exoskeletal anatomy of the madagascar hissing cockroach
TRANSCRIPT

Exoskeletal Anatomy of the Madagascar Hissing CockroachGromphadorhina portentosa1'2
PATRICK J. DAILEY AND ROBERT C. GRAVESDepartment of Biological Sciences, Bowling Green State University, Bowling Green, OH 43403
ABSTRACTThe exoskeletal anatomy of the head and cervical tentosa, are discussed and illustrated. Sexual differences
region, thoracic region, abdominal region, and tracheaeof $ and 9 hissing cockroaches, Gromphadorhina por-
between males and females are also described, but notthe genitalia.
Gromphadorhina portentosa (Schaum) is a speciesof large, wingless, hissing cockroach native to Mada-gascar. Although this species has been widely used asa laboratory animal for research and teaching, littleis known about its general anatomy.
Van Herrewege (1973) has reviewed the genus.However, he has indicated in personal communica-tions that a thorough and complete revision of Grom-phadorhina would require investigations in Mada-gascar because museum specimens are often insuffi-cient and laboratory populations are subject to geneticdrift. Van Herrewege (1973) stated that some labo-ratory cultures referred to as G. portentosa by variousresearchers in the United States may actually be G.picea Van Herrewege.
The large size of this species (5-6 cm as adults)makes it extremely useful for classroom dissection orphysiological research. The external anatomy is illus-trated in this paper, and the gross internal anatomy,including the digestive, circulatory, nervous, and re-productive systems, is considered in Dailey andGraves (1976). We hope these studies will be usefulin future work on the comparative morphology andevolution of cockroaches.
RESULTS AND DISCUSSION
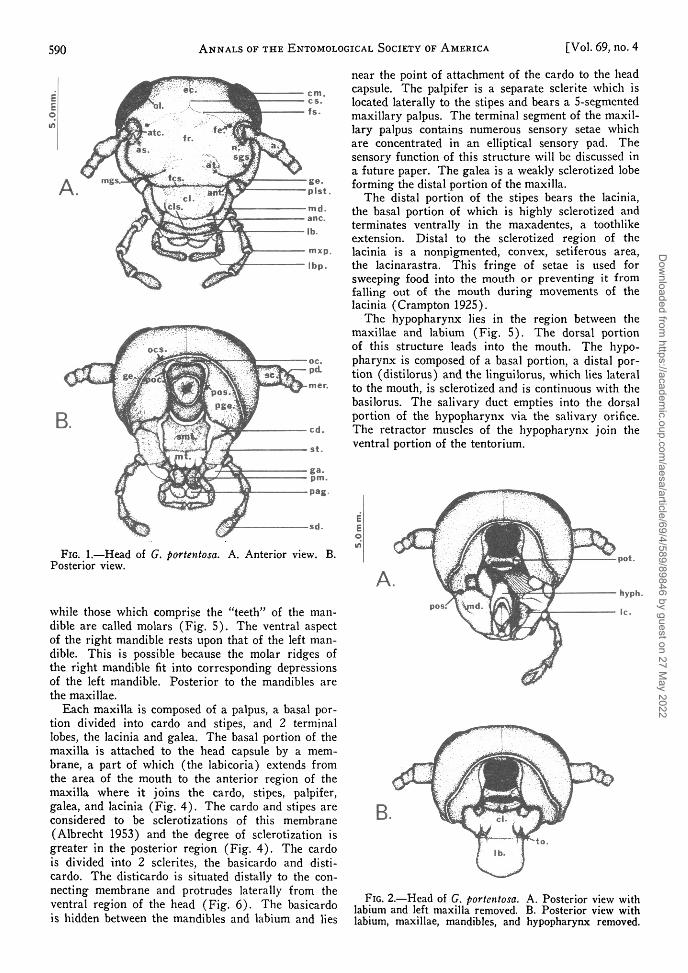
/. The Head and Cervical Region.—Cockroachheads are relatively unspecialized and most scleritesare readily visible. As in other cockroaches, the headof G. portentosa is hypognathous and located ventralto the large shieldlike pronotum. The head capsuleand mouthparts are hard and heavily sclerotized. Thehead is attached to a very flexible neck or cervicumand can be moved extensively.
The frons, clypeus, vertex, genae, postgenae, occi-put, and postocciput form the head capsule (Fig. 1and 14). The vertex occupies the dorsal roof of thehead between the compound eyes and is bordered bythe frons and antennal sutures anteriorly and theoccipital suture and genae posteriorly. The lateralregion of the vertex is limited by the ocular sutures.Immediately posterior to the vertex are the genaewhich are bordered by the subgenal, ocular, mando-genal, and occipital sutures. The oculata or ocularsuture separates the compound eye from the headsclerites. Between the cuticular covering of the eye
1 Blattaria: Blaberidae: Oxyhaloinae.2 Received for publication Feb. 5, 1976.
or cornea and the ocular suture there is a small ocu-lar sclerite (Fig. 14).
The occiput is a small sclerite lying between thevertex and the occipital foramen (Fig. IB). Its ta-pering edges are terminated by the postocciput whichis confined to a small area rimmed by the occipitalforamen, the postgenae, and dorsally by the occiput.The neck membrane or cervicum is attached to thehead at the postocciput.
The epicranium lies anterior to the vertex. In thisdescription it includes the anterior aspect of the ver-tex before it reaches the frontal suture (Fig. 1)which branches ventrolaterally and terminates at thefenestrae. The frontal suture also marks the dorsallimit of the frons which is bordered by the fronto-clypeal suture ventrally, the subgenal suture laterally,and by the frontal, antennal, and ocular suturesdorsally.
The anterior tentorial pits, which indicate the in-ternal attachments of the anterior tentorial arms, arebarely visible exteriorly.
The antennal sutures mark the outer boundary ofthe antennal sockets. The antennal scape is articu-lated to the rim of the antennal socket by a ventralprocess termed the antennifer (Fig. 1), which acts asa pivot for the antenna. Between the scape and theantennal suture is the antacoria, a membrane whichpermits the antenna to move in a semicircular manner.
The clypeus lies ventral to the frontoclypeal suture.The dorsal portion is partially sclerotized but lessthan the frons. Ventral to the clypeus, and confluentwith it, is a non-pigmented area termed the ante-clypeus (Fig. 1). The clypeus is one of the movablesclerites of the head capsule to which is attached amovable preoral lobe, the labrum. The labrum is im-mediately ventral to the anteclypeus and separatedfrom the clypeus by the clypeolabral suture; it isalmost entirely sclerotized except for a small non-pigmented ventral portion. Small sclerites, the tor-mae, occur posteriorly between the clypeus and lab-rum (Fig. 2).
The mandibles are highly sclerotized structuresused for masticating food (Fig. 2 and 5). They ar-ticulate anteriorly at the point where genae and fronsmeet and the clypeus begins, and posteriorly at themost ventral portion of the postgenae (Fig. 2). Eachmandible possesses large teethlike ridges on the rimapproaching the vertical midline of the head capsule.The less prominent ridge is known as the incisor
589
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
590 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA [Vol. 69, no. 4
pag.
sd.
FIG. 1.—Head of G. portentosa. A. Anterior view.Posterior view.
B.
while those which comprise the "teeth" of the man-dible are called molars (Fig. 5). The ventral aspectof the right mandible rests upon that of the left man-dible. This is possible because the molar ridges ofthe right mandible fit into corresponding depressionsof the left mandible. Posterior to the mandibles arethe maxillae.
