fas-mediated killing of primary prostate cancer cells is increased by mitoxantrone and docetaxel
TRANSCRIPT

Fas-mediated killing of primary prostate cancer cells isincreased by mitoxantrone and docetaxel
Juliane C. Symes,1 Michael Kurin,1
Neil E. Fleshner,3 and Jeffrey A. Medin1,2,4
1Department of Medical Biophysics and 2Institute of MedicalSciences, University of Toronto; 3Division of Urology, UniversityHealth Network; 4Division of Stem Cell and DevelopmentalBiology, Ontario Cancer Institute, Toronto, Ontario, Canada
AbstractTherapies for prostate cancer based on Fas (CD95)modulation have been under active development at thepreclinical stage using immortalized cell lines. To addressclinical applicability, the potential of 11 cultures ofprimary prostate cancer cells to be killed by Fas-mediated apoptosis was investigated. In addition, theeffect of the chemotherapeutic agents mitoxantrone anddocetaxel on this killing was determined. Apoptosis wasinduced in patient-derived, primary prostate cancer cellsusing effector cells engineered by recombinant lentivirusinfection to express Fas ligand (FasL) and measured by51Cr release assays. All cultured prostate cells werefound to undergo Fas-mediated killing; cytotoxicityranged from 12% to 87% after 6 h. These cells weresignificantly more sensitive to FasL-mediated killing thanPC-3 cells. The basal expression of Fas or the expressionof five inhibitors of apoptosis (c-FLIP, survivin, cellularinhibitors of apoptosis protein 1 and 2, and bcl-2) wasnot found to correlate with susceptibility to Fas-mediated killing. Both mitoxantrone and docetaxel wereable to induce Fas receptor expression on primaryprostate cancer cells, which translated into a 1.5- to3-fold enhancement of apoptosis mediated by FasL.Whereas mitoxantrone increased the Fas-induced apop-totic response of all cultured prostate cells tested,docetaxel pretreatment was found to preferentiallyenhance the killing of bcl-2-expressing cells. Thesefindings show that cultured primary prostate cancercells are sensitive to Fas-mediated apoptosis. Further-more, the incidence of apoptosis was found to beimproved by combining Fas-mediated therapy with
standard chemotherapeutic agents. These findings mayhave significant implications for prostate cancer therapy.[Mol Cancer Ther 2008;7(9):3018–28]
IntroductionSince the discovery of Fas-mediated apoptosis, there hasbeen sustained interest in exploiting this pathway intherapeutic strategies against prostate cancer (1–7). Apop-totic signaling is induced in cells on binding of Fas ligand(FasL) to its receptor, Fas. A caspase cascade is initiated,leading to death of the Fas-expressing cell. Approachesmanipulating this pathway are growing more sophisticatedand functionally efficacious while at the same time limitingoff-target toxicity mediated by this ligand/receptor axis(8–11).
Although these advances are bringing FasL-based ther-apy closer to clinical reality, the effectiveness of suchtherapies against prostate cancer has not been thoroughlyinvestigated. Functional studies conducted to date makeuse of immortalized prostate cancer cell lines as models,which may not accurately reflect the potential clinicalresponse of prostate tumors to FasL-mediated killing.Indeed, the majority of these cell lines, including PC-3,LNCaP, and DU145 cells, were derived from metastases orwere artificially created by oncogene overexpression andare likely intrinsically resistant to apoptosis. In additionto immortalization, extensive culture periods may havefurther altered the phenotype of these commonly usedmodels. Primary prostate cancer cells are thus a morerepresentative model to study the response of prostatecancer to Fas-mediated killing in a preclinical setting.
Immunohistochemical studies of primary human pros-tate cancer tissues have shown expression of key apoptoticpathway proteins, including Fas and caspases (12, 13). Onthe other hand, abundant expression of inhibitors ofapoptosis, such as survivin and cellular inhibitors ofapoptosis protein 1 and 2 (cIAP1 and cIAP2, respectively),have also been documented (14, 15). Without functionalstudies, it is not known whether the Fas pathway isoperational in primary malignant prostate cells. In fact,based on expression profiling of such apoptotic mediators,it has been predicted that apoptotic signaling may bedramatically interrupted in prostate cancer tissues (12). Todate, no functional studies have been conducted looking atFas-mediated killing of patient-derived primary prostatetumor cells.
Various chemotherapeutic drugs, such as camptothecincompounds, have been shown to enhance the sensitivity ofprostate tumor cell lines to Fas-mediated apoptosis (16, 17).Certain other anticancer agents, including mitoxantrone,are known to increase cellular Fas expression in LNCaP butnot PC-3 or DU145 cells (18). Reports suggest that
Received 4/8/08; revised 6/5/08; accepted 6/9/08.
Grant support: Prostate Cancer Research Foundation of Canada.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
Requests for reprints: Jeffrey A. Medin, 67 College Street, Room 406,Toronto, Ontario, Canada, M5G 2M1. Phone: 416-340-4745;Fax: 416-340-3453. E-mail: [email protected]
Copyright C 2008 American Association for Cancer Research.
doi:10.1158/1535-7163.MCT-08-0335
3018
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

increasing Fas receptor expression in this way can leadto heightened susceptibility of cells to apoptosis in some(19, 20) but not all (21) cell types. This must be examined ona case-by-case basis.
In this study, we investigated the susceptibility ofcultured primary prostate cells to Fas-mediated apoptosis.Primary prostate cells were maintained in short-termcultures (22), which may more accurately reflect thecharacteristics of prostate cancer cells in situ . The effect ofmitoxantrone and docetaxel, chemotherapeutics currentlyused against prostate cancer, on tumor cell killing by FasLwas also explored. Successful combination of novel FasL-based treatments with existing anti-prostate cancer chemo-therapeutics that results in increased tumor cell apoptosiswould be an important advance for therapy of prostatecancer.
Materials andMethodsCulture of CellsProstate tissue was obtained from needle biopsies from
patients undergoing radical prostatectomy. Tissue frompalpable tumors were collected and cells were processedand cultured as described (23). Briefly, tissue was finelyminced, digested 12 to 18 h in 100 units/mL collagenase Iin PrEGM medium (Lonza), and pipetted to disperseclumps. Cells were washed in PBS and cultured oncollagen-coated (PureCol; Inamed) tissue-culture plates(BD Biosciences) in PrEGM medium. This protocol favorsthe growth of epithelial cells. Cell lines were named T1 toT11 and used within 3 weeks of culture. Characterizationstudies were done on the resulting cell lines (see below).All tissues were collected under an approved UniversityHealth Network ethics protocol from patients grantinginformed consent. PrEC cells were obtained from Cloneticsand cultured in PrEGM medium. PC-3 cells were obtainedfrom the American Type Culture Collection and cultured inF-12 Ham’s medium (Sigma) supplemented with 10% FCS,2 mmol/L L-glutamine, 100 units/mL penicillin, and 100mg/mL streptomycin (Sigma).
