estudio de la variaciÓn somaclonal en la
TRANSCRIPT

CENTRO DE INVESTIGACIÓN Y ASISTENCIA EN TECNOLOGÍA Y DISEÑO DEL ESTADO DE JALISCO, A.C.
ESTUDIO DE LA VARIACIÓN SOMACLONAL EN
LA MORFOGÉNESIS DE Bletia purpurea (Lam.).
TESIS
PARA OBTENER EL GRADO ACADÉMICO DE
MAESTRO EN CIENCIAS DE LA FLORICULTURA
QUE PRESENTA
JULIA ESTHER YAH CHULIM
MÉRIDA, YUCATÁN, JUNIO 2011


A
LIBERACIÓN DEL SÍNODO
Mérida, Yucatán a 28 de junio de 2011
Dr. Rodrigo Barba González Coordinador Académico de la Maestría en Ciencias de la Floricultura Guadalajara, Jalisco
Los que suscriben integrantes del jurado de examen de grado de la estudiante Julia Esther Yah Chulim, una vez leída y revisada la Tesis titulada “ESTUDIO DE LA VARIACIÓN SOMACLONAL EN LA MORFOGÉNESIS DE Bletia purpurea (Lam.)” aceptamos que la referida tesis revisada y corregida sea presentada por la alumna para aspirar al grado de Maestro en Ciencias de la Floricultura durante el examen correspondiente.
Y para que así conste firmamos la presente a los veintiocho días del mes de junio del año 2011.
_______________________________ ___________________________ Dra. Ingrid Mayanín Rodríguez Buenfil Dr. Eduardo Villanueva Couoh
Presidente Secretario
_______________________ __________________________ Dra. Ana Luisa Ramos Díaz Dra. Eunice Gómez Uc
Vocal Vocal
______________________ Dra. Guadalupe López Puc
Vocal

B

C
DEDICATORIAS
A:
Dios
Primeramente, le agradezco permitirme la vida, darme la inteligencia y la bendición de emprender nuevos caminos.
Antonio y Lucía, mis padres
Por la confianza, apoyo y motivación a lo largo de mi vida.
Rosy, Lissie, Nurmy, Mary, Alfredo; mis hermanos y sobrinos Ruby, Samuel, Paola
Por su cariño, apoyo incondicional y confianza que siempre han tenido en mí.
Mis amigos Guadalupe: Pati, Nancy, Kati, Minerva, Zazil, Nory, Josué, Rene, David y Luís
Les agradezco su compañía, motivación y apoyo a lo largo de la carrera.
Mis compañeras: Doris Canul, Lyndia Perera, Adriana González y Paola Balam
Les agradezco su apoyo y amistad incondicional durante el transcurso de la maestría.
Soli Deo Gloria

D

E
AGRADECIMIENTOS
A CONACYT y al CIATEJ por la beca otorgada. Al Fondo mixto Gobierno del Estado de Campeche-CONACYT, por el apoyo
otorgado a través del proyecto “Establecimiento de un banco de germoplasma in vitro
de orquídeas nativas del Estado de Campeche para su uso sustentable” clave 96713.
Al Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco, A.C. Unidad Sureste por facilitar el uso de las instalaciones y equipos para la realización de esta tesis. A la Dra. Guadalupe López Puc por la confianza depositada en mí para la realización de este trabajo. Al Dr. Rodrigo Barba González por formar parte importante de la Maestría en Ciencias de la Floricultura. A la Dra. Ingrid Rodríguez Buenfil le agradezco su tiempo, guía y paciencia en el análisis de datos. A la Dra. Lourdes Reyes Escogido por su tiempo, ayuda, paciencia y delicadeza con la que me enseño y guió, durante mi trabajo en el laboratorio y en la redacción del escrito. Al Sínodo: Dra. Eunice Gómez Uc, Dra. Ingrid Mayanín Rodríguez Buenfil, Dra. Ana Luisa Ramos Díaz, Dra. Julia Cano Sosa y Dr. Eduardo Villanueva Couoh, por su tiempo, recomendaciones y comentarios, que fueron de gran ayuda para enriquecer este trabajo. A mis profesores: Dra. Rosa María Escobedo, Dra. Ángeles Sánchez, Dr. Luis Pinzón, Dr. Zahaed Evangelista, Dr. Ernesto Prado, Dr. Alberto Uc, Dr. Gabriel Rincón, que con su experiencia y entrega me enseñaron, guiaron y motivaron a seguir adelante. A todas aquellas personas que de una u otra manera apoyaron e hicieron que este trabajo sea una realidad, mis padres, hermanos, amigos, Dra. Edith González Mondragón, QI. Liliana Pérez, Ruth Valencia.

F

i
CONTENIDO
Página
Lista de tablas. vii
Lista de figuras. xi Lista de abreviaturas. xv RESUMEN. 1 ABSTRACT. 3 I INTRODUCCIÓN. 5 II ANTECEDENTES. 9 2.1 Características generales de las orquídeas. 11 2.2 Importancia comercial de las orquídeas. 12 2.3 Bletia purpurea (Lam.) DC. 13 2.3.1 Descripción botánica. 13 2.3.2 Distribución geográfica de la especie. 14 2.4 Propagación vegetativa de las orquídeas. 15 2.5 Cultivo de tejidos vegetales. 16 2.5.1 Morfogénesis in vitro. 17 2.5.1.1 Embriogénesis somática. 18 2.5.1.2 Organogénesis. 18 2.5.2 Función de los reguladores de crecimiento vegetal en el
desarrollo in vitro. 18
2.5.2.1 Auxinas. 19 2.5.2.2 Citocininas. 20 2.5.3 Cultivo de tejidos vegetales en orquídeas. 21 2.6 Metodología de superficie de respuesta (MSR). 23 2.7 Variación somaclonal. 25 2.7.1 Ventajas y desventajas de la variación somaclonal. 25 2.7.2 Detección de la variación somaclonal. 26 2.7.2.1 Amplificación aleatoria del ADN polimórfico (RAPD). 28 2.7.2.2 Análisis genómico utilizando marcadores moleculares
RAPDs 30

ii

iii
III HIPÓTESIS Y OBJETIVOS 33 3.1 Hipótesis 35 3.2 Objetivo general 35 3.3 Objetivos específicos 35 IV MATERIALES Y MÉTODOS. 37 4.1 Procedimientos generales. 40 4.2 Establecimiento de las condiciones para la inducción
morfogénica in vitro de B. purpurea (Lam.). 40
4.2.1 Material vegetal inicial. 40 4.2.2 Medios y condiciones de cultivo 40 4.3 Diseño experimental. 40 4.4 Estandarización de las técnicas para el estudio molecular. 42 4.4.1 Extracción de ADN. 42 4.4.2 Amplificación al azar del polimorfismo del ADN (RAPD). 43 4.4.2.1 Selección de oligonucleótidos. 43 4.4.2.2 Condiciones de amplificación del ADN genómico de Bletia
purpurea (Lam.). 44
V RESULTADOS Y DISCUSIÓN. 47 5.1 Establecimiento de las condiciones para la inducción
morfogénica in vitro de B. purpurea (Lam.). 49
5.2 Diseño experimental. 51 5.3 Estandarización de las técnicas para el estudio molecular
utilizando extractos de ADN de Sphatoglotis plicata. 57
5.3.1 Extracción de ADN a partir de brotes de Sphatoglotis
plicata. 57
5.3.2 Amplificación al azar del polimorfismo del ADN (RAPD). 59 5.3.2.1 Selección de oligonucleótidos. 60 5.4 Estudio de la variación somaclonal en Bletia purpurea
(Lam.). 65
VI CONCLUSIONES. 71

iv

v
VII RECOMENDACIONES. 75 VIII REFERENCIAS BIBLIOGRÁFICAS. 79
IX ANEXOS. 93 Anexo 1. Diagramas de flujo de cada una de la técnicas probadas para
la extracción del ADN genómico a partir de brotes de
Sphatoglotis plicata.
95

vi

vii
LISTA DE TABLAS
Página Tabla 2.1 Comparación de técnicas de marcadores moleculares
frecuentemente utilizados (Agarwal et al., 2008). 28
Tabla 4.1 Factores y dominio experimental para la inducción morfogénica de Bletia Purpurea (Lam.).
41
Tabla 4.2 Tratamientos de la MSR para la inducción morfogénica de B. purpurea (Lam.) con variable codificada y real.
41
Tabla 4.3 Secuencias de oligonucleótidos RAPD. 44
Tabla 5.1 Resultados obtenidos con los tratamientos del primer diseño, en la inducción morfogénica a partir de pseudobulbos de Bletia purpurea (Lam.), a los 45 días de cultivo.
52
Tabla 5.2 Resultados obtenidos con los tratamientos del segundo diseño, en la inducción morfogénica a partir de pseudobulbos de Bletia purpurea (Lam.), a los 45 días de cultivo.
53
Tabla 5.3 Ecuaciones cuadráticas para cada una de las variables de respuesta, obtenidas del análisis de regresión con los resultados de todos los tratamientos planteados para la inducción morfogénica de Bletia purpurea (Lam.).
53
Tabla 5.4 Resultados de la cuantificación espectrofotométrica de los extractos de ADN obtenidos a partir de brotes de
Spathoglottis plicata.
59
Tabla 5.5 Número de bandas observadas en gel de agarosa de los productos de PCR amplificados de Spathoglottis plicata, para la selección de oligonucleótidos.
62

viii

ix
Tabla 5.6 Resultados de la cuantificación espectrofotométrica de los
extractos de ADN obtenidos a partir de plántulas obtenidas mediante la germinación in vitro y plántulas generadas mediante la organogénesis en explante de pseudobulbo de Bletia purpurea (Lam.).
66
Tabla 5.7 Número de bandas observadas en gel de agarosa de los productos de PCR amplificados de Bletia purpurea (Lam.), para la selección de oligonucleótidos.
68

x

xi
LISTA DE FIGURAS
Página Figura 2.1 Morfología de B. purpurea (Lam.). 13 Figura 2.2 Hábitat típico de Bletia purpurea (Lam.) en Florida. 15 Figura 2.3 Estructura química de algunas auxinas naturales y
sintéticas. 20
Figura 2.4 Estructura química de algunas citocininas naturales y
sintéticas. 21
Figura 2.5 Esquema que muestra los principales pasos de la técnica de
RAPD (Valadez y Kahl, 2000). 29
Figura 4.1 Diagrama de flujo de la estrategia experimental. 39 Figura 5.1 Organogénesis de Bletia purpurea (Lam.) a los 15 días de
la inducción morfogénica, formación directa de brotes sobre explante de pseudobulbo.
50
Figura 5.2 Superficie y contorno de la superficie estimado para la
variable de respuesta: número de brotes. 54
Figura 5.3 Superficie y contorno de la superficie estimado para la
variable de respuesta: número de hojas. 55
Figura 5.4 Superficie y contorno de la superficie estimado para la
variable de respuesta: número de raíces. 56
Figura 5.5 Gel de agarosa para extractos obtenidos por método CTAB a partir de brotes de Spathoglottis plicata.
59
Figura 5.6 Perfiles RAPDs obtenidos de los productos de PCR
amplificados del ADN de Spathoglottis plicata para la selección de oligonucleótidos.
61
Figura 5.7 Perfiles RAPDs comparativos para la estandarización de la
técnica. 63

xii

xiii
Figura 5.8 Gel de agarosa para extractos obtenidos por método CTAB a partir de brotes de Bletia purpurea (Lam.).
66
Figura 5.9 Perfiles RAPDs de los productos de PCR amplificados del
ADN de plántulas de Bletia purpurea (Lam.), realizado para la selección de oligonucleótidos.
67
Figura 5.10 Perfiles RAPDs comparativo de los productos de PCR
amplificados del ADN de plántulas madre e hijas de Bletia
purpurea (Lam.), realizado con oligonucleótidos seleccionados.
69

xiv

xv
LISTA DE ABREVIATURAS
Acetato de sodio Ácido desoxirribonucleico Ácido clorhídrico Ácido indol acético Ácido indol-3-butírico Ácido 2, 4-diclorofenoxiacético Ácido naftalenacético Ácido etilen diamino tetra acético Amplificación aleatoria del ADN polimórfico Bencil amino purina (N6 Benziladenina) Bromuro de hexadeciltrimetilamonio Bromuro de etidio Cloruro de sodio Cloruro de potasio Cultivo de tejidos vegetales Cuerpos tipo protocormos Etanol Hidróxido de sodio Hipótesis nula Isopentenil-adenina (N6-2-Isopentil adenina) Mercaptoetanol Metodología de superficie de respuesta Murashige y Skoog Polivinilpirrolidona Ribonucleasa A Reacción en cadena de la polimerasa Reguladores de crecimiento vegetal Sulfato de amonio Sulfato de magnesio Tidiazuron Hidroximetil amino metano (2-Amino-2-hidroximetil-propano-1,3-diol)
NaOAc ADN
HCl AIA IBA
2,4-D ANA
EDTA RAPD
BAP CTAB
BrEt NaCl KCl
CTV PLBs EtOH
NaOH Ho
2-iP ME
MSR MS
PVP RNAsa
PCR RCV
(NH4)2SO4 MgSO4
TDZ TRIS


1
RESUMEN El cultivo in vitro es una herramienta útil para la propagación de plantas en peligro de extinción y consideradas bajo amenaza como es el caso de Bletia. Purpurea (Lam.). En este trabajo el objetivo fue establecer un protocolo de micropropagación para B.
purpurea, mediante el cual, no se generará variación genética entre las plántulas madre y las plántulas regeneradas. Para lograrlo, se empleó la metodología de superficie de respuesta (MSR) para optimizar la concentración del Ácido Indol Acético (AIA) y la Bencil Amino Purina (BAP), sobre la morfogénesis in vitro de Bletia purpurea (Lam.) y la aplicación de la Amplificación Aleatoria del ADN Polimórfico (RAPD, por sus siglas en inglés) para la evaluación genética de las plántulas micropropagadas. En los experimentos planteados para la morfogénesis se obtuvo la organogénesis directa de brotes sobre el explante de pseudobulbo y mediante la MSR se obtuvo que el valor óptimo para lograrlo fue 1.8 mg L-1 del AIA. Se establecieron las condiciones para la evaluación genética de esta especie de orquídea, mediante la técnica RAPD, seleccionando un protocolo de extracción de ADN genómico y la estandarización de las condiciones de amplificación. Se obtuvo un rendimiento de 20 �g por cada 300 mg de tejido y los geles comparativos de la estandarización de la Reacción en cadena de la polimerasa (PCR, por sus siglas en inglés) demostraron que la concentración de 25 ng de ADN genómico, 50 ng de oligonucleótidos y una temperatura de hibridación de 40 ºC fueron las óptimas para obtener bandas representativas del genoma. De los 20 oligonucleótidos probados se seleccionaron 6 (OPJ-4, OPJ-7, OPJ-13, OPJ-15, OPJ-16 y OPJ-18) para el análisis genético. Al comparar los perfiles obtenidos de la plántula madre e hijas, no se encontró cambio en el genoma, lo cual sugiere que los pseudobulbos de B. purpurea pueden ser utilizados para regenerar brotes múltiples directamente del explante sobre medio de cultivo MS suplementado con 1.8 mg L-1 del AIA, por lo cual de acuerdo a los resultados obtenidos en este estudio, podemos asumir que se mantiene las características genéticas de las plántulas obtenidas en la primera generación. Por lo tanto, en el presente trabajo se logró establecer las bases para la aplicación de la técnica RAPD para estudios posteriores en esta especie de orquídea y las plántulas regeneradas in vitro podrían ser utilizadas como material vegetal para otro tipo de investigaciones. Palabras clave: Variación genética; PCR; análisis RAPDs; Bletia purpurea; pseudobulbos; organogénesis.

2

3
ABSTRACT
In vitro culture is a useful tool for the propagation of endangered plant species and considered under threat as is the case for Bletia purpurea (Lam.). In this work, the goal was to establish a micropropagation protocol for Bletia purpurea (Lam.), through which not generate genetic variation among regenerated plants and mother plant. To achieve this, was employed the Response Surface Methodology (RSM) to optimize the concentration of Indole Acetic Acid (IAA) and Benzyl Amino Purine (BAP) on in vitro morphogenesis of Bletia purpurea (Lam.) and the implementation of Random Amplified Polymorphic DNA (RAPD) technique for genetic evaluation. The results obtained revealed direct shoot organogenesis on explant of pseudobulbs and the RSM showed that to achieve the highest number of shoots was necessary a concentration of 1.8 mg L-1 of the AIA. Conditions were established for genetic evaluation of this orchid species by RAPD technique, selecting a protocol for genomic DNA extraction and amplification conditions. A yield of 20 µg per 300 mg of tissue and gels comparative standardization of Polymerase Chain Reaction (PCR) showed that the concentration of 25 ng genomic DNA, 50 ng oligonucleotide and a hybridization temperature of 40 ° C were optimal for obtain representative bands of the genome. Of the 20 primers tested were selected 6 (OPJ-4, OPJ-7, OPJ-13, OPJ-15, OPJ-16 y OPJ-18) for genetic analysis. By comparing the profiles obtained from the mother and daughters seedling, there was no change in the genome, suggesting that the pseudobulbs B. purpurea can be used to regenerate multiple shoots directly from the explant of pseudobulbs on MS medium supplemented with 1.8 mg L-1 of the IAA, so according to the results obtained in this study, we assume that is kept the genetic characteristics of seedlings obtained in the first generation. Therefore, in this study were able to establish the basis for the application of RAPD technique for further studies in species of orchids and the regenerated plantlets in vitro could be used as vegetal material for other research.
Keywords: Genetic variation, PCR; RAPDs analysis, Bletia purpurea, pseudobulb, organogenesis.

4

5
I. INTRODUCCIÓN

6

7
Bletia purpurea (Lam.), es una especie silvestre que se encuentra bajo amenaza debido a la urbanización y alteración de su hábitat, la disminución de sus poblaciones hace necesario buscar alternativas para su propagación (Dutra et. al., 2008). B.
purpurea se propaga por semillas y vegetativamente por división de plantas. La propagación por semillas requiere de la asociación micorrízica y la progenie no es idéntica a la planta madre. La propagación vegetativa es lenta y no es suficiente para superar la amenaza de extinción. Una herramienta muy útil es el cultivo de tejidos vegetales (CTV), debido a que partir de una célula o grupo de células de algún explante de tejido podría regenerarse una planta completa y para el desarrollo óptimo del cultivo es necesario que además de los nutrientes, se incluya en el medio de crecimiento una o más sustancias reguladoras del crecimiento, como las auxinas y/o citocininas. Sin embargo, las condiciones de cultivo pueden llegar a ser estresantes para las células y es posible obtener variación genética. La variación somaclonal se define como la variación genética o epigenética que se genera durante el cultivo in
vitro de plantas que provenga de células somáticas, así como en plantas regeneradas de los mismos. La variación generalmente es espontánea y los cambios pueden ser heredables o no. Cuando la variación somaclonal es heredable se le asocia con rearreglos cromosomales, deleciones y mutaciones (Sánchez et al., 2003; Noro et al., 2007). Por otro lado, cuando la variación somaclonal es epigenética, puede ser resultado de un cambio en la expresión de los genes, reversible y no heredable (Pierik, 1997; Kaeppler et al., 2000; Jain, 2001; Smulders, 2005), que puede estar asociado a alteraciones en los patrones de metilación del ADN (Kubis et al., 2003). La variación somaclonal puede detectarse por cambios fenotípicos visibles o por medio de marcadores bioquímicos y moleculares (Brar y Brar, 1998; Sahijram et al., 2003). Durante los últimos 20 años se han desarrollado diferentes técnicas moleculares, muchas de ellas son una herramienta útil para detectar variación somaclonal de una manera más precisa y a nivel de ADN (Gupta y Varshney, 1999, Cassells y Curry, 2001; Polanco y Ruiz, 2002; Martín et al., 2002; Mujib, 2004), así como para determinar niveles y los mecanismos por los cuales ocurre la variación somaclonal. Los marcadores moleculares detectan directamente las diferencias genéticas entre individuos, obteniéndose un perfil molecular o huella genética característico para cada variedad e independiente de las condiciones de crecimiento de las plantas (Narváez et al., 2000). Los métodos basados en la técnica de PCR identifican perfiles únicos de fragmentos de ADN al variar la naturaleza del oligonucleótido iniciador y las condiciones de su alineamiento. Entre ellos se encuentra el método RAPD, el cual difiere de otros métodos en que utiliza decanucleótidos de secuencias al azar para amplificar por PCR áreas específicas distribuidas al azar en el genoma. Los oligonucleótidos pequeños y la baja temperatura de alineamiento (36 ºC) aseguran

8
que se una a un gran número de secuencias en el genoma para conseguir amplificar muchos fragmentos de ADN. Con los fragmentos se obtienen perfiles electroforéticos que varían según el polimorfismo de los distintos individuos o grupo de individuos y proporcionan una huella característica (Williams et al., 1990; Lara, 2005; Narváez et
al., 2000; Agarwal et al., 2008). En vista de lo anterior, se planteó como el objetivo general de este trabajo: estudiar mediante marcadores RAPDs la posible variación entre las plántulas madre y las estructuras morfogénicas generadas in vitro; para lograrlo, se empleó la metodología de superficie de respuesta (MSR) con la finalidad de optimizar la concentración de una auxina (Ácido Indol Acético) y una citocinina (Bencil Amino Purina), en la inducción morfogénica de Bletia purpurea (Lam.); así como, la obtención del perfil genético mediante la técnica RAPD de las estructuras morfogénicas regeneradas a partir del tratamiento evaluado para la validación del modelo experimental y comparar con el de la plántula madre para verificar posibles polimorfismos; con la finalidad de obtener un protocolo de micropropagación apropiado que no produzca variación genética durante inducción morfogénica para la regeneración de la especie ya sea para estudios o bien para su posible uso en la multiplicación masiva de la especie.

