experimental cutaneous leishmaniasis: transmission electron microscopy of the inoculation site
TRANSCRIPT

Experimental cutaneous leishmaniasis: transmissionelectron microscopy of the inoculation site
CIDIA VASCONCELLOS AND MIRIAN N. SOTTOInfectious Disease Pathology Laboratory-LIM 50, Department of Pathology,
University of Sao Paulo Medical School, Sao Paulo, SP, Brazil
Received for publication 9 October 1996
Accepted for publication 18 December 1996
Summary. Tissue response against inoculation of Leishmania (Leishmania)amazonensis promastigotes in the hind footpad was quite different betweentwo strains of mice: in BALB/c animals there was parasitism of perineurialcells by the 8th week post inoculation (WPI) and heavy parasitism ofmacrophages, as well as degenerated extracellular parasites close to col-lagen fibers at the 39th WPI, whereas in C57Bl/6j mice there was heavyparasitism of macrophages at 6th WPI, dermal vessels with high endothelialcell at 21st WPI and well preserved intracellular amastigote forms by 51stWPI. In both animals there was no parasitism of keratinocytes or Langerhanscells. Thus BALB/c mice were useful as an experimental model for diffusecutaneous leishmaniasis and showing a new feature, parasitism of perineurialcells, whereas C57Bl/6j animals show hypersensitivity signs, together with afew preserved parasites, only late in the course of infection. From a morpho-logical point of view, there were no differences in macrophages, or in theinteraction between this target cell and the parasite, between the animalmodels studied. This suggests that the difference in the response of the hoststowards the parasite could depend on the way in which they activate acellular, i.e. lymphocyte mediated immune, response.
Keywords: cutaneous experimental leishmaniasis, animal models,electron microscopy
There have been several ultrastructural studies docu-menting the identification of Leishmania parasitism ofinflammatory cells (Pearson & Steigbigel 1981; Ridley &Wells 1986; Pimenta et al. 1987; Bittencourt et al. 1990)and the effect of anti-Leishmania agents (Bhatnagar etal. 1991). Neural compromise has seldom been studied,and only light microscopy was used (Satti et al. 1989; El-Hassan et al. 1992). Some authors (Mann & Harman1984) have drawn attention to the fact that paraesthesiais also present in other granulomatous processes of skin
besides leprosy, e.g. leishmaniasis, but have notexplored this further.
The aim of this study was to determine the ultrastruc-tural characteristics of the sequential tissue responseagainst Leishmania (Leishmania) amazonensis (Lainson& Shaw 1987) at the inoculation site in two strains ofmice: BALB/c (sensitive) and C57Bl/6j (resistant).
Materials and methods
Animals
Female 40-day-old inbred mice C57Bl/6j and BALB/c
Int. J. Exp. Path. (1997), 78, 81–89
81q 1997 Blackwell Science Ltd
Correspondence: Dr Cidia Vasconcellos, Rua Campevas, 639.CEP: 05016-010. Sao Paulo, SP, Brazil.
strains were used, both obtained from University of SaoPaulo Medical School General Colony. The animalswere maintained in plastic cages with wood shelvesand received proper food and water ad libitum.
Parasites
Leishmania (Leishmania) amazonensis (WHOM/BR/00 LTB 0016) (G. Grimaldi, Fiocruz, RJ). The strainwas sustained in the laboratory through sequentialpassages in BALB/c mice, culture in NNN/BHI medium
and reinoculation at the hind footpad in mice(Giorgio 1989).
Infection
Nine experimental groups of each strain, each withthree animals, were inoculated in the right hind footpadunder ether anaesthesia with 50 ml of saline solutioncontaining 107 promastigotes in the stationary growthphase. There was one control animal in each experi-mental group, inoculated under the same conditionswith 50 ml of sterile saline solution.