Immunofluorescence of Primary Cell CulturesT3, T7, and T8 cells were cultured on chamber slides (BD
Falcon), fixed with acetone/methanol (1:1) for 10 min at�20jC, and rehydrated in PBS. Nonspecific binding wasblocked by incubating cells with 1% goat serum in PBS +1% bovine serum albumin for 30 min. Mouse anti-humancytokeratin 8 + 18 and/or anti-cytokeratin 5 + 14 antibodies(Abcam) were used at 1:50 or 1:100, respectively, in 3%bovine serum albumin in PBS + 0.1% Tween 20 for 1 h.Alexa 488 goat anti-mouse IgG1 or Alexa 568 goat anti-mouse IgG2a (Invitrogen) were added at 1:500 in PBS +0.1% Tween 20 + 1% bovine serum albumin for 1 h. Whenslides were costained for cytokeratins 5 + 14 and 8 + 18,staining was done sequentially and an additional blockingstep with 5% mouse serum was done following addition ofthe first secondary antibody. Nuclei were visualized with1 Ag/AL 4,6¶-diamidino-2-phenylindole (Sigma). Slideswere mounted with Fluoromount-G (Southern Biotech)
and analyzed on a Zeiss Axioskop 2 fluorescent micro-scope. Mismatched combinations of primary and second-ary antibodies were used as a negative staining control, andMCF-7 cells were used as a positive control.
MTTGrowth AssayThe growth rates of all prostate tumor cultures were
compared using a MTT assay. Here, 4,000 cells per wellwere plated in a 96-well plate and allowed to adhereovernight. Ten microliters of 5 mg/mL MTT reagent(Sigma) were added followed 4 h later by 100 ALsolubilization solution (10% SDS in 0.01 mol/L HCl). Insome samples, recombinant FasL (0.01-10 ng/mL; Super-FasL; Alexis) was added for 24 to 72 h to assess Fas-induced proliferation. Plates were analyzed at 570 nm ona o Max Kinetic microplate reader (MDS AnalyticalTechnologies).
Flow Cytometric Analysis for Fas and FasL Expres-sion
Cells were harvested and blocked with 10% mouseserum. Cells were labeled with PE-anti-FasL antibodyclone NOK-1 (eBioscience) or APC- or FITC-anti-Fasantibody clone DX2 (BD Biosciences) and 7-AAD. Cellswere analyzed for Fas and FasL expression using aFACSCalibur flow cytometer (BD Biosciences) and Cell-Quest Pro software. Increases in Fas expression wereassessed following 4, 24, 48, or 72 h treatment withirradiation (2, 4, or 8 Gy), mitoxantrone (10, 100 or 1,000nmol/L), or docetaxel (100 or 1,000 nmol/L). Meanfluorescence intensity (MFI) of Fas staining was recorded.To correlate MFI with the number of Fas receptors on thecell surface, a series of Quantum Simply Cellular anti-mouse IgG microspheres were labeled with anti-Fasantibody for 30 min and a calibration curve was establishedusing QuickCal software (Bangs Laboratories). MFI read-ings for the treated and untreated cells were converted toantibody-binding capacity (ABC) values based on astandard curve.
Western BlottingProtein lysates were prepared from 80% confluent
plates of cells. Protein (30 Ag) was loaded onto 10%SDS-polyacrylamide gels and transferred to polyvinyli-dene difluoride membranes (Bio-Rad). Membranes wereblocked using 10% nonfat dry milk in PBS + 0.1% Tween20. Anti-bcl-2 (Cell Signaling Technologies), anti-chro-mogranin A (Abcam), anti-smooth muscle actin (Abcam),anti-cIAP1 (R&D Systems), anti-cIAP2 (R&D Systems),anti-survivin (Novus Biologicals), anti-c-FLIP (KamiyaBiomedical), anti-p53 (Cell Signaling Technologies),anti-actin (Chemicon International), horseradish peroxi-dase–linked anti-rabbit (Santa Cruz Biotechnology), orhorseradish peroxidase – linked anti-mouse IgG (GEHealthcare) antibodies were applied in PBS + 0.1%Tween 20 with 5% nonfat dry milk. Immunoreactivebands were visualized using Western Lightning Chemi-luminescence Reagent Plus (Perkin-Elmer). 293T cellsoverexpressing bcl-2 were used as a positive control forthe bcl-2 Western blot (provided by E. Zacksenhaus;University Health Network).
Molecular Cancer Therapeutics 3019
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

RNA Extraction and Quantitative Reverse Transcrip-tion-PCR Analyses of Prostate Tumor Markers andCharacteristic Apoptosis Pathway Proteins
RNA was extracted from cells using the Trizol reagent(Invitrogen). cDNA was synthesized from 5 Ag total RNAusing a SuperScript II Reverse Transcription kit (Invitrogen).Quantitative real-time reverse-transcription PCR was doneon a Rotor-Gene 3000 thermal cycler (Corbett Life Science)using the following primer sets (Invitrogen): cIAP1 forward5¶-TGTTGTCAACTTCAGATACCACTGG-3¶ and reverse5¶-CATCATGACAGCATCTTCTGAAGA-3¶, cIAP2 forward5¶-GGACAGGAGTTCATCCGTCAAG-3 ¶ and reverse5¶-TGGATAATTGATGACTCTGCATTTTC-3¶, survivin for-ward 5¶-CCCATAGAGGAACATAAAAAGCATTC-3¶ andreverse 5¶-TCAAAAATTCACCAAGGGTTAATTCT-3¶,c-FLIP forward 5¶-CCTAGGAATCTGCCTGATAATCGA-3¶and reverse 5¶-TGGGATATACCATGCATACTGAGATG-3¶,and 18srRNA forward 5¶-CGGCTACCACATCCAAGGAA-3¶ and reverse 5¶-GCTGGAATTACCGCGGCT-3¶. PCR wasdone using SYBR Green PCR Master Mix (Applied Bio-systems) and 0.3 Amol/L primers. Melt curve analyses andgel electrophoresis of PCR products were used to verify thepresence of a single PCR product of appropriate size.
RetroviralTransduction and Selection of K562 CellsK562 cells, a human erythroleukemia cell line, were
cultured in RPMI (Sigma) supplemented with 10% FCS,2 mmol/L L-glutamine, 100 units/mL penicillin, and100 mg/mL streptomycin (Sigma). Cells were transducedwith lentiviral particles produced from pHR¶-cPPT-EF-enGFP-W-SIN (24) or pHR¶-cPPT-ncFasL-W-SIN vectorsto engineer expression of enGFP or noncleavable FasL(ncFasL). ncFasL was created by removing the proteolyticcleavage site at amino acids 103 to 136, rendering FasLmembrane-bound for increased expression and potency (8).pHR¶-cPPT-ncFasL-W-SIN was constructed by replacingthe enGFP cDNA with the ncFasL cDNA. Viral particleswere produced by transient transfection of 293T cellsas described previously (25). K562 cells were transducedin the presence of 8 Ag/mL protamine sulfate for 16 h.Transgene expression was assessed by flow cytometry72 h post-transduction. Clonal populations of enGFP-expressing (K562-enGFP) and ncFasL-expressing (K562-ncFasL) cells were isolated by limiting dilution.51Cr ReleaseAssaysPrimary prostate cells (3 � 105) were labeled with 40 ACi
51Cr-sodium chromate (GE Healthcare) and plated intriplicate in round-bottomed 96-well plates at 104 cells perwell. K562-ncFasL or control K562-enGFP cells were addedat effector-to-target cell ratios of 1:1, 10:1, or 25:1.Background control wells contained only labeled prostatecells, whereas maximum release control wells additionallycontained 1% Triton X-100. After 6 h co-incubation, counts/min from 100 AL medium from each well was determinedon a gamma counter (1277 GammaMaster; Perkin Elmer).Percent cytotoxicity was calculated as (experimentalcounts/min - background release counts/min) / (maxi-mum release counts/min - background release counts/min) � 100. To verify that cytotoxicity was directly Fas
mediated, FasL activity was blocked by the addition of25 Ag/mL anti-FasL NOK-1 or isotype control antibody(BD Biosciences) to K562-enGFP or K562-ncFasL cells 30min before use in 51Cr release assay.
