evaluation of a tissue culture-based approach for differentiating between virulent and avirulent...
TRANSCRIPT

348
Journal of Food Protection VoL 70 No2 2007 Pages 348-354Copyrlghl C InlarnalionalAssociation tor Food Protection
Evaluation of a Tissue Culture-Based Approach forDifferentiating between Virulent and Avirulent Vibrioparahaemolyticus Strains Based on Cytotoxicity
P S MARIE YE Gt MARTIN WIEDMAN 2 AND KA HRY J BOOR2
Biological Sciences Department California Polytechnic State University San Luis Obispo California 93407 and 20epartment of Food ScienceCornell University Ithaca New York 14853 USA
MS 06-333 Received 15 June 2oo6Accepted 30 September 2006
ABSTRACT
The ability of only a subset of Vibrio parahaemolyticus strains to cause human infection under cores the need for ananalytical method that can effectively differentiate between pathogenic strains and tho e that do not cau e disease We testedthe fea ibility of a ti sue culture-based as ay to determine whether clinical isolates could be differentiated from nonclinicali elates based on relative isolate cytopathogenicity To screen for cytotoxic capability we mea ured relative extracellular lactatedehydrogenase as an indicator of host cell damage in five different mammalian cell lines in the presence of V parahaemolyticusIsolate originating from clinical sources exhibited 155 to 593 relative cytotoxicity whereas those originating from foodsource exhibited 44 to 549reJative cytotoxicity In the presence of 12 X 106 cells cytotoxicity was 16- to 35-foldhigher (P lt 005) for clinical isolates than for nonclinical isolates in L2 Henle 407 and Caco-2 cell lines Vparahaemolyticusserotype 03K6 clinical i elates had 16- to 2l-fold higher cytotoxicity than did the non-03K6 clinical i olate with signif-icantly higher cytotoxicity in Hela J774A1 and Henle 407 cells than in L2 and Caco-2 ceUs Because V parahaemolyticusoften i found in oyster the effect of the presence of an oyster matrix on assay efficacy was al 0 tested with L2 cells Thecytotoxicity elicited by a highly cytotoxic V parahaemolyticus isolate was not affected by the presence of oyster ti suesuggesting that an oyster matrix wUJ not interfere with assay sensitivity In the present format this as ay can detect thepre ence of gt lOS cells of a virulent V parahaemolyticus strain in an oyster matrix
As a natural inhabitant of estuarine marine water Vib-rio parahaemoiyticus i widely di tributed in in hare ma-rine waters throughout the world Some V parahaemoiy-ticus strain are capable of cau ing human illness and aub et of the e trains has been implicated in outbreak of
foodborne disease No ingle genetic determinant has beenidentified as both necessary and sufficient for V parahae-molyticus virulence although many factor have been pro-posed to be involved (45) Thermostable direct hemolysin(TDH) i one factor that ha long been con idered a majorV parahaemolyticus virulence marker (29 32) A V para-haemolyticus train bearing a 10 -of-function mutation inthe Idh gene ha attenuated virulence in the rabbit ileal loopmodeJ (32) However loss ofTDH did not impair V para-haemolyticus virulence in all model system (9 11) indi-cating that TDH is not the ole contributor to V parahae-molyticus pathogenicity A TDH-related hemolysin (TRH)also ha been Jinked to V parahaemolyticus ga troenteritis(16 37 38) However V parahaemoiyticus strains lackingTDH have been associated with foodborne disease illnesses(10) and some clinical isolates posse neither tdh nor trh(23 26) Other putative factors associated with V para-haemolyticus pathogenesis include urea hydrolysis the pro-duction of vibrioferrin and the presence of a type III e-cretion system (14 24 44) Currently standard methods
bull Author for correspondence Tel 805-756-2498 Fax 805-756-1419E-mail pmyeungcalpolyedu
for isolation and differentiation of V parahaemolyticus dic-tate that isolates determined to be tdh po itive by PCR orcolony hybridization (40) are potential human pathogenHowever these conventional detection strategie that pe-cifically detect tdh alone may fail to recognize potentiallyvirulent tdh-negative V parahaemolyticus strain
Because V parahaemolyticu trains can exchange ge-netic material (24) and hence genetic marker (1 33) wehypothesized that measurement of V parahaemolyticus vir-ulence-as ociated characteri tic i a more reliable and ro-bu t approach for detection of pathogenic V parahaemo-iyticus strain than is the pre ence or ab ence of specificgenetic markers In support of this concept previous workdemonstrated the potential of tis ue culture-based as aysfor detection of cytotoxic effects of pathogenic Vibriostrains (8) For example V parahaemolyticus serotype 03K6 strain have a clearly detectable cytopathogenic phe-notype in human epithelial HeLa cell (47)
A cytotoxicity assay was developed to measure releaseof lactate dehydrogenase (LDH) from mammalian cell asan indicator of cell damage in the presence of V parahae-molyticus The objectives of this tudy were to (i) quantifythe relative cytotoxicity of various V parahaemoiyticus iso-lates from food or clinical ource in multiple ti sue celllines representing different host cell types and (ii) assewhether a ti ue cuJture-based method could be u ed todetect the presence of pathogenic V parahaemolyticus pres-ent in a eafood matrix
-
J Food Prot Vol 70 No2 CYTOTOXICITIES OF V PARAHAEMOLYTlCVS 349
TABLE 1 Vibrio parahaemolyticu isolates used in this study
Laboratory no FDA idenuficanon Serotype Source Place of isolation Year of isolation rdll res-
FSL-YI-OOJ 92000713 (I) 08 Food (clam) Unknown 1992FSL-YI-002 NY477 04K8 Food (oy ter) New York 1977 +FSL-Y1-003 T3980 04K13 Clinical Japan Unknown +FSL-Yl-004 CRAB Unknown Food Wa hington 1972FS L-Y 1-00Sb SC-1C 01 Food (oy ter) Wa hington 1988FSL-YI-006 M3S0A OS ood (oyster) Washington 1994FSL- Y 1-0 LOb T-3979 05K15 Clinical Japan Unknown +FSL-Y iou- 8332924 0lK56 Food (oyster) Gulf 1983FSL-YI-012 48432 04K12 Clinical Washington 1991FSL-YI-013 47978 06K18 Clinical Washington 1991FSL- Y 1-0 [4b AN-5034 04K68c Clinical Bangladesh 1996 +FSL-YI-01S AN-16000 OIKUFd Clinical Bangladesh 1998 +FSL-Yl-016 TX-2103 03K6 Clinical Texas 1998 +FSL- Y1-0 17b NY-30M 03K6 Clinical New York 1998 +FSL-Yl-021b U-5474 Old 03K6 Clinical Bangladesh 1980 +FSL- Y 1-023b VP86 03K6 Clinical Calcutta 1996 +FSL- Y 1-0241gt VP199 03K6 Clinical Calcutta 1997 +FSL-Yl-025 VP208 03K6 Clinical Calcutta 1997 +FSL- Y 1-026b VPISS 03K6 Clinical Calcutta 1996 +FSL-Y1-036 FIHE 98VI-32-4 03K6 Clinical Japan 1998 +FSL-YI-046 BAC-98-03255 03K6 Clinical New York Unknown +FSL-YI-059 98- 792-807 (27) 08K74 Food (oyster) Galve ton Bay 1998FSL-YI-068 DI-A6-031699 04K9 Food (oyster) Alabama Unknown +FSL-YI-069 Dl-B 1l-031699 04K22 Food (oy ter) Alabama UnknownFSL-YI-073 DI-F8-031699 OIIKUTd Food (oy ter) Alabama UnknownFSL-YI-081 DI A6-020800 0J1K61 Food (oy ter) Alabama Unknown
a Results were compiled from a previou study (47) and from umpublished datab Isolates tested for CHO elongation activityc Strain deemed genetically similar to 03K6 train by arbitrarily primed PCR ribotype and pulsed-field gel electrophoresis pattern
analyses (5 26)d UT untypeablee 03K6 strain isolated before 1996 that is genetically different from the 03K6 strain i elated after 1996
MATERIAL A D METHOD
Bacterial isolates V parahaemolyticus i elate u ed in thistudy were provided by the US Food and Drug Administration(FDA) (Table I) Serotype were determined previously by theCenter for Disease Control and Prevention Upon receipt all iso-late were treaked on tryptic soy agar (Difco Becton DickinsonSparks Md) upplemented with 2 NaCl (TSAS) to obtain singlecolonies Isolates were tored in tryptic soy broth upplementedwith 2 NaCI (TSBS) and 20 glycerol at -80degC Isolate weresubcultured on TSAS at lea t once before each experiment
Tissue cell lines V parahaemolyticus cytotoxicity was de-termined in human epithelial HeLa cells (ATCC CCL-2) mousemacrophage-like J774AI cells (ATCC TIB-67) human epithelialHenle 407 cells (ATCC CCL-6) rat epithelial L2 cells (ATCCCCL-149) and the human colorectal epithelial cells Caco-2(ATCC HTB-37) The HeLa J774Al Henle 407 and L2 cellsWere maintained in Dulbeccos modified Eagles medium (GibcoBRL Invitrogen Gaither burg Md) upplemented with 10 fetalbOVine erum (FBS Gibco) aco-2 cells were maintained in min-imal es ential medium (Gibco) upplemented with 20 FBS TheCHO elongation factor a ay were conducted u ing Chine e ham-ster ovary cell CHO-IC6 (ATCC CRL-1793) that were main-tained in Hams Fl2K medium (Gibco) with 10 FBS All tis uecells were incubated at 37degC with 5 CO2
Cytotoxicity a ay A tissue culture-based as ay was usedto measure host cell cytotoxicity induced in the pre ence of sta-tionary-phase V parahaemolyticus cells A 500-11 aliquot of Vparahaemolyticus cell (8 X 108 to 2 X 109 CFUml a deter-mined by standard plate count in each experiment) that had beengrown overnight in TSBS with haking (250 rpm) at 37degC wascentrifuged at 17900 X g for 3 min and the pellet was re upended in 500 JL1of pho phate-buffered saline (PBS) The resupen ion was diluted 10-fold in PBS and a 15-11aliquot (ie toyield 15 JLIof the undiluted u pen ion) wa used to infect con-fluent monoJayers of tissue cells grown in 100 11 of the appro-priate medium in 96-well rnicrotiter plate (Fisher Scientific In-ternational Inc Hampton NH) At 2 h postinfection cytotoxicitywa asses ed by mea urement of relea ed ho t cell LDH using theCytoTox 96 on-Radioactive Cytotoxicity A say (Promega Mad-i on Wis) LDH oncentration wa mea ured by reading ab or-bance at 490 nm u ing the Fu ion Univer a1 microplate analyzer(Packard Instrument Co Meriden Conn) Relative cytotoxicitycalculations were ba ed on the following formula cytotoxicity() 100 X (Aample A pomaneolls)(AlOlUJ Aspolltaneous) whereAsample i the ab orbance of infected ti ue cells and Aspontaneou isthe ab orbance of uninfected tissue cells To determine Alot1gt themaximal relea e of LDH ti sue cell were lysed by adding 15 jLlof lysis solution according to the manufacturers instruction Theamount of lysis olution added was equivalent to a final concen-
-
--
- -
350 YEU GET AL J Food Prot Vol 70 o 2
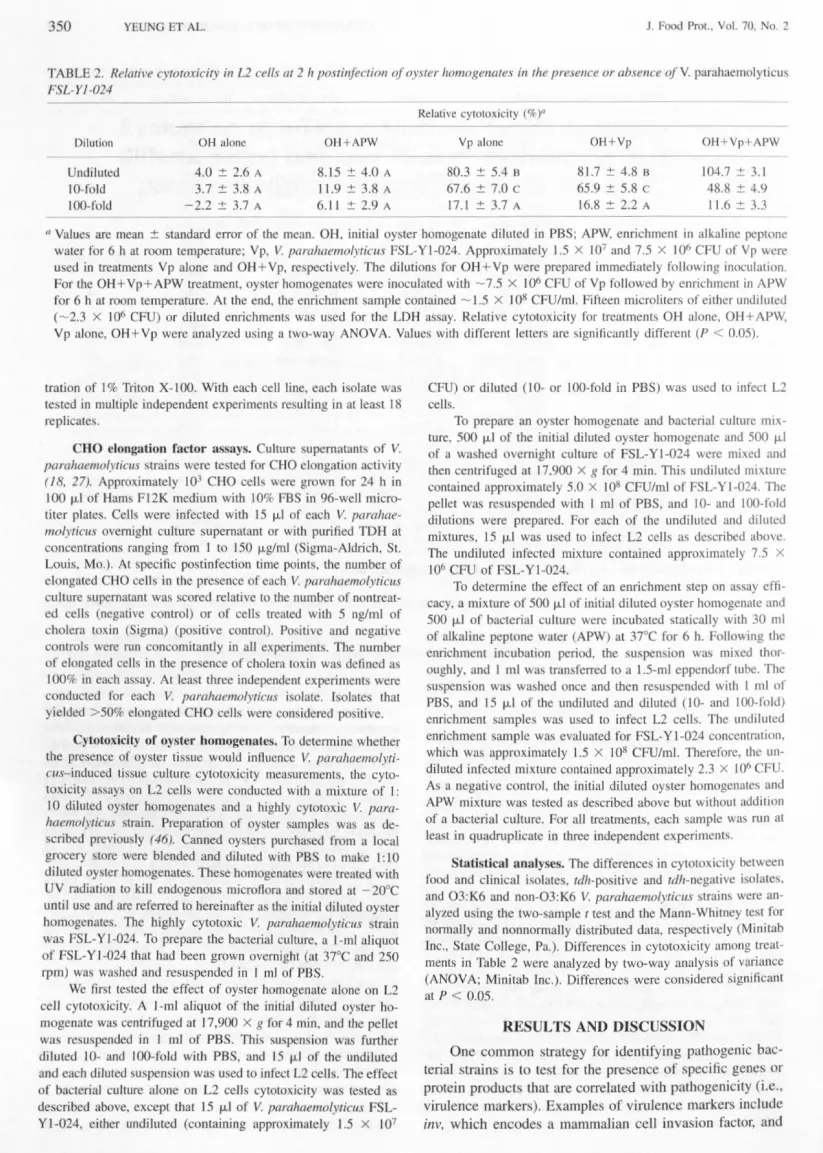
TABLE 2 Relative cytotoxicity in L2 cells at 2 Iipostinfection of oyster homogenate ill the presence or absence of Y parahaernolyticuFSL-YJ-024
Relative cytotoxicity ()
Dilution OH alone OH+APW Yp alone OH+Yp OH+Vp+APW
Undiluted 40 26 A 815 40 A 803 54 B 817 48 B 1047 31IMoid 37 38 A 119 38 A 676 t 70 c 659 58 c 488 49100-fold -22 37 A 611 29 A 171 37 A 168 22 A 116 33
II Value are mean tandard error of the mean OH initial oyster homogenate diluted in PBS APW enrichment in alkaline peptonewater for 6 h at room temperature Vp V parahaemoiyticus FSL-Yl-024 Approximately 15 x 101 and 75 X 106 CFU of Vp wereu ed in treatments Vp alone and OH + Vp respectively The dilutions for OH + Vp were prepared immediately following inoculationFor the OH+Vp+APW treatment oyster homogenates were inoculated with -75 X 106 CFU of Vp followed by enrichment in APWfor 6 h at room temperature At the end the enrichment ample contained -15 X J 08 CFUmJ Fifteen microliters of either undiluted(-23 X 106 CFU) or diluted enrichments was u ed for the LDH assay Relative cytotoxicity for treatments OH alone OH + APWVp alone OH+Vp were analyzed using a two-way ANOVA Values with different letters are significantly different (P lt 005)
tration of I Triton X-I 00 With each cell line each isolate wastested in multiple independent experiments reulting in at least 18replicates
HO elongation factor assays Culture upernatants of Vparahaemolyticus trains were tested for CHO elongation activity(18 27) Approximately 103 CHO cell were grown for 24 h In100 jJ1of Hams Fl2K medium with 10 FBS in 96-well micro-titer plate Cells were infected with 15 jJJof each V parawe-molyticu overnight culture supernatant or with purified TDH atconcentrations ranging from 1 to 150 ugrnl (Sigma-Aldrich StLouis Mo) At specific postinfection time points the number ofelongated CHO cells in the presence of each V parahaemolyticusculture upernatant was scored relative to the number of nontreat-ed cells (negative control) or of cell treated with 5 ngml ofcholera toxin (Sigma) (po itive control) Po itive and negativecontrols were run concomitantly in all experiment The numberof elongated cells in the pre ence of cholera toxin was defined asl00~ in each assay At lea t three independent experiments wereconducted for each V parahaemolyticus isolate Isolates thatyielded gt50~ elongated CHO cell were considered po itive
Cytotoxicity of oyster homogenates To determine whetherthe presence of oyster tissue would influence V parahaemoiyti-Cts-induced tissue culture cytotoxicity measurements the cyto-toxicity assays on L2 cell were conducted with a mixture of I10 diluted oy ter homogenates and a highly cytotoxic V para-haemolyticus train Preparation of oy ter ample wa as de-cribed previously (46) Canned oyster purcha ed from a local
grocery lore were blended and diluted with PBS to make 110diluted oyster hornogenates The e hornogenatcs were treated withUV radiation to kill endogenous microflora and stored at -20degCuruil u e and are referred to hereinafter a the initial diluted oy telhomogenate The highly cytotoxic V parahaemolyticus strainwas FSL-YI-024 To prepare the bacterial culture a l-rnl aliquotof FSL- YJ -024 that had been grown overnight (at 37degC and 250rpm) was washed and resuspended in I ml of PBS
We fir t tested the effect of oyster homogenate alone on L2cell cytotoxicity A l-ml aliquot of the initial diluted oyster ho-mogenate wa centrifuged at 17900 X g for 4 min and the pelletwas re upended in 1 mJ of PBS This suspen ion was furtherdiluted 10- and 100-fold with PBS and 15 fJ-1 of the undilutedand each diluted u pension wa u ed to infect L2 cell The effectof bacterial culture alone on L2 cell cytotoxicity wa te ted asdescribed above except that 15 jJ1of V parahaemolyeicus FSL-YI-024 either undiluted (containing approximately 15 x 107
CFU) or diluted (10- or 100-fold in PBS) was used to infect L2cell
To prepare an oyster homogenate and bacterial culture mix-ture 500 jJI of the initial diluted oyster homogenate and 500 jL1of a washed overnight culture of FSL- Y 1-024 were mixed andthen centrifuged at 17900 X g for 4 min This undiluted mixturecontained approximately 50 X 108 CFUlml of FSL-YI-024 Thepellet was resu ipended with 1 rnl of PBS and lO- and laO-folddilutions were prepared For each of the undiluted and dilutedmixtures 15 ul wa used to infect L2 cells as de cribed aboveThe undiluted infected mixture contained approximately 75 X106 CFU of FSL- Y] -024
To determine the effect of an enrichment step on assay effi-cacy a mixture of 500 fJ-1 of initial diluted oyster homogenate and500 fJ-1 of bacterial culture were incubated tatically with 30 mlof alkaline peptone water (APW) at 3rC for 6 h Following theenrichment incubation period the su pension wa mixed thor-oughly and I rnl wa tran ferred to a 15-1111eppendorf tube Thesuspension wa wa hed once and then resuspended with I rnl ofPBS and 15 jJ1of the undiluted and diluted (10- and lOa-fold)enrichment samples was used to infect L2 cell The undilutedenrichment sample was evaluated for FSL- Y 1-024 concentrationwhich was approximately 15 X 108 CFUm Therefore the un-diluted infected mixture contained approximately 23 X 106 CFUAs a negati ve control the initial diluted oyster homogcnates andAPW mixture wa tested a described above but without additionof a bacterial culture For all treatments each sample was run atleast in quadruplicate in three independent experiment
Statistical analyse The differences in cytotoxicity betweenfood and clinical i olate tdh-positive and edit-negative i elate and 03K6 and non-03K6 V parahaemolyticus trains were an-alyzed u ing the two- ample e test and the Mann-Whitney te t fornormally and nonnorrnally distributed data respectively (MinitabInc State College Pa) Differences in cytotoxicity among treat-ments in Table 2 were analyzed by two-way analysis of variance(ANOVA Minitab Inc) Differences were considered significantat P lt 005
RE ULTS AND DISCUSSIO
One common strategy for identifying pathogenic bac-terial strain i to te t for the presence of pecific gene orprotein products that are correlated with pathogenicity (ievirulence marker ) Examples of virulence marker includeinv which encode a mammalian cell invasion factor and