82 C. Vasconcellos and M.N. Sotto
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89
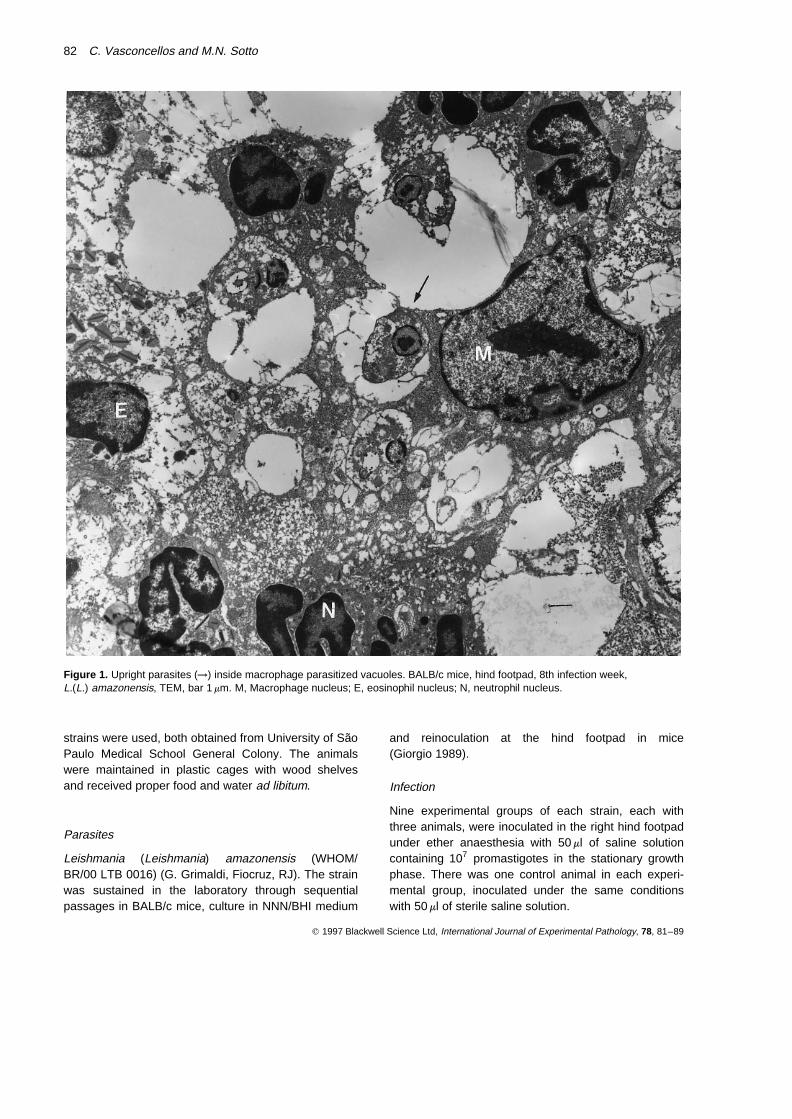
Figure 1. Upright parasites (→) inside macrophage parasitized vacuoles. BALB/c mice, hind footpad, 8th infection week,L.(L.) amazonensis, TEM, bar 1 mm. M, Macrophage nucleus; E, eosinophil nucleus; N, neutrophil nucleus.
Three inoculated animals and one control were killed1, 2, 3, 6, 8, 11, 21, 39 (for BALB/c mice) and 51 (forC57Bl/6j mice) weeks post inoculation (WPI).
Technique
All animals were autopsied with gross examination ofviscera.
Fragments from right hind footpad, skin, right pop-liteal lymph node, liver, spleen, kidneys, heart andlungs were processed for routine haematoxylin-eosinstaining, in order to rule out other diseases, mostlyinfections.
Fragments from the inoculated hind footpad wereprocessed for transmission electron microscopy (TEM)(Duarte et al. 1992).