Mitochondrial Membrane Depolarization AssayTetramethylrhodamine ethyl ester (TMRE) dye (Invitro-
gen) was used to measure mitochondrial outer membraneintegrity of prostate cancer cells after induction of Fas-mediated apoptosis (26). Target cells (2.5 � 106-3 � 106)were co-incubated for 6 h at a 20:1 effector-to-target cellratio with K562-enGFP or K562-ncFasL cells that had beenlabeled with 5 Amol/L carboxyfluorescein succinimidylester (Invitrogen). Following incubation, cells were har-vested and incubated with 100 nmol/L TMRE in culturemedium for 30 min at 37jC. The percentage of carboxy-fluorescein succinimidyl ester–negative cells retainingTMRE dye was assessed by flow cytometry. Z-VAD-FMK(BD PharMingen), a general caspase inhibitor, wasincluded at 10 Amol/L in a subset of cocultures of targetcells with K562-ncFasL cells as a negative control.
Incubation of Primary Prostate Cancer Cells withMitoxantrone and Docetaxel
Mitoxantrone (Sigma) was reconstituted in water andstored at room temperature; docetaxel (Fluka) was pre-pared in DMSO and stored at �20jC. The IC50 for docetaxelwas determined to be 1,000 nmol/L after 48 h exposureusing T4 and T9 cells and analysis by MTT assays (data notshown). For Fas up-regulation and cytotoxicity assays,10 or 100 nmol/L mitoxantrone or 100 or 1,000 nmol/Ldocetaxel was added to plates of 70% confluent primaryprostate cells 24, 48, or 72 h before 51Cr release or flowcytometry assays.
p53 Inhibition AssayCells were cultured with 30 Amol/L pifithrin a (Sigma) for
24 h before the addition of 100 nmol/L mitoxantrone or1,000 nmol/L docetaxel and refreshed every 24 h. Forty-eight hours after mitoxantrone and docetaxel addition, cellswere collected for use in 51Cr release assays as before. Celllysates were probed for p53 expression by Western blotting.
Statistical AnalysisData are presented as mean F SD. Sample means were
compared using two-tailed unpaired Student’s t tests, witha significance level of P < 0.05. Correlation was measuredusing Spearman rank-order correlation on SigmaStatsoftware with P < 0.05.
ResultsAcquisition and Characterization of ProstateTissueCell cultures were established from needle biopsy cores
of tissue from palpable prostate tumors from 11 menundergoing radical prostatectomy. Ten of these cultureswere derived from Gleason score 7 tumors, whereas oneculture (T9) was derived from a Gleason score 9 (4, 5)tumor from a patient who had been receiving luteinizinghormone-releasing hormone therapy. To characterize thetype of cells populating the resulting cultures, immunoflu-orescence analysis was done to measure levels of basal
FasL-Induced Apoptosis in Primary Prostate Cancer Cells3020
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

epithelial cell markers (cytokeratins 5 + 14) and luminalepithelial cell markers (cytokeratins 8 + 18). Cell lines T3,T7, and T8 were analyzed as representative samples. Allcells were found to express both cytokeratins 5 + 14 and 8 +18 (Fig. 1A). Although 100% of the cells from the testedgroups co-expressed these cytokeratins, a small number ofcells were found to express cytokeratins 5 + 14 morestrongly than cytokeratins 8 + 18. To exclude the presenceof neuroendocrine or stromal cells in the primary prostatecancer cell cultures, chromogranin A, a neuroendocrine cellmarker, and smooth muscle actin, a stromal cell marker,levels were measured by Western blot. T3, T7, and T8 cellswere found to have undetectable levels of chromogranin Aand very low levels of a smooth muscle actin (Fig. 1B).These data, combined with the immunofluorescence data,suggest that the cultures were composed almost exclusivelyof epithelial cells with an insignificant presence of stromalcells in the cultures. Based on this expression profiling,the cultures consisted of cells that resemble an intermediatetransitional epithelial type (27).
During carcinogenesis, activation of pathways that driveproliferation can render cells resistant to apoptosis (28). Todetermine whether the rate of proliferation of primary
prostate cells might influence the incidence of apoptosis inthese cell lines, their relative rates of growth weremeasured by MTT assay. As expected, growth rates variedsomewhat between cell lines (Fig. 1C).
Primary Prostate Cell Cultures ExpressHigh Levels ofSurface Fas but No FasL
The first protein in the Fas-mediated apoptosis cascadeis the Fas receptor. To verify that cultured prostate cancercells express Fas, cells were analyzed by flow cytometry.High, uniform expression of Fas receptor was observed inall tested cells (MFI = 409.9 F 166.1 for Fas versus 10.7 F3.8 for isotype control; P < 0.01; Fig. 2A). A calibrationcurve to translate MFI values into ABC was generatedusing antibody-labeled microspheres with known numb-ers of antibody-binding sites. ABC was then used as asurrogate measure of the number of molecules of Fas onthe cell surface. The Fas expression on the prostate cancercell lines ranged from 52,475 ABC (T11) to 208,726 ABC(T4), with an average of 96,346 F 42,450 ABC. Of note,the Gleason score 9 tumor-derived cells (T9) hadsignificantly reduced levels of Fas receptor comparedwith the overall mean of all tested samples (MFI = 311.8for T9 versus 409.9 average; P < 0.05) as has been
Figure 1. Characterization of cultured human prostate cancer cells. A, expression of the basal epithelial cell marker cytokeratin 5 + 14 (red ) and theluminal epithelial cell marker cytokeratin 8 + 18 (green ) in T3 cells as measured by immunofluorescence. Overlay shows co-expression of these markers inthe majority of cells. At least four fields from each of cytokeratin 5 + 14– labeled, cytokeratin 8 + 18– labeled, and colabeled cells were analyzed.Magnification, �400. Similar results were obtained for T7 and T8 cells. B, expression of the neuroendocrine marker chromogranin A is not detectable in T3,T7, or T8 cell lysates by Western blot, and very low levels of the stromal marker a smooth muscle actin were detected after 40 min exposure.Chromogranin A–positive control: murine pancreatic lysate (panc. ); negative control: HeLa cell lysate; a-smooth muscle actin–positive control: NIH 3T3cell lysate. C, relative rates of growth of individual tumor cell lines were compared in a MTT assay. Cells (4 � 103) were plated in a 96-well plate and MTTassays performed 24 h later. Growth rates of PC-3 and nonmalignant PrEC cells are included for comparison. Average F SD of at least six replicates.