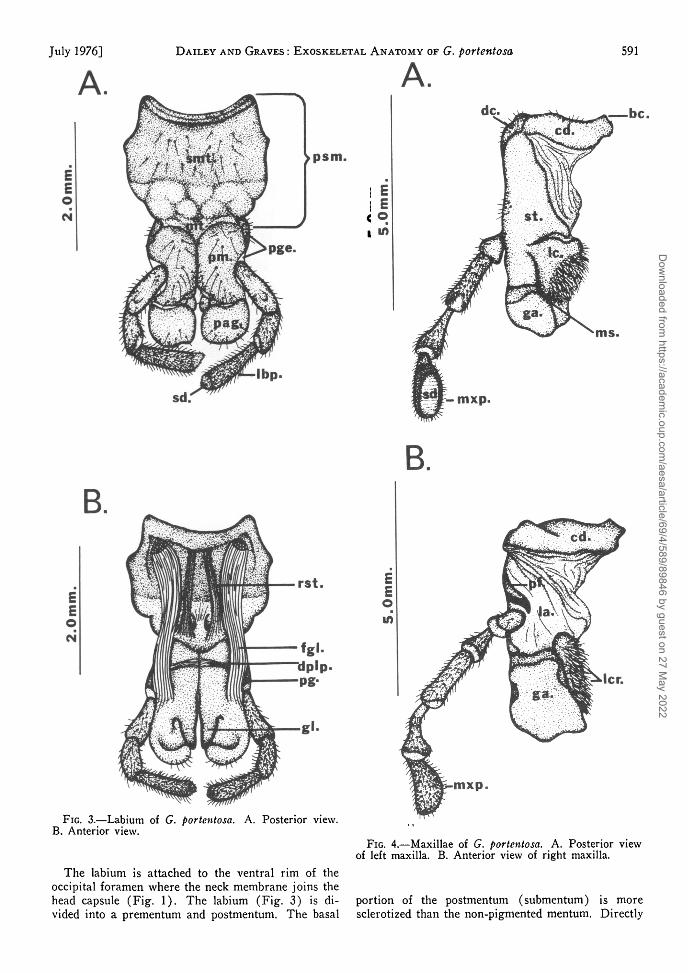
Each maxilla is composed of a palpus, a basal por-tion divided into cardo and stipes, and 2 terminallobes, the lacinia and galea. The basal portion of themaxilla is attached to the head capsule by a mem-brane, a part of which (the labicoria) extends fromthe area of the mouth to the anterior region of themaxilla where it joins the cardo, stipes, palpifer,galea, and lacinia (Fig. 4). The cardo and stipes areconsidered to be sclerotizations of this membrane(Albrecht 1953) and the degree of sclerotization isgreater in the posterior region (Fig. 4). The cardois divided into 2 sclerites, the basicardo and disti-cardo. The disticardo is situated distally to the con-necting membrane and protrudes laterally from theventral region of the head (Fig. 6). The basicardois hidden between the mandibles and labium and lies
near the point of attachment of the cardo to the headcapsule. The palpifer is a separate sclerite which islocated laterally to the stipes and bears a 5-segmentedmaxillary palpus. The terminal segment of the maxil-lary palpus contains numerous sensory setae whichare concentrated in an elliptical sensory pad. Thesensory function of this structure will be discussed ina future paper. The galea is a weakly sclerotized lobeforming the distal portion of the maxilla.
The distal portion of the stipes bears the lacinia,the basal portion of which is highly sclerotized andterminates ventrally in the maxadentes, a toothlikeextension. Distal to the sclerotized region of thelacinia is a nonpigmented, convex, setiferous area,the lacinarastra. This fringe of setae is used forsweeping food into the mouth or preventing it fromfalling- out of the mouth during movements of thelacinia (Crampton 1925).
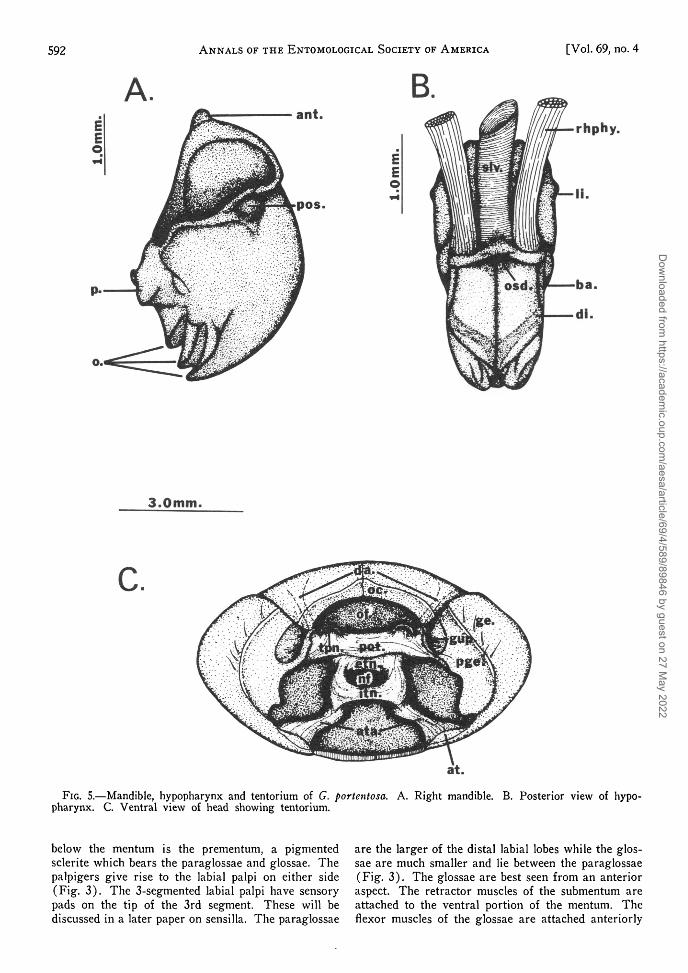
The hypopharynx lies in the region between themaxillae and labium (Fig. 5). The dorsal portionof this structure leads into the mouth. The hypo-pharynx is composed of a basal portion, a distal por-tion (distilorus) and the linguilorus, which lies lateralto the mouth, is sclerotized and is continuous with thebasilorus. The salivary duct empties into the dorsalportion of the hypopharynx via the salivary orifice.The retractor muscles of the hypopharynx join theventral portion of the tentorium.
pot.
hyph.
FIG. 2.—Head of G. portentosa. A. Posterior view withlabium and left maxilla removed. B. Posterior view withlabium, maxillae, mandibles, and hypopharynx removed.
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
July 1976]
A.
B.
DAILEY AND GRAVES : EXOSKELETAL ANATOMY OF G. portentosa
A.
>psm.
Ibp.
rst.
FIG. 3.—Labium of G. portentosa. A. Posterior view.B. Anterior view.
EE©in
591
k —be.
ms.
B.
FIG. 4.—Maxillae of G. portentosa. A. Posterior viewof left maxilla. B. Anterior view of right maxilla.
The labium is attached to the ventral rim of theoccipital foramen where the neck membrane joins thehead capsule (Fig. 1). The labium (Fig. 3) is di- portion of the postmentum (submentum) is morevided into a prementum and postmentum. The basal sclerotized than the non-pigmented mentum. Directly
more
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
592 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA
B.ant.
pos.
[Vol. 69, no. 4
rhphy.
3.0mm.
c.
