0.50% 1.60% 0.04%
0.38% 1.10% 0.02%
0.01 % 0.97% 0.03% 0.01% 0.00% 0.00%
8 6 1
-
0.01 % 0.69% 0.02% 0.01 % 0.00% 0.00%
0.06% 0.07% 0.00%
0.04% 0.03% 0.00%

Orcter % =
Artiodactyla Cervidae Cen~us elaphus
L~isc. (small mammal 1 (7 10.05% 10.04% I
Common Name artiodactyls
t & 9
axon Artiodactyla
Odocoileus sp. Antilocapra americana Ovis canadensis medium artiodactyls large artiodactyls
I (medium mammal 1203 11 SO% 11.10%
deer family wapiti
(large mammal I TOTAL I
Deer pronghorn bighorn sheep deer size artiodactyls wapitilbison sized
. - I I . - - . - . - - . - total identified I I I I I I
- %all Taxa 1.02%
NlSP 191 57 4
The faunal collection at Simon Fraser University does not have examples of all of
% of Mam. ,
1.40%
581 2 6 3 7
these species in the comparative collection. Therefore, it was not possible to identify
0.40% 0.03%
Lagomorpha specimens beyond designating them as Lepus or Sylvilagus. Also, many
0.30% 0.01%
4.40% 0.01 % 0.05% 0.02% 0.05%
of the distinctions between species are not readily recognizable in skeletal remains.
3.10% 0.01 % 0.03% 0.02% 0.04%
Size of skeletal elements was the primary characteristic used to distinguish between
Lepus and Sylvilagus. No lagomorphs smaller than Sylvilagus, such as pika, were
recovered. Sylvilagus sp. were the most common form of lagomorph in the Shields
Pueblo faunal assemblage with a NlSP of 8,054 fragments; while Lepus sp. totaled
Rodents are the next most common order with some 1,857 fragments (14% of all
mammals). Sciuridae, Cynomys sp., Thomomys sp., and Neotoma sp. (squirrels, prairie
dog, pocket gopher, and wood rat) are the most abundant rodent taxa. Small rodents
including Muridae, Peromyscus, and Microtus were found in smaller quantities. There
were also a number of nonspecific small rodents (smaller than woodrats). Large

rodents, represented by Castor canadensis (beaver) and Erethizon dorsatum
(porcupine) were also present in the collection.
The next most common taxonomic group is the Artiodactyla, consisting of a total
of 851 fragments. Artiodactyls constitute some 6.4% of the mammal total. Artiodactyls
were categorized into eight different taxonomic assignments: the order designation,
Artiodactyla; Cervidae (indicating the deer family); C e n m elaphus (wapiti); Odocoileus
sp. (either mule deer or white-tailed deer); Antilocapra americana (antelope); Ovis
canadensis (Bighorn Sheep); medium artiodactyls; and large artiodactyls.
Carnivora make up approximately 3.8% of mammals with some 500 fragments.
Canids (including dogs and foxes) are the most common carnivores with a NlSP of 421.
Cats are the next most common group with 24 specimens. Other carnivores present in
small numbers include Mustela sp., Mustela frenata, Gulo luscus, and Taxidea taxus
(weasel, long-tailed weasel, wolverine, and badger respectively). Some fragments could
not be identified to species, but were clearly carnivore. These were categorized by size,
small carnivore for animals smaller than a fox and medium carnivore for fox-sized or
larger.
Miscellaneous taxa (small, medium, and large mammals) are represented by 220
fragments. Fragments within this category were generally fetal, juvenile, or fragments of
bone (such as rib bodies) that could not be assigned to a specific species, but were
definitely mammal.
The least common order in the mammalian category are Insectivores. There are
two fragments identified in this category, one identified as lnsectivora and the other a
Soricidae.

Birds
Among the birds there are 19 taxa present (Table 8). The vast majority of these
specimens belong to Meleagris gallopavo and the large bird categories. Turkey and
large bird have been combined, as "large bird." This designation was used to identify
elements that were probably turkey, but were highly fragmented. In total, turkey and
large bird have a NlSP of 5,219 (some 96.5% of the birds and 27.8% of all identified
taxa). Other species of bird are present in small numbers. There are 91 medium-sized
birds (mallard sized and smaller) accounting for 1.7% of all birds. Small birds (robin
sized and smaller) have an NlSP of 27 (and a frequency of 0.50%). Falconiformes and
Strigiformes make up 0.66% with a total of 36 fragments. 12 Piciformes are identified
along with 6 Passeriformes (including one raven) and Columbiformes (NISP=4).
Table 8. Relative Frequency of Bird Taxa
I 1 ~athartes aura ]turkey vulture I 1 (0.02% 10.00% l~aliaeetus leucocephalus lbald eagle 1 2 10.03% 10.00%
Qrder Falconiformes
I l ~ c c i ~ i t e r gentilis lgoshawk ( 1 (0.02% 10.00% 1
NlSP 13
Taxon Falconiformes
I 1 ~eleagr is gallopavo lturkey (4195 (75.60% 122.35%
% of Bird 0.24%
Common Name vultures, hawks, eagles
Galliformes
% all Taxa 0.07%
l~asseriformes l~asseriformes herchina birds 1 2 10.03% 10.00% 1
Buteo jamaicensis Galliformes Tetraonidae spp.
Columbiformes Strigiformes
Piciformes
red-tailed hawk grouse, etc. grouse
Columbiformes Strigiformes Bubo virginianus Asio otus Piciformes Picoides ~ubescens
I corvus corax
total identified l taxa I
raven 1 4 10.07% (0.02% Miscellaneous
TOTAL
5 2 9
Pigeons and doves owls great horned owl long-eared owl woodpeckers downv wood~ecker
small bird medium bird large bird
0.09% 0.03% 0.17%
4 3 5 6 11 1
0.02% 0.00% 0.05%
smaller than robin smaller than mallard larger than mallard
0.07% 0.06% 0.09% 0.1 1% 0.20% 0.02%
0.02% 0.01% 0.03% 0.03% 0.06% 0.00%
27 91
1024 5406
0.49% 1.70% 19.94% 100.00%
0.14% 0.48% 5.46% 28.80%

Reptiles, Amphibia, and Gastropoda
There are only a few elements identified as belonging to the Reptilia, Amphibia,
or Gastropoda classes (shown in Table 6). Reptiles consist of a single element
categorized as reptile and 96 snake vertebrae for a total of 0.51 % of all identified taxa.
There are 7 elements identified to the category of amphibian. Gastropoda is the least
common taxa with only 6 total shell fragments.
Preservation/Taphonomic Biases
Preservation of bone is of utmost importance in regards to analysis and
interpretation of faunal remains. Taphonomic analysis allows interpretation of the
formation of an archaeological site and provides a measure for comparison between
sites. The basis of taphonomic analysis is to distinguish between natural and cultural
forces that both cause and affect site development (Davis 1987; Hesse and Wapnish
1985; Reitz and Wing 1999). Natural taphonomic forces are actors in the natural realm,
such as weather, geomorphic/geologicaI activities, and biotic activity (specifically the
activities of plants and animals) (Davis 1987; Hesse and Wapnish 1985; Reitz and Wing
1999). Cultural taphonomic forces are those perpetuated by human activity. These
include (but are not restricted to) activities such as hunting, butchering, cooking, and
discarding trash (Davis 1987; Hesse and Wapnish 1985; Reitz and Wing 1999).
There are several different kinds of taphonomic processes and effects that act
upon or change the appearance of a bone from the time the animal is living until it is
analyzed by an archaeologist. These processes affect the bone in different ways.
Processes might include accumulation (natural or cultural) or exposure. Likewise, there
are taphonomic effects such as, weathering (caused by sun, water, or acid in the soil),
erosion, decomposition, and biotic disturbance (carnivore scavenging, rodent activity,
root etching, etcetera) (Reitz and Wing 1999). Many of these processes and effects can

create similar patterns in the assemblage. This phenomenon is called equifinality. For
example, spiral fractures to a bone can be caused through cultural activity (such as
breaking and twisting during marrow extraction). Additionally, these breaks can be the
result of trampling (Haynes l983), natural traumatic injury to the animal (Lyman l984),
or through carnivore activity (Binford 1981). The problem of equifinality can be
overcome through various kinds of analyses used in the study of taphonomy.
Traditionally, taphonomy has been relegated to the realm of middle-range theory in
archaeology. Experimentation, observation, and ethnographic analogy have proven
useful for understanding how faunal remains enter the archaeological record and in what
condition (these methods are summarized in Lyman 1994).
Natural Accumulation
As mentioned above, bone may accumulate through natural or cultural
processes. Some of the bones found on archaeological sites are present because they
were purposefully brought to the site, modified in some way (butchered, cooked,
etcetera) and finally were deposited as garbage. Some bone, however, enters an
archaeological site through natural processes (Driver 1985: 18). Burrowing organisms
(particularly rodents) are common bioturbators of archaeological sites, often dying in situ
within burrows. Animals may also be brought to an archaeological site by carnivores or
scavengers at a later date (Brain 1980, 1981 ; Binford 1981 ; D' Andrea and Gotthardt
1984; Haynes 1980). In order to interpret whether faunal remains are naturally occurring
or cultural in origin it is necessary to distinguish natural from cultural accumulations.
Both of these will be discussed by defining or describing the "method" of deposition and
by detailing how these can be identified in the archaeological record. The Shields
Pueblo Faunal assemblage will then be analyzed (with these factors in mind) and
compared with other sites in the immediate region when possible.

Canid Accumulation
Naturally deposited bone is often the result of the scavenging or hunting activities
of predatory animals. This is often demonstrated by the presence of tooth marks (both
punctures and rounded grooves) and the absence of articular ends accompanied by the
aforementioned indicators (Binford 1981). Digestion may also alter bone. Bones that
have passed through the digestive tracts of carnivores are generally broken into "bite-
sized" pieces and often show evidence of chemical erosion (Korth 1979; Andrews 1990).
Carnivores (especially canids) will also lick the ends of bone, effectively polishing the
articular surfaces (Haynes 1980; Binford 1981 ).
In order to determine the extent of canid damage to the Shields Pueblo
assemblage all evidence for canid damage (tooth marks, chewing, digestion, and licking)
were recorded during analysis of faunal remains. These data are illustrated in Table 9
for all of the major taxonomic groups.
Table 9. Frequency of Carnivore Modification-Shields Pueblo
I I
Sciuridae ( I 324) 17 1 0.5%
Lagomorph (74)
Lepus sp. (1 685)
Sylvilagus sp. (8054)
I I
M. gallopavo (52 1 9) 1 141 12.7%
0
40
62
0
2.4%
0.8%

Over all, the frequency of carnivore damage is quite low. If one includes all
identified and unidentified taxon only 0.87% of the assemblage has been damaged by
carnivores. Table 10 shows the frequency of carnivore scavenging for Woods Canyon
Pueblo, Castle Rock Pueblo, Yellow Jacket Pueblo, and Sand Canyon Pueblo. Such as
Shields
Table 10. Frequency of Carnivore Modification for Selected Taxa (site comparison)
Lagomorph I 6.25 1 2.00
I Lepus I 9.09 3 2.83
Sylvilagus 2 .995 12 1.42
Sciuridae 0 1 1.05
M. gallopavo 9 3.65 21 7.39
(Driver 2000,2002; Muir 1999:56; Muir and Driver 200:
Pueblo, the frequency of carnivore damage never rises into the double digits. Evidence
of scavenging or canid damage does not necessarily prove that a bone is present
through natural accumulation. The Anasazi kept domestic dogs (Szuter 2000). If we
consider ethnographic accounts, it is clear that dogs had free range through habitation
areas and would have been able to access garbage including faunal remains. Damage
from domestic dogs is indistinguishable from wild dogs and other carnivores. In this
instance, then, the evidence of canid scavenging does not necessarily indicate the
natural accumulation of faunal bone. Instead, it implies something about how garbage
was deposited, namely that it was not easily accessible to domestic dogs. It also

implies that the bone is culturally accumulated and not the result of wild canid
huntinglscavenging.
Raptor Accumulation
Wild and domestic canids are not the only sources of naturally accumulated
faunal assemblages. Raptors (or birds of prey) are also known to "build" assemblages
of various rodents and lagomorphs. Not all bones consumed by predators show
evidence of chewing. Owls typically consume rodents whole and deposit fairly complete
skeletal remains in owl pellets (Dodson and Wexlar 1979; Kusmer 1986; Andrews 1990).
Raptors are also known to take larger prey and create assemblages of mixed rabbit
remains (Hockett 1991, 1995; Quirt-Booth and Cruz-Uribe 1996). No owl pellets were
recovered from Shields Pueblo, nor were there instances of large mixed species of
"complete" rodent skeletons. The possible effects of raptor activity will be discussed
further (in reference to the rabbit assemblage) below in the discussion of skeletal part
frequencies.
Other Natural Accumulations
The final category of naturally accumulated bone is from animals that died on site
by natural death. Most natural deaths are those of burrowing animals that have tunneled
into the site and simply died in their burrows (Driver 1985). Animals that have died in this
manner should therefore show very little damage and should be fairly complete with
intact skeletons.
Accumulation due to natural death can be analyzed by looking at the
completeness of rodent bones in the assemblage. The frequencies of completeness
and fragmentation for selected burrowing rodent species is illustrated in Table 11.

The frequencies of complete bones at Shields Pueblo are comparatively low,
none going above 60%. The smallest species of rodents, Muridae and Microtus, have
unexpectedly low frequencies of complete bone. In comparison, rodent species at Sand
Canyon Pueblo are between 67.6% and 84.3% complete (Muir 199956). At Shields
Pueblo, all of the burrowing rodents are only slightly less complete than the burrowing
species from Sand Canyon Pueblo. Thus, the Shields Pueblo rodents are likely present
through natural processes.
Table 11. Frequency of Complete and Fragmented Bone - Rodents
Cultural Modification
Understanding cultural modification is of utmost importance to zooarchaeological
research because it is direct evidence of human behavior. Culturally deposited faunal
remains are identified by the presence of cut marks, breakage patterns, grinding,
polishing, and evidence of burning (Driver 1999; Fisher 1995; Olsen 1980). Each of
these will be discussed in turn below. Included in each discussion will be the results of
the Shields Pueblo faunal analysis and how these results compare with other sites in the
McElmo Dome region.
Damage due to butchering practices is easily identifiable as evidence of human
activity (Fisher 1995). Cut marks are linear and v-shaped in cross-section (quite
different from u-shaped canine tooth marks) and, such as canid chewing, are typically
found at the proximal or distal ends of bone. This pattern is typical of disarticulation and
Neotoma sp.(205)
Small Rodents (52)
92 44.9%
33 63.5%
113 55.1%
19 36.5%

meat removal (Binford 1981). Cut marks are not purposefully applied, instead they are
"accidents of motion" (Turner and Turner 1999:18). Therefore, the placement and
patterning of cut marks are simply artifacts of particular butchering techniques. Table 12
shows the frequency of occurrences of cutmarks for selected species from Shields
Pueblo, Woods Canyon Pueblo, Castle Rock Pueblo, Yellow Jacket Pueblo, and Sand
Canyon Pueblo.
Certain breakage patterns are also indicative of human modification (Driver
1999). Breakage can occur through impact blows. Zierhut (1967) describes the process
of marrow extraction from long bones using hammers and the blunt ends of axes.
Breaking bone open in this manner results in spiral fractures, longitudinally splintered
breaks, and finely broken bone chips. Zierhut also noted that after marrow extraction,
bone was further broken up for degreasing, leaving only the distal and proximal ends
Table 12. Culturally Modified Bone - Cut Marks
(Driver 2000,2002; Muir l999:56; Muir and Driver 2003)

identifiable (1 967:35). Bone breakage may also occur through snapping and twisting
(especially with smaller bird and lagomorph bones).
Despite the possibility of natural causes (through trampling) (Binford 1981 ;
Bonnichsen 1973; Haynes 1983), spiral fractures have commonly used to identify
culturally modified bone. Spiral fractures were included in this research for the purposes
of providing other researchers with data for comparison. Likewise, given that the faunal
assemblage represents a cultural deposit, and would not have been effected by
trampling of large animals, spiral fractures are likely cause by human breakage. Type
of breakage was recorded for each specimen in the Shields Pueblo faunal assemblage.
Table 13 provides the frequency of spiral fractures among selected species for Shields
Pueblo as well as sites in the immediate region. The highest frequencies of spiral
fractures among Shields Pueblo fauna are found among the carnivore, medium bird,
falconiformes, and rabbits. Artiodactyl is lower than would be expected (compared to
Sand Canyon Pueblo). However, as noted by Zierhut (1 967) deer remains may undergo
far more processing than other taxa, resulting in greater fragmentation and obliteration of
spiral fractures.
The presence of either spiral fractures or splintered breaks are often more
frequent in economically important species (Muir 1999:56). These species undergo far
more processing through butchering, removal of bone marrow, fat extraction, and
manufacture into tools. Therefore, economically important species are often more
fragmented than naturally deposited species. Table 14 shows the frequency of complete
and fragmented bones for selected species for Shields Pueblo. Overall, this table
illustrates that sites with smaller samples show more variation, while the larger site
samples are fairly similar. In general there is a high frequency of breakage among

Table 13. Culturally Modified Bone - Spiral Fractures
Artiodactyla Lepus sp. Sylvilagus sp. E. dorsatum
(Driver 2000,2002; MZ 1999:56; Muir and Driver 2003). -
C. canadensis Rodentia Sciuridae
Table 14. Frequency of Breakage for Selected Taxa at Shields Pueblo Compared with Other Sites on the McElmo Dome
13 1.5 35 2.1 227 2.8
-
* S.P.= Shields Pueblo; W.C.P.= Woods Canyon Pueblo; C.R.P.= Castle Rock Pueblo; Y.J.P.= Yellow Jacket Pueblo; and S.C.P.= Sand Canyon Pueblo (Driver 2000,2002; Muir 1999:56; Muir and Driver 2003)
- 1 0.6
-
economically important species. Artiodactyla show the highest frequency of breakage at
- 4 36.3 15 7.4
nearly 78% followed by Sylvilagus sp. at 75%, Lepus sp. at 70%, and Meleagris
- -
gallopavo at 61%. This seems to be strong evidence of cultural deposition. However, if
1 1.8 6 5.6 57 6.7
-
these frequencies are compared to the frequency of fragmentation among the Sciuridae
- 1 4.1 3 3.1
(60.3%) it is clear that there are other factors influencing fragmentation. If the frequency
4 1.7 37 14.3 117 10.1
-
25 3.7 13 9.6 125 5.3
4 10.0 -
2 2.0 -
9 0.9

of breakage at Shields Pueblo is compared with Woods Canyon Pueblo, Castle Rock
Pueblo, Yellow Jacket Pueblo, and Sand Canyon Pueblo, then it becomes clear that all
of these sites and their faunal remains show similar breakage patterning.
Grinding and polishing are also indicative of human activity. Bone may be
ground and polished during tool or artifact manufacture or through repetitive use (i.e.
wear marks) (Olsen 1980). All evidence of grinding and polishing was recorded for the
Shields Pueblo faunal assemblage. Table 15 presents these data for Shields Pueblo
and other sites in the McElmo region. In general, the artiodactyls have the most frequent
occurrences of grindinglpolishing among the Shields Pueblo fauna, followed by turkey
and Lepus sp. When compared to other sites in the region, Shields differs slightly (in its
lack of bobcat, for example). However, there are some basic patterns across all sites.
Artiodactyls, turkeyllarge bird, and Lepus sp. are common sources of bone for tools
throughout the region.
Table 15. Culturally Modified Bone - GroundIPolished
T m n
Artiodactyla Lepus sp. Sylvilagus sp. E. dorsatum C. canadensis Rodentia Sciuridae
Woods Canyon Pueblo N % 1 25.0
Shields PwMo
N % 58 6.8
Neotoma sp Thomomys sp. Cynomys sp.
27 1.6 17 0.2
- - -
Carnivore Canis sp. Lynx sp. Falconiforme M. gallopavo Medium bird Large bird
-
- Sand Canyon Pueblo N % 59 8.8
N %
3 5.4
- -
- -
(Driver 2000,2002; Muir 1999:56; Muir and Driver 2003)
3 1.4 -
96 2.3
3 0.3
Puclabk, N % 9 3.8
-
3 28.3 2 0.2
-
1 25.0 -
13 5.3
9 3.5
- -
2 0.8 -
- -
-
- 34 11.9
4 0.9
5 3.7 4 0.2
-
-
- - -
-
-
21 4.2 -
16 7.6 8 19.0
132 9.1
-

Burning is the final example of human modification considered herein. Bone is
typically burned unintentionally through the cooking process. Cooking meat with the
bone in results in a particular pattern of burning. Articular ends (where bones are
generally separated during butchery and left bare) are often burned while the shaft
remains undamaged. This localized pattern of burning is described by Driver (1999:24).
Bone may also be burned after disposal (thrown into the cooking hearth or by being
deposited in a structure that is then burned). In these cases bone may be carbonized
(partially or completely) or even calcined (grey or white) if the temperature of the fire is
great enough (Shipman et al. 1984). Bones with evidence of burning (e.g., localized
burning, carbonization, and calcination) were recorded for Shields Pueblo and are
compared with the frequencies from sites in the McElmo Dome (Table 16). While
Shields Pueblo seems to have a higher number of species with evidence of burning, it
follows the basic pattern seen among other sites in the region. Artiodactyls, canids,
Lepus, Sylvilagus, and turkeyllarge bird are all more frequently burned than other
species.
If Tables 12 through 16 are considered, some basic conclusions about the
cultural modification of the Shields Pueblo faunal assemblage can be made. In general,
the frequencies for each category of human modification lie within the ranges of
frequencies from Woods Canyon Pueblo, Castle Rock Pueblo, Yellow Jacket Pueblo,
and Sand Canyon Pueblo. However, Shields Pueblo fauna typically shows less
modification if compared to the average frequencies from the other sites. Cultural
modification of major species for all sites is summarized in Figure 6.
Some of the frequencies from these pueblos are much higher than from Shields
Pueblo. For example, at Woods Canyon Pueblo 25% of all Artiodactyla have burning
and polishing, but this is due primarily to the small size of the sample. At Shields

Pueblo, only a few species (such as Lynx, "carnivore", medium mammal, and porcupine
[Erethizon dorsatum]) have frequencies in the double digits (again, because the number
Table 16. Culturally Modified Bone - Burned
Canis sp. Lynx sp. Falconiforme M. gallopavo Medium bird Large bird (Driver 2000,2002; Muir 1999:56; Muir and Driver 2003)
7 3.4 2 22.2
290 6.9 7 7.7 27 6.9
1 4.7
2 0.8
2 0.8
1 50.0
29 10.2 1 1.6 29 10.2
2 18.2
21 4.2
21 4.2
9 4.3 5 11.9
89 6.2
89 6.2

Frequency of Cultural Modification for Selected Species
.Shields Pueblo L=J
Figure 6. Frequency of Cultural Modification for Selected Species. *Please note that the combined relative frequencies for Woods Canyon Pueblo, Castle Rock Pueblo, Yellow Jacket Pueblo, and Sand Canyon Pueblo are presented as average frequencies for each species.
of specimens is very small). The rest of the hunted and tended species seem to show
less evidence of modification. Taxa that were generally very economically important to
the Anasazi (Artiodactyla, the Lagomorphs, and Meleagris gallopavo), while more
frequent in the assemblage, have lower frequencies of modification than would be
expected. Generally, one would expect that economically valuable faunal sources would
be culturally deposited, and therefore, should have a high frequency of cultural
modification.
That the frequency of cultural modification does not rise above 9% is not unlike
the frequencies from the other pueblos. This could be due to various taphonomic effects
(such as the extent of human processing and the amount of postdepositional breakage).
While much of the modified bone and bone tools identified at Shields Pueblo were
recognized as coming from specific species, they could not be identified to element and

as per the instructions in the CCAC Manual for Description of Vertebrate Remains, were
recorded as unidentifiable (Driver 1999). If they could have been recorded as
identifiable species, the frequency of modification for artiodactyls, rabbits, and turkey
would have been higher.
Sullegic Processes (Archaeological Decisions)
Another taphonomic process affecting the Shields Pueblo assemblage is the so-
called sullegic process, which includes all of the archaeological decisions that affect
what is recovered from a site (Efremov 1940). The most common of these are sampling
procedures and excavation methods. The objective of the Shields Pueblo excavation
was to identify and investigate the subsurface deposits. While some separate midden
areas were excavated, the majority of excavation focused on habitationlritual structures
(refer to Duff and Ryan 2000). It is possible that the focus on habitationlritual structures
affected the faunal assemblage by skewing the faunal counts in favor of species
associated with structures. However, it is important to note that the pit structures
themselves were often used as refuse sites upon their abandonment. Therefore, the fill
above floor contexts can still tell us something about refuse practices. Choosing to
focus on structures does not necessarily affect the outcome of the faunal analysis, it
might, however, affect the understanding of site layout and spatial analysis.
Excavation decisions (including screen size) are far more likely to have affected
the Shields Pueblo faunal assemblage. In order to understand how screen size affects
an archaeological assemblage it is necessary to look at the size of the specimens
recovered. Above it was noted that larger fragments are more frequent in the
assemblage than smaller ones. Likewise, if the size of species is considered, larger
species are more common than smaller ones and larger bone elements are more
common than smaller ones (this will be discussed further in Skeletal Part Frequencies).

Thus, small specimens are more frequently lost, while large ones are easier to locate
and do not slip through screens.
Post-depositional destruction of bone
Once deposited bone can be destroyed by various topic processes. Physical as
well as chemical agents may further affect the condition or survivability of bone (Clark
and Kietzke 1967). The Shields Pueblo assemblage seems to have been affected to
some extent by postdepositional breakage (due to historic farming among other things)
and, to a lesser extent, by weathering. Both of these factors will be analyzed and
discussed for the Shields Pueblo assemblage and compared with sites in the McElmo
region. The faunal remains from Shields Pueblo may seem to be quite fragmentary in
nature (as indicated by the number of unidentified taxa and in the amount of specific
types of bone breakage). Of the 40,952 fragments recorded at Shields Pueblo, 22,182
were fragmented beyond recognition. Figure 7 shows the relative frequency of complete
and fragmented bone by taxon for Shields Pueblo. Approximately 54% of the
assemblage was unidentifiable. However, 40% to 60% of assemblages from sites in the
Northern San Juan region are also unidentifiable (see Driver et a/. 1999; Muir 1999; Muir
and Driver 2003). Therefore, the rate of identification at Shields Pueblo is similar to
other large sites in the region. The site of Shields Pueblo, unlike other pueblo sites in
the region, is predominantly subsurface. Very little remains of surface architecture (with
the exception of room-block structures in architectural block 100). The site lies on land
with a long history of agricultural use. The plow zone ranges in depth from 20 to 40 cm
across the site. This may explain the lack of surface architecture and fragmentation and
mixing of deposits within the top 40 cm, but it does not explain the amount of bone

breakage found throughout the assemblage.
Relative Frequency of Complete and Fragmented Bone
Taxa
Figure 7. Relative Frequency of Complete and Fragmented Bone: Shields Pueblo
Size of fragments may be an indication of postdepositional factors. The average length
of a bone fragment in this collection is 3.43 cm, which is similar to the average lengths of
fragments from Woods Canyon Pueblo, Castle Rock Pueblo, Yellow Jacket Pueblo, and
Sand Canyon Pueblo) (Driver 2002; Muir 1999; Muir and Driver 2003). Therefore, some
other factor (common to all sites) is indicated. Some species, because of their size,
density, etc., are more likely to be fragmented (and become unidentifiable elements) or
are destroyed, and thus are not preserved in the archaeological record.
In addition to breakage, bone may be destroyed by exposure to the elements.
The degree of destruction due to weathering depends upon several factors: the time
span between deposition and burial, the size of the element, the density of cortical
structure, amount of grease in the bone, and various cultural processes (Muir 1999:49).