Molecular Cancer Therapeutics 3021
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

reported by others for advanced tumors (13). Surfaceexpression of FasL was not detected by flow cytometryunder these conditions (MFI = 4.0 F 1.0 for FasL versus5.3 F 1.8 for isotype control; Fig. 2A).
Characterization of Expression of Inhibitors ofApoptosis
Although expression of the Fas receptor is necessary forinduction of apoptosis by FasL, complex modulation of thispathway is asserted by protein inhibitors of apoptosis.Malignant cells often acquire dysfunctions in apoptoticsignaling caused by mutations in pro-apoptotic proteins orby overexpression of inhibitors of apoptosis (29). To evaluatethe levels of expression of several such inhibitors, Westernblots for bcl-2, c-FLIP, survivin, cIAP1, and cIAP2 proteinswere performed. The expression of p53 protein was detected
in all primary cell lines (Fig. 2B). Bcl-2 is often up-regulatedin prostate cancers (30), and robust expression of this proteinwas observed in T2, T4, T5, T6, T9, and T11 cells at levelscomparable with that of PC-3 cells (Fig. 2B). Conversely,low or no expression of bcl-2 was seen in T1, T8, T3, T7, orT10 cells. Expression of c-FLIP, survivin, cIAP1, and cIAP2mRNA was measured by quantitative reverse transcription-PCR (Fig. 2C). At both protein and mRNA levels, variableexpression of cIAP1, cIAP2, survivin, and c-FLIP mRNA wasobserved in the primary cell lines. Co-expression of cIAP1and cIAP2 or survivin and c-FLIP protein was stronglycorrelated (Spearman rank-order correlation rs z 0.6, P <0.05). Protein and mRNA expression of all tested inhibitorsof apoptosis was independent of levels of Fas expression(Spearman rank-order correlation, P > 0.05).
Figure 2. Characterization of ex-pression of pro-apoptotic and anti-apoptotic proteins. A, surface FasLand Fas expression was measured byflow cytometry on all 11 prostate celllines. Data shown are from T1 cellsand are representative of all samples.Solid histograms, cells labeled withPE-anti-FasL or APC-anti-Fas anti-bodies; outline histograms, isotypecontrols. B, representative Westernblot analysis of expression of bcl-2,c-FLIP, cIAP1, cIAP2, survivin, p53,and h-actin in primary prostate cells.293T cells engineered to overex-press bcl-2 (293T bcl-2 ) were usedas a positive control for the bcl-2Western. Densitometry values forthe inhibitors of apoptosis are shownnormalized for h-actin and relative toT1 levels. C, expression of inhibitorsof apoptosis was measured by quan-titative reverse transcription-PCR forc-FLIP, survivin, cIAP1, and cIAP2.Columns, averages of at least sixreplicates; bars, SD. mRNA expres-sion (ng/reaction) was normalized tothat of 18s rRNA in each case.
FasL-Induced Apoptosis in Primary Prostate Cancer Cells3022
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from
Prostate Cell Cultures Undergo Fas-MediatedApoptosis
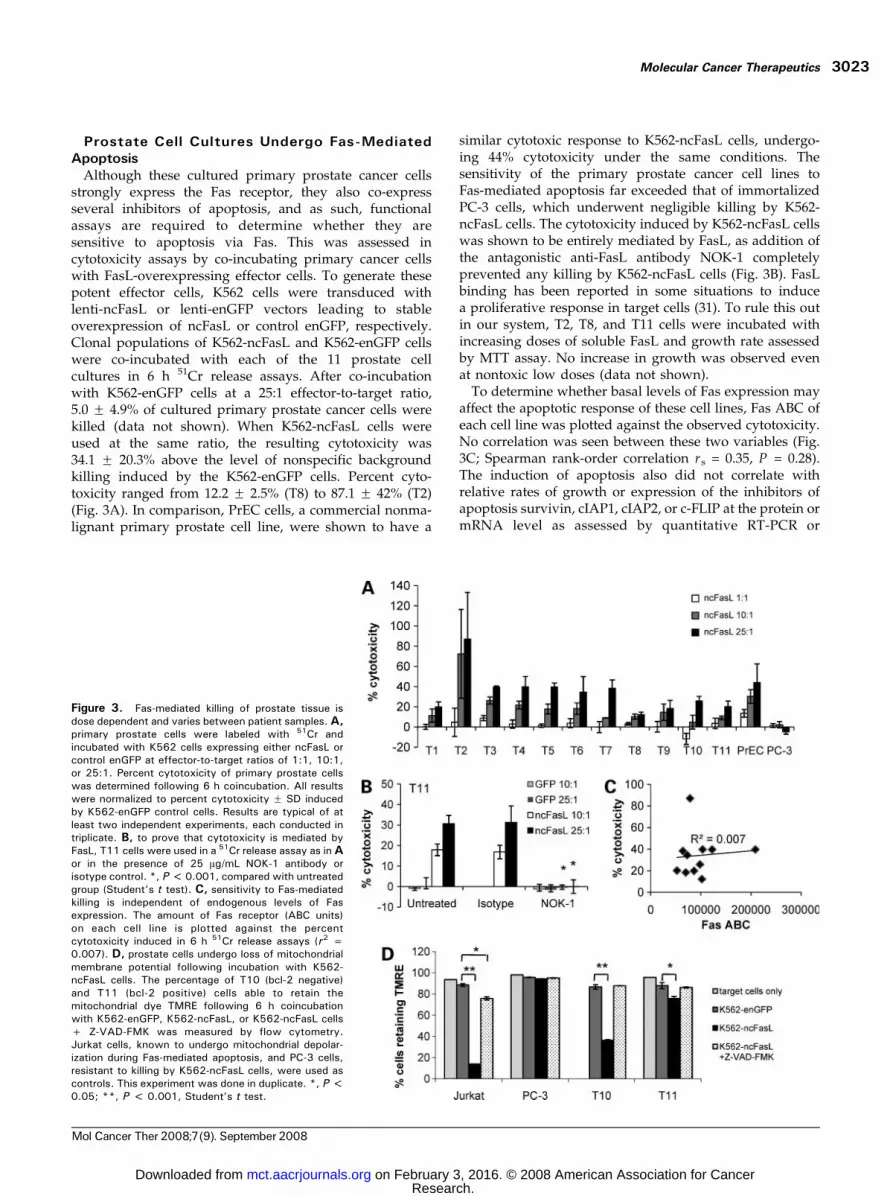
Although these cultured primary prostate cancer cellsstrongly express the Fas receptor, they also co-expressseveral inhibitors of apoptosis, and as such, functionalassays are required to determine whether they aresensitive to apoptosis via Fas. This was assessed incytotoxicity assays by co-incubating primary cancer cellswith FasL-overexpressing effector cells. To generate thesepotent effector cells, K562 cells were transduced withlenti-ncFasL or lenti-enGFP vectors leading to stableoverexpression of ncFasL or control enGFP, respectively.Clonal populations of K562-ncFasL and K562-enGFP cellswere co-incubated with each of the 11 prostate cellcultures in 6 h 51Cr release assays. After co-incubationwith K562-enGFP cells at a 25:1 effector-to-target ratio,5.0 F 4.9% of cultured primary prostate cancer cells werekilled (data not shown). When K562-ncFasL cells wereused at the same ratio, the resulting cytotoxicity was34.1 F 20.3% above the level of nonspecific backgroundkilling induced by the K562-enGFP cells. Percent cyto-toxicity ranged from 12.2 F 2.5% (T8) to 87.1 F 42% (T2)(Fig. 3A). In comparison, PrEC cells, a commercial nonma-lignant primary prostate cell line, were shown to have a
similar cytotoxic response to K562-ncFasL cells, undergo-ing 44% cytotoxicity under the same conditions. Thesensitivity of the primary prostate cancer cell lines toFas-mediated apoptosis far exceeded that of immortalizedPC-3 cells, which underwent negligible killing by K562-ncFasL cells. The cytotoxicity induced by K562-ncFasL cellswas shown to be entirely mediated by FasL, as addition ofthe antagonistic anti-FasL antibody NOK-1 completelyprevented any killing by K562-ncFasL cells (Fig. 3B). FasLbinding has been reported in some situations to inducea proliferative response in target cells (31). To rule this outin our system, T2, T8, and T11 cells were incubated withincreasing doses of soluble FasL and growth rate assessedby MTT assay. No increase in growth was observed evenat nontoxic low doses (data not shown).