Experimental cutaneous leishmaniasis: TEM of inoculation site 83
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89
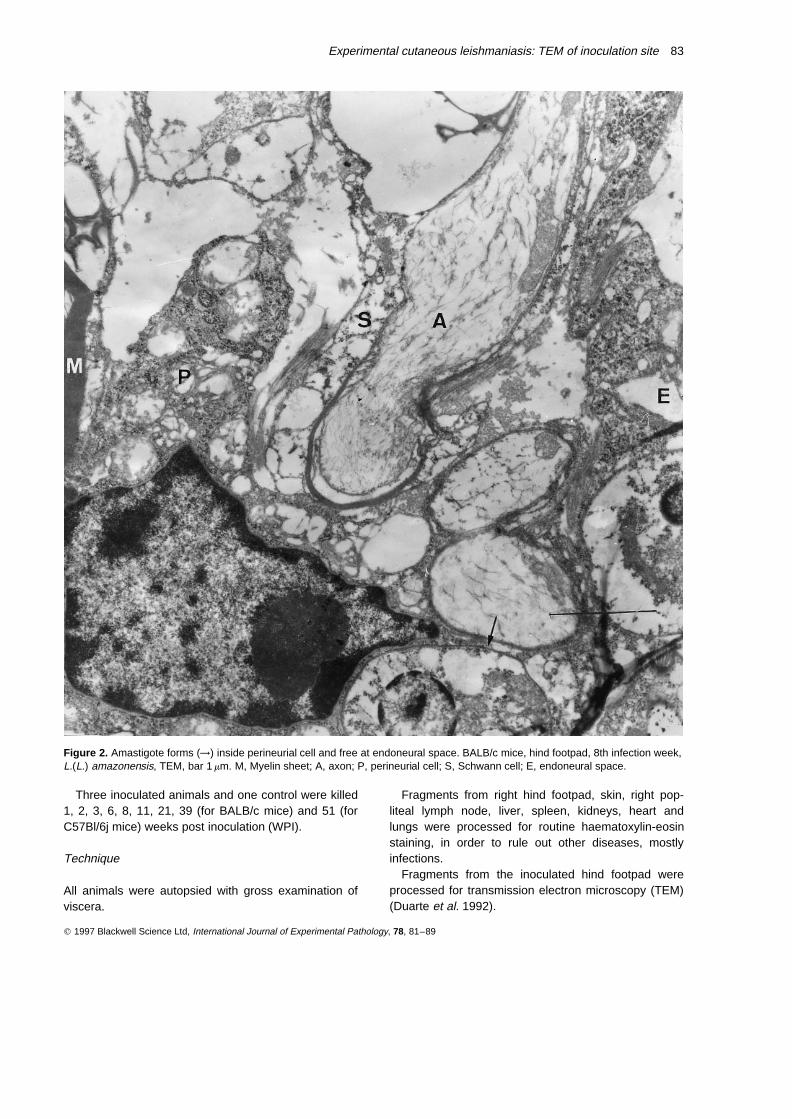
Figure 2. Amastigote forms (→) inside perineurial cell and free at endoneural space. BALB/c mice, hind footpad, 8th infection week,L.(L.) amazonensis, TEM, bar 1 mm. M, Myelin sheet; A, axon; P, perineurial cell; S, Schwann cell; E, endoneural space.
Results
BALB/c mice
Light microscopy examination of the inoculated hindfootpad showed intense macrophage parasitism. Thesecells were predominant in the dermal infiltrate. At the
electron microscope the upright parasites were groupedor isolated inside the vacuoles of parasitized cells(Figure 1). There were also extracellular parasitesamong the cellular debris. Other inflammatory elementswere neutrophils and eosinophils. Extracellular amasti-gote forms were observed closely related to delaminated
84 C. Vasconcellos and M.N. Sotto
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89
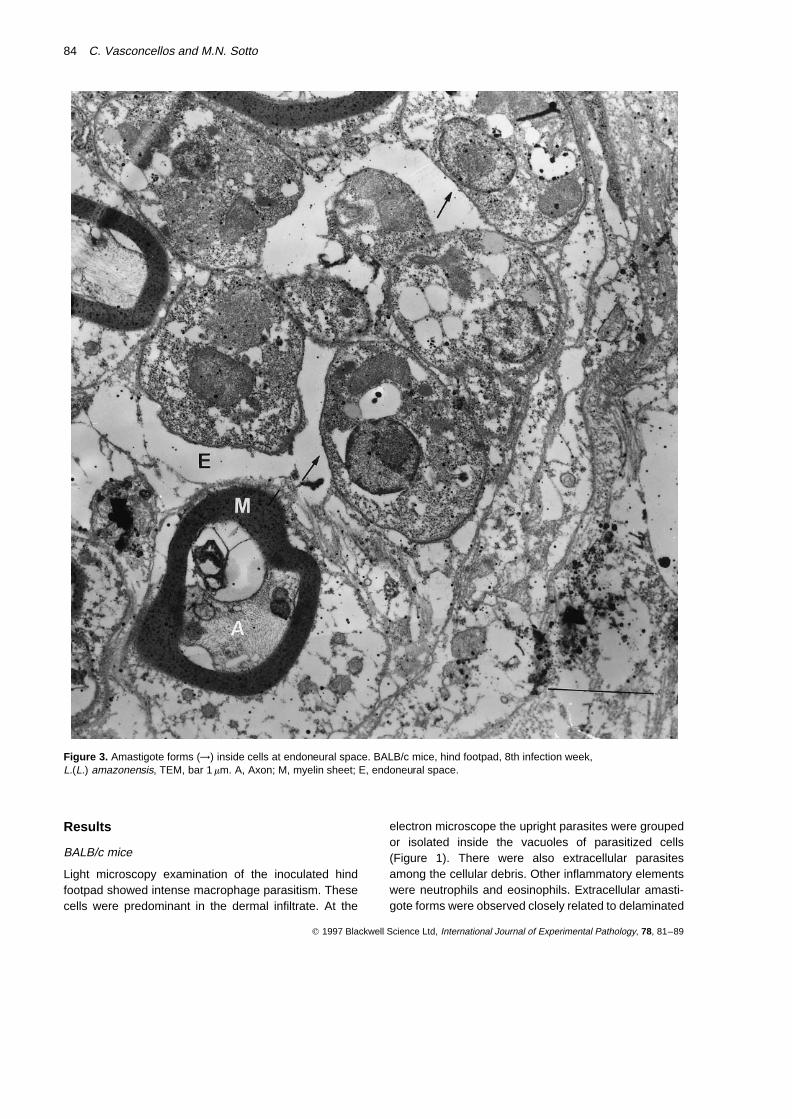
Figure 3. Amastigote forms (→) inside cells at endoneural space. BALB/c mice, hind footpad, 8th infection week,L.(L.) amazonensis, TEM, bar 1 mm. A, Axon; M, myelin sheet; E, endoneural space.