Evidence of weathering was recorded in regard to particular breakage patterns as either
"D", for eroded break, or "P" describing a splintered break. Degree of weathering
(e.g., light to heavy) was not recorded. Table 17 illustrates the frequency of weathered
breaks among selected taxa at Shields Pueblo. Table 18 shows the frequency of
weathered breaks among selected taxa for Woods Canyon Pueblo, Castle Rock Pueblo,
Yellow Jacket Pueblo, and Sand Canyon Pueblo. Over all, artiodactyls were more
frequently weathered than other fauna. Muir argues that this pattern is due to the
deposition of particular fauna on the roofs of tower structures (1 999:93). This practice is
likely associated with ritual hunting for religious events. At this point it is unclear if there
are any towers at Shields Pueblo, although a greater frequency of weathered artiodactyl
remains may indicate the placement of artiodactyl remains on the roofs of houses
(similar to what White (1 974) noted among the Sia).
Table 17. Frequency of weathering among faunal remains, Shields Pueblo.
Table 18. Frequency of weathering among faunal remains (site comparison)
T m n
Artiodactvla . - ., -
Lagomorph Lepus sp. Sylvilagus s p. M. gallopavo Sciuridae
Yellow &&et Puablo PI %
18 7.8
Sand Canyon Puebfo N %
1 64 24.6
WOOCIS. Canyon Pueblo
N % 1 50.0
(Driver 2002, Muir and Driver 2003; Muir 1999)
1 6.25 1 9.09 1 .49 27 5.4 0
Castle Rock Pueblo N %
7 12.7 5 10.0 3 2.83 33 3.9 23 3.3 1 1.05
1 1.4 10 3.9 40 3.5 50 4.5 1 .98
-
NID 1 0.7 17 0.7 107 3.1 3 0.3

Summary of the Taphonomy of Shields Pueblo Faunal Remains
It is clear from the nature of the Shields Pueblo faunal collection, that the
distinction between culturally accumulated bone and naturally accumulated bone cannot
be based upon a single criterion. Instead, several lines of evidence (particularly human
modification) must also be considered. The economically important species
(Artiodactyla, Lagomorphs, and Turkey) all have relatively high frequencies of
fragmentation. It is clear that fragmentation, whether due to prehistoric cultural activity
or more recent taphic processes, greatly influences the general frequencies of cultural
modification because of the vast quantity of bone fragments. So, instead of focusing on
the frequencies of modification in "cultural" taxa, it is important to look at these
frequencies among species that are naturally occurring to a site. Rodent bone by and
large, shows less evidence of human modification (refer to Tables 12-16). Rodent bone
shows no evidence of cut marks nor grinding and polishing. None of the tools or other
bone artifacts are comprised of rodent bone (while the majority of identifiable modified
bone comes from Meleagris gallopavo, Artiodactyla, and Lagomorph). One of the
burrowing rodents, prairie dog (Cynomys sp.), has a spiral fracture frequency of 1.1 %
and burned frequency of 4%. While prairie dog is a burrowing rodent and may disturb
an archaeological site, it was also a prey species for the Anasazi, so it is no surprise that
a small percentage of these animals show evidence of human modification. If only
small and medium-sized rodents are considered, the frequency of spiral fracture falls
considerably. Only a single small rodent bone has a spiral fracture. Spiral fractures,
while common in culturally modified bone are not caused by human activity exclusively.
Carnivore damage, natural injury, and trampling can cause spiral fractures as well
(Binford 1981 ; Haynes 1983; Lyman 1984). It is interesting that there seems to be a low
frequency of spiral fractures among "cultural" taxa. However, this is likely due to a

combination of the high level of processing these species have undergone and post
depositional factors.
Skeletal Part Frequencies
Skeletal part frequencies are recorded in order to measure the degree to which
specific taxa are affected by taphonomic forces, in other words, to ascertain whether
certain elements preserve in higher frequencies than others. Both cultural and natural
processes influence skeletal part frequencies. Cultural processes include (but are not
limited to): the "Schlep Effect" (Marean et a/. 1992), when only meat-bearing skeletal
elements are removed from a kill or butchery site and carried back to the habitation site;
and the actual butchery process (how an animal is apportioned out into cuts) (Perkins
and Daly 1968; White 1953). The degree of completeness of a particular animal or
species can reflect human activity. For example, large-bodied taxa that are relatively
complete were likely killed near the habitation site. Butchering practices vary depending
upon prey size. Cooking practices may also influence which elements are more
frequently found in archaeological sites (Goody 1982). Groups that cook meat by
roasting or by cooking over open flame are more likely to leave the meat package in
larger units. Groups that cook meat by boiling or stewing are more likely to cut meat into
smaller portions or to filet meat from the bone. Likewise, the size of cooking pots may
influence the size of cuts of meat (Goody 1982).
Natural processes may also influence skeletal part frequencies. Animals, such
as carnivores and even rodents affect faunal remains in particular ways. Carnivores are
very selective in the elements they choose (Marean et a/. 1992). Bones containing
marrow or grease are particularly attractive to scavengers, so certain long bones and
spongy bone are less likely to be recovered due to removal or destruction. Raptors are
also responsible for creating faunal assemblages. Hockett (I 991, 1995) describes the

characteristics of raptor created lagomorph assemblages. Assemblages of rabbit in
raptor pellets typically consist of more forelimbs than hindlimbs, more subadults, larger
quantities of vertebrae, and are generally incomplete; assemblages associated with
nesting areas are typified by more hindlimb than forelimb, more adults, fewer vertebrae,
and are mostly complete (Hockett 1995). Rodents are also active in changing element
frequencies. Typically, small burrowing mammals collect small elements (phalanges,
carpals, and other small bones), so these may also be absent from the faunal record
(Hoffman and Hays 1987). Other natural forces include geomorphological forces (e.g.,
eolian or alluvial deposition). Elements that are less dense are more likely to be carried
off or broken by these forces. For example, many studies done in East and South Africa
show that water and simple gravity differentially remove or destroy small, lightweight
bones and may cause bones of varying sizes to be sorted spatially (Butzer 1982;
Hanson 1980).
Shields Pueblo Skeletal Part Frequencies
For the purposes of this study, only certain taxa have been chosen for further
analysis. These taxa include Artiodactyla, Lepus sp., Sylvilagus sp., and Meleagris
gallopavo. These are the most common species found among the Shields Pueblo fauna
and are also considered more economically important as food and for their secondary
products (Gnabasik 1981). For some analyses, other taxa (Canidae, Sciuridae,
Cynomys sp., and all combined "wild bird") will be used for comparative purposes.
Skeletal part frequencies will be broken down by taxa and then by element group
(cranial, axial, pectoral girdle, forelimbs, pelvic girdle, hindlimbs, and extremities). These
frequencies are illustrated in Table 19 as MNE. Minimum number of elements (MNE),
first employed by Binford (1978), is a basic measure of skeletal part frequency. For
example, if an assemblage of artiodactyls has 15 proximal humerii and 25 distal humerii,

the MNE is the greater of the two values, 25. This can be calculated for separate
elements or for skeletal regions. Generally, MNE counts are lower for regions of the
body with fewer elements. The reverse is true for regions with multiple elements. This
pattern holds true for all of the taxon with the exception of the rodents. In these two
cases, the extremities are underrepresented due to the size of these elements and the
techniques used to recover bone during excavation. Over and under-representations
of elements can be eliminated by calculating the MAU (minimal animal units). MAU is
calculated by dividing the MNE by the number of times the element occurs in the body
(Lyman 1994). The MAU values have been calculated to describe the rate of
preservation. Minimal animal unit values for Lepus sp., Sylvilagus sp., and M. gallopavo
are presented in Table 20a,b, and c (note that thoracic and lumbar elements are not
included in the M, gallopavo MAU because these elements are typically fused together).
Artiodactyls have been separated because they yield the densest cortical bone (Table
21). These are given along with Brain's (1981) rate of survivability and Lyman's (1994)
average bone mineral densities for deer. The typical pattern that emerges shows that
denser bone is well represented, while lower density bone is less so. This is particularly
evident in the Artiodactyla remains, but can be seen throughout the collection of smaller
mammals.
Table 19. Skeletal Part Frequencies - MNE counts by skeletal region for major taxa
Lepus 1 93 1 192 ( 72 1 180 1 69 1 126 1 306 Svlvilaaus 1 61 3 1 523 1 493 1 1169 1 746 1 732 1 988
Artiodactyla Canidae
I Sciuridae 1 52
8 11
7 1 77
36 10
56 33
17 9
20 19
159 55

Table 20. MAU Values: Lepus sp. (A); Sylvilagus sp. (B); and M. gallopavo (C).
Femur
Lumbar
Glenoid 30 78.90% Distal 30 78.90% Proximal 21.5 56.60%
Proximal 32.40% Distal 28.40% Proximal 21.00% Centrum 20.80% Centrum 2.90%
Femur Femur Radius
Metatarsal l ~ i s t a l
Proximal Distal Proximal
57.11 20.70% Humerus Lumbar Thoracic
136 118
105.5
Proximal Centrum Centrum
49.20% 42.70% 38.20%
48.5 21.3 5.8
17.50% 7.70% 2.10%

Table 21. MAU Values: Artiodactyls.
Scapula Tibia Meta~odial Radius Metapodial Humerus
Glenoid Distal Distal
Femur Tibia Lumbar
Proximal Proximal Distal
Femur Humerus Thorasic Rib
The Shields Pueblo faunal assemblage seems to correlate with preservational
studies such as Brain's experimental studies in Africa (1981) and Lyman's (1994) study
on density and preservation (as indicated in Table 21). The Shields Pueblo Artiodactyl
assemblage is more closely correlated with Brain's skeletal ranking. This is likely due to
the fact that identical forces (humans and natural processes) affected the Shields Pueblo
assemblage. Shields Pueblo's correlation with Lyman's index is slightly less (this is
especially notable with the distal rib). However, it must be noted that Lyman deals
exclusively with density as a factor for preservation/loss (Lyman 1994). Obviously, the
preservation or loss of the Shields Pueblo assemblage has been influenced by more
than simply bone density.
The Shields Pueblo assemblage has likely been primarily affected by post-
depositional forces. This is clearly illustrated in the data for artiodactyls, as well as in
smaller mammals. Table 22 illustrates that the weaker long bone ends of cottontails and
prairie dogs are less frequent in the Shields Pueblo assemblage than the stronger distal
ends. This is typical for other sites in the region (Woods Canyon Pueblo, Castle Rock
6.5 6.5 6.3
Distal Proximal Centrum
I
6 5 4
Proximal Proximal Centrum Dorsal
Radius ( ~ i s t a l
100.00% 100.00% 96.90%
3.5 3
2.4
0.51 7.70%(1ntermediate 1 0.43
92.30% 76.90% 61.50%
2 1.5 0.9 0.9
High High Hiah
53.80% 46.20% 36.90%
0.36 0.5 0.5
High High High
30.70% 23.10% 13.80% 13.80%
0.5 0.5
0.39 Intermediate Low Intermediate
0.28 0.3 0.3
Intermediate Low Low Low
0.36 0.24 0.24 0.25

Pueblo, Yellow Jacket Pueblo, and Sand Canyon Pueblo) (Driver 2000, 2002; Muir
1999; Muir and Driver 2003).
Table 22. Frequency of Weaker Long Bone Ends for Sylvilagus and Cynomys
Age Estimation: Seasonality and Evidence of Turkey Production
Estimating age at time of death in faunal remains has been an important aspect
of zooarchaeological research since its development as a subfield of archaeology. Age
at death is useful for researching both hunted species and domestic species as it
provides information about seasonality and husbandry practices. Estimations can be
made using several different techniques concentrating on the dentition and epiphyseal
union of long bones. Studies using dentition as a measure of age typically look at tooth
eruption and tooth attrition. The majority of research in dental eruption and attrition have
been based upon studies of various domestic animals (pigs, sheep, cattle, and goats)
(Andrews 1973; Chaplin 1973; Davis 1987; Deniz and Payne 1982; Gibson 1993; Grant
1982; Hesse and Wapnish 1985; Moran and O'Connor 1994; Payne 1987; Wenham
and Fowler 1973). However, there is some research on eruption patterns for wild North
American species including mule deer (Main and Owens 1995) and buffalo (Frison and
Reher 1970). Only a few mandible fragments have been recovered from Shields Pueblo
and these are generally small and thus do not provide an accurate measure.
S@iIagus
Tib.
Hum. And Tib.
Hum.
Cynornys
Prox,
25
274
371
Prox.
97
f DM. %weaker end
455
1008
D M 553 78
' %waaker end ,
17.5 32.0
37.6
36.9
17
42
39
117
43.9
35.9
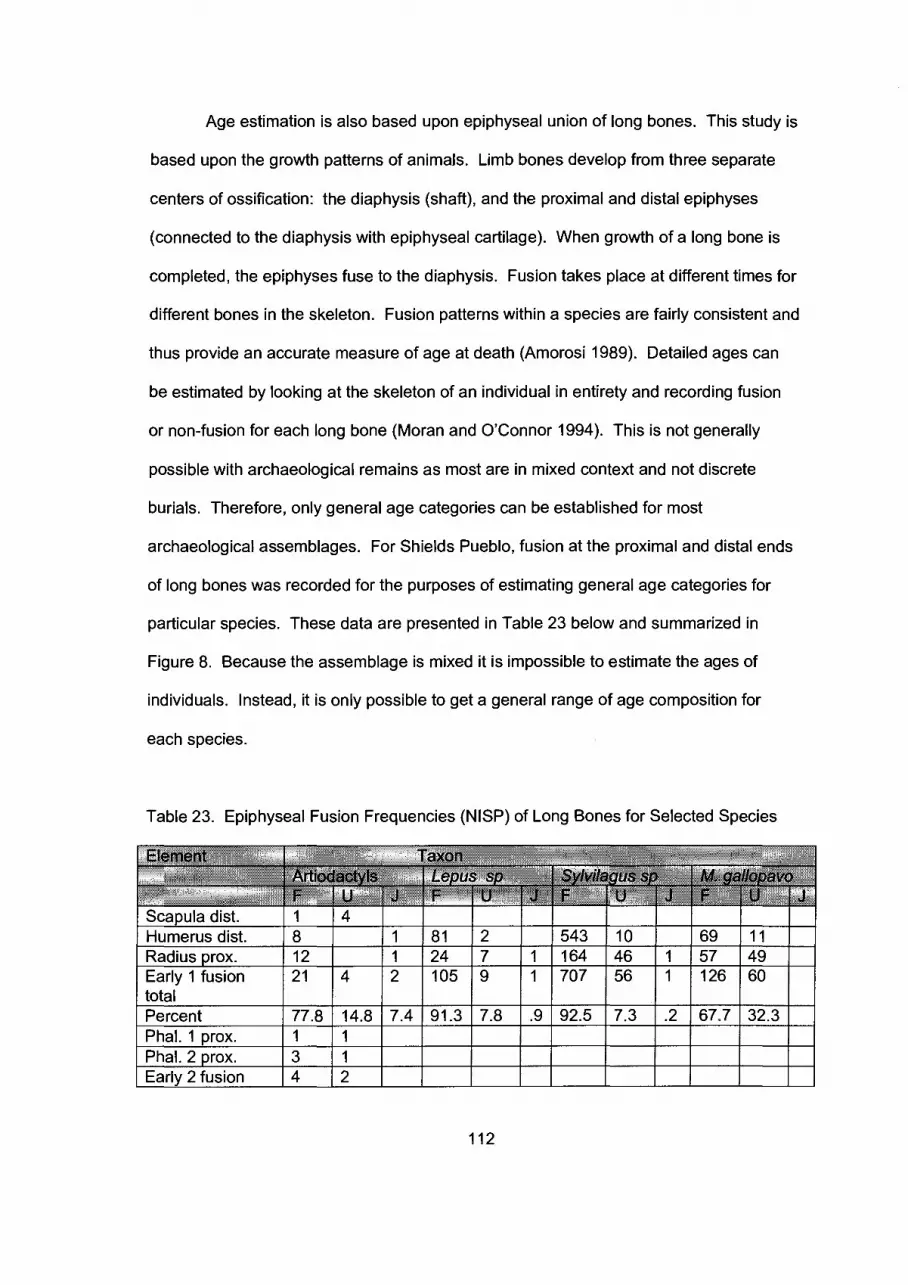
Age estimation is also based upon epiphyseal union of long bones. This study is
based upon the growth patterns of animals. Limb bones develop from three separate
centers of ossification: the diaphysis (shaft), and the proximal and distal epiphyses
(connected to the diaphysis with epiphyseal cartilage). When growth of a long bone is
completed, the epiphyses fuse to the diaphysis. Fusion takes place at different times for
different bones in the skeleton. Fusion patterns within a species are fairly consistent and
thus provide an accurate measure of age at death (Amorosi 1989). Detailed ages can
be estimated by looking at the skeleton of an individual in entirety and recording fusion
or non-fusion for each long bone (Moran and O'Connor 1994). This is not generally
possible with archaeological remains as most are in mixed context and not discrete
burials. Therefore, only general age categories can be established for most
archaeological assemblages. For Shields Pueblo, fusion at the proximal and distal ends
of long bones was recorded for the purposes of estimating general age categories for
particular species. These data are presented in Table 23 below and summarized in
Figure 8. Because the assemblage is mixed it is impossible to estimate the ages of
individuals. Instead, it is only possible to get a general range of age composition for
each species.
Table 23. Epiphyseal Fusion Frequencies (NISP) of Long Bones for Selected Species


The majority of specimens at Shields Pueblo (regardless of species) are fused.
Lepus sp. and Sylvilagus sp. have the greatest percentage of fused elements at 83.2%;
the artiodactyl elements have a 72.5% rate of fusion; and Turkey a rate of 63%. If bones
with different fusion rates (indicating different age categories) are considered, a different
pattern emerges. This is particularly evident among the artiodactyls where fused
epiphyses are far more common than unfused for: early 1 fusing bones (those fusing
between 6-1 0 months), early 2 fusing bones (fusing between 13-1 6 months), and middle
fusing bones (those fusing between18-28 months). However, if the late fusing bones
(those fusing between 36-42 months) are considered, fused and unfused epiphyses are
nearly equal. This means that just over half of artiodactyls were killed before they
reached maturity. Specific age ranges for fusion are not available for lagomorphs or
turkey, thus these species are presented in relative age categories based upon early,
middle, and late fusing bones. Among the lagomorphs, epiphyseal fusion of late fusing
bones indicates that approximately 113 of Lepus and Sylvilagus were killed before they
reached maturity. Overall, fusion rates indicate that among selected mammal taxa, few
very young juveniles were hunted. However, fusion rates among turkey differ. Early
fusing bone rates indicate that 113 of turkeys were killed as very young juveniles, while
only 115 were killed as older juveniles. So there does seem to be a slight preference for
very young birds when juveniles are killed.
Change in Taxon Frequency Over Time
The main focus of this dissertation is to understand how the sexual division of
labor (specifically concerning meat procurement) changes through time at Shields
Pueblo. In order to map changes in meat procurement strategies, the complete
habitation sequence (A.D. 725 - 1280) is considered with a particular interest in the
Pueblo II and Pueblo Ill sub-assemblages. The general frequencies (NISP) for

artiodactyls, all lagomorphs, and turkeyllarge bird are presented in Table 24. Figure 9
illustrates these relative frequencies. This provides the raw data for an analysis of
taxonomic ratios.
The purpose of this study is to understand the relationship between prey species,
thus faunal indices (based upon Bayham 1982; Bayham and Hatch 1985; Broughton
1994a and b) are used to measure the change in relative frequency over time. The
indices of the selected taxa are presented in Table 25. Three indices are used. The
artiodactyl index summarizes the extent to which artiodactyls contribute to the
assemblage. This is calculated by dividing the total number of Artiodactyla by the total
number of Artiodactyla plus the total number of lagomorphs. Figure 10 displays the
relationship between the artiodactyl index and time. The lagomorph index measures the
contribution of Sylvilagus in the assemblage. This has been calculated in different ways.
Bayham and Hatch (1985) use the following formula: Sylvilagusl total lagomorphs. This
is problematic for the Shields faunal data because a nonspecific category, "lagomorph",
has been used for specimens that could not be positively identified to cottontail or
jackrabbit. Since the category "lagomorph" is indeterminate, only specimens identified
Table 24. Relative Frequency of Selected Taxa - Change through Time
I 1 TurkevILarae Bird 1 263 32.6%


Artiodactyl Index
725-800 1020-1060 1060-1150 1150-1225 1225-1280
Years A.D.
Figure 10. Artiodactyl Indices: Shields Pueblo
as Sylvilagus or Lepus were used to calculate the lagomorph index. Thus, the
lagomorph index was calculated by dividing the total number of Sylvilagus by the sum of
Sylvilagus and Lepus (O'Hara 1994; Quirt-Booth and Cruz-Uribe 1997). Figure 1 1
shows the relationship between the lagomorph index and time. The turkeyllarge bird
index measures the proportional contribution of turkey to the assemblage by dividing the
total number of turkey and large bird by the sum of turkeyllarge bird and all of the
lagomorphs. Figure 12 illustrates the relationship between the turkey index and time.
Any index that approaches 1 .O, shows a high proportion of turkey relative to lagomorphs.
Values closer to 0.0 are indicative of higher 1agomorph:turkey ratios.

1 Lagomorph Index
~ Years A.D.
Figure 11. Lagomorph Index: Shields Pueblo
Turkey Indices per Sub Period
725-800 1020-1060 1060-1150 1150-1225 1225-1280
Years A.D.
Figure 12. TurkeylLarge Bird Index: Shields Pueblo.

LAC
Figure 13. Relationship between the artiodactyl index and lagomorph index. r= -0.669 (perfect negative correlation = -1 .o);?= 0.448.
There is an interesting pattern to note with change in species indices over time.
The ratio of artiodactyls to lagomorphs declines as the ratio of Sylvilagus to Lepus (the
lagomorph index) increases. Figure 13 illustrates this by plotting the artiodactyl index
against the lagomorph index. The correlation coefficient (r) is -0.669, and while not a
perfect correlation, this does indicate a strong degree of negative correlation between
the artiodactyl and lagomorph indices. Some 45% (? = 0.448) of the variance of the
lagomorph index can be explained by the artiodactyl index. There are no discernible
relationships between the lagomorph index and turkey index, nor between the artiodactyl
index and turkey index.
Another method of measuring the distribution of taxa through time includes
contingency table analysis. Contingency analysis tests the significance of variation in

taxon frequencies within different categories (time in this instance) to their expected
frequencies. Table 26 illustrates the observed (or actual) frequencies (NISP) of selected
taxa. Expected frequencies were calculated by pooling the data within each
chronological category. In order to perform this, the average frequency was established
for each taxonomic group. These were then used to extrapolate expected frequencies
for each taxon for each time period (Drennan 1996: 187-1 9 1). Expected frequencies
assume perfect distribution so that expected frequencies would be a constant
percentage of the total for each sub period (Table 27). Table 28 presents the
standardized residuals for taxonomic groups by sub period. Standardized residuals
were calculated by dividing the observed frequency minus the expected frequency by
the square root of the expected frequency [(observed-expected) I d expected].
The standardized residuals demonstrate high variability for each taxonomic
group over time, although, turkeyllarge bird and artiodactyls are more variable than
lagomorphs. If just the sub periods are considered, an interesting pattern appears.
Variability among the different sub periods differs quite drastically. Sub periods 3 and 9
show the least variability, while sub periods 7 and 8 vary the most.
Table 26. Observed Frequency (NISP) For Selected Species by Sub Period
Artiodactyls
Lagomorphs
TurkeyILBI
Total
10
147
35
192
101
342
280
723
400
3916
714
5030
7 1
2365
171 6
4152
20
524
263
807
602
7294
3008
1 0904

Table 27. Expected Frequency (NISP) for Selected Species by Sub Period
Artiodactyls 1 10.6 1 39.9 1 277.7 1 229.2 1 44.6
Lagomorphs 128.4 483.6 3364.7 2777.4 539.8
TurkeyILBI 53 199.4 1387.6 1 145.4 222.6
Total 192 723 5030 4152 807
Table 28. Standardized Residuals* for Selected Species by Sub Period
Pearson Chi-square= 1123.52; d.f.= 8; phi-square= 0.32; Cramer's V= 0.23; P=0.000 *Calculated as: (Observed-Expected)/&xpected. Values which fall beyond one standard deviation of the mean standardized residual are in bold. Totals represent sums of absolute values.
Artiodactyls Through Time
During the earliest period of occupation, sub period 3 (A.D. 725-800), the
artiodactyl index is approximately 0.06. This demonstrates a lower frequency relative to
lagomorphs. During early Pueblo II times, Shields Pueblo had been depopulated for a
period of 220 years (A.D. 800 - 1020). It was re-settled during the middle of the Pueblo
II period (sub period 6, A.D. 1020 - 1060). During this period of occupation the
artiodactyl index jumps dramatically to 0.2. This is the highest index achieved by
Artiodactyls at Shields Pueblo. After sub period 6, the artiodactyl index drops
precipitously. During the later half of Pueblo II (sub period 7, A.D. 1060 - 1150) the
index drops to 0.07. During the first half of Pueblo Ill (sub period 8, A.D. 1150 - 1225)

the artiodactyl index bottoms out at 0.03. This index remains unchanged during the sub
period just prior to depopulation (sub period, A.D. 1225 - 1280).
The pattern in the relative frequency of artiodactyl remains is not surprising given
the 220-year period of occupational hiatus at Shields Pueblo. Population during Pueblo I
in the Northern San Juan increased dramatically over the previous period (Rohn 1989).
The beginning of Pueblo ll saw a number of population relocations. During this period of
Shields Pueblo's depopulation, the local artiodactyl populations would have had time to
rebound if the site and surrounding area were not being used. When the site was
reoccupied in the middle part of Pueblo II, deer and other ungulates were more plentiful
and therefore made up a larger portion of the diet. Once human populations began to
grow in the region, there was a dramatic drop in the number of artiodactyls hunted,
evidenced by a precipitous drop in artiodactyl frequency in the faunal assemblage at
Shields Pueblo, and an increase in Sylvilagus and turkey.
This phenomenon is quite common in the assemblages from other sites across
the Northern San Juan region (Driver 2002). This is, in fact, evidence of resource
intensification. Based upon prey choice model, high ranked prey (prey whose net caloric
value is higher than the time and energy spent in pursuit and processing) will be taken
whenever possible, while low ranked prey (prey that is costly to pursue or process) is
taken based upon the availability of high-ranked prey (Bettinger 1991). As high-ranked
prey becomes scarce, lower ranked prey becomes more important as a resource (thus
increasing diet breadth, but decreasing foraging efficiency). Broughton (1994 a and b)
illustrates this trend in the Sacramento Valley. He is able to illustrate how the frequency
of large and medium-sized game (in this case artiodactyls, lagomorphs, and
anadromous fish) decrease while small fauna (small local fishes) increase over time.
This resource intensification is associated with an increase in sedentism and intensified
use of smaller site catchment areas (Broughton I994b:512).

This pattern is very similar to what is described in the Northern San Juan during
late Pueblo Ill. Driver (2002) points out that the decline in artiodactyls is "most obvious"
in highly circumscribed areas. Driver (2002:160) concludes by arguing that this scarcity
led deer to become a more highly valued prey, thus prompting individuals or groups to
garner control over artiodactyl resources. This same activity (controlling access to
scarce resources) has been argued to be evidence of conservation, such that as
population rises and resource availability falls, leaders or groups consciously decide to
limit resource access (Smith and Wishnie 2000). Ethnographic evidence in the Pueblo
Southwest indicates that certain individuals (War Chiefs) had control over when deer
hunts would take place and over the duration of the hunt (Anell 1969; Lange 1959;
Parsons 1918; White 1932). Therefore, it is certainly possible that village leaders could
have exercised the same privilege. It is unclear whether control over access to
resources is a conscious act of conservation or if conservation is simply a byproduct of
resource control. Regardless, it is clear that as the population rose, availability of
artiodactyls declined. Some have argued the inverse of this argument. Speth and
Scott (1 989) argue that reliance on large game increases as a population becomes more
aggregated. They note several sites in the greater American Southwest that show an
increase in artiodactyls and decrease in Lagomorphs as villages begin to grow and
socioeconomic conditions change. In these cases, large villages with large numbers of
people will better be able to organize communal deer hunts, even if game has become
further removed in space. While this is an interesting explanation and may account for
initial increases of large game in villages during initial population growth, it does not
explain the dramatic decrease in large game that is seen across the northern San Juan
area as human populations reach their peak, nor the high frequency of cottontails in
relation to other taxa. The inhabitants of the northern San Juan were not able to
intensify communal artiodactyl hunting activities, and thus, seem to have taken another

approach to intensification of meat procurement by increasing the production of
domesticated turkey and by focusing more on local hunting (Driver 2002; Neusius 1996;
Stratton 1999).
Lagomorphs through Time
Lagomorph frequencies vary quite a bit over time at Shields Pueblo. Among
artiodactyls and turkeyllarge bird, lagomorphs in general consistently dominate as a prey
choice. The lagomorph index is consistently high, indicating that among all of the
lagomorphs, Sylvilagus is far more predominant than Lepus. The highest lagomorph
index is found in sub period 3 when it reaches 0.9. Sub period 6 (mid Pueblo II) sees a
substantial decrease in the lagomorph index (0.76) indicating a drop in Sylvilagus and a
rise in hunting of Lepus. It is also during this time period that the relative frequency of
lagomorphs drops to its lowest percentage (23.2%). During the later half of Pueblo II
(sub period 7) lagomorphs increase in frequency to 67% of the total faunal assemblage.
The lagomorph index also begins to rebound (at 0.82), indicating a rise in cottontails
over other lagomorphs. This trend continues through early and late Pueblo Ill (sub
periods 8 and 9), rising slightly until depopulation.
There is an interesting trend in taxonomic indices. As the artiodactyl index
increases, the ratio of Sylvilagus to all other lagomorphs decreases, and as artiodactyls
begin to move downward again cottontails become more and more dominant over
jackrabbits. This could be a product of localized environmental change or a change in
access to productive hunting grounds. When the artiodactyl index increases the number
of Lepus in relation to Sylvilagus also increases. Both antelope and jackrabbits are
common in more open environments, while deer and cottontails prefer a more brushy
environment (Szuter and Bayham 1989). Therefore, this change in artiodactyl and
lagomorph indices may indicate a change from an open environment to one that is more

conducive to cottontails. This pattern could also reflect changes in access to "wild"
hunting grounds. As communities grow and become increasingly circumscribed on the
landscape, their resource catchment areas become highly bounded (effectively
decreasing catchment area) (Varien 2002). Once preferred game is depleted in a local
area, hunters can either exploit land away from settlement crowding, or they can
intensify resource activities locally (by focusing on local, lower-ranked prey, or by
increasing domestic turkey production). Based upon the increase in Sylvilagus sp. and
domestic turkey (discussed below) the latter seems to explain the faunal pattern. Thus,
as human populations increase: 1) more land is cleared (which results in the creation of
habitat favorable to Sylvilagus), and 2) preferred game species become over-hunted
(which results in an increasing reliance on the more locally abundant Sylvilagus).
Turkey and Large Bird through Time
Turkeyllarge bird frequencies do not appear to be patterned in relation to
chronology. Most problematic are mid Pueblo II (A.D. 1020-1060), because they are far
more frequent than expected; and late Pueblo II (A.D. 1060-1 l5O), which seems to be
lower than expected. The seeming over-abundance of turkey in the first half of Pueblo II
is due to the occurrence of a number of probable turkey burials (of at least 5 individuals)
coming from structure 1308. If the turkey burials are removed from consideration, the
index changes to 0.21, which is only slightly higher than early Pueblo Ill (Figure 14).
There is a noticeable drop in the frequency of turkey in the later half of Pueblo II. Even
with the lower turkey index of 0.21, the late Pueblo II index of 0.13 seems quite low.
There does not seem to be any environmental or population-based explanation for this
disparity. Given the increasing population and decreasing site catchments size, turkey
should be higher in the latter half of Pueblo II. Some other factor (decrease in maize
surplus, disease, etc.) must be responsible for the low index.

Turkey Indices per Sub Period
725-800 1020-1060 1060-1150 1150-1225 1225-1280
Years A.D.
Figure 14. "Corrected" * TurkeyILarge Bird Indices.* Index for A.D. 1020-1 060 refigured by removing turkey burials.
Change in Fauna and Human Population
One of the research questions stated in Chapter 1 asked if change in faunal
patterning over time is a result of change in size of population at Shields Pueblo. In
order to look at this it is necessary to have a proxy for population. There are a number
of methods that have been used in the past to estimate prehistoric populations. These
methods have included calculating the amount of arable land and extrapolating the
number of individuals who could be sustained (Fisher 1936); using number of rooms,
total structure area, and total floor space (Naroll 1962; Drager 1976); and calculating the
number of "households" in a pueblo and extrapolating the population using ethnographic
data on average family size of Pueblo Indians (Hayes 1981). In the case of Shields
Pueblo, the lack of surface architecture makes this difficult, thus another method must

be employed. One method that has been developed to roughly estimate population is to
use pottery weights for dated contexts (Ortman et a/. 2000). This method relies on the
assumption that the rate of pottery deposition is constant. To use this method, first the
pottery weights must be "corrected" by establishing the rate of deposition (i-e., how
much pottery is deposited per year). This is done by dividing the total pottery weight (in
grams) by the number of years in the sub-phase, thus providing the grams per year
deposition rate. Next, the area excavated (m2) is considered by dividing the grams per
year by the total area excavated associated with a given sub period, which allows
analysis of both temporal as well as spatial patterning (Table 29). The idea behind this
procedure is that the rate of pottery disposal is a constant. Therefore, if the rate of
deposition of pottery for time period A is lower than the rate of deposition for time period
6, then the population should be lower in time period A. This obviously does not give an
exact population estimate but simply a guide against which to compare faunal
frequencies. In fact there are several problems with estimating population using this
method. First, it assumes that there is a constant weight per pot. Second, it assumes a
constant rate of breakage. These assumptions become problems if there are any
changes to ceramic technology. For example, if the size of vessels changes or if thinner
higher fired pottery becomes prevalent, these changes would affect pottery weight.
Thus, it is stressed here that pottery weight is being used only as a rouah means of
estimating change in human population over time.
Before looking at change in specific taxa in relation to pottery weights, it is
necessary to check if the frequencies of all faunal categories follow the pattern of
ceramic weight frequencies over time (Table 30). This is done by calculating the rate of
deposition of faunal remains, using the same method as above with frequency (NISP)
instead of weight frequencylyrlm2. Figure 15 illustrates the changes in frequencies for
both ceramics and fauna for each time period.