To determine whether basal levels of Fas expression mayaffect the apoptotic response of these cell lines, Fas ABC ofeach cell line was plotted against the observed cytotoxicity.No correlation was seen between these two variables (Fig.3C; Spearman rank-order correlation rs = 0.35, P = 0.28).The induction of apoptosis also did not correlate withrelative rates of growth or expression of the inhibitors ofapoptosis survivin, cIAP1, cIAP2, or c-FLIP at the protein ormRNA level as assessed by quantitative RT-PCR or
Figure 3. Fas-mediated killing of prostate tissue isdose dependent and varies between patient samples. A,primary prostate cells were labeled with 51Cr andincubated with K562 cells expressing either ncFasL orcontrol enGFP at effector-to-target ratios of 1:1, 10:1,or 25:1. Percent cytotoxicity of primary prostate cellswas determined following 6 h coincubation. All resultswere normalized to percent cytotoxicity F SD inducedby K562-enGFP control cells. Results are typical of atleast two independent experiments, each conducted intriplicate. B, to prove that cytotoxicity is mediated byFasL, T11 cells were used in a 51Cr release assay as inAor in the presence of 25 Ag/mL NOK-1 antibody orisotype control. *, P <0.001, compared with untreatedgroup (Student’s t test). C, sensitivity to Fas-mediatedkilling is independent of endogenous levels of Fasexpression. The amount of Fas receptor (ABC units)on each cell line is plotted against the percentcytotoxicity induced in 6 h 51Cr release assays (r2 =0.007). D, prostate cells undergo loss of mitochondrialmembrane potential following incubation with K562-ncFasL cells. The percentage of T10 (bcl-2 negative)and T11 (bcl-2 positive) cells able to retain themitochondrial dye TMRE following 6 h coincubationwith K562-enGFP, K562-ncFasL, or K562-ncFasL cells+ Z-VAD-FMK was measured by flow cytometry.Jurkat cells, known to undergo mitochondrial depolar-ization during Fas-mediated apoptosis, and PC-3 cells,resistant to killing by K562-ncFasL cells, were used ascontrols. This experiment was done in duplicate. *, P <0.05; **, P < 0.001, Student’s t test.
Molecular Cancer Therapeutics 3023
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

Western blot (Spearman rank-order correlation, P > 0.13 inall cases; data not shown).
Fas-mediated apoptosis can proceed through a mito-chondrial-dependent or mitochondrial-independent path-way depending on the cell type. Loss of mitochondrialmembrane potential is a hallmark of apoptotic signalingthrough the mitochondria. To examine the pathwayfollowed by Fas-mediated apoptosis in these prostatecancer cells, the integrity of the mitochondrial outermembrane was measured by the ability of these cells toretain the mitochondrial dye TMRE after co-incubationwith K562-ncFasL cells. Co-incubation of T10 and T11 cellswith K562-ncFasL cells resulted in a significant reduction inthe number of cells retaining TMRE dye, indicating a loss ofmembrane potential in these cells (Fig. 3D). The addition ofZ-VAD-FMK to block caspase activation and thus apoptoticsignaling prevented the loss of mitochondrial membranepotential.
Chemotherapeutic Drugs Mitoxantrone and Doce-taxel Increase Fas Expression
Chemotherapeutic agents have been observed to induceincreases in Fas receptor expression on various cancer celllines, which in some cases translates into increased Fas-mediated apoptosis (18). The potential of the anti-prostatecancer chemotherapeutics docetaxel and mitoxantrone toenhance cell surface expression of Fas on primary prostatecancer cell lines was investigated. Cell cultures wereexposed to increasing doses of mitoxantrone (1-100 nmol/L) or docetaxel (10-1,000 nmol/L) for 24, 48, or 72 h.Changes in Fas expression were measured by flowcytometry and calculated as the change in Fas MFI fortreated cells over that of untreated cells. The greatestincrease in Fas expression by T1 and T8 cells was seenfollowing a 72 h exposure to 100 nmol/L mitoxantrone (3.2-fold increase in MFI; Fig. 4A and B). Similar results werefound for T3 and T4 cells (data not shown). Pretreatmentof prostate cells with docetaxel resulted in a 1.5-foldincrease in Fas expression at both 100 and 1,000 nmol/Lconcentrations, with no evidence of dose-dependency atthese concentrations (Fig. 4C, T8 cells). Enhancement ofFas expression in this manner significantly increased theABC of T8 cells to 215,138 F 9,172 ABC after 48 hexposure to 100 nmol/L docetaxel compared to the basalexpression level of 139,385 F 10,283 ABC (P < 0.05). Inaccordance with published reports (32), irradiation ofprostate cells was also found to lead to increased Fasexpression in these cells, with a 1.4-fold increase in MFI inT1 cells 72 h after irradiation with 8 Gy in our hands(data not shown).
Combination of Mitoxantrone and Docetaxel withFasLTherapy Leads to Increased Cell Killing
The effect of mitoxantrone- and docetaxel-mediated Fasup-regulation on the effectiveness of Fas-induced apoptosiswas tested on T3, T4, T5, T8, and T11 cells. Increases in Fas-mediated apoptosis of mitoxantrone-treated T3, T8, andT11 cells peaked following 48 h treatment with 100 nmol/Lmitoxantrone, resulting in a 1.5- to 3-fold enhancementof cytotoxicity following combination treatment (Fig. 5A).