9
II. ANTECEDENTES

10

11
México es el cuarto país con mayor diversidad de plantas con flores. De las plantas con flores, la familia Orquideaceae, es una de las más grandes y diversas, a esta pertenecen las orquídeas. Taxonómicamente, representa una de las familias más desarrolladas entre las monocotiledóneas con aproximadamente 750 géneros, 25 000 especies, y más de 30 000 híbridos son cultivados en todo el mundo. En nuestro país existen alrededor de 1000 especies, y de ellas pueden ser encontradas poco más del 10 % en la Península de Yucatán. Sin embargo, muchas de estas especies aún no se han estudiado y aprovechado en toda su magnitud, por lo cual es importante sumar esfuerzos y generar conocimiento que nos permita preservar los recursos silvestres (Hew y Clifford, 1993; Sánchez et al., 2002; Li y Ge, 2006; Luan et al., 2006). 2.1 Características generales de las orquídeas Generalmente las orquídeas tienen un aspecto herbáceo, aunque existen en una gran variedad de formas, dependiendo del clima y hábitos característicos de su medio natural. El crecimiento de la planta se presenta de dos formas: Monopodial: se da cuando el crecimiento es por la yema apical, siempre aumentando la altura. Simpodial: Se presenta por el rizoma que al avanzar va originando nuevos brotes de la planta. El hábitat de las orquídeas es muy variado, es común encontrarlas en las ramas de los árboles, epifitas; las rocas, rupícolas; el suelo, hojarasca o arena, aún en terrenos pantanosos; terrestres. La mayoría de las orquídeas simpodiales son terrestres. Las orquídeas de regiones frías y muy templadas son terrestres, y también pueden encontrarse en climas más cálidos. En cambio, las epifitas solo crecen en regiones cálidas (Echeverri et al., 2001; Bellido, 2007). Estas plantas constan de raíces, tallos, hojas y flores. La mayoría de las orquídeas epífitas poseen una estructura más gruesa en la base de sus hojas, llamado pseudobulbo. En general, el pseudobulbo es la porción más gruesa del tallo del cual surgen todas las hojas y las inflorescencias. El pseudobulbo puede ser clasificado, independientemente de la forma, como homoblástico (muchos entrenudos) y heteroblástico (entrenudo único) en base al tipo y número de entrenudos que lo forman. Estudios realizados a pseudobulbos de varias orquídeas han revelado la ausencia de estomas. Sin embargo, el rol de los pseudobulbos como órganos de almacenamiento de agua y alimentos es bien conocido (Hew y Yong, 2004). El pseudobulbo tiene una estructura única, el órgano es cubierto por una cutícula gruesa carente de estomas. La epidermis consiste de dos, tres o cuatro paredes de capas gruesas de células parenquimatosas. La matriz no es diferenciada claramente y

12
no hay corteza discernible. Los sistemas vasculares están dispersos de manera irregular en toda la masa. Dos tipos principales de células se han reportado para varias especies de orquídeas. Hay pequeñas células de asimilación que están vivas y contienen predominantemente cloroplastos y granos de almidón; y grandes células muertas que son de forma irregular con paredes plegadas. En comparación con la parte externa, la parte interna es de un tono de verde más ligero; esto se le atribuye a la distribución de las células vivas, las células vivas más cercanas a la epidermis son ricas en cloroplastos, pero carentes de granos de almidón cerca del centro son ricas en granos de almidón y carecen de cloroplastos. En base a estudios anatómicos la posible función de las células vivas más pequeñas es como fuente de almacenamiento de almidón mientras que las células grandes pueden ser utilizados para el almacenamiento de agua (Hew y Yong, 2004).
Las flores son la parte más distintiva de la planta y esencial para la definición y diferenciación de las especies; generalmente emergen en la punta o a lo largo de un pedúnculo que recibe el nombre de escapo floral, consta de tres sépalos y tres pétalos (tépalos). Los pétalos son dos iguales y un tercero diferenciado que recibe el nombre de labelo. En la mayoría de las especies los órganos reproductores masculinos y femeninos están unidos en una sola estructura conocida como columna. El polen de estas flores forma masas compactas que reciben el nombre de polinios, casi siempre se presentan en pares. El fruto es una cápsula dehiscente, es decir, que abre sola al madurar y esparce las semillas (Sánchez, 1998). 2.2 Importancia comercial de las orquídeas La venta de orquídeas como flores de corte o plantas de maceta contribuye significativamente al comercio nacional e internacional. El desarrollo de nuevos híbridos y el cultivo comercial es una gran industria en algunos países como Europa, Hawái, Australia, Japón, Tailandia, Singapur, Sri Lanka y Kenia (Luan et al., 2006). El éxito comercial de las orquídeas se debe a que las flores muestran remarcada longevidad; así como diversidad de formas, tamaños, colores, estructuras, número y fragancia de flores. Algunas especies se utilizan con propósitos terapéuticos como tonificantes. Debido a sus características las orquídeas son depredadas masivamente para el comercio o por coleccionistas. Y si a esto, le sumamos la destrucción de sus hábitats ya sea por actividades antropogénicas o naturales, podemos decir que la población de las orquídeas está en constante disminución (Hew y Clifford, 1993; Lo et al., 2004; Li y Ge, 2006).

2.3 Bletia purpurea Bletia purpurea (Lam.) urbanización y alteración de su hábitat. cual fue descrito por los botánicos Ruíz y Pavón, en el año de 1883; diversificado en el centro de México,intermedias consta de aproximadamente 35et al., 2008). Subfamilia: Epidendroideae LindleyTribu: Arethuseae LindleySubtribu: Bletiinae BenthamSinonimias:
Bletiana RafinesqueGyas Salisb. Jimenesia RafinesqueRegnellia B. R. Thiebautia Colla
2.3.1 Descripción botánica Las plantas de B. purpurea
con un corto cormo, pseudobulbos: comprimidos en la parte superior, de 3.8 cm de diámetro máximo, casi siempre producidos en largas cadenas, subterráneos o parcialmente expuestos, con pocas hojas. elíptico-lanceolada, plicatas, largamenteescapo, de 20 a 90 cm de largo y de 1 a 5 cm de ancho
Figura 2.1. Morfología de (c) Cápsulas maduras. Fuente: Dutra
13
(Lam.) DC
(Lam.) una especie considerada bajo amenaza urbanización y alteración de su hábitat. Pertenece al género Bletia Ruiz & Pav., el
fue descrito por los botánicos Ruíz y Pavón, en el año de 1883; diversificado en el centro de México, por lo general en las elevaciones altas o intermedias consta de aproximadamente 35-40 especies (Carnevali et al.,
Epidendroideae Lindley Arethuseae Lindley
Bletiinae Bentham
Rafinesque
Rafinesque
Colla
botánica
B. purpurea son terrestres o litófitas (Figura 2.1 a), glabrasseudobulbos: comprimidos en la parte superior, de 3.8 cm de
diámetro máximo, casi siempre producidos en largas cadenas, subterráneos o parcialmente expuestos, con pocas hojas. Las hojas son lineales a estrechamente
lanceolada, plicatas, largamente acuminadas, la parte más baja envainando al escapo, de 20 a 90 cm de largo y de 1 a 5 cm de ancho (Sánchez et al., 2002).
Morfología de Bletia purpurea (Lam.). (a) Plantas de Bletia purpurea
Fuente: Dutra et al., 2008.
debido a la Ruiz & Pav., el
fue descrito por los botánicos Ruíz y Pavón, en el año de 1883; se ha por lo general en las elevaciones altas o
et al., 2001; Dutra
, glabras, erectas, seudobulbos: comprimidos en la parte superior, de 3.8 cm de
diámetro máximo, casi siempre producidos en largas cadenas, subterráneos o lineales a estrechamente
acuminadas, la parte más baja envainando al , 2002).
Bletia purpurea (b) Flores

14
La inflorescencia abierta, con pocas o muchas flores (Figura 2.1 b), un racimo simple o paniculado, que nace de un pedúnculo lateral largo y delgado; el pedúnculo está provisto con vainas tubulares remotas de 0.27 a 1.7 m de largo y las brácteas florales principalmente pequeñas, de ovado-triangulares a ovado-lanceoladas, de agudas a acuminadas, de 2 a 8 mm de largo. Las flores rosas, rosa-purpura o purpura oscuro, rara vez casi blancas, vistosas, con ovarios delgados pedicelados, de 9 a 18 mm de largo, variable en tamaño, generalmente abren en forma sucesiva (Sánchez et al., 2002). Los sépalos son dorsales de oblongo-elíptico a ovado-lanceolado, de semiobtuso a agudo, de 1.5 a 2.6 de largo y de 5 a 9 mm de ancho; sépalos laterales de oblicuamente ovado-oblongo a elíptico-oblongo de abruptamente agudos a acuminados, márgenes involutos, de 1.2 a 2 cm de largo y de 5 a 8 mm de ancho (Sánchez et al., 2002). Los pétalos de oblicuamente ovado-oblongos a elípticos u oblongo-lanceolados, de obtusos a agudos, de 1.2 a 2.1 de largo y de 7 a 11 mm de ancho, forman una capucha sobre el labelo (Sánchez et al., 2002). El labelo es ampliamente cordado, ovado-cordado o cordado-semicuadrado, cuando está extendido hacia afuera, conspicuamente trilobulado encima de la parte media, fuertemente recurvado en sus posición natural, con la base truncada o subauriculada, de 1 a 1.8 cm de largo y de 8 a 14 mm de ancho, a través de los lóbulos laterales, los cuales son incurvados en sus posición natural, ampliamente redondeados en la base, con el ápice de ahusado, a triangular-obtuso o redondeado; el lóbulo medio suborbicular, de truncado a profundamente emarginado en el ápice, márgenes ondulado-crenulados, de 5 a 10 mm de ancho; disco venoso, con cinco a siete laminillas que se extienden de cerca de la base del labelo hasta cerca del ápice del lóbulo medio, con dos laminillas más cortas sobre los lóbulos laterales (Sánchez et
al., 2002). La columna es fuertemente arqueada, clavelada, con alas laterales estrechas de 8 a 12 mm de largo. El fruto es una capsula oblicuamente cilíndrica (Figura 2.1 c), erecta, rojiza a chocolate-marrón, con 2 a 4.5 cm de largo y 8 a 10 mm de diámetro (Sánchez et al., 2002). 2.3.2 Distribución geográfica de la especie. B. purpurea se distribuye desde Florida en Estados Unidos de América (Figura 2.2), México (Jalisco, Michoacán, México, Morelos, Guerrero, Oaxaca, Tamaulipas, San

Luis Potosí, Veracruz, Puebla, Tabasco, Chiapas)Colombia, Ecuador. Se encuentran en bosques secos y en llanuras, también puede encontrársele en la base de troflotando en agua, sobre grupos de hierbas o pastos, en altitudes de hasta 2000 m.s.n.m (Sánchez et al., 2002). lugares abiertos como irrigación, especialmentepoblaciones de B. purpurea
(Carnevali et al., 2001necesario desarrollar un método de propagación que sea eficiente y ecológicamente viable (Dutra et al., 2008)
Figura 2.2 Hábitat típico de
2.4 Propagación vegetativa de las orquídeas Las orquídeas requieren de una multitud de factores para la continua reproducción en la naturaleza. La propagación de las través de métodos convencionales es un proceso lento. a través de división de matas o rizomas, esquejes oproducidos a partir de la madre o después de 2-3 años. A pesar de que las orquídeas producen frutos cmillones de semillas Orquídeas son liberadas de sus cápsulas cuando estas maduran y son llevadas por el viento, pudiendo germinar y crecer si eventualmente caen en un medio adecuado la germinación (características específicas del árbol hospedero y de condiciones
15
Potosí, Veracruz, Puebla, Tabasco, Chiapas) hasta América Central, Venezuela, Se encuentran en bosques secos y en llanuras, también puede
la base de troncos de árboles, sobre trozas y tocones en pantanos, o flotando en agua, sobre grupos de hierbas o pastos, en altitudes de hasta 2000 m.s.n.m
., 2002). Es una especie terrestre y silvestre crece generalmente en s como suelos arenosos cerca de los bordes de las aguas para la
irrigación, especialmente en el sureste de la Península de Yucatán. Las flores de las B. purpurea en la Península de Yucatán son bastante variables
2001). La disminución de las poblaciones en América necesario desarrollar un método de propagación que sea eficiente y ecológicamente
2008).
típico de Bletia purpurea en Florida. Fuente: Dutra et al., 2008
vegetativa de las orquídeas
Las orquídeas requieren de una multitud de factores para la continua reproducción en La propagación de las orquídeas silvestres, endémicas y nativas,
través de métodos convencionales es un proceso lento. Con la propagación vegetativa n de matas o rizomas, esquejes o la separación de vástagos
producidos a partir de la madre o pseudobulbos, solo se obtienen unas cuantas A pesar de que las orquídeas producen frutos con cientos hasta por cápsula, bajo condiciones naturales, las semillas de
Orquídeas son liberadas de sus cápsulas cuando estas maduran y son llevadas por el viento, pudiendo germinar y crecer si eventualmente caen en un medio adecuado
(características específicas del árbol hospedero y de condiciones
América Central, Venezuela, Se encuentran en bosques secos y en llanuras, también puede
ncos de árboles, sobre trozas y tocones en pantanos, o flotando en agua, sobre grupos de hierbas o pastos, en altitudes de hasta 2000 m.s.n.m
crece generalmente en arenosos cerca de los bordes de las aguas para la
as flores de las son bastante variables
oblaciones en América hace necesario desarrollar un método de propagación que sea eficiente y ecológicamente
2008
Las orquídeas requieren de una multitud de factores para la continua reproducción en silvestres, endémicas y nativas, a
a propagación vegetativa la separación de vástagos
pseudobulbos, solo se obtienen unas cuantas plantas on cientos hasta
, bajo condiciones naturales, las semillas de Orquídeas son liberadas de sus cápsulas cuando estas maduran y son llevadas por el viento, pudiendo germinar y crecer si eventualmente caen en un medio adecuado para
(características específicas del árbol hospedero y de condiciones

16
ambientales favorables). Sin embargo, dichas semillas tienen pocas posibilidades de sobrevivir, debido a que muchas de las especies de orquídeas sólo son capaces de crecer en presencia de un hongo simbiótico: Rhizoctonia spp. Una manera de salvaguardar las orquídeas es por medio de los métodos de cultivo in vitro (Luan et
al., 2006; Romero et al., 2007; Pedroza, 2009). 2.5 Cultivo de tejidos vegetales Pérez, et al. (1999), definieron al Cultivo de tejidos Vegetales (CTV) como al conjunto de técnicas que permiten el establecimiento, mantenimiento y desarrollo de brotes y/o embriones somáticos a partir de cualquier célula, tejido u órgano de una planta, bajo condiciones artificiales, axénicas y controladas. El objetivo de la propagación de plantas vía cultivo de tejidos, llamado micropropagación, es una propagación de plantas fiel al tipo, esto es, obtener clones. Los sistemas de CTV se han convertido en una herramienta indispensable para estudios de fisiología y bioquímica vegetal, obtención de plantas libres de organismos patógenos, propagación masiva de plantas de interés, creación y conservación de bancos de germoplasma, producción y biosíntesis de metabolitos secundarios y fármacos de interés económico, mejoramiento genético mediante la inducción de mutaciones y la selección in vitro, la ingeniería genética, el incremento de poblaciones de especies que se encuentren en peligro de extinción (Boon, 1991; Hurtado y Merino,1994; Jiménez, 1998; Arias, 2000; Taniguchi et al., 2008). De acuerdo a Alonso (2002), los métodos de propagación vegetativa in vitro se pueden clasificar en función del tejido que se ponga en cultivo. 1. Cultivo de plantas intactas: siembra de una semilla in vitro, obteniéndose primero una plántula y finalmente una planta. 2. Cultivo de embriones: cultivo de embriones aislados después de retirar el resto de los tejidos de la semilla. 3. Cultivo de meristemos y yemas terminales o laterales. 4. Cultivo de órganos aislados: raíces, anteras o bien porciones de tejidos u órganos aislados de la planta (explantes). 5. Cultivo de callo: cultivo de tejido desdiferenciado in vitro formándose órganos adventicios o embriones somáticos. 6. Cultivo de células aisladas de un tejido, un callo o un tejido en suspensión, obtenidas con la ayuda de enzimas o mecánicamente. 7. Cultivo de protoplastos que se obtienen por digestión enzimática de la pared celular.