Table 29. Rate of Pottery Deposition: Shields Pueblo
Table 30. Rate of Faunal Deposition (NISP): Shields Pueblo
Change in Relative Deposition Rates Over Time
Sub Periods (A.D.)
Figure 15. Change in Relative Depositional Rates of Ceramics and Fauna Over Time (by Sub Period). (P= Rate of deposition of pottery [g/yr/m2 values on the left], F= Rate of deposition of faunal remains [ ~ l ~ ~ / y r / r n ~ values on the right])

Figure 15 illustrates an interesting pattern. Population, represented by the
weighted rate of deposition (the solid line), declines slightly from Early Pueblo 1 (A.D.
725-800) when the site is resettled in Mid Pueblo II (A.D. 1020-1060), increases slightly
through Late Pueblo II (A.D. 1060-1 150), but begins to rise significantly during the Late
Pueblo IIIEarly Pueblo Ill transition (A.D. 1150-1225), and is followed by a rapid increase
in population during Late Pueblo Ill (A.D. 1225-1280). The relative rate of faunal
deposition (represented by the dashed line) has an inverse relationship to population
from Early Pueblo I through Early Pueblo Ill. As population falls from Early Pueblo I to
Mid Pueblo II, faunal deposition rates rise and the inverse is true as population rises
between Late Pueblo II to early Pueblo Ill. As population spikes in Late Pueblo Ill, the
faunal deposition rate begins to rise again.
Taxonomic Indices and Pottery Deposition Rates Over Time
Sub Periods (A.D.)
Fiaure 16. S~ecific Taxonomic Indices and Pottery Deposition Rates Over Time. - 9
'(Pottery deiosition rates g/yrlm2 on left; taxonomic indices on right. Raw data can be found in Table 25 (for ART, LAG, and MEG), Corrected Turkey Indices on p.121 and Figure 14, and Pottery Deposition Rates in Table 29)

If each major faunal index is considered in reference to change in population
over time, some interesting patterns emerge (Figure 16). Among the taxa, artiodactyls
have the most interesting relationship to population. While the population falls from
Pueblo I to Mid Pueblo II, the artiodactyls index is at its highest. However, the
artiodactyl index declines at a steady rate as population begins to rebound. As the
population begins to climb significantly from Early to Late Pueblo Ill, artiodactyls fall to
their lowest index.
If the lagomorph index is considered, a more parallel correlation occurs. While
lagomorphs remain high throughout, the "dips" in the index over time follow the same
basic curve as population. The Turkey index seems to rise rapidly as human population
increases from late Pueblo II to Early Pueblo Ill. From Early Pueblo Ill to Late Pueblo Ill
(during the height of population) the turkey index falls slightly, as the lagomorph index
increases again.
Spatial Patterning of Faunal Remains
As stated previously, Shields Pueblo consists primarily of subsurface kiva
structures. Only a partial room block in architectural block 100 remains of the surface
architecture. Even with the absence of surface architecture, there should still be some
spatial patterning to faunal deposition based upon the sub-period in which an
architectural block was occupied and based upon structure and non-structure contexts.
Other spatial analysis that has been done in the region has focused on species
patterning between different architectural blocks and between structural and
nonstructural contexts (Muir 1999). Muir found that artiodactyls tended to be
concentrated in kivas and on the roofs of towers at Sand Canyon Pueblo indicating the
location of processing, storage, and perhaps redistribution (1999:154). The purpose of
looking for spatial patterning at Shields Pueblo is to see if artiodactyls follow the same

pattern as Sand Canyon Pueblo. If so, then it may be possible to identify processing or
storage areas among the Shields Pueblo kivas. Table 31 shows the frequency of major
taxa by excavation block (also illustrated in Figure 17). The 100, 200 and 1300 blocks
have the highest numbers of excavated structures and non-structures and the highest
frequencies of the selected taxa. These blocks also have the longest periods of
occupation. The 100 block has Pueblo I deposits, as well as late Pueblo II and early
Pueblo Ill. Structures 110 and 141 contain the early Pueblo I deposits. The majority of
Sylvilagus from the dated sub assemblage comes from Structure 110. The remainder of
the excavated structures are subterranean pit structures. Block 200 contains late

Tab
le 3
1. F
requ
ency
of S
elec
ted
Tax
a fro
m e
ach
Exc
avat
ion
Blo
ck.
SY
L M
EG
35
9 33
5 42
.8%
'4
0.0
%
176
68
68.5
%
26.5
%
740
624
38.5
%
32.4
%
510
841
34.9
%
57.5
%
12
7 46
.2%
26
.9%


Frequency of Taxa Per Block Area
8100 B200 8400 8800 81100 81200 81300 81400 81500
Excavation Blocks
Figure 17. Frequency of Taxa Per Excavation Block
l ART El LEP
SYL
{OMEG
Pueblo II through Pueblo Ill deposits. Structures 222 and 234 contain the largest
amount of lagomorph remains for the entire site. Block 1300 has the longest continuous
occupation, containing deposits from each sub period (from both structures and non-
structures). The high frequency of faunal remains found in blocks 100 and 200 can be
explained by the fact that both have the largest number of excavated structures. Block
100 also has the only surface structures and consequently is probably better preserved.
Both of these architectural blocks are located along the bottom of the "horseshoe" of the
pueblo. This portion has the earliest occupations and the most consecutive occupation.
Table 32 presents the standardized residual values for each of the major
taxonomic groups by excavation block. Sylvilagus and turkeyllarge bird display the
greatest variability in frequencies, represented by the absolute sum of standardized
residuals. Turkeyllarge birds are far more abundant in excavation block 400 and 1400

(dating from late in Pueblo Ill), and far more scarce in block 100 (with earlier dates).
Sylvilagus is abundant in block 100 and far less common in blocks 1 100,1300,and 1400.
Artiodactyls also show a high degree of variation. They are far more abundant in block
1300 and far scarcer in blocks 200, 1100, and 1400. Artiodactyls are probably more
abundant in block 1300 because this block had dense trash-filled Pueblo I and II
pitstructures, while blocks 200, 1100, and 1400 date primarily from Pueblo Ill (Duff,
personal communication). If excavation blocks are considered, block 1400 and 1300
display the greatest amount of variation. Block 1400 has a total absolute residual of
43.1. All of the taxa in this block have standardized values beyond one standardized
deviation of the mean standardized residual (indicated in bold in Table 32). Turkeyllarge
bird is the only taxon that occurs far more frequently than expected. Sylvilagus, Lepus,
and artiodactyls all occur far less frequently in this block. Block 1300 also shows
significant deviation from expected frequencies. Artiodactyls are much more abundant,
while Sylvilagus are far less abundant. Blocks 400, 800, 1200, and 1500 display the
least amount of variation among blocks and do not have significantly higher or lower
taxon frequencies.
Because the vast majority of site is subsurface, little can be said about ritual
versus domestic disposal. However, it is possible to consider structure versus non-
structure contexts (Table 33 and Figurel8). Table 33 and Figure 18 illustrate an
interesting pattern. The Chi-square value of 20.0 indicates a confidence level between
98% and 99%, with a significance probability just over 0.01. Overall, the selected taxa
as well as ceramics are found far more frequently in the non-structural context. This is
similar to deposition patterns at Sand Canyon Pueblo (Muir 1999:144). The
artiodactyls are strongly associated with non-structural contexts. The lagomorphs,

Table 33. Relative Frequency (NISP) of Selected Taxa and Pottery Weights from Structures and Non-structures.
Pearson Chi-square= 20.0; d.f.=8; phi-square=2.0; Cramer's V= 1 .O; P ~ 0 . 0 1
Frequency of Taxa and Pottery Weights: Structure Vs. Non-Structure
Structure NonStructure
Figure 18. Frequency of Taxa and Pottery Weights: Structure vs. Non-structure.
NSYL MEG
- --
turkeyllarge bird, and pottery are tightly correlated in frequencies. Even if the possible
turkey burials in Structure 1308 are removed from consideration, they are still found in
non-structural contexts at a frequency of 70.3%.

Chapter Summary
The purpose of this chapter has been to present the Shields Pueblo faunal
assemblage so that various patterns (in preservation, frequencies over time, and in
relation to population) can be recognized among the fauna. Overall, Shields Pueblo
follows patterns similar to other sites in the region (Driver 2002). The faunal assemblage
is heavily weighted towards certain species, namely lagomorphs, domestic turkey, and
artiodactyls. While other species of fauna are present, this "trinity" of economically
important species are by far the most numerous species present within the assemblage.
Additionally, it is important to establish the extent to which the Shields Pueblo
assemblage was affected by taphonomic forces. This was done by measuring and
comparing the effects of both natural and cultural activities upon faunal remains. Shields
Pueblo's faunal assemblage shows very little carnivore damage. The most frequently
chewed species is turkey and only 2.7% of these are modified by canids. By far the
most frequently culturally modified species at Shields Pueblo are the artiodactyls with
nearly 20% modification. Turkey is the next most commonly modified at 10%. Some 8%
of Lepus are modified and 5% of Sylvilagus are modified. Most of the modified rabbit
bone is burned. All economically important species are highly fragmented (broken), thus
suggesting a high level of faunal processing (indicative of resource intensification).
Preservation of the faunal assemblage was further analyzed by calculating
skeletal part frequencies. The basic pattern is that denser bone is better preserved than
less dense bone. While nearly all skeletal units are represented among the lagomorphs
and turkey, the majority of artiodactyl elements came from the forelimb, pectoral girdle,
and hindlimb, supporting the notion that deer and other ungulates were being brought
into the site from some distance. However, these elements are also more dense, thus,
preservational bias cannot be ruled out.

Age at death estimates show an interesting pattern. When the later bone fusion
stages are considered, over 50% of artiodactyls and 30% of lagomorphs are killed
before they reach maturity. This likely mirrors the demographics of the hunted
population of rabbits and artiodactyls. The overall turkey assemblage is made up of
approximately 40% immature birds. When fusionlage categories are considered, very
young turkey seem to be killed preferentially (30%) over late juveniles (20%). This
could indicate culling for flock expansion. Further analysis in establishing sex and tighter
age categories would have to be done to investigate this argument.
Change in frequency and species indices over time, again showed a pattern that
is typical of the Northern San Juan. Artiodactyls are more common in earlier
occupations (with an artiodactyl index of .06 in sub period 3 and 0.2 in sub period 6) and
then are replaced by increasing numbers of Sylvilagus and turkey (evidenced by the
dropping artiodactyl index 0.07 in sub period 7, 0.03 in sub period 8, and 0.03 in sub
period 9) as populations expand at the site and in the general region. Sylvilagus is by
far the most common of rabbit genera with high indices throughout the occupation of the
site. The ceramic deposition rates seem to indicate that there was also a drastic rise in
population during Pueblo Ill at Shields Pueblo. At this time the artiodactyl index is at
0.03, the lagomorph index at 0.89, and turkey at 0.33. So population seems to affect the
availability of artiodactyls, causing increased reliance on Sylvilagus and turkey.
While the Shields Pueblo fauna followed the basic trend of deposition outside of
structures, there were no strong correlations between artiodactyls and kivas. In fact, the
reverse was true as some 80% of artiodactyl remains were recovered from nonstructure
contexts. This could possibly be explained by ethnographic accounts of disposal of
artiodactyls. Once remains had been displayed on ritual structures for a period of time,
they were removed and deposited in the trash middens (White 1974b). This might also

be indicated by the higher frequency of weathering among artiodactyl remains illustrated
in Table 17.
Some excavation blocks show significantly higher frequencies of particular taxa.
Block 100 has a significant abundance of Sylvilagus, block 1300 of artiodactyls, and
block 1400 of turkeyllarge bird. Certain species (Lepus, in this instance) were far less
variable than other species with a low sum of absolute standard deviations. The lack of
a correlation between artiodactyls and specialized structures (as noted by Muir 1999 in
reference to towers) can be explained by the lack of preservation of surface structures.
Connections between the spatial patterning of faunal deposits and gender will be
discussed in Chapter 6.
Overall, Shields Pueblo follows patterns typical of other sites in the region.
Chapter 5 furthers this discussion by outlining the greater regional faunal patterns and
how Shields Pueblo fits within these parameters.

CHAPTER 5
SHIELDS PUEBLO IN A REGIONAL CONTEXT
Shields Pueblo is important within the Monument/McElmo drainage region for its
long period of occupation. While there are Pueblo I and II sites within the immediate
region, most archaeological sites date from Pueblo Ill. In order to fully understand the
faunal pattern at Shields Pueblo, it must be put into context with sites in the greater
northern San Juan region. This chapter will put Shields Pueblo in a regional context by
comparing the relative frequencies and indices of selected species with those from sites
in the immediate region as well as the greater Northern San Juan region. This chapter
will conclude with a discussion positing explanations for these trends.
Changes in Faunal Patterning at a Regional Scale
Chapter 3 defined the Monument/McElmo drainage region as the area bounded
in the south by McElmo Creek, running northwest to the reaches of Monument Creek at
the Utah border (refer to Figures 2 and 3 in Chapter 3, and Adams and Petersen [1999]).
This is the immediate region around Shields Pueblo and has many sites, both small and
large, that can be used for comparative purposes. Large village sites include: Sand
Canyon Pueblo, Castle Rock Pueblo, Woods Canyon Pueblo, and Yellow Jacket Pueblo.
There are numerous smaller sites in the Sand Canyon locality. Most of the sites found in
this region are Pueblo Ill sites (many, specifically late Pueblo Ill). However, there are
some sites with older components and these are used for comparison with Shields
Pueblo for the Pueblo I and II sub-assemblages.
The greater northern San Juan region lies north of the San Juan River (in
Northwestern New Mexico). It includes land as far east as the Animas River and runs as
far west as Cottonwood Wash (in Utah). The northern San Juan Region is bounded in

the north by the La Plata Mountains and the northern reaches of the Dolores River (refer
to Figure 2 in Chapter 3).
For this chapter, only three taxonomic groups have been chosen for
consideration: artiodactyls, lagomorphs (including cottontails and jackrabbits), and
turkey (including turkey and elements identified as large bird). These particular
taxonomic groups were chosen because they are by far the most common and
economically important species in faunal assemblages from Pueblo I through Pueblo Ill.
Other species of fauna are represented in the Shields Pueblo assemblage, but many
occur only in small numbers. Other research has included carnivore frequencies as well
as the above species in order to look at human exploitation of the "wild" versus
"domestic" environment (Driver 2002) or to further explore inter-site variation of ritually
important species (Muir 1999). Because most of the carnivores occurring at Shields
Pueblo are canids (which are not necessarily wild), and because all other carnivore
species occur at such low frequencies they were not included in this analysis.
Pueblo I Sites (A. D. 750-900)
Pueblo I sites in the Monument/McElmo drainage region are rare. Of the sites
with published faunal assemblage data, only two have primarily Pueblo I components.
The Duckfoot Site is the larger of the two sites in the McElmo Dome region. This site
was a small 19-room hamlet with 4 pit structures (Lightfoot and Etzkorn 1993). Duckfoot
was excavated by Crow Canyon Archaeological Center. It is the most completely
excavated Pueblo I site on the McElmo Dome. Occupation dates between A.D. 850 and
A.D. 880 (Lightfoot and Etzkorn 1993:l). In addition to the Duckfoot Site, there is a
small Pueblo I site in Yellow Jacket Canyon (5MT 8838) (Akins 1988). Table 34 gives
the relative frequencies of artiodactyls, lagomorphs, and turkey for the Duckfoot Site,
5MT8838, and Shields Pueblo in the immediate region and several sites from the

Dolores River area from the greater northern San Juan region. The Dolores River region
seems to have been the focus of settlement during Pueblo I, as it contains the majority
of occupation sites in the Northern San Juan. Figure 19 illustrates the relationships
between taxonomic frequencies at each of the Pueblo I sites. Overall, there does not
appear to be any patterning of sites (i.e., none of the sites located in different
environments cluster together). The only characteristic shared between these sites is
the generally low frequency of turkey. The relationship between sites is discussed in
greater detail below.
Table 34. Northern San Juan Region Pueblo I sites: Relative Frequencies and Indices
Dolores
(Akins 1988; Applegarth and Feldman 1981 ; Fetterman and Honeycutt 1982; Neusius l986a, l986b,l986c, l986d; Neusius and Gould 1988; Walker 1993: Table 8.1 :24O).
5MT8838
Shields Pueblo
5MT4007
Grass Mesa Hamlet
Windy Wheat Hamlet
Permian Hamlet
6.8%
5.2%
26.4%
45.1 %
20.6%
22.7%
88.1%
76.6%
70.4%
53.7%
78.6%
74.8%
5.1%
18.2%
3.2%
1.2%
0.7%
2.5%
.07
.06
.27
.47
.21
.23
.65
.90
.44
.58
.49
.74
.05
.19
.04
.02
.O1
.03

Figure 19. Taxonomic Frequencies: Pueblo I, Northern San Juan Region. (Arrow indicates Shields Pueblo) *Note that Lagomorph frequencies are read along the horizontal gridline, Turkey along the right-hand diagonal gridline and Artiodactyls along the bottom diagonal gridline.
Table 34 illustrates that Shields Pueblo has the lowest artiodactyl index of all of
the northern San Juan region sites, the highest lagomorph index, and the highest turkey
index during Pueblo I. Sites within the McElmo drainage during Pueblo I do not appear
to be tightly correlated. In fact, in Figure 19, Shields Pueblo appears to be somewhat of
an outlier in comparison to Duckfoot and Yellow Jacket Pueblo. This is primarily due to
Shields Pueblo's low artiodactyl index, the high lagomorph index, and most notably the
high turkey index in comparison to the other sites. During Pueblo I, Shields Pueblo was
small in size. Only two structures were identified and dated to Pueblo I (Structures 110

and 141). Structure I 10 is a subterranean pit structure dated to about A.D. 770 and
Structure 141 is a rectangular subterranean room dating between A.D 600 to 950 (Duff
and Ryan 1999). No other structures at Shields date this early. In contrast to the
larger Duckfoot Site and Dolores area sites, Shields seems to have been a small farming
hamlet. The low frequency of artiodactyls during Pueblo I is not what is expected based
upon frequencies from other sites. The region was not heavily populated and villages
were concentrated in the Alkali Ridge area in the far western portion of the
Montezuma/McElmo Drainage, to the southeast in the Animas Drainage and in the
Dolores River area (Wilshusen 1999:Figure 7.5:211). The lack of artiodactyls in later
periods has been argued to be a function of over-hunting of game by large, spatially
circumscribed populations (Driver 2002). Given that most Pueblo I sites in the northern
San Juan were likely hamlets and farmsteads, over-hunting is not a plausible
explanation. Instead, difference in artiodactyl frequencies between assemblages may
depend upon the size of local populations and their ability to procure large amounts of
big game. If populations were low, communal hunting may not have occurred, or may
not have yielded as many deer or other large game as hunts performed by larger
communities. Another explanation is that there simply are no good Pueblo I trash
deposits at Shields Pueblo due to destruction and mixing of archaeological deposits.
This lack of deposits would certainly justify low artiodactyl frequencies.
Typically, Pueblo I faunal assemblages are dominated by Lagomorphs, although
half of the site assemblages in the region contain more than 20% artiodactyls remains
(Driver 2002:154). According to Driver (2002: 155, Figure 7.5), the Lagomorph index
tends to indicate a focus on cottontail with close to 50% of sites having a majority of
cottontail, although there is some variation depending on site location.
The basic faunal pattern of Pueblo I sites in the northern San Juan is notable in
the apparent lack of turkey and large birds in the assemblages. During this period,

turkey was not a dietary staple, and was primarily used for religious purposes (Munro
1994). Most sites in the area have very low counts of turkey (some have none at all).
Only a few sites have notable amounts of turkey in their assemblages during this period
(including Shields Pueblo), and these could be present in larger quantities because they
represent turkey burials and not domestic meal refuse. However, no discrete burials
were identified for Pueblo I contexts. Therefore it is not possible to "correct" the turkey
frequency for this time period by removing turkey burials.
Pueblo I1 Sites (A.D. 900-1 150)
There are few excavated Pueblo II sites in the MonumentlMcElmo region. These
sites include Yellow Jacket Pueblo, three sites in the Yellow Jacket Locality (5MT8827,
5MT8839, and 5MT8371), another site in the McElmo Dome region (5MT1786), and a
series of sites in the greater region. While sites dating to the Pueblo I period tend to be
located in the Dolores region, they begin to spread southward and westward during
Pueblo II (Varien 2002). Table 35 provides the relative frequencies for the artiodactyls,
lagomorphs, and turkey. Pueblo II taxonomic frequencies are fairly widely scattered
across the ternary plot (Figure 20). There does not seem to be much grouping of sites in
relation to their frequencies. Some sites, such as 5MT8371, have a relatively high
frequency of artiodactyls especially in comparison to the lagomorph index, and a high
turkey index. So, individuals at this site seem to have focused on artiodactyls and turkey
and did not utilize rabbits to the same extent as others. On the opposite side of the
spectrum, the assemblage from 5MT1786 (Gnatsville) site, located reasonably close to
Shields Pueblo, has no Artiodactyla whatsoever; instead it contains mostly rabbit, and
some turkey. Shields Pueblo lies in between these two extremes.

Table 35. Northern San Juan Region Pueblo II Sites: Relative Frequencies and Indices
LA65029 LA65030
(Akins 1988; Anderson 1966; Bertram 1991 ; Driver 1996; Emslie 1978; Hayes 1984; Kent 1989a; Mick O'Hara 1994; Muir and Driver 2003; Neusius 1986c; Nielson et al. 1985; Nordby 1973; Rhone 1971 ; Thompson 1985)
55.4% 14.8%
27.7% 56.2%
16.9% 29.0%
.67
.21 .28 .50
.38
.34

Figure 20. Taxonomic Frequencies: Pueblo II, Northern San Juan. (Shields Pueblo indicated by arrow). *Note that Lagomorph frequencies are read along the horizontal gridline, Turkey along the right-hand diagonal gridline and Artiodactyls along the bottom diagonal gridline.
One might argue that the taxonomic frequencies at 5MT8371 support Speth and
Scott's (1 989) assertion that rabbits are the preferred game and artiodactyls become the
predominant focus of hunting activities when populations reach environmental carrying
capacity and people must exert more energy in communal hunting for artiodactyls farther
away from settlements. This idea is discussed in further
detail below in relation to Pueblo Ill faunal patterns. If this argument is true, then one
would expect 5MT8371 to be a large site with an equally large prehistoric population.

This is not the case. 5MT8371 is described by Dykeman (1986) as an isolated pit
structure, likely a farmstead.
It is also unclear if environmental differences are responsible for all of the
variations in taxonomic indices. All of these sites lie at around 2000 meters in elevation.
Likewise most sites on the McElmo Dome are situated in the Pinyon-Juniper woodland
mixed with sagebrush- saltbrush today. This environment is likely similar to the
paleoenvironment extant during Pueblo II (Adams and Petersen 1999) and would have
provided access to both large and small game animals (see Table 3 in Chapter 3 for
specific species). Some other factor must, therefore, be causing the difference seen in
taxonomic usage among the site assemblages. However, it is clear that the low
lagomorph indices in the La Plata region are based on environmental differences and
lower elevations. This region is characterized by open grassland, thus more attractive to
Lepus than Sylvilagus (Adams and Petersen 1999; Szuter and Bayham 1989).
Pueblo II tends to have a widely variable faunal pattern (Driver 2002: 153, Figure
7.3). However, there is a basic trend in an overall increase in Turkey usage. Pueblo II
saw the beginning of the use of turkey as a food source (Munro 1994). The high level of
variability in relative frequencies of artiodactyls, lagomorphs, and turkey is probably due
to local site factors, such as location on the landscape or site catchment size, and also
to social factors such as community size and the ability to organize labor for communal
hunting.
Pueblo 111 Sites (A. D. 1 150 - 1300)
Pueblo Ill sites are quite numerous in the Monument/McElmo region. There are
several larger sites ( Woods Canyon Pueblo, Castle Rock Pueblo, Yellow Jacket Pueblo,
and Sand Canyon Pueblo) and many smaller sites in the Sand Canyon Locality (Mad
Dog Tower, Saddlehorn Hamlet, Shorlene's Site, Roy's Ruin, Lillian's Site, Troy's Tower,

Catherine's Site, Kenzie Dawn Hamlet, Lester's Site, Lookout House, Stanton's Site, and
G and G Hamlet). Because of the large number of smaller sites, these have been
combined to establish mean frequencies and indices for the Sand Canyon Locality.
Table 36 shows the relative frequencies of the selected taxa from Pueblo Ill
assemblages in the McElmo Dome region and Figure 21 illustrates the relationship
between taxonomic frequencies and indices for each site.
Table 36. Northern San Juan Pueblo Ill Sites: Relative Frequencies and Indices.
(Anderson 1966; Bertram 1991 ; Driver 1996; Driver, et al. 1999: Table 18.1 ; Mick O'Hara 1994; Muir and Driver 2003: Table 12; Nickens 1981 ; Rohn 1971 ; Shelley 1993; Thompson 1990; White 1977)

Figure 21. Taxonomic Frequencies: Pueblo Ill, Northern San Juan (Shields Pueblo indicated by arrow). *Note that Lagomorph frequencies are read along the horizontal gridline, Turkey along the right-hand diagonal gridline and Artiodactyls along the bottom diagonal gridline.
During Pueblo Ill faunal assemblages in the McElmo dome seem to cluster with
moderate frequencies of lagomorphs and turkeys and low frequencies of artiodactyls.
Most sites show a distinctive lack of artiodactyl remains, but differ in the relationship
between lagomorphs and turkey. For this time period many of the sites (most notably,
Woods Canyon Pueblo and Castle Rock Pueblo) show an increased reliance on turkey
in relation to lagomorphs. Cottontails are by far the most common of lagomorphs found
in the McElmo Dome site assemblages during Pueblo Ill. This does not follow the same
pattern described in other regions of the Southwest. Szuter and Bayham (1989) claim
that cottontails should be found in small sites and less populated regions, while

jackrabbits should be found in increasingly deforested or cleared environments around
larger, more densely populated villages. The McElmo dome was certainly densely
populated during Pueblo Ill. Wilshusen (2002 :Table 5.6) estimates that there were
between 22.6 to 28.9 persons per square km. in the Mesa Verde Region. This high
density of settlement causes site catchment areas around settlements to decrease in
size, become more bounded by other community catchments, and to become
increasingly overlapping (Varien etal. 2000, Varien 2002). It is clear that given the
increasing aggregation of settlements in the McElmo Dome, that deforestation and land
clearing were factors in environmental change (Kohler 1992, Kohler and Matthews
1988). However, instead of a decrease in cottontail and increase in jackrabbits, the
reverse is true, thus indicating that land clearing did not result in creating grassy
environments, but scrubby environments.
Pueblo Ill is by far the most significant period in terms of cultural and economical
change in the northern San Juan. It is a period of high population densities and
community aggregation. Because of these factors, the faunal record indicates some
major changes to hunting and domestic animal usage. Over-all turkey usage remains
high, however, there are several sites where turkey is the predominant protein source,
such as Roundtree Pueblo, site 499, and Knobby Knee Pueblo (Anderson 1966, Bertram
1991). The frequency of artiodactyls in the faunal assemblages decreases substantially
in most locations. Over 70% of sites in the northern San Juan region have between a
1 % and 10% artiodactyl frequency, and several have no artiodactyl remains at all (Driver
2002: 153-155). Lagomorphs are by far the most dominant taxonomic family during this
time period. Cottontail in particular, is dominant as evidenced by the change to higher
lagomorph indices for northern San Juan sites. This is by far the most interesting
finding, as it does not follow the pattern of lagomorph indices in other parts of the greater
Southwestern region (Szuter and Bayhan 1989; Quirt-Booth and Cruz-Uribe 1996).

There is also some indication of geographic patterning of faunal frequencies.
The majority of La Plata and McElmoICortez sites cluster together tightly (refer to Figure
21, cluster at arrow) with low artiodactyl frequencies, and moderate to high lagomorph
and turkey frequencies. Mesa Verde and Mancos sites tend to have lower frequencies
of lagomorphs, with moderate frequencies of artiodactyls and turkey. These clusters of
sites may indicate similarities in environment. For example, Mesa Verde is primarily
Gambel Oak scrubland on the mesa top with Pinyon-Juniper woodland in lower
elevations, while Mancos is primarily Pinyon-Juniper woodland (both productive
environments for artiodactyls) (Adams and Petersen 1999: 15-1 7). However, La Plata
and McElmoI Cortez sites also tend to cluster together (especially in terms of low
artiodactyl frequencies). While the La Plata sites and the Cortez site are located in
Sagebrush-Saltbrush biotic communities, the McElmo region sites are found in a variety
of environments (such as sagebrush/satlbrush on mesa tops, and Pinyon-Juniper in
canyons, talus slopes, and drainages). In the latter region, the low frequency of
artiodactyls in an environment that is typically attractive to artiodactyls, is likely caused
by other factors such as over-hunting and population pressure (discussed below).
Discussion
While there are points of variation, in general, Shields Pueblo follows the same
basic faunal pattern seen throughout the northern San Juan region. Artiodactyls decline
substantially in Pueblo Ill, lagomorphs are by far the most frequent taxon during each of
the time periods, and turkey becomes increasingly common as populations in the region
grow and hunting declines. Various arguments have been made concerning population
size and game preference. Speth and Scott (1989) argue that large game becomes
more important while small game (such as lagomorphs) declines in importance as
human population numbers rise and resources become increasingly scarce. The idea is

that communal hunting is economically effective at harvesting a large amount of meat
when groups are forced to hunt a great distance away from the settlement, the caveat
being that this requires more labor as well as risk. Therefore, local populations must be
high enough and resources scarce enough to make communal hunting worthwhile. The
situation described by Speth and Scott (for the Chaco area) does not fit with the
demographic, environmental, and faunal data for Pueblo II in the northern San Juan
region (as a whole) nor the immediate McElmo Dome. While the population in the
McElmo dome expanded during Pueblo II (from 4.9 people/km2 in early Pueblo II to
about 7 people/km2), it did not reach the levels seen in Pueblo Ill (nearly 15 people/km2
to as many as 22 people/km2) (Mahoney et al. 2000:Figures 2 and 3: 74-75). Likewise,
site catchment areas were fairly widely spread with no overlap between communities
(Varien et a/. 2000). And yet, many site sub-assemblages dating to this period display
their highest artiodactyl frequencies. Therefore it does not seem as if resource stress
was a factor.
The McElmo region seems to have been depopulated (or at least very sparsely
settled) during mid to late Pueblo I. When this area was repopulated in Pueblo II, there
would have been a period of time for game to rebound locally. There is no drop in
artiodactyl counts until populations rise to their maximum levels during Pueblo Ill.
Speth and Scott's (1989) hypothesis is completely counterintuitive to what is
evident in the archaeological record for this region. While there is truth to the argument
that communal hunting requires a certain size population and level of labor organization,
it is not clear why Speth and Scott argue that this is a negative or costly endeavor.
Hunting with larger numbers of people, while certainly requiring more planning and
organization beforehand, does not seem to require any more energy per person than a
typical solitary or small scale hunt as many of the same hunting tactics are used.
Instead, it is probably a riskier enterprise than single hunts in terms of success rates.