1 Food Prot Vol 70 o 2
ial which is the inva ion-associated locu The pre ence ofinv is associated with pathogenic strain of Yer inia pseu-dotuberculosis (12 39) and ial is highly specific for Shi-gella and enteroinvasive Escherichia coli (22) The reli-ability of virulence markers for differentiation of pathogen-ic strains i dependent on the universality of the presenceof the marker in strains capable of causing disease
The current standard method for detection of patho-genic V parahaemolyticus strain i ba ed on the presenceof tdh (encoding the TDH protein) and Irh (encoding TDH-related hemolysin) (40) Not all pathogenic V parahae-molyticus are positive for tdh and trh and the ability ofvibrios to exchange genetic material and genetic markers(I 33) trongly under core the need for development ofanalytical tools that can mea ure the functional capabilityof a pecific train to cau e di ea e We reasoned that ati ue culture approach that allowed mea urement of therelative cytopathogenicity of clinical V parahaemolyticusstrain and of strains not a ociated with human illnemight provide such a tool The relative cytotoxicity of Vparahaemofyticus isolates wa examined by measuringLDH relea e in several different ho t cell line We al atested the fea ibility of u ing LDH release a a detectionmethod for V parahaemoiyticus in eafood sample
Cytopathogenicity a say and groupings of isolateBecau e microorgani ill interact differently with host tisues from different source (15 41) we chose multiple tisue cell lines representing differ nt host celJ type includ-ing the HeLa cell line that has been used in previous tudieon V parahaemolyticus (28 35 47) to measure relativecytotoxicity of the isolate in our V parahaemolyticus col-lection Specifically we mea ured host cell release ofLDHan indigenou enzyme found in human and animal cells inthe presence of the V parahaemotyticus cultures The quan-tity of LDH released i proportional to the degree of dam-age to the host cell membrane (31) incurred by exposureto V parahaemolyticus
In Vibrio cholerae and Salmonella enterotoxigenicityi associated with production of a factor that cau es elon-gation of CHO cells (7 36) Therefore we al 0 conductedthe CHO cell elongation assay (i) to determine whether Vparahaemolyticus possesses a CHO cell elongation factorand if so (ii) to determine the fea ibiJity of this assay fordifferentiating between pathogenic and nonpathogenic bac-teria
The isolates used in this rudy were cla Hied into var-iou categories Clinical isolates were obtained from patienttool sample and food i olates (ie nonclinical isolates)
were obtained from food amples that had not been as 0-
ciated with any clinical ca e Isolate were also classifiedby serotype into 03K6 and non-03K6 groups V para-haemotvucus serotype 03K6 is an emerging pathogen hy-pothesized to have enhanced virulence relative to non-03K6 strains 03K6 trains and the closely related clonalgroups have caused a number of human disease outbreakworldwide since 1996 (3 4 6 20 34) Because TDH halong been regarded a a virulence marker for V parawe-
CYTOTOXICITTES OF V PARAHAEMOLYTfCUS 351
A) 700
600 ~
500
~ 400middot 300gt- -
lit ~n200
100
00l2j
HeLa J774AI L2 Henle 407 Ceeomiddot2
jocllnlcai 545 596 500 369 155
IFoOd 438 549 262 224 44
B) 000
000 ---700
gt- 000-
il 50000 400o
J)Oo200
100
00H9n1e407 ~2I-tLa J774Al L2
o Oinical031lt6 673 685 610 450 192
Oinical non-03K6 314 435 325 239 112
C) 700
600
500~ 400x
300
200
100
00HeLa J774Al
o tdh-positive 530 577 341 151
ldh-ne atlve 458 573 257 58
FIGURE 1 Cytotoxicity ( relative to complete chemical celllysis) in different cell lines at 2 h postinfection with V parahae-molyticu isolates Cytotoxicity wa measured by host LDH re-lease Error bars represent the standard error of the mean Meancytotoxicity percentages are noted in table below each figureFor statistical analyses isolates were grouped (A) food and clin-ical isolates (B) clinical non-03K6 versus 03K6 isolates and(C) tdh + versus tdh isolates An asterisk indicates that the cy-totoxicity difference between each group for a given cell line issignificantly different (P lt 005)
molyticus (29 32) we also compared the relative cytotox-icity of i olates with and without tdh
V parahaemolyticus isolates exbibit different cyto-toxic capabilities When grouped by source food and clin-ical isolates had different cytotoxic capabilitie For aU tis-sue culture cell lines the mean cytotoxicity of clinical i 0-
late was greater than that for food isolate (Fig IA) Clin-ical isolate exhibited 545 596 500 369 and 155relative cytotoxicity in HeLa J774Al L2 Henle 407 andCaco-Z cell line respectively whereas food i olates ex-hibited 438 549 262 224 and 44 relative cytotox-icity in th e respective cell line The relative cytotoxicityfor these two group wa ignificantly different (P lt 005)
--
- - -----~ ~
-
~
~
-
-
~
352 YEUNG ET AL
in the L2 Henle 407 and Caco-2 cell lines Cytotoxicityof the V parahaemolyticus 03K6 and non-03K6 i olatealso was compared Because all 03K6 isolates in our col-lection were from clinical sources only clinical non-03K6i olate were compared with clinical 03K6 isolates Cy-totoxicity differences between the e two group were ig-nificant in Henle 407 cells (450 with the 03K6 groupvel us 239 with the non-03K6 group) HeLa cells (673versus 314) and J774AI cell (685 versu 435) (FiglB) The differences in L2 cells (610 versu 325) weremarginally significant (P lt 010) Although tdh-positiveisolates appeared to have slightly higher cytotoxicity thantdh-negative i olates (Fig IC) the differences were signif-icant only for the Caco-2 cell line
Our findings from the LDH as ays support conclusionfrom previou tudies which suggested that the presenceof tdh alone does not fully correlate with cytotoxicity fluidaccumulation and other virulence measurements (9 2335) therefore tdh alone i not sufficient as a predictor forV parahaemotyticus pathogenicity Our re ult further ug-ge t that the apparent enhanced virulence of V parahae-molyticus 03K6 strain could be a sociated with the en-hanced cytotoxic effects that these cells have on host cell When cla ified into group by ource clinical isolates hadhigher relative cytotoxicity than did food isolates in threeho t cell lines In its current format however this LDHassay cannot reliably discriminate between clinical andnonclinical i olates at the strain level because the relativecytotoxicity percentage in these two group of isolatesoverlap Nevertheless with further development a ti ueculture-ba ed assay may be a useful approach for the de-tection of pathogenic trains becau e the outcome of suchan assay will be more dependent on the ability of a trainto cau e di ease than on the pre ence or absence of one ormore specific virulence determinant
CHO elongation factor is not useful for evaluationof V parahaemolyticus pathogenicity Although V para-haemolyticus i olates clearly differed in their cytotoxic ef-fects on host cells the organism does not appear to possessa CHO elongation factor under the conditions tested NineV parahaemolyticus isolates (Table 1) were creened forthe presence of the CHO elongation factor but none werepositive Thus the CHO elongation assay is not useful forcharacterization of V parahaemolyticus pathogenicity
Development of a tissue culture-based assay to de-tect cytotoxic V parahaemolyticus Becau e clinical iso-lates generally were more cytotoxic than were nonclinicali olates we further tested the efficacy of the LDH assay asa detection tool to screen for the pre ence of V parahae-molyticus strains with pathogenic potential in food sample Our goal with the e experiment wa to as e the abilityof our as ay to detect the presence of pathogenic V para-haemolyticus directly from an oyster tissue matrix Theability to detect potentially dangerou pathogens directlyfrom food samples could ignificantly reduce the analyticaltime needed to make deci ions related to hazards that maybe a ociated with bacterial contamination such as deter-mining the afety of oy ter bed or the need to issue a
J Food Prot Vol 70 o 2
product recal However food rnarrice can interfere withanalytical outcomes For example inhibitory sub tancepresent in food can lead to false-negative results with ornedetection technologic uch as tho e involving PCRs (19)Oyster sample were te ted becau e V parahaemolyticusinfection have been a ociated frequently with can ump-tion of raw or undercooked bivalve shellfi b We selectedL2 cells for a say development because the cytotoxicity dif-ference between clinical and nonclinical isolates was themost pronounced with thi cell line ie 6 (462) of 13clinical isolates but none of the food isolate had gt60relative cytotoxicity The fast-growing L2 cell line al a isea y to maintain V parahaemolyticu FSL- Y 1-024 was se-lected for the development of the screening a say becauseof it relative evere cytotoxic effects in all cell lines (datanot hown)
Approximately 5 X 108 CFU of FSL- YI-024 culturewa used to artificially inoculate the initial diluted oysterhomogenates and then 15 J-LIof the mixture (undiluted ordiluted 10- or 100-fold) were used to infect L2 cell eitherwith or without an enrichment step Results are summarizedin Table 2 A low level of cytotoxicity wa detected inoy tel homogenates and APW (treatments OR alone andOH + APW in Table 2) pre umably because of nonspecificLDH release from the oyster samples Thi nonspecificLDH relea e can tituted the background level for our assayWhen L2 cell were inoculated with 15 X 107 CFU ofVparahaemolyticus FSL- Yl-024 alone a strong response(803 cytotoxicity) wa detected A reduced response(676 cytotoxicity) wa detected with to-fold diluted cul-ture ie 15 X 106 CFU The cytotoxicity of the lOO-folddiluted culture was not significantly different from thebackground Component pre ent in oy ter homogenate including endogenous LDH did not appear to affect detec-tion of V parahaemoiyticus similar cytotoxicity was de-tected for FSL- Y1-024 either with or without oy tel ho-mogenates (treatments OH+Vp and Vp alone)
Although the addition of an enrichment tep to a de-tection assay increases the overall time required for a aycompletion thi step can improve assay sensitivity Wedemon trated previously that a 6-h enrichment period waoptimum for detecting a larger collection of V parahae-molyticus with a PCR method (46) Enrichment at an ap-propriate temperature Can allow injured cells to recover andviable-but-nonculturable (VBNC) cell to re u citate (1321 25) In thi regard an enrichment tep may be partic-ularly useful for V parahaemolyticu because of its abilityto enter into the VBNC tage (2 30) Wong et aI (43)found that VB C cell exhibited delayed mouse lethalityand enteropathogenicity compared with log-pha e cells Inthe natural environment V parahaemoiyticus al 0 may beinjured or starved Wong and Chang (42) found that a hortincubation wa neces ary to enable starved V parahae-molyticus cell to reach the same cytopathogenicity as ex-hibited by the log-pha e cells To balance the benefits ofenrichment with the need for a rapid assay a 6-h enrich-ment period wa selected for the pre ent study FSL- Yl-024 numbers increased by 126 log CFU following this en-richment Stati tical comparison of the treatm nt with an
1 Fo d Pror Vol 70 o 2
enrichment step (OH +Vp+ APW) and the other treatmentswa not conducted because bacterial cell number differedby gt05 log C U
Cytotoxicity mea urements following enrichment maynot be comparable to tho e without enrichment In thistudy V parohaemolyticus cell after the 6-h enrichment
were likely in late Jog to early tationary phase in contra tto the late tationary pha cell u ed in other treatm nt There i increa ing evidenc that virulen e of bacterialpathogen u h a Listeria monocytogene (15) and Strep-tococcus pneumoniae (17) is influenced by environmentaltr e and growth pha e Thus it i also pos ible that V
parahaemolyticu cytopath genicity may be influenced bygrowth phase Further tudy will b neces ary to asses theffect of growth pha on V parahaemolyticus virulencecharacteri tic
In its current format our LDH a say can differ ntiateclinical V parahaemolyticus strains only at the populationlevel uture a ay optimization also must tak into on id-eration the pre nee of other microfl ra in a typical t tmatrix Improved n itivity and pecificity will be neces-ary for application of thi approach a a routin detection
procedureonventi nal detection tools designed t e rabli h the
pre ence of ne genetic determinant or phenotypi char-acteri tic in a given i olate are likely to b of only limitedutility for det cting emerging path genic V parahaemoly-ticus train In th pre ent rudy we amined the utilityof a ti ue culture-based as ay for d velopment of detecti nmethod for pathogenic V parahaemolyticu train A agroup clinical i olate are more cytotoxic than nonclinieali alate Ba ed on LDH relea e re ult -106 cells of cy-totoxic V parahaemolyticus were detect d An ptimizedenrichment tep might further enhance the en itivity of thiassay Further insight into factor contributing to V para-haemolytieus pathogen si will enable development of newdetection method with increased sen itivity and discrimi-natory capabil itie
AK OWLEDGME TWe are grateful to Angelo DePa la and Charle Kay ner (US Food
and Drug Administrati n) r providing the V parahaemolyri us isolatcsu ed in this ludy We al 0 thank Helene Marqui and Edward Dubovi(Cornell University Ithaca Y) for as i tance with cell lines ThL workwa supported by a gram from tJle New York ea Gram pI gram (RI HH-J3) 10 K J Boor and M Wiedmann
REF RE EI Baba K H hirai A Terai K Kumagai Y Takeda and M Nish-
ibuclti 1991 Similarity of the rdh gene-bearing pIa mids of Vibrioeholerae noo-Ol and Vibrio parthaemolyriclls Microb PatllOg 1061-70
2 Bate T C and J D Oliver 2004 The viable but nonculturabletate of Kanagawa positive and negati e train of Vibrio parahaemiddot
l1lolytieus J Microbial 44274-793 Chi u C S Y Hsu I hiu T K Wang and C S Chao
2000 Vibrio parahaemolyriws serovar 03K6 as cause of unusuaJlyhigh incidence offood-bome di ease outbreak in Taiwan from 1996to 1999 J Clin Microbial 384621-4625
4 ChOWdhuryA M [ hiba hi V D Thiem D T Tuy t T V TungB T Chien L von eidlein D G Canh J Clemen D D Trachand M i hibuchi 2004 Emergence and erovar tran ition of Vibrio
CYTOTOXICIT1ES OF V PARAHAEMOLYTICUS 353
parahaemolyticus pandemic strains isolated during a diarrhea out-break in Vietnam between 1997 and 1999 Microbial lmmunol 48319-327
5 howdhury N R S Chakraborty T Ramamunhy M ishibuchiS Yama aki Y Takeda and G B Nair 2000 Molecular evidenceof clonal Vibrio parahaemolvti IS pandemic strains Emerg InfectDis 6631-636
6 Daniels N A B Ray A Easton Marano E Kahn A LMcShan II L Del Ro ario T Baldwin M A Kingsley N D PuhrJ G Wells and E J Angulo 2000 Emergence of a new Vibriopurahaemotyticus serotype in raw oy ters a prevenri n quandaryJAMA 2841541-1545
7 Guerrant R L L L Brunton T C chnaitrnan L I Rebhun andA G Gilman 1974 Cyclic adenosine rnonophosphate and alterationof hinese hamster ovary cell morphology a rapid sen itive in vitroas ay for the enterotoxins of Vibrio cholerae and Escherichia coliInfect lrnmun JO320-327
8 Hackney R E G Kleeman B Ray and M L Speck 1980Adherence as a method of differentiating virulent and avirulenttrains of Vibrio parahaemoiyticus Appl Environ Microbial 40
652-6589 Hoa hi K K Ogata H Taniguchi H Yamashita K Tsuji Y Mi-
zuguchi and Ohtorno 1990 Pathogenesis of Vibrio parahae-molyticus intraperitoneal and oroga tric challenge experiment inmice Microbiol Immunol 34355-366
10 Hondo l G 101 Minemal u Ikeda Asano M IshibashiY Kino hita ishibuchi T Honda and T Miwaiani 1987 Gatr emeriti due to Kanagawa negative Vibrio parahaemolyticus UII-
cet i33 1-332J J lijima Y H Yamada and hinoda 19 I Adherence of Vibrio
parahaemolyticu and it relation to pathogenicity Call J Micro-bioi 271252-1259
12 I berg R D L Vo rhi and Falkow 1987 Identification ofinvasin a protein thaL allows enteric bacteria to penetrate culturedmammalian cell Cell 50769-778
13 Johnston M D and M H Brown 2002 An investigation into thechanged physiological state of Vibrio bacteria as a survival mechamiddotni m in rc p n e to cold temperature and studie on their sen itivilyto heating and freezing J Appl Mi robiol 92 1066-1077