17
Los métodos de propagación vegetativa que se seleccionen en la multiplicación in
vitro dependen de la planta que se vaya a multiplicar y de los objetivos finales del esquema de micropropagación. Se debe elegir el tejido del que se quiere partir para la micropropagación; así como, la forma de diferenciación y de regeneración de órganos. La mejor opción no es siempre la que mayor número de brotes produce (Qureshi y Saxena, 1992). 2.5.1 Morfogénesis in vitro La morfogénesis implica la formación de órganos o embriones somáticos a partir de células somáticas de los tejidos de los órganos aislados. Puede ser indirecta si existe una etapa intermedia y se forma a partir de callos o directa sin la formación de callo. En la morfogénesis indirecta, las células y/o los tejidos se desdiferencian para formar callo. Si el callo se disgrega, se originan grupos de células (agregados), y/o células aisladas (cultivos en suspensión). La inducción de los procesos morfogénicos se inducen fundamentalmente mediante el uso de reguladores de crecimiento como auxinas y/o citocininas en el medio nutritivo (Pierik, 1990; Alonso, 2002). De manera general, se sabe que bajo las condiciones adecuadas, ciertas células dentro de un explante comienzan a dividirse cambiando su estado de diferenciación y dando lugar a regiones llamadas meristemoides. Los meristemoides están compuestos por células pequeñas, isodiamétricas, de pared celular delgada, varias vacuolas periféricas pequeñas y un núcleo grande en posición central, lo cual corresponde a las características de las células meristemáticas comunes. Cada uno de estos meristemoides es capaz de desarrollarse para dar lugar a brotes o embriones somáticos (Pérez et al., 1999). El punto crítico en este proceso es el momento en que una célula dentro del explante, usualmente parenquimatosa y vacuolada, comienza a dividirse para dar lugar al grupo de células que componen el meristemoide. A este proceso en concreto se le da el nombre de inducción. Existen dos teorías básicas acerca del proceso de inducción; la primera de ellas, propone que en un momento dado ciertas células dentro del explante sufren un aislamiento físico y/o fisiológico con respecto al resto del tejido, lo cual acarrea una pérdida de los controles externos y permite que las células aisladas expresen su potencial de desarrollo y dividiéndose y formando los meristemoides (Pérez et al., 1999). La segunda teoría, propone que son los cambios cuantitativos en la disponibilidad de nutrientes y reguladores de crecimiento en una célula particular, lo que induce a esta a dividirse y formar los meristemoides. En la mayoría de los casos estudiados, los

18
brotes o embriones se originan a corta distancia del medio de cultivo, o bien del tejido vascular, lo cual parece indicar que los gradientes fisiológicos que se dan dentro del explante tiene mucho que ver con el desarrollo de los meristemas. De acuerdo a los conocimientos actuales, la segunda teoría puede ser la más acertada (Pérez et al., 1999). Existen varios factores que deben ser considerados para la manipulación exitosa de la morfogénesis. Entre ellos se menciona el tamaño y la edad fisiológica de los explantes, pues se ha comprobado que los tejidos juveniles presentan una mayor aptitud para la morfogénesis que los tejidos que provienen de órganos adultos o senescentes. También la luz, la temperatura, consistencia del medio de cultivo, el pH y los reguladores de crecimiento desempeñan un papel fundamental en el desarrollo de la respuesta (Pérez et al., 2008). 2.5.1.1 Embriogénesis somática. La embriogénesis somática es el proceso en el cual las células somáticas se desarrollan para dar lugar a plantas diferenciadas pasando por los estados embriogénicos pero sin la fusión de gametos. Los embriones somáticos proceden de células individuales, son bipolares y no están conectados al sistema vascular de la planta madre. Los embriones somáticos tienen la ventaja de ser verdaderos clones, y no quimeras, al proceder de una sola célula; además, se puede aumentar la eficiencia en la formación de plantas al producirse con menor número de pasos y en mayores cantidades con mayor uniformidad tanto morfológica como citológicamente (Bajaj, 1991). 2.5.1.2 Organogénesis. La organogénesis constituye una de las posibles vías morfogénicas para la diferenciación de plantas de novo. Consiste en la formación de raíces o brotes adventicios en los explantes cultivados in vitro, que puede ocurrir en forma directa a partir de células, tejidos u órganos vegetales iniciadores del cultivo in
vitro o indirecta a partir de callos. Aun cuando la organogénesis es un proceso muy utilizado para la propagación de plantas, es muy poco lo que se sabe acerca de los fenómenos fisiológicos, bioquímicos y genéticos que lo hacen posible (Pérez et al.,
2008). 2.5.2 Función de los reguladores de crecimiento vegetal en el desarrollo in vitro
Para el desarrollo óptimo del cultivo in vitro de tejidos y órganos es necesario que en adición a los nutrientes, se incluya en el medio de crecimiento una o más sustancias reguladoras. Las primeras hormonas y reguladores sintéticos con efectos sobre el

19
crecimiento de las plantas, fueron identificados como hormonas del crecimiento, reguladores del crecimiento, sustancias reguladoras del crecimiento o sustancias del crecimiento (Jankiewicz, 2003). Los requerimientos de estas sustancias varían considerablemente con los tipos de tejidos y los niveles endógenos de estos reguladores, así como con la finalidad del cultivo in vitro (Lara, 2005). Los reguladores del crecimiento vegetal (RCV) se clasifican en 5 grupos básicos, dependiendo de su estructura química y su efecto fisiológico: auxinas, citocininas, giberelinas, ácido abscísico y etileno. Actualmente se han aislado y estudiado otras sustancias fuera de estos grupos, que podrían ser considerados como RCV, por ejemplo: las poliaminas, jasmonatos, ácido salicílico y los brasinoesteroides (Davies, 1995). Los RCV más utilizados en los cultivos in vitro son los pertenecientes a los grupos de las auxinas y citocininas, de los cuales se conoce regulan en gran medida los procesos de crecimiento y desarrollo. En menor grado se utilizan algunos reguladores no pertenecientes a estos grupos como las giberelinas y el ácido abscísico (Pérez et al., 1999). 2.5.2.1 Auxinas. Las auxinas son un grupo de compuestos derivados comúnmente del triptófano, sintetizados por lo general en los ápices; ejercen un control sobre la replicación del ADN y se participan en muchos procesos del desarrollo vegetal: crecimiento, dominancia apical, enraizamiento, partenocarpia, tropismos, abscisión, elongación celular, entre otros. En cultivo in vitro son utilizadas principalmente para la diferenciación de raíces y frecuentemente en la inducción de callo para embriogénesis y organogénesis en cultivos en suspensión. En la Figura 2.3, se muestran las estructuras químicas de algunas auxinas las de la parte superior e inferior son naturales y sintéticas respectivamente. Las más utilizadas son: AIB (acido indol-3-butírico), ANA (ácido naftalenacético), AIA y 2,4-D (ácido diclorofenoxiacético). El AIB y ANA son usados frecuentemente para enraizamiento. El 2,4-D es más efectivo para la inducción de callos (Salisbury y Ross, 2000; Lara, 2005; Condemarín et al., 2007).

Figura 2.3 Estructura química de algunasal., 2008.
2.5.2.2 Citocininas. Las citocininas afecta a una amplia gama de procesos de crecimiento y desarrollo de las plantas, entre lo que cabe citar la iniciación del desarrollo del cloroplasto, fotosíntesis, la expansión de los cotiledones, así comvegetales. Están implicadas en la división celularacontecimientos que conducela diferenciación de tallos y otros. citocininas para promover la división celular y la inducción de yemas adventicias en callos y órganos. Además se usan estos compuestos para la proliferación de tallos axilares por la ruptura de la dominkinetina (KIN) y isopentenilestructuras químicas de algunas naturales y sintéticas respectivamenteCondemarín et al., 2007;
20
Estructura química de algunas auxinas naturales y sintéticas. Fuente: George
Las citocininas estimulan la división y diferenciación celular que afecta a una amplia gama de procesos de crecimiento y desarrollo de las plantas, entre lo que cabe citar la iniciación del desarrollo del cloroplasto, fotosíntesis, la expansión de los cotiledones, así como la inducción de la formación de brotes en cultivo
stán implicadas en la división celular parecen ejercer control en lacontecimientos que conducen a la mitosis, la modificación de la dominancia apical,
iación de tallos y otros. En los medios para cultivo in vitro para promover la división celular y la inducción de yemas adventicias en
callos y órganos. Además se usan estos compuestos para la proliferación de tallos axilares por la ruptura de la dominancia apical, las citocininas más usadas son: BAP,
isopentenil-adenina (2-iP). En la Figura 2.4, se muestran las estructuras químicas de algunas citocininas las de la parte superior e inferior son naturales y sintéticas respectivamente (Salisbury y Ross, 2000;
., 2007; George et al., 2008).
Fuente: George et
estimulan la división y diferenciación celular que afecta a una amplia gama de procesos de crecimiento y desarrollo de las plantas, entre lo que cabe citar la iniciación del desarrollo del cloroplasto, fotosíntesis, la expansión
o la inducción de la formación de brotes en cultivo recen ejercer control en los
modificación de la dominancia apical, se incorporan
para promover la división celular y la inducción de yemas adventicias en callos y órganos. Además se usan estos compuestos para la proliferación de tallos
s usadas son: BAP, , se muestran las
las de la parte superior e inferior son 2000; Lara, 2005;

Figura 2.4 Estructura química de algunas citocininal., 2008
2.5.3 Cultivo de tejidos vegetales en orquídeas Con la aplicación de los métodos de cultivo orquídeas silvestres, nativas y endémicas amenazadas o en peligro de extinción. Actualmente se están desarrollando investigaciones sobre la germinación simbiótica y asimbiótica in vitro. La germinación asimbióticala producción comercial producción, conservación y propósitos de reintroducción. vitro de semillas se puede obtener información básica del crecimiento y desarrollo temprano de la planta (Su Dutra et al., 2008). Como se ha mencionadovitro sea eficiente la presencia de los RCV y la concentración de ellos en el medio juegan un papel importante.multiplicación de callos embriogénicos, con la combinación de mg L-1 de cada uno, paraextinción.
21
ructura química de algunas citocininas naturales y sintéticas. Fuente: George
Cultivo de tejidos vegetales en orquídeas
Con la aplicación de los métodos de cultivo in vitro ha sido posible la propagación de orquídeas silvestres, nativas y endémicas amenazadas o en peligro de extinción. Actualmente se están desarrollando investigaciones sobre la germinación simbiótica y
. La germinación asimbiótica de las semillas ha sido utilizada para comercial de orquídeas y ha mostrado ser una herramienta útil para la
producción, conservación y propósitos de reintroducción. Mediante la germinación de semillas se puede obtener información básica del crecimiento y desarrollo
de la planta (Su et al., 2006; Johnson et al., 2007; Romero
Como se ha mencionado anteriormente para que la propagación o regeneración sea eficiente la presencia de los RCV y la concentración de ellos en el medio
importante. Lee et al. (2007) reportaron un aumento multiplicación de callos embriogénicos, con la combinación de ANA, BAP y AIA, 2
para Laelia anceps spp. Dawsonii, especie en grave peligro de
Fuente: George et
propagación de orquídeas silvestres, nativas y endémicas amenazadas o en peligro de extinción. Actualmente se están desarrollando investigaciones sobre la germinación simbiótica y
de las semillas ha sido utilizada para de orquídeas y ha mostrado ser una herramienta útil para la
Mediante la germinación in
de semillas se puede obtener información básica del crecimiento y desarrollo ; Romero et al., 2007;
anteriormente para que la propagación o regeneración in
sea eficiente la presencia de los RCV y la concentración de ellos en el medio umento en la
NA, BAP y AIA, 2 , especie en grave peligro de

22
Wu et al. (2004) reportaron que para optimizar la formación de embriones a partir de callos de Oncidium `Gower Ramsey` probaron el efecto por separado de cinco auxinas (2,4 D, AIA, AIB, ANA y picloram) y cinco citocininas (2iP, BAP, KIN, TDZ y Zeatina) y encontraron que la KIN (0.5 y 2 mg L-1) y zeatina (0.5 mg L-1) fueron los RCV más efectivos. Por su parte, Murthy en 2005 describió un método de multiplicación de Aerides
crispum a través de cultivo de protocormos y hojas en medio MS suplementado con auxinas, citocininas (AIA, ANA, BAP, KIN y TDZ) solas y combinadas con agua de coco a las concentraciones de 0.5, 1.0, 2.0, 5.0 µM y endospermo líquido de coco a 5, 10, 15 %, v/v. Los explantes desarrollaron cuerpos tipo protocormos (PLBs) que a las 5-8 semanas desarrollaron a plántulas, las cuales fueron reintroducidas a su hábitat natural. Para el cultivo de tejidos en orquídeas se ha reportado el uso de diferentes explantes como son: hojas, tallos florales, nudos y rizomas con la finalidad de inducir brotes, callos embriogénicos, embriones somáticos y PLBs (Nayak et al., 1997; Sheelavantmath et al., 2000; Tokuhara y Mii, 2001; Chen et al., 2002a, b; Su et al., 2006; Gow et al., 2008; Janarthanam y Seshadri, 2008). El uso de pseudobulbos como explante ha sido reportado por Sharma et al. (2007), que desarrollaron un protocolo para la propagación clonal de Dendrobium
microbulbon A. Rich, una orquídea con propiedades etnomedicinales para su conservación y reintroducción a su hábitat natural; utilizaron medio MS suplementado con 2 mg L-1 de BAP para la formación de brotes. Sunitibala y Rajkumar (2009), reportaron un protocolo para la micropropagación de Dendrobium transparens en segmentos de pseudobulbos derivados de la germinación in vitro. Para la multiplicación utilizaron medio MS con diferentes concentraciones de BAP y en combinación con 1 mg L-1 ANA. El uso de 2 mg L-1 de BAP les dio la mejor respuesta para la producción de callos y para la inducción de brotes múltiples fue el uso de 1 mg L-1 ANA y 2 mg L-1 BAP. Un reporte más reciente fue realizado por Mata et al. (2010), este grupo reportó un protocolo para la propagación in vitro a partir de secciones de pseudobulbos de Lycaste aromatica (Graham ex Hook) Lindl., una especie de orquídea ornamental y con fragancia. Evaluaron el efecto de la KIN, Metatopolina (mT), BAP y TDZ. Mencionan que todos los tratamientos en los cuales se utilizaron secciones apical y basal del pseudobulbo generaron brotes y donde se utilizaron la sección media en medio suplementado con BAP se desarrollaron cuerpos tipo protocormo, el máximo número de brotes se obtuvo con la sección apical en medio con 2.2, 4.4 y 8.87 µM de

23
TDZ. Sin embargo, debido a que la sección media fue el mejor explante, ellos recomiendan que el pseudobulbo sea dividido en dos secciones, la basal podría ser cultivado en MS con 4.4 µM y la apical con 2.2 µM de TDZ. Los brotes que obtuvieron los pasaron a medio sin RCV para su desarrollo y enraizamiento. La razón de supervivencia que tuvieron bajo condiciones de invernadero fue del 90 %. Basker y Narmatha, (2006) desarrollaron un protocolo para la micropropagación de Coelogyne stricta, una especie de orquídea endémica de la India; emplearon segmentos de pseudobulbos de plántulas de un año de edad germinadas in vitro. Los segmentos respondieron positivamente produciendo brotes directamente del explante en medio MS suplementado con ANA y BAP individualmente y en combinación. La combinación de ANA (1 mg L-1) + BAP (2 mg L-1) y BAP solo, favoreció el desarrollo de brotes múltiples. El número de raíces se incrementaron con ANA (1 y 2 mg L-1) en el medio MS. Las plántulas obtenidas fueron exitosamente transferidas a macetas. Cabe señalar que en Bletia purpurea (Lam.) es poca la información científica relacionada con los requerimientos necesarios para la germinación de las semillas, crecimiento, desarrollo y regeneración de la especie (Dutra et al., 2008). Para la regeneración de esta especie de orquídea por cultivo in vitro podría ser necesario el uso de los RCV. Sin embargo, el uso de los RCV y en general, las condiciones de cultivo puede llegar a ser estresantes para las células y es posible obtener variación somaclonal. En especies como B. purpurea en el cual los pseudobulbos son obtenidos en una etapa temprana del desarrollo, podrían ser una excelente fuente de explante. 2.6 Metodología de superficie de respuesta (MSR). Grima et al. (2004), mencionan que uno de los deseos de la humanidad es modelar la realidad, es decir, buscar funciones matemáticas que expliquen los fenómenos. Este deseo de encontrar modelos responde principalmente a dos aplicaciones inmediatas: la capacidad de predecir y optimizar la respuesta. En la práctica, es muy difícil intentar resolver este problema de encontrar un modelo único. En su lugar, se intenta aproximar tales relaciones de manera local a través de funciones polinómicas de primero y segundo orden. Además, el análisis de estas aproximaciones permitirá localizar la región óptima de una manera secuencial. La MSR, es una colección de técnicas matemáticas y estadísticas útiles para analizar problemas en que varias variables independientes influyen en una variable dependiente o respuesta, y la meta es optimizar esta respuesta (Montgomery, 2007).

24
La MSR puede ser definida como, un método estadístico que usa datos cuantitativos de diseños experimentales apropiados para determinar y simultáneamente resolver ecuaciones multivariables, ecuaciones que puedan ser representadas gráficamente como una superficie de respuesta la cual puede ser utilizada para describir como las variables probadas afectan la respuesta, para determinar las interacciones entre ellas y para describir el afecto combinado de todas las variables probadas de respuesta (Nuñez, 1995). Básicamente la MSR es un proceso de cuatro pasos: 1. Identificación de las variables: identificar dos o tres variables críticas del proceso bajo estudio, esto es, los factores que tiene mayor influencia sobre la respuesta analizada. Estos pueden ser determinados por experimentos preliminares, bibliografía o por la experiencia del investigador. El número de variables estudiadas se limita generalmente a dos o tres para simplificar el modelo estadístico. 2. Definición de los niveles de las variables: definir los rangos a probarse para cada variable. Si estos rangos son muy amplios y el óptimo no es definido claramente por MSR, entonces se hace un segundo experimento, usando un rango más estrecho. 3. Diseño experimental: establecer los experimentos a realizarse usando un diseño experimental adecuado. Estos diseños seleccionan un grupo de experimentos, de todos los experimentos posibles, lo cual reduce el número total de pruebas. Después de que los experimentos son especificados, son realizados y así obtenemos lo datos cuantitativos que se utilizaran en el análisis estadístico. 4. Análisis de datos: analizando los datos cuantitativos por medio de un adecuado programa de computación, las conclusiones hechas de estos análisis deberán de ser confirmados con pruebas posteriores, en donde se trabajara con los valores óptimos encontrados para las variables (Nuñez, 1995). La elaboración de gráficas de superficie de respuesta, pueden usarse para determinar las condiciones óptimas de operación, y una vez que las ecuaciones de regresión han sido obtenidas, el proceso puede ser modelado para una amplia variedad de situaciones (Nuñez, 1995). Al analizar una superficie de respuesta puede representar 1) un punto de respuesta máxima, 2) un punto de respuesta mínima o 3) un punto silla (minimax), dependiendo de la relación que exista entre las variables probadas (Montgomery, 2007).

25
2.7 Variación somaclonal La idea original del CTV fue que todas las plantas regeneradas de una célula o tejido tiene una constitución genética idéntica al de la planta original. Sin embargo, el comportamiento celular normal es el resultado de una compleja cascada de programas genéticos que son sensibles a la disrupción por estrés biótico y abiótico. El cultivo in
vitro per se puede llegar a ser muy estresante para las células vegetales y puede involucrar procesos mutagénicos durante el establecimiento del explante, la inducción de callo, la formación de embriones o durante la formación de órganos. Por estas vías es posible obtener variación somaclonal de origen nuclear y/o citoplasmático (Larkin, 2004; Chen et al., 2005). El proceso denominado variación somaclonal, por Larkin y Scowcroft (1981), es la variación genética o epigenética que se genera durante el cultivo in vitro de plantas (cultivos celulares de tejidos u órganos) que provenga de células somáticas, así como en plantas regeneradas de los mismos. La variación somaclonal generalmente es espontánea y los cambios pueden ser heredables o no. Cuando la variación somaclonal es heredable se le asocia con rearreglos cromosomales, deleciones y mutaciones (Sánchez et al., 2003; Noro et al., 2007). Por otro lado, cuando la variación somaclonal es epigenética, puede ser resultado de un cambio en la expresión de los genes, reversible y no heredable (Pierik, 1997; Kaeppler et al., 2000; Jain, 2001, Smulders, 2005), que puede estar asociado a alteraciones en los patrones de metilación del ADN (Kubis et al., 2003; Cardone et al., 2004). Los mecanismos por los cuales ocurre la variación somaclonal no han sido completamente elucidados, pero entre sus causas se mencionan alteraciones en el cariotipo, mutaciones puntuales, recombinación somática e intercambio de cromátidas hermanas, elementos genéticos transponibles y alteración en los patrones de metilación del ADN (Phillips et al., 1994; Kaeppler et al., 2000; Cardone et al., 2004). La variación somaclonal obtenida a partir del cultivo de tejidos depende de factores como: genotipo, nivel de ploidía, tipo y grado de diferenciación del explante, tiempo de cultivo, la vía o sistema de regeneración, y las condiciones, concentración y tipo de RCV; así como la composición del medio de cultivo (Novak, 1990; Skirvin et al., 1994; Duncan, 1997; Cardone et al., 2004; Podwyszyńska, 2005)
2.7.1 Ventajas y desventajas de la variación somaclonal La variación somaclonal puede ser utilizada como herramienta para inducir variabilidad genética y, en el mejor de los casos, obtener características agronómicas deseables (Araújo et al. 2001; Cassells y Curry, 2001). Esta técnica ha sido aplicada

26
para generar variabilidad, especialmente en plantas con propagación vegetativa, como el ajo (Al-Zahim et al., 1999). Nuevos cultivares con alto valor comercial han sido obtenidos también por medio de la variación somaclonal en arroz (Oryza sativa) (Araújo et al., 2001), chile dulce (Capsicum annuum) (Anu et al., 2004), palma aceitera (Elaeis guineensis) (Tregear et al., 2002), crisantemo (Chrysanthemum sp.) (Martín et al., 2002), Hypericum perforatum (planta con propiedades farmacológicas) (Halušková y Košuth, 2003). En los casos mencionados anteriormente, las características agronómicas generadas por la variación somaclonal fueron heredables. Otras variantes somaclonales han sido utilizadas comercialmente por sus características de resistencia a enfermedades, mejora en la apariencia de la planta, flores y frutos, tamaño, sabor, fragancia y presencia de metabolitos de interés. Tal es el ejemplo de variantes somaclonales de fresa (Fragaria x ananassa), resistentes a antracnosis (Hammerschlag et al., 2006). En algunos casos la variación somaclonal es indeseable, cuando el objetivo es mantener la uniformidad y características con valor agronómico (Ahloowalia, 1998; Brar y Brar, 1998; Sahijram et al., 2003). En estos casos es deseable reducir o, al menos, estimar la tasa de variación somaclonal (Gupta y Varshney, 1999). El banano (Musa spp.) es un ejemplo donde la variación somaclonal puede ser no deseable, ya que ocurre frecuentemente durante la propagación clonal en la cual interesa mantener la estabilidad del material (Sahijram et al., 2003). 2.7.2 Detección de la variación somaclonal La variación somaclonal puede ser caracterizada por diversos tipos de marcadores: morfológicos, bioquímicos (isoenzimáticos y proteicos) y moleculares (Sánchez y Jiménez, 2009). Los marcadores morfológicos se basan en las características fenotípicas como: altura, color, forma de las hojas, tipo de inflorescencia, etc. Sin embargo, algunas de estas características varían en respuesta a factores ambientales, bióticos o endógenos, y es necesario que la planta se encuentre en una fase adulta para que sean evaluados (Azofeifa-Delgado, 2006). Los marcadores bioquímicos, fueron los primeros que se utilizaron en la comparación de variedades o clones de plantas, identifican las variantes de una proteína, estas variaciones pueden ser el resultado del polimorfismo de un locus o por modificaciones postraduccionales de la proteína. A pesar de ser una técnica sencilla y rápida, actualmente es poco usada ya que son pocas las enzimas que se pueden emplear como marcadores, ya que como requerimiento estas no deben presentar cambios en su acumulación o composición química en respuesta a factores externos o endógenos. Por lo tanto, aunque sería factible identificar dos variedades de una