However, any risks can be underwritten by communal surpluses given larger group
structures (such as lineages or clans). Therefore, the dramatic decline of artiodactyls in
Pueblo Ill assemblages supports the classic population and environmental models, in
that they do decline in areas of high human population densities and that they tend to
decline through out the region.
Late Pueblo Ill is argued to have been a period of resource depression. Several
studies have looked at landscape changes, such as deforestation and changes to the
arable landscape (Ahlstrom et a/. 1995; Huckleberry and Billman 1998; Kohler and
Matthews 1988; Szuter and Gillespie 1994). Deforestation has been shown to have
caused changes to hydrology regimes, soil, and disruption to large game
populations/migrations and to human hunting strategies (Kohler and Matthews
l988:54O; Szuter and Gillespie 1994). Decreases in the artiodactyl resource base could
have been caused by these changes to the landscape as well as by an increase in the
number of people preying upon them. The increase in population combined with greater
aggregation of communities in productive areas creates conditions for over-hunting.
Driver (2002:146) suggests six trends in faunal assemblages due to over-hunting: a) a
decrease in preferred species; b) an increase in species diversity; c) a focus on locally
available species; d) intensification of the production of domestic turkeys; e) intensive
processing of animal remains in order to extract fats and proteins; and finally f) evidence
of trade in preferred meat resources. Given that many of the sites in the northern San
Juan region were crowded into a smaller geographic area during this time period, it is
clear that the deer population would be stressed in these areas. What becomes
problematic is explaining why sites along the periphery of this crowding follow the same
pattern when they would have had larger catchment areas to exploit. In Chapter 1, I
discussed a few different hypotheses that address this problem. Among them, social or
political control and conflict or warfare both seem to answer specific issues in the

northern San Juan region, namely why people hunted within their local catchment area
and did not exploit seemingly more productive environments.
Driver's (1996) argument that larger sites were able to control access to
resources may account for the lack of artiodactyls in small sites around larger
community centers (such as Sand Canyon). These larger community centers certainly
would have been able to influence settlements around them. But what gave these
centers their political power? I would argue that the rising level of violence in the region
(probably due to competition over land and resources) and the old adage of safety in
numbers are the lynch pins to explaining why larger community centers have much
larger frequencies of artiodactyls. Smaller communities simply did not have the numbers
of people to allow a large contingent of warriors to leave on a hunting expedition. This
would have left the community vulnerable to attack. Large community centers, on the
other hand, would have had high enough populations for a contingent to remain at home
for security.
Shields Pueblo, while large, does not have the same frequency of artiodactyls as
other large sites on the McElmo dome. In fact, it is substantially lower and more closely
resembles the smaller sites in artiodactyl frequencies. Shields Pueblo lies in a more
peripheral location (e.g., not as completely bounded as Sand Canyon or Yellow Jacket
Pueblo) (Varien 2002). Given that the pueblo is unbounded to the east and would have
had access to those hunting areas, the inhabitants of Shields Pueblo should have been
able to procure higher number of artiodactyls. Either Shields Pueblo was not as highly
populated as its size suggests, or it was simply too dangerous for hunters to leave in
great numbers for communal artiodactyl hunts. Another explanation is that Goodman
Point Pueblo was the community center during Pueblo Ill and Shields had only a
nominal population. Until Goodman Point is excavated and analyzed, little can be
known of Shields' prominence. Finally, it is possible that the relevant surface structures

were not excavated at Shields Pueblo, given that artiodactyls were more commonly
associated with towers at Sand Canyon Pueblo and Yellow Jacket Pueblo.
Along a similar vein as Speth and Scott's (1989) hypothesis, Szuter and Bayham
(1994) conclude that the lagomorph index should be high in small farmsteads and low in
larger hamlets and villages. The brunt of this argument is that cottontails prefer a more
closed in, heavily vegetated environment that has not been substantially affected
through intensive farming practices. However, if the population grows, resulting in more
land being cleared for farming, the lagomorph index should drop in favor of jackrabbits.
This effect should increase as population densities and occupation spans increase. The
northern San Juan region does not follow this pattern of faunal change. Such as Speth
and Scott's (1989) artiodactyl hypothesis, the reverse pattern is indicated. Instead of
large quantities of jackrabbit overtaking cottontail, cottontail becomes the predominant
rabbit genus in the faunal record.
The problem with Szuter and Bayham's (1994) argument does not lie with the
environmental preference of rabbit species, but with the assumption that farming
practices always result in the creation of open grassland environments. While this
assumption is certainly true of some geographic regions, I suspect that shrubby species
of plants are just as common colonizers of depopulated fields as grasses. In fact, big
sagebrush (Arfemisia tridentate) and rabbitbrush (Chrysothamnus nauseosus) are early
successional plants that typically take over depopulated fields or disturbed environs
(Adams and Bowyer 2002). Shields Pueblo, such as most of the sites in the northern
San Juan, focused primarily on cottontail throughout its occupation. Changes in
population densities over time or any environmental changes brought about by human
activities on the landscape, simply do not affect the lagomorph index. Cottontails were
probably more available around settlements (including Shields Pueblo), and therefore,
more common than jackrabbits.

Turkey indices are interesting in what they could imply about access to hunted
animal resources. For various reasons, hunted game is often considered more
desirable than domesticated animals. In fact, many groups categorize animals
differently based upon whether they are wild or domesticated (Kent 1989). Wild animals
are often anthropomorphized and are believed to be related to humans in some way.
Some groups believe that animals behave in similar ways to humans (take for instance
"Coyote" in the Dine Bahane [a creation myth of the Navajo] who speaks and plays
practical jokes on both humans and other animals [Zolbrod 19881). Other groups such
as the Wari of Brazil believe that wild animals (especially tapirs) are the reincarnated
souls of their ancestors (Conklin 1995). Kent (1989) describes a series of conditions
under which wild and domestic animals are categorized as separate beings. Societies
who keep domestic animals, but do not typically hunt, tend to categorize all animals
together and do not recognize them as intellectual beings. Groups that have no
domestic animals and rely on wild, hunted game nearly always associate animals with
humans as sentient beings. Among cultures that keep domestic animals, but regularly
hunt, wild animals are nearly always categorized as intellectual entities, while domestic
animals are not (Kent 1989b: 13). The Anasazi probably fell in the latter category.
Ethnographic records from Puebloan groups repeatedly describe communal hunting
activities (for both large and small game) as highly ritualized, and even mention that
animals taken during small hunts by single hunters were treated in ritualized ways
(Gnabasik 1981). Once turkey became a primarily domestic animal (and not as ritually
important), it must have lost a certain amount of esteem becoming somewhat "common."
If this is the case, then increased frequencies of turkey would mean that there was less
access to preferred wild meat resources, thus causing people to rely more heavily on a
less preferred species.

Throughout its occupation, Shields Pueblo falls within the mean frequency of
turkey for northern San Juan sites. However, some sites (both large and small) do show
much greater frequencies of turkey (especially during the late Pueblo Ill period) than
Shields Pueblo. These sites are generally located in clusters of highly aggregated
settlements decreasing their catchment areas, effectively limiting their access to large
game. (Access to small game would not be affected to the same degree for the reasons
given above). Communities that were not able to hunt enough game to meet their
protein requirements supplemented their diets with turkey. Again, because Shields
Pueblo lies along the edge of community clusters, inhabitants would have had more
access to wild resources, so that turkey played a less prominent role than what is seen
in other communities.
Chapter Summary
While sites are relatively scarce during Pueblo I for the region as a whole, there
are numerous sites in the Dolores River region to the northeast. Species frequencies
vary greatly between the McElmo and Dolores sites, probably because they lie in
different biotic communities. The McElmo sites have an average artiodactyl frequency of
9.7% (Shields Pueblo's is 5.2%), an average lagomorph frequency of 83% (Shields
Pueblo's is 76.6%), and a turkey frequency of 7.3% (with Shields Pueblo at 18.2%). The
Dolores region sites have a much higher artiodactyl frequency (39.8%), resulting in lower
relative lagomorph frequency (56.7%) and even lower turkey frequency (1.2%). Dolores
lies in a mixture of biotic communities including pine-Douglas-fir, Gambel oak scrubland,
and pinyon juniper. While pinyon-juniper (which makes up the majority of the McElmo
region) is relatively productive for large game, the pine-Douglas-fir and Gambel oak
scrublands are especially rich in large game (Adams and Petersen 1999: 17). Turkey, in
general, is quite low during Pueblo I. It was not a dietary staple and therefore, is not

expected to dominate the assemblages. Shields Pueblo varies most from other sites in
the region with its relatively higher turkey frequency (18.2%) and its relatively high turkey
index (.19). It is unclear why this is the case. It is possible that these early deposits are
contaminated with later ones, or that inhabitants of Shields Pueblo were eating turkey
earlier than other sites in the region.
As expected (based on Driver 2002), Pueblo II has a widely variable faunal
pattern. Artiodactyl frequencies range from 0 to 72%. Lagomorphs and turkey also
follow this highly variable pattern. Some of the variation can be explained by
environment. The La Plata region sites have a low lagomorph index, indicative of a
predominance of jackrabbit rather than cottontail, while all other regions focus on
cottontail. This makes sense given the more open grassland environment that would
have dominated the region. In most cases, variability in frequencies is not so easily
explained and is probably due to catchment size or social factors (such as the ability of a
community to organize labor).
Pueblo Ill, by far, has the most numerous sites. This is especially true of the
McElmo region. The most notable change during this period is the decline in artiodactyls
(with most frequencies ranging from .5% to 16%). Only a few sites retain relatively high
artiodactyl frequencies during this time period. These sites differ due to their locations,
which limit crowding (to a certain extent) and over-hunting (Driver 2002). Shields Pueblo
(such as the majority of sites in the Northern San Juan region) has a low artiodactyl
frequency (1.9%), a high lagomorph frequency (58.2%) and index (.86), and a moderate
frequency of turkey (at 39.9%). Other sites in the region have much higher turkey
frequencies with lower frequencies of lagomorphs. In these instances, it is clear that
population densities, landscape modification, and resource depression all play significant
roles in explaining the Pueblo Ill faunal pattern.

CHAPTER 6
GENDERED ANALYSIS OF FAUNAL DATA
In order to look at faunal change from a gendered perspective for Shields
Pueblo, it is necessary to look specifically at accounts of meat procurement. Chapter 2
discussed gender in faunal analysis and introduced the argument that small domestic
animals, those kept in the household, are routinely raised by women, and are therefore a
product of female labor. This chapter will further investigate this notion by discussing
cultural beliefs associated with hunting and domestic production. The results of the
stable isotope analysis will be presented along with intra-site analysis. This chapter will
conclude with a gendered analysis of Shields Pueblo meat procurement.
Meat Procurement
Puebloan peoples of the American Southwest ate a variety of wild and
domesticated meat sources. At Shields Pueblo some 35 taxa have been identified to
species level. Of these, approximately 15 species (including large rodents and
woodrats) were commonly eaten by pueblo peoples in historic times; some 17 species
were commonly procured solely for ritual or medicinal purposes; and at least 3 of the
rodent species are typically intrusive and probably not culturally deposited (Gnabasik
1981). Wild meat was procured through either individual or communal hunts. Deer
would have been available in the higher elevations and even in the lower sagebrush
desert habitats surrounding Shields Pueblo. Deer and other artiodactyls can be hunted
by individuals, small groups or communities. There are some reports of large-scale
antelope drives requiring up to 70 individuals (White 1932). Large game was hunted
exclusively by men.

War and hunting were often closely connected by the Puebloan people.
Communal hunts were organized by war captains and performed by warriors (generally
defined as all able-bodied men of the community) (Anell l969:6l ;Lange 1959: 128-1 29;
Parsons 191 8:l73; White l974a:I 26,301,302). Communal hunts among the historic
pueblos were associated with various ritual events and with particular ritual societies.
This has been evidenced time and again by the connection of Puebloan ritual and
ungulate hunts. For example, ethnographic records for Sia Pueblo indicate that meat
from communal hunts was dried and kept for Katsina rituals (White 1974a:l26, 301,
302). Men gained positions in the warrior society by being the first to touch the felled
animal (Lange 1959; Parsons 1920; White 1974b). At the Santa Ana, an animal-killer
Opi (member of the warrior society) was also considered a man-killer Opi, even if he had
not taken a human life in battle (Parsons 1920: 66-67).
Lagomorphs were by far the most abundant meat source for the Anasazi and
such as large game could be hunted in large groups or by individuals. Even rabbits
were generally hunted by men. When rabbit drives were called by the war chief to
supply meat for ceremonial purposes, only men could participate (Parsons 191 8, 1920,
1921, 1970; White 1974a:144-145). Non-ceremonial, or general hunts were more open
to women in some communities. At Acoma, San Felipe, and Santo Domingo women
were allowed to attend general hunts as retrievers and at San Felipe a fall rabbit hunt
consisted of young unmarried girls (these hunts were held in the horticultural fields of the
pueblo) (Lange 1959: 129; Parsons 191 8:l73; White l932:52). It is also important to
note that there are scenes of communal drives on Mimbres pottery as well as preserved
drive nets, indicating that such drives occurred prehistorically as well (Szuter 2000).
One in particular shows a group of people (of unidentifiable sex) using nets, sticks, and
crooks to drive jackrabbits (Shaffer and Gardener 1995). While images on pottery are

not necessarily indicative of daily activities, they do illustrate the existence of communal
hunting practices during prehistoric times.
Communal drives consisted of groups of hunters with various weapons (clubs,
bow and arrow, throwing sticks) and nets. Rabbits were surrounded and often forced
into smaller, tighter groups (or even into nets or "corrals") and then dispatched by armed
individuals. Both jackrabbits and cottontail rabbits were procured by this method and
continue to be hunted in the same fashion at Taos Pueblo today. Rabbits (particularly
cottontails) were also taken during "garden hunts." Agricultural fields provide ideal
habitat for cottontail rabbits, so these were likely caught, snared, or killed with throwing
sticks or bow and arrow to provide meat for family meals. Rabbits were also taken
expediently, while men were performing other tasks away from the pueblo and
surrounding fields (White 1974b).
Turkey Production
There are only a few ethnographic accounts of Puebloan groups keeping
domestic turkey or fowl in corrals or other forms of enclosures by each family (Reed
1951 ; Henderson and Harrington 1914). Historic and ethnographic records indicate that
turkey was kept both for food and for secondary products, namely feathers. Reed (1951 :
199-200) even mentions that women (specifically) wore turkey feather blankets. In order
to discuss prehistoric turkey production, it is necessary to first establish the evidence
that turkeys in an archaeological assemblage are, in fact, domesticated and not wild
hunted. This can be established by looking at the spatial distribution of turkey at an
archaeological site or by analyzing their diet.

Spatial evidence of turkey production
Several of the questions asked during analysis concern evidence of turkey
domestication. One question is whether or not turkeys were kept in pens associated
with particular households, or if turkeys were allowed free range. Physical evidence of
turkey pens should be indicated by: 1) architectural enclosures (such as the bird boxes
seen in the Casas Grandes region), or postholes indicating "corrals"; 2) deposits of bird
droppings; 3) large deposits of eggshell, and areas with increased frequencies of fetal or
very young turkeys. Turkey manure may show up in the archaeological record as actual
"lenses" of droppings in the stratigraphy (which has been noted for other domestic birds
in the American Southwest by Minnis et al. 1993). Turkey manure contains high levels
of nitrogen, phosphorus, and potassium (www.discover~farms.or~ 2005;
www.dirtdoctor.com 2003, 2004). Thus, soil in areas where turkeys were kept should
have higher levels of these substances. Higher frequencies of eggshell and fetal or very
young turkey remains would also indicate nesting areas.
While no definitive turkey pens can be identified at Shields Pueblo due to lack of
surface architecture and the lack of plazas, basic spatial analysis of the distribution of
eggshell is possible. More turkeyllarge bird eggshell is associated with non-structure
contexts than with structures. At Shields Pueblo some 522 (58%) eggshell fragments
come from midden deposits, while 380 (42%) come from structural contexts. Non-
structures 1409, dating from sub-period 8 (primarily secondary midden refuse) and 152,
dating from sub-period 7 (primarily from walllroof fall contexts) have the highest number
of turkeyllarge-bird eggshell among the non-structural contexts; while structure 139,
dating from A.D. 1060-1 150 has the highest among the structures. Eggshell comes from
two contexts in this structure, some 200 fragments were recovered from walllroof fall,
while 82 were recovered from a primary refuse context from an undefined feature. In
most of these cases, eggshell is not in a primary context. Instead, these deposits of

eggshell are in contexts described as "secondary deposits" by Crow Canyon. While
there does not seem to be any evidence of primary deposition of eggshell at Shields
Pueblo, this does not rule out the existence of turkey pens. Instead, such as much of
the spatial analysis at Shields Pueblo, it may simply point to poor preservation of surface
contexts.
Changes in the frequency of eggshell deposition over time may indicate changes
in turkey production. Such as turkey bone, increases in the rate of deposition may
indicate intensification of turkey production. In order to understand the true amount of
eggshell deposited during each sub period, it is necessary to calculate the rate of
eggshell deposition (Table 37). Figure 22 illustrates the change in frequency of
eggshell deposition over time compared with population. Periods 7 and 9 have the
highest relative rates of eggshell deposition. In both cases, the majority of fragments
come from particular "non-structure" and "structure" contexts. Because of the nature of
eggshell, simple counts of fragments are not reliable in estimating actual number of
offspring. However, all of the eggshell fragments are in the same size category (e.g.,
between one and two centimeters in length). Measuring the mass of eggshell, would
provide a better gauge of relative frequency. The largest single context sample of
eggshell comes from structure 139 (with 200 fragments ). This sample weighs
approximately 10 grams. Unfortunately, no other samples were weighed (mainly
because most samples simply contained only a few fragments). Using pottery
deposition as a comparison, there seems to be an inverse relationship between
deposition of eggshell and pottery except from Sub period 8 to 9, where both rates begin
to rise.

Table 37. Rate of Deposition of Eggshell ~ l y r l m ~ .
Relative Rate of Deposition of Eggshell Over Time
725- 1020- 1060- 1150- 1225- 800 1060 1150 1225 1280
Sub Periods (Years A.D.)
Pottery I Eggshell ! --
Figure 22. Relative Rate of Deposition of Eggshell per Sub Period compared to Pottery deposition: Shields Pueblo. '(Pottery deposition rates glyr.m2 are on the left; eggshell deposition rates ~ l ~ ~ l y r l r n ~ are on the right).
Occurrences of fetal turkey remains have also been identified and analyzed at
Shields Pueblo. Only a small number of turkey fetal remains was identified (NISP of
17). This low number is not surprising. Given the size and density of fetal bone, it is not
expected to survive site formation processes as well as adult bone. The majority of fetal
turkey bone comes from non-structures 1418 (35.2%) and 1310 (41.2%). Such as much

of the eggshell, the fetal turkey deposit in non-structure 1418 is associated with walllroof
fall. The fetal turkey deposit in non-structure 131 0 comes from the contents of a
"surface" feature. In this case, "surface" is not used to describe ground surface, but an
archaeological surface, although, it is unlikely that this constitutes a turkey enclosure
feature. This area is, in fact, believed to be associated with prehistoric soil excavation
for mortar manufacture (Duff, personal communication).
Turkey and Gender
The most important question asked in this dissertation is whether or not turkey
were raised by women. In Chapter 2, ethnographic evidence from around the world was
discussed in order to illustrate the basic pattern of care associated with small,
household-based domestic animals. Proof of gendered care of turkey in the
archaeological record is difficult to find. One of the most important patterns noted in c
Chapter 2 was that animals fed from household stores of food, and leftovers of
household meals, were invariably fed and cared for by the women of the house (as they
had control over the distribution of domestic grains and other plant foods, and were also
typically responsible for cooking meals and cleaning up refuse afterward). If turkey were
raised by the women of the household, then we should expect to see proof of their being
fed from domestic stores of food.
Since the primary domestic food source was maize, isotopic analysis of turkey
bone should show that they were eating a diet dominated by C4 plants (Matson and
Chisholm 1991). Results of the stable isotope analysis do in fact show that turkeys were
ingesting large quantities of maize (Table 38). In addition to the carbon isotope analysis,
nitrogen was also measured. If turkeys were allowed free range through the puebloan
fields, they would be able to browse for food (mainly insects or other invertebrates). All
of the turkey samples had nitrogen levels between 6.12 and 8.23, so that the carbon to

Table 38. Isotope values for samples of jackrabbit (LEP), cottontail (SYL) and turkey (MEG) collagen. Note the anomalous value for LEP-16
nitrogen ratios ranged from 3.08 to 2.93 respectively. These nitrogen levels indicate that
SFU ID MEG-1
the turkey sampled were primarily vegetarians (Ambrose 1993). Given both the results
of the carbon isotope analysis and nitrogen isotope analysis, it seems likely that turkeys
- %V~.?DB&O) -8.41
were confined or given limited movement and fed surplus household grain. While this is
not direct evidence of care by women, it does bolster the argument that women raised
. - 9 4 v AIR (%) 7.1 1
turkeys.
If lagomorphs are considered, a different diet is indicated by the carbon isotope
C t W 1 N (X) 45.94 1 15.76
analysis. With the exception of one jackrabbit, all of the lagomorphs have carbon
C:N 2.91
values indicative of C3 plants (local shrubs and grasses). The single jackrabbit with an
anomalous reading seemed to have a diet mainly consisting of C4 plants (maize, in this

case). Similar results were obtained on carbon isotope analysis of turkey from Sand
Canyon Pueblo (Driver, personal communication).
Discussion
Recently, there has been an increase in gendered studies of space in reference
to labor organization and household organization in the southwest (Hegmon et a/. 2000;
Nelson et a/. 2002; Ortman 1998). Much of this has focused on certain aspects of
women's labor, namely grinding corn and craft production. This research has looked at
the prominence, location, or visibility of women's task areas in order to rate the emic
importance of women and women's activities. This line of research is not new to
anthropology. For example, Bourdieu (1973) was one of the first anthropologists to use
household layout as a proxy of women's status. While this source is outdated, it is
notable for its attempt to equate status with spatial organization. Bourdieu (1973)
argued that the low social status of women in the Berber culture was signified by the
"marginal" placement of women's work areas. Since Bourdieu's initial study, Hegmon et
a/. (2000); Nelson et a/. (2002); and Ortman (1998) have used this line of reasoning to
argue that women and women's labor in the prehistoric Southwest held a certain amount
of cultural prestige. In the Southwest women's work spaces (i-e., cooking hearths,
metates, and mealing binslrooms) have been identified in prominent, central locations
and often shared space in kivas. The amount of space allocated may also be used to
"measure" its social significance. Nelson et a/. (2002) attempt to show that the
positioning and "commitment" of space used by women is related to the scale of craft
production and thus, its importance to household economies. They use the highland
Mayan community of San Mateo lxtatan as an ethnographic analogue for Southwestern
Puebloan groups. While their findings do not necessarily support their hypothesis (as
they found women's surplus craft production to be fairly invisible), they do lead to the

question, "Is it possible to gauge the cultural significance of women's meat production?"
In fact it should be possible to "measure" status based upon spatial organization
Ethnographic descriptions of where turkeys were kept are scant. Reed
(1951 : I 99) mentions early Spanish records that indicate large numbers of turkeys were
held in corrals. In general, very little research has been done in an attempt to find
archaeological signatures of prehistoric turkey pens. In fact, the only sites in the
southwest with evidence of domestic bird housing is found in the Casas Grandes region
of northern Chihuahua. The site of Paquime (or Casas Grandes), as well as other sites
in the region have been identified as having bird enclosures (Di Peso 1974; Minnis etal.
1993, Maxwell 2002). Both turkey and macaws were kept within adobe bird boxes
located in plazas associated with sets of roomblocks (possibly indicating households)
(see Figures 23 and 24).
Figure 23. Macaw boxes, complete with box plugs, Paquime, Chihuahua, Mexico (personal photograph).

Figure 24. Turkey and macaw boxes in a residential plaza, Paquime, Chihuahua, Mexico (personal photograph).
While macaws were raised for ritual use and (even more importantly) for trade
throughout the Southwest, it is unclear if turkey production was for exchange. Given the
ubiquity of turkey remains throughout the Puebloan Southwest, production for exchange
is unlikely. However, turkey boxes are often located along-side the smaller macaw
boxes, in fairly prominent positions within the community. Therefore, it seems likely that
turkey and the raising of turkey by women was considered an important part of
community life, both for economic reasons (turkey as a food product or source of
feathers for blankets) and ritual (as a sacrifice or for fetishes and prayer sticks).
If the significance of turkey production is to be understood in the northern San
Juan region, then some effort must be made to identify the production of turkey spatially.

There does not seem to be any architectural evidence of turkey production (i.e., stone or
adobe bird boxes or enclosures) outside of the Casas Grandes region. While turkey
corrals or coops in the northern San Juan were likely ephemeral in nature (such as
fenced areas or perhaps brush structures) there should be some physical evidence of
turkey housing. These are discussed above in relation to proof of turkey domestication.
Another line of investigation into the importance of women's turkey production
might be seen in other forms of spatial analysis. The spatial analysis of the Shields
Pueblo faunal assemblage, discussed in Chapter 4, suggested some patterning of
artiodactyl and turkey remains. Artiodactyls were far more common in excavation block
1300, while turkey was more common in blocks 400 and 1400 where deer was either
very low or absent. Previously, it has been shown that artiodactyl remains are strongly
associated with particular structures and thus with certain ritual behaviors (Muir 1999;
Muir and Driver 2002). Ethnographic evidence backs this up, as many groups displayed
the remains of artiodactyls on the roofs of their houses as evidence of their standing
within the hunting societies (White 1974:302-304). It is possible then, that excavation
block 1300 contained such a structure, thus associating this area with male hunting
societies or ritual.
Turkey predominates in excavation blocks 400 and 1400 where it makes up
59.3% and 57.5% (respectively) of the economically important fauna. It is unlikely that
these higher turkey frequencies indicate ritual display or deposition, as both areas date
from early and late Pueblo Ill (well after turkey became a dietary staple). They are not
turkey burials, but simply deposits of turkey in secondary contexts, or refuse midden.
Instead, these high turkey frequencies may indicate intrasite socio-economic or
household differences. The fact that hunted fauna are low in these areas may indicate a
lack of access to hunted fauna (both large and small), or it may indicate that households
in these blocks were able to keep larger numbers of turkey due to surplus corn

production. In order to answer these proposals it is necessary to investigate other
archaeological lines of evidence. First, it is necessary to identify households at Shields
Pueblo. Given the lack of surface architecture, a proxy should be used. In this instance
kivas may be used to delineate household groups, as each kiva -as well as other
architectural suites, such as mealing rooms- are associated with a particular room block
as part of the Prudden unit (Ortman 1998; Varien 1999). Second, it is necessary to look
at the frequencies of other types of artifacts throughout the site (especially artifacts
associated with prestige or with feastinglritual events, such as decorated bowls).
Households with higher frequencies of "prestige" goods are likely more "powerful" within
their community. If high frequencies of turkeys are associated with lower levels of
"prestige" goods, then this would support the first hypothesis and indicate that these
households had less access to hunted game and may indicate a lower socioeconomic
standing. If high frequencies of turkey are associated with higher levels of "prestige"
goods, then this would support the latter argument and that these households may have
had a higher standing in their community. At the time of the writing of this dissertation,
the Shields Pueblo site report and artifact databases are not complete. Therefore,
household comparison is not possible at this time.
Much of the ethnographic literature stresses the importance of artiodactyl hunting
and the prestige gained by the hunter (Anell 1969; Lange 1959; Parsons 191 8, 1920,
1921, 1970; White 1974), but little mention is made of the importance of domestic turkey.
It is clear from the association of turkey feathers and turkey images with religious
iconography that it was an important ritual species. Because turkeys are domesticated,
they have been deemed as less important by many ethnographers (such that little if any
mention is ever made of turkey raising among Puebloan communities). Even more
modern research has associated greater prestige with wild versus domestic animals.
Kent (1989) argues that as animals become domesticated, they are placed in separate

categories than wild animals. Wild animals are often placed within the same category as
humans by many hunting societies, such that wild animals are often anthropomorphized.
Domestic animals, by contrast, are represented in a different category from human and
wild animals, that is lesser, non-intellectual, and therefore not as important as hunted
game (Kent 1989). However, this research does not include cultural examples where
domestic animals are highly valued as evidence of wealth, power and prestige, and as
necessary parts of particular ritual events (i.e., funeral rights, weddings, war rituals) such
as pigs among the Melanesians (Feil 1984; Rubel and Rosman 1978; Strathern 1971,
Strathern 1972). Because of their use and importance in various religious artifacts and
iconography, turkey were likely highly prized for their feathers as well as for their meat
and may have had a different but "equal" categorization with large game animals. Just
because turkey was domesticated does not mean that it was relegated to a lesser status
than hunted game. As dependence upon domestic turkey as a source of meat
increased through Pueblo II and Ill, it would have become even more economically as
well as socially important to households. While large game animals were associated
with the idea of "masculinity" and men's activities outside of the pueblo (as
hunterslwarriors); turkeys may have symbolized "femininity" and the
productivelsupportive capacities of women within the community.
Both of these aspects, masculine and feminine were equally important to Pueblo
life. This idea of complementarity is not new. Roscoe (in reference to 3rd and 4th
genders) has argued that males and females have equally important roles within
Puebloan societies, extending through all aspects of daily and ritual life (1991,1998).
Even rituals associated with large game hunting involved the inclusion of women through
the sprinkling of sacred corn meal over slain animals (Parsons 1920:footnote 5: 65-66;
Hegmon et a/. 2000). Furthermore, the location of women's activity spaces (i.e., mealing
rooms) in direct association with kivas, is indicative of their importance in household

ritual life as well as economic life (Mobley-Tanaka 1997). Other aspects of
Southwestern iconography and ritual behavior also support this viewpoint. In addition to
being associated with women, bird images, macaws, and turkeys are also associated
with shamanic journeys as helpers, guides, or caretakers (VanPool and VanPool2006:
68). This is supported by images of shamanic trance and travel into the spiritual realm
and by ethnographic accounts of the wives or sisters of shamans caring for and
protecting his body during trance (Wilbert 1987: 157-1 58; Thevet 2001 [ I 5571; VanPool
and VanPool2006:68-69). While Paquime is located a great distance from the Anasazi
and can not be used for direct comparison, VanPool and VanPool's study does seem to
indicate a fairly widespread pattern of women's connection with birds in the Southwest.
It is clear, given the regional faunal patterns over time, that turkey became an
increasingly important resource during Pueblo Ill (A.D. 1150-1300). In fact, turkey was
the primary faunal species found at many sites, including Sand Canyon Pueblo, Knobby
Knee Pueblo, Roundtree Pueblo, Nancy Patterson Village, among others, during Pueblo
Ill (refer to Table 35, Chapter 5). Turkey, as a domestic source of meat is dependable
with controllable production. Therefore, as hunted sources of large game became
increasingly scarce or too risky to pursue, turkey would have become more valuable.
As control over turkey resources lay with the household unit (Reed 1951) and ostensibly
with the women of the household, an increase in the importance of turkey would likely
have increased women's economic and social standing within the pueblo. it also seems
likely that a woman's economic and social prestige could have been increased based
upon the household's "rate" of turkey production. Moreover, the "rate" of turkey
production would affect various household decisions, such as how much corn was
planted each season, and how much of this corn would be available for turkey feed, thus
giving women greater control over household economic activities.

During Pueblo Ill, men became increasingly preoccupied with "outside" pursuits
such as warfare, protectionldefense, raiding, etcetera. This may have afforded women
increased responsibilities and more control over household as well as community
decisions. The decline in male artiodactyl hunting associated with particular
rituallreligious events would have been supplemented or replaced by turkey production.
It is likely that women may have taken a greater role in religious events (i.e., beyond
food preparation and serving) and were probably involved in organizing household
contributions, and redistributionlfeasting for these events. The archaeological signatures
of these behaviors would be difficult to identify. However, there are some examples of
women with achieved status during the protohistoric period at Hawaikku (Howell
1995,1996). Here some 14 burials were uncovered that showed an incredible amount
and richness of burial goods (Howell and Kintigh 1996). Just under half of these burials
were of women, with at least one of these women interpreted as a medicine sodality
leader (or religious leader) (Howell 1996, 2001; Howell and Kintigh 1996). However,
during Pueblo Ill individual status was not expressed in burials. Kantner (2004) argues
that there were no identifiable inequalities between inhabitants of Pueblo Ill
communities, nor among households. Instead, inequalities seem to be expressed
between different sized communities (Adler et a/. 1996; Lipe and Varien 1999). Thus
burial analysis and even household comparison may yield little in the way of evidence of
changes in women's status. Thus, it would be best to try and reconstruct the levels of
production between households. This could be measured by comparing differential
frequencies of large storage jars between households, by looking at the numbers of
grinding implements, the size and prominence of mealing rooms, or the sizelnumber of
turkey pens between different households in the community. If particular households
seem to be producing maize or turkey at a larger scale, then this could indicate greater

involvement in producing foods for feasting (or it could simply indicate larger
households).
Chapter Summary
Meat procurement is an activity performed by both men and women. While men
primarily hunt (ostensibly for prestige, as a means for maintaining offensive and
defensive capacities, as well as meat), women are involved in the production of
domestic meat sources (turkey in this instance). Ethnographic resources indicate that
Puebloan communal hunts involved women as well. However, there were caveats to
their participation. First, women could only participate in hunts for small game. Second,
women could not participate in hunts for "ritual" purposes. Third, women generally acted
as retrievers of felled animals. And finally, women participated in communal hunts within
reasonable proximity to the pueblo. Therefore, when meat procurement was tied to
male prestige and associated with warrior societies, women would not take place in
those hunting activities. When meat was domestic, coming from domestic sources or
was hunted for household purposes only, women may have actively procured/produced
meat resources.
Turkey were likely raised and tended by women. The turkeys of Shields Pueblo
were likely penned, as evidenced by stable isotope analysis of a sample of turkey bone.
The results indicate a vegetarian diet based upon maize and not local C3 plants. If
turkey were allowed free forage there should be higher levels of C3 as well as greater
levels of nitrogen (indicating that turkey were ingesting insects and other invertebrates).
Furthermore, ethnographic evidence supports this with descriptions of "household"
turkey flocks. Because turkeys were fed maize from household stores and possibly
meal remnants, their care was probably the responsibility of women.