14 Kay ner C A C Abeyta Jr P A Tro t J H Wetherington KC Jinneman W E Hill and M M Wekell 1994 Urea hydrolysiscan predict the potential pathogenicity of Vibrio parahaemolyticlsstrain iolated in the Pacific Norlhwe t Appl Environ Microbiol603020-3022
15 Kim H H Marquis and K 1 Boor 2005 (18 contribute to Lisreriamono yrogenes inva ion by controlling expre sion of inlA and inlBMicmbiology 151 32]5-3222
16 Ki hishita M N Matsuoka K Kumagai S Yamasaki Y Takedaand M Ni hibuchi 1992 Sequence variation in the thermostabledirect hemoly in-related hemolysin (rrh) gene of Vibrio paralwe-molyricus Appl Environ Microbiol 582449-2457
17 Ko K S S Park W Oh J Y uh T Oh S Ahn J Chun andJ H Song 2006 omparative analysi of growth-phase dependentgene expre sion in virulent and avirulent Srreptocoecls pllelmoniaeu ing a high-den ity D A microarray Mol Cells 2182-88
18 Kothary M E R Claverie M D Milioti J M Madden and H Richard on J995 Purification and characLerizatjon of a Chineseham ter ovary cell elongation factor of Vibrio hollisae hrfeel 1m-IIlUII 632418-2423
19 Lantz P G W Abu almiddotSoud R Knuts on B HahnmiddotHagerdal andP Rad trom 2000 Biotechnical u of polymerase chain reactionfor microbiological analy i of biological sample BioteclllJol AmllRev 587-130
20 Laohaprertthi an v A Chowdhury U Kongmuang S Kalnaumiddotwakul M Ishibashi C Matsumoto and M i hibuchi 2003 Prev-alence and erodiversity of the pandemic clone among the clinicaltrains of Vibrio parahaemolyticus isolated in southern Thailand Ep-
idemiol Illfecr 130395-40621 Liao C H and W F Fen 2005 Re uscitation of acid-injured Sal-
~
-
354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909

J Food Prot Vol 70 No2 CYTOTOXICITIES OF V PARAHAEMOLYTlCVS 349
TABLE 1 Vibrio parahaemolyticu isolates used in this study
Laboratory no FDA idenuficanon Serotype Source Place of isolation Year of isolation rdll res-
FSL-YI-OOJ 92000713 (I) 08 Food (clam) Unknown 1992FSL-YI-002 NY477 04K8 Food (oy ter) New York 1977 +FSL-Y1-003 T3980 04K13 Clinical Japan Unknown +FSL-Yl-004 CRAB Unknown Food Wa hington 1972FS L-Y 1-00Sb SC-1C 01 Food (oy ter) Wa hington 1988FSL-YI-006 M3S0A OS ood (oyster) Washington 1994FSL- Y 1-0 LOb T-3979 05K15 Clinical Japan Unknown +FSL-Y iou- 8332924 0lK56 Food (oyster) Gulf 1983FSL-YI-012 48432 04K12 Clinical Washington 1991FSL-YI-013 47978 06K18 Clinical Washington 1991FSL- Y 1-0 [4b AN-5034 04K68c Clinical Bangladesh 1996 +FSL-YI-01S AN-16000 OIKUFd Clinical Bangladesh 1998 +FSL-Yl-016 TX-2103 03K6 Clinical Texas 1998 +FSL- Y1-0 17b NY-30M 03K6 Clinical New York 1998 +FSL-Yl-021b U-5474 Old 03K6 Clinical Bangladesh 1980 +FSL- Y 1-023b VP86 03K6 Clinical Calcutta 1996 +FSL- Y 1-0241gt VP199 03K6 Clinical Calcutta 1997 +FSL-Yl-025 VP208 03K6 Clinical Calcutta 1997 +FSL- Y 1-026b VPISS 03K6 Clinical Calcutta 1996 +FSL-Y1-036 FIHE 98VI-32-4 03K6 Clinical Japan 1998 +FSL-YI-046 BAC-98-03255 03K6 Clinical New York Unknown +FSL-YI-059 98- 792-807 (27) 08K74 Food (oyster) Galve ton Bay 1998FSL-YI-068 DI-A6-031699 04K9 Food (oyster) Alabama Unknown +FSL-YI-069 Dl-B 1l-031699 04K22 Food (oy ter) Alabama UnknownFSL-YI-073 DI-F8-031699 OIIKUTd Food (oy ter) Alabama UnknownFSL-YI-081 DI A6-020800 0J1K61 Food (oy ter) Alabama Unknown
a Results were compiled from a previou study (47) and from umpublished datab Isolates tested for CHO elongation activityc Strain deemed genetically similar to 03K6 train by arbitrarily primed PCR ribotype and pulsed-field gel electrophoresis pattern
analyses (5 26)d UT untypeablee 03K6 strain isolated before 1996 that is genetically different from the 03K6 strain i elated after 1996
MATERIAL A D METHOD
Bacterial isolates V parahaemolyticus i elate u ed in thistudy were provided by the US Food and Drug Administration(FDA) (Table I) Serotype were determined previously by theCenter for Disease Control and Prevention Upon receipt all iso-late were treaked on tryptic soy agar (Difco Becton DickinsonSparks Md) upplemented with 2 NaCl (TSAS) to obtain singlecolonies Isolates were tored in tryptic soy broth upplementedwith 2 NaCI (TSBS) and 20 glycerol at -80degC Isolate weresubcultured on TSAS at lea t once before each experiment
Tissue cell lines V parahaemolyticus cytotoxicity was de-termined in human epithelial HeLa cells (ATCC CCL-2) mousemacrophage-like J774AI cells (ATCC TIB-67) human epithelialHenle 407 cells (ATCC CCL-6) rat epithelial L2 cells (ATCCCCL-149) and the human colorectal epithelial cells Caco-2(ATCC HTB-37) The HeLa J774Al Henle 407 and L2 cellsWere maintained in Dulbeccos modified Eagles medium (GibcoBRL Invitrogen Gaither burg Md) upplemented with 10 fetalbOVine erum (FBS Gibco) aco-2 cells were maintained in min-imal es ential medium (Gibco) upplemented with 20 FBS TheCHO elongation factor a ay were conducted u ing Chine e ham-ster ovary cell CHO-IC6 (ATCC CRL-1793) that were main-tained in Hams Fl2K medium (Gibco) with 10 FBS All tis uecells were incubated at 37degC with 5 CO2
Cytotoxicity a ay A tissue culture-based as ay was usedto measure host cell cytotoxicity induced in the pre ence of sta-tionary-phase V parahaemolyticus cells A 500-11 aliquot of Vparahaemolyticus cell (8 X 108 to 2 X 109 CFUml a deter-mined by standard plate count in each experiment) that had beengrown overnight in TSBS with haking (250 rpm) at 37degC wascentrifuged at 17900 X g for 3 min and the pellet was re upended in 500 JL1of pho phate-buffered saline (PBS) The resupen ion was diluted 10-fold in PBS and a 15-11aliquot (ie toyield 15 JLIof the undiluted u pen ion) wa used to infect con-fluent monoJayers of tissue cells grown in 100 11 of the appro-priate medium in 96-well rnicrotiter plate (Fisher Scientific In-ternational Inc Hampton NH) At 2 h postinfection cytotoxicitywa asses ed by mea urement of relea ed ho t cell LDH using theCytoTox 96 on-Radioactive Cytotoxicity A say (Promega Mad-i on Wis) LDH oncentration wa mea ured by reading ab or-bance at 490 nm u ing the Fu ion Univer a1 microplate analyzer(Packard Instrument Co Meriden Conn) Relative cytotoxicitycalculations were ba ed on the following formula cytotoxicity() 100 X (Aample A pomaneolls)(AlOlUJ Aspolltaneous) whereAsample i the ab orbance of infected ti ue cells and Aspontaneou isthe ab orbance of uninfected tissue cells To determine Alot1gt themaximal relea e of LDH ti sue cell were lysed by adding 15 jLlof lysis solution according to the manufacturers instruction Theamount of lysis olution added was equivalent to a final concen-
-
--
- -
350 YEU GET AL J Food Prot Vol 70 o 2
TABLE 2 Relative cytotoxicity in L2 cells at 2 Iipostinfection of oyster homogenate ill the presence or absence of Y parahaernolyticuFSL-YJ-024
Relative cytotoxicity ()
Dilution OH alone OH+APW Yp alone OH+Yp OH+Vp+APW
Undiluted 40 26 A 815 40 A 803 54 B 817 48 B 1047 31IMoid 37 38 A 119 38 A 676 t 70 c 659 58 c 488 49100-fold -22 37 A 611 29 A 171 37 A 168 22 A 116 33
II Value are mean tandard error of the mean OH initial oyster homogenate diluted in PBS APW enrichment in alkaline peptonewater for 6 h at room temperature Vp V parahaemoiyticus FSL-Yl-024 Approximately 15 x 101 and 75 X 106 CFU of Vp wereu ed in treatments Vp alone and OH + Vp respectively The dilutions for OH + Vp were prepared immediately following inoculationFor the OH+Vp+APW treatment oyster homogenates were inoculated with -75 X 106 CFU of Vp followed by enrichment in APWfor 6 h at room temperature At the end the enrichment ample contained -15 X J 08 CFUmJ Fifteen microliters of either undiluted(-23 X 106 CFU) or diluted enrichments was u ed for the LDH assay Relative cytotoxicity for treatments OH alone OH + APWVp alone OH+Vp were analyzed using a two-way ANOVA Values with different letters are significantly different (P lt 005)
tration of I Triton X-I 00 With each cell line each isolate wastested in multiple independent experiments reulting in at least 18replicates
HO elongation factor assays Culture upernatants of Vparahaemolyticus trains were tested for CHO elongation activity(18 27) Approximately 103 CHO cell were grown for 24 h In100 jJ1of Hams Fl2K medium with 10 FBS in 96-well micro-titer plate Cells were infected with 15 jJJof each V parawe-molyticu overnight culture supernatant or with purified TDH atconcentrations ranging from 1 to 150 ugrnl (Sigma-Aldrich StLouis Mo) At specific postinfection time points the number ofelongated CHO cells in the presence of each V parahaemolyticusculture upernatant was scored relative to the number of nontreat-ed cells (negative control) or of cell treated with 5 ngml ofcholera toxin (Sigma) (po itive control) Po itive and negativecontrols were run concomitantly in all experiment The numberof elongated cells in the pre ence of cholera toxin was defined asl00~ in each assay At lea t three independent experiments wereconducted for each V parahaemolyticus isolate Isolates thatyielded gt50~ elongated CHO cell were considered po itive
Cytotoxicity of oyster homogenates To determine whetherthe presence of oyster tissue would influence V parahaemoiyti-Cts-induced tissue culture cytotoxicity measurements the cyto-toxicity assays on L2 cell were conducted with a mixture of I10 diluted oy ter homogenates and a highly cytotoxic V para-haemolyticus train Preparation of oy ter ample wa as de-cribed previously (46) Canned oyster purcha ed from a local
grocery lore were blended and diluted with PBS to make 110diluted oyster hornogenates The e hornogenatcs were treated withUV radiation to kill endogenous microflora and stored at -20degCuruil u e and are referred to hereinafter a the initial diluted oy telhomogenate The highly cytotoxic V parahaemolyticus strainwas FSL-YI-024 To prepare the bacterial culture a l-rnl aliquotof FSL- YJ -024 that had been grown overnight (at 37degC and 250rpm) was washed and resuspended in I ml of PBS
We fir t tested the effect of oyster homogenate alone on L2cell cytotoxicity A l-ml aliquot of the initial diluted oyster ho-mogenate wa centrifuged at 17900 X g for 4 min and the pelletwas re upended in 1 mJ of PBS This suspen ion was furtherdiluted 10- and 100-fold with PBS and 15 fJ-1 of the undilutedand each diluted u pension wa u ed to infect L2 cell The effectof bacterial culture alone on L2 cell cytotoxicity wa te ted asdescribed above except that 15 jJ1of V parahaemolyeicus FSL-YI-024 either undiluted (containing approximately 15 x 107
CFU) or diluted (10- or 100-fold in PBS) was used to infect L2cell
To prepare an oyster homogenate and bacterial culture mix-ture 500 jJI of the initial diluted oyster homogenate and 500 jL1of a washed overnight culture of FSL- Y 1-024 were mixed andthen centrifuged at 17900 X g for 4 min This undiluted mixturecontained approximately 50 X 108 CFUlml of FSL-YI-024 Thepellet was resu ipended with 1 rnl of PBS and lO- and laO-folddilutions were prepared For each of the undiluted and dilutedmixtures 15 ul wa used to infect L2 cells as de cribed aboveThe undiluted infected mixture contained approximately 75 X106 CFU of FSL- Y] -024
To determine the effect of an enrichment step on assay effi-cacy a mixture of 500 fJ-1 of initial diluted oyster homogenate and500 fJ-1 of bacterial culture were incubated tatically with 30 mlof alkaline peptone water (APW) at 3rC for 6 h Following theenrichment incubation period the su pension wa mixed thor-oughly and I rnl wa tran ferred to a 15-1111eppendorf tube Thesuspension wa wa hed once and then resuspended with I rnl ofPBS and 15 jJ1of the undiluted and diluted (10- and lOa-fold)enrichment samples was used to infect L2 cell The undilutedenrichment sample was evaluated for FSL- Y 1-024 concentrationwhich was approximately 15 X 108 CFUm Therefore the un-diluted infected mixture contained approximately 23 X 106 CFUAs a negati ve control the initial diluted oyster homogcnates andAPW mixture wa tested a described above but without additionof a bacterial culture For all treatments each sample was run atleast in quadruplicate in three independent experiment
Statistical analyse The differences in cytotoxicity betweenfood and clinical i olate tdh-positive and edit-negative i elate and 03K6 and non-03K6 V parahaemolyticus trains were an-alyzed u ing the two- ample e test and the Mann-Whitney te t fornormally and nonnorrnally distributed data respectively (MinitabInc State College Pa) Differences in cytotoxicity among treat-ments in Table 2 were analyzed by two-way analysis of variance(ANOVA Minitab Inc) Differences were considered significantat P lt 005
RE ULTS AND DISCUSSIO
One common strategy for identifying pathogenic bac-terial strain i to te t for the presence of pecific gene orprotein products that are correlated with pathogenicity (ievirulence marker ) Examples of virulence marker includeinv which encode a mammalian cell invasion factor and
1 Food Prot Vol 70 o 2
ial which is the inva ion-associated locu The pre ence ofinv is associated with pathogenic strain of Yer inia pseu-dotuberculosis (12 39) and ial is highly specific for Shi-gella and enteroinvasive Escherichia coli (22) The reli-ability of virulence markers for differentiation of pathogen-ic strains i dependent on the universality of the presenceof the marker in strains capable of causing disease
The current standard method for detection of patho-genic V parahaemolyticus strain i ba ed on the presenceof tdh (encoding the TDH protein) and Irh (encoding TDH-related hemolysin) (40) Not all pathogenic V parahae-molyticus are positive for tdh and trh and the ability ofvibrios to exchange genetic material and genetic markers(I 33) trongly under core the need for development ofanalytical tools that can mea ure the functional capabilityof a pecific train to cau e di ea e We reasoned that ati ue culture approach that allowed mea urement of therelative cytopathogenicity of clinical V parahaemolyticusstrain and of strains not a ociated with human illnemight provide such a tool The relative cytotoxicity of Vparahaemofyticus isolates wa examined by measuringLDH relea e in several different ho t cell line We al atested the fea ibility of u ing LDH release a a detectionmethod for V parahaemoiyticus in eafood sample
Cytopathogenicity a say and groupings of isolateBecau e microorgani ill interact differently with host tisues from different source (15 41) we chose multiple tisue cell lines representing differ nt host celJ type includ-ing the HeLa cell line that has been used in previous tudieon V parahaemolyticus (28 35 47) to measure relativecytotoxicity of the isolate in our V parahaemolyticus col-lection Specifically we mea ured host cell release ofLDHan indigenou enzyme found in human and animal cells inthe presence of the V parahaemotyticus cultures The quan-tity of LDH released i proportional to the degree of dam-age to the host cell membrane (31) incurred by exposureto V parahaemolyticus
In Vibrio cholerae and Salmonella enterotoxigenicityi associated with production of a factor that cau es elon-gation of CHO cells (7 36) Therefore we al 0 conductedthe CHO cell elongation assay (i) to determine whether Vparahaemolyticus possesses a CHO cell elongation factorand if so (ii) to determine the fea ibiJity of this assay fordifferentiating between pathogenic and nonpathogenic bac-teria
The isolates used in this rudy were cla Hied into var-iou categories Clinical isolates were obtained from patienttool sample and food i olates (ie nonclinical isolates)
were obtained from food amples that had not been as 0-
ciated with any clinical ca e Isolate were also classifiedby serotype into 03K6 and non-03K6 groups V para-haemotvucus serotype 03K6 is an emerging pathogen hy-pothesized to have enhanced virulence relative to non-03K6 strains 03K6 trains and the closely related clonalgroups have caused a number of human disease outbreakworldwide since 1996 (3 4 6 20 34) Because TDH halong been regarded a a virulence marker for V parawe-
CYTOTOXICITTES OF V PARAHAEMOLYTfCUS 351
A) 700
600 ~
500
~ 400middot 300gt- -
lit ~n200
100
00l2j
HeLa J774AI L2 Henle 407 Ceeomiddot2
jocllnlcai 545 596 500 369 155
IFoOd 438 549 262 224 44
B) 000
000 ---700
gt- 000-
il 50000 400o
J)Oo200
100
00H9n1e407 ~2I-tLa J774Al L2
o Oinical031lt6 673 685 610 450 192
Oinical non-03K6 314 435 325 239 112
C) 700
600
500~ 400x
300
200
100
00HeLa J774Al
o tdh-positive 530 577 341 151