When the same cells were pretreated with docetaxel before51Cr release assays, no increase in Fas-mediated killing ofT3 or T8 cells was seen despite similar increases in cellsurface Fas expression (Fig. 5B). In fact, small butsignificant decreases were actually seen in Fas-mediatedapoptosis following docetaxel treatment in these cells (P <0.05). When T11 cells were used in this assay, again nodifference in apoptosis induction was seen at 24 h (Fig. 5B).However, increases in cytotoxicity of 1.4-fold were ob-served by 48 and 72 h after addition of 1,000 nmol/Ldocetaxel (Fig. 5B; P < 0.05). One activity of docetaxel is theinhibition of bcl-2 (33). Whereas T11 prostate cancer cells
Figure 4. Fas expression is up-regulated on primary human prostateepithelial cells following clinically relevant treatment. A, T1 cells werelabeled with anti-Fas antibody and examined by flow cytometry forexpression of Fas receptor 24 and 48 h after treatment with mitoxantrone(filled histogram, untreated; gray line, 1 nmol/L; black line, 10 nmol/L;dashed line, 100 nmol/L). Relative increase in Fas MFI of T8 cellsfollowing treatment with (B) mitoxantrone (1, 10, or 100 nmol/L) or (C)docetaxel (100 and 1,000 nmol/L) compared with untreated cells. Flowcytometry was done 24, 48, or 72 h following addition of drug. M,mitoxantrone (n = 3); D, docetaxel (n = 2). *, P < 0.05, compared withuntreated controls for each time point (Student’s t test).
FasL-Induced Apoptosis in Primary Prostate Cancer Cells3024
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

expressed high levels of bcl-2, T3 and T8 cells expressedlow levels or no detectable bcl-2 (Fig. 2B). Similar increasesin Fas-mediated cytotoxicity were obtained when bcl-2-expressing T4 cells were pretreated with docetaxel (datanot shown). No further improvements in FasL-inducedcytotoxicity of T8 or T11 cells were observed whenmitoxantrone and docetaxel were added simultaneously(data not shown).
Mechanism of Sensitization to Fas-Mediated Apop-tosis Involves p53
Induction of p53 by DNA-damaging agents has beenreported to stimulate expression of the Fas gene and torender certain cell types more sensitive to Fas-mediatedapoptosis (34). To verify that the mechanism of sensitiza-tion of primary prostate cancer cells to Fas-mediated killingis dependent on p53 activity, apoptosis experiments wererepeated in the presence of pifithrin a, a chemical inhibitorof p53. T11 cells were treated with pifithrin a for 24 h beforethe commencement of 48 h mitoxantrone or docetaxeltreatment. Both the basal expression level of p53 and itsinduction by mitoxantrone and docetaxel were reduced bypifithrin a treatment (Fig. 6A). When 51Cr release assayswere done using pifithrin a– treated cells, a generalreduction in the level of cytotoxicity was observed. As
well, any increase in cytotoxicity induced by docetaxelwas completely prevented, whereas sensitization to Fas-mediated apoptosis by mitoxantrone treatment was limited(Fig. 6B). These data show the important role of p53 insensitizing primary prostate cancer cells to Fas-mediatedapoptosis.
DiscussionThis study shows for the first time that primary humanprostate cancer cells are sensitive to Fas-mediated killingand that this killing can be further improved by pretreat-ment of the cells with mitoxantrone and, in select cases,docetaxel. Fas signaling induces depolarization of themitochondrial outer membrane, suggesting a mitochondri-al-dependent mechanism behind this killing. Cytotoxicitystudies revealed no difference in the ability of six of ourprimary tumor cell lines and PrEC normal prostate epithelialcells to undergo apoptosis in response to FasL. This suggeststhat malignant transformation may not necessarily lead toacquisition of resistance to Fas-mediated killing at this stageof disease. The sensitivity of all 11 primary prostate cancercell lines to apoptosis was found to exceed that of PC-3 cells,revealing that the use of immortalized prostate cancer cells
Figure 5. Pretreatment of primaryhuman prostate cancer cells withmitoxantrone increases sensitivity toFas-mediated killing. Bcl-2-negativeT8 and T3 cells and bcl-2-expressingT11 cells were treated for 24, 48, or72 h with (A) 10 or 100 nmol/Lmitoxantrone or (B) 100 or 1,000nmol/L docetaxel before assessmentof sensitivity to FasL-mediated killingin 6 h 51Cr release assays. Drug-treated T3, T8, and T11 cells wereco-incubated with K562-ncFasL orK562-enGFP cells at 1:1, 10:1, or25:1 effector-to-target cell ratios.Percent cytotoxicity was calculatedas in Fig. 3, and target cell cytotox-icity induced following incubationwith K562-ncFasL cells at a 25:1ratio is shown, normalized to cyto-toxicity values for target cells nottreated with drug. Triplicate samplesfrom one of two representative experi-ments are shown. *, P < 0.05,Student’s t test.
Molecular Cancer Therapeutics 3025
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

in functional assays may underestimate the responsivenessof prostate cancer to Fas-mediated apoptosis. This resultprovides important evidence validating the suitability ofFasL-based therapies for prostate cancer and suggests thatnovel combinatorial approaches with chemotherapy mightbe most effective.
Encouragingly, we have shown that apoptosis occursreadily in Gleason score 7 prostate tumor-derived cellsdespite the expression of inhibitors of apoptosis in many ofthese primary lines. c-FLIP expression has been shown tocause resistance of prostate DU145 cells to Fas-mediatedapoptosis (35) and is sufficient to induce TRAIL resistancein prostate cancer cells (36). IAP expression is alsoassociated with resistance to apoptosis (37). In this study,simultaneous expression of a number of apoptosis inhib-itors was observed; however, we did not show a correlationbetween expression of inhibitors and induction of Fas-mediated killing. Furthermore, the primary human prostatecell lines with the lowest response to Fas, T1 (19.9%) and T8(12.2%), were found to be bcl-2 negative, whereas the mostsensitive cell line, T2 (87.1%), expressed abundant bcl-2.These observations suggest that measurement of expres-
sion of only these inhibitors of apoptosis is not a validpredictor of apoptotic responses in these primary cells.
The observed lack of correlation between apoptosisinduction and inhibitor expression may be due to thelimited sample size studied or may result from the complexcellular mechanisms controlling apoptosis. In addition tothe inhibitors studied, overexpression of other bcl-2 familyproteins, dysregulation of mitogen-activated protein kinaseor phosphatidylinositol 3-kinase activity, expression ofdecoy receptors, or deletion/mutation of caspases havebeen described in cancer (38). Thus, although expression ofIAP proteins or c-FLIP may limit apoptosis in some celllines, other factors may be more influential in regulatingthis important cellular process. In addition, protein andmRNA expression was measured from pooled lysates andmRNA. It is possible that any cells surviving after 6 h in ourassays will express these proteins at higher levels than thecells that were readily killed. Future studies will addressthis issue.