FIG. 5.—Mandible, hypopharynx and tentorium of G. portentosa. A. Right mandible. B. Posterior view of hypo-pharynx. C. Ventral view of head showing tentorium.
below the mentum is the prementum, a pigmentedsclerite which bears the paraglossae and glossae. Thepalpigers give rise to the labial palpi on either side(Fig. 3). The 3-segmented labial palpi have sensorypads on the tip of the 3rd segment. These will bediscussed in a later paper on sensilla. The paraglossae
are the larger of the distal labial lobes while the glos-sae are much smaller and lie between the paraglossae(Fig. 3). The glossae are best seen from an anterioraspect. The retractor muscles of the submentum areattached to the ventral portion of the mentum. Theflexor muscles of the glossae are attached anteriorly
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
July 1976] DAILEY AND GRAVES : EXOSKELETAL ANATOMY OF G. portentosa 593
at a point just below the palpiger. The depressormuscles of the labial palpi are located between thepalpigers.
If the mouthparts are removed and the head cap-sule is placed in 50% KOH for 3-4 days to dissolvethe muscles and other soft parts, it is possible toview the tentorium (Fig. 5). According to Crampton(1925) the tentorium is an internal framework whichserves as a place for muscle attachment and as abrace to support the head and protect fragile internalstructures such as the brain. The posterior arms ofthe tentorium arise as invaginations of the gular pitswhich lie beneath the tentorial prominences. Theposterior arms unite to form the body of the ten-torium (eutentorium). The anterior tentorial armsattached at the anterior tentorial pits (previously dis-cussed) unite in the mid-region of the head to formthe intertentorium. The opening formed from thefusion of the intertentorium with the eutentorium iscalled the neuroforamen through which pass the cruracerebri of the ventral nerve cord. Anterior prolonga-tions of the eutentorium (dorsal arms) extend dor-sally to reach the region of the antennal sclerites.
The head is joined to the thorax by the cervicumor cervical membrane (Fig. 14). The cervical scle-rites of the neck region are located in this membrane.The dorsal cervical sclerites connect anteriorly withthe occiput and posteriorly to the dorsal tergite of theprothorax. The posterior portion of the dorsal cervi-cal sclerite is characterized by a very thin cuticularplate which attaches to the prothorax. At the junc-tion of the dorsal cervical sclerite with this cuticularplate a pair of thin cuticular arms protrude. Thesearms probably make contact with the prothoracic cu-ticle and secure the dorsal cervical sclerite to thatstructure.
A lightly sclerotized area called the lateral cervicalsensory patch can be seen on either side of the dorsalcervical sclerite near its attachment to the posteriorregion of the head.
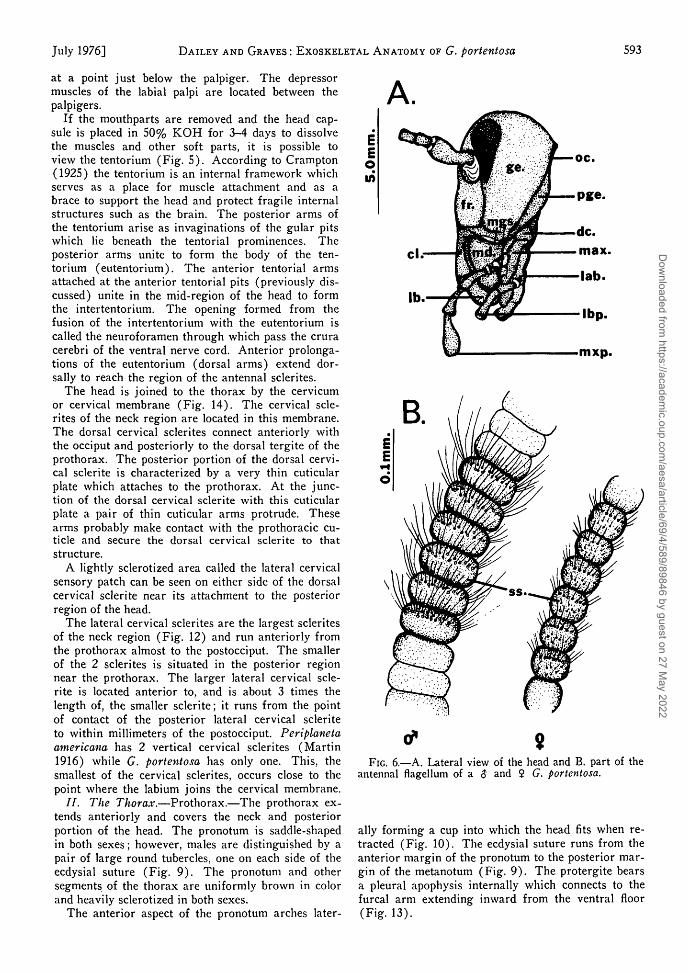
The lateral cervical sclerites are the largest scleritesof the neck region (Fig. 12) and run anteriorly fromthe prothorax almost to the postocciput. The smallerof the 2 sclerites is situated in the posterior regionnear the prothorax. The larger lateral cervical scle-rite is located anterior to, and is about 3 times thelength of, the smaller sclerite; it runs from the pointof contact of the posterior lateral cervical scleriteto within millimeters of the postocciput. Periplanetaamericana has 2 vertical cervical sclerites (Martin1916) while G. portentosa has only one. This, thesmallest of the cervical sclerites, occurs close to thepoint where the labium joins the cervical membrane.
//. The Thorax.—Prothorax.—The prothorax ex-tends anteriorly and covers the neck and posteriorportion of the head. The pronotum is saddle-shapedin both sexes; however, males are distinguished by apair of large round tubercles, one on each side of theecdysial suture (Fig. 9). The pronotum and othersegments of the thorax are uniformly brown in colorand heavily sclerotized in both sexes.
The anterior aspect of the pronotum arches later-
oc.
Pge.
mxp.
B
FIG. 6.—A. Lateral view of the head and B. part of theantennal flagellum of a $ and $ G. portentosa.
ally forming a cup into which the head fits when re-tracted (Fig. 10). The ecdysial suture runs from theanterior margin of the pronotum to the posterior mar-gin of the metanotum (Fig. 9). The protergite bearsa pleural apophysis internally which connects to thefurcal arm extending inward from the ventral floor(Fig. 13).
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
594 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA [Vol. 69, no. 4
10.0mm.
me.
Mesl.