ldh-ne atlve 458 573 257 58
FIGURE 1 Cytotoxicity ( relative to complete chemical celllysis) in different cell lines at 2 h postinfection with V parahae-molyticu isolates Cytotoxicity wa measured by host LDH re-lease Error bars represent the standard error of the mean Meancytotoxicity percentages are noted in table below each figureFor statistical analyses isolates were grouped (A) food and clin-ical isolates (B) clinical non-03K6 versus 03K6 isolates and(C) tdh + versus tdh isolates An asterisk indicates that the cy-totoxicity difference between each group for a given cell line issignificantly different (P lt 005)
molyticus (29 32) we also compared the relative cytotox-icity of i olates with and without tdh
V parahaemolyticus isolates exbibit different cyto-toxic capabilities When grouped by source food and clin-ical isolates had different cytotoxic capabilitie For aU tis-sue culture cell lines the mean cytotoxicity of clinical i 0-
late was greater than that for food isolate (Fig IA) Clin-ical isolate exhibited 545 596 500 369 and 155relative cytotoxicity in HeLa J774Al L2 Henle 407 andCaco-Z cell line respectively whereas food i olates ex-hibited 438 549 262 224 and 44 relative cytotox-icity in th e respective cell line The relative cytotoxicityfor these two group wa ignificantly different (P lt 005)
--
- - -----~ ~
-
~
~
-
-
~
352 YEUNG ET AL
in the L2 Henle 407 and Caco-2 cell lines Cytotoxicityof the V parahaemolyticus 03K6 and non-03K6 i olatealso was compared Because all 03K6 isolates in our col-lection were from clinical sources only clinical non-03K6i olate were compared with clinical 03K6 isolates Cy-totoxicity differences between the e two group were ig-nificant in Henle 407 cells (450 with the 03K6 groupvel us 239 with the non-03K6 group) HeLa cells (673versus 314) and J774AI cell (685 versu 435) (FiglB) The differences in L2 cells (610 versu 325) weremarginally significant (P lt 010) Although tdh-positiveisolates appeared to have slightly higher cytotoxicity thantdh-negative i olates (Fig IC) the differences were signif-icant only for the Caco-2 cell line
Our findings from the LDH as ays support conclusionfrom previou tudies which suggested that the presenceof tdh alone does not fully correlate with cytotoxicity fluidaccumulation and other virulence measurements (9 2335) therefore tdh alone i not sufficient as a predictor forV parahaemotyticus pathogenicity Our re ult further ug-ge t that the apparent enhanced virulence of V parahae-molyticus 03K6 strain could be a sociated with the en-hanced cytotoxic effects that these cells have on host cell When cla ified into group by ource clinical isolates hadhigher relative cytotoxicity than did food isolates in threeho t cell lines In its current format however this LDHassay cannot reliably discriminate between clinical andnonclinical i olates at the strain level because the relativecytotoxicity percentage in these two group of isolatesoverlap Nevertheless with further development a ti ueculture-ba ed assay may be a useful approach for the de-tection of pathogenic trains becau e the outcome of suchan assay will be more dependent on the ability of a trainto cau e di ease than on the pre ence or absence of one ormore specific virulence determinant
CHO elongation factor is not useful for evaluationof V parahaemolyticus pathogenicity Although V para-haemolyticus i olates clearly differed in their cytotoxic ef-fects on host cells the organism does not appear to possessa CHO elongation factor under the conditions tested NineV parahaemolyticus isolates (Table 1) were creened forthe presence of the CHO elongation factor but none werepositive Thus the CHO elongation assay is not useful forcharacterization of V parahaemolyticus pathogenicity
Development of a tissue culture-based assay to de-tect cytotoxic V parahaemolyticus Becau e clinical iso-lates generally were more cytotoxic than were nonclinicali olates we further tested the efficacy of the LDH assay asa detection tool to screen for the pre ence of V parahae-molyticus strains with pathogenic potential in food sample Our goal with the e experiment wa to as e the abilityof our as ay to detect the presence of pathogenic V para-haemolyticus directly from an oyster tissue matrix Theability to detect potentially dangerou pathogens directlyfrom food samples could ignificantly reduce the analyticaltime needed to make deci ions related to hazards that maybe a ociated with bacterial contamination such as deter-mining the afety of oy ter bed or the need to issue a
J Food Prot Vol 70 o 2
product recal However food rnarrice can interfere withanalytical outcomes For example inhibitory sub tancepresent in food can lead to false-negative results with ornedetection technologic uch as tho e involving PCRs (19)Oyster sample were te ted becau e V parahaemolyticusinfection have been a ociated frequently with can ump-tion of raw or undercooked bivalve shellfi b We selectedL2 cells for a say development because the cytotoxicity dif-ference between clinical and nonclinical isolates was themost pronounced with thi cell line ie 6 (462) of 13clinical isolates but none of the food isolate had gt60relative cytotoxicity The fast-growing L2 cell line al a isea y to maintain V parahaemolyticu FSL- Y 1-024 was se-lected for the development of the screening a say becauseof it relative evere cytotoxic effects in all cell lines (datanot hown)
Approximately 5 X 108 CFU of FSL- YI-024 culturewa used to artificially inoculate the initial diluted oysterhomogenates and then 15 J-LIof the mixture (undiluted ordiluted 10- or 100-fold) were used to infect L2 cell eitherwith or without an enrichment step Results are summarizedin Table 2 A low level of cytotoxicity wa detected inoy tel homogenates and APW (treatments OR alone andOH + APW in Table 2) pre umably because of nonspecificLDH release from the oyster samples Thi nonspecificLDH relea e can tituted the background level for our assayWhen L2 cell were inoculated with 15 X 107 CFU ofVparahaemolyticus FSL- Yl-024 alone a strong response(803 cytotoxicity) wa detected A reduced response(676 cytotoxicity) wa detected with to-fold diluted cul-ture ie 15 X 106 CFU The cytotoxicity of the lOO-folddiluted culture was not significantly different from thebackground Component pre ent in oy ter homogenate including endogenous LDH did not appear to affect detec-tion of V parahaemoiyticus similar cytotoxicity was de-tected for FSL- Y1-024 either with or without oy tel ho-mogenates (treatments OH+Vp and Vp alone)
Although the addition of an enrichment tep to a de-tection assay increases the overall time required for a aycompletion thi step can improve assay sensitivity Wedemon trated previously that a 6-h enrichment period waoptimum for detecting a larger collection of V parahae-molyticus with a PCR method (46) Enrichment at an ap-propriate temperature Can allow injured cells to recover andviable-but-nonculturable (VBNC) cell to re u citate (1321 25) In thi regard an enrichment tep may be partic-ularly useful for V parahaemolyticu because of its abilityto enter into the VBNC tage (2 30) Wong et aI (43)found that VB C cell exhibited delayed mouse lethalityand enteropathogenicity compared with log-pha e cells Inthe natural environment V parahaemoiyticus al 0 may beinjured or starved Wong and Chang (42) found that a hortincubation wa neces ary to enable starved V parahae-molyticus cell to reach the same cytopathogenicity as ex-hibited by the log-pha e cells To balance the benefits ofenrichment with the need for a rapid assay a 6-h enrich-ment period wa selected for the pre ent study FSL- Yl-024 numbers increased by 126 log CFU following this en-richment Stati tical comparison of the treatm nt with an
1 Fo d Pror Vol 70 o 2
enrichment step (OH +Vp+ APW) and the other treatmentswa not conducted because bacterial cell number differedby gt05 log C U
Cytotoxicity mea urements following enrichment maynot be comparable to tho e without enrichment In thistudy V parohaemolyticus cell after the 6-h enrichment
were likely in late Jog to early tationary phase in contra tto the late tationary pha cell u ed in other treatm nt There i increa ing evidenc that virulen e of bacterialpathogen u h a Listeria monocytogene (15) and Strep-tococcus pneumoniae (17) is influenced by environmentaltr e and growth pha e Thus it i also pos ible that V
parahaemolyticu cytopath genicity may be influenced bygrowth phase Further tudy will b neces ary to asses theffect of growth pha on V parahaemolyticus virulencecharacteri tic
In its current format our LDH a say can differ ntiateclinical V parahaemolyticus strains only at the populationlevel uture a ay optimization also must tak into on id-eration the pre nee of other microfl ra in a typical t tmatrix Improved n itivity and pecificity will be neces-ary for application of thi approach a a routin detection
procedureonventi nal detection tools designed t e rabli h the
pre ence of ne genetic determinant or phenotypi char-acteri tic in a given i olate are likely to b of only limitedutility for det cting emerging path genic V parahaemoly-ticus train In th pre ent rudy we amined the utilityof a ti ue culture-based as ay for d velopment of detecti nmethod for pathogenic V parahaemolyticu train A agroup clinical i olate are more cytotoxic than nonclinieali alate Ba ed on LDH relea e re ult -106 cells of cy-totoxic V parahaemolyticus were detect d An ptimizedenrichment tep might further enhance the en itivity of thiassay Further insight into factor contributing to V para-haemolytieus pathogen si will enable development of newdetection method with increased sen itivity and discrimi-natory capabil itie
AK OWLEDGME TWe are grateful to Angelo DePa la and Charle Kay ner (US Food
and Drug Administrati n) r providing the V parahaemolyri us isolatcsu ed in this ludy We al 0 thank Helene Marqui and Edward Dubovi(Cornell University Ithaca Y) for as i tance with cell lines ThL workwa supported by a gram from tJle New York ea Gram pI gram (RI HH-J3) 10 K J Boor and M Wiedmann
REF RE EI Baba K H hirai A Terai K Kumagai Y Takeda and M Nish-
ibuclti 1991 Similarity of the rdh gene-bearing pIa mids of Vibrioeholerae noo-Ol and Vibrio parthaemolyriclls Microb PatllOg 1061-70
2 Bate T C and J D Oliver 2004 The viable but nonculturabletate of Kanagawa positive and negati e train of Vibrio parahaemiddot
l1lolytieus J Microbial 44274-793 Chi u C S Y Hsu I hiu T K Wang and C S Chao
2000 Vibrio parahaemolyriws serovar 03K6 as cause of unusuaJlyhigh incidence offood-bome di ease outbreak in Taiwan from 1996to 1999 J Clin Microbial 384621-4625
4 ChOWdhuryA M [ hiba hi V D Thiem D T Tuy t T V TungB T Chien L von eidlein D G Canh J Clemen D D Trachand M i hibuchi 2004 Emergence and erovar tran ition of Vibrio
CYTOTOXICIT1ES OF V PARAHAEMOLYTICUS 353
parahaemolyticus pandemic strains isolated during a diarrhea out-break in Vietnam between 1997 and 1999 Microbial lmmunol 48319-327
5 howdhury N R S Chakraborty T Ramamunhy M ishibuchiS Yama aki Y Takeda and G B Nair 2000 Molecular evidenceof clonal Vibrio parahaemolvti IS pandemic strains Emerg InfectDis 6631-636
6 Daniels N A B Ray A Easton Marano E Kahn A LMcShan II L Del Ro ario T Baldwin M A Kingsley N D PuhrJ G Wells and E J Angulo 2000 Emergence of a new Vibriopurahaemotyticus serotype in raw oy ters a prevenri n quandaryJAMA 2841541-1545
7 Guerrant R L L L Brunton T C chnaitrnan L I Rebhun andA G Gilman 1974 Cyclic adenosine rnonophosphate and alterationof hinese hamster ovary cell morphology a rapid sen itive in vitroas ay for the enterotoxins of Vibrio cholerae and Escherichia coliInfect lrnmun JO320-327
8 Hackney R E G Kleeman B Ray and M L Speck 1980Adherence as a method of differentiating virulent and avirulenttrains of Vibrio parahaemoiyticus Appl Environ Microbial 40
652-6589 Hoa hi K K Ogata H Taniguchi H Yamashita K Tsuji Y Mi-
zuguchi and Ohtorno 1990 Pathogenesis of Vibrio parahae-molyticus intraperitoneal and oroga tric challenge experiment inmice Microbiol Immunol 34355-366
10 Hondo l G 101 Minemal u Ikeda Asano M IshibashiY Kino hita ishibuchi T Honda and T Miwaiani 1987 Gatr emeriti due to Kanagawa negative Vibrio parahaemolyticus UII-
cet i33 1-332J J lijima Y H Yamada and hinoda 19 I Adherence of Vibrio
parahaemolyticu and it relation to pathogenicity Call J Micro-bioi 271252-1259
12 I berg R D L Vo rhi and Falkow 1987 Identification ofinvasin a protein thaL allows enteric bacteria to penetrate culturedmammalian cell Cell 50769-778
13 Johnston M D and M H Brown 2002 An investigation into thechanged physiological state of Vibrio bacteria as a survival mechamiddotni m in rc p n e to cold temperature and studie on their sen itivilyto heating and freezing J Appl Mi robiol 92 1066-1077
14 Kay ner C A C Abeyta Jr P A Tro t J H Wetherington KC Jinneman W E Hill and M M Wekell 1994 Urea hydrolysiscan predict the potential pathogenicity of Vibrio parahaemolyticlsstrain iolated in the Pacific Norlhwe t Appl Environ Microbiol603020-3022
15 Kim H H Marquis and K 1 Boor 2005 (18 contribute to Lisreriamono yrogenes inva ion by controlling expre sion of inlA and inlBMicmbiology 151 32]5-3222
16 Ki hishita M N Matsuoka K Kumagai S Yamasaki Y Takedaand M Ni hibuchi 1992 Sequence variation in the thermostabledirect hemoly in-related hemolysin (rrh) gene of Vibrio paralwe-molyricus Appl Environ Microbiol 582449-2457
17 Ko K S S Park W Oh J Y uh T Oh S Ahn J Chun andJ H Song 2006 omparative analysi of growth-phase dependentgene expre sion in virulent and avirulent Srreptocoecls pllelmoniaeu ing a high-den ity D A microarray Mol Cells 2182-88
18 Kothary M E R Claverie M D Milioti J M Madden and H Richard on J995 Purification and characLerizatjon of a Chineseham ter ovary cell elongation factor of Vibrio hollisae hrfeel 1m-IIlUII 632418-2423
19 Lantz P G W Abu almiddotSoud R Knuts on B HahnmiddotHagerdal andP Rad trom 2000 Biotechnical u of polymerase chain reactionfor microbiological analy i of biological sample BioteclllJol AmllRev 587-130
20 Laohaprertthi an v A Chowdhury U Kongmuang S Kalnaumiddotwakul M Ishibashi C Matsumoto and M i hibuchi 2003 Prev-alence and erodiversity of the pandemic clone among the clinicaltrains of Vibrio parahaemolyticus isolated in southern Thailand Ep-
idemiol Illfecr 130395-40621 Liao C H and W F Fen 2005 Re uscitation of acid-injured Sal-
~
-
354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909
350 YEU GET AL J Food Prot Vol 70 o 2
TABLE 2 Relative cytotoxicity in L2 cells at 2 Iipostinfection of oyster homogenate ill the presence or absence of Y parahaernolyticuFSL-YJ-024
Relative cytotoxicity ()
Dilution OH alone OH+APW Yp alone OH+Yp OH+Vp+APW
Undiluted 40 26 A 815 40 A 803 54 B 817 48 B 1047 31IMoid 37 38 A 119 38 A 676 t 70 c 659 58 c 488 49100-fold -22 37 A 611 29 A 171 37 A 168 22 A 116 33
II Value are mean tandard error of the mean OH initial oyster homogenate diluted in PBS APW enrichment in alkaline peptonewater for 6 h at room temperature Vp V parahaemoiyticus FSL-Yl-024 Approximately 15 x 101 and 75 X 106 CFU of Vp wereu ed in treatments Vp alone and OH + Vp respectively The dilutions for OH + Vp were prepared immediately following inoculationFor the OH+Vp+APW treatment oyster homogenates were inoculated with -75 X 106 CFU of Vp followed by enrichment in APWfor 6 h at room temperature At the end the enrichment ample contained -15 X J 08 CFUmJ Fifteen microliters of either undiluted(-23 X 106 CFU) or diluted enrichments was u ed for the LDH assay Relative cytotoxicity for treatments OH alone OH + APWVp alone OH+Vp were analyzed using a two-way ANOVA Values with different letters are significantly different (P lt 005)
tration of I Triton X-I 00 With each cell line each isolate wastested in multiple independent experiments reulting in at least 18replicates
HO elongation factor assays Culture upernatants of Vparahaemolyticus trains were tested for CHO elongation activity(18 27) Approximately 103 CHO cell were grown for 24 h In100 jJ1of Hams Fl2K medium with 10 FBS in 96-well micro-titer plate Cells were infected with 15 jJJof each V parawe-molyticu overnight culture supernatant or with purified TDH atconcentrations ranging from 1 to 150 ugrnl (Sigma-Aldrich StLouis Mo) At specific postinfection time points the number ofelongated CHO cells in the presence of each V parahaemolyticusculture upernatant was scored relative to the number of nontreat-ed cells (negative control) or of cell treated with 5 ngml ofcholera toxin (Sigma) (po itive control) Po itive and negativecontrols were run concomitantly in all experiment The numberof elongated cells in the pre ence of cholera toxin was defined asl00~ in each assay At lea t three independent experiments wereconducted for each V parahaemolyticus isolate Isolates thatyielded gt50~ elongated CHO cell were considered po itive