The categorization of meat procurement/production activities into "male" and
"female" illustrates the idea that these activities complemented each other both in the
acquisition of protein into the diet as well as in ritual and cultural importance. The
connection of men with large game hunting (interchangeable with warfare) and of
women with producing domestic animals simply reifies their position within the
community and within their cultural worldview. Men were associated more with the wild
or with the world outside the pueblo (and therefore dangerous environment), while
women were associated with the domestic and the life inside the pueblo. This
connection and the increasing importance of turkey as a meat resource may have also
increased women's economic power within the household and community at large as
well as increasing their social prestige.

CHAPTER 7
CONCLUSIONS
The purpose of this dissertation has been to analyze the faunal data from Shields
Pueblo and to investigate gendered roles in meat procurement. Another goal has been
to understand how Shields Pueblo fits into the regional context and how multiple
environmental, demographic, and cultural factors have changed the Anasazi culture.
This has been accomplished by considering several lines of evidence. First, gendered
faunal analysis was discussed and a method of investigation and reconstruction
established. Cross-cultural studies were used to investigate the roles women and men
play in the care of domestic animals. Secondly, the physical and cultural history of
Shields Pueblo was summarized in order to place the site and its assemblage into
context. Next, the analyzed faunal assemblage was discussed. Species frequencies
were discussed and an analysis of taphonomic factors was performed in order to
understand the changes to the faunal assemblage and how these affected preservation.
Then, both time and population were considered in reference to faunal change and
spatial analysis was employed to locate any patterns in species distribution across the
site. Next, the Shields Pueblo faunal assemblage was compared with other sites in the
Northern San Juan region in order to see how Shields fits into the general regional
pattern. Finally, a gendered analysis of the faunal assemblage was carried out to track
changes in meat procurement, using ethnographic, archaeological, and dietary
reconstruction data. All of the results of these analyses will be discussed in order to
address each of the research questionslobjectives stated in Chapter 1.
1) Is there variability in species frequencies from Pueblo I through Pueblo Ill?
2) If so, what is the specific relationship between hunted and tended fauna over
time?

3) Is there a decrease in large game over time and if so, is this a result of short-
term residentiaVpopulation decline or a decline in hunting?
Research expectations
Chapter 1 outlines a series of expectations concerning faunal patterning. These can be
summarized as follows:
There should be a decline in artiodactyls and communally hunted Lepus over time There should be an increase in the frequency of Sylvilagus in relation to other hunted species. Tended fauna should be distinguished from hunted fauna by diet (i.e., turkey should have a maize-based diet and not one based on "wild foods"). There should be a decline in large hunted game from PI1 through PIII. There should be a significant increase in turkey as hunted game declines.
Each expectation will be addressed by discussing faunal variability, hunted versus
tended fauna, and how faunal patterning relates to population at Shields Pueblo.
Faunal Variability
Chapter 4 presents the analysis of Shields Pueblo's faunal assemblage. There is
significant variability among species over time. Table 26 illustrates the standardized
residuals for artiodactyl, lagomorph, and turkey frequencies over time. Artiodactyls and
turkey vary the most over all time periods with total standardized residuals of 31.4 and
45.9 (respectively).
Artiodactyls are relatively rare in the Shields Pueblo assemblage across all time
periods. During early Pueblo I (A.D. 725-800) artiodactyls make up approximately 5% of
the total selected taxa. Mid Pueblo II (A.D. 1020-1060) has the highest frequency of
artiodactyls at 14%. Artiodactyls then decline during late Pueblo II (AD. 1060-1 150)
(7.9%); early Pueblo Ill (AD. 1150-1225) (1.7%); and then increase by less than a single
percent (2.5%) during the final period of occupation, late Pueblo Ill (A.D. 1225-1280).

Turkey is far more common in the Shields Faunal assemblage than artiodactyls
and shows the most variability in frequencies over time. During early Pueblo I, turkey
comprise just over 18% of the total of selected taxa. Their frequency rises to 38.7%
during mid Pueblo It, drops to 14.2% in late Pueblo II, rises during early Pueblo Ill to
41.3%, and then falls to 32.6% in late Pueblo II.
Lagomorphs also vary over time, however they tend to retain relatively high
frequencies throughout. Frequencies range from a low of 47.3% to a high of 77.9% of
the total selected taxa.
Overall, these trends follow those described by Driver (2002) for the Northern
San Juan region as a whole (with a few exceptions). The majority of Pueblo I sites in the
region are dominated by lagomorphs and artiodactyls. Shields Pueblo differs most in its
low frequency of artiodactyls and higher frequency of turkey. Pueblo II sites vary greatly
in faunal frequencies, although lagomorphs tend to dominate in most assemblages.
Artiodactyls and turkey vary greatly among sites and (in this period) do not seem to be
associated with site location. Pueblo Ill shows the strongest patterning among sites in
the Northern San Juan region. The majority of assemblages show a decline in
artiodactyl frequencies during this time period. There is also a general increase in the
frequency of turkey across many sites (especially among small sites). Shields Pueblo
has slightly fewer turkey during this final period of occupation and more lagomorphs,
however it still fits into the pattern seen by other sites in the immediate region (i.e., the
McElmo dome). The relationship between artiodactyls, lagomorphs and turkey can be
better explored by looking at specific relationships between hunted and tended species.
Hunted versus Tended Fauna
Specific taxonomic indices were used to understand the relationships between
particular taxa. Relationships between hunted taxa were explored with the artiodactyl

index and the lagomorph index. The artiodactyl index considers the relationship
between large game (artiodactyls) and small game (lagomorphs). The lagomorph index
focuses on the relationship between Sylvilagus and Lepus (cottontails: jackrabbits).
Both are helpful in understanding the effects of local environment, climatic change, and
population on hunted species. The relationship between hunted and tended taxa was
investigated with the turkey index, which measures the ratio of turkey and large bird to
all lagomorphs over time. This index is particularly important to this dissertation because
it provides a measure of the intensity of women's turkey production during each sub
period.
The relationship between large hunted game and small hunted game follows the
expected pattern (discussed previously by Driver [1996, 2000, 20021; Muir [1999]; Muir
and Driver [2001]) of a decline in large game from Pueblo II to Pueblo Ill. For reasons
discussed elsewhere, large game is preferable to small game and its frequency in
archaeological sites is controlled to a large extent by its availability.
Such as the general artiodactyl frequency, the artiodactyl index is lower in Pueblo
I than would be expected. The ratio of artiodactyls to lagomorphs is 0.06 for this time
period, which is lower than the indices from other contemporaneous sites. While this
seems problematic, it is important to consider the low artiodactyl index in an
environmental context. Most of the Pueblo I sites are found in the Dolores River region.
The Dolores region provides a mixture of environments with the highest productivity of
large game species. Shields Pueblo, is more similar to the other early Pueblo I sites in
the McElmo region. These sites sit primarily in the pinyon-juniper biotic community, and
while large game is found in this community, they are not necessarily as plentiful as
those found in other communities.
Artiodactyl indices are higher in mid Pueblo II. In fact, mid Pueblo II has the
highest artiodactyl index among all subsequent time periods (0.20). This index is

achieved after a period of depopulation at Shields Pueblo, thus it is not surprising.
During a period of settlement hiatus, the lack of hunting would allow for a rebound in
local game populations (assuming there are no other "active" sites in the locality).
Once the artiodactyl index reaches its peak in mid Pueblo II, it begins a steady
descent. By the end of Pueblo II the ratio of artiodactyls to lagomorphs falls to 0.07,
then bottoms out at 0.03, during early Pueblo Ill, where it remains for the duration of site
occupation. This indicates that small game became the primary hunted species at
Shields Pueblo during the last 140 years of occupation. This follows the pattern seen at
other sites in the region. Most sites have a low ratio of artiodactyls to lagomorphs.
Various hypotheses have been developed to explain this phenomenon. Environmental
and climatic studies indicate that Pueblo Ill was a period of cooler dryer weather, which
affected local environments to varying degrees. Human populations were also growing
exponentially during this time period. Human population growth combined with declining
environmental productivity caused communities to crowd into productive environments.
Several population studies have indicated that population doubled and even tripled (in
many cases) from late Pueblo II to late Pueblo Ill (Wilshusen 2002: Table 5.6).
Mahoney et a/. estimate that the momentary population of the Goodman Point
community rose from 101 to 330 persons from late Pueblo II to Late Pueblo 111 (2000:78).
Site aggregation also increased during this time period, with the McElmo region showing
some 78% overlap of 2 km site catchment boundaries (Varien 2002; Varien et a/. 2000:
Figures 2-4). While these population trends have been shown for other areas, the exact
relationship of population size to large game at Shields Pueblo still needs to be
established (this will be discussed below).
The relationship among lagomorphs is also enlightening. Szuter and Bayham
(1989) focus on the relationship of Lepus to Sylvilagus, in relation to site size. They
argue that Lepus should be more prominent in large sites where larger amounts of land

have been cleared for farming, while Sylvilagus should dominate at smaller sites, with
less clearing. The Shields Pueblo lagomorph index does not support this argument. In
fact, Sylvilagus dominate throughout the history of occupation at Shields Pueblo. The
lagomorph index only declines during early Pueblo II after a period of depopulation.
Others as well, have noted that the ratio of Sylvilagus to Lepus is high at large sites in
the McElmo region (Driver 2002; Muir and Driver 2001). It becomes apparent, that
Szuter and Bayham's (1994) lagomorph index measures local environmental conditions.
In certain environments, clearing of land, use and subsequent depopulation of
agricultural fields will result in creating a more open grassland environment. In the
McElmo region, clearing and subsequent depopulation of fields results in the
colonization of shrubby plant species (Adams and Bowyer 2002), thus providing the
shrubby cover that is more attractive to Sylvilagus than Lepus.
The turkey index provides a measure of the importance of tended domestic fauna
compared to small hunted fauna (lagomorphs in this instance). The ratio of turkey to
lagomorphs directly measures the change in the degree of women's meat production in
relation to hunting (a primarily male activity, although women do often hunt and trap
small game). The turkey index at Shields Pueblo rises and falls from one period to the
next in an interesting pattern. During initial settlement the turkey index stands at 0.19
(higher than other sites in the region, but not completely uncommon). During early
Pueblo II, the initial index sits at 0.61, substantially higher than the previous period.
Once the possible turkey burials are removed from the equation, the index is only slightly
higher than Pueblo I at 0.21. This seems reasonable, given the fact that Pueblo II is the
time period when turkey first began to be used as a food source (Munro 1994).
However, in the later portion of Pueblo II, the turkey index falls to 0.13, which, given the
increase in population, seems low. On the other hand, considering the variation in
turkey indices during this time period it is not wholly unexpected. It is not until early

Pueblo Ill that the turkey index begins to rise significantly in relation to lagomorphs
indicating a greater reliance on women's meat production. During this sub period, the
index is 0.42. This drops slightly in late Pueblo Ill, down to 0.33 (which is similar to the
turkey index in the Sand Canyon assemblage). This rise and fall in the prominence of
tended fauna (thus, reliance on women's labor) raises some interesting questions. Is the
dependence upon domestic turkey a function of the availability of hunted game or does
the availability of surplus maize for feed play a role?
Population and the Decline of Large Game at Shields Pueblo
The final research question asked whether or not the decrease in large game
was the result of an increase in population at Shields Pueblo or the result of short-term
residential depopulation. Because the majority of site population estimates rely on the
use of number of rooms, roomblocks, or kivas per room block, it was necessary to find
another proxy for population at Shields Pueblo. Pottery deposition rates have been used
by others in the region to provide rough estimates of population growth and decline
(Ortman et al. 2000). Once pottery weights were converted into rate of deposition it was
possible to measure the relative rate of deposition (thus, population) over time. The
resulting curve (refer to Figure 14) indicates that Pueblo I deposition rates were 12.5% of
the total rate of deposition. This falls to 4.9% in early Pueblo II, after the period of
occupational hiatus (found throughout the McElmo region during this time period [Duff
and Wilshusen 2000; Mahoney et a/. 2000; Varien 2002; and Wilshusen 20021). From
this point in time the rate of pottery deposition rises. There is a small increase in the rate
of pottery deposition during the late Pueblo II sub period, at 6.3% of the total. Early
Pueblo Ill is even higher at 18.1 % of the total deposition rate. These rates of pottery
deposition over time do not indicate a decline in the population after mid Pueblo II. It
has already been established that large game frequencies drop from late Pueblo II

through late Pueblo Ill. Therefore, it seems the drop in large game was not caused by a
decrease in human populations. The trend in declining artiodactyl frequencies, therefore,
seems to have a direct inverse correlation to the increase in population., so that as
human population increases, artiodactyl frequencies drop.
Discussion
Most of the research expectations are met (or met with some caveats). There is
a significant drop in large game over time. It was expected that there would be a larger
frequency of artiodactyls dating to the Pueblo I period. This was not the case. Instead,
artiodactyls are most frequent in the Mid Pueblo II sub-assemblage. However, they do
decline (as expected) from late Pueblo II onward. It was initially expected that Lepus
would also decline over time. This expectation, such as the previous, was partially met.
Throughout the occupation of the site, Sylvilagus is the predominant lagomorph species.
Such as artiodactyls, it was expected that Lepus would be highest in the earliest periods
and decline in frequency over time. Instead, jackrabbits are highest during mid Pueblo II
and at their lowest during early Pueblo I (refer to Figure 11). Such as artiodactyls, once
they reached their peak frequency in mid Pueblo II, Lepus declined through each
successive time period. Sylvilagus was expected to increase in frequency over time.
Again, this expectation is met with the caveat that only mid Pueblo II and subsequent
periods be considered. During Pueblo I the lagomorph index is at its highest (0.9),
indicating that Sylvilagus are far more frequent than Lepus.
It was expected that tended fauna and hunted fauna should have different diets.
Tended fauna were expected to have a diet consisting primarily of maize. This
expectation was met. Turkeys have been shown to have had diets rich in C4 plants and
not the local C3 vegetation. Likewise, it is indicated that turkeys did not eat many

invertebrates (as their nitrogen ratios indicate a predominately vegetarian diet), and as
such, were likely restricted in their movements.
The final two expectations were that the frequencies of hunted game should
decline, while the frequency of tended meat sources increase. These expectations
were, by and large met. The rise and fall of turkey in the assemblage is related to a rise
and fall in hunted game. However, turkey does not rise in frequency incrementally over
time. Instead it rises and falls in relation to the frequency of lagomorphs in the
assemblage and does not seem to be correlated with large game frequencies.
Therefore, the inclusion of turkey in the diet seems to act as a supplement to rabbit.
Potential Causal Factors
In Chapter 1 several potential causal factors for the decline of large game and
increase in domestic turkey were discussed, as were the expected faunal patterns for
each explanation. Each hypothesis and its resultant faunal pattern will be addressed in
comparison to the Shields Pueblo faunal pattern.
Environment
The most commonly cited explanation for faunal change and depopulation in the
Anasazi region is the "great drought" or "Little Ice Age." This was a period of cooler and
dryer than average weather that is argued to have caused a decline in maize production
by shortening the growing season and interfering with the cycle of bi-annual precipitation
(Petersen 1986, 1988, 1992). While some have questioned the severity of the '"reat
drought" (such as Van West and Lipe 1992), it continues to be used as a factor that
exacerbated declining agricultural and environmental productivity. If environment is the
sole factor in cultural and faunal changes during the Pueblo II to Pueblo Ill time periods,
then certain faunal patterns should be present among the assemblages. First, there

should be a decline in large game at all sites within the study area. Likewise, there
should be higher frequencies of artiodactyls at sites located near permanent water
sources. Large game should decline in relation to domestic fauna. Finally, there should
be shifts in the faunal pattern for every climatic shift.
While most of these expectations are met at Shields Pueblo, they are not
necessarily true at the regional scale. There is an overall decline in large game during
Pueblo Ill at Shields Pueblo, which can be seen throughout the Northern San Juan
region. However, sites on or near Mesa Verde, in Johnson Canyon, and in the Mancos
region all have high frequencies of artiodactyls in their Pueblo Ill assemblages.
Proponents of the environmental hypothesis might argue that these sites are located
near permanentlreliable water sources. However, most sites in the region were situated
near seeps or springs, if no permanently running rivers or creeks were present (the
closest spring to Shields Pueblo was located at Goodman Point Pueblo). So, lack of
water resources cannot really be used to explain why artiodactyls are absent at the
majority of sites in the region. If the frequency of large game to domestic fauna is
considered, Shields Pueblo (such as most sites in the region) shows a decline in large
game and an increase in domestic turkey. Again, however, there are exceptions to this
trend. Sites such as Hovenweep 53, Hovenweep 94, Wallace Ruin, and Salmon Ruin
while they have low frequencies of large game, also have extremely low frequencies of
domestic turkey and much higher frequencies of small game, such as lagomorphs.
These discrepancies at Wallace Ruin and Salmon Ruin may be due to the emphasis on
architectural, not midden excavations. In regards to the final expectation, that each
climatic shift should result in a faunal shift, there simply are not enough data on the
paleoclimate of Shields Pueblo to either support or dispel this argument. However, the
paleoclimatic changes for the larger region are well understood. Pollen records indicate
that there were many little oscillations in climate from A.D. 750 to 1300. Some,

occurring in the mid A.D. 800s and A.D. 900-IOOOs, are quite dramatic (Davis
1996:Figure 12-5). However, large game does not decline in a predictable pattern
during these time periods. Instead, artiodactyls have a high frequency at many sites in
the region (refer to Tables 32 and 33). Therefore, while climate certainly has had an
effect on faunal assemblages, the actual affects are not completely clear and it is likely
that there are other forces at work.
Population Pressure
Population growth or pressure is another factor that influences faunal patterns
over time. If population pressure is the primary cause of faunal change, then there
should be a distinctive drop in large game. This should then be accompanied by an
increase in domestic fauna. Smaller sites, or those located in sparsely populated areas,
or in areas that are less bounded, should have higher frequencies of artiodactyls than
sites located in densely populated areas. While it is true that Shields Pueblo shows an
increase in artiodactyls when population is low, and a decrease in large game
frequencies when population rises; it is clear that the relationship between fauna and
population is more complicated when turkey are considered. Instead of a steady rise in
turkey as population increases, turkey frequencies rise and fall over time, and in fact, fall
as population reaches its peak in late Pueblo Ill. Rabbits dominate the assemblage
throughout the site's occupation. Another difficulty with the population pressure
argument lies with the location of Shields Pueblo in relation to other community centers.
While the McElmo region is densely populated, causing intensive crowding of site
catchment areas, Shields Pueblo is located along the eastern boundary of the McElmo
cluster. There are no sites located to the east of Shields Pueblo during Pueblo Ill. Sites
in the Dolores group (e.g., Emerson, Reservoir, and Escalante) were depopulated by
early Pueblo Ill, while sites in the area around present-day Cortez (Mitchell Springs and

Mud Springs) were depopulated by late Pueblo Ill (Varien 2002; Varien et a/. 2000).
Once these areas were depopulated, opening up this area for exploitation, why were
they not utilized by Shields Pueblo? Other sites in the McElmo region also seem to
"ignore" hunting grounds outside their immediate areas. Yellow Jacket Pueblo is
unbounded along its northern border with vast amounts of space between it and the
Dolores region, and yet artiodactyl frequencies are low (Driver 2002). These low
frequencies indicate that some other factor kept hunters close to home, exploiting
"crowded" local environments.
Social Factors
One of the trends noted in faunal patterning at the intersite level, was that large
regional centers maintained a higher frequency of artiodactyls than did smaller sites.
Driver (1996) ascribes this phenomenon to control over resources by larger influential
sites. If certain communities were able to control access to large game, then there
should be differential distribution of highly valuable resources (artiodactyls in this case)
among large sites. Small sites within the proximity of larger communities should have
low frequencies of artiodactyls and much higher frequencies of local small game and
domestic turkey. Shields Pueblo has a low frequency of artiodactyls and a relatively
higher frequency of turkey. Thus, it would seem as if Shields Pueblo was a small, less
influential site. In fact, Duff and Ryan (2000) have argued that Goodman Point Pueblo
was the community center during late Pueblo Ill. Unfortunately, this site has not been
excavated, so its faunal assemblage has not been analyzed. However, other sites in the
McElmo region have been analyzed in reference to their size and relationship to one
another. Sand Canyon Pueblo, along with Yellow Jacket Pueblo, Woods Canyon
Pueblo, and Castle Rock Pueblo are all large sites within the McElmo region. Driver and
Muir (2002) analyze these sites at the household, community, and sub-regional scales in

order to better understand the social and economic factors influencing the frequencies of
artiodactyls, lagomorphs, and turkey. In particular, tower blocks at Sand Canyon Pueblo
and Yellow Jacket Pueblo contained higher frequencies of artiodactyls than other
structures (Muir and Driver 2003). Muir (1999) suggests that these structures were
associated with hunting societies and may have been associated with communal hunts
(not unlike as those described in ethnographic records). The Sand Canyon area
contains several sites, the large Sand Canyon Pueblo and 13 smaller sites located within
close proximity. Muir (1999) is able to show that the smaller sites within the Sand
Canyon locality had higher frequencies of turkey with lower frequencies of both
artiodactyls and lagomorphs over time. In addition, Muir argues that Sand Canyon
Pueblo has a significant abundance of large game and lagomorphs. Greater artiodactyl
frequencies and the association of artiodactyl remains with tower blocks could suggest
the presence of "elites" at Sand Canyon as well as Yellow Jacket Pueblo (Muir and
Driver 2003). Certainly, some level of organization and leadership is required to plan
and carry out communal activities. However, pinpointing specific individuals as "elites" is
very difficult. Muir (1999) points out that while artiodactyls are typically found in certain
structures, there does not seem to be any differential distribution of meaty elements
across the site.
Economic specialization has also been suggested as a causal factor in faunal
change. With specialization, there should be evidence of increasing dependence upon
turkey and a decline in all hunted species. This is simply not the case at Shields Pueblo.
Even as artiodactyls decline, lagomorphs remain the dominant species. Likewise, no
evidence of flock management has been found. Munro (1994), in her investigation of
turkey production in the Sand Canyon locality, argues that there is equal distribution of
male and female birds. Munro (1994:147-148) further determines that 94% of turkey
assemblages are made up of adult individuals. If turkeys were being raised to maximize

production, there should be greater numbers of young males in the assemblage, as they
would have been preferentially culled from the flock. The Shields Pueblo turkey
assemblage has a slightly different ratio of adult to juvenile individuals. Approximately
60% of turkey in the assemblage is adult, while some 40% are juvenile. Since sex was
not determined for turkey specimens, it is unknown if this pattern would have differed
from Sand Canyon locality assemblages. It is unclear if turkeys were culled to control
populations. However, given the difference in age categories between Munro's (1994)
work in the Northern San Juan region and Shields Pueblo, it is possible that some culling
did occur. Another possibility is that the people of Shields Pueblo were having to kill
more birds to meet their needs and were killing animals before they could reach - maturity.
Warfare
Warfare cannot be considered as a singular cause for faunal change. Instead it
influences hunting decisions based upon the duration of the hunting trip and the distance
hunters are willing to travel to access game. Thus, there should be a decrease in game
acquired long distances away from communities who were susceptible to attack.
Smaller sites, with lower populations should be more greatly affected by this than larger
sites, as they would not have the population capacity to allow a group of hunters to leave
the community while simultaneously keeping a large enough contingent of men at home
for protection. This is essentially what is indicated at Shields Pueblo and other sites in
the region. Dependence upon artiodactyls declined, while dependence upon locally
acquired game (lagomorphs) and domestic turkey increased as the level of violence
increased during Pueblo Ill. Sites such as Sand Canyon Pueblo and Yellow Jacket
Pueblo were certainly large enough to maintain enough men to hunt and stay behind for
protection. There are several sites in the Northern San Juan region with evidence of

violence. Burnt Mesa, Cottonwood Wash, Sambrito Village, Teec Nos Pos 4, Castle
Rock Pueblo and both Sand Canyon Pueblo and Yellow Jacket Pueblo show evidence
of violence and even massacre (Kuckelman et a/. 2000). While human skeletal remains
were not excavated at Shields Pueblo, there is indirect evidence of violence in the
presence of tunnels. Tunnels connecting kivas to other structures have been likened to
"escape routes", allowing people to leave a subterranean kiva without moving out on to
more open, potentially dangerous kiva roofs (Kuckelman 2002). Previously, it was noted
that Shields Pueblo and Yellow Jacket Pueblo did not seem to utilize "open", productive
environments beyond the boundaries of their "local" catchment areas. These large,
empty spaces between communities are indicative of buffer zones. These "no-mans-
lands" tend to be located between communities who are habitually at war with each
other (Aikens 1966; Fairly 1989; Kuckelman 2002; LeBlanc 1999; Matson et a/. 1988;
Rohn 1989; Steward 1941). Several of these buffer zones have been identified in the
Northern Sand Juan region dating to late Pueblo Ill times (Haas and Creamer 1993;
Jewett 1989; Mera 1935; Wilcox 1981). The area to the east of Shields Pueblo could
have ostensibly acted as a buffer zone and as such, would not have been utilized for
hunting activities, thus keeping hunters close to home. Entering a "no-mans-land" could
have been viewed by enemies as an act of war, thus residents avoided such areas.
Discussion
It is clear that no single causal factor can explain the decrease in large game and
increase in small game and domestic fauna at Shields Pueblo. Instead, a combination
of climatic and population pressure, change in social organizationlsite hierarchy, and the
"complication" of warfare acted together to decrease the availability of large game at
Shields Pueblo and sites throughout the Northern San Juan Region. Because small
game was locally available, it did not significantly decline over time. While turkey shows

an over all increase in importance as a source of meat, there is no clear evidence that it
was bred for increasing stock production. Previous research in turkey usage indicates
that there were no patterns of kill off based on age or sex (Munro 1994). Turkeys were
seemingly killed off at random (although adults were killed more frequently). Turkey
frequencies vary over time, as does the relative amount of hunted lagomorphs. The
threat of violence would certainly have kept men closer to home, and while rabbits are a
quickly renewing resource, increasing populations would likely have put stress on
lagomorphs, requiring an additional source of meat.
Women are the likely choice to have raised turkey. Cross-cultural research has
shown that women are far more likely to care for small, household domestic animals
than are men. This can be explained primarily by the diet of most small domestic
animals. Pigs, poultry, and guinea pigs all share one thing in common; they are
traditionally fed from household stores of food (whether this is surplus agricultural
production or the "leftovers" from family meals). As women typically "control" access to
stored food and cook for the household, they are more likely to be responsible for
feeding household domestic animals. Based upon isotopic analysis it is evident that the
turkeys of Shields Pueblo were fed primarily on maize. Because they were fed from
household stores of grain, they were likely the responsibility of the female head of
household. Therefore, women were probably responsible for a sizable percentage of the
meat in the Pueblo II and Pueblo Ill diet.
Furthermore, the cultural significance of turkey production has been argued in
this dissertation to be high among the Anasazi of the northern San Juan region. Turkey
was used as both a ritual species as well as a source of meat. Its importance as a ritual
species is illustrated by the use of turkey feathers in prayer sticks, fetishes, altars, and
Katsina figures; the ubiquity of turkey imagery in pueblo iconography from petroglyphs,
pictograms, and pottery; and by its prehistoric use as an offering or sacrifice made

during certain rituals and events (i.e., the ritual closure of a kiva) as evidenced by turkey
burials. Turkey's significance as a domestic household animal is shown by its primary
use as an important source of meat for the household, and by its secondary uses (such
as bone for tools and feathers for turkey robes). Cultural importance has also been
illustrated by the location of turkey penslboxes within some pueblo communities. The
centralized, "public" placement of bird boxes at the site of Paquime and others in the
Casas Grandes region is demonstrative of the economic and ritual importance of these
birds. Because domestic birds were raised by women, their importance was certainly
reflected in the importance of women in pueblo society. Thus, production of turkey may
have provided women with added of prestige or status within the community. As the
availability of artiodactyls and men's hunting of large game declined, the importance of
turkey likely increased. Therefore, the ability to raise and keep large numbers of turkey
would indicate a household's ability to produce surpluses of corn (signifying its economic
success) and may also have been tied into ideals of religious success and power or the
ability to supply meat for feasts. Women belonging to these households were able to
gain both household as well as personal prestige. As turkey was the primary meat
resource for many sites in the Northern San Juan region this increase in the importance
of turkey and of women as meat producers probably meant greater participation of
women within the economic, social, and religious aspects of community life.
Intensification of production would mean a necessary increase in the amount of maize
produced for turkey consumption. Thus, women would have had greater control over
household agricultural production -especially concerning how much of the harvest would
be relegated to turkey feed. Likewise, as men became further engrossed in activities
associated with warfare and their role as meat providers for ritual events declined,
women's production of turkey would have held even greater ritual importance. This may
have involved an increase in women's involvement in planning religious events (or

feasts), such as organizing household contributions and even redistribution of turkey for
these events.
Evaluation and Suggestions For Research
This dissertation contributes a few different methodological and theoretical
arguments to research in gender, as well as Southwestern archaeology. First, it
provides a new model for other gender studies through the use of stable isotope data.
This line of evidence has been shown to be useful in reconstructing the care of domestic
household animals and thus, their likely caregivers. Second, the cross-cultural survey
conducted illustrates a strong cultural pattern in the care of household domestic fauna,
proving an important analogical argument for this and future research. Third, in regards
to temporal and regional faunal patterns, this dissertation has supported previous
research. Many studies have been somewhat hampered by the small size of their faunal
samples, thus limiting analysis and bringing results into question. Shields Pueblo is the
largest faunal assemblage in the McElmo region, and as such, provides more
statistically reliable results. Even with this large sample, however, it must be noted that
many factors have contributed to the potential for error. Chapter 4 laid out a series of
taphonomic factors that have potentially biased the results of this analysis. It was
previously noted that larger faunal species are far more common than small species.
Preservation as well as excavation techniques are largely responsible for this bias.
While some small rodent species have been identified, there is a distinct lack of other
small species, particularly among birds, reptiles, and amphibians. This lack of small
fauna has given the impression of a rather narrow diet base. While certain species were
likely preferred (and therefore more common in the assemblage), others (including song
birds, migratory birds, and reptiles) were also likely part of the diet.