perineurial collagen sheets. There was parasitism ofperineurial cells (Figure 2) and of cells inside dermalneural endings with subcellular characteristics that didnot permit affirmation of their histogenetic nature(Figure 3). Those cells in endoneural spaces were veryclose to Schwann cells. The latter were well preservedand free of parasites. There were also dermal nerveendings with subcellular structures apparently preserved.
C57Bl/6j mice
C57Bl/6j mice disclosed a better organized tissueresponse at the inoculation site, defined after the 8thweek of infection by granuloma together with para-sitism of macrophages. After this time, there wasmorphologic evidence of mechanisms related tocellular immune response (organization of granulomaand dermal blood vessels with high endothelial cell)
Experimental cutaneous leishmaniasis: TEM of inoculation site 85
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89
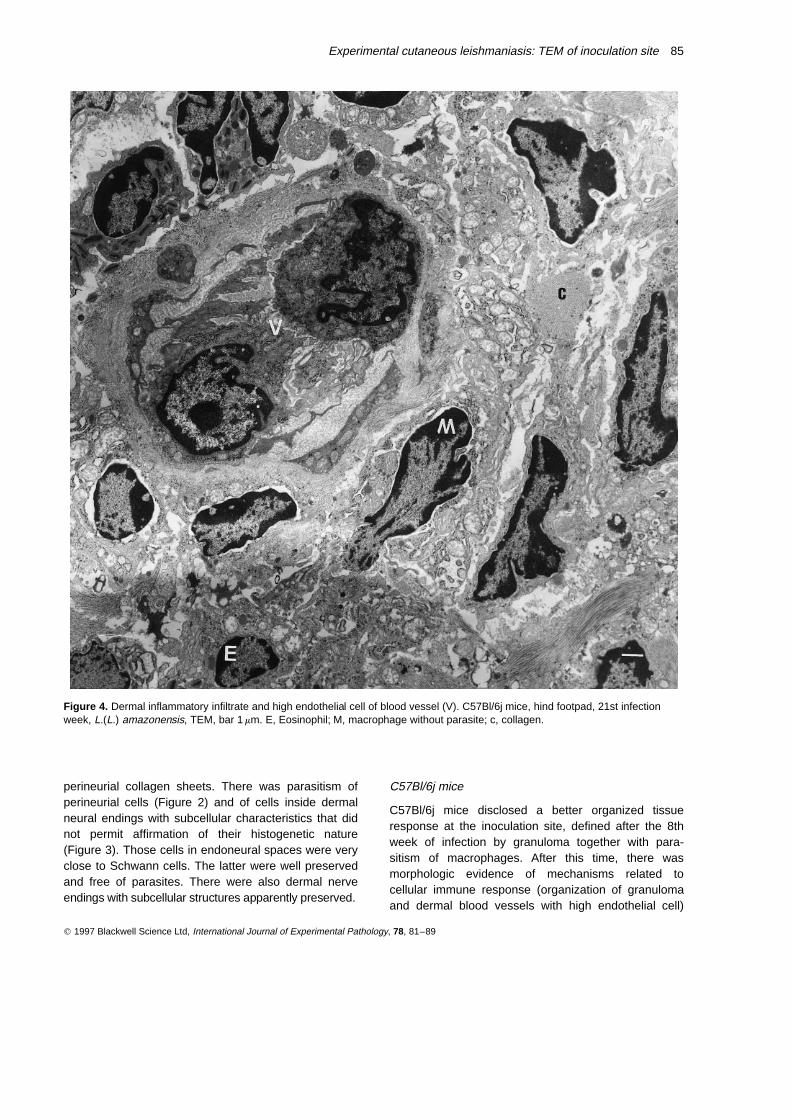
Figure 4. Dermal inflammatory infiltrate and high endothelial cell of blood vessel (V). C57Bl/6j mice, hind footpad, 21st infectionweek, L.(L.) amazonensis, TEM, bar 1 mm. E, Eosinophil; M, macrophage without parasite; c, collagen.