Metl. i
Prl- 4
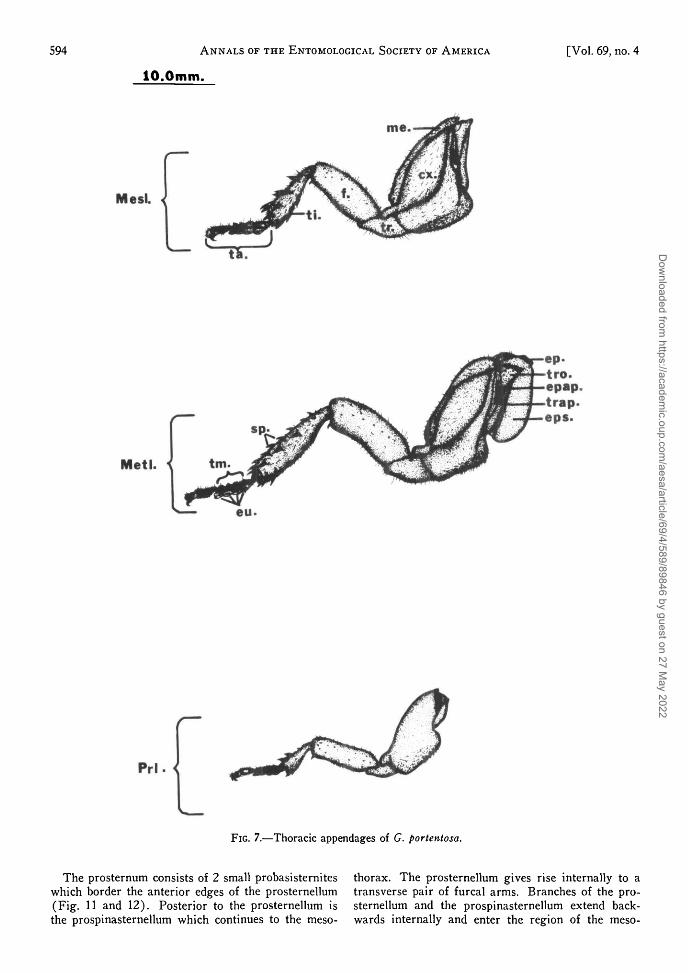
FIG. 7.—Thoracic appendages of G. portentosa.
The prosternum consists of 2 small probasisterniteswhich border the anterior edges of the prosternellum(Fig. 11 and 12). Posterior to the prosternellum isthe prospinasternellum which continues to the meso-
thorax. The prosternellum gives rise internally to atransverse pair of furcal arms. Branches of the pro-sternellum and the prospinasternellum extend back-wards internally and enter the region of the meso-
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
July 1976] DAILEY AND GRAVES : EXOSKELETAL ANATOMY OF G. portentosa 595
ts.
>pr.
td.
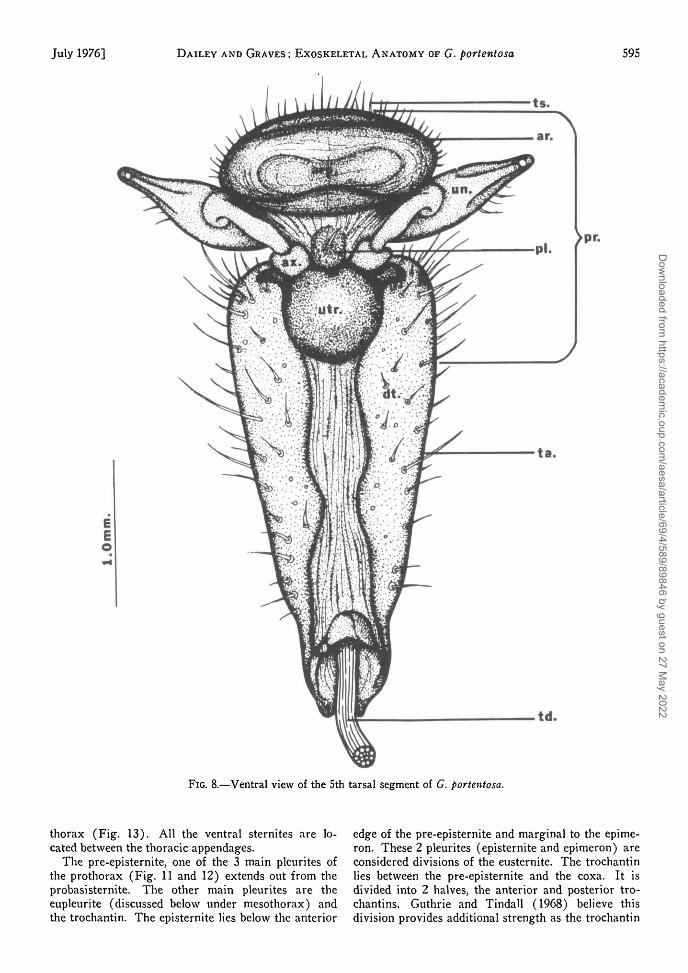
FIG. 8.—Ventral view of the 5th tarsal segment of G. portentosa.
thorax (Fig. 13). All the ventral sternites are lo-cated between the thoracic appendages.
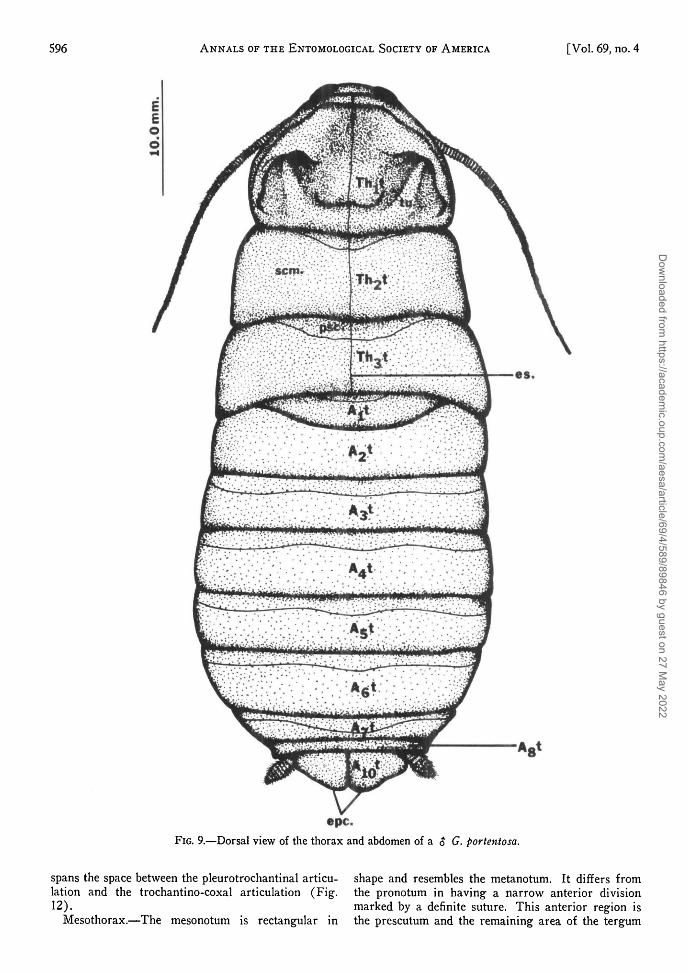
The pre-episternite, one of the 3 main pleurites ofthe prothorax (Fig. 11 and 12) extends out from theprobasisternite. The other main pleurites are theeupleurite (discussed below under mesothorax) andthe trochantin. The episternite lies below the anterior
edge of the pre-episternite and marginal to the epime-ron. These 2 pleurites (episternite and epimeron) areconsidered divisions of the eusternite. The trochantinlies between the pre-episternite and the coxa. It isdivided into 2 halves, the anterior and posterior tro-chantins. Guthrie and Tindall (1968) believe thisdivision provides additional strength as the trochantin
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
596 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA [Vol. 69, no. 4
epc.