During the preparation of this dissertation a few other avenues for future
research have been recognized. It is clear that there needs to be a better understanding
of local site paleoenvironments, as these local conditions greatly influence the
frequencies of particular species. For example, Szuter's and Bayham's (1989) study
argues for an increase in jackrabbits as human population and cleared land around
settlements increases. This is clearly not the case in the Northern San Juan Region,
where site microenvironments apparently differ significantly from regions to the south.
More needs to be learned about exactly which plant species colonize cleared and
depopulated agricultural fields in particular regions and how this change in the floral
environment affects local fauna. Paleobotanical studies for the region would certainly be
helpful in this line of research by looking for the presence of colonizing species in the
archaeological record. This could also be done using experimental archaeology (i-e.,
clearing segments of land in different environments, elevations, and regions). This
would be a large study and would certainly require collaboration among different
researchers.
Yet another avenue for research concerns the affects of population, site size, and
aggregation on faunal patterning. While this relationship is well understood for later
time periods (especially Pueblo Ill), earlier periods are assumed to have small
populations and greater numbers of large game. Shields Pueblo's early Pueblo I faunal
assemblage does not follow the expected pattern (nor do other sites in the region), as it
has a very low frequency of artiodactyls. There needs to be more research into early
sites with particular attention to population estimates for early, and late Pueblo I
occupations. Particular attention to number of households per site or number of rooms
per site should provide archaeologists with population estimates for early sites.
Much of the follow-up research will require additional cross-cultural analysis,
continuing the sampling began in this dissertation. It is also important to explore current

ethnobiolological and ethnoarchaeological research that is currently being performed in
the region. It would be ideal to interview descendant community members on their
interpretation of the faunal data collected from Shields Pueblo. If the latter research
suggestion is done, then these findings could be published with the Native community in
mind.
In regards to how this research was conducted, I would not have changed any of
my methods. I feel that the research methods used in the preparation of this dissertation
were adequate for the data. The cross-cultural analysis was both interesting and was
able to provide needed information. Stable isotope analysis proved to be an important
test of a "new" method in gender research. It would have been preferable to take more
samples from a larger population of animals. Larger samples would have provided a
more "accurate" account of faunal diet. The only aspects of this research that I feel
could have been changed, were the sampling procedures used by Crow Canyon. While
their sampling methodology (trenching or opening a unit in the center of structures) is the
common method used, it gives a very limited view of each structure. Opening up the
complete floor of structures may have provided more evidence that could be used to
reconstruct the gendered use of space. A better understanding of the location and
number of mealing features could have provided information on how women's work was
perceived by the community and on variation between households (both important for
interpreting women's status).
This dissertation has attempted to describe and explain patterns shown by major
taxa at Shields Pueblo, to place these changes into both a temporal and regional
context, and ultimately to situate meat procurement within a gendered perspective. The
latter has been accomplished by investigating the ethnographic record (of various
Puebloan as well as other analogue cultures) as well as archaeological evidence from
the Southwest. Through this research, a basic pattern in the sexual division of labor

associated with meat procurement and production has come to light. While this pattern
seems to be consistent across cultures, it cannot be considered direct proof of women's
involvement in meat production at Shields Pueblo. Instead, it is simply a baseline, used
to suggest a gendered interpretation of archaeological data. Obviously, there is room for
future research. The ethnographic record is vast, and by no means was this resource
exhausted during the research phase of this dissertation. First and foremost, it is
recognized that a larger ethnographic survey of meat procurementlproduction is needed
in order to "test" the findings of this dissertation. Furthermore, the lack of access to
artifact databases for this site, and the inability to clearly identify turkey production areas
at Shields Pueblo needs to be addressed. In order to truly understand the cultural
significance of women's roles in turkey production, turkey production areas need to be
identified. This can be accomplished by analyzing other sites in the northern San Juan
in reference to the predicted archaeological signatures of turkey housing areas
(discussed in Chapter 6), namely deposits of turkey droppings; high levels of nitrogen,
phosphorus, and potassium in associated soil deposits; significant deposits of eggshell;
and bones of fetal turkey. The switch from communally hunted large game to turkey
seems to indicate movement away from communal-based meat procurement to
increasingly "domestic" household-driven meat production. The socioeconomic
inferences of turkey production also need to be explored. For example, is there a
relationship between the amount of turkey bone recovered from a household and its
socioeconomic status? Are households with higher frequencies of turkey more
successful or more powerful than households with lower frequencies? Again,
understanding the relationship between turkey production and other aspects of
economic and social life would allow us to better understand the cultural significance of
turkey, of women's labor, and changes to economic and social power in these
communities. Some have argued that large pueblos were able to continue communal

hunting of large game far longer than smaller communities (Muir and Driver 2002).
While this is certainly the case, the faunal assemblages of many of these large, late
Pueblo Ill sites (Sand Canyon Pueblo and Yellow Jacket Pueblo) were dominated by
domestic turkey (Muir 1999; Muir and Driver 2002; Muir and Driver 2003). If household-
based economies became increasingly important in late Pueblo Ill at large central sites,
then does this indicate the beginning of a shift away from highly aggregated sites back to
smaller, dispersed settlements? It has been argued elsewhere (Duff 1998) that migration
decisions were made at the household level. Perhaps the increasing household
independence meant that maintaining large communities (in an over-populated, over-
hunted, and increasingly violent area) was no longer viable, and that depopulation of the
Northern San Juan may very well have been "one household at a time."

REFERENCES CITED
Adams, E.C. 1989 The Case for Conflict during the Late Prehistoric and Protohistoric
Periods in the Western Pueblo Area of the American Southwest. In Cultures in Conflict: Current Archaeological Perspectives. Proceedings of the Twentieth Annual Conference of the Archaeological Association of the University of Calgary. Pp. 103-1 1 1. The University of Calgary, Calgary.
1991 The Origins and Development of the Pueblo Katsina Cult. University of Arizona Press, Tucson.
Adams, E.C., and A.I. Duff (eds.) 2004 Settlement Clusters and the Pueblo IV Period. In The Protohistoric
Pueblo World, A.D. 1275-1600, pp. 3-16. The University of Arizona Press, Tucson.
Adams, K.R., and V.L. Bohrer 1998 Archaeobotanical Indicators of Seasonality: Examples from Arid
Southwestern United States. In Seasonality and Sedentism. Archaeological Perspectives from Old and New World Sites, edited by T.R. Rocek and 0. Bar-Yosef. Peabody Museum Bulletin 6:129-141. Peabody Museum of Archaeology and Ethnology, Harvard University, Cambridge, MA.
Adams, K.R., and K.L. Petersen
Adams, R.K. 1980
Adler, M.A. 1990
Environment. Chapter 2 in Colorado Prehistory: A Context for the Southern Colorado River Basin, edited by W.D. Lipe, M.D. Varien, and R.H. Wilshusen, pp. 14-50. Colorado Council of Professional Archaeologist, Denver.
Salmon Ruin: Site Chronology and Formation Processes. In Investigations at the Salmon Site: the Structure of Chacoan Society in the Northern Southwest, edited by C. Irwin-Williams and P.H. Shelley, pp. 185-272. Eastern New Mexico University, Portales
Communities of Soil and Stone: An Archaeological Investigation of Population Aggregation Among the Mesa Verde Region Anasazi, A. D. 900-1300. Unpublished Ph.D. dissertation, Department of Anthropology, University of Michigan, Ann Arbor.
Adler, M.A., and M.D. Varien 1994 The Changing Face of the Community in the Mesa Verde Region A.D.
1000-1 300. In Proceedings of the Anasazi Symposium, 1991, compiled by A. Hutchinson and J.E. Smith, pp. 83-97. Mesa Verde Museum Association, Mesa Verde National Park, Colorado.

I996 The Prehistoric Pueblo World, A. D. 11 50-1 350. University of Arizona Press, Tucson.
Adriani, N., and A.C. Kruijt 1951 The Bare-speaking Toradja of central Celebes (the East Toradja).
Noord-Hallandsche Uitgeves Maatsschappij, Amsterdam.
Ahlstrom, R.V.N., C.R. Van West, and J.S. Dean 1995 Environmental and Chronological Factors in the Mesa Verde-Northern
Rio Grande Migration. Journal of Anthropological Archaeology 14: 125- 142.
Akins, N.J. 1988 Faunal Assemblages from the South Canal Sites. In Archaeological
lnvestigations in the Bodo Canyon Area, La Plata County, Colorado, prepared by S.L. Fuller, pp. 421-433. UMTRA Archaeological Report, no. 25. Complete Archaeological Service Associates, Cortez, Colorado.
Ambler, R.J. 1989 The Anasazi. Museum of Northern Arizona, Flagstaff.
Ambrose. S.H.
Amorosi, T. 1989
Amyot, J. I976
Anderson, E. 1966
Anderson, M. I978
Andrews A.H. 1973
Isotopic analysis of paleodiets: methodological and interpretive considerations. In lnvestigations of ancient human tissue: chemical analysis in anthropology, edited by M.K. Sandford, pp. 59-130. Gordon and Breach Science Publishers, Philadelphia.
A Postcranial guide to domestic neo-natal and juvenile mammals. British Archaeological Reports International Series 533, Oxford.
Village Ayuttha: social and economic conditions of a rural population in central Thailand. Chulalongkorn University Social Science Institute, Bangkok.
Animal Remains. In Contributions to Mesa Verde Archaeology, 111: Site 866, and the Cultural Sequence at Four Villages in the Far View Group, Mesa Verde National Park, Colorado, by R.H. Lister, pp. 113-1 15. University of Colorado Studies Series in Anthropology, no. 12. University of Colorado Press, Boulder.
Saami ethnoecology: resource management in Norwegian Lapland. University Microfilms International, Ann Arbor.
A survey of the relationship between age and the development of the anterior teeth in cattle. The Vetrinarv Record 92:275-283.

Andrews, P. 1990 Owls, Caves and Fossils. University of Chicago Press, Chicago.
Anell, B. 1969 Running down and driving of game in North America. Studia
Ethnographica Upsaliencia XX. Berlingska Boktrijckeriel, Sweden.
Applegarth, J., and L.H. Feldman 1981 Analysis of Faunal Materials. In A Final Contract Report on the Salvage
Excavation of Archaeological Sites impacted by the Colorado Division of Highways Project No. RS0184, Mancos North, by S.M. Riches and R.W. Biggs, Appendix E.
Armelagos, G.J. 1994 "You are what you eat." In Paleonutrition: the diet and health of
prehistoric Americans, edited by K.D. Sobolik, pp. 235-44. Carbondale: Southern Illinois University at Carbondale Occasional Paper 22, Carbondale.
Auban, J.B., C.M. Barton, and M.P. Ripoll 2001 A Taphonomic Perspective on Neolithic Beginnings: Theory,
Interpretation, and Empirical Data in the Western Mediterranean. Journal of Archaeological Science 28: 597-61 2.
Bagwell, G. 1975 Colorado Mountain College Archaeological Expedition, Excavation Log,
1975. Field notes on file, Crow Canyon Archaeological Center, Cortez.
1976 Colorado Mountain College Archaeological Expedition, Excavation Log, 1976. Field notes on file, Crow Canyon Archaeological Center, Cortez.
1977 Colorado Mountain College Archaeological Expedition, Excavation Log, 1977. Field notes on file, Crow Canyon Archaeological Center, Cortez
Bartram, L.E., and C.W. Marean 1999 Explaining the "Klasies Pattern": Kua Ethnoarchaeology, the
Die Kelders Middle Stone Age Archaeofauna, Long Bone Fragmentation and Carnivore Ravaging. Journal of Archaeological Science 26: 9-29.
Basso, K.H. (ed.) 1971 Western Apache Raiding and Warfare. University of Arizona Press,
Tucson.
Bayham, F.E. I982 A diachronic analysis of prehistoric animal exploitation at Ventana Cave.
Unpublished Ph.D. dissertation, Department of Anthropology, Arizona State University, Tempe, Arizona.
Bayham, F.E., and P. Hatch 1985 Hohokam and Salado animal utilization in the Tonto Basin. In Studies in
the Hohokam and Salado Tonto Basin, edited by G. Rice, pp. 191 -21 0.

Office of Cultural Resource Management. Arizona State University, Tempe, Arizona.
Beaglehole, E. 1937 Notes on Hopi economic life. Yale University Press, New Haven.
Beaglehole, E., and P. Beaglehole 1964 Hopi of the second mesa. Kraus Reprint Corp., New York.
Bender, B. 1989 The roots of inequality. In Domination and Resistance, edited by D. Miller,
M. Rowlands and C. Tilley, pp.83-95. Unwin Hyman, London.
Bennett, W.C. 1935 The Tarahumara: an Indian tribe of northern Mexico. The University of
Chicago Press, Chicago.
Bergfeld, A., R. Bergmann, and P. von Sengbusch
Bertram, J.B. 1991
Botany Online - The lnternet Hypertextbook. The University of Hamburg, Germany. http://www.bioloqie.uni-hamberq.de/b-online/eco/default.htm. Accessed April 29,2006.
An Analysis of Archeofaunal Assemblages, Nonhuman Burials, and Worked Bone Items: Hovenweep Lateral Project. In Archaeological Excavations on the Hovenweep Laterals, by J.N. Morris, pp. 991 -1 104. Four Corners Archaeological Project Report , no 16. Complete Archaeological Service Associates, Cortez, Colorado.
Betancourt, J.L. 1984 Late Quaternary Plant Zonation and Climate in Southeastern Utah. Great
Basin Naturalist 44: 1 -35.
Betancourt, J.L., and N. Biggard 1985 Preliminary Assessment of Late Quaternary Vegetation and Climate of
Southeastern Utah Based on the Analysis of Packrat Middens. BMIIONI- 570, Battelle Memorial Institute, Columbus, OH.
Betancourt, J.L., T.R. Van Devander, and P.S. Martin (editors) 1990 Packrat Middens, the Last 40,000 Years of Biotic Change. University of
Arizona Press, Tucson.
Bettinger, R.L. 1991 Hunter-gatherers: archaeological and evolutionary theory. Plenum Press,
New York.
Billman, B.R., P.M. Lambert, and B.L. Leonard 2000 Cannibalism, Warfare, and Drought in the Mesa Verde Region During the
Twelfth Century A.D. American Antiquity 65(1) 145-1 78

Binford, L.R. 1978 Nunamiut ethnoarchaeology. Academic Press, New York.
1981 Bones: ancient men and modern myths. Academic Press, New York.
Blinman, E. 1989 Potluck in the Protokiva: Ceramics and Ceremonialism in Pueblo I
Villages. In The Architecture of Social Integration in Prehistoric Pueblos, edited by W. Lipe and M. Hegmon, pp. 113-124. Occasional Papers No. I. Crow Canyon Archaeological Center, Cortez, CO.
Bohannan, P. 1968 Tiv Economy. Northwestern University Press, Evanston, IL.
Bokonyi, S. 1970 A new method for the determination of the number of individuals in animal
bone material. American Journal of Archaeology 74: 291 -92.
Bonnichsen, R.
Bradley, B.A. 1988
Brain, C.K. 1969
Bringa, T. 1995
Some operational aspects of human and animal bone alteration. In Mammalian osteo-archaeology: North America, edited by B.M. Gilbert, pp. 9-24. Archaeological Society, University of Missouri, Columbia.
Wallace Ruin Interim Report. Southwest Lore 54(2): 8-33.
The contribution of Namib Desert Hottentots to an understanding of australopithecine bone accumulations. Scientific Papers of the Namib Desert Research Station 39, 13-22.
Some Criteria for the Recognition of Bone Collecting Agencies in African Caves. In Fossils in the Making, edited by A.K. Behrensmyer and A. Hill, pp. 107-1 30. University of Chicago Press, Chicago.
The Hunters of the Hunted. University of Chicago Press, Chicago.
Being Muslim the Bosnian way: identity and community in a central Bosnian village. Princeton University Press, Princeton.
Broughton, J.M. 1994a Declines in Mammalian Foraging Efficiency during the Late Holocene,
San Francisco Bay, California. Journal of Anthropological Archaeology l3:3i'l-4Ol.
1994b Late Holocene Resource Intensification in the Sacramento Valley, California: The Vertebrate Evidence. Journal of Archaeological Science 21 :5OI-514.

Brown, D.E. 1982 Plains and Great Basin Grasslands. In Biotic Communities of the
American Southwest-United States and Mexico, edited by D.E. Brown, pp. 1 15-121. Desert Plants 4(1-4).
Brumbach, H., and R. Jarven~a
Brumfiel E.M. 1991
Bunzel, R. 1938
Butzer, K.W. 1982
Carlson, R.L 1970
Woman the ~un te r : Ethnoarchaeological Lessons from Chipewyan Life-Cycle Dynamics. In Women in Prehistory: North America and Mesoamerica, edited by C. Classen and R.A. Joyce, pp 17- 32. University of Pennsylvania Press, Philadelphia.
Weaving and Cooking: women's Production in Aztec Mexico. In Engendering Archaeology: Women and Prehistory, edited by J.M. Gero and M.W. Conkey, pp.224-254. Basil Blackwell, Oxford.
The Economic Organization of Primitive Peoples. In General Anthropology, edited by F. Boas. Heath, Washington D.C.
Archaeology as human ecology. Cambridge University Press, Cambridge.
White Mountain Redware: A Pottery Tradition of East-Central Arizona and Western New Mexico. Anthropological Papers of the University of Arizona, no. 19. University of Arizona Press, Tucson.
Carrara, P.E., D.A. Trimble, and M. Rubin 1991 Holocene Timberline Fluctuation in the Northern San Juan Mountains,
Colorado, U.S.A., as Indicated by Radiocarbon-dated Conifer Wood. Arctic and Alpine Research 23:233-246.
Casteel, R.W. 1978 Faunal assemblages and the "weigmethod" or weight method. Journal of
Field Archaeology 5: 7 1-77.
Chaplin, R.E. 1971 The study of animal bone from archaeological sites. Seminar Press,
London.
Chase, P.G., and R.M. Hagman 1987 Minimum number of individuals and its alternatives: a probability theory
perspective. Ossa 13, 75-86.
Childs, G.M. 1949 Umbundu kinship & character: being a description of social structure and
individual development of the Ovimbundu of Angola with observations concerning the bearing on the enterprises of Christian Missions of certain phases of life and culture description. International African Institute, Oxford University Press, London.

Chisholm, B., and R.G. Matson 1994 Carbon and Nitrogen Isotopic Evidence on Basketmaker II Diet at Cedar
Mesa, Utah. Kiva 60:239-255.
Claassen,C. 1991 Gender, Shellfishing and the Shell Mound Archaic. In Engendering
Archaeology: Women and Prehistory, edited by J.M. Gero and M.W. Conkey, pp. 276-300. Blackwell, Oxford.
Clark J,. and K.K. Kietzke 1967 Palaeoecology of the Lower Nodule Zone, Brule Formation, in the Big
Badlands of South Dakota. In Oligocene sedimentation, stratigraphy and palaeoclimatology in the Big Badlands of South Dakota, edited by J. Clark, J.R. Beerbower, and K.K. Kietzke, pp. 11 1-137. Fieldianna Geology Memoir 5.
Cohen, R. 1967 The Kanuri of Bornu. Holt, Reinhart and Winstonlnc., New York.
Conkey, M. W. 1991 Contexts of Action, Contexts for Power: Material Culture and
Gender in the Magdalenian. In Engendering Archaeology: Women and Prehistory, edited by J.M. Gero and M.W. Conkey, pp. 57-92. Basil Blacwell, Oxford.
Conkey, M.W., and J.M. Gero 1991 Tensions, Pluralities, and Engendering Archaeology An
Introduction to Women and Prehistory. In Engendering Archaeology: Women and Prehistory, edited by J.M Gero and M.W. Conkey, pp. 3-30. Basil Blackwell, Oxford.
Conkey, M.W., and J. Spector 1984 Archaeology and the Study of Gender. Advances in Archaeological
Method and Theory 7: 1-38.
Conklin, B.A. 1995 "Thus Are Our Bodies, Thus, Was Our Custom": Mortuary Cannibalism in
an Amazonian Society. American Ethnologist 22(1): 75-1 01.
Connolly, M. 1992 The Goodman Point Historic Land-Use Study. In The Sand Canyon
Archaeological Project: A Progress Report, edited by W.D. Lipe, pp. 33- 44. Occasional Paper No. 2. Crow Canyon Archaeological Center, Cortez, Colorado.
Cordell, L.S. 1984 Prehistory of the Southwest. Academic Press, New York.

Cordell, L.S., and G.J. Gumerman 1989 Cultural Interaction in the Prehistoric Southwest. In Dynamics of
Southwestern Prehistory, edited by L.S. Cordell and G.J. Gumerman, pp. 1-1 8. Smithsonian Institution Press, Washington D.C.
Crabtree, P.J.
Crotty, H.K. 2001
Cucchiari, S. 1986
Cushing, F.H 1882
Zooarchaeology and Complex Societies: some uses for faunal analysis for the study of trade, social status, and ethnicity. In Archaeological method and theory, vol. 2, edited by M.B. Schiffer, pp. 155-205. University of Arizona Press, Tucson.
Shields, Shield Bearers, and Warfare Imagery in Anasazi Art, 1200-1500. In Deadly Landscapes: Case Studies in Prehistoric Southwestern Warfare, edited by G.E. Rice and S.A. LeBlanc, pp. 65-83. The University of Utah Press, Salt Lake City.
The gender revolution and the transition from bisexual horde to patrilocal band: the origins of gender hierarchy. In Sexual meanings, edited by S. Ortner and H. Whitehead, pp. 31-79. Cambridge University Press, Cambridge.
My Adventures in Zuni. Reprint, American West Publishing Company, Palo Alto, CA. Reprint 1941, The Peripatetic Press, Santa Fe.
Zuni: Selected Writings of Frank Hamilton Cushing. Edited by J. Green. University of Nebraska Press, Lincoln.
Cushing, F.N., J.W. Fewkes, and E.C. Parsons 1922 Contributions to Hopi History. American Anthropologist v. 24: 253-298.
D' Andrea, A.C., and R.M. Gotthardt 1984 Predator and Scavenger Modification of Recent Equid Skeletal
Assemblages. Arctic 37:276-283.
Davis, O.K. 1996 The Impact of Climatic Change on Available Moisture in Arid Lands:
Examples From the American Southwest. In Diachronic Climatic Impacts on Water Resources, edited by A.N. Angelakis and A.S. Issar, pp. 283- 300. NATO AS1 Series, Vol. 1 36. Springer-Verlag, Berlin.
Davis, S.J.M. 1987 The archaeology of animals. Batsford, London.
Davis-Kimball, J. 1997 Warrior Women of the Eurasian Steppes. Archaeology
JanuarylFebruary: 44-48.

Dean, J.S. 1996 Demography, Environment and Subsistence Stress. In Evolving
Complexity and Environmental Risk in the Prehistoric Southwest, edited by J.A. Tainter and 8.8. Tainter, pp. 25-56. Proceedings Vol. 24, Santa Fe Institute Studies in the Sciences of Complexity. Addison-Weley, Reading, MA.
Dean, J.S., and W. J. Robinson 1977 Dendroclimatic Variability in the American Southwest, A. D. 680 to 1970.
Final report to the National Park Service, Department of the Interior. Laboratory of Tree-Ring Research, University of Arizona, Tucson.
1 978 Expanded Tree-Ring Chronologies for the Southwestern United States (Chronology Series 111). Laboratory of Tree-Ring Research, University of Arizona, Tucson.
Dean, J.S., W.H. Doelle, and J.D. Orcutt 1994 Adaptive Stress: Environment and Demography. In Themes in Southwest
Prehistory, edited by G.J. Gumerman, pp. 53-86. School of American Research Press, Santa Fe, New Mexico.
Di Peso, C.G. (editor) 1974 Casas Grandes: a fallen trading center of the Gran Chichimeca. Amerind
Foundation, Dragoon, and Norhtland Press, Flagstaff, AZ.
Discovery Farms 2006 College of Agriculture & Life Sciences, University of Wisconsin Extension.
http://www..discoven/farms.or~. Accessed December 27,2005.
Dodson, P., and D. Wexlar 1979 Taphonomic Investigations of Owl Pellets. Paleobiology 5275-284.
Dohm, K.M. 1994 The Search for Anasazi Village Origins: Basketmaker II Dwelling
Aggregation on Cedar Mesa. Kiva 60: 257-276.
Douglas, A.E. 1929
1935
Drager, D.L. 1976
The secret of the Southwest Solved by Talkative Tree Rings. National Geographic 56: 736-770.
Dating Pueblo Bonito and Other Ruins of the Southwest. Contributed Technical Papers, Pueblo Bonito Series, No.1. National Geographic Society, Washington D.C.
Anasazi population estimates with the aid of data derived from photogrammetric maps. In Remote sensing experiments in cultural resource studies. Assembled by T.R. Lyons. Reports of the Chaco Center, no. 1, pp. 157-71. Division of Culture Research, National Park Service, Albuquerque.

Drennan, R.D.
Driver, J.C. 1985
Statistics for Archaeologists: A Commonsense Approach. Plenum Press, New York.
Zooarchaeology of Six Prehistoric Sites in the Sierra Blanc Region, New Mexico. University of Michigan, Museum of Anthropology, Technical Report No. 17, Ann Arbor.
Social Complexity and Hunting Systems in Southwestern Colorado. In Debating Complexity: proceedings of the twenty-sixth annual Chocmool Conference, edited by D.A. Meyer, P.C. Dawson, and D.T. Hanna, pp. 364-374. The Archaeological Association of the University of Calgary, Calgary.
Archaeology and Social Organization in Non-State Societies. Anthropologica No. 25-26: 79-84.
Crow Canyon Archaeological Center Manual For Description of Vertebrate Remains 5th edition. Crow Canyon Archaeological Center, Cortez.
Hunting strategies and horticultural communities in southeastern New Mexico. In Animal Bones, Human Societies. Edited by P. Rowley-Conway, pp.115-123. Oxbow Books, Oxford.
Faunal Variation and Change in the Northern San Juan Region. In Seeking the Center Place: Archaeology and Ancient Communities in the Mesa Verde Region. Edited by M.D. Varien and R.H. Wilshusen, pp. 143- 160. The University of Utah Press, Salt Lake City.
Driver, J. C., M.J. Brand, L. Lester, and N. D. Munro 1999 Faunal Studies. In The Sand Canyon Archaeological Project: Site
Testing. Crow Canyon Archaeological Center, Cortez.
Dubisch, J. 1986 Culture enters through the kitchen: women, food, and social boundaries in
rural Greece. In Gender and power in Rural Greece, edited by J. Dubisch, pp. 195-214. Princeton University Press, Princeton.
Duff, A.I., and S.C. Ryan 1999 The Shields Pueblo Research Project: Annual Report, 1999 Fieldwork.
Crow Canyon Archaeological Center, Cortez.
2000 The Shields Pueblo Research Project: Annual Report, 2000 Fieldwork. Crow Canyon Archaeological Center, Cortez.
Duff, A.I., and R.H. Wilshusen 2000 Prehistoric Population Dynamics in the Northern San Juan Region, A.D.
950-1 300. Kiva 66:167-189.

Durkheim, E. 1964 The Division of Labor in Society, translated by G. Simpson. Free Press,
New York.
Dykeman, D.D.
Dyson, W. 2002
Eddy, F.W. 1972
Excavations at 5MT8377, an isolated Pueblo I1 Pit Structure in Montezuma County, Cory of human modification lie within the ranges of frequencies from Woods Canyon Pueblo, San Juan County Museum Association, Farmington, NM.
The Ovimbundu of Angola. Originally published in 1934. Human Relations Area Files. Computer Files, New Haven.
Culture; Ecology and the Prehistory of the Navajo Reservoir District, New Mexico and Colorado. American Antiquity 39: 75-84.
Efremov, J.A. 1940 Taphonomy: new branch of palaeontology. Pan-American Geologist 74:
8 1-93.
Ellis, F.H. 1959 An outline of Laguna Pueblo history and social organization.
Southwestern Journal of Anthropology 15: 325-347.
Elmendorf, M.L.
Emslie, S.D. I978
Euler, R.C. I988
Fairley, H.C. 1989
Nine Mayan Women: a village faces change. Schenkman Publishing Co., Cambridge, Mass.
Vertebrate Fauna from Site 5MTUMR 2837, Mancos Canyon, Colorado. In Excavations at Site 5MTUMR 2837, Mancos Canyon, Ute Mountain
Homelands, Colorado. Report of file, Bureau of Indian Affairs, Washington D.C.
Demography and cultural dynamics on the Colorado plateaus. In The Anasazi in a changing environment, edited by G.J. Gumerman, pp. 192- 229. Cambridge University Press, Cambridge.
Culture History. In Man, Models and Management: An Overview of the Archaeology of the Arizona Strip and the Management of Its Cultural Resources, edited by J. H. Altshul and H.C, Fairley, pp. 85-152. Statistical Research, Tucson.

Fallers, L.A. 1964 Social Stratification in traditional Buganda. In The King's men: leadership
and status in Buganda on the eve of independence. Oxford University Press, London.
Farmer, M.F. 1957 A Suggested Typology of Defensive Systems of the Southwest.
Southwestern Journal of Anthropology 1 3: 249-266.
Feil, D.K. 1984 Ways of Exchange: The Enga Tee of Papua New Guinea. University of
Queensland Press, St. Lucia.
Fetterman, J., and L. Honeycutt 1982 Testing and Excavation Report, MAPCO's Rocky Mountain Liquid
Hydrocarbons Pipeline, Southwest Colorado. Report on file, Anasazi Heritage Center, Dolores, Colorado.
Fieller, N.R.J., and A. Turner 1982 Number estimation in vertebrate samples. Journal of Archaeological
Science 9(1), pp. 49-62.
Fish, P.R., S.K. Fish, G.J. Gumerman, and J.J Reid 1994 Toward an Explanation for Southwestern "Depopulation's ." In Themes is
Southwest Prehistory, edited by G.J. Gumerman, pp 135-1 64. School of American Research Press, Santa Fe, New Mexico.
Fisher, J.W., Jr. 1995 Bone surface modifications in zooarchaeology. Journal of Archaeological
Method and Theory 2(1): 7-68.
Frison, G.C., and C.A. Reher
Garrett, H. 2003
Geib, P.R. 1990
Gero, J.M. 1991
Age determination of buffalo by teeth eruption and wear. In Glenrock Buffalo Jump, edited by G.C. Frison. Plains Anthropologist Memoirs, pp. 46-50.
Natural Organic Gardening and Living. http://www.dirtdoctor.com. Accessed December 27, 2005.
A Basketmaker Wooden Tool Cache from Lower Glen Canyon. Kiva 55: 265-278.
Genderlithics: Women's Roles in Stone Tool Production. In Engendering Archaeology: Women and Prehistory, edited by J.M Gero and M.W. Conkey, pp. 163-193. Basil Blackwell, Oxford.
Gifford-Gonzalez, D. 1993 Gaps in the Zooarchaeological Analysis of Butchery: Is Gender

an Issue? In From Bones to Behavior: Ethnoarchaeological and Experimental Contributions to the Interpretations of Faunal Remains, edited by J. Hudson, pp. 181-199. Southern Illinois University at Carbondale Center for Archaeological Investigations, Carbondale.
Gilbert, A.S., B.H. Singer, and D. Perkins jr. 1982 Quantification experiments in computer simulated faunal collections.
Ossa 8, 79-94.
Gilchrist, R. 1999 Gender and Archaeology: Contesting the Past. Routledge, London.
Gilliland, M.K. 1986 The maintenance of family values in a Yugoslav town. University of
Michigan Microfilms, Ann Arbor.
Glacken, C.J. 1953 Studies of Okinawan village life. Pacific Science Board, National
Research Council, Washington D.C.
Gladwin, H.S. 1957 A History of the Ancient Southwest. Bond, Wheelwright Co., Portland,
ME.
Gnabasik, V.R. 1981 Faunal Utilization by the Pueblo Indians. Unpublished M.A. thesis,
Department of Anthropology, Eastern New Mexico University.
Gomes, E.H. 191 1 Seventeen years among the Sea Dyaks of Borneo: a record of intimate
association with the natives of the Bornean jungles. Seeley and Co. Ltd., London.
Goody, J. 1982 Cooking, Cuisine, and Class. Cambridge University Press, Cambridge.
Grayson, D.K. 1979 On the quantification of vertebrate archaeofaunas. In Advances in
Archaeological Method and Theory vol. 2, edited by M.B. Schiffer, pp. 199-237. Academic Press, New York.
1984 Quantitative Zooarchaeology. Academic Press, London.
1988 Danger Cave, Last Supper Cave and Hanging Rock Shelter: the faunas. American Museum of Natural History Anthropological Papers 66(1): 1- 130.
Gullick, C.J.M.R. 1976 Exiled from St. Vincent: the development of Black Carib culture in Central
America up to 1945. Progress Press, Malta.