Cytotoxicity of oyster homogenates To determine whetherthe presence of oyster tissue would influence V parahaemoiyti-Cts-induced tissue culture cytotoxicity measurements the cyto-toxicity assays on L2 cell were conducted with a mixture of I10 diluted oy ter homogenates and a highly cytotoxic V para-haemolyticus train Preparation of oy ter ample wa as de-cribed previously (46) Canned oyster purcha ed from a local
grocery lore were blended and diluted with PBS to make 110diluted oyster hornogenates The e hornogenatcs were treated withUV radiation to kill endogenous microflora and stored at -20degCuruil u e and are referred to hereinafter a the initial diluted oy telhomogenate The highly cytotoxic V parahaemolyticus strainwas FSL-YI-024 To prepare the bacterial culture a l-rnl aliquotof FSL- YJ -024 that had been grown overnight (at 37degC and 250rpm) was washed and resuspended in I ml of PBS
We fir t tested the effect of oyster homogenate alone on L2cell cytotoxicity A l-ml aliquot of the initial diluted oyster ho-mogenate wa centrifuged at 17900 X g for 4 min and the pelletwas re upended in 1 mJ of PBS This suspen ion was furtherdiluted 10- and 100-fold with PBS and 15 fJ-1 of the undilutedand each diluted u pension wa u ed to infect L2 cell The effectof bacterial culture alone on L2 cell cytotoxicity wa te ted asdescribed above except that 15 jJ1of V parahaemolyeicus FSL-YI-024 either undiluted (containing approximately 15 x 107
CFU) or diluted (10- or 100-fold in PBS) was used to infect L2cell
To prepare an oyster homogenate and bacterial culture mix-ture 500 jJI of the initial diluted oyster homogenate and 500 jL1of a washed overnight culture of FSL- Y 1-024 were mixed andthen centrifuged at 17900 X g for 4 min This undiluted mixturecontained approximately 50 X 108 CFUlml of FSL-YI-024 Thepellet was resu ipended with 1 rnl of PBS and lO- and laO-folddilutions were prepared For each of the undiluted and dilutedmixtures 15 ul wa used to infect L2 cells as de cribed aboveThe undiluted infected mixture contained approximately 75 X106 CFU of FSL- Y] -024
To determine the effect of an enrichment step on assay effi-cacy a mixture of 500 fJ-1 of initial diluted oyster homogenate and500 fJ-1 of bacterial culture were incubated tatically with 30 mlof alkaline peptone water (APW) at 3rC for 6 h Following theenrichment incubation period the su pension wa mixed thor-oughly and I rnl wa tran ferred to a 15-1111eppendorf tube Thesuspension wa wa hed once and then resuspended with I rnl ofPBS and 15 jJ1of the undiluted and diluted (10- and lOa-fold)enrichment samples was used to infect L2 cell The undilutedenrichment sample was evaluated for FSL- Y 1-024 concentrationwhich was approximately 15 X 108 CFUm Therefore the un-diluted infected mixture contained approximately 23 X 106 CFUAs a negati ve control the initial diluted oyster homogcnates andAPW mixture wa tested a described above but without additionof a bacterial culture For all treatments each sample was run atleast in quadruplicate in three independent experiment
Statistical analyse The differences in cytotoxicity betweenfood and clinical i olate tdh-positive and edit-negative i elate and 03K6 and non-03K6 V parahaemolyticus trains were an-alyzed u ing the two- ample e test and the Mann-Whitney te t fornormally and nonnorrnally distributed data respectively (MinitabInc State College Pa) Differences in cytotoxicity among treat-ments in Table 2 were analyzed by two-way analysis of variance(ANOVA Minitab Inc) Differences were considered significantat P lt 005
RE ULTS AND DISCUSSIO
One common strategy for identifying pathogenic bac-terial strain i to te t for the presence of pecific gene orprotein products that are correlated with pathogenicity (ievirulence marker ) Examples of virulence marker includeinv which encode a mammalian cell invasion factor and
1 Food Prot Vol 70 o 2
ial which is the inva ion-associated locu The pre ence ofinv is associated with pathogenic strain of Yer inia pseu-dotuberculosis (12 39) and ial is highly specific for Shi-gella and enteroinvasive Escherichia coli (22) The reli-ability of virulence markers for differentiation of pathogen-ic strains i dependent on the universality of the presenceof the marker in strains capable of causing disease
The current standard method for detection of patho-genic V parahaemolyticus strain i ba ed on the presenceof tdh (encoding the TDH protein) and Irh (encoding TDH-related hemolysin) (40) Not all pathogenic V parahae-molyticus are positive for tdh and trh and the ability ofvibrios to exchange genetic material and genetic markers(I 33) trongly under core the need for development ofanalytical tools that can mea ure the functional capabilityof a pecific train to cau e di ea e We reasoned that ati ue culture approach that allowed mea urement of therelative cytopathogenicity of clinical V parahaemolyticusstrain and of strains not a ociated with human illnemight provide such a tool The relative cytotoxicity of Vparahaemofyticus isolates wa examined by measuringLDH relea e in several different ho t cell line We al atested the fea ibility of u ing LDH release a a detectionmethod for V parahaemoiyticus in eafood sample
Cytopathogenicity a say and groupings of isolateBecau e microorgani ill interact differently with host tisues from different source (15 41) we chose multiple tisue cell lines representing differ nt host celJ type includ-ing the HeLa cell line that has been used in previous tudieon V parahaemolyticus (28 35 47) to measure relativecytotoxicity of the isolate in our V parahaemolyticus col-lection Specifically we mea ured host cell release ofLDHan indigenou enzyme found in human and animal cells inthe presence of the V parahaemotyticus cultures The quan-tity of LDH released i proportional to the degree of dam-age to the host cell membrane (31) incurred by exposureto V parahaemolyticus
In Vibrio cholerae and Salmonella enterotoxigenicityi associated with production of a factor that cau es elon-gation of CHO cells (7 36) Therefore we al 0 conductedthe CHO cell elongation assay (i) to determine whether Vparahaemolyticus possesses a CHO cell elongation factorand if so (ii) to determine the fea ibiJity of this assay fordifferentiating between pathogenic and nonpathogenic bac-teria
The isolates used in this rudy were cla Hied into var-iou categories Clinical isolates were obtained from patienttool sample and food i olates (ie nonclinical isolates)
were obtained from food amples that had not been as 0-
ciated with any clinical ca e Isolate were also classifiedby serotype into 03K6 and non-03K6 groups V para-haemotvucus serotype 03K6 is an emerging pathogen hy-pothesized to have enhanced virulence relative to non-03K6 strains 03K6 trains and the closely related clonalgroups have caused a number of human disease outbreakworldwide since 1996 (3 4 6 20 34) Because TDH halong been regarded a a virulence marker for V parawe-
CYTOTOXICITTES OF V PARAHAEMOLYTfCUS 351
A) 700
600 ~
500
~ 400middot 300gt- -
lit ~n200
100
00l2j
HeLa J774AI L2 Henle 407 Ceeomiddot2
jocllnlcai 545 596 500 369 155
IFoOd 438 549 262 224 44
B) 000
000 ---700
gt- 000-
il 50000 400o
J)Oo200
100
00H9n1e407 ~2I-tLa J774Al L2
o Oinical031lt6 673 685 610 450 192
Oinical non-03K6 314 435 325 239 112
C) 700
600
500~ 400x
300
200
100
00HeLa J774Al
o tdh-positive 530 577 341 151
ldh-ne atlve 458 573 257 58
FIGURE 1 Cytotoxicity ( relative to complete chemical celllysis) in different cell lines at 2 h postinfection with V parahae-molyticu isolates Cytotoxicity wa measured by host LDH re-lease Error bars represent the standard error of the mean Meancytotoxicity percentages are noted in table below each figureFor statistical analyses isolates were grouped (A) food and clin-ical isolates (B) clinical non-03K6 versus 03K6 isolates and(C) tdh + versus tdh isolates An asterisk indicates that the cy-totoxicity difference between each group for a given cell line issignificantly different (P lt 005)
molyticus (29 32) we also compared the relative cytotox-icity of i olates with and without tdh
V parahaemolyticus isolates exbibit different cyto-toxic capabilities When grouped by source food and clin-ical isolates had different cytotoxic capabilitie For aU tis-sue culture cell lines the mean cytotoxicity of clinical i 0-
late was greater than that for food isolate (Fig IA) Clin-ical isolate exhibited 545 596 500 369 and 155relative cytotoxicity in HeLa J774Al L2 Henle 407 andCaco-Z cell line respectively whereas food i olates ex-hibited 438 549 262 224 and 44 relative cytotox-icity in th e respective cell line The relative cytotoxicityfor these two group wa ignificantly different (P lt 005)
--
- - -----~ ~
-
~
~
-
-
~
352 YEUNG ET AL
in the L2 Henle 407 and Caco-2 cell lines Cytotoxicityof the V parahaemolyticus 03K6 and non-03K6 i olatealso was compared Because all 03K6 isolates in our col-lection were from clinical sources only clinical non-03K6i olate were compared with clinical 03K6 isolates Cy-totoxicity differences between the e two group were ig-nificant in Henle 407 cells (450 with the 03K6 groupvel us 239 with the non-03K6 group) HeLa cells (673versus 314) and J774AI cell (685 versu 435) (FiglB) The differences in L2 cells (610 versu 325) weremarginally significant (P lt 010) Although tdh-positiveisolates appeared to have slightly higher cytotoxicity thantdh-negative i olates (Fig IC) the differences were signif-icant only for the Caco-2 cell line
Our findings from the LDH as ays support conclusionfrom previou tudies which suggested that the presenceof tdh alone does not fully correlate with cytotoxicity fluidaccumulation and other virulence measurements (9 2335) therefore tdh alone i not sufficient as a predictor forV parahaemotyticus pathogenicity Our re ult further ug-ge t that the apparent enhanced virulence of V parahae-molyticus 03K6 strain could be a sociated with the en-hanced cytotoxic effects that these cells have on host cell When cla ified into group by ource clinical isolates hadhigher relative cytotoxicity than did food isolates in threeho t cell lines In its current format however this LDHassay cannot reliably discriminate between clinical andnonclinical i olates at the strain level because the relativecytotoxicity percentage in these two group of isolatesoverlap Nevertheless with further development a ti ueculture-ba ed assay may be a useful approach for the de-tection of pathogenic trains becau e the outcome of suchan assay will be more dependent on the ability of a trainto cau e di ease than on the pre ence or absence of one ormore specific virulence determinant
CHO elongation factor is not useful for evaluationof V parahaemolyticus pathogenicity Although V para-haemolyticus i olates clearly differed in their cytotoxic ef-fects on host cells the organism does not appear to possessa CHO elongation factor under the conditions tested NineV parahaemolyticus isolates (Table 1) were creened forthe presence of the CHO elongation factor but none werepositive Thus the CHO elongation assay is not useful forcharacterization of V parahaemolyticus pathogenicity
Development of a tissue culture-based assay to de-tect cytotoxic V parahaemolyticus Becau e clinical iso-lates generally were more cytotoxic than were nonclinicali olates we further tested the efficacy of the LDH assay asa detection tool to screen for the pre ence of V parahae-molyticus strains with pathogenic potential in food sample Our goal with the e experiment wa to as e the abilityof our as ay to detect the presence of pathogenic V para-haemolyticus directly from an oyster tissue matrix Theability to detect potentially dangerou pathogens directlyfrom food samples could ignificantly reduce the analyticaltime needed to make deci ions related to hazards that maybe a ociated with bacterial contamination such as deter-mining the afety of oy ter bed or the need to issue a
J Food Prot Vol 70 o 2
product recal However food rnarrice can interfere withanalytical outcomes For example inhibitory sub tancepresent in food can lead to false-negative results with ornedetection technologic uch as tho e involving PCRs (19)Oyster sample were te ted becau e V parahaemolyticusinfection have been a ociated frequently with can ump-tion of raw or undercooked bivalve shellfi b We selectedL2 cells for a say development because the cytotoxicity dif-ference between clinical and nonclinical isolates was themost pronounced with thi cell line ie 6 (462) of 13clinical isolates but none of the food isolate had gt60relative cytotoxicity The fast-growing L2 cell line al a isea y to maintain V parahaemolyticu FSL- Y 1-024 was se-lected for the development of the screening a say becauseof it relative evere cytotoxic effects in all cell lines (datanot hown)
Approximately 5 X 108 CFU of FSL- YI-024 culturewa used to artificially inoculate the initial diluted oysterhomogenates and then 15 J-LIof the mixture (undiluted ordiluted 10- or 100-fold) were used to infect L2 cell eitherwith or without an enrichment step Results are summarizedin Table 2 A low level of cytotoxicity wa detected inoy tel homogenates and APW (treatments OR alone andOH + APW in Table 2) pre umably because of nonspecificLDH release from the oyster samples Thi nonspecificLDH relea e can tituted the background level for our assayWhen L2 cell were inoculated with 15 X 107 CFU ofVparahaemolyticus FSL- Yl-024 alone a strong response(803 cytotoxicity) wa detected A reduced response(676 cytotoxicity) wa detected with to-fold diluted cul-ture ie 15 X 106 CFU The cytotoxicity of the lOO-folddiluted culture was not significantly different from thebackground Component pre ent in oy ter homogenate including endogenous LDH did not appear to affect detec-tion of V parahaemoiyticus similar cytotoxicity was de-tected for FSL- Y1-024 either with or without oy tel ho-mogenates (treatments OH+Vp and Vp alone)
Although the addition of an enrichment tep to a de-tection assay increases the overall time required for a aycompletion thi step can improve assay sensitivity Wedemon trated previously that a 6-h enrichment period waoptimum for detecting a larger collection of V parahae-molyticus with a PCR method (46) Enrichment at an ap-propriate temperature Can allow injured cells to recover andviable-but-nonculturable (VBNC) cell to re u citate (1321 25) In thi regard an enrichment tep may be partic-ularly useful for V parahaemolyticu because of its abilityto enter into the VBNC tage (2 30) Wong et aI (43)found that VB C cell exhibited delayed mouse lethalityand enteropathogenicity compared with log-pha e cells Inthe natural environment V parahaemoiyticus al 0 may beinjured or starved Wong and Chang (42) found that a hortincubation wa neces ary to enable starved V parahae-molyticus cell to reach the same cytopathogenicity as ex-hibited by the log-pha e cells To balance the benefits ofenrichment with the need for a rapid assay a 6-h enrich-ment period wa selected for the pre ent study FSL- Yl-024 numbers increased by 126 log CFU following this en-richment Stati tical comparison of the treatm nt with an
1 Fo d Pror Vol 70 o 2
enrichment step (OH +Vp+ APW) and the other treatmentswa not conducted because bacterial cell number differedby gt05 log C U
Cytotoxicity mea urements following enrichment maynot be comparable to tho e without enrichment In thistudy V parohaemolyticus cell after the 6-h enrichment
were likely in late Jog to early tationary phase in contra tto the late tationary pha cell u ed in other treatm nt There i increa ing evidenc that virulen e of bacterialpathogen u h a Listeria monocytogene (15) and Strep-tococcus pneumoniae (17) is influenced by environmentaltr e and growth pha e Thus it i also pos ible that V
parahaemolyticu cytopath genicity may be influenced bygrowth phase Further tudy will b neces ary to asses theffect of growth pha on V parahaemolyticus virulencecharacteri tic