27
planta, no se recomienda en estudios somaclonales donde la variación entre una planta madre y los clones podría deberse a una mutación puntual (Azofeifa-Delgado, 2006) Los marcadores moleculares, analizan la variación de la secuencia del ADN entre individuos localizadas en regiones que evidencian polimorfismos (Picca et al., 2004; Azofeifa-Delgado, 2006) y que idealmente son representativos a nivel del genoma completo (Agarwal et al., 2008). En comparación con otro tipo de marcadores, los moleculares son estables y detectables en todos los tejidos sin importar el crecimiento, diferenciación, desarrollo o estatus de defensa de la célula, no son afectados por el ambiente, ni por los efectos pleitrópicos y epistáticos (Picca et al., 2004; Azofeifa-Delgado, 2006; Agarwal et al., 2008). Las técnicas de marcadores moleculares, pueden clasificarse en dos categorías. La primera categoría, se basan en la técnica conocida como hibridación tipo Southern. Esta técnica tiene el propósito de explorar las variaciones en la longitud de los fragmentos del ADN, ocasionados por la restricción del genoma con alguna endonucleasa particular. La segunda categoría, se basan en la amplificación de segmentos de ADN, son los más socorridos debido a la simplicidad, rapidez y requiere poca cantidad de ADN genómico en comparación con otros métodos.
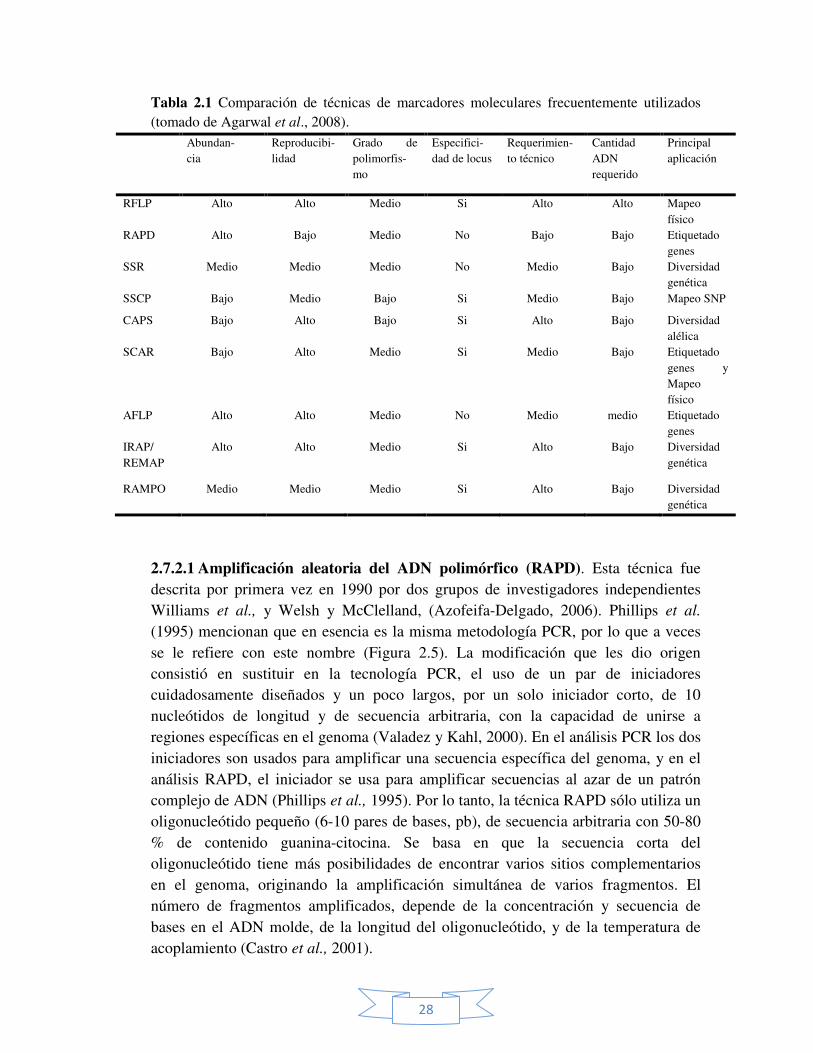
Agarwal et al., (2008), mencionan que desafortunadamente no existe una técnica molecular para cada situación; las técnicas difieren una de otras con respecto a características importantes tales como: abundancia genómica, nivel de polimorfismo detectado, especificidad de locus, reproducibilidad requerimiento técnico y costos (Tabla 2.1) por lo que su aplicación depende de las necesidades. Por lo tanto, la elección de la técnica depende del investigador y de la resolución genética necesitada, así como también del presupuesto con que se cuenta y la experiencia técnica disponible.
28
Tabla 2.1 Comparación de técnicas de marcadores moleculares frecuentemente utilizados (tomado de Agarwal et al., 2008).
Abundan- cia
Reproducibi-lidad
Grado de polimorfis-mo
Especifici-dad de locus
Requerimien-to técnico
Cantidad ADN requerido
Principal aplicación
RFLP Alto Alto Medio Si Alto Alto Mapeo físico
RAPD Alto Bajo Medio No Bajo Bajo Etiquetado genes
SSR Medio Medio Medio No Medio Bajo Diversidad genética
SSCP Bajo Medio Bajo Si Medio Bajo Mapeo SNP
CAPS Bajo Alto Bajo Si Alto Bajo Diversidad alélica
SCAR Bajo Alto Medio Si Medio Bajo Etiquetado genes y Mapeo físico
AFLP Alto Alto Medio No Medio medio Etiquetado genes
IRAP/ REMAP
Alto Alto Medio Si Alto Bajo Diversidad genética
RAMPO Medio Medio Medio Si Alto Bajo Diversidad genética
2.7.2.1 Amplificación aleatoria del ADN polimórfico (RAPD). Esta técnica fue descrita por primera vez en 1990 por dos grupos de investigadores independientes Williams et al., y Welsh y McClelland, (Azofeifa-Delgado, 2006). Phillips et al.
(1995) mencionan que en esencia es la misma metodología PCR, por lo que a veces se le refiere con este nombre (Figura 2.5). La modificación que les dio origen consistió en sustituir en la tecnología PCR, el uso de un par de iniciadores cuidadosamente diseñados y un poco largos, por un solo iniciador corto, de 10 nucleótidos de longitud y de secuencia arbitraria, con la capacidad de unirse a regiones específicas en el genoma (Valadez y Kahl, 2000). En el análisis PCR los dos iniciadores son usados para amplificar una secuencia específica del genoma, y en el análisis RAPD, el iniciador se usa para amplificar secuencias al azar de un patrón complejo de ADN (Phillips et al., 1995). Por lo tanto, la técnica RAPD sólo utiliza un oligonucleótido pequeño (6-10 pares de bases, pb), de secuencia arbitraria con 50-80 % de contenido guanina-citocina. Se basa en que la secuencia corta del oligonucleótido tiene más posibilidades de encontrar varios sitios complementarios en el genoma, originando la amplificación simultánea de varios fragmentos. El número de fragmentos amplificados, depende de la concentración y secuencia de bases en el ADN molde, de la longitud del oligonucleótido, y de la temperatura de acoplamiento (Castro et al., 2001).

Figura 2.5 Esquema que muestra los principalKahl, 2000). Para que ocurra la amplificación con la técnica RAPD, los oligonucleótidos deben alinearse en secuencias repetidas en cadenas opuestas. con la técnica RAPD se denominan marcadores RAPDcualquier cambio en la secuencia o sitio de unión del iniciador (mutación puntual), lo cual impide que el iniciador se una a la cadena, o también pueden ser el producto de cambios que alteren el tamaño o impidan la exitosa amplificación del ADN de la muestra. Como regla general, el tamaño de las variantes se detectan muy escasamente y los productos de la amplificación individuales representan un alelo por locus. En estudios de herencia, los productos se compartan como marcadores dominanteslo tanto, los RAPDs son marcadores que se heredan con naturaleza dominante, es decir, la variación detectada mediante lcada gen (como varios loci del es posible distinguir entre individuos heterocigotos y homlimitación en la información genética obtenidaSosa et al., 2004).
29
Esquema que muestra los principales pasos de la técnica de RAPD
Para que ocurra la amplificación con la técnica RAPD, los oligonucleótidos deben alinearse en secuencias repetidas en cadenas opuestas. Los polimorfismos producidos con la técnica RAPD se denominan marcadores RAPDs, y pueden resultar de cualquier cambio en la secuencia o sitio de unión del iniciador (mutación puntual), lo cual impide que el iniciador se una a la cadena, o también pueden ser el producto de
alteren el tamaño o impidan la exitosa amplificación del ADN de la muestra. Como regla general, el tamaño de las variantes se detectan muy escasamente y los productos de la amplificación individuales representan un alelo por locus. En
los productos se compartan como marcadores dominantesson marcadores que se heredan con naturaleza dominante, es
decir, la variación detectada mediante los mismos no permite distinguircomo varios loci del genoma son amplificados por cada oligonucleótido
es posible distinguir entre individuos heterocigotos y homocigóticos)limitación en la información genética obtenida (Lara, 2005; Azofeifa-Delgado,
es pasos de la técnica de RAPD (Valadez y
Para que ocurra la amplificación con la técnica RAPD, los oligonucleótidos deben Los polimorfismos producidos
, y pueden resultar de cualquier cambio en la secuencia o sitio de unión del iniciador (mutación puntual), lo cual impide que el iniciador se una a la cadena, o también pueden ser el producto de
alteren el tamaño o impidan la exitosa amplificación del ADN de la muestra. Como regla general, el tamaño de las variantes se detectan muy escasamente y los productos de la amplificación individuales representan un alelo por locus. En
los productos se compartan como marcadores dominantes. Por son marcadores que se heredan con naturaleza dominante, es
os mismos no permite distinguir los alelos de son amplificados por cada oligonucleótido, no
ocigóticos), y existe una Delgado, 2006;

30
La técnica RAPD dispone de ventajas considerables respecto a otros marcadores moleculares. Por un lado proporciona un número ilimitado de loci (fragmentos) y el nivel de polimorfismo es considerado. Además, requiere muy poca cantidad de material biológico, por lo que es una técnica no destructiva, ideal en especies amenazadas y puede secarse utilizando gel de sílice después de su colecta en campo, lo cual lo convierte en una técnica muy versátil. No requiere conocimientos previos del genoma de la especie estudiada, y aunque la naturaleza de las secuencias de ADN amplificadas mediante esta técnica se desconoce, la mayor parte de los fragmentos amplificados parecen corresponder a regiones no codificantes, por lo que probablemente son marcadores neutrales. Finalmente, es una técnica rápida, relativamente barata y sencilla de aplicar (Lara, 2005; Sosa et al., 2004). Su desventaja es que el perfil es dependiente de las condiciones de reacción de amplificación, calidad y concentración de ADN, lo que influye en la reproducibilidad de los datos, diferentes condiciones entre laboratorios con pequeñas alteraciones en los parámetros de amplificación pueden dar origen a resultados diferentes (Lara, 2005, Narváez et al., 2000; Agarwal et al., 2008).
2.7.2.2 Análisis genómico utilizando marcadores moleculares RAPDs. La técnica RAPDs ha sido aplicada para evaluar las relaciones filogenéticas, diversidad genética, identificación de cultivar y detección de hibridación en plantas ornamentales como Azaleas y Orquídeas. En Azaleas el perfil RAPDs fue útil para identificar el cultivar y selección del hibrido. En las orquídeas las bandas fueron específicas para subgéneros o especies (Takashi, 1998). Para identificar el locus que controla el tipo de flor en clavel (Scovel et al., 1998). También se han aplicado para evaluar la variabilidad genética en plantas micropropagadas de cultivos agronómicos importantes como Prunus dulcis (Martins, 2004) y estudiar la diversidad genética existente en un grupo de 50 cultivares de vid (Narváez et al., 2000). En el 2008, Taniguchi y colaboradores establecieron un método para la propagación in vitro de Cypripedium macranthos var. Rebunense una orquídea en peligro de extinción localizada en Hokkaido, Japón; a partir de semillas maduras. Este grupo utilizo técnicas moleculares para analizar el proceso de desarrollo con el objetivo de tener una técnica de propagación confiable para tener material vegetal para reintroducir esta orquídea a su hábitat natural. Para el análisis molecular mediante RAPDs, inicialmente utilizaron 20 primers de Operon tecnologies para la amplificación, de los cuales seleccionaron 5: OPM01, 5´-GTTGGTGGCT-3´; OPM06, 5´-CTGGGCAACT-3´; OPN04, 5´-GACCGACCCA-3´; OPM05, 5´-GGGAACGTGT-3´; y OPN01, 5´-CTCACGTTGG-3`. Ellos obtuvieron fragmentos polimórficos al analizar tanto para las plantas obtenidas del cultivo in vitro así como

31
para las especies silvestres por lo que concluyen que las plantas en su hábitat natural son heterogéneas genéticamente y que la introducción de plantas propagadas in vitro con un fondo genético similar al de este hábitat podría ajustarse a la diversidad genética de la población existente. Rout et al. (1998), utilizaron la técnica RAPDs para determinar la estabilidad genética de plantas micropropagadas de Zingiber oficinales Rose (ginger), utilizaron 15 oligonucleótidos de secuencia aleatoria para amplificar el ADN de plantas in vivo e in vitro, los perfiles que obtuvieron de las plantas micropropagadas a partir del cultivo de meristemos fueron monomórficos y similares al de las plantas control; por lo tanto, concluyen que aplicando esta técnica de análisis genético se puede asegurar la estabilidad genética de la plantas derivadas del cultivo in vitro. Otro ejemplo, es el reportado por Bridg (2001), quien evaluó la estabilidad genética de plantas regeneradas in vitro de Annona cherimola y A. muricata, evaluaron 29 iniciadores, de los cuales seleccionaron seis por su patrón de repetición en ambas especies. Los resultados confirmaron la estabilidad genética de las plantas micropropagadas. De Melo et al. (2006), utilizaron el análisis con RAPDs para checar posibles alteraciones genéticas en plantas propagadas in vitro usando yemas axilares en presencia de tidiazuron (TDZ) originadas mediante seis subcultivos Dendrobium Hybrid (Orchidaceae). Ellos probaron 20 primers con los cuales obtuvieron 172 fragmentos, de los cuales entre 5-12 bandas fueron distintas y reproducibles. Durante el cultivo observaron 2 alteraciones fenotípicas: hojas facioladas y elongadas. A pesar de estas alteraciones, el análisis RAPDs no mostró bandas polimórficas entre las plantas que fenotípicamente presentaron algunas de estas alteraciones, lo que indica que la variación detectada no necesariamente corresponde a alteraciones en la secuencia de ADN, de acuerdo con el set de oligonucleótidos utilizados. Por lo tanto, que concluyeron que el protocolo que utilizaron para la propagación clonal de este híbrido fue eficiente y no se observó algún polimorfismo Chen et al. (1998), estudiaron las posibles variaciones morfológicas y genéticas en somaclones de Phalaenopsis True Lady "B79-19" derivados de la inducción de brotes adventicios a partir de tallos florales. Evaluaron 1360 somaclones en floración, no se encontró diferencia aparente en la forma de las hojas, mientras que las flores de algunos somaclones se veían deformes. Seleccionaron 38 cebadores para amplificar el ADN por la técnica molecular RAPDs. Los datos mostraron diferencias genéticas entre los somaclones y las plantas madre.

32

33
III. HIPOTESIS Y OBJETIVOS.

34

35
3.1 HIPOTESIS Si durante la inducción morfogénica se presento la variación somaclonal entonces, el perfil genético de las estructuras morfogénicas obtenidas y las plántulas madre de Bletia purpurea (Lam.) presentan diferencia en los patrones de bandeo.
3.2 OBJETIVO GENERAL. Estudiar mediante marcadores RAPDs la posible variación somaclonal entre las plántulas madre y las estructuras obtenidas de la inducción morfogénica in vitro de Bletia purpurea (Lam.). 3.3 OBJETIVOS ESPECÍFICOS. Optimización de la concentración de la auxina (Acido Indol Acético) y la citocinina (Bencil Amino Purina) sobre la morfogénesis in vitro de Bletia purpurea (Lam.). Validación de las ecuaciones obtenidas para el número de brotes, hojas y raíces en la morfogénesis de Bletia purpurea (Lam.). Obtener los perfiles genéticos de la plántula madre y de las estructuras obtenidas del tratamiento óptimo validado para la inducción morfogénica de Bletia purpurea, por RAPDs.

36

37
IV. MATERIALES Y MÉTODOS.

38
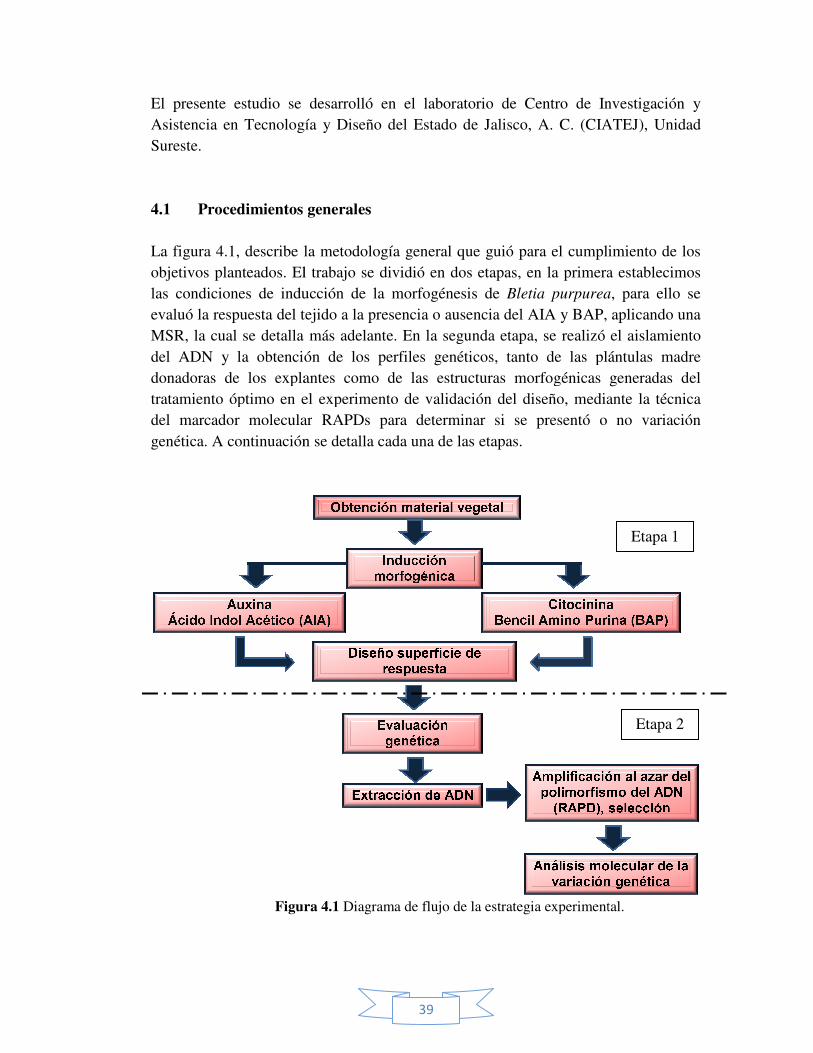
El presente estudio se desarrollóAsistencia en Tecnología y Diseño del Estado de Jalisco, A. C. (CIATEJSureste.
4.1 Procedimientos generales La figura 4.1, describe la metodología generalobjetivos planteados. El trabajo las condiciones de inducción de la morfogénesisevaluó la respuesta del tejido a la presencia o MSR, la cual se detalla del ADN y la obtención de losdonadoras de los explantes tratamiento óptimo en el experimento de validación del diseño, del marcador moleculargenética. A continuación
Figura 4
39
El presente estudio se desarrolló en el laboratorio de Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco, A. C. (CIATEJ
Procedimientos generales
escribe la metodología general que guió para el cumplimiento de los El trabajo se dividió en dos etapas, en la primera
inducción de la morfogénesis de Bletia purpurea
evaluó la respuesta del tejido a la presencia o ausencia del AIA y BAP, , la cual se detalla más adelante. En la segunda etapa, se realizó el aislamiento
la obtención de los perfiles genéticos, tanto de las plántulasdonadoras de los explantes como de las estructuras morfogénicas
óptimo en el experimento de validación del diseño, medianteel marcador molecular RAPDs para determinar si se presentó o no
A continuación se detalla cada una de las etapas.
ura 4.1 Diagrama de flujo de la estrategia experimental
en el laboratorio de Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco, A. C. (CIATEJ), Unidad
l cumplimiento de los la primera establecimos
Bletia purpurea, para ello se aplicando una
se realizó el aislamiento plántulas madre
de las estructuras morfogénicas generadas del mediante la técnica
terminar si se presentó o no variación
experimental.
Etapa 1
Etapa 2