Gumerman, G.J. (editor) 1988 The Anasazi in a Changing Environment. The School of American
Research and Cambridge University Press, Cambridge.
Gumerman, G.J. 1994 Patterns and Perturbations in Southwest Prehistory. In Themes in
Southwest Prehistory, edited by G.J Gumerman, pp 3-1 0. School of American Research Press, Santa Fe, New Mexico.
Gumerman, G.J., and J.S. Dean 1989 Prehistoric Cooperation and Competition in the Western Anasazi Area. In
Dynamics of Southwest Prehistory, edited by L.S. Cordell and G.J. Gumerman, pp. 99-148. Smithsonian Institution Press, Washington D.C.
Haas, J., and W. Creamer 1996 The Role of Warfare in the Pueblo Ill Period. In The Prehistoric Pueblo
World, A.D. 1150-1350, edited by M.A. Adler, pp. 205-213. University of Arizona Press, Tucson.
Hack, J.T. 1942 The Changing Physical Environment of the Hopi Indians of Arizona.
Papers of the Peabody Museum of American Archaeology and Ethnology 35(1). Harvard University, Cambridge Massachusetts.
Halperin, R.H. 1994 Cultural Economies: Past and Present. University of Texas
Press, Austin.
Hammack, N.S., and R. Walkenhorst 1991 Facility Site and Ancillary Construction Features. In Cultural Resource
Monitoring and Testing, Sand Canyon C02 Project, Montezuma County, edited by N.S, Hammack, pp. 25-55. Complete Archaeological Service Associates, Cortez, CO.
Hampton, O.W. 1995 Rock Quarries and the Manufacture, Trade, and Use of Stone Tools and
Symbolic Stones in the Central Highlands of lrian Jaya, Indonesia: Ethnographic Perspectives. PhD thesis, Texas A and M University.
Hanson, C.B. 1980 Fluvial Taphonomic Processes: Models and Experiments. In Fossils in
the Making, edited by A.K. Behrensmeyer and A.P. Hill, pp. 156-181. University of Chicago Press, Chicago.
Harris, M. 1971 Culture, Man, and Nature: An Introduction to General
Anthropology. Thomas Crowell Company, New York.
Hastorf, C.A. 1991 Gender, Space, and Food in Prehistory. In Engendering

Archaeology: Women and Prehistory, edited by J.M Gero and M.W. Conkey, pp. 132-1 59. Basil Blackwell, Oxford.
Hayden, B. 1995 Pathways to Power: Principles for Creating Socioeconomic Inequalities.
In Foundations of Social Inequality, edited by T. Price and G. Feinman, pp. 15-86. Plenum Press, New York.
Haynes, G. 1983 A guide for differentiating mammalian carnivore taxa responsible for gnaw
damage to herbivore long bones. Paleobiology 9:164-172.
Hayes, A.C. 1981 A survey of Chaco Canyon archaeology. In Archaeological surveys of
Chaco Canyon, New Mexico. By A.C. Hayes, D.M. Brugge, and W.L. Judge. National Park Service Archaeological Series, no. 17A, Chaco Canyon Studies, pp. 1-68. Washington D.C.
1984 Two Raven House, Mesa Verde National Park, Colorado. Manuscript on file, Mesa Verde National Park Research Center, Colorado.
Hayes, A.C., and C.C. Chapell 1962 A Copper Bell from Southwestern Colorado. Plateau 35: 53-56.
Haynes, G. 1980 Evidence of Carnivore Gnawing on Pleistocene and Recent Mammalian
Bones. Paleobiology 6: 341 -351.
1983 Frequencies of spiral and green-bone fractures on ungulate limb bones in modern surface assemblages. American Antiquity 48: 102-1 14.
Hays-Gilpin, K., and D.S. Whitley 1997 Introduction. Reader in Gender Archaeology, edited by K.
Hays-Gilpin and D.S. Whitley, pp. 3-10. Routledge, London.
Henderson, J., and J.P. Harrington 191 4 Ethnozoology of the Tewa Indians. Bureau of American Ethnology,
Bulletin 56. Washington D.C.
Hendon, J.A. 1997 Women's Work, Women's Space, and Women's Status Among
The Classic Period Maya Elite of the Copan Valley, Honduras. In Women in Prehistory: North America and Mesoamerica, edited by C. Classen and R.A. Joyce, pp. 33-46. University of Pennsylvania Press, Philadelphia.
Herzfeld, M. 1985 The poetics of manhood: contest and identity in a Cretan mountain
village. Princeton University Press, Princeton.
Hesse, B., and P. Wapnish 1985 Animal bone archaeology. Taraxacum Press, Washington D.C.

Hewitt, E.L. 1906
Hewitt, N.J. I983
Hevly, R.H. 1981
Antiquities of the Jemz Plateau, New Mexico. Bureau of American Ethnology Bulletin 32. Smithsonian Institution, Washington D.C.
Excavation of Sagehill Hamlet (Site 5MT2198), a Basketmaker Ill-Pueblo I Habitation Site. In Dolores Archaeological Program: Field Investigation and Analysis-7978, compiled by D.A. Breternitz, pp. 103-138. U.S. Department of the Interior, Bureau of Reclamation, Engineering and Research Center, Denver.
Pollen Production, Transport and Preservation: Potentials and Limitations in Archaeological Palynology. Journal of Ethnobiology 1:39-54.
Prehistoric Vegetation and Paleoclimates on the Colorado Plateaus. In The Anasazi in a Changing Environment, edited by G. J. Gumerman, pp. 92-1 18. School of American Research Advanced Seminar Series. Cambridge University Press, Cambridge.
Hevly, R.H., R.E. Kelly, G. Anderson, and S.J. Olsen 1979 Comparative Effects of Climatic Change, Cultural Impact and Volcanism
in the Paleoecology of Flagstaff, Arizona, A.D. 900-1300. In Volcanic Activity and Human Ecology, edited by P.D. Sheets and D.K. Greyson, pp. 487-523. Academic Press, New York.
Hill, E. 2000 The Contextual Analysis of Animal Interments and Ritual Practice in
Southwestern North America. Kiva 65: 361-398.
Hodge, H.C. 1877 Arizona as It Is; or, The Coming Country. Hurd and Houghton, New York.
Hoffman, R., and C. Hays I987 The Eastern Wood Rat (Neotoma floridana) as a Taphonomic Factor in
Archaeological Sites. Journal of Archaeological Science 14:325-337.
Howell, T.L. 1996 Identifying leaders at Hawikku. Kiva 62: 61-82.
2001 Foundations of political power in ancestral Zuni society. In Ancient burial practices in the American Southwest, edited by D.R. Mitchell and J. L. Brunson-Hadley, pp. 149-166. University of New Mexico Press, Albuquerque.
Howell, T.L., and K.W. Kintigh 1996 Archaeological identification of kin groups using mortuary and biological
data: an example from the American Southwest. American Antiquity 61: 537-554.

Huckleberry, G. A., and B.R. Billman 1998 Floodwater Farming, Discontinuous Ephemeral Streams, and Puebloan
Depopulation in Southwestern Colorado. American Antiquity 63(4):595- 61 6.
Hughs, M.K. and H.F. Diaz (editors) 1994 The Medieval Warm Period. Kluwer, Dordrect, Netherlands.
Hurlbut, S.A. 2001 The Taphonomy of Cannibalism: A Review of Anthropogenic Bone
Modification in the American Southwest. International Journal of Osteoarchaeology 10: 4-26.
Janetski, J.C. 1994 Recent Transitions in Eastern Great Basin Prehistory: The
Archaeological Record. In Across the West: Human Population Movement and the Expansion of the Numa, edited by D.B. Madsen and D. Rhode, pp. 157-178. University of Utah Press, Salt Lake City.
Jennings, J.D. 1980 Cowboy Cave. Anthropological Papers No. 104. University of Utah, Salt
Lake City.
Jewett,R. 1989 Distance, Interaction, and Complexity: A Pan-Regional Comparison of the
Spatial Organization of Fourtenth-Century Settlement Clusters in the American Southwest. In Sociopolitical Structure of Prehistoric Southwestern Societies, edited by S. Upham, K. Lightfoot, and R. Jewett. Westview Press, Boulder CO.
Joyce, R.A. 1997 Women in the Ancient Americas: Archaeologists, Gender, and
the Making of Prehistory, In Women in Prehistory: North America and Mesoamerica, edited by C. Classen and R.A. Joyce, pp.1-14. University of Pennsylvania Press, Philadelphia.
Joyce, R.A., and C.Classen Women in the Ancient Americas: Archaeologists, ender, and the Making of Prehistory. In Women in Prehistory: North America and Mesoamerica, edited by C. Classen and R.A. Joyce, pp. 1-14. University of Pennsylvania Press, Philidelphia.
Judge, W.J. 1989 Chaco: Current Views of Prehistory and the Regional System. In Chaco and Hohokam: Prehistoric Regional Systems in the American Southwest, edited by P.L. Crown and J.W. Judge. pp. 1 1-30. School of American Research, Santa Fe.
Kantner, J. 2004 Ancient Puebloan Southwest. Cambridge University Press, Cambridge.

Katzenberg, M.A., and J.H. Kelley 1991 Stable Isotope Analysis of Prehistoric Bone from the Sierra Blanca
Region of New Mexico. In Mogollon V, pp.207-219. COAS Publishing, Las Cruces, NM.
Keeley, L.H. 1996 War Before Civilization: The Myth of the Peaceful Savage. Oxford
University Press, New York
Keesing R.M. 1975 Kin Groups and Social Structure. Harcourt and Brace
Jovanovich College Publishers, Austin.
Kennedy, J.G. 1 978 Tarahumara of the Sierra Madre: beer, ecology, and social organization.
A.H.M. Publishing Corporation, Arlington Heights, Illinois.
Kent, S. 1989a Site 5MT1786, a Final Report of Excavations and Analysis. Report on
file, Anasazi Heritage Center, Dolores, Colorado.
1989b Cross-cultural perceptions of farmers as hunters and the value of meat. In Farmers as Hunters: The Implications of sedentism, edited by S. Kent, pp. 1-1 7. Cambridge University Press, Cambridge.
1998 Invisible Gender - Invisible Foragers: Southern African Hunter- Gatherer Spatial Patterning and the Archaeological Record. In Gender in African Prehistory, edited by S. Kent, pp. 39-67. Alta Mira Press, Walnut Creek.
Kidder, A.V. 1924 An Introduction to the Study of Southwestern Archaeology, with a
Preliminary Account of the Excavations at Pecos. Papers of the Phillips Academy Southwestern Expedition 1. Yale University Press, New Haven.
1927 Southwestern Archaeological Conference. Science 66: 489-491.
Kintz, E. 1990 Life under the tropical canopy: tradition and change among the Yucatec
Maya. Holt, Rinehart and Winston, Fort Worth, TX.
Klein, R.G., and K. Cruz-Uribe 1984 The Analysis of Animal Bones from Archaeological Sites. Prehistoric
Archaeology and Ecology Series, edited by K.W. Butzer and L.G. Freeman. University of Chicago Press, Chicago.
Koch, C.P. 1990 Bone Breakage, differential preservation and "Theropithecus" butchery at
Olorgesailie, Kenya. In Tempus: archaeology and material culture studies in anthropology, pp. 158-66.

Kohler, T.A. 1992 Prehistoric impact on the environment in the upland southwest.
Population and Environment 13:255-268.
Kohler, T.A., and M. Matthews 1988 Long-term Anasazi land use and forest reduction: A case study from
southwestern Colorado. American Antiquity 53:537-564.
Kohler, T.A., and C.R. Van West I996 The Calculus of Self-Interest in the Development of Cooperation:
Sociopolitical Development and Risk Among the Northern Anasazi. In Evolving Complexity and Environmental Risk in the Prehistoric Southwest, edited by J. Tainter and B. Tainter, pp. 169-196. Santa Fe Institute Studies in the Sciences of Complexity 24. Addison-Wesley, Reading MA.
Korth, W.W. 1979 Taphonomy of Microvertebrate Fossil Assemblages. Annals of the
Carnegie Museum 48: 235-285.
Kremensek, S. 1983 On the fringe of the town. Urban life in Mediterranean Europe:
anthropologicalperspectives, edited by M. Kenny and D.I. Kertzer, pp. 282-289. University of Illinois Press, Urbana.
Kreutz, K.J, P.A. Mayewski, L.D. Meeker, M.S. Twickler, S.I. Whitlow, and 1.1. Pittalwala 1997 Bipolar Changes in Atmospheric Circulation During the Little Ice Age.
Science 277: 1294-1 296.
Kuckelman, K.A. 1997 Preliminary Results of Testing and Report of 1997 Excavations at Yellow
Jacket Pueblo (Site 5Mt5). Crow Canyon Archaeological Center, Cortez, CO. Electronic publication. Htp://www.crowcanyon.orglResearchReportsNJP/Reprt.htm. Accessed June 15,2005.
Kusmer, K.D. 1986 Microvertebrate Taphonomy in Archaeological Sites: An Examination of
Owl Deposition and the Taphonomy of Small Mammals from Sentinel Cave, Oregon. Unpublished M.A. Thesis, Department of Archaeology. Simon Fraser University, Burnaby.
Lambert, P.M., B.R. Billman, and B.L. Leonard 2001 Explaining Variability in Mutilated Human Bone Assemblages from the
American Southwest: A Case Study from the Southern Piedmont of Sleeping Ute Mountain, Colorado. International Journal of Osteoarchaeology 1 0: 49-64.

Lange, C.H. 1959 Cochiti: a New Mexico pueblo, past and present. University of Texas
Press, Austin.
Larson, D.O. and J. Michaelson 1990 Impacts of Climatic Variability and Population Growth on Virgin Branch
Anasazi Cultural Developments. American Antiquity 55: 227-249.
Larson, D.O., H. Neff, D. Graybill, J. Michaelson and E. Ambos 1996 Risk, Climatic Variability, and the Study of Southwestern Prehistory: An
Evolutionary Perspective. American Antiquity 61 (2):217-241.
Lee, R.B., and I. Devore 1968 Man the Hunter. Aldine, Chicago.
LeBlanc, S.A. 1999 Prehistoric Warfare In the American Southwest. University of Utah Press,
Salt Lake City.
Leonard, R.D., and H.E. Reed 1993 Population Aggregation in the Prehistoric American Southwest: A
Selectionist Model. American Antiquity 58 (4): 648-661.
Lepofsky, D. 1986 Preliminary Analysis of Floatation Samples From the Turkey Pen, Cedar
Mesa, Utah. Ms on file, Laboratory of Archaeology, University of British Colombia, Vancouver.
Lightfoot, R.R., and Etzkorn 1993 Introduction. In The Duckfoot Site Volume 1: Descriptive Archaeology.
Occasional Papers of the Crow Canyon Archaeological Center, edited by W.D. Lipe. Crow Canyon Archaeological Center: Cortez, CO.
Lightfoot, R.R., and K.A. Kuckelman 2000 A Case of Warfare in the Mesa Verde Region. In Deadly Landscapes:
Case Studies in Prehistoric Southwestern Warfare, edited by G.E. Rice and S.A. LeBlanc, pp. 51-64 The University of Utah Press. Salt Lake City.
Lipe, W. D. 1992 Summary and Concluding Comments. In The Sand Canyon
Archaeological Project: a Progress Report, edited by W.D. Lipe, pp. 1 2 1 - 133. Occasional Paper No. 2, Cortez.
The Depopulation of the Northern San Juan: Conditions in the Turbulent 1200's. Journal of Anthropological Archaeology 14: 143-1 69.
Chapter 5. Basketmaker 11 (1000 B.C. - A.D. 500). In Colorado Prehistory: A Context for the Southern Colorado River Basin, edited by W.D. Lipe, M.D. Varien, and R.H. Wilshusen. C.C.P.A., Denver.

Lipe, W.D., and M.D. Varien 1999a Chapter 8. Pueblo II (A.D. 900-1 150). In Colorado Prehistory: A Context
for the Southern Colorado River Basin, edited by W.D. Lipe, M.D. Varien, and R.H. Wilshusen, pp. 242-289. C.C.P.A., Denver.
1999b Chapter 9. Pueblo Ill (A.D. 1150-1300). In Colorado Prehistory: A Context for the Southern Colorado River Basin, edited by W.D. Lipe, M.D. Varien, and R.H. Wilshusen, pp. 290-352. C.C.P.A., Denver.
Lipe, W.D., M.D. Varien, and R.H. Wilshusen (eds.) 1999 Colorado Prehistory: A Context for the Southern Colorado River Basin.
C.C.P.A., Denver.
Lister, R.H. 1978 Mesoamerican Influence at Chaco Canyon, New Mexico. In Across the
Chichimec Sea, edited by C.L. Riley and B.C. Hedrick. Southern Illinois University Press, Carbondale.
Lockwood, W.G.
Lyman, R.L. I982
European Moslems: economy and ethnicity in western Bosnia. Academic Press, New York.
Archaeofauna and subsistence studies. In Advances in Archaeological Method and Theory 5, edited by M.B. Schiffer, pp. 331-393. Academic Press, New York.
Broken Bones, Bone Expediency Tools, and Bone Pseudotools: Lessons from the Blast Zone around Mount St. Helens, Washington. American Antiquity 49: 3 15-333.
Was there a Last Supper at the Last Supper Cave? In Danger Cave, Last Supper Cave, and Hanging Rock Shelter: The Faunas, edited by D.K. Greyson, pp. 81 -1 04. American Museum of Natural History Anthropological Papers 66(1).
Vertebrate Taphonomy. Cambridge University Press, Cambridge.
Maher, L.J., Jr. 1961 Pollen Analysis and Postglacial Vegetation History in the Animas Valley
Region, Southern San Juan Mountains. Ph. D. dissertation, University of Minnesota, Minneapolis. University Microfilms, Ann Arbor.
Mahoney, N.M., M.A. Adler, and J.W. Kendrick 2000 The changing scale and configuration of Mesa Verde communities. Kiva
66:67-90.
Main, M.B., and R. Owens 1995 Estimating mule deer age from measurements of incisor wear. Northwest
Naturalist 76: 130-1 32.

Marean, C.W., L.M. Spencer, R.J. Blumenschine, and S.D. Capaldo 1992 Captive Hyena Bone Choice and Destruction, the Schlep Effect and
Olduvai Archeofaunas. Journal of Archaeological Science 19: 101 -1 2 1.
Martin, D.L., and N.J. Akins
Mathien, F.J. 1981
Matson, R.G. 1991
Unequal treatment in life as in death: trauma and mortuary behavior at La Plata (A.D. 1000-1 300). In Troubled times: violence and warfare in the past, edited by D.L. Martin and D.W. Frayer, pp. 45-76. Gordon and Breach Publishers, Amsterdam.
Economic Exchange Systems in the San Juan Basin. Unpublished Ph.D. Dissertation, University of New Mexico. Manuscript, Chaco Center, Albuquerque.
The Origins of Southwestern Agriculture. University of Arizona Press, Tucson.
Matson, R.G., and B. Chisholm 1991 Basketmaker II Subsistence: Carbon Isotope and other Dietary Indicators
from Cedar Mesa, Utah. American Antiquity 56(3) 444-459.
Matson, R.G., W.D. Lipe, and W.R. Haas 1988 Adaptational Communities and Occupation Discontinuities: The Cedar
Mesa Anasazi. The Journal of Field Archaeology 15: 245-64.
Maxwell, T.D. 2002 Casas Grandes Region: Prehistoric Life in the Chihuahuan Desert. El
Palacio 1 O7(3): 1 2-1 9.
Mc Brearty, S., and M. Moniz 1991 Prostitutes or Providers? Hunting, Tool Use, and Sex Roles in
Earliest Homo. In The Archaeology of Gender, edited by D. Walde and N.D. Willows, pp. 71-82. Archaeological Association of the University of Calgary, Calgary.
Mc Cormack, A.P. 1964 Khasis. In Ethnic groups of mainland Southeast Asia, edited by F.M.
Lebar, G.C. Hickey, and J.K Musgrave, pp. 105-1 12. Human Relations Area Files Press, New Haven.
Mc Kusick, C.R. 1980 Avian Remains. In Long House Mesa Verde National Park, Colorado,
edited by G.J. Cattanach, pp. 391-398. Publications in Archaeology 7H. National Park Service, United States Department of the Interior, Washington D.C.
Meggitt, M.J. 1977 Blood is Their Argument. Mayfield, Palo Alto, CA.

Mera, H.P. 1935 Ceramic Clues to the Prehistory of North Central New Mexico.
Laboratory of Anthropology Technical Series Bulletin no. 8. Santa Fe.
1940 Population Changes in the Rio Grande Glaze Paint Area. Laboratory of Anthropology Technical Series Bulletin no. 9. Santa Fe.
Mick O'Hara, L.S. 1994 Nutritional Stability and Changing Faunal Resource Use in La Plata
Valley Prehistory. Unpublished Ph.D. dissertation, Department of Anthropology University of New Mexico, Albuquerque.
Minnis, P.E., and C.L. Redman (eds.) 1 990 Perspectives on Southwester Prehistory. Westview Press, Boulder
Minnis, P.E., M.E. Whalen, J.H. Kelley, and J.D. Stewart 1993 Prehistoric Macaw Breeding in the North American Southwest. American
Antiquity 58(2):270-276.
Mobley-Tanaka, J.L. 1997 Gender and Ritual Space During the Pithouse to Pueblo Transition:
Subterranean Mealing Rooms in the North American Southwest. American Antiquity 62(3): 473-448.
Monks, G.G. 1981 Seasonality studies. In Advances in archaeological method and theory
vo1.4, edited by M.B. Schiffer, pp. 177-240. Academic Press, New York.
Morales 1995 The guinea pig: healing, food, and ritual in the Andes. University of
Arizona Press, Tucson.
Moran, N.C. and T.P. O'Connor 1994 Age attribution in domestic sheep by skeletal and dental maturation: a
pilot study of available resources. International Journal of Osteoarchaeology 4:267-285.
Morena-Garcia, M. 2004 Hunting practices and consumption patterns in rural communities in the
Rif Mountains (Morocco)- some ethno-zoological notes. In Behavior behind Bones: The Zooarchaeology of ritual, religion, status and identity, edited by S.J. O'Day, W. Van Neer, and A. Ervynck, pp. 327-334. Oxbow Books, Oxford.
Morris, E.H., and R.F. Burgh 1954 Basketmaker I1 Sites Near Durango, Colorado. Publication 604, Carnegie
Institution of Washington, Washington D.C.

Morris, J.N. 1991 Archaeological Excavations on the Hovenweep Laterals. Four Corners
Archaeological Project Report No. 16. Complete Archaeological Service Associates, Cortez, CO.
Muir, R.J. 1999 Zooarchaeology of Sand Canyon Pueblo, Colorado. Ph.D. thesis. Simon
Fraser University.
Muir, R.J., and J.C. Driver
Muller, V. I985
Munro, N.D. 1994
Munson, M.K. 2000
Naroll, R. I962
Scale of Analysis and Zooarchaeological Interpretation: Pueblo Ill Faunal Variation in the Northern San Juan Region. Journal of Anthropological Archaeology 21, pp. 165-1 99.
Faunal Remains. In The Archaeology of Yellow Jacket Pueblo: Excavations at a Large Community Center in Southwestern Colorado, edited by K.A. Kuckelman. Crow Canyon Archaeological Center, Cortez.
Origins of Class and Gender Hierarchy in Northwestern Europe. Dialectical Anthropology 10: 93-1 05.
An Investigation of Anasazi Turkey Production in Southwestern Colorado. Unpublished Masters Thesis. Simon Fraser University.
Sex, Gender, and Status: Human Images from Classic Mimbres. American Antiquity 65(1): 127-1 43.
Floor area and settlement pattern. American Antiquity 27:587-89.
Nelson, M., D. Glowacki, and A. Smith 2002 The Impact of Women on Household Economies: A Mayan Case Study.
In Pursuit of Gender, Worldwide Archaeological Approaches, edited by S.M. Nelson and M. Rosen-Ayalon. Altimira Press: Walnut Creek, CA.
Nelson, S.M, 1997 Gender in archaeology-analyzing power and prestige. Altamira Press,
London.
Nemeth, D.J. 1998 Privy-Pigs in Prehistory? A Korean Analogy for Neolithic Chinese
Subsistence Practices. MASCA Research Papers in Science and Archaeology Vol. 15. Ancestors for the Pigs: Pigs in Prehistory, edited by S.M. Nelson. Museum of Applied Science Center for Archaeology University of Pennsylvania Museum of Archaeology and Anthropology, Philadelphia.

Neusius, S.W. 1986a Faunal Remains from Windy Wheat Hamlet. In Dolores Archaeological
Program: Anasazi Communities at Dolores: Early Anasazi Sites in the Sagehorn Flats, compiled by A.E. Kane and G.T. Gross, pp.855-862. Bureau of Reclamation, Engineering and Research Center, Denver.
1986b Faunal Remains from Permian Hamlet Area 1. In Dolores Archaeological Program: Anasazi Communities at Dolores: Middle Canyon Area, compiled by A.E. Kane and C.K. Robinson, pp. 187-197. Bureau of Reclamation, Engineering and Research Center, Denver.
1986c Faunal Remains from the Le Moc Shelter. In Dolores Archaeological Program: Anasazi Communities at Dolores: Early Small Settlements in the Dolores River Canyon and Western Sagehen Flats Area, compiled by T.A Kohler, W.D. Lipe, and A.E. Kane, pp. 279-296. Bureau of Reclamation, Engineering and Research Center, Denver.
1986d Faunal Remains at Prince Hamlet. In Dolores Archaeological Program: Anasazi Communities at Dolores: Early Small Settlements in the Dolores Canyon Area and the Western Sagehen Flats Area, compiled by T.A. Kohler, W.D. Lipe, and A.E. Kane, pp. 483-496. Bureau of Reclamation, Engineering and Research Center, Denver.
Neusius, S.W., and M. Gould 1988 Faunal Remains: Implications for Dolores Anasazi Adaptations. In
Dolores Archaeological Program: Anasazi Communities at Dolores: Grass Mesa Village, compiled by W.D. Lipe, J.N. Morris, and T.A. Kohler, pp. 1049-1 135. Bureau of Reclamation, Engineering and Research Center, Denver.
Nickens, P.R. 1981 Pueblo 111 Communities in Transition: Environment and Adaptation in
Johnson Canyon. Memoirs, no. 2. Colorado Archaeological Society, Boulder.
Nielson, A.S., J.C. Janetski, and J.D. Wilde 1985 Final Recapture Wash Archaeological Project 1981-1983, San Juan
County, Utah. Museum of Peoples and Cultures Technical Series, nos. 85-87. Brigham Young University, Provo, Utah.
Nordby, L.V. 1973 Salvage Excavations in Mancos Canyon, Ute Mountain Ute Homelands,
Colorado, during the Summer of 1972. Unpublished Mater's thesis, Department of Anthropology, University of Colorado, Boulder.
O'Connell J.F., and K. Hawkes 1984 Food choice and foraging sites among the Alyawara. Journal of
Anthropological Research 40: 435-504.

O'Connor, T. 2000 The archaeology of animal bones. Texas A&M University Press, College
Station.
Ogilvie, M.D, and C.E. Hilton 2001 Ritualized Violence in the Prehistoric American Southwest. International
Journal of Osteoarchaeology 10: 27-48.
Oliver, D.L. 1955 A Solomon Island Society: Kinship and Leadership among the Siuai of
Bouganville. Harvard University Press, Cambridge.
Olsen, S.L. 1980 Bone Artifacts from Kinishba Ruin: Their Manufacture and Use. Kiva, 46
(1 -2):39-67.
Orcutt, J.D., E. Blinman, and T.A. Kohler 1990 Explanations of Population Aggregation in the Mesa Verde Region Prior
to A.D. 900. In Perspectives on Southwestern Prehistory, edited by P.E. Minnis and C.L. Redman, pp. 196-21 2. Westview Press, Boulder.
Ortman, S.G. 1998 Corn Grinding and Community Organization in the Pueblo Southwest,
A.D. 1 150-1 55-. In Migration and Reorganization: The Pueblo IV Period in the American Southwest, edited by K.A. Spielmann, pp. 165-1 92. Anthropological Research Papers 51. Arizona State University, Tempe.
Ortman, S.G., D.M. Glowacki, M.J. Churchill, and K.A. Kuckelman 2000 Pattern and Variation in Northern San Juan Village Histories. In Kiva,
vo1.66, no. 1.
Payne, S. 1987 Reference codes for wear stages in the mandibular cheek teeth of sheep
and goats. Journal of Archaeological Science 14:609-614
Parsons, E.C. 191 8 Notes on Acoma and Laguna. American Anthropologist 20: 162-1 86.
1920 Notes on Isleta, Santa Ana, and Acoma. American Anthropologist 22: 56- 59.
1921 Further Notes on Isleta. American Anthropologist 23: 149-169.
1970 Taos Pueblo. Johnson Reprint, New York.
1977 The Pueblo of Jemez. University Microfilms, Ann Arbor.
Patterson-Rudolph, C. 1993 Petroglyphs and Pueblo Myths of the Rio Grande. Avanyu Publishing Inc,
Albuquerque.

Pavlovic, J.M. 1973 Folk life and customs in the Kragujevac region of the Jasenica in
Sumdaija. Human Resource Area Files, 1997 HRAF Computer files, New Haven.
Pennington, C.W. 1963 The Tarahumara of Mexico: their environment and material culture.
University of Utah Press, Salt Lake City.
Perkins, D. and P. Daly 1968 A Hunter's Village in Neolithic Turkey. Scientific American 219(5):96-106.
Petersen, K.L.
Plog, F.G. 1979
Plug, C. and I 1990
Climatic Reconstruction for the Dolores Project Area. In Dolores Archaeological Program: Studies in Environmental Archaeology, compiled by K.L. Petersen and V.L. Clay, pp. 207-216. U.S. Department of the Interior, Bureau of Reclamation, Engineering and Research Center, Denver.
Climatic Reconstruction for the Dolores Project. In Dolores Archaeological Program: Final Synthetic Report. Compiled by D.A. Breternitz, C.K. Robinson, and G.T. Gross, pp. 312-325. United States Department of the Interior, Bureau of Reclamation, Engineering and Research Center, Denver.
Climate and the Dolores River Anasazi: A Paleoenvironmental Reconstruction from a 70,000-Year Pollen Record, La Plata Mountains, Southwestern Colorado. Anthropological Papers No. 113. University of Utah, Salt Lake City.
A Warm and Wet Little Climatic Optimum and a Cold and Dry Little Ice Age in the Southern Rocky Mountains, U.S.A. Paper prepared for the U.S. Department of Energy, Office of Environmental Restoration and Waste Management. Westinghouse Hanford Company, Richland, WA.
A Warm and Wet Little Climatic Optimum and a Cold and Dry Little Ice Age in the Southern Rocky Mountains, U.S.A. Climatic Change 26:243- 269.
Prehistory: Western Anasazi. In Southwest, edited by A. Ortiz, pp. 108- 130. Handbook of North American Indians vol. 9, W.C/ Sturtevant, general editor. Smithsonian Institution, Washington D.C.
Archaeological Bulletin 45, 53-57.
Plug MNI counts as estimates of species abundance. South African

Potter, J.M. 1997 Communal Ritual and Faunal Remains: an Example from the Dolores
Anasazi. Journal of Field Archaeology 24: 353-364.
2000 Pots, Parties, and Politics: Communal Feasting In the American Southwest. American Antiquity 65(3): 471 -492.
Quirt-Booth, T., and K. Cruz-Uribe 1997 Analysis of Leporid Remains from Prehistoric Sinagua Sites, Northern
Arizona. Journal of Archaeological Science 24:945-960. Rautman, A.E
1993 Resource Variability, Risk, and the Structure of Social Networks: An Example from the Prehistoric Southwest. American Antiquity 58(3): 403- 424.
Reyman, J.E. 1978 Pochteca Burials at Anasazi Sites? In Across the Chichimec sea, edited
by C.L. Riley and B.C. Hedrick. Southern Illinois University Press, Carbondale.
Reed, E.K. 1951 Turkeys in southwestern archaeology. El Palacio 58: 195-205.
Reitz, E.J., and E.S. Wing 1 999 Zooarchaeology. Cambridge University Press, Cambridge.
Rice, P. 1991 Women and Prehistoric Pottery Production. In The
Archaeology of Gender: Proceedings of the Twenty-second Annual Chokmool Conference of the Archaeological Association of the University of Calgary, edited by 0 . Walde and N.D. Willows, pp. 436-443. Archaeological Association, University of Calgary, Calgary.
Richards, A.I. 1939 Land, labour and diet in Northern Rhodesia: an economic study of the
Bemba tribe. International Institute of African Languages & Cultures, Oxford University Press, London.
Ringrose, T.J.
Rohn, A.H. 1971
Bone counts and statistics: a critique, Journal of Archaeological Science 20, 121-157.
Mug House, Mesa Verde National Park, Colorado. Archaeological Research Series, no. 7-0. National Park Service, Washington D.C.
A stockaded Basketmaker Ill Village at Yellow Jacket, Colorado. The Kiva 40: 1 13-1 19.
Northern San Juan Prehistory. In Dynamics of Southwestern Prehistory, edited by L.S. Cordell and G.J. Gumerman, pp. 149-177. Smithsonian Institution Press. Washington D.C.