In our study, mitoxantrone, a topoisomerase II inhibitorand molecule that integrates into DNA, along withdocetaxel, a microtubule-stabilizing agent, were both
Figure 6. Fas-upregulation and sensitization to Fas-mediated apoptosis can be prevented in primary humanprostate cancer cells by inhibition of p53. A, treatmentof cells with 30 Amol/L pifithrin a reduces basal p53expression in T8 and T11 cells and limits p53accumulation after 48 h mitoxantrone and docetaxeltreatment as measured by Western blot. Densitometryvalues are plotted relative to untreated cells. B, both thebasal cytotoxicity and the increase in apoptosis seenfollowing mitoxantrone and docetaxel treatment arelimited when pifithrin a is included in culture of T11cells. Triplicate samples from one of three representa-tive experiments are shown. U, untreated; M, 100nmol/L mitoxantrone; D, 1,000 nmol/L docetaxel; pa,pifithrin a; p + M, pifithrin a and mitoxantrone; p + D,pifithrin a and docetaxel. *, P < 0.05, compared withuntreated (Student’s t test).
FasL-Induced Apoptosis in Primary Prostate Cancer Cells3026
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

effective at increasing Fas receptor expression on primaryprostate cancer cells. However, only mitoxantrone was ableto improve the susceptibility of all tested primary prostatecultures to Fas-mediated apoptosis. In the majority of thecells tested, docetaxel did not improve sensitivity toapoptosis despite also enhancing Fas expression. Oneexplanation for this difference could be the existence of athreshold effect, as mitoxantrone was more effective atincreasing Fas expression than docetaxel (3.2-fold versus1.5-fold induction). Alternatively, mitoxantrone and doce-taxel may differentially affect the expression of multiplecell survival or apoptosis proteins. Indeed, we haveobserved a change in expression in 110 proteins in primaryprostate cancer cells on addition of mitoxantrone.5 Thisconcept is backed up by the lack of correlation in this studybetween basal Fas expression and cytotoxicity (Fig. 3C).Furthermore, one observed mechanism of action ofdocetaxel is the inactivation of bcl-2 (39). In our study,docetaxel pretreatment was only found to increase apopto-sis induction by FasL-based therapy in bcl-2-expressingcells (Fig. 5B). In contrast, mitoxantrone was able tosensitize both bcl-2-negative and bcl-2-expressing cells toFas-mediated killing. It is thus possible that docetaxel-induced inactivation of bcl-2 is a more potent sensitizerto apoptosis than docetaxel-induced Fas up-regulation.Further investigation into the effect of these agents onapoptotic pathways is warranted, as the molecular profileof an individual cancer is likely to influence the success ofany apoptosis-based therapeutic strategy. It is also possiblethat no single apoptosis-sensitizing agent will be optimalfor all tumors.
Expression of p53 protein was detected in all cell linestested by Western blot analyses (Fig. 2B). Addition ofpifithrin a, an inhibitor of p53, to cultures of T8 and T11 cellslimited the induction of apoptosis and the sensitization toFas-mediated killing by docetaxel and mitoxantrone. p53has been implicated in control of Fas expression in severalcell models (20). Other studies show that combinationcisplatin treatment and Fas-triggering induced apoptosis inp53-mutant cell lines (19). In the present study, apoptosiswas not completely inhibited by pifithrin a, suggesting thatalthough p53 plays a role in sensitivity to apoptosis, loss ofp53 activity does not completely abrogate this process.
FasL-based therapies will require careful design to avoidoff-target toxicity. This can be addressed by introducingstringent delivery, expression, and activation controls onFasL. Several such approaches have been used withsuccess, including the development of gene therapy-basedstrategies engineering tissue-specific FasL expression (8),bispecific anti-Fas antibodies that aggregate and becomeactive only in the presence of a specified tumor antigen(9, 10), and recombinant FasL prodrug constructs that arefunctional only at the tumor site (11). These innovationsin FasL-based drug design are likely to culminate in amolecule with a more favorable toxicity profile and are
bolstering enthusiasm for these types of signaling-directedanticancer approaches. In addition, as we have shown thatchemotherapy is able to improve the therapeutic efficacy ofFasL, controlled delivery of chemotherapeutic agents couldbe used to further direct the specificity of Fas-mediatedtoxicity. For example, nanoparticle-based drug deliverymethods are under development, which, by incorporationof antigen- or receptor-targeting molecules into the particlesurface, can deliver high levels of localized drug to specifictarget cells (40). In this way, tumor-specific targeting can beachieved, reducing toxicity to nonmalignant cells.
In conclusion, we have shown in primary prostate cellsthat Fas-based approaches to prostate cancer cell killing arepossible. Mitoxantrone and docetaxel were shown tofurther improve this killing. Although neither mitoxan-trone nor docetaxel is curative for prostate cancer inthemselves, their use in combination with FasL-basedtherapies could potentially increase anticancer efficacy.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Acknowledgments
We thank prostate cancer patients at the University Health Network forcontributing samples and Karen Hersey and Susie Lehnerz for assistance inobtaining informed consent.
References
1. Nakanishi H, Mazda O, Satoh E, et al. Nonviral genetic transfer of Fasligand induced significant growth suppression and apoptotic tumor celldeath in prostate cancer in vivo . Gene Ther 2003;10:434–42.
2. Hedlund TE, Meech SJ, Srikanth S, et al. Adenovirus-mediatedexpression of Fas ligand induces apoptosis of human prostate cancercells. Cell Death Differ 1999;6:175–82.
3. Rubinchik S, Wang D, Yu H, et al. A complex adenovirus vector thatdelivers FASL-GFP with combined prostate-specific and tetracycline-regulated expression. Mol Ther 2001;4:416–26.
4. Schimmer AD, Thomas MP, Hurren R, et al. Identification of smallmolecules that sensitize resistant tumor cells to tumor necrosis factor-family death receptors. Cancer Res 2006;66:2367–75.
5. Hyer M, Sudarshan S, Schwartz DA, Hannun Y, Dong J, Norris JS.Quantification and characterization of the bystander effect in prostatecancer cells following adeno-virus-mediated FasL expression. Cancer GeneTher 2003;10:330–9.
6. Rokhlin OW, Bishop GA, Hostager BS, et al. Fas-mediated apoptosis inhuman prostatic carcinoma cell lines. Cancer Res 1997;57:1758–68.
7. Hedlund TE, Duke RC, Schleicher MS, Miller GJ. Fas-mediatedapoptosis in seven human prostate cancer cell lines: correlation withtumor stage. Prostate 1998;36:92–101.
8. Aoki K, Akyurek LM, San H, et al. Restricted expression of anadenoviral vector encoding Fas ligand (CD95L) enhances safety for cancergene therapy. Mol Ther 2000;1:555–65.
9. Herrmann T, Grosse-Hovest L, Otz T, Krammer PH, Rammensee HG,Jung G. Construction of optimized bispecific antibodies for selectiveactivation of the death receptor CD95. Cancer Res 2008;68:1221–7.
10. Samel D, Muller D, Gerspach J, et al. Generation of a FasL-basedproapoptotic fusion protein devoid of systemic toxicity due to cell-surfaceantigen-restricted Activation. J Biol Chem 2003;278:32077–82.
11. Watermann I, Gerspach J, Lehne M, et al. Activation of CD95L fusionprotein prodrugs by tumor-associated proteases. Cell Death Differ 2007;14:765–74.
12. Sasaki Y, Ahmed H, Takeuchi T, Moriyama N, Kawabe K. Immuno-histochemical study of Fas, Fas ligand and interleukin-1b convertingenzyme expression in human prostatic cancer. Br J Urol 1998;81:852–5.5 Manuscript under revision.