40
4.2 Establecimiento de las condiciones para la inducción morfogénica de B.
purpurea (Lam.) 4.2.1 Material vegetal inicial Las plántulas que se utilizaron para los experimentos fueron micropropagadas a partir de la germinación in vitro (López Puc, 2010 datos no publicados) quien germinó las semillas in vitro. Las vitroplantas utilizadas fueron de 7 meses de edad, seleccionando aquellas de tamaño uniforme a partir de las cuales se obtuvieron diferentes tejidos: hojas, tallos y pseudobulbos. 4.2.2 Medios y condiciones de cultivo Para la inducción morfogénica in vitro de B. purpurea el medio de cultivo utilizado fue el MS (Murashige y Skoog, 1962), suplementado con mioinositol (100 mg L-1), niacina (0.5 mg L-1), piridoxina-HCl (0.5 mg L-1), tiamina-HCl (0.1 mg L-1), glicina (2 mg L-1), peptona (1 mg L-1), fosfato de sodio (170 mg L-1), sacarosa 20 %, 2 g L-1 de carbón activado y solidificados con 2.2 g L-1 de gel rite (Phytagel®). Los RCV fueron añadidos al medio de acuerdo a los tratamientos de la MSR. El pH fue ajustado a 5.7 (con NaOH o HCl al 1 N). Se dosificaron 20 mL del medio de cultivo en tubos de ensaye PYREX® de 25 x 150 mm Rimless. El medio fue esterilizado a 121 ºC a 1.5 atmósferas de presión durante 15 min. (Nayak et al., 1997; Chen et al., 2002b; Su et al., 2006; Gow et al., 2008).
Los tubos de cada tratamiento con la muestra se cultivaron a 23 ± 2 ºC bajo una lámpara de luz blanca fría fluorescente de 40 W con fotoperiodo 16/8 hrs de luz/oscuridad (Nayak et al., 1997; Wu et al., 2004). La evaluación de los cultivos durante la inducción morfogénica, se realizó a los 15, 30 y 45 días de cultivo. A los 45 días de la inducción se realizó la evaluación porque los brotes ya se encontraban más desarrollados y se procedió al análisis estadístico de los datos. 4.3 Diseño experimental Para el análisis estadístico, se eligió la MSR para optimizar las concentraciones de los RCV en la inducción morfogénica de B. purpurea. Primero, se realizaron unos ensayos preliminares para seleccionar las variables del cultivo y se definieron los
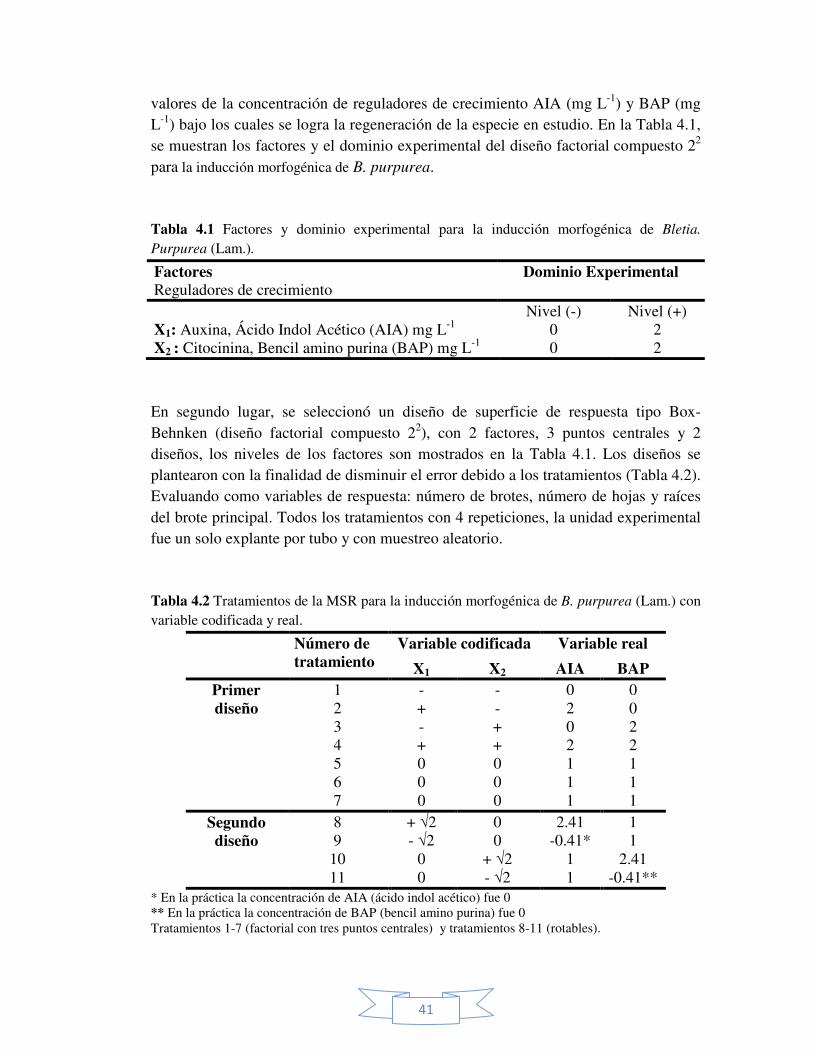
41
valores de la concentración de reguladores de crecimiento AIA (mg L-1) y BAP (mg L-1) bajo los cuales se logra la regeneración de la especie en estudio. En la Tabla 4.1, se muestran los factores y el dominio experimental del diseño factorial compuesto 22 para la inducción morfogénica de B. purpurea. Tabla 4.1 Factores y dominio experimental para la inducción morfogénica de Bletia.
Purpurea (Lam.).
Factores Reguladores de crecimiento
Dominio Experimental
Nivel (-) Nivel (+) X1: Auxina, Ácido Indol Acético (AIA) mg L-1 0 2 X2 : Citocinina, Bencil amino purina (BAP) mg L-1 0 2
En segundo lugar, se seleccionó un diseño de superficie de respuesta tipo Box-Behnken (diseño factorial compuesto 22), con 2 factores, 3 puntos centrales y 2 diseños, los niveles de los factores son mostrados en la Tabla 4.1. Los diseños se plantearon con la finalidad de disminuir el error debido a los tratamientos (Tabla 4.2). Evaluando como variables de respuesta: número de brotes, número de hojas y raíces del brote principal. Todos los tratamientos con 4 repeticiones, la unidad experimental fue un solo explante por tubo y con muestreo aleatorio. Tabla 4.2 Tratamientos de la MSR para la inducción morfogénica de B. purpurea (Lam.) con variable codificada y real.
Número de tratamiento
Variable codificada Variable real
X1 X2 AIA BAP Primer diseño
1 - - 0 0 2 + - 2 0 3 - + 0 2 4 + + 2 2 5 0 0 1 1 6 0 0 1 1 7 0 0 1 1
Segundo diseño
8 + √2 0 2.41 1 9 - √2 0 -0.41* 1
10 0 + √2 1 2.41 11 0 - √2 1 -0.41**
* En la práctica la concentración de AIA (ácido indol acético) fue 0 ** En la práctica la concentración de BAP (bencil amino purina) fue 0 Tratamientos 1-7 (factorial con tres puntos centrales) y tratamientos 8-11 (rotables).

42
Los experimentos del primer diseño fueron útiles para verificar su ajuste al modelo lineal. El análisis de regresión y el ANOVA revelaron un valor de p mayor a 0.05, por lo tanto, se aceptó la hipótesis nula (Ho) que afirma: no existe un ajuste al modelo lineal. No habiendo ajuste a lineal, se realizaron los experimentos del segundo diseño (Tabla 4.2). La regresión de segundo orden, se llevó a cabo considerando los tratamientos probados en los dos diseños para verificar si se estaba en la zona óptima. Una vez hallando ajuste a modelo cuadrático, se realizó un análisis canónico para determinar los valores óptimos. Finalmente, se generaron los contornos y la superficie de respuesta para cada variable de respuesta. Estos pueden tener diferentes formas dependiendo de la relación hallada entre las variables X1 y X2. Los valores óptimos fueron llevados a experimentación para verificar el ajuste del modelo. El análisis de resultados se realizó auxiliándose del software para análisis estadísticos StatGraphics Centurion XV.1, obteniéndose de este el ajuste a comportamiento lineal y/o cuadrático. El análisis molecular fue realizado al tratamiento óptimo del experimento realizado para la validación del diseño experimental, seleccionando con la ayuda del programa estadístico y tomando un 99 % de confianza, los tubos y el número de plántulas a evaluar.
4.4 Estandarización de las técnicas para el estudio molecular 4.4.1 Extracción de ADN Para estandarizar la técnica de extracción del ADN se utilizó como material vegetal brotes germinados in vitro de Spathoglottis plicata. Se evaluaron cuatro protocolos reportados previamente (Allen et al., 2006; Saghai y Moroof et al., 1984; Zambrano et al., 2002; Li et al., 2007). Se eligió el protocolo reportado por Allen et al., 2006, basado en el método Bromuro de hexadeciltrimetilamonio (CTAB) La muestra de 200 a 400 mg de tejido se maceraron sobre un mortero con un volumen del buffer de extracción (2 % CTAB, 1 M Tris base pH 8,0, 0.5 M EDTA, 5 M NaCl y 1 % ME), el macerado se colocó en tubos eppendorf de 1.5 mL, se agitaron por inversión y se incubaron a 65 ºC durante 30 min con agitación cada 10 min. Posteriormente se centrifugó a 13500 rpm durante 10 min y a cada tubo se le adicionó un volumen igual de Fenol/Cloroformo/Alcohol isoamílico (25:24:1), se mezcló por inversión, se dejó reposar por 20 min y se centrifugó. Transcurrido el tiempo de la centrifugación, la fase acuosa fue separada con micropipeta y transferida a tubo

43
nuevo eppendorf. Se repitió por segunda vez la fase de lavado con Fenol/Cloroformo/Alcohol isoamílico (25:24:1). Después, a la fase acuosa se le adicionó un volumen de isopropanol frío, se mezcló e incubó por 10 min a temperatura ambiente. Luego de centrifugar se decantó y al pellet se le añadió 250 µL de TE (Tris 2 M y EDTA 0.5 M) y 2 µL RNAsa, se incubó a 37 ºC por 30 min. Luego, se adicionó 25 µL NaOAc 3 M y EtOH al 100 % frío e incubó a -20 ºC por 20 min. Los lavados se realizaron con EtOH al 70 %. Por último, las muestras se dejaron secar a temperatura ambiente y se resuspendieron en agua destilada estéril. Se almacenaron a -20 ºC para su posterior uso. Para estimar la concentración final de ADN genómico, se realizó una dilución 1:1000 (muestra/agua destilada) y se leyó en un espectrofotómetro UV-Vis (Eppendorf biophotometer) a densidades ópticas (DO) 260 y 280 nm. La calidad del ADN extraído fue verificada realizando una electroforesis en geles de agarosa al 1 % en amortiguador TAE 1X (Tris base 40 mM, NaOAc 5 mM. EDTA 1 mM pH 8), a 80 Volts durante 45 min. Teñido con bromuro de etidio (0.5 µg mL-1) y visualizado en el sistema fotodocumentador de geles marca UVP modelo DIGIDOC-IT. 4.4.2 Amplificación al azar del polimorfismo del ADN (RAPD) 4.4.2.1 Selección de oligonucleótidos. Como primer paso se realizó un escrutinio para identificar aquellos oligonucleótidos capaces de amplificar en el genoma de Spathoglottis plicata. Para ello se empleó el protocolo descrito por Williams et al. (1990) con algunas modificaciones propuestas por Scovel et al. (1998). Se evaluó un kit de 20 oligonucleótidos de Operon Technologies, Inc. (Tabla 4.3), utilizando un oligonucleótido por reacción. De aquellos oligonucleótidos que amplificaron se seleccionaron 5 para la estandarización de la técnica RAPD. Para estandarizar la técnica se evaluaron dos concentraciones de los oligonucleótidos (25 y 50 ng) y diferentes concentraciones de ADN (20, 25, 35, 100, 150 y 200 ng); así como tres temperaturas de hibridación (35, 40 y 45 ºC). Las condiciones finales se utilizaron para la selección de oligonucleótidos para la especie en estudio.
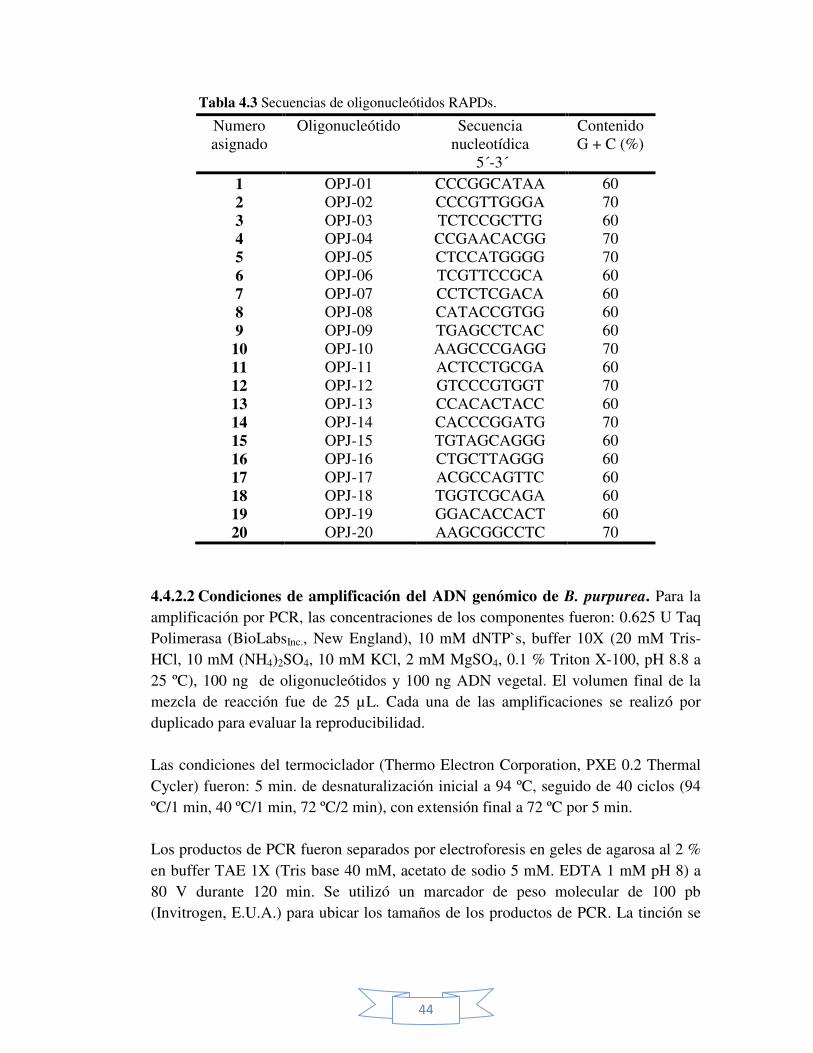
44
Tabla 4.3 Secuencias de oligonucleótidos RAPDs.
Numero asignado
Oligonucleótido Secuencia nucleotídica
5´-3´
Contenido G + C (%)
1 OPJ-01 CCCGGCATAA 60 2 OPJ-02 CCCGTTGGGA 70 3 OPJ-03 TCTCCGCTTG 60 4 OPJ-04 CCGAACACGG 70 5 OPJ-05 CTCCATGGGG 70 6 OPJ-06 TCGTTCCGCA 60 7 OPJ-07 CCTCTCGACA 60 8 OPJ-08 CATACCGTGG 60 9 OPJ-09 TGAGCCTCAC 60
10 OPJ-10 AAGCCCGAGG 70 11 OPJ-11 ACTCCTGCGA 60 12 OPJ-12 GTCCCGTGGT 70 13 OPJ-13 CCACACTACC 60 14 OPJ-14 CACCCGGATG 70 15 OPJ-15 TGTAGCAGGG 60 16 OPJ-16 CTGCTTAGGG 60 17 OPJ-17 ACGCCAGTTC 60 18 OPJ-18 TGGTCGCAGA 60 19 OPJ-19 GGACACCACT 60 20 OPJ-20 AAGCGGCCTC 70
4.4.2.2 Condiciones de amplificación del ADN genómico de B. purpurea. Para la amplificación por PCR, las concentraciones de los componentes fueron: 0.625 U Taq Polimerasa (BioLabsInc., New England), 10 mM dNTP`s, buffer 10X (20 mM Tris-HCl, 10 mM (NH4)2SO4, 10 mM KCl, 2 mM MgSO4, 0.1 % Triton X-100, pH 8.8 a 25 ºC), 100 ng de oligonucleótidos y 100 ng ADN vegetal. El volumen final de la mezcla de reacción fue de 25 µL. Cada una de las amplificaciones se realizó por duplicado para evaluar la reproducibilidad.
Las condiciones del termociclador (Thermo Electron Corporation, PXE 0.2 Thermal Cycler) fueron: 5 min. de desnaturalización inicial a 94 ºC, seguido de 40 ciclos (94 ºC/1 min, 40 ºC/1 min, 72 ºC/2 min), con extensión final a 72 ºC por 5 min. Los productos de PCR fueron separados por electroforesis en geles de agarosa al 2 % en buffer TAE 1X (Tris base 40 mM, acetato de sodio 5 mM. EDTA 1 mM pH 8) a 80 V durante 120 min. Se utilizó un marcador de peso molecular de 100 pb (Invitrogen, E.U.A.) para ubicar los tamaños de los productos de PCR. La tinción se

45
realizó con bromuro de etidio (0.5 µg mL-1), finalmente fueron visualizados en el sistema fotodocumentador de geles marca UVP modelo DIGIDOC-IT. La evaluación se realizó mediante la observación de las fotografías de los geles RAPDs, en los cuales cada individuo tiene un patrón de bandeo específico. Las bandas con el mismo peso molecular en distintos individuos madre e hijo pueden ser asumidas como idénticas.

46

47
V. RESULTADOS Y DISCUSIÓN

48

49
5.1 Establecimiento de las condiciones para la inducción morfogénica in vitro de B. purpurea (Lam.) Inicialmente para la inducción morfogénica, se realizó un experimento preliminar tomando segmentos de hojas de 1 cm2 en los cuales no se observó formación de brotes, ni callos, y al paso del tiempo en estos se vio un cambio en la coloración del verde al pardo obscuro. Este resultado nos hizo suponer que el explante de hoja de B.
purpurea desprende compuestos fenólicos en cultivo; por lo que se tomó la decisión de añadir carbón activado para el montaje de los experimentos finales. Dado que es uno de los compuestos utilizados en el cultivo para contrarrestar el efecto de los compuestos orgánicos e inorgánicos, contenidos tanto en el medio de cultivo como los liberados por las mismos explantes (Pierik, 1990). Además, Margara (1998), afirma que el carbón activado administrado en bajas concentraciones a los medios de cultivo, en dosis que pueden variar desde los 0.5-5 g L-1, estimula los procesos de desarrollo morfogénico en diferentes orquídeas. Los reportes de Vij et al. (1994) y Chen et al. (1999) observaron que el carbón activado mejora el desarrollo tanto de protocormos como de plántulas, especialmente a concentraciones inferiores a 2 g L-1 en el desarrollo de plántulas de Dendrobium (Zhou et al., 1995). Sharma et al. (2007), mencionan que en el cultivo de tejidos existe el riesgo de contaminación lo cual puede afectar, en particular en los casos en los que se busca la conservación del germoplasma elite de algunas plantas raras o en peligro de extinción. En este trabajo, solo un 2 % de los explantes se perdió debido a la contaminación a las tres semanas de haberse iniciado el cultivo. Para el primer diseño experimental (tratamientos 1-7), se evaluaron diferentes tejidos hojas, tallos y pseudobulbos. La respuesta morfogénica de los tratamientos fue evaluado a los 15, 30 y 45 días de cultivo, las variables de respuesta evaluadas fueron número de brotes, hojas y raíces. A los 15 días de haber iniciado los experimentos para la inducción de la morfogénesis, se observó que solamente en los explantes de pseudobulbos de B. purpurea se formaron brotes directamente sin la formación de callos (Figura 5.1 a) por lo que concluimos que se logro la organogénesis directa. De forma similar Sharma et al., 2007; Sunitibala y Rajkumar, 2009 y Mata et al., 2010 obtuvieron organogénesis directa a partir de pseudobulbos en Dendrobium microbulbon A. Rich, Dendrobium transparens y Lycaste aromatica respectivamente.