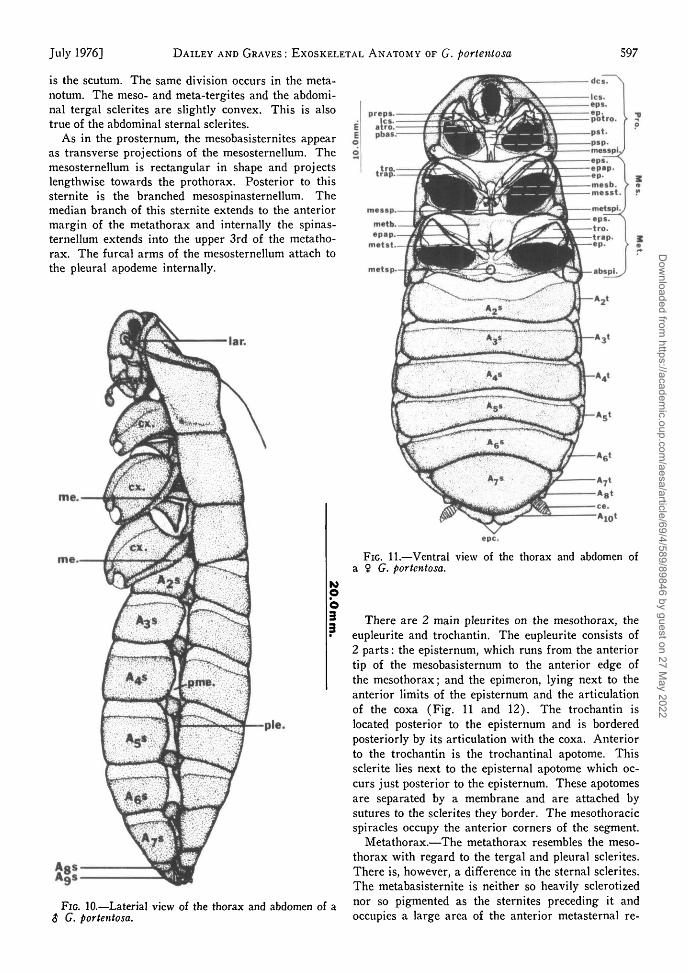
FIG. 9.—Dorsal view of the thorax and abdomen of a $ G. portentosa.
spans the space between the pleurotrochantinal articu- shape and resembles the metanotum. It differs fromlation and the trochantino-coxal articulation (Fig. the pronotum in having a narrow anterior division12). marked by a definite suture. This anterior region is
Mesothorax.—The mesonotum is rectangular in the prescutum and the remaining area of the tergum
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
July 1976] DAILEY AND GRAVES : EXOSKELETAL ANATOMY OF G. portentosa 597
is the scutum. The same division occurs in the meta-notum. The meso- and meta-tergites and the abdomi-nal tergal sclerites are slightly convex. This is alsotrue of the abdominal sternal sclerites.
As in the prosternum, the mesobasisternites appearas transverse projections of the mesosternellum. Themesosternellum is rectangular in shape and projectslengthwise towards the prothorax. Posterior to thissternite is the branched mesospinasternellum. Themedian branch of this sternite extends to the anteriormargin of the metathorax and internally the spinas-ternellum extends into the upper 3rd of the metatho-rax. The furcal arms of the mesosternellum attach tothe pleural apodeme internally.
lar.
me.
me.
FIG. 10.—Laterial view of the thorax and abdomen of a$ G. portentosa.
epc.
FIG. 11.—Ventral view of the thorax and abdomen ofa ? G. portentosa.
There are 2 main pleurites on the mesothorax, theeupleurite and trochantin. The eupleurite consists of2 parts: the episternum, which runs from the anteriortip of the mesobasisternum to the anterior edge ofthe mesothorax; and the epimeron, lying next to theanterior limits of the episternum and the articulationof the coxa (Fig. 11 and 12). The trochantin islocated posterior to the episternum and is borderedposteriorly by its articulation with the coxa. Anteriorto the trochantin is the trochantinal apotome. Thissclerite lies next to the episternal apotome which oc-curs just posterior to the episternum. These apotomesare separated by a membrane and are attached bysutures to the sclerites they border. The mesothoracicspiracles occupy the anterior corners of the segment.
Metathorax.—The metathorax resembles the meso-thorax with regard to the tergal and pleural sclerites.There is, however, a difference in the sternal sclerites.The metabasisternite is neither so heavily sclerotizednor so pigmented as the sternites preceding it andoccupies a large area of the anterior metasternal re-
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
598 ANNALS OF THE ENTOMOLOGICAL SOCIETY OF AMERICA
Ics.
[Vol. 69, no. 4
Mes.<
Met/
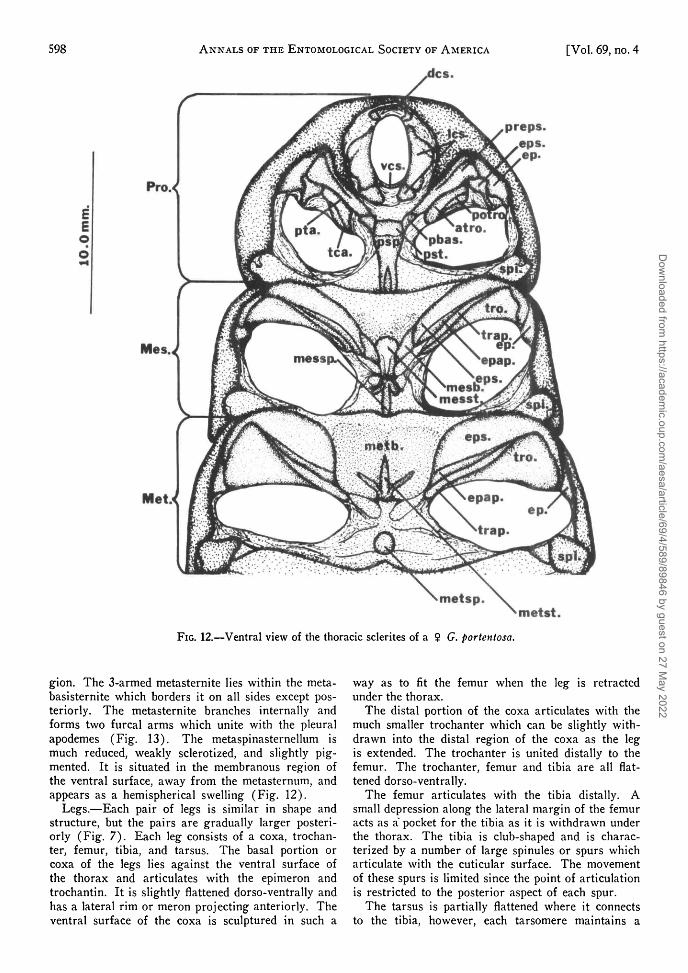
metst.FIG. 12.—Ventral view of the thoracic sclerites of a $ G. portentosa.
gion. The 3-armed metasternite lies within the meta-basisternite which borders it on all sides except pos-teriorly. The metasternite branches internally andforms two furcal arms which unite with the pleuralapodemes (Fig. 13). The metaspinasternellum ismuch reduced, weakly sclerotized, and slightly pig-mented. It is situated in the membranous region ofthe ventral surface, away from the metasternum, andappears as a hemispherical swelling (Fig. 12).
Legs.—Each pair of legs is similar in shape andstructure, but the pairs are gradually larger posteri-orly (Fig. 7). Each leg consists of a coxa, trochan-ter, femur, tibia, and tarsus. The basal portion orcoxa of the legs lies against the ventral surface ofthe thorax and articulates with the epimeron andtrochantin. It is slightly flattened dorso-ventrally andhas a lateral rim or meron projecting anteriorly. Theventral surface of the coxa is sculptured in such a
way as to fit the femur when the leg is retractedunder the thorax.