(Figure 4) and morphologic characteristics suggestive ofhypersensitivity (fibrinoid degeneration of blood vesselsand epidermal infiltration by inflammatory cells). Necro-sis was more intense in this animal model with transepi-dermic elimination of parasites by the 21st week ofinfection. The granuloma surrounded foci of necrosis,being partially substituted by fibrous tissue.
The epidermis was also more involved in this model
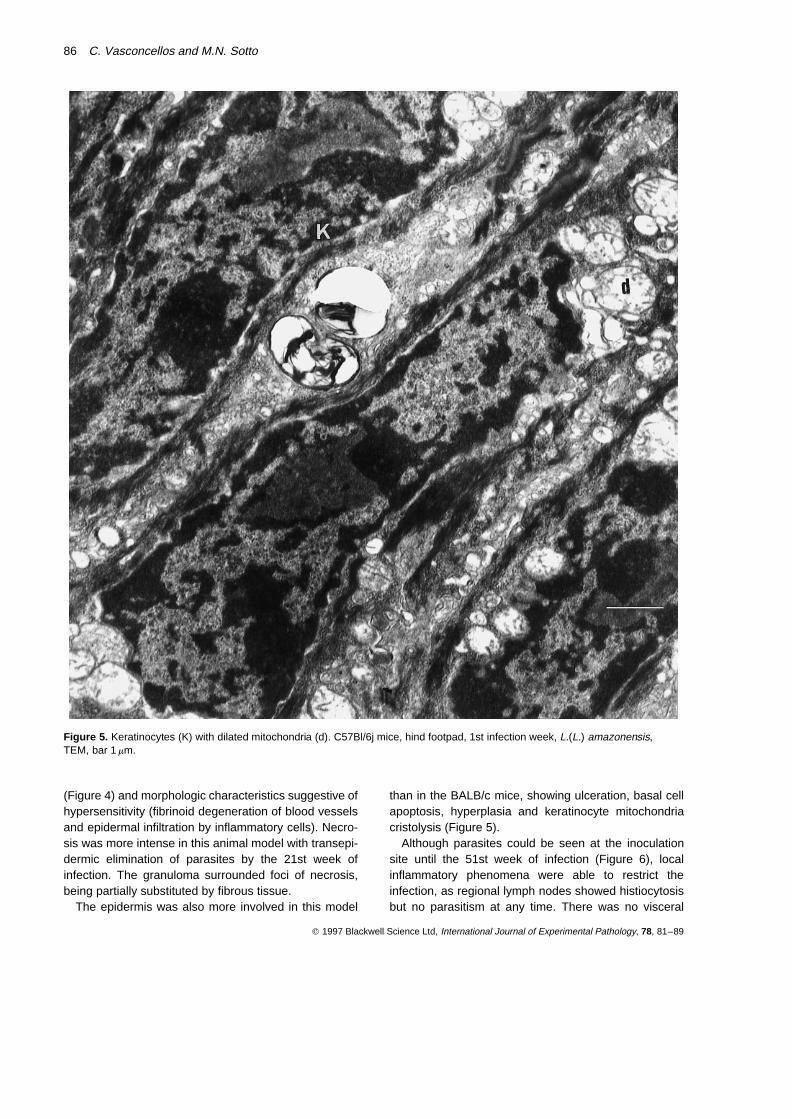
than in the BALB/c mice, showing ulceration, basal cellapoptosis, hyperplasia and keratinocyte mitochondriacristolysis (Figure 5).