Figura 5.1 Organogénesis de morfogénica, formación directa de brotes sobre explante de pseudobulboesteroscopio. b) plántulas regeneradas a los 30 días de cultivo. días de cultivo con raíz y hojas bien desarrolladas.
Una vez inducida la formación dedesarrollo a plántula fue observándose al paso de los díasFigura 5.1 b a los 30 días explante como plántula completa con hojas y raíces. experimentos con hojas y tallos se verde al amarillo o pardo obscuro respectivamente. La regeneración de plantas reportada en la literatura para diferentes especies tejidos de la planta donadora del explante (2000; Tokuhara y Mii, 2001; ChenJanarthanam y Seshadri, 2008los pseudobulbos son los que mostraron una respuesta Para la obtención de la respuesta un factor crucial es la elección deltamaño, la edad y estado fisiológico de los órganos y células de la planta madre son importantes para la competencia de regeneración Narmatha, 2006). El contenido de reguladores de crecimiento endógenos sonliberación de algunas rutas metabólicas implicadas en el desbloqueo y funcionamiento de procesos involucrados en la regeneración (ha sido documentado que la morfogénesis exógenos. Los reguladores de crecimientoregulación de señales del desarrollo. Las auxinas y las citocininas son los principales RCVs involucrados en la regulación de
50
Organogénesis de Bletia purpurea (Lam.). a) a los 15 días de la inducción morfogénica, formación directa de brotes sobre explante de pseudobulbo
plántulas regeneradas a los 30 días de cultivo. c) Plántula regenerada a los 60 días de cultivo con raíz y hojas bien desarrolladas.
la formación de los brotes en los explantes de los pseudobulbos, su desarrollo a plántula fue observándose al paso de los días, como se muestra en la
b a los 30 días y a los 60 días (Figura 5.1 c) ya se pudo separar del como plántula completa con hojas y raíces. Los explantes
experimentos con hojas y tallos se cultivaron durante 60 días y el tejido paso del verde al amarillo o pardo obscuro respectivamente.
La regeneración de plantas utilizando las técnicas de cultivo de tejidos ha sido reportada en la literatura para diferentes especies de orquídeas a partir de diferentes tejidos de la planta donadora del explante (Nayak et al., 1997; Sheelavantmath
Tokuhara y Mii, 2001; Chen et al., 2002a, b; Su et al., 2006; GowJanarthanam y Seshadri, 2008). De los diferentes explantes probados en este estudio, los pseudobulbos son los que mostraron una respuesta organogénica.
Para la obtención de la respuesta un factor crucial es la elección deltamaño, la edad y estado fisiológico de los órganos y células de la planta madre son
competencia de regeneración del tejido (Bajaj, 1991
El contenido de reguladores de crecimiento endógenos son un factor que regula la liberación de algunas rutas metabólicas implicadas en el desbloqueo y funcionamiento de procesos involucrados en la regeneración (Bajaj, 1991)ha sido documentado que la morfogénesis in vitro es estimulada por
reguladores de crecimiento son los candidatos más viables en la regulación de señales del desarrollo. Las auxinas y las citocininas son los principales
involucrados en la regulación de la división y diferenciación celular (Lee
días de la inducción
morfogénica, formación directa de brotes sobre explante de pseudobulbo, fotografía de Plántula regenerada a los 60
en los explantes de los pseudobulbos, su , como se muestra en la
ya se pudo separar del Los explantes de los
60 días y el tejido paso del
cultivo de tejidos ha sido orquídeas a partir de diferentes
Sheelavantmath et al., ., 2006; Gow et al., 2008;
De los diferentes explantes probados en este estudio,
Para la obtención de la respuesta un factor crucial es la elección del explante: el tamaño, la edad y estado fisiológico de los órganos y células de la planta madre son
1991; Basker y
un factor que regula la liberación de algunas rutas metabólicas implicadas en el desbloqueo y
Bajaj, 1991). También es estimulada por los RCV
son los candidatos más viables en la regulación de señales del desarrollo. Las auxinas y las citocininas son los principales
la división y diferenciación celular (Lee et al.,

51
2007). Su efecto sobre la respuesta fue vista en este trabajo. A pesar, que no fue muy marcado en el número de brotes es aceptable ya que en este tipo de respuesta organogénica es un poco más limitado el número obtenido que por otras vías (Bajaj, 1991). En el caso, específico de las orquídeas se ha reportado que el uso de citocininas contribuye a tener una mejor respuesta morfogénica, ya sea que se utilicen solas (Kuo et al., 2005; Gow et al., 2008; Hong et al., 2010; Mata et al., 2010) o combinadas con alguna auxina (Lee et al., 2007, Nayak et. al. 1998; Janarthanam y Seshadri, 2008; Sunitibala y Rajkumar, 2009). Sin embargo, en este estudio de acuerdo, al diseño experimental planteado el tratamiento con auxinas fue mejor. Es conocido, que las citocininas son sintetizadas en las raíces y transportados a las partes aéreas de la planta (Hew y Clifford, 1993). De aquí, podemos inferir que el contenido endógeno de citocininas en el pseudobulbo es mayor y por lo tanto, la adición exógena de auxina fue necesaria para el equilibrio hormonal. Sunitibala y Rajkumar (2009) y Mata et al. (2010), asumen que los brotes múltiples generados en su estudio posiblemente fueron derivados de los brotes axilares presentes en los segmentos de los pseudobulbos y que en las hojas y tallos cercanos a las hojas no se observó formación de brotes, lo que indica que en esta parte del tallo no queda ningún resto del meristemo apical, sino solamente envoltura foliar. Por lo tanto, la capacidad de los pseudobulbos de B. purpurea para regenerar brotes múltiples puede estar asociado a los brotes axilares presentes en los pseudobulbos influenciados por las concentración de los reguladores de crecimiento y no fue afectado por la edad de la planta madre de donde se tomaron los explantes. Por lo cual, se sugiere que el cultivo de pseudobulbos pueden ser empleados para la rápida inducción ajustando adecuadamente los nutrientes y los reguladores de crecimiento. 5.2 Diseño experimental En este estudio se encontró que el desarrollo directo de brotes sobre el explante de pseudobulbo fue promovido por las diferentes concentraciones de AIA y BAP (0-2 mg L-1) solos o en combinación. Se obtuvo de 1-3 brotes en promedio por explante, una altura del brote de 50.32-69.84 mm, con 3-4 hojas y 1-2 raíces (Tabla 5.1). Sharma et al. (2007), reportan que obtuvieron de 8-9 brotes por explante de pseudobulbo en Dendrobium con 7.5 mg L-1 de AIA y 20 mg L-1 de BAP y una longitud de brote de 0.5 cm. Por su parte, Sunitibala y Kishor (2009) obtuvieron 7 brotes por explante de pseudobulbo de D. transparens L. con la combinación de BAP (2 mg L-1) y ANA (1 mg L-1). Mata el al. (2010), con 4.4, 8.87 y 2.2 µM de TDZ
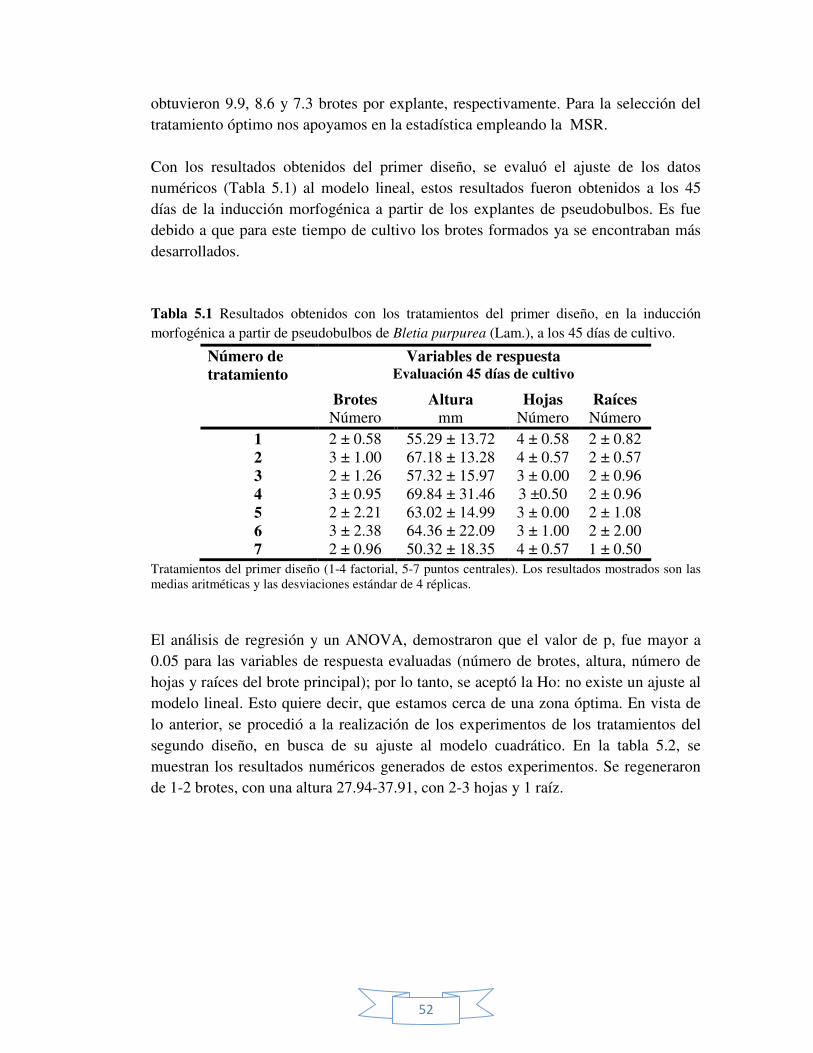
52
obtuvieron 9.9, 8.6 y 7.3 brotes por explante, respectivamente. Para la selección del tratamiento óptimo nos apoyamos en la estadística empleando la MSR. Con los resultados obtenidos del primer diseño, se evaluó el ajuste de los datos numéricos (Tabla 5.1) al modelo lineal, estos resultados fueron obtenidos a los 45 días de la inducción morfogénica a partir de los explantes de pseudobulbos. Es fue debido a que para este tiempo de cultivo los brotes formados ya se encontraban más desarrollados. Tabla 5.1 Resultados obtenidos con los tratamientos del primer diseño, en la inducción morfogénica a partir de pseudobulbos de Bletia purpurea (Lam.), a los 45 días de cultivo.
Número de tratamiento
Variables de respuesta Evaluación 45 días de cultivo
Brotes Altura Hojas Raíces Número mm Número Número
1 2 ± 0.58 55.29 ± 13.72 4 ± 0.58 2 ± 0.82 2 3 ± 1.00 67.18 ± 13.28 4 ± 0.57 2 ± 0.57 3 2 ± 1.26 57.32 ± 15.97 3 ± 0.00 2 ± 0.96 4 3 ± 0.95 69.84 ± 31.46 3 ±0.50 2 ± 0.96 5 2 ± 2.21 63.02 ± 14.99 3 ± 0.00 2 ± 1.08 6 3 ± 2.38 64.36 ± 22.09 3 ± 1.00 2 ± 2.00 7 2 ± 0.96 50.32 ± 18.35 4 ± 0.57 1 ± 0.50
Tratamientos del primer diseño (1-4 factorial, 5-7 puntos centrales). Los resultados mostrados son las medias aritméticas y las desviaciones estándar de 4 réplicas.
El análisis de regresión y un ANOVA, demostraron que el valor de p, fue mayor a 0.05 para las variables de respuesta evaluadas (número de brotes, altura, número de hojas y raíces del brote principal); por lo tanto, se aceptó la Ho: no existe un ajuste al modelo lineal. Esto quiere decir, que estamos cerca de una zona óptima. En vista de lo anterior, se procedió a la realización de los experimentos de los tratamientos del segundo diseño, en busca de su ajuste al modelo cuadrático. En la tabla 5.2, se muestran los resultados numéricos generados de estos experimentos. Se regeneraron de 1-2 brotes, con una altura 27.94-37.91, con 2-3 hojas y 1 raíz.
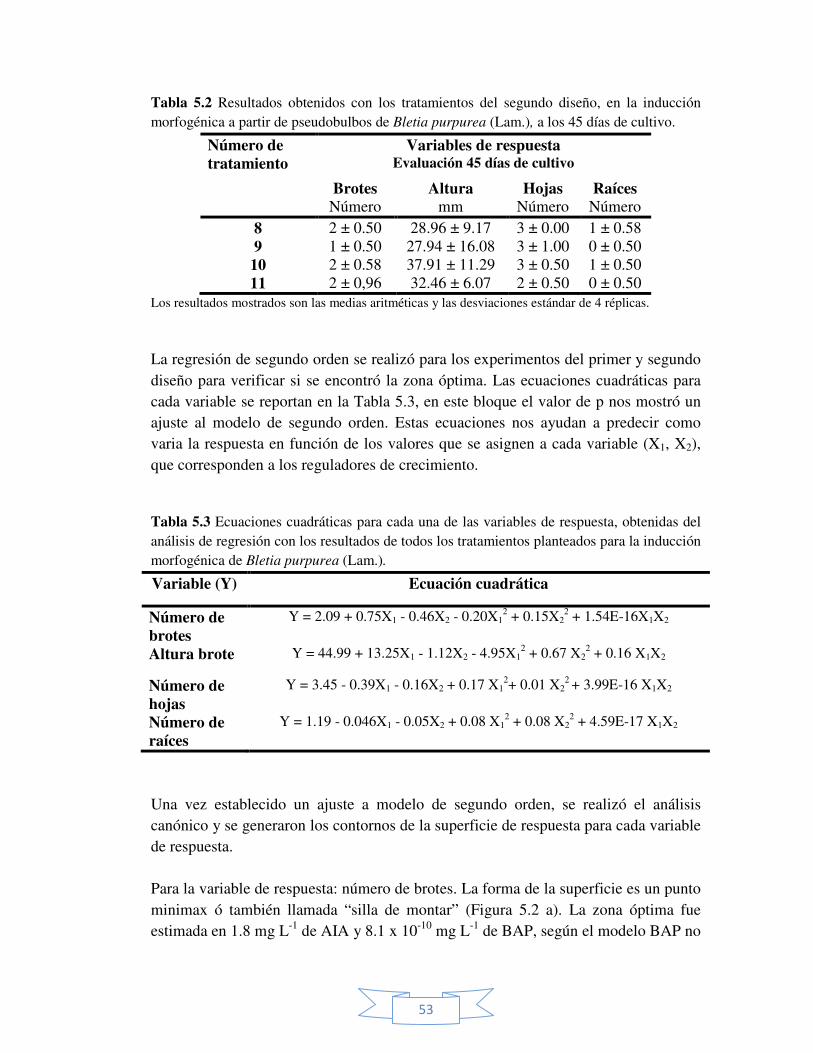
53
Tabla 5.2 Resultados obtenidos con los tratamientos del segundo diseño, en la inducción morfogénica a partir de pseudobulbos de Bletia purpurea (Lam.), a los 45 días de cultivo.
Número de tratamiento
Variables de respuesta Evaluación 45 días de cultivo
Brotes Altura Hojas Raíces Número mm Número Número
8 2 ± 0.50 28.96 ± 9.17 3 ± 0.00 1 ± 0.58 9 1 ± 0.50 27.94 ± 16.08 3 ± 1.00 0 ± 0.50
10 2 ± 0.58 37.91 ± 11.29 3 ± 0.50 1 ± 0.50 11 2 ± 0,96 32.46 ± 6.07 2 ± 0.50 0 ± 0.50
Los resultados mostrados son las medias aritméticas y las desviaciones estándar de 4 réplicas. La regresión de segundo orden se realizó para los experimentos del primer y segundo diseño para verificar si se encontró la zona óptima. Las ecuaciones cuadráticas para cada variable se reportan en la Tabla 5.3, en este bloque el valor de p nos mostró un ajuste al modelo de segundo orden. Estas ecuaciones nos ayudan a predecir como varia la respuesta en función de los valores que se asignen a cada variable (X1, X2), que corresponden a los reguladores de crecimiento. Tabla 5.3 Ecuaciones cuadráticas para cada una de las variables de respuesta, obtenidas del análisis de regresión con los resultados de todos los tratamientos planteados para la inducción morfogénica de Bletia purpurea (Lam.).
Variable (Y) Ecuación cuadrática
Número de brotes
Y = 2.09 + 0.75X1 - 0.46X2 - 0.20X12 + 0.15X2
2 + 1.54E-16X1X2
Altura brote Y = 44.99 + 13.25X1 - 1.12X2 - 4.95X12 + 0.67 X2
2 + 0.16 X1X2
Número de hojas
Y = 3.45 - 0.39X1 - 0.16X2 + 0.17 X12+ 0.01 X2
2 + 3.99E-16 X1X2
Número de raíces
Y = 1.19 - 0.046X1 - 0.05X2 + 0.08 X12 + 0.08 X2
2 + 4.59E-17 X1X2
Una vez establecido un ajuste a modelo de segundo orden, se realizó el análisis canónico y se generaron los contornos de la superficie de respuesta para cada variable de respuesta. Para la variable de respuesta: número de brotes. La forma de la superficie es un punto minimax ó también llamada “silla de montar” (Figura 5.2 a). La zona óptima fue estimada en 1.8 mg L-1 de AIA y 8.1 x 10-10 mg L-1 de BAP, según el modelo BAP no
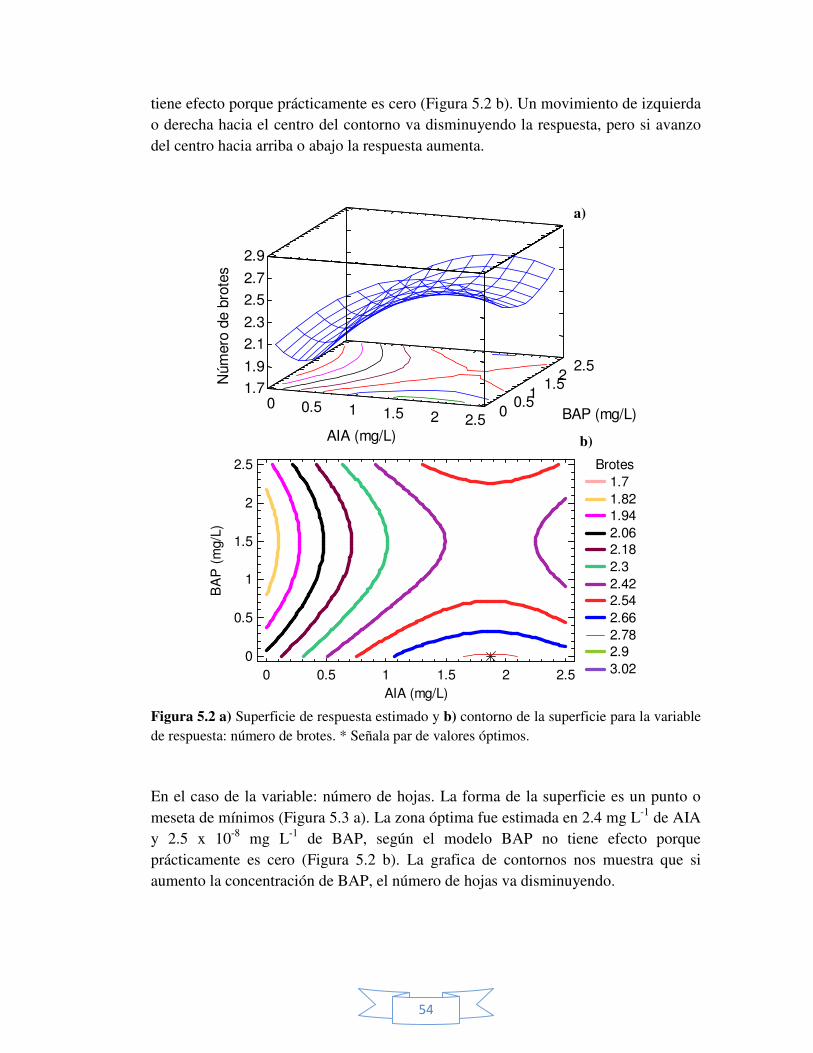
54
tiene efecto porque prácticamente es cero (Figura 5.2 b). Un movimiento de izquierda o derecha hacia el centro del contorno va disminuyendo la respuesta, pero si avanzo del centro hacia arriba o abajo la respuesta aumenta.
Figura 5.2 a) Superficie de respuesta estimado y b) contorno de la superficie para la variable de respuesta: número de brotes. * Señala par de valores óptimos.
En el caso de la variable: número de hojas. La forma de la superficie es un punto o meseta de mínimos (Figura 5.3 a). La zona óptima fue estimada en 2.4 mg L-1 de AIA y 2.5 x 10-8 mg L-1 de BAP, según el modelo BAP no tiene efecto porque prácticamente es cero (Figura 5.2 b). La grafica de contornos nos muestra que si aumento la concentración de BAP, el número de hojas va disminuyendo.
AIA (mg/L)
BAP (mg/L)
Núm
ero
de b
rote
s
0 0.5 1 1.5 2 2.50
0.51
1.52
2.5
1.7
1.9
2.1
2.3
2.5
2.7
2.9
AIA (mg/L)
BA
P (
mg/L
)
0 0.5 1 1.5 2 2.5
0
0.5
1
1.5
2
2.5 Brotes
1.7
1.82
1.94
2.06
2.18
2.3
2.42
2.54
2.66
2.78
2.9
3.02
a)
b)
55
Figura 5.3 a) Superficie de respuesta estimado y b) contorno de la superficie para la variable de respuesta: número de hojas. * Señala par de valores óptimos. Para la variable: número de raíces. La forma de la superficie al igual que para la variable número de hojas es una meseta de mínimos (Figura 5.4 a). La zona óptima fue estimada en 2.4 mg L-1 de AIA y 2.4 mg L-1 de BAP (Figura 5.4 b). Un movimiento hacia el centro del contorno va disminuyendo la respuesta, pero si aumento las concentraciones a la par, la respuesta va en aumento. Sin embargo, esto en la práctica no sería viable dado que se aumentan los costos por la cantidad de RCV empleados.
BAP (mg/L)
AIA (mg/L)
Núm
ero
de h
oja
s
0 0.5 1 1.5 2 2.50
0.51
1.52
2.5
2.8
3
3.2
3.4
3.6
AIA (mg/L)
BA
P (
mg/L
)
0 0.5 1 1.5 2 2.5
0
0.5
1
1.5
2
2.5 Hojas
2.8
2.88
2.96
3.04
3.12
3.2
3.28
3.36
3.44
3.52
3.6
3.68
b)
a)
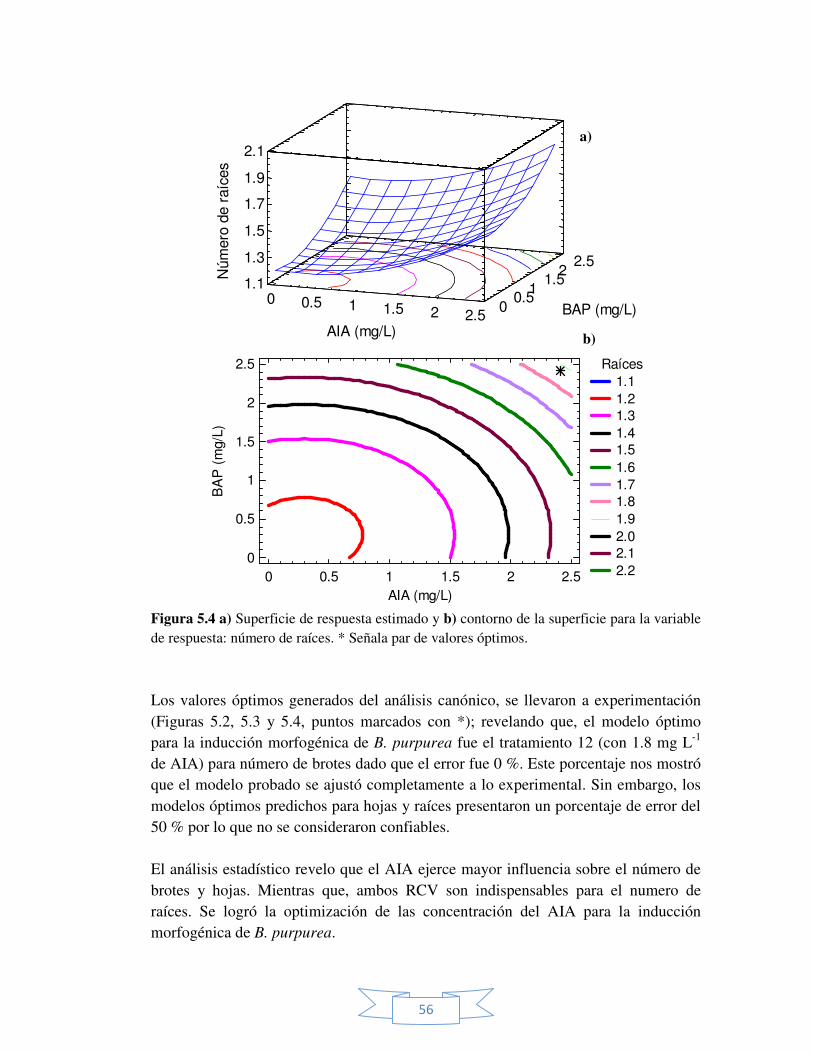
56
Figura 5.4 a) Superficie de respuesta estimado y b) contorno de la superficie para la variable de respuesta: número de raíces. * Señala par de valores óptimos.
Los valores óptimos generados del análisis canónico, se llevaron a experimentación (Figuras 5.2, 5.3 y 5.4, puntos marcados con *); revelando que, el modelo óptimo para la inducción morfogénica de B. purpurea fue el tratamiento 12 (con 1.8 mg L-1 de AIA) para número de brotes dado que el error fue 0 %. Este porcentaje nos mostró que el modelo probado se ajustó completamente a lo experimental. Sin embargo, los modelos óptimos predichos para hojas y raíces presentaron un porcentaje de error del 50 % por lo que no se consideraron confiables. El análisis estadístico revelo que el AIA ejerce mayor influencia sobre el número de brotes y hojas. Mientras que, ambos RCV son indispensables para el numero de raíces. Se logró la optimización de las concentración del AIA para la inducción morfogénica de B. purpurea.
AIA (mg/L)
BAP (mg/L)
Núm
ero
de r
aíc
es
0 0.5 1 1.5 2 2.50
0.51
1.52
2.5
1.1
1.3
1.5
1.7
1.9
2.1
AIA (mg/L)
BA
P (
mg/L
)
0 0.5 1 1.5 2 2.5
0
0.5
1
1.5
2
2.5 Raíces
1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
1.9
2.0
2.1
2.2
a)
b)