The distal portion of the coxa articulates with themuch smaller trochanter which can be slightly with-drawn into the distal region of the coxa as the legis extended. The trochanter is united distally to thefemur. The trochanter, femur and tibia are all flat-tened dorso-ventrally.
The femur articulates with the tibia distally. Asmall depression along the lateral margin of the femuracts as a pocket for the tibia as it is withdrawn underthe thorax. The tibia is club-shaped and is charac-terized by a number of large spinules or spurs whicharticulate with the cuticular surface. The movementof these spurs is limited since the point of articulationis restricted to the posterior aspect of each spur.
The tarsus is partially flattened where it connectsto the tibia, however, each tarsomere maintains a
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
July 1976] DAILEY AND GRAVES : EXOSKELETAL ANATOMY OF G. portentosa 599
rounded appearance. Each tarsus is divided into 5segments or tarsomeres, the 1st 4 of which havewhite, cushion-like ventral surfaces known as tarsalpulvilli or euplantulae. These structures are utilizedby the cockroaches when climbing slippery surfacesor walking upside down on glass. The 5th tarsomereends in a clawed structure known as the pretarsus(Fig. 8). This appendage bears 2 lateral claws orungues and an arolium. The claws are articulateddorsally to the unguifer and ventrally to the ungui-tractor place. The arolium is the terminal structureon the pretarsus; its ventral surface is modified intoan aroliar pad similar to the cushionlike euplantulaeof the preceding tarsomeres.
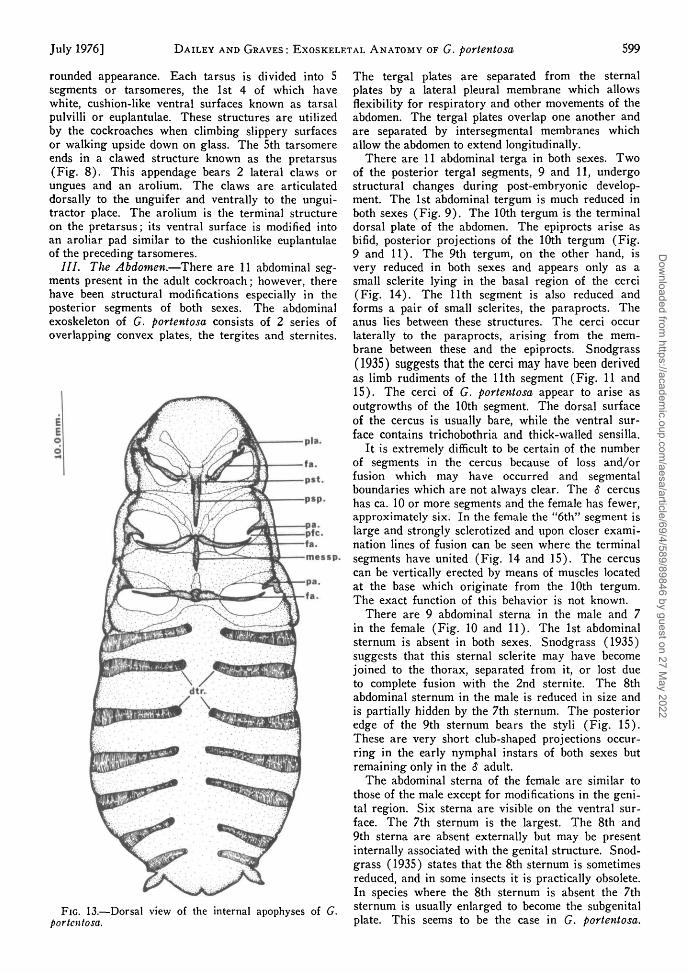
/ / / . The Abdomen.—There are 11 abdominal seg-ments present in the adult cockroach; however, therehave been structural modifications especially in theposterior segments of both sexes. The abdominalexoskeleton of G. portentosa consists of 2 series ofoverlapping convex plates, the tergites and sternites.
pla.
messp.
FIG. 13.—Dorsal view of the internal apophyses of G.portentosa.
The tergal plates are separated from the sternalplates by a lateral pleural membrane which allowsflexibility for respiratory and other movements of theabdomen. The tergal plates overlap one another andare separated by intersegmental membranes whichallow the abdomen to extend longitudinally.
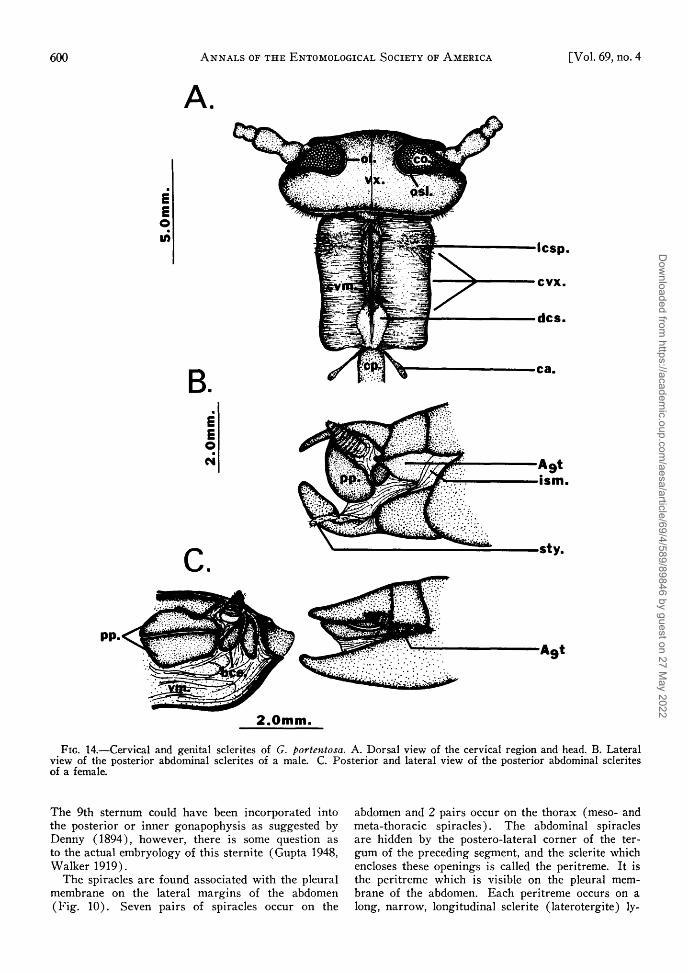
There are 11 abdominal terga in both sexes. Twoof the posterior tergal segments, 9 and 11, undergostructural changes during post-embryonic develop-ment. The 1st abdominal tergum is much reduced inboth sexes (Fig. 9). The 10th tergum is the terminaldorsal plate of the abdomen. The epiprocts arise asbifid, posterior projections of the 10th tergum (Fig.9 and 11). The 9th tergum, on the other hand, isvery reduced in both sexes and appears only as asmall sclerite lying in the basal region of the cerci(Fig. 14). The 11th segment is also reduced andforms a pair of small sclerites, the paraprocts. Theanus lies between these structures. The cerci occurlaterally to the paraprocts, arising from the mem-brane between these and the epiprocts. Snodgrass(1935) suggests that the cerci may have been derivedas limb rudiments of the 11th segment (Fig. 11 and15). The cerci of G. portentosa appear to arise asoutgrowths of the 10th segment. The dorsal surfaceof the cercus is usually bare, while the ventral sur-face contains trichobothria and thick-walled sensilla.