In its current format our LDH a say can differ ntiateclinical V parahaemolyticus strains only at the populationlevel uture a ay optimization also must tak into on id-eration the pre nee of other microfl ra in a typical t tmatrix Improved n itivity and pecificity will be neces-ary for application of thi approach a a routin detection
procedureonventi nal detection tools designed t e rabli h the
pre ence of ne genetic determinant or phenotypi char-acteri tic in a given i olate are likely to b of only limitedutility for det cting emerging path genic V parahaemoly-ticus train In th pre ent rudy we amined the utilityof a ti ue culture-based as ay for d velopment of detecti nmethod for pathogenic V parahaemolyticu train A agroup clinical i olate are more cytotoxic than nonclinieali alate Ba ed on LDH relea e re ult -106 cells of cy-totoxic V parahaemolyticus were detect d An ptimizedenrichment tep might further enhance the en itivity of thiassay Further insight into factor contributing to V para-haemolytieus pathogen si will enable development of newdetection method with increased sen itivity and discrimi-natory capabil itie
AK OWLEDGME TWe are grateful to Angelo DePa la and Charle Kay ner (US Food
and Drug Administrati n) r providing the V parahaemolyri us isolatcsu ed in this ludy We al 0 thank Helene Marqui and Edward Dubovi(Cornell University Ithaca Y) for as i tance with cell lines ThL workwa supported by a gram from tJle New York ea Gram pI gram (RI HH-J3) 10 K J Boor and M Wiedmann
REF RE EI Baba K H hirai A Terai K Kumagai Y Takeda and M Nish-
ibuclti 1991 Similarity of the rdh gene-bearing pIa mids of Vibrioeholerae noo-Ol and Vibrio parthaemolyriclls Microb PatllOg 1061-70
2 Bate T C and J D Oliver 2004 The viable but nonculturabletate of Kanagawa positive and negati e train of Vibrio parahaemiddot
l1lolytieus J Microbial 44274-793 Chi u C S Y Hsu I hiu T K Wang and C S Chao
2000 Vibrio parahaemolyriws serovar 03K6 as cause of unusuaJlyhigh incidence offood-bome di ease outbreak in Taiwan from 1996to 1999 J Clin Microbial 384621-4625
4 ChOWdhuryA M [ hiba hi V D Thiem D T Tuy t T V TungB T Chien L von eidlein D G Canh J Clemen D D Trachand M i hibuchi 2004 Emergence and erovar tran ition of Vibrio
CYTOTOXICIT1ES OF V PARAHAEMOLYTICUS 353
parahaemolyticus pandemic strains isolated during a diarrhea out-break in Vietnam between 1997 and 1999 Microbial lmmunol 48319-327
5 howdhury N R S Chakraborty T Ramamunhy M ishibuchiS Yama aki Y Takeda and G B Nair 2000 Molecular evidenceof clonal Vibrio parahaemolvti IS pandemic strains Emerg InfectDis 6631-636
6 Daniels N A B Ray A Easton Marano E Kahn A LMcShan II L Del Ro ario T Baldwin M A Kingsley N D PuhrJ G Wells and E J Angulo 2000 Emergence of a new Vibriopurahaemotyticus serotype in raw oy ters a prevenri n quandaryJAMA 2841541-1545
7 Guerrant R L L L Brunton T C chnaitrnan L I Rebhun andA G Gilman 1974 Cyclic adenosine rnonophosphate and alterationof hinese hamster ovary cell morphology a rapid sen itive in vitroas ay for the enterotoxins of Vibrio cholerae and Escherichia coliInfect lrnmun JO320-327
8 Hackney R E G Kleeman B Ray and M L Speck 1980Adherence as a method of differentiating virulent and avirulenttrains of Vibrio parahaemoiyticus Appl Environ Microbial 40
652-6589 Hoa hi K K Ogata H Taniguchi H Yamashita K Tsuji Y Mi-
zuguchi and Ohtorno 1990 Pathogenesis of Vibrio parahae-molyticus intraperitoneal and oroga tric challenge experiment inmice Microbiol Immunol 34355-366
10 Hondo l G 101 Minemal u Ikeda Asano M IshibashiY Kino hita ishibuchi T Honda and T Miwaiani 1987 Gatr emeriti due to Kanagawa negative Vibrio parahaemolyticus UII-
cet i33 1-332J J lijima Y H Yamada and hinoda 19 I Adherence of Vibrio
parahaemolyticu and it relation to pathogenicity Call J Micro-bioi 271252-1259
12 I berg R D L Vo rhi and Falkow 1987 Identification ofinvasin a protein thaL allows enteric bacteria to penetrate culturedmammalian cell Cell 50769-778
13 Johnston M D and M H Brown 2002 An investigation into thechanged physiological state of Vibrio bacteria as a survival mechamiddotni m in rc p n e to cold temperature and studie on their sen itivilyto heating and freezing J Appl Mi robiol 92 1066-1077
14 Kay ner C A C Abeyta Jr P A Tro t J H Wetherington KC Jinneman W E Hill and M M Wekell 1994 Urea hydrolysiscan predict the potential pathogenicity of Vibrio parahaemolyticlsstrain iolated in the Pacific Norlhwe t Appl Environ Microbiol603020-3022
15 Kim H H Marquis and K 1 Boor 2005 (18 contribute to Lisreriamono yrogenes inva ion by controlling expre sion of inlA and inlBMicmbiology 151 32]5-3222
16 Ki hishita M N Matsuoka K Kumagai S Yamasaki Y Takedaand M Ni hibuchi 1992 Sequence variation in the thermostabledirect hemoly in-related hemolysin (rrh) gene of Vibrio paralwe-molyricus Appl Environ Microbiol 582449-2457
17 Ko K S S Park W Oh J Y uh T Oh S Ahn J Chun andJ H Song 2006 omparative analysi of growth-phase dependentgene expre sion in virulent and avirulent Srreptocoecls pllelmoniaeu ing a high-den ity D A microarray Mol Cells 2182-88
18 Kothary M E R Claverie M D Milioti J M Madden and H Richard on J995 Purification and characLerizatjon of a Chineseham ter ovary cell elongation factor of Vibrio hollisae hrfeel 1m-IIlUII 632418-2423
19 Lantz P G W Abu almiddotSoud R Knuts on B HahnmiddotHagerdal andP Rad trom 2000 Biotechnical u of polymerase chain reactionfor microbiological analy i of biological sample BioteclllJol AmllRev 587-130
20 Laohaprertthi an v A Chowdhury U Kongmuang S Kalnaumiddotwakul M Ishibashi C Matsumoto and M i hibuchi 2003 Prev-alence and erodiversity of the pandemic clone among the clinicaltrains of Vibrio parahaemolyticus isolated in southern Thailand Ep-
idemiol Illfecr 130395-40621 Liao C H and W F Fen 2005 Re uscitation of acid-injured Sal-
~
-
354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909
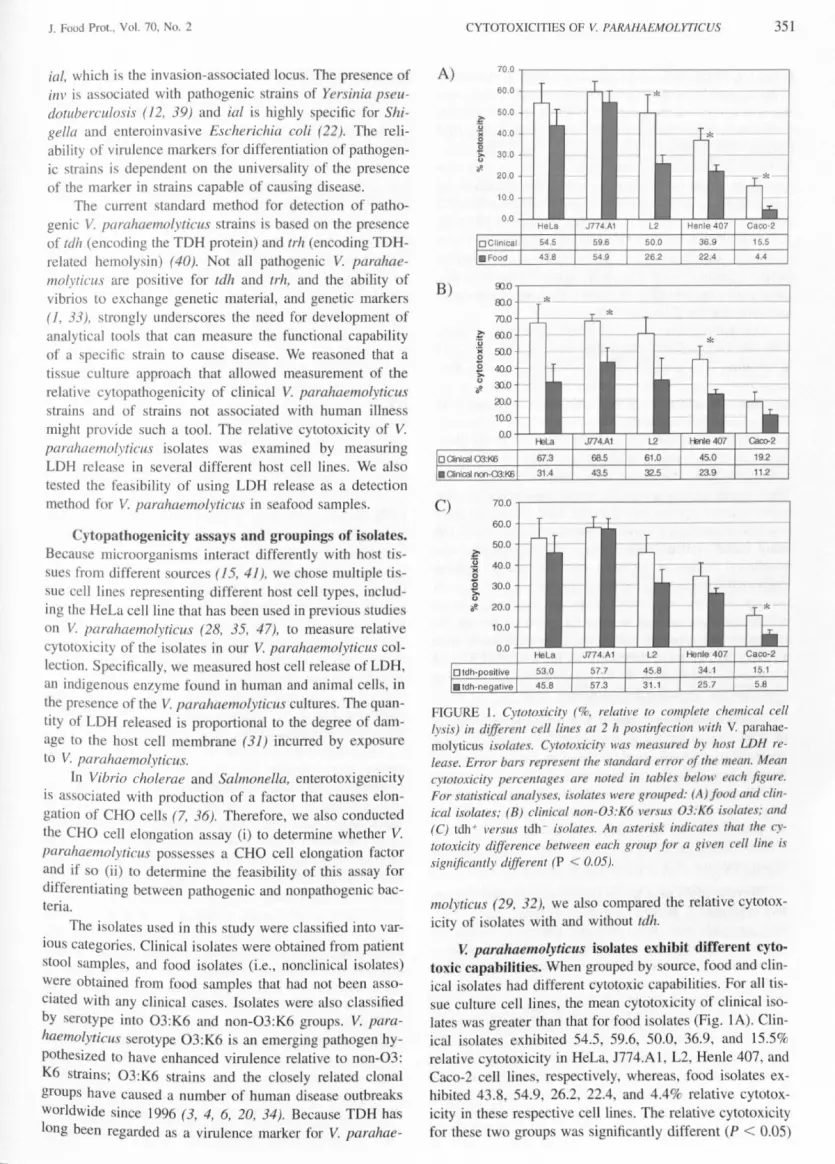
1 Food Prot Vol 70 o 2
ial which is the inva ion-associated locu The pre ence ofinv is associated with pathogenic strain of Yer inia pseu-dotuberculosis (12 39) and ial is highly specific for Shi-gella and enteroinvasive Escherichia coli (22) The reli-ability of virulence markers for differentiation of pathogen-ic strains i dependent on the universality of the presenceof the marker in strains capable of causing disease
The current standard method for detection of patho-genic V parahaemolyticus strain i ba ed on the presenceof tdh (encoding the TDH protein) and Irh (encoding TDH-related hemolysin) (40) Not all pathogenic V parahae-molyticus are positive for tdh and trh and the ability ofvibrios to exchange genetic material and genetic markers(I 33) trongly under core the need for development ofanalytical tools that can mea ure the functional capabilityof a pecific train to cau e di ea e We reasoned that ati ue culture approach that allowed mea urement of therelative cytopathogenicity of clinical V parahaemolyticusstrain and of strains not a ociated with human illnemight provide such a tool The relative cytotoxicity of Vparahaemofyticus isolates wa examined by measuringLDH relea e in several different ho t cell line We al atested the fea ibility of u ing LDH release a a detectionmethod for V parahaemoiyticus in eafood sample
Cytopathogenicity a say and groupings of isolateBecau e microorgani ill interact differently with host tisues from different source (15 41) we chose multiple tisue cell lines representing differ nt host celJ type includ-ing the HeLa cell line that has been used in previous tudieon V parahaemolyticus (28 35 47) to measure relativecytotoxicity of the isolate in our V parahaemolyticus col-lection Specifically we mea ured host cell release ofLDHan indigenou enzyme found in human and animal cells inthe presence of the V parahaemotyticus cultures The quan-tity of LDH released i proportional to the degree of dam-age to the host cell membrane (31) incurred by exposureto V parahaemolyticus
In Vibrio cholerae and Salmonella enterotoxigenicityi associated with production of a factor that cau es elon-gation of CHO cells (7 36) Therefore we al 0 conductedthe CHO cell elongation assay (i) to determine whether Vparahaemolyticus possesses a CHO cell elongation factorand if so (ii) to determine the fea ibiJity of this assay fordifferentiating between pathogenic and nonpathogenic bac-teria
The isolates used in this rudy were cla Hied into var-iou categories Clinical isolates were obtained from patienttool sample and food i olates (ie nonclinical isolates)
were obtained from food amples that had not been as 0-
ciated with any clinical ca e Isolate were also classifiedby serotype into 03K6 and non-03K6 groups V para-haemotvucus serotype 03K6 is an emerging pathogen hy-pothesized to have enhanced virulence relative to non-03K6 strains 03K6 trains and the closely related clonalgroups have caused a number of human disease outbreakworldwide since 1996 (3 4 6 20 34) Because TDH halong been regarded a a virulence marker for V parawe-
CYTOTOXICITTES OF V PARAHAEMOLYTfCUS 351
A) 700
600 ~
500
~ 400middot 300gt- -
lit ~n200
100
00l2j
HeLa J774AI L2 Henle 407 Ceeomiddot2
jocllnlcai 545 596 500 369 155
IFoOd 438 549 262 224 44
B) 000
000 ---700
gt- 000-
il 50000 400o
J)Oo200
100
00H9n1e407 ~2I-tLa J774Al L2
o Oinical031lt6 673 685 610 450 192
Oinical non-03K6 314 435 325 239 112
C) 700
600
500~ 400x
300
200
100
00HeLa J774Al
o tdh-positive 530 577 341 151
ldh-ne atlve 458 573 257 58
FIGURE 1 Cytotoxicity ( relative to complete chemical celllysis) in different cell lines at 2 h postinfection with V parahae-molyticu isolates Cytotoxicity wa measured by host LDH re-lease Error bars represent the standard error of the mean Meancytotoxicity percentages are noted in table below each figureFor statistical analyses isolates were grouped (A) food and clin-ical isolates (B) clinical non-03K6 versus 03K6 isolates and(C) tdh + versus tdh isolates An asterisk indicates that the cy-totoxicity difference between each group for a given cell line issignificantly different (P lt 005)
molyticus (29 32) we also compared the relative cytotox-icity of i olates with and without tdh
V parahaemolyticus isolates exbibit different cyto-toxic capabilities When grouped by source food and clin-ical isolates had different cytotoxic capabilitie For aU tis-sue culture cell lines the mean cytotoxicity of clinical i 0-
late was greater than that for food isolate (Fig IA) Clin-ical isolate exhibited 545 596 500 369 and 155relative cytotoxicity in HeLa J774Al L2 Henle 407 andCaco-Z cell line respectively whereas food i olates ex-hibited 438 549 262 224 and 44 relative cytotox-icity in th e respective cell line The relative cytotoxicityfor these two group wa ignificantly different (P lt 005)
--
- - -----~ ~
-
~
~
-
-
~
352 YEUNG ET AL
in the L2 Henle 407 and Caco-2 cell lines Cytotoxicityof the V parahaemolyticus 03K6 and non-03K6 i olatealso was compared Because all 03K6 isolates in our col-lection were from clinical sources only clinical non-03K6i olate were compared with clinical 03K6 isolates Cy-totoxicity differences between the e two group were ig-nificant in Henle 407 cells (450 with the 03K6 groupvel us 239 with the non-03K6 group) HeLa cells (673versus 314) and J774AI cell (685 versu 435) (FiglB) The differences in L2 cells (610 versu 325) weremarginally significant (P lt 010) Although tdh-positiveisolates appeared to have slightly higher cytotoxicity thantdh-negative i olates (Fig IC) the differences were signif-icant only for the Caco-2 cell line
Our findings from the LDH as ays support conclusionfrom previou tudies which suggested that the presenceof tdh alone does not fully correlate with cytotoxicity fluidaccumulation and other virulence measurements (9 2335) therefore tdh alone i not sufficient as a predictor forV parahaemotyticus pathogenicity Our re ult further ug-ge t that the apparent enhanced virulence of V parahae-molyticus 03K6 strain could be a sociated with the en-hanced cytotoxic effects that these cells have on host cell When cla ified into group by ource clinical isolates hadhigher relative cytotoxicity than did food isolates in threeho t cell lines In its current format however this LDHassay cannot reliably discriminate between clinical andnonclinical i olates at the strain level because the relativecytotoxicity percentage in these two group of isolatesoverlap Nevertheless with further development a ti ueculture-ba ed assay may be a useful approach for the de-tection of pathogenic trains becau e the outcome of suchan assay will be more dependent on the ability of a trainto cau e di ease than on the pre ence or absence of one ormore specific virulence determinant
CHO elongation factor is not useful for evaluationof V parahaemolyticus pathogenicity Although V para-haemolyticus i olates clearly differed in their cytotoxic ef-fects on host cells the organism does not appear to possessa CHO elongation factor under the conditions tested NineV parahaemolyticus isolates (Table 1) were creened forthe presence of the CHO elongation factor but none werepositive Thus the CHO elongation assay is not useful forcharacterization of V parahaemolyticus pathogenicity
Development of a tissue culture-based assay to de-tect cytotoxic V parahaemolyticus Becau e clinical iso-lates generally were more cytotoxic than were nonclinicali olates we further tested the efficacy of the LDH assay asa detection tool to screen for the pre ence of V parahae-molyticus strains with pathogenic potential in food sample Our goal with the e experiment wa to as e the abilityof our as ay to detect the presence of pathogenic V para-haemolyticus directly from an oyster tissue matrix Theability to detect potentially dangerou pathogens directlyfrom food samples could ignificantly reduce the analyticaltime needed to make deci ions related to hazards that maybe a ociated with bacterial contamination such as deter-mining the afety of oy ter bed or the need to issue a
J Food Prot Vol 70 o 2
product recal However food rnarrice can interfere withanalytical outcomes For example inhibitory sub tancepresent in food can lead to false-negative results with ornedetection technologic uch as tho e involving PCRs (19)Oyster sample were te ted becau e V parahaemolyticusinfection have been a ociated frequently with can ump-tion of raw or undercooked bivalve shellfi b We selectedL2 cells for a say development because the cytotoxicity dif-ference between clinical and nonclinical isolates was themost pronounced with thi cell line ie 6 (462) of 13clinical isolates but none of the food isolate had gt60relative cytotoxicity The fast-growing L2 cell line al a isea y to maintain V parahaemolyticu FSL- Y 1-024 was se-lected for the development of the screening a say becauseof it relative evere cytotoxic effects in all cell lines (datanot hown)
Approximately 5 X 108 CFU of FSL- YI-024 culturewa used to artificially inoculate the initial diluted oysterhomogenates and then 15 J-LIof the mixture (undiluted ordiluted 10- or 100-fold) were used to infect L2 cell eitherwith or without an enrichment step Results are summarizedin Table 2 A low level of cytotoxicity wa detected inoy tel homogenates and APW (treatments OR alone andOH + APW in Table 2) pre umably because of nonspecificLDH release from the oyster samples Thi nonspecificLDH relea e can tituted the background level for our assayWhen L2 cell were inoculated with 15 X 107 CFU ofVparahaemolyticus FSL- Yl-024 alone a strong response(803 cytotoxicity) wa detected A reduced response(676 cytotoxicity) wa detected with to-fold diluted cul-ture ie 15 X 106 CFU The cytotoxicity of the lOO-folddiluted culture was not significantly different from thebackground Component pre ent in oy ter homogenate including endogenous LDH did not appear to affect detec-tion of V parahaemoiyticus similar cytotoxicity was de-tected for FSL- Y1-024 either with or without oy tel ho-mogenates (treatments OH+Vp and Vp alone)