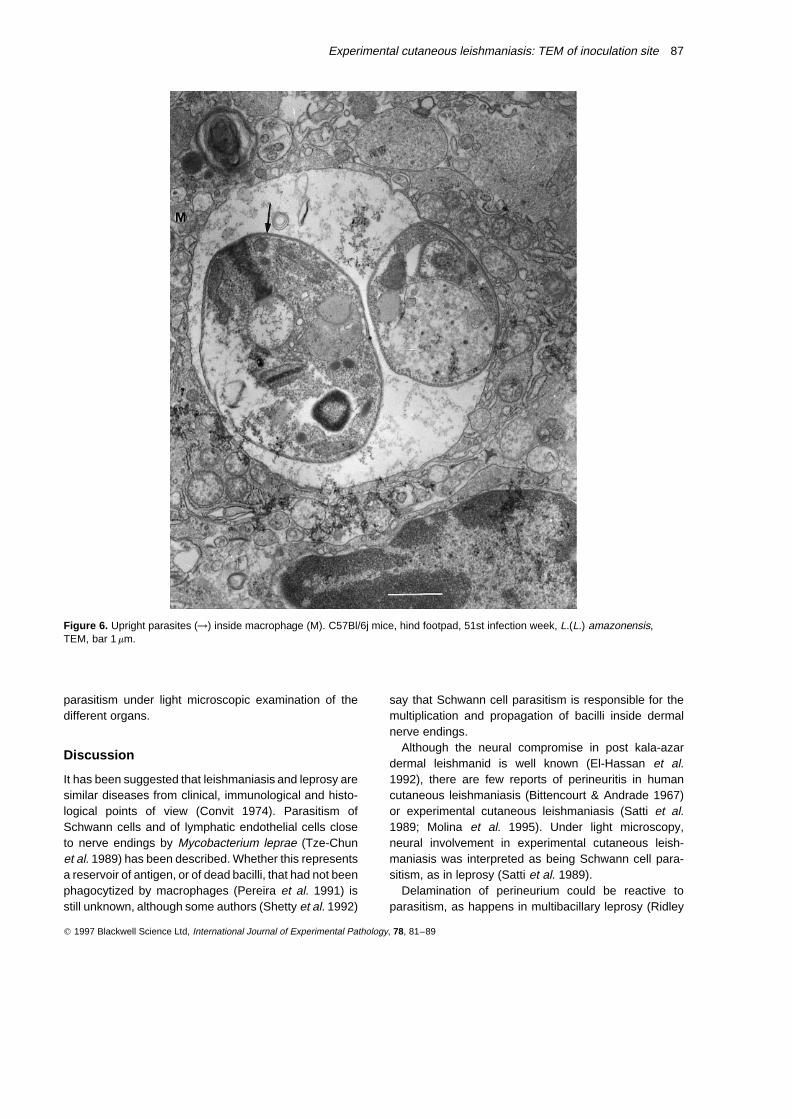
Although parasites could be seen at the inoculationsite until the 51st week of infection (Figure 6), localinflammatory phenomena were able to restrict theinfection, as regional lymph nodes showed histiocytosisbut no parasitism at any time. There was no visceral
86 C. Vasconcellos and M.N. Sotto
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89
Figure 5. Keratinocytes (K) with dilated mitochondria (d). C57Bl/6j mice, hind footpad, 1st infection week, L.(L.) amazonensis,TEM, bar 1 mm.
parasitism under light microscopic examination of thedifferent organs.
Discussion
It has been suggested that leishmaniasis and leprosy aresimilar diseases from clinical, immunological and histo-logical points of view (Convit 1974). Parasitism ofSchwann cells and of lymphatic endothelial cells closeto nerve endings by Mycobacterium leprae (Tze-Chunet al. 1989) has been described. Whether this representsa reservoir of antigen, or of dead bacilli, that had not beenphagocytized by macrophages (Pereira et al. 1991) isstill unknown, although some authors (Shetty et al. 1992)
say that Schwann cell parasitism is responsible for themultiplication and propagation of bacilli inside dermalnerve endings.
Although the neural compromise in post kala-azardermal leishmanid is well known (El-Hassan et al.1992), there are few reports of perineuritis in humancutaneous leishmaniasis (Bittencourt & Andrade 1967)or experimental cutaneous leishmaniasis (Satti et al.1989; Molina et al. 1995). Under light microscopy,neural involvement in experimental cutaneous leish-maniasis was interpreted as being Schwann cell para-sitism, as in leprosy (Satti et al. 1989).