It is extremely difficult to be certain of the numberof segments in the cercus because of loss and/orfusion which may have occurred and segmentalboundaries which are not always clear. The $ cercushas ca. 10 or more segments and the female has fewer,approximately six. In the female the "6th" segment islarge and strongly sclerotized and upon closer exami-nation lines of fusion can be seen where the terminalsegments have united (Fig. 14 and 15). The cercuscan be vertically erected by means of muscles locatedat the base which originate from the 10th tergum.The exact function of this behavior is not known.
There are 9 abdominal sterna in the male and 7in the female (Fig. 10 and 11). The 1st abdominalsternum is absent in both sexes. Snodgrass (1935)suggests that this sternal sclerite may have becomejoined to the thorax, separated from it, or lost dueto complete fusion with the 2nd sternite. The 8thabdominal sternum in the male is reduced in size andis partially hidden by the 7th sternum. The posterioredge of the 9th sternum bears the styli (Fig. 15).These are very short club-shaped projections occur-ring in the early nymphal instars of both sexes butremaining only in the $ adult.
The abdominal sterna of the female are similar tothose of the male except for modifications in the geni-tal region. Six sterna are visible on the ventral sur-face. The 7th sternum is the largest. The 8th and9th sterna are absent externally but may be presentinternally associated with the genital structure. Snod-grass (1935) states that the 8th sternum is sometimesreduced, and in some insects it is practically obsolete.In species where the 8th sternum is absent the 7thsternum is usually enlarged to become the subgenitalplate. This seems to be the case in G. portentosa.
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
600 A N N A L S OF T H E E N T O M O L O G I C A L S O C I E T Y OF A M E R I C A [ V o l . 69, no. 4
The 9th sternum could have been incorporated into the posterior or inner gonapophysis as suggested by Denny ( 1 8 9 4 ) , however , there is some question as to the actual embryology of this sternite (Gupta 1948, Walker 1919) .
The spiracles are found associated with the pleural membrane on the lateral margins of the abdomen ( F i g . 1 0 ) . Seven pairs of spiracles occur on the
abdomen and 2 pairs occur on the thorax (meso- and meta-thoracic spi rac les) . T h e abdominal spiracles are hidden by the postero-lateral corner of the ter-gum of the preceding segment, and the sclerite which encloses these openings is called the peritreme. It is the peritreme which is visible on the pleural membrane of the abdomen. Each peritreme occurs on a long, narrow, longitudinal sclerite (laterotergite) ly-
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
July 1976] D A I L E Y A N D G R A V E S : E X O S K E L E T A L A N A T O M Y OF G. portentosa 601
ing between the tergum and sternum of each segment ( F i g . 1 0 ) .
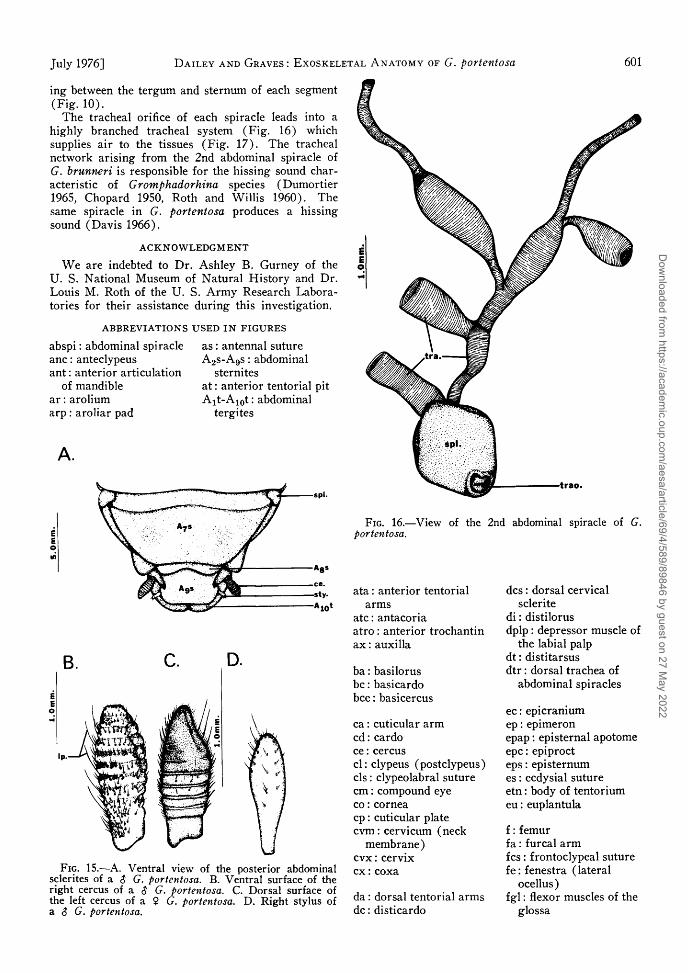
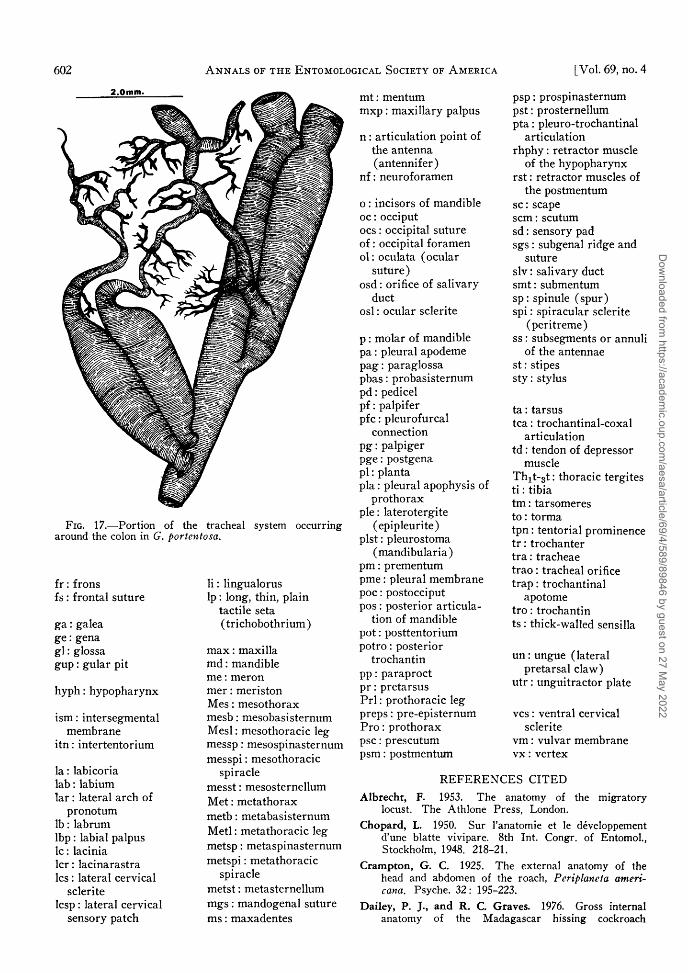
T h e tracheal orifice of each spiracle leads into a highly branched tracheal system ( F i g . 16) which supplies air to the tissues ( F i g . 1 7 ) . T h e tracheal network arising from the 2nd abdominal spiracle of G. brunneri is responsible for the hissing sound characteristic of Gromphadorhina species (Dumor t ie r 1965, Chopard 1950, Roth and Wi l l i s 1960) . The same spiracle in G. portentosa produces a hissing sound ( D a v i s 1966) .