Although the addition of an enrichment tep to a de-tection assay increases the overall time required for a aycompletion thi step can improve assay sensitivity Wedemon trated previously that a 6-h enrichment period waoptimum for detecting a larger collection of V parahae-molyticus with a PCR method (46) Enrichment at an ap-propriate temperature Can allow injured cells to recover andviable-but-nonculturable (VBNC) cell to re u citate (1321 25) In thi regard an enrichment tep may be partic-ularly useful for V parahaemolyticu because of its abilityto enter into the VBNC tage (2 30) Wong et aI (43)found that VB C cell exhibited delayed mouse lethalityand enteropathogenicity compared with log-pha e cells Inthe natural environment V parahaemoiyticus al 0 may beinjured or starved Wong and Chang (42) found that a hortincubation wa neces ary to enable starved V parahae-molyticus cell to reach the same cytopathogenicity as ex-hibited by the log-pha e cells To balance the benefits ofenrichment with the need for a rapid assay a 6-h enrich-ment period wa selected for the pre ent study FSL- Yl-024 numbers increased by 126 log CFU following this en-richment Stati tical comparison of the treatm nt with an
1 Fo d Pror Vol 70 o 2
enrichment step (OH +Vp+ APW) and the other treatmentswa not conducted because bacterial cell number differedby gt05 log C U
Cytotoxicity mea urements following enrichment maynot be comparable to tho e without enrichment In thistudy V parohaemolyticus cell after the 6-h enrichment
were likely in late Jog to early tationary phase in contra tto the late tationary pha cell u ed in other treatm nt There i increa ing evidenc that virulen e of bacterialpathogen u h a Listeria monocytogene (15) and Strep-tococcus pneumoniae (17) is influenced by environmentaltr e and growth pha e Thus it i also pos ible that V
parahaemolyticu cytopath genicity may be influenced bygrowth phase Further tudy will b neces ary to asses theffect of growth pha on V parahaemolyticus virulencecharacteri tic
In its current format our LDH a say can differ ntiateclinical V parahaemolyticus strains only at the populationlevel uture a ay optimization also must tak into on id-eration the pre nee of other microfl ra in a typical t tmatrix Improved n itivity and pecificity will be neces-ary for application of thi approach a a routin detection
procedureonventi nal detection tools designed t e rabli h the
pre ence of ne genetic determinant or phenotypi char-acteri tic in a given i olate are likely to b of only limitedutility for det cting emerging path genic V parahaemoly-ticus train In th pre ent rudy we amined the utilityof a ti ue culture-based as ay for d velopment of detecti nmethod for pathogenic V parahaemolyticu train A agroup clinical i olate are more cytotoxic than nonclinieali alate Ba ed on LDH relea e re ult -106 cells of cy-totoxic V parahaemolyticus were detect d An ptimizedenrichment tep might further enhance the en itivity of thiassay Further insight into factor contributing to V para-haemolytieus pathogen si will enable development of newdetection method with increased sen itivity and discrimi-natory capabil itie
AK OWLEDGME TWe are grateful to Angelo DePa la and Charle Kay ner (US Food
and Drug Administrati n) r providing the V parahaemolyri us isolatcsu ed in this ludy We al 0 thank Helene Marqui and Edward Dubovi(Cornell University Ithaca Y) for as i tance with cell lines ThL workwa supported by a gram from tJle New York ea Gram pI gram (RI HH-J3) 10 K J Boor and M Wiedmann
REF RE EI Baba K H hirai A Terai K Kumagai Y Takeda and M Nish-
ibuclti 1991 Similarity of the rdh gene-bearing pIa mids of Vibrioeholerae noo-Ol and Vibrio parthaemolyriclls Microb PatllOg 1061-70
2 Bate T C and J D Oliver 2004 The viable but nonculturabletate of Kanagawa positive and negati e train of Vibrio parahaemiddot
l1lolytieus J Microbial 44274-793 Chi u C S Y Hsu I hiu T K Wang and C S Chao
2000 Vibrio parahaemolyriws serovar 03K6 as cause of unusuaJlyhigh incidence offood-bome di ease outbreak in Taiwan from 1996to 1999 J Clin Microbial 384621-4625
4 ChOWdhuryA M [ hiba hi V D Thiem D T Tuy t T V TungB T Chien L von eidlein D G Canh J Clemen D D Trachand M i hibuchi 2004 Emergence and erovar tran ition of Vibrio
CYTOTOXICIT1ES OF V PARAHAEMOLYTICUS 353
parahaemolyticus pandemic strains isolated during a diarrhea out-break in Vietnam between 1997 and 1999 Microbial lmmunol 48319-327
5 howdhury N R S Chakraborty T Ramamunhy M ishibuchiS Yama aki Y Takeda and G B Nair 2000 Molecular evidenceof clonal Vibrio parahaemolvti IS pandemic strains Emerg InfectDis 6631-636
6 Daniels N A B Ray A Easton Marano E Kahn A LMcShan II L Del Ro ario T Baldwin M A Kingsley N D PuhrJ G Wells and E J Angulo 2000 Emergence of a new Vibriopurahaemotyticus serotype in raw oy ters a prevenri n quandaryJAMA 2841541-1545
7 Guerrant R L L L Brunton T C chnaitrnan L I Rebhun andA G Gilman 1974 Cyclic adenosine rnonophosphate and alterationof hinese hamster ovary cell morphology a rapid sen itive in vitroas ay for the enterotoxins of Vibrio cholerae and Escherichia coliInfect lrnmun JO320-327
8 Hackney R E G Kleeman B Ray and M L Speck 1980Adherence as a method of differentiating virulent and avirulenttrains of Vibrio parahaemoiyticus Appl Environ Microbial 40
652-6589 Hoa hi K K Ogata H Taniguchi H Yamashita K Tsuji Y Mi-
zuguchi and Ohtorno 1990 Pathogenesis of Vibrio parahae-molyticus intraperitoneal and oroga tric challenge experiment inmice Microbiol Immunol 34355-366
10 Hondo l G 101 Minemal u Ikeda Asano M IshibashiY Kino hita ishibuchi T Honda and T Miwaiani 1987 Gatr emeriti due to Kanagawa negative Vibrio parahaemolyticus UII-
cet i33 1-332J J lijima Y H Yamada and hinoda 19 I Adherence of Vibrio
parahaemolyticu and it relation to pathogenicity Call J Micro-bioi 271252-1259
12 I berg R D L Vo rhi and Falkow 1987 Identification ofinvasin a protein thaL allows enteric bacteria to penetrate culturedmammalian cell Cell 50769-778
13 Johnston M D and M H Brown 2002 An investigation into thechanged physiological state of Vibrio bacteria as a survival mechamiddotni m in rc p n e to cold temperature and studie on their sen itivilyto heating and freezing J Appl Mi robiol 92 1066-1077
14 Kay ner C A C Abeyta Jr P A Tro t J H Wetherington KC Jinneman W E Hill and M M Wekell 1994 Urea hydrolysiscan predict the potential pathogenicity of Vibrio parahaemolyticlsstrain iolated in the Pacific Norlhwe t Appl Environ Microbiol603020-3022
15 Kim H H Marquis and K 1 Boor 2005 (18 contribute to Lisreriamono yrogenes inva ion by controlling expre sion of inlA and inlBMicmbiology 151 32]5-3222
16 Ki hishita M N Matsuoka K Kumagai S Yamasaki Y Takedaand M Ni hibuchi 1992 Sequence variation in the thermostabledirect hemoly in-related hemolysin (rrh) gene of Vibrio paralwe-molyricus Appl Environ Microbiol 582449-2457
17 Ko K S S Park W Oh J Y uh T Oh S Ahn J Chun andJ H Song 2006 omparative analysi of growth-phase dependentgene expre sion in virulent and avirulent Srreptocoecls pllelmoniaeu ing a high-den ity D A microarray Mol Cells 2182-88
18 Kothary M E R Claverie M D Milioti J M Madden and H Richard on J995 Purification and characLerizatjon of a Chineseham ter ovary cell elongation factor of Vibrio hollisae hrfeel 1m-IIlUII 632418-2423
19 Lantz P G W Abu almiddotSoud R Knuts on B HahnmiddotHagerdal andP Rad trom 2000 Biotechnical u of polymerase chain reactionfor microbiological analy i of biological sample BioteclllJol AmllRev 587-130
20 Laohaprertthi an v A Chowdhury U Kongmuang S Kalnaumiddotwakul M Ishibashi C Matsumoto and M i hibuchi 2003 Prev-alence and erodiversity of the pandemic clone among the clinicaltrains of Vibrio parahaemolyticus isolated in southern Thailand Ep-
idemiol Illfecr 130395-40621 Liao C H and W F Fen 2005 Re uscitation of acid-injured Sal-
~
-
354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909

352 YEUNG ET AL
in the L2 Henle 407 and Caco-2 cell lines Cytotoxicityof the V parahaemolyticus 03K6 and non-03K6 i olatealso was compared Because all 03K6 isolates in our col-lection were from clinical sources only clinical non-03K6i olate were compared with clinical 03K6 isolates Cy-totoxicity differences between the e two group were ig-nificant in Henle 407 cells (450 with the 03K6 groupvel us 239 with the non-03K6 group) HeLa cells (673versus 314) and J774AI cell (685 versu 435) (FiglB) The differences in L2 cells (610 versu 325) weremarginally significant (P lt 010) Although tdh-positiveisolates appeared to have slightly higher cytotoxicity thantdh-negative i olates (Fig IC) the differences were signif-icant only for the Caco-2 cell line
Our findings from the LDH as ays support conclusionfrom previou tudies which suggested that the presenceof tdh alone does not fully correlate with cytotoxicity fluidaccumulation and other virulence measurements (9 2335) therefore tdh alone i not sufficient as a predictor forV parahaemotyticus pathogenicity Our re ult further ug-ge t that the apparent enhanced virulence of V parahae-molyticus 03K6 strain could be a sociated with the en-hanced cytotoxic effects that these cells have on host cell When cla ified into group by ource clinical isolates hadhigher relative cytotoxicity than did food isolates in threeho t cell lines In its current format however this LDHassay cannot reliably discriminate between clinical andnonclinical i olates at the strain level because the relativecytotoxicity percentage in these two group of isolatesoverlap Nevertheless with further development a ti ueculture-ba ed assay may be a useful approach for the de-tection of pathogenic trains becau e the outcome of suchan assay will be more dependent on the ability of a trainto cau e di ease than on the pre ence or absence of one ormore specific virulence determinant
CHO elongation factor is not useful for evaluationof V parahaemolyticus pathogenicity Although V para-haemolyticus i olates clearly differed in their cytotoxic ef-fects on host cells the organism does not appear to possessa CHO elongation factor under the conditions tested NineV parahaemolyticus isolates (Table 1) were creened forthe presence of the CHO elongation factor but none werepositive Thus the CHO elongation assay is not useful forcharacterization of V parahaemolyticus pathogenicity
Development of a tissue culture-based assay to de-tect cytotoxic V parahaemolyticus Becau e clinical iso-lates generally were more cytotoxic than were nonclinicali olates we further tested the efficacy of the LDH assay asa detection tool to screen for the pre ence of V parahae-molyticus strains with pathogenic potential in food sample Our goal with the e experiment wa to as e the abilityof our as ay to detect the presence of pathogenic V para-haemolyticus directly from an oyster tissue matrix Theability to detect potentially dangerou pathogens directlyfrom food samples could ignificantly reduce the analyticaltime needed to make deci ions related to hazards that maybe a ociated with bacterial contamination such as deter-mining the afety of oy ter bed or the need to issue a
J Food Prot Vol 70 o 2
product recal However food rnarrice can interfere withanalytical outcomes For example inhibitory sub tancepresent in food can lead to false-negative results with ornedetection technologic uch as tho e involving PCRs (19)Oyster sample were te ted becau e V parahaemolyticusinfection have been a ociated frequently with can ump-tion of raw or undercooked bivalve shellfi b We selectedL2 cells for a say development because the cytotoxicity dif-ference between clinical and nonclinical isolates was themost pronounced with thi cell line ie 6 (462) of 13clinical isolates but none of the food isolate had gt60relative cytotoxicity The fast-growing L2 cell line al a isea y to maintain V parahaemolyticu FSL- Y 1-024 was se-lected for the development of the screening a say becauseof it relative evere cytotoxic effects in all cell lines (datanot hown)
Approximately 5 X 108 CFU of FSL- YI-024 culturewa used to artificially inoculate the initial diluted oysterhomogenates and then 15 J-LIof the mixture (undiluted ordiluted 10- or 100-fold) were used to infect L2 cell eitherwith or without an enrichment step Results are summarizedin Table 2 A low level of cytotoxicity wa detected inoy tel homogenates and APW (treatments OR alone andOH + APW in Table 2) pre umably because of nonspecificLDH release from the oyster samples Thi nonspecificLDH relea e can tituted the background level for our assayWhen L2 cell were inoculated with 15 X 107 CFU ofVparahaemolyticus FSL- Yl-024 alone a strong response(803 cytotoxicity) wa detected A reduced response(676 cytotoxicity) wa detected with to-fold diluted cul-ture ie 15 X 106 CFU The cytotoxicity of the lOO-folddiluted culture was not significantly different from thebackground Component pre ent in oy ter homogenate including endogenous LDH did not appear to affect detec-tion of V parahaemoiyticus similar cytotoxicity was de-tected for FSL- Y1-024 either with or without oy tel ho-mogenates (treatments OH+Vp and Vp alone)
Although the addition of an enrichment tep to a de-tection assay increases the overall time required for a aycompletion thi step can improve assay sensitivity Wedemon trated previously that a 6-h enrichment period waoptimum for detecting a larger collection of V parahae-molyticus with a PCR method (46) Enrichment at an ap-propriate temperature Can allow injured cells to recover andviable-but-nonculturable (VBNC) cell to re u citate (1321 25) In thi regard an enrichment tep may be partic-ularly useful for V parahaemolyticu because of its abilityto enter into the VBNC tage (2 30) Wong et aI (43)found that VB C cell exhibited delayed mouse lethalityand enteropathogenicity compared with log-pha e cells Inthe natural environment V parahaemoiyticus al 0 may beinjured or starved Wong and Chang (42) found that a hortincubation wa neces ary to enable starved V parahae-molyticus cell to reach the same cytopathogenicity as ex-hibited by the log-pha e cells To balance the benefits ofenrichment with the need for a rapid assay a 6-h enrich-ment period wa selected for the pre ent study FSL- Yl-024 numbers increased by 126 log CFU following this en-richment Stati tical comparison of the treatm nt with an
1 Fo d Pror Vol 70 o 2
enrichment step (OH +Vp+ APW) and the other treatmentswa not conducted because bacterial cell number differedby gt05 log C U
Cytotoxicity mea urements following enrichment maynot be comparable to tho e without enrichment In thistudy V parohaemolyticus cell after the 6-h enrichment
were likely in late Jog to early tationary phase in contra tto the late tationary pha cell u ed in other treatm nt There i increa ing evidenc that virulen e of bacterialpathogen u h a Listeria monocytogene (15) and Strep-tococcus pneumoniae (17) is influenced by environmentaltr e and growth pha e Thus it i also pos ible that V
parahaemolyticu cytopath genicity may be influenced bygrowth phase Further tudy will b neces ary to asses theffect of growth pha on V parahaemolyticus virulencecharacteri tic
In its current format our LDH a say can differ ntiateclinical V parahaemolyticus strains only at the populationlevel uture a ay optimization also must tak into on id-eration the pre nee of other microfl ra in a typical t tmatrix Improved n itivity and pecificity will be neces-ary for application of thi approach a a routin detection
procedureonventi nal detection tools designed t e rabli h the
pre ence of ne genetic determinant or phenotypi char-acteri tic in a given i olate are likely to b of only limitedutility for det cting emerging path genic V parahaemoly-ticus train In th pre ent rudy we amined the utilityof a ti ue culture-based as ay for d velopment of detecti nmethod for pathogenic V parahaemolyticu train A agroup clinical i olate are more cytotoxic than nonclinieali alate Ba ed on LDH relea e re ult -106 cells of cy-totoxic V parahaemolyticus were detect d An ptimizedenrichment tep might further enhance the en itivity of thiassay Further insight into factor contributing to V para-haemolytieus pathogen si will enable development of newdetection method with increased sen itivity and discrimi-natory capabil itie
AK OWLEDGME TWe are grateful to Angelo DePa la and Charle Kay ner (US Food
and Drug Administrati n) r providing the V parahaemolyri us isolatcsu ed in this ludy We al 0 thank Helene Marqui and Edward Dubovi(Cornell University Ithaca Y) for as i tance with cell lines ThL workwa supported by a gram from tJle New York ea Gram pI gram (RI HH-J3) 10 K J Boor and M Wiedmann
REF RE EI Baba K H hirai A Terai K Kumagai Y Takeda and M Nish-