Molecular Cancer Therapeutics 3027
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

13. Jiang J, Ulbright TM, Zhang S, et al. Fas and Fas ligand expression iselevated in prostatic intraepithelial neoplasia and prostatic adenocarcino-ma. Cancer 2002;95:296–300.
14. Krajewska M, Krajewski S, Banares S, et al. Elevated expression ofinhibitor of apoptosis proteins in prostate cancer. Clin Cancer Res 2003;9:4914–25.
15. McEleny KR, Watson RW, Coffey RN, O’Neill AJ, Fitzpatrick JM.Inhibitors of apoptosis proteins in prostate cancer cell lines. Prostate2002;51:133–40.
16. Costa-Pereira AP, McKenna SL, Cotter TG. Activation of SAPK/JNKby camptothecin sensitizes androgen-independent prostate cancer cells toFas-induced apoptosis. Br J Cancer 2000;82:1827–34.
17. Chatterjee D, Schmitz I, Krueger A, et al. Induction of apoptosis in9-nitrocamptothecin-treated DU145 human prostate carcinoma cellscorrelates with de novo synthesis of CD95 and CD95 ligand and down-regulation of c-FLIP(short). Cancer Res 2001;61:7148–54.
18. Liu QY, Rubin MA, Omene C, Lederman S, Stein CA. Fas ligand isconstitutively secreted by prostate cancer cells in vitro . Clin Cancer Res1998;4:1803–11.
19. Bagnoli M, Balladore E, Luison E, et al. Sensitization of p53-mutated epithelial ovarian cancer to CD95-mediated apoptosis is syner-gistically induced by cisplatin pretreatment. Mol Cancer Ther 2007;6:762–72.
20. Muller M, Wilder S, Bannasch D, et al. p53 activates the CD95 (APO-1/Fas) gene in response to DNA damage by anticancer drugs. J Exp Med1998;188:2033–45.
21. Romano C, De Fanis U, Sellitto A, et al. Induction of CD95upregulation does not render chronic lymphocytic leukemia B-cellssusceptible to CD95-mediated apoptosis. Immunol Lett 2005;97:131–9.
22. Peehl DM, Stamey TA. Serum-free growth of adult human prostaticepithelial cells. In Vitro Cell Dev Biol 1986;22:82–90.
23. Peehl D. Growth of prostatic epithelial and stromal cells in vitro . In:Russel P, Jackson P, Kingsley E, editors. Prostate cancer methods andprotocols. Totowa (NJ): Humana Press; 2003. p. 41–57.
24. Siatskas C, Underwood J, Ramezani A, Hawley RG, Medin JA.Specific pharmacological dimerization of KDR in lentivirally transducedhuman hematopoietic cells activates anti-apoptotic and proliferativemechanisms. FASEB J 2005;19:1752–4.
25. Yoshimitsu M, Sato T, Tao K, et al. Bioluminescent imaging of amarking transgene and correction of Fabry mice by neonatal injection ofrecombinant lentiviral vectors. Proc Natl Acad Sci U S A 2004;101:16909–14.
26. Jayaraman S. Flow cytometric determination of mitochondrialmembrane potential changes during apoptosis of T lymphocytic and
pancreatic h cell lines: comparison of tetramethylrhodamineethylester(TMRE), chloromethyl-X-rosamine (H2-CMX-Ros) and MitoTracker Red580 (MTR580). J Immunol Methods 2005;306:68–79.
27. Verhagen AP, Ramaekers FC, Aalders TW, Schaafsma HE, DebruyneFM, Schalken JA. Colocalization of basal and luminal cell-type cytoker-atins in human prostate cancer. Cancer Res 1992;52:6182–7.
28. Mimeault M, Pommery N, Henichart JP. Synergistic antiproliferativeand apoptotic effects induced by epidermal growth factor receptor andprotein kinase A inhibitors in human prostatic cancer cell lines. Int J Cancer2003;106:116–24.
29. McKenzie S, Kyprianou N. Apoptosis evasion: the role of survivalpathways in prostate cancer progression and therapeutic resistance. J CellBiochem 2006;97:18–32.
30. Byrne RL, Horne CH, Robinson MC, et al. The expression of waf-1,p53 and bcl-2 in prostatic adenocarcinoma. Br J Urol 1997;79:190–5.
31. Li H, Cai X, Fan X, Moquin B, Stoicov C, Houghton J. Fas Ag-FasLcoupling leads to ERK1/2-mediated proliferation of gastric mucosal cells.Am J Physiol Gastrointest Liver Physiol 2008;294:G263–75.
32. Shimada K, Nakamura M, Ishida E, Kishi M, Konishi N. Androgen andthe blocking of radiation-induced sensitization to Fas-mediated apoptosisthrough c-jun induction in prostate cancer cells. Int J Radiat Biol 2003;79:451–62.
33. Pienta K. Preclinical mechanisms of action of docetaxel and docetaxelcombinations in prostate cancer. Semin Oncol 2001;28:3–7.
34. Muller M, Strand S, Hug H, et al. Drug-induced apoptosis inhepatoma cells is mediated by the CD95 (APO-1/Fas) receptor/ligandsystem and involves activation of wild-type p53. J Clin Invest 1997;99:403–13.
35. Hyer ML, Sudarshan S, Kim Y, et al. Downregulation of c-FLIPsensitizes DU145 prostate cancer cells to Fas-mediated apoptosis. CancerBiol Ther 2002;1:401–6.
36. Zhang X, Jin TG, Yang H, DeWolf WC, Khosravi-Far R, Olumi AF.Persistent c-FLIP(L) expression is necessary and sufficient to maintainresistance to tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis in prostate cancer. Cancer Res 2004;64:7086–91.
37. LaCasse EC, Baird S, Korneluk RG, MacKenzie AE. The inhibitors ofapoptosis (IAPs) and their emerging role in cancer. Oncogene 1998;17:3247–59.
38. de Thonel A, Eriksson JE. Regulation of death receptors—relevance incancer therapies. Toxicol Appl Pharmacol 2005;207:123–32.
39. Haldar S, Chintapalli J, Croce C. Taxol induced bcl-2 phosphorylationand death of prostate cancer cells. Cancer Res 1996;56:1253–5.
40. Haley B, Frenkel E. Nanoparticles for drug delivery in cancertreatment. Urol Oncol 2008;26:57–64.
FasL-Induced Apoptosis in Primary Prostate Cancer Cells3028
Mol Cancer Ther 2008;7(9). September 2008
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from

2008;7:3018-3028. Mol Cancer Ther Juliane C. Symes, Michael Kurin, Neil E. Fleshner, et al. increased by mitoxantrone and docetaxelFas-mediated killing of primary prostate cancer cells is
Updated version
http://mct.aacrjournals.org/content/7/9/3018
Access the most recent version of this article at:
Cited articles
http://mct.aacrjournals.org/content/7/9/3018.full.html#ref-list-1
This article cites 39 articles, 15 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on February 3, 2016. © 2008 American Association for Cancermct.aacrjournals.org Downloaded from