Roscoe, W. 1991 The Zuni Man-Woman. University of New Mexico Press, Albuquerque.
Rowlands, M.J. 1973 Defense: A Factor in the Organization of Settlements. In Territoriality and
Proxemics: Archaeological and Ethnographic Evidence for the Use and Organization of Space, edited by R. Tringham, pp. 1-16. Warner Modular Publications, Andover.
Rowlely-Conwy, P. (editor) 2000 Animal Bones, Human Societies. Oxbow Books, Oxford.
Rubel, P.G., and A. Rosman 1978 Your Own Pigs You May Not Eat: A Comparative Study of New Guinea
Societies. The University of Chicago Press, Chicago.
Schlanger, S.H. 1988 Patterns of Population Movement and Long-term Population Growth on
Southwestern Colorado. American Antiquity 53: 773-793.
Schneider, D,M., and J.M Roberts 1965 Zuni Daily Life. Human Relations Area Files Press, New Haven.
Schoeninger, M.J., and K. Moore 1992 Bone stable isotope studies in archaeology. Journal of World Prehistory
6(2): 247-296.
Schoenwetter, J. 1966 A Re-Evaluation of the Navajo Reservoir Pollen Chronology. El Palacio
73: 19-26.
Schram, L. 1954 The Monguors of the Kansu-Tibetan frontier: their origin, history and
social organization. American Philosophical Society, Philadelphia.
Schroeder, A.H. 1965 A Brief History of the Southwestern Utes. Southwestern Lore 3053-78.
Sebastian, L. 1992 The Chaco Anasazi: sociopolitical evolution in the prehistoric Southwest.
Cambridge University Press, Cambridge.
Shaffer, B.S. 1996 Prehistoric Small Game Snare Trap Technology, Deployment Strategy,
and Trapper Gender Depicted in Mimbres Pottery. Journal of Ethnobiology 16(2): 145-1 55.

Shaffer, B.S., and K.M. Gardner 1995 The Rabbit Drive through time: Analysis of the North American
Ethnographic and Prehistoric Evidence. Utah Archaeology 8:13-25.
Shapiro, J. 1983 Anthropology and the study of Gender. In A Feminist
Perspective in the Academy: the Difference I t Makes, edited by E. Langland and W. Gove, pp 110-129. University of Chicago Press, Chicago.
Shelley, S.D. 1993 Analysis of the Faunal Remains from the Wallace Ruin: A Chacoan
Outlier near Cortez, Colorado. Unpublished Ph.D. dissertation, Department of Anthropology, Washington State University, Pullman.
Shipman, P., G. Foster, and M. Schoeninger 1984 Burnt Bones and Teeth: An Experimental Study of Color, Morphology,
Crystal Structure and Shrinkage. Journal of ~rchaeolo~ica l science 1 11307-325.
Shore, 6. 1981
Shostak, M. 1981
Siemer, E.G. 1977
Skoggard, I. 2001
Sexuality and Gender in Samoa: Conceptions and Missed Conceptions. In Sexual Meanings: The Cultural Construction of Gender and Sexuality, edited by S. Ortner and H. Whitehead, pp. 192-215. Cambridge University Press, Cambridge.
Nisa: the Life and Worlds of a !Kung Woman. Random House inc., New York.
Colorado Climate. Colorado Experiment Station, Colorado State University, Fort Collins.
Cultural Summary: Ovimbundu. Human Relations Area Files. Computer Files, New Haven.
Smith, E.A., and M. Wishnie 2000 Conservation and Subsistence in Small-Scale Societies. Annual Review
of Anthropology 29:493-524.
Sorensen, M.L.S. 1988 Is there a feminist contribution to archaeology? Archaeological Review
from Cambridge 7(1): 9-20.
Spector, J. 1982 Male and Female Task Differentiation Among the Hidatsa:
Toward the Development of an Archaeological Approach to the Study of Gender. In Hidden Half: Studies of Plains Indian Women, edited by P

Albers and B. Medicine, pp. 77-100. University Press of America, Washington DC.
Speth, J.D., and S.L. Scott 1989 Horticulture and large-mammal hunting: the role of resource depletion
and constraints of time and labor. In Farmers as hunters: the implications of sedentism, edited by S. Kent, pp. 71-79. Cambridge University Press, Cambridge.
Sponheimer, M., T. Robinson, L. Ayliffe, B. Roede, J. Hammer, B. Passey, A. West, T Cerlings, D. Dearing, and J. Ehleringer
2003 Nitrogen isotopes in mammalian herbivores: hair 6 '=N values from a controlled feeding study. International Journal of Osteoarchaeology l3(1-2): 80-87.
Stahl, A.B. 1992
Stark, M.T. 1993
Concepts of Time and approaches to analogical reasoning in historical perspective. American Antiquity 58: 235-260.
Refitting the "cracked and broken fa~ade": the case for empiricism in post-processual ethnoarchaeology. In Archaeological Theory: Who Sets the Agenda?, edited by N. Yoffee and A. Sherratt, pp. 93-104. Cambridge University Press, Cambridge.
Stevenson, M.C. 1894 The Sia. Eleventh Annual Report of the Bureau of American Ethnology
1889-90, pp. 3-1 57. Washington D.C.
Steward, J.H. 1941 Archaeological Reconnaissance of Southern Utah. Anthropological
Paper 18, Bureau of American Ethnology Bulletin 128. Smithsonian Institution, Washington D.C.
Strathern, A. 1971 The Rope of Moka: Big-men and Ceremonial Exchange in Mount Hagen
New Guinea. University Press, Cambridge.
Strathern, M. 1972 Women in Between: Female Roles in a Male Worlds: Mount Hagen, New
Guinea. Seminar Press, London.
Statton, S.K. 1999 Reconstructing the Role of Faunal Resource Use During the
Occupational History of Grass Mesa Village, Southwestern Colorado. PhD. Dissertation, University of New Mexico, Albuquerque.
Szuter, C.R. 2000 Gender and Animals: Hunting Technology, Ritual, and

Subsistence in the Greater Southwest. In Women & Men in the Prehispanic Southwest: Labor, Power, and Prestige, edited by P. Crown, pp. 169-196. School of American Research, Santa Fe.
Szuter, C.R., and F.E. Bayham 1989 Sedentism and prehistoric animal procurement among desert
horticulturalists of the North American Southwest. In Farmers as hunters: The implications of sedentism, edited by S. Kent, pp.80-95. Cambridge University Press: Cambridge
Szuter, C.R., and W.B. Gillespie
Thevet, A. 2001
Thiessen, I. 2001
Interpreting Use of Animal Resources at Prehistoric American Southwest Communities. In The Ancient Southwestern Community, edited by W.H. Wills and R.D. Leonard, pp. 67-76. University of New Mexico Press, Albuquerque.
Evoking the Devil: Fasting with Tobacco to Learn How to Cure (originally published in 1557). In Shamans Through Time: 500 Years on the Path to Knowledge, edited by J. Narby and F. Huxley, pp. 13-15. Jeremy P. TarcherIPutnam, New York.
The Social Construction of Gender: Female Cannibalism in Papua New Guinea. Anthropos 96: 141-1 56.
Thompson, C. 1990 Anasazi Social Boundaries and Resource Access: The Faunal Evidence
from Nancy Patterson Village, Utah. Unpublished Master's thesis, Department of Anthropology, Brigham Young University, Provo, Utah.
Trigger, B.G. 1982 Ethnohistory: problems and prospects. Ethnohistory 29(l/4): 1-1 9.
Tringham, R.E. 1991 Household with Faces: The Challenges of Gender in Prehistoric
Architectural Remains. In Engendering Archaeology: Women and Prehistory, edited by J.M. Gero and M.W. Conkey, pp. 93-1 30. Basil Blackwell, Cambridge.
Tschopik, H. 1946 The Ayamara. Smithsonian Institution, Washington D.C.
Turner, C.G. and J.A. Turner 1999 Man Corn: Cannibalism and Violence in the Prehistoric American
Southwest. The University of Utah Press, Salt Lake City.
Upham, S. 1982 Polities and Power: An Economic and Political History of the Western
Pueblo. Academic Press, New York.

1984 Adaptive Diversity and Southwestern Depopulation. Journal of Anthropological Research 40: 235-256.
Upham, S., and P.F. Reed 1989 Inferring the Structure of Anasazi Warfare. In Cultures in Conflict:
Current Archaeological Perspectives. Proceedings of the Twentieth Annual Conference of the Archaeological Association of the University of Calgary. pp. 163-1 72. The University of Calgary, Calgary.
U.S. Geological Survey 2005 U.S. Department of the Interior. http://www.usc~s.c~ov. Accessed March
17,2004.
Van der Merwe, N.J. I982 Carbon Isotopes, photosynthesis, and archaeology. American Scientist
70: 596-606
Van Pelt, N.S. I978 Woodlands and Parks in Southeastern Utah, Unpublished Master's
Thesis, Department of Geography, University of Utah, Salt Lake City.
VanPool, C.S., and T.L. VanPool 2006 Gender in Middle Range Societies: A Case Study in Casas Grandes
Iconography. American Antiquity 71 (1 ): 53-75.
Van West, C.R., and W.D. Lipe
Varien, M.D. 1999
Modeling prehistoric Climate and Agriculture in southwestern Colorado. In The Sand Canyon Archaeological Project: a Progress Report, edited by W.D. Lipe, pp105-120. Crow Canyon Archaeological Center Occasional Paper No. 2. Cortez.
Sedentism and Mobility in a Social Landscape: Mesa Verde and Beyond. University of Arizona Press, Tucson.
Persistent Communities and Mobile Household: Population Movement in the Central Mesa Verde Region, A.D. 950 to 1290. In Seeking the Center Place: Archaeology and Ancient Communities in the Mesa Verde Region. Edited by M.D. Varien and R.H. Wilshusen, pp.163-184. The University of Utah Press, Salt Lake City.
Varien, M.D., K.A. Kuckelman and J.H. Kleidon 1992 The Site Testing Program. In The Sand Canyon Archaeological Project:
a Progress Report, edited by W.D. Lipe, pp. 45-67. Crow Canyon Archaeological Center Occasional Paper No. 2. Cortez.
Varien, M.D., W. Lipe, M. Adler, I. Thompson, and B. Bradley 1996 Southwest Colorado and Southeastern Utah Settlement Patterns: A.D.
1 100 to 1300. In Prehistoric Pueblo World A.D. 1150-1350, edited by M. Adler, pp 86-1 13. University of Arizona Press, Tucson.

Varien, M.D., C.R. Van West, and G.S. Patterson 2000 Competition, Cooperation and Conflict: Agricultural Production and
Community Catchments in the Central Mesa Verde Region. In Kiva, Vol. 66, NO. 1, pp. 45-66.
Villa Rojas, A.
Wadley, L. 1999
Walker, D.N. 1993
Walker, M.K. 1974
The Maya of east central Quintana Roo. Carnegie Institution, Washington D.C.
The Invisible Meat Providers: Women if the Stone Age of South Africa. In Gender in African Prehistory, edited by S. Kent, pp. 69-82. Alta Mira Press, Walnut Creek.
Faunal Remains. In The Duckfoot Site Volume 1: Descriptive Archaeology, edited by R.R. Lightfoot and M.C. Etzkorn, pp. 239-252. Occasional Papers, no. 3. Crow Canyon Archaeological Center, Cortez.
Social constraints, individuals, and social decisions ins a Scottish rural community. University Microfilms International, Ann Arbor.
Wenham, G., and V.R. Fowler 1973 A radiographic study of age changes in the skull, mandible and teeth of
pigs. Journal of Agricultural Science 1 7: 187-1 96.
Western Regional Climate Center 2005 www.wrcc.dri.edu. Accessed September 19,2004.
White, T.D. 1977
White, L.A. 1932
An Analysis of the 1976 Faunal Remains. In Hovenweep 1976, by J.C. Winter, pp. 237-242. Archaeological Report, no. 3. San Jose State University, San Jose California.
Prehistoric Cannibalism at Mancos 5MTUMR-2346. Princeton University Press, Princeton.
The Pueblo of San Felipe. Memoirs of the American Anthropological Association, No. 38, Washington D.C.
A method for calculating the dietary percentages of various food animals utilized by aboriginal peoples. American Antiquity 18, 368-398.
1974a The pueblo of Santo Domingo, New Mexico. Kraus Reprint, Millwood, N.Y. (originally published in 1935, Memoirs of the American Anthropological Association No. 43.)
1974b Zia-the sun symbol pueblo. University of Albuquerque, Albuquerque.

Wilbert, J. 1987
Wilcox, D.R. 1981
Wilcox, D.R., 1994
Tobacco and Shamanism in South America. Yale University Press, New Haven.
Changing Perspectives in the Protohistoric Pueblos, A.D. 1450-1700. In The Protohistoric Periods in the North American Southwest, edited by D.R. Wilcox and W.B. Masse, pp. 378-409. Research Paper no. 24. Arizona State University, Tempe.
nd J. Haas The Scream of the Butterfly: Competition and Conflict in the Prehistoric Southwest. In Themes in Southwest Prehistory, edited by G.J/ Gumerman, pp. 21 1-238. School of American Research, Santa Fe.
Wilshusen, R.H. 1999a Chapter 6. Basketmaker Ill (A.D. 500-750). In Colorado Prehistory: A
Context for the Southern Colorado River Basin, edited by W.D. Lipe, M.D. Varien, and R.H. Wilshusen, pp. 166-195. C.C.P.A., Denver.
1999b Chapter 7. Pueblo 1 (A.D. 750-900). In Colorado Prehistory: A Context for the Southern Colorado River Basin, edited by W. D. Lipe, M. D. Varien, and R.H. Wilshusen. Pp. 196-241. C.C.P.A., Denver.
2002 Estimating Population in the Central Mesa Verde Region. In Seeking the Center Place: Archaeology and Ancient Communities in the Mesa Verde Region, edited by M.D. Varien and R.H. Wilshusen, pp. 101-120. University of Utah Press, Salt Lake City.
Wilshusen, R.H., and E. Blinman 1992 Pueblo I Village Formation: A Reevaluation of Sites Recorded by Earl
Morris on Ute Mountain Tribal Lands. The Kiva 77: 251-269.
Wilshusen, R.H., M.J. Churchill, and J.M. Potter 1997 Prehistoric Reservoirs and Water Basins in the Mesa Verde Region:
Intensification of Water Collection Strategies during the Great Pueblo Period. American Antiquity 62(4):664-681.
Wilshusen, R.H., and S.G. Ortman 1999 Rethinking the Pueblo I Period in the San Juan Drainage: Aggregation,
Migration, and Cultural Diversity. The Kiva 64: 369-399.
Wilshusen R.H., and R.H. Towner 1999 Post Puebloan Occupation (A.D. 1300-1 840). In Colorado Prehistory:
A Context for the Southern Colorado River Basin, edited by W.D. Lipe, M.D. Varien, and R.H. Wilshusen, pp. 353-369. C.C.P.A., Denver.
Wilson, C.D., and E. Blinman 1993 Upper San Juan Region Pottery Typology. Archaeology Notes 80.
Museum of New Mexico, Anta Fe.

Winter, J.C. 1978 Anasazi Agriculture at Hovenweep I: Field Systems. In Limited Activity
and Occupation Sites: A Collection of Conference Papers, compiled and edited by A.E. Ward, pp. 83-97. Contributions to Anthropological Studies No. 1. Center for Anthropological Studies, Albuquerque.
1981 Anasazi Agriculture at Hovenweep II: The Development and Use of Towers. Contract Abstracts and CRM Archaeology 2(2): 28-36
Wobst, H.M. 1978 The Archaeo-ethnology of Hunter-Gatherers, or the Tyranny of
the Ethnographic Record in Archaeology. American Antiquity 43: 303-9.
Wright, H.E., Jr., A.M. Bent, B.S. Hansen, and L.J. Maher, Jr.
Wylie, A. 1992
Zierhut, N.W. I967
Zihlman, A. I989
Present and Past Vegetation of the Chuska Mountains, Northwestern New Mexico. Bulletin 84:1155-1188. Geological Society of America.
Feminist Theories of Social Power: Some Implications for a Processual Archaeology. Norwegian Archaeological Review 25(l):5 1 -67.
Bone Breakage Activities of the Calling Lake Cree. Alberta Anthropologist 1:33-36.
Woman the Gatherer: The Role of Women in Early Hominid Evolution. In Gender and Anthropology: Critical Reviews for Research and Teaching, edited by S. Morgan, pp. 21-40. American Anthropological Society, Washington DC.
Gender: The View from Physical Anthropology. In The Archaeology of Gender: Proceedings of the Twenty-second Annual Chacmool Conference of the Archaeological Association of the University of Calgary, edited by D. Walde and N. Willows, pp. 4-1 0. Archaeological Association of the University of Calgary, Calgary.
Zolbrod, P.G. (Translator) 1988 Dine bahane: The Navajo Creation Story. University of New Mexico
Press, Albuquerque.

APPENDIX A:
CROW CANYON ARCHAEOLOGICAL CENTER ZOOARCHAEOLOGICAL
IDENTIFICATION AND RECORDING STANDARDS
(After Driver 1999)
Taxon
The identification of bone fragments is a complex process, and different zooarchaeologists approach this task in different ways. It is important to observe the following rules:
1. The only bones which are to be considered "identifiable" are those for which the element can be specified. No identification to any taxonomic level (even of the "large mammal" or "small bird" variety) will be allowed unless the element is identified. Terms such as "long bone" or "axial" do not qualify as element descriptions.
2. It is important to define a "universe" of species from which the animal remains are assumed to derive. (most zooarchaeologists do this unconsciously, and rarely make their decisions explicit). This is because virtually all zooarchaeological identification presupposes that certain animals are likely to be represented at a certain time and location. For example, when we are working on 13 '~ century Anasazi sites, we will assume that the bears found may be black bears or grizzlies; we will not bother to check our archaeological specimens against polar bears or Old World bear species, even though it might be difficult to distinguish those species from North American species on the basis of Osteology. For analyses of faunal assemblages of the last few thousand years, it will be assumed that the extant historically know faunas of southern Colorado and Utah and northern New Mexico and Arizona provide the universe from which our specimens are drawn.
Definition of this universe does not preclude the possibility of more exotic species being identified. However, these will normally only be identified when it can be positively demonstrated that an Anasazi area species cannot be represented by a particular bone.
3. Identification may be made to standard zoological classification, such as species, genus, family etc. Zooarchaeologists often use less formal categories such as "large bird", "medium artiodactyl", and terms such as these can be used.
4. In order to be confident of identifications, you must be able to justify your choice of taxon. This is best done by comparing your specimen with all taxa from the local faunal "universe ." In practice this is achieved rapidly, because your general knowledge of anatomy will allow you to eliminate most taxa from consideration. However, you should only identify to a particular "level of identifiability" if you are sure that the identification will bear scrutiny. You can only identify to the species level if you can definitely exclude all other species of the same genus in the study area. You can only identify to genus if you can exclude all other genera from the same family in the study area. You can only identify to family if you can exclude all other families from the same order in the study area etc. etc.
5. Each bone or bone fragment must be identified on its own merits. For example, if a burial of a dog was excavated, some bones would be referred to species while others (e.g. the ribs and vertebrae) would be referable only to the genus or family

level. You can use the "comments" section to note the presence of articulating specimens.
6. Remember that there is no disgrace in not being able to identify bone fragments to the species level. Most species are defined by a host of characters, most of which will not preserve in the skeleton. It is much better to be conservative that over- confident. Once the analysis is finished and interpretation begins, you may wish to make some assumptions about the bones identified. For example, if all the artiodactyls identified to species are from deer, you may wish to assume (perhaps for the purposes of studying body parts represented) that all "medium artiodactyls" are also deer. This can be stated in the faunal report, and would be quire a reasonable assumption; it would be unreasonable to make such an assumption while bone fragments were being identified.
Element Element refers to the whole bone of which you may either find a complete
specimen or a fragment. There are fairly well standardized names for most of the individual bones in vertebral skeletons, although fish bones are not particularly well standardized and there is still controversy about which system should be used.
Although we should ideally be able to specify elements fairly exactly, this is not always possible. For example, we may be able to identify the proximal phalanx of a deer, yet not determine whether it is from digit Ill or digit IV.
Cranial fragments present something of a problem because the cranium is composed of many named bones. When coding cranial fragments, use the names of individual bones if the majority of the fragment is made up of a particular bone; otherwise use the general code for cranial fragment.
Part These codes refer to the portion of the element that is represented. For each
major type of element there are a series of numeric codes to designate different portions. The code "1"always refers to a complete element, but others vary depending on the element being described.
Confusion may result when dealing with bones in which epiphyses are not fused. For example, a complete mammal long bone with unfused epiphyses would be coded as "1" (i.e. complete) even if the unfused epiphyses were not recovered, because it is likely that the bone was originally deposited as a complete unit. If unfused epiphyses are present and can be fitted back to the diaphysis, they should be considered as part of a single element, and should not be coded as separate fragments.
Side These can be coded as left, right, irrelevant (e.g. for the vertebral column), or as
"unknown ."
Fusion Each fragment must receive a two letter code for fusion (one for the proximal end
and one fro the distal end). Fusion can be coded as fused, unfused, just fused, and fetal.
Breakage Each fragment receives a two-letter code indicating type of breakage for each
end. Types of breaks include: intact (I); broken during excavation (E); made into an artifact (A); chewed by carnivores (C); eroded (D); splintered (P), with a series of

transverse fractures terminating at different points; gnawed by rodents (R); spiral fractures (S), with a distinctive spiral morphology and smooth break surfaces; transverse fractures (T), with breaks perpendicular to the long axis of the bone; and irregular fractures (V), that display a zigzag pattern.
Modification This refers to either natural or cultural alteration to the bone. More than one
letter code may be used to describe a number of alterations. For example, a bone may display burning and rodent gnawing, each of which requires a code.
Length Each fragment is coded according to a simple scale. Exact lengths are not
required. For example, fragments with a length less than 1 cm are coded as "1 ." Fragments with a length between 1 to 1.99 cm are coded as "2" etc. etc.
Cortical thickness This is measured (in mm) only for long bones. It is designed mainly to allow the
analyst to assign a size range for otherwise unidentifiable long bone fragments. This will assist in interpretation of butchery and fragmentation. As cortical thickness varies, use the thickest portion of cortex to define the thickness. The measurement is taken perpendicularly from the outside to the inside (marrow cavity) of the fragment. Thickness is indicated by numeric code. For example, a fragment with a thickness of less than 2 mm is coded as "1 ." Fragments with a thickness of 2 to 3.99 mm are coded as "2" etc. etc.
Comment This field is available for extra comments, which you may wish to make (e.g., tool
type, extent of weathering etc. etc.).

APPENDIX B:
MODIFIED BONE FREQUENCIES FOR A SERIES OF SITES
Table 61 Culturally Modified Bone- Shields Pueblo
I Spiral Fracture Burning Cut Marks GrndlPolished &I I I
Canis sp. (374) 11 1 .267
Artiodactyla (650)
I I
Carnivore (1 7) 11 1 5.88
N
12
Small Carnivore (1 7) 1 0
%
1.84
I I
Lynx sp. (1 7)
Sylvilagus sp. (7289) 1 225 1 3.08
1
.531 Med. Mammal
(1 88)
Lepus (1421 )
I I
Erethizon dorsatum (19) 1 0
29.4
1
I I
Cynomys sp. (557) ( 8 1 1.43
33
I I
Neotoma sp. (202) 1 0
2.32
I I
Rodentia (1 55) 11 1 .645 I I
Sciuridae (1 06) 1 0 I I
Meleagris gallopavo
(3680)
(91)
Falconiforme (1 3)
5.49 Medium Bird 5
1 7.69

Table 82 Culturally Modified Bone- Woods Canyon (Driver 2002)
Taxon (NISP) Spiral Fractures
Artiodactyla ( 0 1
Canidae (21
Lagomorph I 0 I (16) Lepus (1 1 ) 4 36.3 Sylvilagus 1 15 ( 7.46 (201) -
Small mammal 3 11.1
Rodent (2) 10 1 Galiforms (3) 1 33.3 Meleagris 15 6.12 gallopavo (245) Large Bird 52 20.4
Burning Cut marks GroundIPolished

Table B3 Culturally Modified Bone- Castle Rock Pueblo (Driver 2000)
Taxon (NISP)
Artiodactyla (6) Odocoileus (11) Med. Art. (38) Canidae (4) Canis sp (2) Med. Carnivore (3) Med. Mammal (1 9) Lagomorph (50) Lepus (106) Sylvilagus (843) Small mammal (1 73) ~eo ioma (138) Rodentia (24) Sciuridae (95) Spermophilus (22) Muridae (1 8) Small Rodent (22) Buteo sp. (29) Meleagris gallopavo (284) Passeriformes (4 Phasianidae (2) Small Bird (4) Medium Bird (62) Large Bird (408)
Spiral Fractures
3 1 3 2
0 0
0 16
4
1 0 0
1
N 1 3
1 0 2 0
2
2
6 57
0
% 16.7 27.7
2.63
100
10.5
4.00
5.66
2.17 4.16 3.15 9.09
5.63
100
50.0
.245
GroundIPolished Burning
N 1 1
1 0 0 1
1
0
3 2
0
Cut Marks
N 1 2
4 1 1 1
3
1
8 43
12
7 0 2 0
1 13
0 29
1
0 1 1
35
% 16.7 9.09
2.63
33.3
5.26
28.3
-
N 0 0
1 0 0 0
0
0
0 0
0
% 16.7 18.2
10.5 25.0 50.0
33.3
15.8
2.00
7.54 5.10
6.93
%
2.63
5.07
2.10
5.55 59.0
10.2
25.0
25.0 1.61
8.57
6.89 .352
1.62
.245
0 0 0 0
0 0
2 1
0
0 0 1
1
0 0 0 0
0 0
0 34
0
0 0 0
4
11.9
.980

TableB4 Culturally Modified Bone - Yellow Jacket Pueblo (Muir and Driver 2003)
Taxon (NISP) Spiral Fracture Burning Cut Marks Ground1 Polished
N % N O h
Artiodactyla (5) 0 1 20.0 Odocoileus sp. 5 17.9 0 (28) A. americana 1 11.1 0 (9) Ovis 1 33.3 1 33.3 Canadensis (3) Bos taurus (1) Med. Artiodactyl (1 82) Large 1 20.0 0 artiodactyls (5) 1 Canidae (16) Canis SD. (1 1)
Lagomorpha 15.5 4.2 (71 1 Leius (259) 37 14.3 15 5.8 Sylvilagus sp 1 17 10.1 69 6.0
~ c i u i d a e (102) 2 2.0 3 2.9 Ground Squirrel 0 3 20.0 11 51 Spermophilus I 0 11 4.3 sp. (23) Geomyidae 0 1 2.4 (41 ) Neotoma sp 3 2.5 1 0.8
castor Canadensis 140)
retraonidae 1321 Centrocerus ~rophasianus I1 0) Weleagris ~allopavo (500) Passiformes (2)
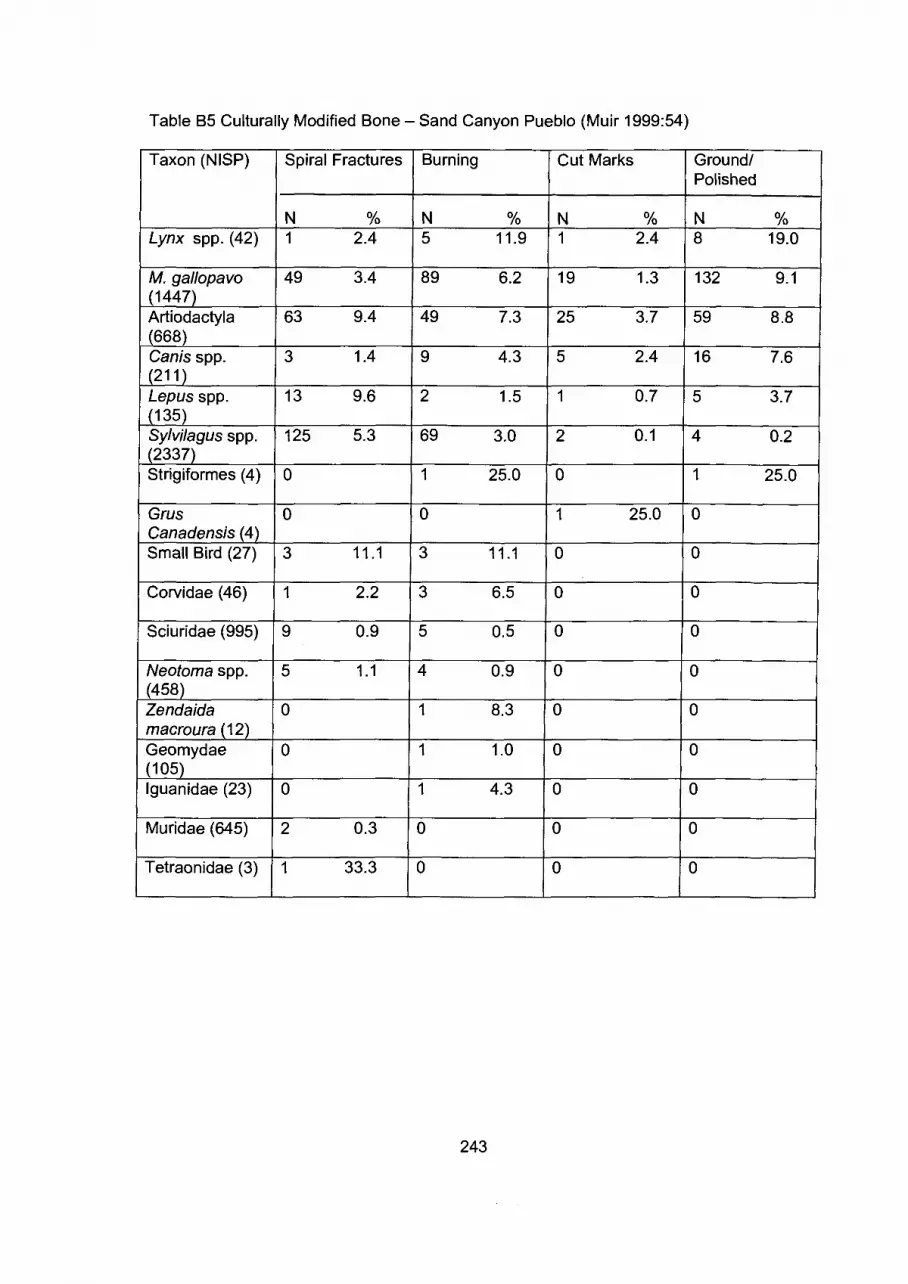
Table B5 Culturally Modified Bone - Sand Canyon Pueblo (Muir 1999:54)
I 1 I I
M, gallopavo ( 49 3.4 1 89 6.2 1 19 1.3 ( 132 9.1
Taxon (NISP)
Lynx spp. (42)
Spiral Fractures
N % 1 2.4
(1 447) .
Artiodactyla (668) Canis spp.
63 9.4
. . (21 1) Lepus spp. (1 35) Sylvilagus spp.
I
Grus 1 0 1 0 11 25.0 1 0
Ground1 Polished
N % 8 19.0
Burning
N % 5 11.9
3 1.4
. .
(2337)- Strigiformes (4)
Cut Marks
N % 1 2.4
49 7.3
13 9.6
1 25 5.3
9 4.3
0
Canadensis (4) Small Bird (27)
25 3.7
2 1.5
69 3.0
Corvidae (46)
I I I I
Neotoma spp. ( 5 1 .I 14 0.9 1 0 1 0
59 8.8
5 2.4
1 25.0
3 11.1
Sciuridae (995)
16 7.6
1 0.7
2 0.1
1 2.2
5 3.7
4 0.2
0
3 11.1
9 0.9
. . (458) Zendaida 1 0
1 25.0
3 6.5
macroura (1 2) Geomydae
0
5 0.5
1 8.3
(105) -
lguanidae (23)
0
0
0
Muridae (645)
Tetraonidae (3)
0
0
0
0
0
0
1 1 .O
2 0.3
1 33.3
1 4.3
0
0
0
0
0 0
0
0
0
0