ibuclti 1991 Similarity of the rdh gene-bearing pIa mids of Vibrioeholerae noo-Ol and Vibrio parthaemolyriclls Microb PatllOg 1061-70
2 Bate T C and J D Oliver 2004 The viable but nonculturabletate of Kanagawa positive and negati e train of Vibrio parahaemiddot
l1lolytieus J Microbial 44274-793 Chi u C S Y Hsu I hiu T K Wang and C S Chao
2000 Vibrio parahaemolyriws serovar 03K6 as cause of unusuaJlyhigh incidence offood-bome di ease outbreak in Taiwan from 1996to 1999 J Clin Microbial 384621-4625
4 ChOWdhuryA M [ hiba hi V D Thiem D T Tuy t T V TungB T Chien L von eidlein D G Canh J Clemen D D Trachand M i hibuchi 2004 Emergence and erovar tran ition of Vibrio
CYTOTOXICIT1ES OF V PARAHAEMOLYTICUS 353
parahaemolyticus pandemic strains isolated during a diarrhea out-break in Vietnam between 1997 and 1999 Microbial lmmunol 48319-327
5 howdhury N R S Chakraborty T Ramamunhy M ishibuchiS Yama aki Y Takeda and G B Nair 2000 Molecular evidenceof clonal Vibrio parahaemolvti IS pandemic strains Emerg InfectDis 6631-636
6 Daniels N A B Ray A Easton Marano E Kahn A LMcShan II L Del Ro ario T Baldwin M A Kingsley N D PuhrJ G Wells and E J Angulo 2000 Emergence of a new Vibriopurahaemotyticus serotype in raw oy ters a prevenri n quandaryJAMA 2841541-1545
7 Guerrant R L L L Brunton T C chnaitrnan L I Rebhun andA G Gilman 1974 Cyclic adenosine rnonophosphate and alterationof hinese hamster ovary cell morphology a rapid sen itive in vitroas ay for the enterotoxins of Vibrio cholerae and Escherichia coliInfect lrnmun JO320-327
8 Hackney R E G Kleeman B Ray and M L Speck 1980Adherence as a method of differentiating virulent and avirulenttrains of Vibrio parahaemoiyticus Appl Environ Microbial 40
652-6589 Hoa hi K K Ogata H Taniguchi H Yamashita K Tsuji Y Mi-
zuguchi and Ohtorno 1990 Pathogenesis of Vibrio parahae-molyticus intraperitoneal and oroga tric challenge experiment inmice Microbiol Immunol 34355-366
10 Hondo l G 101 Minemal u Ikeda Asano M IshibashiY Kino hita ishibuchi T Honda and T Miwaiani 1987 Gatr emeriti due to Kanagawa negative Vibrio parahaemolyticus UII-
cet i33 1-332J J lijima Y H Yamada and hinoda 19 I Adherence of Vibrio
parahaemolyticu and it relation to pathogenicity Call J Micro-bioi 271252-1259
12 I berg R D L Vo rhi and Falkow 1987 Identification ofinvasin a protein thaL allows enteric bacteria to penetrate culturedmammalian cell Cell 50769-778
13 Johnston M D and M H Brown 2002 An investigation into thechanged physiological state of Vibrio bacteria as a survival mechamiddotni m in rc p n e to cold temperature and studie on their sen itivilyto heating and freezing J Appl Mi robiol 92 1066-1077
14 Kay ner C A C Abeyta Jr P A Tro t J H Wetherington KC Jinneman W E Hill and M M Wekell 1994 Urea hydrolysiscan predict the potential pathogenicity of Vibrio parahaemolyticlsstrain iolated in the Pacific Norlhwe t Appl Environ Microbiol603020-3022
15 Kim H H Marquis and K 1 Boor 2005 (18 contribute to Lisreriamono yrogenes inva ion by controlling expre sion of inlA and inlBMicmbiology 151 32]5-3222
16 Ki hishita M N Matsuoka K Kumagai S Yamasaki Y Takedaand M Ni hibuchi 1992 Sequence variation in the thermostabledirect hemoly in-related hemolysin (rrh) gene of Vibrio paralwe-molyricus Appl Environ Microbiol 582449-2457
17 Ko K S S Park W Oh J Y uh T Oh S Ahn J Chun andJ H Song 2006 omparative analysi of growth-phase dependentgene expre sion in virulent and avirulent Srreptocoecls pllelmoniaeu ing a high-den ity D A microarray Mol Cells 2182-88
18 Kothary M E R Claverie M D Milioti J M Madden and H Richard on J995 Purification and characLerizatjon of a Chineseham ter ovary cell elongation factor of Vibrio hollisae hrfeel 1m-IIlUII 632418-2423
19 Lantz P G W Abu almiddotSoud R Knuts on B HahnmiddotHagerdal andP Rad trom 2000 Biotechnical u of polymerase chain reactionfor microbiological analy i of biological sample BioteclllJol AmllRev 587-130
20 Laohaprertthi an v A Chowdhury U Kongmuang S Kalnaumiddotwakul M Ishibashi C Matsumoto and M i hibuchi 2003 Prev-alence and erodiversity of the pandemic clone among the clinicaltrains of Vibrio parahaemolyticus isolated in southern Thailand Ep-
idemiol Illfecr 130395-40621 Liao C H and W F Fen 2005 Re uscitation of acid-injured Sal-
~
-
354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909

1 Fo d Pror Vol 70 o 2
enrichment step (OH +Vp+ APW) and the other treatmentswa not conducted because bacterial cell number differedby gt05 log C U
Cytotoxicity mea urements following enrichment maynot be comparable to tho e without enrichment In thistudy V parohaemolyticus cell after the 6-h enrichment
were likely in late Jog to early tationary phase in contra tto the late tationary pha cell u ed in other treatm nt There i increa ing evidenc that virulen e of bacterialpathogen u h a Listeria monocytogene (15) and Strep-tococcus pneumoniae (17) is influenced by environmentaltr e and growth pha e Thus it i also pos ible that V
parahaemolyticu cytopath genicity may be influenced bygrowth phase Further tudy will b neces ary to asses theffect of growth pha on V parahaemolyticus virulencecharacteri tic
In its current format our LDH a say can differ ntiateclinical V parahaemolyticus strains only at the populationlevel uture a ay optimization also must tak into on id-eration the pre nee of other microfl ra in a typical t tmatrix Improved n itivity and pecificity will be neces-ary for application of thi approach a a routin detection
procedureonventi nal detection tools designed t e rabli h the
pre ence of ne genetic determinant or phenotypi char-acteri tic in a given i olate are likely to b of only limitedutility for det cting emerging path genic V parahaemoly-ticus train In th pre ent rudy we amined the utilityof a ti ue culture-based as ay for d velopment of detecti nmethod for pathogenic V parahaemolyticu train A agroup clinical i olate are more cytotoxic than nonclinieali alate Ba ed on LDH relea e re ult -106 cells of cy-totoxic V parahaemolyticus were detect d An ptimizedenrichment tep might further enhance the en itivity of thiassay Further insight into factor contributing to V para-haemolytieus pathogen si will enable development of newdetection method with increased sen itivity and discrimi-natory capabil itie
AK OWLEDGME TWe are grateful to Angelo DePa la and Charle Kay ner (US Food
and Drug Administrati n) r providing the V parahaemolyri us isolatcsu ed in this ludy We al 0 thank Helene Marqui and Edward Dubovi(Cornell University Ithaca Y) for as i tance with cell lines ThL workwa supported by a gram from tJle New York ea Gram pI gram (RI HH-J3) 10 K J Boor and M Wiedmann
REF RE EI Baba K H hirai A Terai K Kumagai Y Takeda and M Nish-
ibuclti 1991 Similarity of the rdh gene-bearing pIa mids of Vibrioeholerae noo-Ol and Vibrio parthaemolyriclls Microb PatllOg 1061-70
2 Bate T C and J D Oliver 2004 The viable but nonculturabletate of Kanagawa positive and negati e train of Vibrio parahaemiddot
l1lolytieus J Microbial 44274-793 Chi u C S Y Hsu I hiu T K Wang and C S Chao
2000 Vibrio parahaemolyriws serovar 03K6 as cause of unusuaJlyhigh incidence offood-bome di ease outbreak in Taiwan from 1996to 1999 J Clin Microbial 384621-4625
4 ChOWdhuryA M [ hiba hi V D Thiem D T Tuy t T V TungB T Chien L von eidlein D G Canh J Clemen D D Trachand M i hibuchi 2004 Emergence and erovar tran ition of Vibrio
CYTOTOXICIT1ES OF V PARAHAEMOLYTICUS 353
parahaemolyticus pandemic strains isolated during a diarrhea out-break in Vietnam between 1997 and 1999 Microbial lmmunol 48319-327
5 howdhury N R S Chakraborty T Ramamunhy M ishibuchiS Yama aki Y Takeda and G B Nair 2000 Molecular evidenceof clonal Vibrio parahaemolvti IS pandemic strains Emerg InfectDis 6631-636
6 Daniels N A B Ray A Easton Marano E Kahn A LMcShan II L Del Ro ario T Baldwin M A Kingsley N D PuhrJ G Wells and E J Angulo 2000 Emergence of a new Vibriopurahaemotyticus serotype in raw oy ters a prevenri n quandaryJAMA 2841541-1545
7 Guerrant R L L L Brunton T C chnaitrnan L I Rebhun andA G Gilman 1974 Cyclic adenosine rnonophosphate and alterationof hinese hamster ovary cell morphology a rapid sen itive in vitroas ay for the enterotoxins of Vibrio cholerae and Escherichia coliInfect lrnmun JO320-327
8 Hackney R E G Kleeman B Ray and M L Speck 1980Adherence as a method of differentiating virulent and avirulenttrains of Vibrio parahaemoiyticus Appl Environ Microbial 40
652-6589 Hoa hi K K Ogata H Taniguchi H Yamashita K Tsuji Y Mi-
zuguchi and Ohtorno 1990 Pathogenesis of Vibrio parahae-molyticus intraperitoneal and oroga tric challenge experiment inmice Microbiol Immunol 34355-366
10 Hondo l G 101 Minemal u Ikeda Asano M IshibashiY Kino hita ishibuchi T Honda and T Miwaiani 1987 Gatr emeriti due to Kanagawa negative Vibrio parahaemolyticus UII-
cet i33 1-332J J lijima Y H Yamada and hinoda 19 I Adherence of Vibrio
parahaemolyticu and it relation to pathogenicity Call J Micro-bioi 271252-1259
12 I berg R D L Vo rhi and Falkow 1987 Identification ofinvasin a protein thaL allows enteric bacteria to penetrate culturedmammalian cell Cell 50769-778
13 Johnston M D and M H Brown 2002 An investigation into thechanged physiological state of Vibrio bacteria as a survival mechamiddotni m in rc p n e to cold temperature and studie on their sen itivilyto heating and freezing J Appl Mi robiol 92 1066-1077
14 Kay ner C A C Abeyta Jr P A Tro t J H Wetherington KC Jinneman W E Hill and M M Wekell 1994 Urea hydrolysiscan predict the potential pathogenicity of Vibrio parahaemolyticlsstrain iolated in the Pacific Norlhwe t Appl Environ Microbiol603020-3022
15 Kim H H Marquis and K 1 Boor 2005 (18 contribute to Lisreriamono yrogenes inva ion by controlling expre sion of inlA and inlBMicmbiology 151 32]5-3222
16 Ki hishita M N Matsuoka K Kumagai S Yamasaki Y Takedaand M Ni hibuchi 1992 Sequence variation in the thermostabledirect hemoly in-related hemolysin (rrh) gene of Vibrio paralwe-molyricus Appl Environ Microbiol 582449-2457
17 Ko K S S Park W Oh J Y uh T Oh S Ahn J Chun andJ H Song 2006 omparative analysi of growth-phase dependentgene expre sion in virulent and avirulent Srreptocoecls pllelmoniaeu ing a high-den ity D A microarray Mol Cells 2182-88
18 Kothary M E R Claverie M D Milioti J M Madden and H Richard on J995 Purification and characLerizatjon of a Chineseham ter ovary cell elongation factor of Vibrio hollisae hrfeel 1m-IIlUII 632418-2423
19 Lantz P G W Abu almiddotSoud R Knuts on B HahnmiddotHagerdal andP Rad trom 2000 Biotechnical u of polymerase chain reactionfor microbiological analy i of biological sample BioteclllJol AmllRev 587-130
20 Laohaprertthi an v A Chowdhury U Kongmuang S Kalnaumiddotwakul M Ishibashi C Matsumoto and M i hibuchi 2003 Prev-alence and erodiversity of the pandemic clone among the clinicaltrains of Vibrio parahaemolyticus isolated in southern Thailand Ep-
idemiol Illfecr 130395-40621 Liao C H and W F Fen 2005 Re uscitation of acid-injured Sal-
~
-
354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909

354 YEUNG ET AL
monela in enrichment broth in apple juice and on the surfaces oftre h-cut cucumber and apple Lett Appl Microbial 4] 487-492
22 Lindqvist R 1999 Detection of Shigella spp in food with a ne tedPCR method-sensitivity and performance compared with a con-ventional culture method 1 Appl Microbiol 86971-978
23 Lynch T S Livingstone E Buenaventura E Lutter J FedwickA G Buret D Graham and R Devinney 2005 Vibrio parahae-molyucus disruption of epithelial cell tight junction occurs indepen-dently of toxin production infect Immun 73 1275-1283
24 Makino K K O hirna K Kurokawa K Yokoyama T Uda KTagornori Y Iijima M Najima M Nakano A Yamashita Y Ku-bota S Kimura T Yasunaga T Honda H Shinagawa M Hattoriand T lida 2003 Genome sequence of Vibrio parahaemolyticus apathogenic mechani m distinct from that of V cholerae Lancet 361 743-749
25 Ma-Lin C F A and L R Beuchat 1980 Recovery ofchill-stressedVibrio parahaemolyticus from oysters with enrichment broths sup-plemented with magnesium and iron salt Appl Environ Microbial
39 179-18526 Matsumoto c J Okuda M Ishibashi M Iwanaga P Garg T
RammamurthyH C Wong A Depaola Y B Kim M J Albertand M Nishibuchi 2000 Pandemic spread of an 03K6 clone ofVibrio parahaemolyticus and emergence of related strains evidencedby arbitrarily primed PCR and loxRS sequence analyses J ClinMicrobial 38578-585
27 McCardell B A M H Kothary R H Hall and V Sathyamoorthy2000 Identification of a CHO cell-elongating factor produced byVibrio cholerae OJ Microb Pathog 29 1-8
28 Merrell B R R l Walker and S W Joseph 1984 ln vitro and invivo pathologic effects of Vibrio parahaemolyticus on human epi-thelial cells Can J Microbiol 3038J-388
29 Miyamoto Y T Kato Y Obara S Akiyama K Takizawa and SYamai 1969 In vitro hemolytic characteristics of Vibrio parahae-molyticus its close correlation with human pathogenicity J Bacter-ial1001147-1149
30 Mizunoe Y S N Wai T Ishikawa A Takade and S Yoshida2000 Resuscitation of viable but nonculturable cells of Vibrio para-haemolyticus induced at low temperature under starvation FEMSMicrobiol Lett 186 115-120
31 Nachlas M M S L Margulies 1 D Goldberg and A M Selig-man J960 The determination of lactic dehydrogenase with a tet-razolium salt Anal Biochel 10317-326
32 Nishibuchi M A Fasano R G Russell and 1 B Kaper 1992Enterotoxigenicity of Vibrio para-JaemolyticlIs with and withoutgenes encoding thermostable direct hemolysin Infecl Immlln 603539-3545
33 isbibuchi M and 1 B Kaper 1990 Duplication and variation ofthe thermostable direct haemolysin (tch) gene in Vibrio parahaemiddotmolyticlIs Mol Microbiol 487-99
34 Okuda J M Ishibashi E Hayakawa T Nishino Y Takeda A KMukhopadhyay S Garg K Bhattacharya G B air and M
J Food Prot Vol 70 No2
Nishibuchi ]997 Emergence of a unique 03K6 clone of Vibrioparahaemolyticus in Calcutta India and isolation of strain from the
arne clonal group from Southeast Asian travelers arriving in Japan1 CUn Microbial 353150-3155
35 Park K-S T Ono M Rokuda M-H lang T [ida and T Honda2004 Cytotoxicity and enterotoxicity of the thermostable direct he-molysin-deletion mutant of Vibrio parahaemolyticus Microbiallmmunol 48313-318
36 Rumeu M T M A Suarez S Morales and R Rotger 1997 En-terotoxin and cytotoxin production by Salmonella Enteritidis trainisolated from gastroenteritis outbreaks 1 Appl Microbial 8219-31
37 Shirai H H Ito T Hirayama Y akarnoto akabayashi KKumagai Y Takeda and M Nishibuchi 1990 Molecular epide-miologic evidence for association of thermo table direct hemolysin(TDH) and TDHmiddotrelated hemoly in of Vibrio parahaemolyticus withgastroenteriti Inject Immun 583568-3573
38 Sochard M R and R R Colwell 1977 Toxin isolation from aKanagawa-phenornenon negative strain of Vibrio parahaemolyticusMicrobial lmmunol 21 243-254
39 US Food and Drug Admini tration 2001 Yersinia enterocoliticaand Yersinia p eudotuberculosis III Bacteriological anaJytical man-ual online Available at httpwwwcfsanfdagov-ebarnbam-8html Po ted January 2001
40 US Food and Drug Administration 2004 Bacteriological analyticalmanual online Available at htlplwwwcfsanfdagov-ebamlbam-9html Posted May 2004
41 Venkateswaran K H Horiuchi H akano H Matsuda and HHa hirnoto 1991 Vibrio virulence factors and the quantitative anal-ysis of cytotoxicity elaborated by environmental i olates Cytobios66 143-15 I
42 Wong H-C and c- Chang 2005 Hydrophobicity cell adher-ence cytotoxicity and enterotoxigenicity of starved Vibrio parawe-molytlcus 1 Food Prot 68 154-156
43 Wong H-C c-T Shen C-N Chang Y-S Lee and J Oliver2004 Biochemical and virulence characterization of viable but non-culturable cells of Vibrio parahaemolyticus J Food Prot 672430-2435
44 Yamamoto S N Okujo S Miyoshi S Shinoda and S Narimatsu1999 Siderophore production of Vibrio parahaemolyticlls strainfrom different source Microbiol ImmlllwL 43909-912
45 Yeung P S M and K J Boor 2004 EpidemiOlogy pathogenesisand prevention of foodbome Vibrio parahaemolyticus infectionsFoodborne Pathog Dis 174-88
46 Yeung P S M A DePaola C A Kaysner and K J Boor 2003A PCR assay for specific detection of the pandemic Vibrio para-haemolytiClls 03K6 clone from shellfish J Food Sci 681459-1466
47 Yeung P S M M C Hayes A DePaola C A Kaysner L Korn-stein and K J Boor 2002 Comparative phenotypic molecular andvirulence characterization of Vibrio parahemolyticus 03K6 i olate Appl Environ Microbial 68 290]-2909