ACKNOWLEDGMENT
W e are indebted to Dr . Ashley B. Gurney of the U . S. National Museum of Natural His tory and Dr . Louis M . Roth of the U . S. A r m y Research Laboratories for their assistance during this investigation.
ABBREVIATIONS USED IN FIGURES
abspi : abdominal spiracle a s : antennal suture a n c : anteclypeus ant: anterior articulation
of mandible a r : arolium arp : aroliar pad
A 2 s - A 9 s : abdominal sternites
at : anterior tentorial pit A i t - A 1 0 t : abdominal
tergites
FIG. 16.—View of the 2nd abdominal spiracle of G. portentosa.
FIG. 15.—A. Ventral view of the posterior abdominal sclerites of a $ G. portentosa. B. Ventral surface of the right cercus of a $ G. portentosa. C. Dorsal surface of the left cercus of a 9- G. portentosa. D . Right stylus of a S G. portentosa.
ata: anterior tentorial arms
ate: antacoria a t ro : anterior trochantin a x : auxilla
b a : basilorus b e : basicardo b e e : basicercus
c a : cuticular arm c d : cardo c e : cercus c l : c l y p e u s (pos tc lypeus) e ls : clypeolabral suture c m : compound eye c o : cornea c p : cuticular plate c v m : cervicum (neck
membrane) c v x : cervix c x : coxa
da : dorsal tentorial arms d c : disticardo
d e s : dorsal cervical sclerite
d i : distilorus dp lp : depressor muscle of
the labial palp d t : distitarsus dt r : dorsal trachea of
abdominal spiracles
e c : epicranium e p : epimeron epap : episternal apotome e p c : epiproct eps : episternum e s : ecdysial suture e tn: body o f tentorium e u : euplantula
f: femur f a : furcal arm f e s : frontoclypeal suture f e : fenestra (lateral
ocellus ) f g l : flexor muscles of the
glossa
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022
602 A N N A L S OF T H E E N T O M O L O G I C A L S O C I E T Y OF A M E R I C A [ V o l . 69, no. 4
f r : frons f s : frontal suture
g a : galea g e : gena g l : glossa g u p : gular pit
h y p h : hypopharynx
i sm: intersegmental membrane
i tn: intertentorium
la : labicoria l ab : labium lar : lateral arch of
pronotum l b : labrum l b p : labial palpus l c : lacinia l c r : lacinarastra l c s : lateral cervical
sclerite l c sp : lateral cervical
sensory patch
l i : lingualorus l p : long, thin, plain
tactile seta ( t r ichobothr ium)
m a x : maxilla m d : mandible m e : meron m e r : meriston M e s : mesothorax m e s b : mesobasi sternum M e s l : mesothoracic leg messp : mesospinasternum messpi : mesothoracic
spiracle messt : mesosternellum M e t : metathorax m e t b : metabasisternum M e t l : metathoracic leg metsp : metaspinasternum metspi : metathoracic
spiracle metst : metasternellum m g s : mandogenal suture m s : maxadentes
m t : mentum m x p : maxil lary palpus
n : articulation point of the antenna (antennifer)
nf : neuroforamen
o : incisors of mandible o c : occiput o c s : occipital suture o f : occipital foramen o l : oculata (ocular
suture) o s d : orifice of salivary
duct o s l : ocular sclerite
p : molar of mandible p a : pleural apodeme p a g : paraglossa pbas : probasisternum p d : pedicel p f : palpifer p f c : pleurofurcal
connect ion p g : palpiger p g e : postgena p i : planta p la : pleural apophysis of
prothorax p i e : laterotergite
(epipleuri te) plst : pleurostoma
(mandibular ia) p m : prementum p m e : pleural membrane p o c : postocciput p o s : posterior articula
tion of mandible p o t : posttentorium p o t r o : posterior
trochantin p p : paraproct p r : pretarsus P r l : prothoracic leg preps : pre-episternum P r o : prothorax p s c : prescutum p s m : postmentum
p s p : prospinasternum pst : prosternellum pta: pleuro-trochantinal
articulation rhphy: retractor muscle
of the hypopharynx rst: retractor muscles of
the postmentum s c : scape s c m : scutum sd : sensory pad sgs : subgenal ridge and
suture s lv : salivary duct smt : submentum s p : spinule ( spu r ) sp i : spiracular sclerite
(per i t reme) ss : subsegments or annuli
of the antennae s t : stipes s ty : stylus
ta: tarsus tea: trochantinal-coxal
articulation td : tendon o f depressor
muscle T h ^ ^ t : thoracic tergites t i : tibia t m : tarsomeres t o : tor ma tpn: tentorial prominence t r : trochanter t ra : t racheae t r ao : tracheal orifice t rap: trochantinal
apotome t ro : trochantin t s : thick-walled sensilla
un : ungue (lateral pretarsal c l a w )
utr : unguitractor plate
v e s : ventral cervical sclerite
v m : vulvar membrane v x : vertex
R E F E R E N C E S C I T E D
Albrecht, F. 1953. The anatomy of the migratory locust. The Athlone Press, London.
Chopard, L. 1950. Sur l'anatomie et le developpement d'une blatte vivipare. 8th Int. Congr. of Entomol., Stockholm, 1948. 218-21.
Crampton, G. C. 1925. The external anatomy of the head and abdomen of the roach, Pcriplaneta ameri-cana. Psyche. 32 : 195-223.
Dailey, P. J., and R. C. Graves. 1976. Gross internal anatomy of the Madagascar hissing cockroach
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022

July 1976] D A I L E Y A N D G R A V E S : E X O S K E L E T A L A N A T O M Y OF G. portentosa 603
Gromphadorhina portentosa. Ann. Entomol. Soc. Am. 69: 609-16.
Davis, M. C. 1966. Production of sound in the whistling roach, Gromphadorhina portentosa (Schaum) (Blattidae). Turtox News 44: 271.
Denny, A. 1894. On the development of the ovipositor in the cockroach, Periplaneta americana. Rep. British Assoc. Adv. Sci. 63: 818.
Dumortier, B. 1965. L'emission sonore dans le genre Gromphadorhina brunneri (Blattodea, Perisphaeri-idae), etude morphologique et biologique. Bull. Soc. Zool. France. 90: 89-101.
Gupta, P. D. 1948. On the structure, development and homology of the female reproductive organs in orthopteroid insects. Indian J. Entomol. 10: 75-123.
Guthrie, D. M., and A. R. Tindall. 1968. The biology of the cockroach. St. Martin's Press, New York.
Martin, J. F. 1916. The thoracic and cervical sclerites of insects. Ann. Entomol. Soc. Am. 9: 35-88.
Roth, L. M., and E. R. Willis. 1960. Biotic associations of cockroaches. Smithonsian Misc. Coll. 141: 1-470.
Snodgrass, R. E. 1935. Principles of insect morphology. McGraw-Hill Book Company, New York.
Van Herrewege, C. 1973. Contribution a l'etude des Blattaria de la faune Malgache. II. Description de huit especes nouvelles appartenant aux genres Gromphadorhina Brunner v. W . et Elliptorhina gen. nov. Bull. Mens. Soc. Linneenne Lyon. 42 : 75-103.
Walker, E. M. 1919. The terminal abdominal structures of orthopteroid insects. I. Ann. Entomol. Soc. Am. 12: 267-326.
Dow
nloaded from https://academ
ic.oup.com/aesa/article/69/4/589/89846 by guest on 27 M
ay 2022