Delamination of perineurium could be reactive toparasitism, as happens in multibacillary leprosy (Ridley
Experimental cutaneous leishmaniasis: TEM of inoculation site 87
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89
Figure 6. Upright parasites (→) inside macrophage (M). C57Bl/6j mice, hind footpad, 51st infection week, L.(L.) amazonensis,TEM, bar 1 mm.

& Job 1985). We, however, did not find neural destruc-tion at this phase of experimental infection as describedby others (Satti et al. 1989).
Evolution of the infection until the 39th week displayeda spontaneous recovery of the neuritis, the last animalsshowing no parasites inside nerve endings of the dermis.This leads us to propose that the transient perineurial cellparasitism represents a defence mechanism of theinvaded host, that will later destroy Leishmania.
The epidermis is a true participant of immuneresponse through keratinocyte activation as antigen pre-senting cell and/or secretion of cytokines and adhesionmolecules that modulate local immune response (Lugeret al. 1981; Schmitt et al. 1982; Kupper et al. 1986; Tapiaet al. 1989; Fine 1990; Schmitt 1990; Oxholm et al. 1991;Boyce 1994). C57Bl/6j epidermis was more compro-mised than that of BALB/c, in spite of the fact that wecould not see parasitism of keratinocytes or Langerhanscells in either animal.
Thus, BALB/c mice behaved as an experimentalmodel of diffuse cutaneous leishmaniasis, that showeda new feature: parasitism of perineurial cells. C57Bl/6janimals showed hypersensitivity signs together with afew preserved parasites late in the course of the infection(follow-up of 51 weeks).
From a morphological point of view, there were nodifferences in the macrophages, or in the interactionbetween the target cell and the parasite, between thetwo animal models studied. This suggests that differ-ences in the response of the hosts towards the parasitecould depend on the way in which they initiate the cellularlymphocyte response.
Acknowledgements
We are grateful to Ms Monica R. Kauffman andCleusa F. Hirata, who kindly carried out the techniqueused in this paper.
This study was supported by the Fundacao de Amparoa Pesquisa do Estado de Sao Paulo (92/2017-0) andConselho Nacional de Desenvolvimento Cientıfico eTecnologico, Brazil.
References
BHATNAGAR S., MAITRA S.C., GURU P.Y. & KATIYAR J.C.(1991) Ultrastructural study on the antileishmanial effect of1,2-dimethyl-3-methoxycarbonil-4- (2-nitro-4,5-dimetoxy-phenyl) pyrrole in hamsters. Indian J. Med. Res. 93, 147–151.
BITTENCOURT A.L. & ANDRADE Z.A. (1967) Aspectos imunologicosna leishmaniose cutaneo-mucosa. Hospital 71, 89–98.
BITTENCOURT A.L., FREITAS L.A.R., POMPEU M.L., VIEIRA M.L. &BARRAL A. (1990) Distinct ultrastructural aspects in different
biopsies of a single patient with diffuse cutaneous leish-maniasis. Mem. Inst. Oswaldo Cruz 85, 53–59.
BOYCE S.T. (1994) Epidermis as a secretory tissue. J. Invest.Dermatol. 102, 8–10.
CONVIT J. (1974) The Kellersberger Memorial Lecture: Leprosyand leishmaniasis: similar clinical-immunological-pathologicalmodels. Ethiop. Med. J. 12, 187–195.
DUARTE M.I.S., MARIANO O.N., TAKAKURA C.F.H., UIP D.E. &CORBETT C.E.P. (1992) A fast method for processing biologicmaterial for electron microscopic diagnosis in infectiousdisease. Ultrastruct. Pathol. 16, 475–482.
EL-HASSAN A.M., ALI M.S., ZIJLSTRA E., EL-TOUM I.A., GHALIB H.W.& AHMED H.M. (1992) Post-kala-azar dermal leishmaniasisin the Sudan: peripheral neural involvement. Int. J. Dermatol.31, 400–403.
FINE R.M. (1990) Keratinocyte intercellular adhesion molecule-1(ICAM-1) in the pathogenesis of Sezary syndrome and cuta-neous T cell lymphomas. Int. J. Dermatol. 29, 623–624.