57
5.3 Estandarización de las técnicas para el estudio molecular utilizando extractos de ADN de Sphatoglotis plicata. La variación genética es una característica común de las células de las plantas que han sido objeto de protocolos de cultivo de tejidos. Esta variación genética podría estar presente en las células de la planta donadora o bien haber sido originado del proceso de cultivo, principalmente en la fase de callo. La estabilidad de las plantas regeneradas puede ser influenciada por cualquiera de estas condiciones por lo que se ha recurrido al uso de marcadores moleculares y bioquímicos para la detección de esta variación (Vargas et al., 2008). En la actualidad, la aplicación de las técnicas moleculares es de especial interés para la exitosa ejecución de programas tales como el mejoramiento genético y detección de la variación somaclonal de los cultivos, obtenidos por los métodos de cultivo in
vitro. Una técnica utilizada para este tipo de estudios es el método RAPD, la aplicación de esta técnica está comprendida principalmente de dos etapas: la primera es la extracción del ADN, y la segunda es la amplificación del ADN por PCR. La calidad del ADN utilizado es un factor importante a considerar para la reproducibilidad con esta técnica (Stewart y Via, 1993; Rocha, 2002). El objetivo principal en la extracción del material genético de las plantas, es obtener un ADN de buena calidad y en gran cantidad. La calidad se refiere a la integridad del ADN por lo cual aumenta la posibilidad de almacenarlo por tiempo indefinido, manteniendo su estructura y propiedades; además, es un factor importante para la reproducibilidad en experimentos posteriores. La cantidad depende del número y estado de las células propias del tejido a utilizar. Por lo tanto, un buen método de extracción debe mantener la integridad física y bioquímica del ADN e incrementar sus rangos de pureza y concentración (Rocha, 2002). Dado estos requisitos, en este estudio se probaron diferentes técnicas (Anexo 1) para la extracción del ADN genómico con el objetivo de seleccionar y estandarizar una técnica que permitiera la obtención de un ADN con las características mencionadas. Esto se apoyó con la cuantificación espectrofotométrica y la visualización en geles de agarosa. 5.3.1 Extracción de ADN a partir de brotes de Sphatoglotis plicata. Las técnicas ensayadas para la extracción están basadas en el método CTAB, empleando como material vegetal brotes germinados in vitro de Spathoglottis plicata, una especie de orquídea de origen oriental donada, esto mientras se esperaba la respuesta de la especie propuesta para este estudio, considerando que el tejido y secuencia genómica entre ambas especies podría ser similar y brindarnos información en la cual pudiéramos basarnos para establecer las condiciones para B. purpurea. Las

58
técnicas utilizadas se basaron en las propuestas por Saghai y Moroof et al., 1986, Zambrano et al., 2002, Allen et al., 2006 y Li et al., 2007, quienes las utilizaron para la extracción del ADN de plantas diferentes a orquídeas; por lo tanto, podemos esperar que la composición celular difiera y debido a ello también la concentración de cada reactivo requerido para la lisis celular, recuperación y limpieza del ácido nucleico. Se emplearon cuatro técnicas, de las cuales se puede observar una comparación del ADN obtenido por cada una de estas en la Figura 5.5, la cual muestra que por las tres técnicas se recuperó ADN, pero las concentraciones y calidad fueron diferentes. En la Figura 5.2 a, se aprecia una banda mayor a 12 000 kb lo cual indica la presencia de ADN en cada uno de los extractos obtenidos por las diferentes técnicas probadas. También puede notarse que con las técnicas 1 y 3 (Allen et al., 2006 y Zambrano et
al., 2002, respectivamente) se obtiene una mayor cantidad de ADN comparada con la obtenida por la técnica 2 (Saghai y Moroof et al., 1986) pero la pureza y cantidad de los extractos obtenidos por medio de la técnica 1 fue mejor que los obtenidos por las otras técnicas. Cabe mencionar que con la técnica 4 propuesta por Li et al. (2007) no se pudo recuperar ADN, por lo que se decidió descartar. La técnica 1 (Figura 5.5 a) propuesta por Allen et al. (2006) mostró ser útil para la especie de orquídea probada (S. plicata), por lo que se eligió para estudios posteriores, dado que se partió de tejido obtenido de la germinación in vitro. Una vez seleccionada la técnica se procedió a la estandarización de la misma (Figura 5.5 b) utilizando plántulas de S. plicata, se realizaron extracciones por triplicado obteniendo un ADN genómico limpio e íntegro. Con esta técnica se obtuvo ADN genómico como puede observarse en el gel una banda bien definida de buena concentración y calidad, lo cual fue reproducible.

Figura 5.5 Gel de agarosaSpathoglottis plicata. Muestras de 3 gel nativo de agarosa al 1 %, teñido con 2006; 2: técnica Saghai y Moroof obtenidos en la estandarización de la técnica de 1-3 extractos de ADN obtenidos por triplicado.Ladder, Invitrogen).
El rendimiento obtenido de acuerdo a la cuantificación del ADNmg de tejido utilizado (Tabla 5a la obtenida por Allen tabaco, quien reportó que respectivamente. Sin embargo, fue suficiente para las evaluaciones planteadas. Tabla 5.4 Resultados de la
obtenidos a partir de brotes
Muestra:
B B
Brote fresco (B); técnica 1, Allen
5.3.2 Amplificación al azar del polimorfismo del ADN (RAPD) Una vez establecida la técnica para la obtención del ADN genómico, el siguiente paso fue estandarizar las condiciones de amplificación para la
59
a)
agarosa para extractos obtenidos por método CTAB a partir de brotes de . Muestras de 3 µL de ADN fueron fraccionados por e
gel nativo de agarosa al 1 %, teñido con 0.5 µg mL-1 de BrEt . a) 1: técnica Saghai y Moroof et al., 1986; 3: técnica Zambrano et al., 2002
obtenidos en la estandarización de la técnica de extracción seleccionada (Allen extractos de ADN obtenidos por triplicado. M: escalera de ADN (1 Kb plus DNA,
El rendimiento obtenido de acuerdo a la cuantificación del ADN fue de 20(Tabla 5.4). La concentración obtenida fue ligeramente menor
por Allen et al. (2006) para diferentes especies A. thaliana
que por cada 200 mg de tejido se recuperaron 5, 30 y 50 µSin embargo, fue suficiente para las evaluaciones planteadas.
de la cuantificación espectrofotométrica de los extractos de ADN
obtenidos a partir de brotes de Spathoglottis plicata, mediante las técnicas 1 y 3.
Muestra: Técnica utilizada
Concentración (µg mL-1)
1 100.3 3 90.1
técnica 1, Allen et al., 2006; técnica 3, Zambrano et al., 2002
Amplificación al azar del polimorfismo del ADN (RAPD)
Una vez establecida la técnica para la obtención del ADN genómico, el siguiente paso fue estandarizar las condiciones de amplificación para la obtención de los PCR
b)
a partir de brotes de L de ADN fueron fraccionados por electroforesis en
écnica Allen et al., ., 2002. b) Extractos
Allen et al., 2006), escalera de ADN (1 Kb plus DNA,
fue de 20 µg por 300 ligeramente menor
A. thaliana, maíz y 5, 30 y 50 µg
Sin embargo, fue suficiente para las evaluaciones planteadas.
s extractos de ADN
1 y 3.
Una vez establecida la técnica para la obtención del ADN genómico, el siguiente paso obtención de los PCR-