GIORGIO S. (1989) Estudo da imunidade celular e humoral naimunopatalogia e resistencia a infeccao por L. amazonensis.Sao Paulo. 103 p. Dissertacao (Mestrado) Instituto de Cien-cias Biomedicas, Universidade de Sao Paulo.
KUPPER T.S., BALLARD D.W., CHUA A.O., MCGUIRE J.S., FLOOD
P.M., HOROWITZ M.C., LANGDON R., LIGHTFOOT L. & GUBLER U.(1986) Human-keratinocytes contain mRNA indistinguishablefrom monocyte interleukin1a and bmRNA. J. Exp. Med. 164,2095–2100.
LAINSON R. & SHAW J.J. (1987) Evolution, classification andgeographical distribution. In The Leishmaniasis in Biologyand Medicine. Volume 1. Eds W. Peters and Killick-Kendrick,London: Academic Press. pp. 1–120.
LUGER T.A., STADLER B.M., KATZ S.I. & OPPENHEIM J.J. (1981)Epidermal cell (keratinocyte) derived thymocyte-activatingfactor (ETAF). J. Immunol. 127, 1493–1498.
MANN R.J. & HARMAN R.R. (1984) Cutaneous anaesthesia innecrobiosis lipoidica. Br. J. Dermatol. 110, 323–325.
MOLINA S.S., CHINCHILLA M.C. & TUK B.D. (1995) Estudio histo-patologico de la leishmaniasis cutanea experimental enanimales sensibles y resistentes. Patalogıa 33, 143–148.
OXHOLM A., DIAMANT M., OXHOLM P. & BENDTZEN K. (1991) Inter-leukin-6 and tumour necrosis factor alpha are expressedby keratinocytes but not by Langerhans cells. APMIS 99,58–64.
PEARSON R.D. & STEIGBIGEL R.T. (1981) Phagocytosis and kill-ing of the protozoan Leishmania donovani by human poly-morphonuclear leukocytes. J. Immunol. 127, 1438–1443.
PEREIRA J.H., PALANDE D.D. & GSCHMEISSNER S.E. (1991) Myco-bacteria in nerve trunks of long-term treated leprosy patients.Lepr. Rev. 62, 134–142.
PIMENTA P.F., SANTOS M.A. & SOUZA W. (1987) Fine structureand cytochemistry of the interaction between Leishmaniamexicana amazonensis and rat neutrophils and eosinophils.J. Submicrosc. Cytol. 19, 387–395.
RIDLEY D.S. & JOB C.K. (1985) The pathology of leprosy. InLeprosy. Ed. R.C. Hastings. Edinburgh; Churchill Livingstone.pp. 100–133.
RIDLEY M.J. & WELLS C.W. (1986) Macrophage-parasiteinteractions in the lesions of cutaneous leishmaniasis: anultrastructural study. Am. J. Pathol. 123, 79–85.
SATTI M.B., EL-HASSAN A.M., AL-GINDAU Y., OSMAN M.A. &AL-SOHAIBANI M.O. (1989) Peripheral neural involvementin cutaneous leishmaniasis: a pathologic study of human
88 C. Vasconcellos and M.N. Sotto
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89

and experimental animal lesions. Int. J. Dermatol. 28,243–247.
SCHMITT D. (1990) La presentation antigenique au niveau dela peau. Ann. Dermatol. Venereol. 117, 405–413.
SCHMITT D., SOUTEYRAND P., BROCHIER J., CZERNIELEWSKI J. &THIVOLET J. (1982) Phenotype of cells involved in mycosisfungoides and Sezary syndrome (blood and skin lesions):immunomorphological study with monoclonal antibodies.Acta Derm. Venereol. (Stockholm) 62, 193–199.
SHETTY V.P., MUKHERJEE R. & ANTIA N.H. (1992) Ultrastructuralstudy of mouse dorsal root ganglion cultures infected longterm with M. Lepræ. Indian J. Lepr. 64, 293–301.
TAPIA F.J., CACERES-DITTMAR G., ACUNA L. & MOSCA W. (1989)Epidermal Langerhans cells in infectious diseases. Histol.Histopathol. 4, 499–508.
TZE-CHUN L., ZHONG-MIN J. & SKINSNES O.K. (1989) Light- andelectron-microscopic study of M. lepræ-infected armadillonerves. Int. J. Leprosy 57, 65–72.
Experimental cutaneous leishmaniasis: TEM of inoculation site 89
q 1997 Blackwell Science Ltd, International Journal of Experimental Pathology, 78, 81–89