60
RAPDs. El primer paso fue la selección de los oligonucleótidos con los que se logró la amplificación del ADN muestra. 5.3.2.1 Selección de oligonucleótidos. Operon technologies es una compañía que ha diseñado diversas series de 20 oligonucleótidos cortos, los cuales se han utilizado en diversos trabajos ya reportados para RAPDs. Un ejemplo, es el trabajo sobre variación somaclonal en Phalaenopsis reportado por Chen et al. (1998), en el cual utilizaron marcadores RAPDs y oligonucleótidos de las series A, D, G, J, M, Q, S, T, Y de Operon technologies. En este trabajo se utilizaron los oligonucleótidos de la serie OPJ que consta de 20 oligonucleótidos que van de OPJ-1 al OPJ-20. Para la selección de los oligonucleótidos que se utilizarían en el estudio, se realizaron 20 reacciones de PCR, una por cada oligonucleótido utilizando diferentes concentraciones. La primera condición evaluada fue la concentración de oligonucleótidos utilizada para la amplificación, para lo cual se evaluaron dos concentraciones una de 25 ng y otra de 50 ng ambos con 25 ng de ADN. La Figura 5.6 muestra la comparación de los perfiles obtenidos con ambas concentraciones de oligonucleótidos y en la Tabla 5.5 se concentra el número de bandas obtenidas por cada uno en base al perfil de la Figura 5.6. Cabe señalar que se eligieron los oligonucleótidos que presentaron un número de bandas mayor a tres y como referencia uno que presenta una sola banda. De los 20, se seleccionaron los oligonucleótidos OPJ 11, OPJ 12, OPJ 13, OPJ 14 y OPJ 15 para estandarizar las condiciones de la técnica.
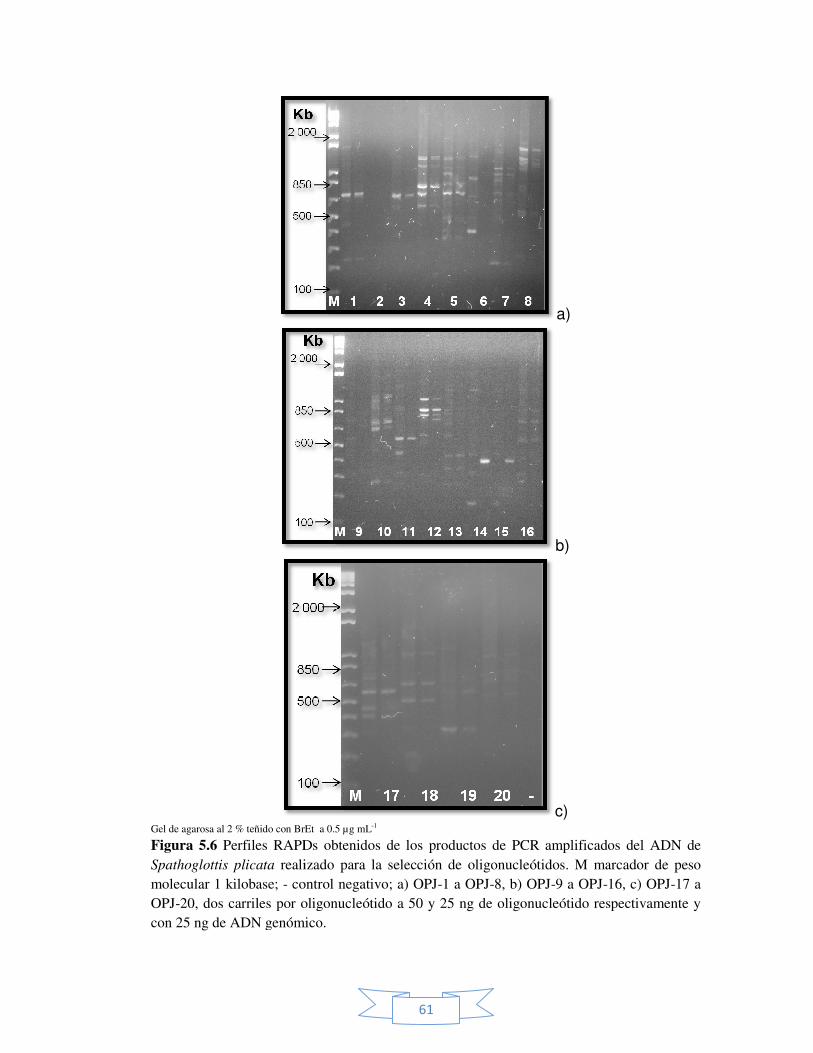
Gel de agarosa al 2 % teñido con BrEt a
Figura 5.6 Perfiles RAPDSpathoglottis plicata realizado para la selemolecular 1 kilobase; - control negativo; OPJ-20, dos carriles por oligonucleótido con 25 ng de ADN genómico
61
a)
b)
c) al 2 % teñido con BrEt a 0.5 µg mL-1
es RAPDs obtenidos de los productos de PCR amplificadosrealizado para la selección de oligonucleótidos. M marcador
control negativo; a) OPJ-1 a OPJ-8, b) OPJ-9 a OPJ-16dos carriles por oligonucleótido a 50 y 25 ng de oligonucleótido respectivamente
genómico.
obtenidos de los productos de PCR amplificados del ADN de marcador de peso
16, c) OPJ-17 a respectivamente y
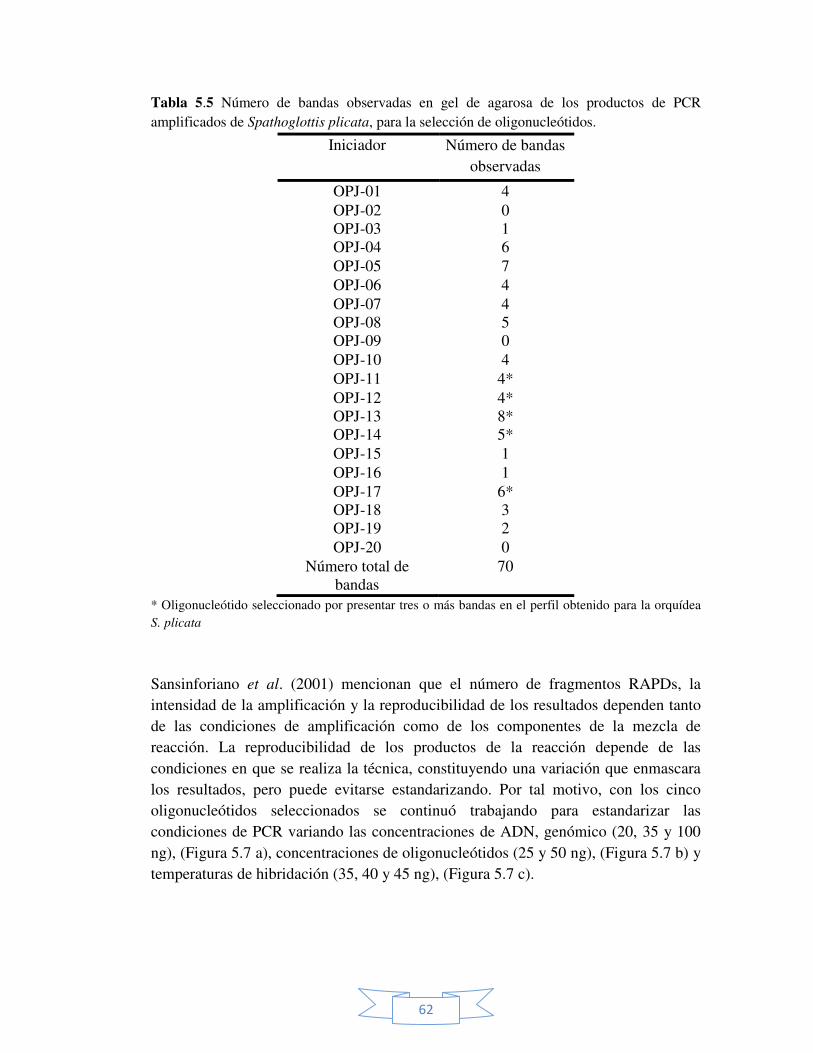
62
Tabla 5.5 Número de bandas observadas en gel de agarosa de los productos de PCR amplificados de Spathoglottis plicata, para la selección de oligonucleótidos.
Iniciador Número de bandas observadas
OPJ-01 4 OPJ-02 0 OPJ-03 1 OPJ-04 6 OPJ-05 7 OPJ-06 4 OPJ-07 4 OPJ-08 5 OPJ-09 0 OPJ-10 4 OPJ-11 4* OPJ-12 4* OPJ-13 8* OPJ-14 5* OPJ-15 1 OPJ-16 1 OPJ-17 6* OPJ-18 3 OPJ-19 2 OPJ-20 0
Número total de bandas
70
* Oligonucleótido seleccionado por presentar tres o más bandas en el perfil obtenido para la orquídea S. plicata
Sansinforiano et al. (2001) mencionan que el número de fragmentos RAPDs, la intensidad de la amplificación y la reproducibilidad de los resultados dependen tanto de las condiciones de amplificación como de los componentes de la mezcla de reacción. La reproducibilidad de los productos de la reacción depende de las condiciones en que se realiza la técnica, constituyendo una variación que enmascara los resultados, pero puede evitarse estandarizando. Por tal motivo, con los cinco oligonucleótidos seleccionados se continuó trabajando para estandarizar las condiciones de PCR variando las concentraciones de ADN, genómico (20, 35 y 100 ng), (Figura 5.7 a), concentraciones de oligonucleótidos (25 y 50 ng), (Figura 5.7 b) y temperaturas de hibridación (35, 40 y 45 ng), (Figura 5.7 c).
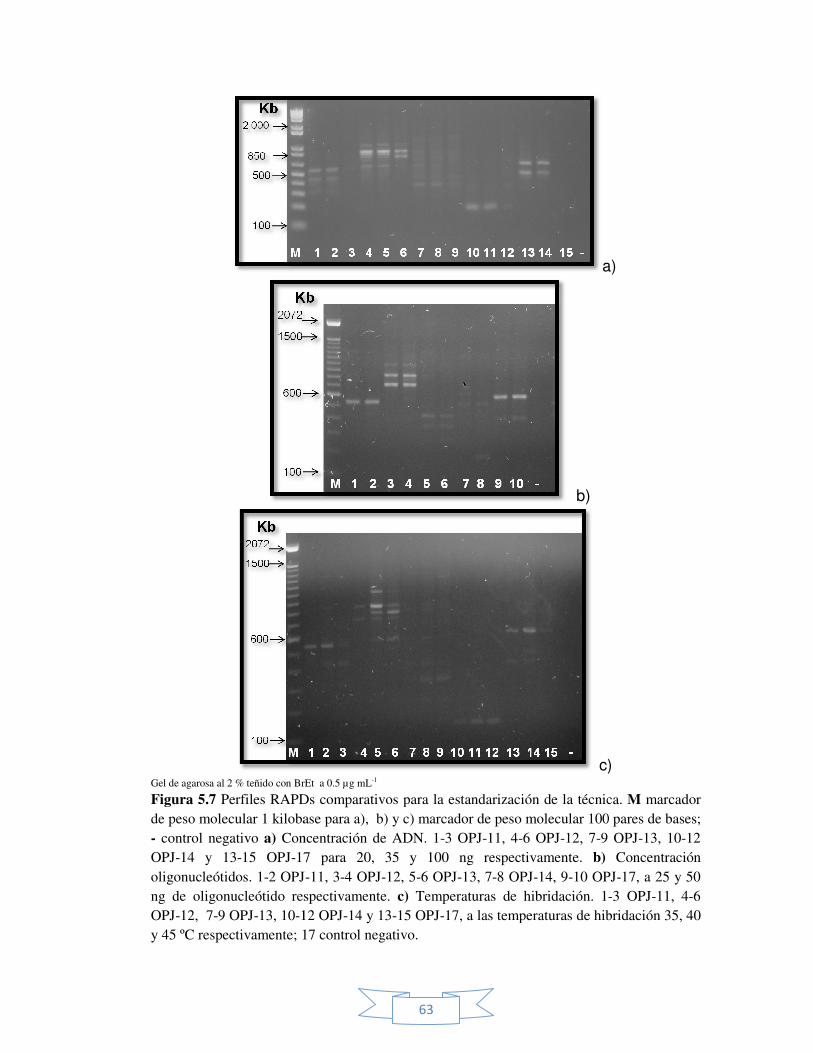
Gel de agarosa al 2 % teñido con BrEt a
Figura 5.7 Perfiles RAPDde peso molecular 1 kilobase para a), - control negativo a) Concentración deOPJ-14 y 13-15 OPJ-17oligonucleótidos. 1-2 OPJng de oligonucleótido respectivamente.OPJ-12, 7-9 OPJ-13, 10-12 OPJy 45 ºC respectivamente; 17 control negativo.
63
a)
b)
c)al 2 % teñido con BrEt a 0.5 µg mL-1
RAPDs comparativos para la estandarización de la técnica.o molecular 1 kilobase para a), b) y c) marcador de peso molecular 100 pares de bases;
Concentración de ADN. 1-3 OPJ-11, 4-6 OPJ-12, 7-9 17 para 20, 35 y 100 ng respectivamente. b)
OPJ-11, 3-4 OPJ-12, 5-6 OPJ-13, 7-8 OPJ-14, 9-10 OPJrespectivamente. c) Temperaturas de hibridación. 1-
12 OPJ-14 y 13-15 OPJ-17, a las temperaturas de hibridación 35, 40 17 control negativo.
a)
c)
para la estandarización de la técnica. M marcador b) y c) marcador de peso molecular 100 pares de bases;
OPJ-13, 10-12 Concentración
OPJ-17, a 25 y 50 -3 OPJ-11, 4-6
temperaturas de hibridación 35, 40

64
La cantidad del ADN genómico que se ha utilizado como templado en la técnica PCR-RAPDs, ha sido variable ya que se han reportado desde 1-100 ng. Por ejemplo, Sheidai el al. (2008) en su análisis molecular en plátano (Musa acuminate L.) utilizaron 1 ng, Tyagi et al. (2007), en su análisis de estabilidad genética en Curcuma
longa L. emplearon 50 ng. Los grupos de investigadores Neveen y Bekheet (2008) y Mohamed (2007) para fresa micropropagada utilizaron 5 y 100 ng, respectivamente. De Melo et al. (2006) reportan que utilizaron 25 ng para el análisis molecular de Dendrobium Hybrid. Por otro lado, para optimizar la concentración de ADN para la técnica PCR-RAPD Sansinforiano et al. (2001), en levaduras evaluaron 50, 100, 150, 200, 250 y 300 ng, de los cuales 150 ng fue la mejor. Por su parte, Ramírez, et al. (2008) en plantas de tomate evaluaron 1, 5, 10, 15, 25, 50, 100, 200, 400 ng y Goh, et
al. (2005) para Phalaenopsis evaluaron 50, 100, 150 y 200 ng de los cuales en ambos estudios la concentración de 50 ng fue la mejor. Por lo cual, en este trabajo se consideró estandarizar una concentración de ADN que permitiera obtener bandas más definidas y menor número de productos inespecíficos. En la Figura 5.7 a) se puede ver que a las concentraciones de 20 y 35 ng las bandas se veían iguales pero que comparado con el gel en donde se utiliza una concentración de 25 ng las bandas se ven más definidas (Figura 5.6). Con 100 ng la reacción se satura y de los cinco oligonucleótidos utilizados solo en uno se logró la amplificación. Estos resultados coinciden con otros publicados por Goh et al. (2005) y Ramírez et al. (2008), los cuales concluyen que altas concentraciones de ADN genómico (mayor de 100 ng) inhiben la actividad de la enzima Taq ADN polimerasa, probablemente por la presencia de una mayor cantidad de contaminantes en la muestra; o bien, un excesivo número de moléculas de ADN en relación con las de la enzima, lo cual satura la reacción. En la reacción PCR-RAPDs la hibridación tiene lugar en varios puntos de la cadena de ADN simultáneamente; haciendo necesario que los oligonucleótidos se añadan a la reacción en un exceso molar sobre el ADN genómico, de manera que la formación del complejo oligonucleótidos-ADN genómico se vea favorecida sobre la reasociación de las dos hebras del ADN en el siguiente paso del ciclo cuando desciende la temperatura (Ávila et al., 1995; Sansinforiano et al., 2001). Así que, para optimizar esta variable se utilizaron 25 y 50 ng. En la Figura 5.6, se observan los perfiles electroforéticos para ambas concentraciones de oligonucleótidos. Con 50 ng se ven bandas más nítidas y definidas, lo cual muestra que a esta concentración la intensidad de la amplificación proporciona productos visibles y poco inespecíficos. Esto permitió seleccionar aquellos oligonucleótidos que mostraron ser reproducibles mostrando un patrón de bandas con mayor resolución. Lo cual puede notarse en la Figura 5.7 b que confirma los resultados para la mayoría de los oligonucleótidos probados.

65
La fase de la hibridación es uno de los puntos más delicados de la reacción RAPD (Sansinforiano et al., 2001). La temperatura media más reportada es de 36 ºC para diferentes especies de plantas tanto crecidas de manera natural así como obtenidas mediante el cultivo in vitro (Goh et al., 2005; Tyagi et al., 2007; Lakshmanan et al., 2007; Mohamed, 2007; Ramírez et al., 2008). En este estudio, para conseguir la mayor especificidad posible en los productos se evaluaron distintas temperaturas (35, 40 y 45 ºC), como puede observarse en la Figura 5.7 c. Para los productos amplificados con los cinco oligonucleótidos y las tres temperaturas, las bandas se ven más definidas a la temperatura de 40 ºC, lo cual puede ser debido a que al aumentar la temperatura aumenta también la especificidad de los oligonucleótidos. Sansinforiano et al. (2001), menciona que al incrementar la temperatura de hibridación se reducen las uniones inespecíficas oligonucleótidos-ADN genómico y la incorporación errónea de nucleótidos, logrando de esta manera un producto más específico. Una vez evaluadas todas las condiciones; se seleccionaron las siguientes: 25 ng ADN, 50 ng de oligonucleótidos y una temperatura de hibridación de 40 ºC. 5.4 Estudio de la variación somaclonal en B. purpurea (Lam.). Una vez obtenido el material vegetal propuesto para este estudio se procedió a la extracción del ADN genómico, con el fin de cumplir con los objetivos planteados. Utilizando las condiciones seleccionados previamente con S. plicata se procedió al tratamiento de las muestras. En la Figura 5.8, se muestran los extractos de ADN obtenidos tanto para las plántulas madre como de las plántulas hijas de B. purpurea,
donde podemos observar la calidad del ADN, se obtuvo una banda mayor a 12 000 pb para todos los extractos, lo cual también nos muestra que el método seleccionado es reproducible para otras especies de orquídeas. La concentración obtenida varía en el rango de 5-10 µg, esto debido principalmente a la cantidad de tejido utilizado para la extracción. En la Tabla 5.6, se muestra un resumen de las concentraciones y relaciones A260/A280 y A230/A260 que respaldan la conclusión que el ADN no contiene proteínas ni otro compuesto que interfiera con las reacciones. Lo cual sugiere que se obtuvo una preparación libre de proteínas, compuestos de polifenólicos y polisacáridos, similar a los resultados obtenidos por Li et al. (2007) y Zambrano et al. (2002).
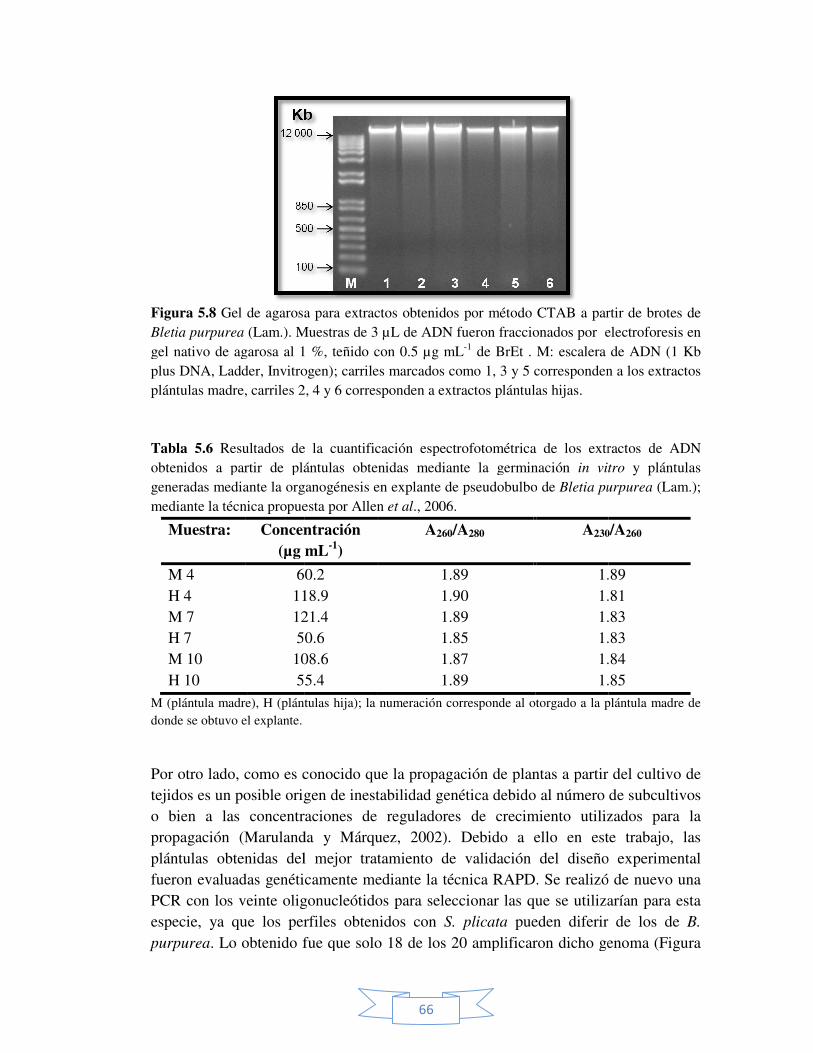
Figura 5.8 Gel de agarosaBletia purpurea (Lam.). Muestras de 3 gel nativo de agarosa al 1 %, teñido con plus DNA, Ladder, Invitrogen); plántulas madre, carriles 2, 4 Tabla 5.6 Resultados de laobtenidos a partir de plántulas obtenidas mediante la germinación generadas mediante la organogénesis en explante de pseudobulbomediante la técnica propuesta por Allen
Muestra:
Concentración(µg
M 4 60.2H 4 118.9M 7 121.4H 7 50.6M 10 108.6H 10 55.4
M (plántula madre), H (plántulas hija); la numeración corresponde al otorgado a la plántula madre de donde se obtuvo el explante.
Por otro lado, como es conocido que la propagación de plantas a partir del cultivo de tejidos es un posible origen de inestabilo bien a las concentraciones de reguladores de crecimiento utilizados para la propagación (Marulanda y Márquez, 2002). Debido a ello en este trabajo, plántulas obtenidas del mejor tratamientofueron evaluadas genéticamente medianPCR con los veinte oligonucleótidos para seleccionar las que se utilizarían para esta especie, ya que los perfiles obtenidos con purpurea. Lo obtenido fue que s
66
agarosa para extractos obtenidos por método CTAB a partir de brotes de
. Muestras de 3 µL de ADN fueron fraccionados por egel nativo de agarosa al 1 %, teñido con 0.5 µg mL-1 de BrEt . M: escalera de ADN (1 Kb plus DNA, Ladder, Invitrogen); carriles marcados como 1, 3 y 5 corresponden a los extractos
rriles 2, 4 y 6 corresponden a extractos plántulas hijas.
de la cuantificación espectrofotométrica de los extractos de ADN obtenidos a partir de plántulas obtenidas mediante la germinación in vitro
generadas mediante la organogénesis en explante de pseudobulbo de Bletia purpurea
mediante la técnica propuesta por Allen et al., 2006.
Concentración (µg mL-1)
A260/A280 A230
60.2 1.89 1.89118.9 1.90 1.81121.4 1.89 1.8350.6 1.85 1.83
108.6 1.87 1.8455.4 1.89 1.85
(plántula madre), H (plántulas hija); la numeración corresponde al otorgado a la plántula madre de
Por otro lado, como es conocido que la propagación de plantas a partir del cultivo de tejidos es un posible origen de inestabilidad genética debido al número de subcultivos o bien a las concentraciones de reguladores de crecimiento utilizados para la
anda y Márquez, 2002). Debido a ello en este trabajo, plántulas obtenidas del mejor tratamiento de validación del diseño fueron evaluadas genéticamente mediante la técnica RAPD. Se realizó dePCR con los veinte oligonucleótidos para seleccionar las que se utilizarían para esta especie, ya que los perfiles obtenidos con S. plicata pueden diferir de los de
Lo obtenido fue que solo 18 de los 20 amplificaron dicho genoma
a partir de brotes de L de ADN fueron fraccionados por electroforesis en
: escalera de ADN (1 Kb carriles marcados como 1, 3 y 5 corresponden a los extractos
cuantificación espectrofotométrica de los extractos de ADN in vitro y plántulas
urpurea (Lam.);
230/A260
1.89 1.81 1.83 1.83 1.84 1.85
(plántula madre), H (plántulas hija); la numeración corresponde al otorgado a la plántula madre de
Por otro lado, como es conocido que la propagación de plantas a partir del cultivo de idad genética debido al número de subcultivos
o bien a las concentraciones de reguladores de crecimiento utilizados para la anda y Márquez, 2002). Debido a ello en este trabajo, las
del diseño experimental realizó de nuevo una
PCR con los veinte oligonucleótidos para seleccionar las que se utilizarían para esta rir de los de B.
olo 18 de los 20 amplificaron dicho genoma (Figura

5.9), obteniendo de 1-5 bandas por oligonucleótido con un total de 51 5.8). En base a este experimento se seleccionaron OPJ-13, OPJ-15, OPJ-16 y OPJamplificados que fueron reproducibles. En el caso específico del OPJseleccionado debido a querápidamente la ausencia o presencia del amplificado. Los tamaños de los fragmentos amplificados oscilaron entre 100coincide con lo reportado en la literatura (< 2000 pb) cuando esta técnica es utilizada en estudios similares con otras especies de (2002), para verificar la calidad de los extractos de ADN realiza la amplificación de extractos de ADN de Saccharum
las tres especies productos de amplificación que oscil
Gel de agarosa al 2 % teñido con BrEt a
Figura 5.9 Perfiles RAPDBletia purpurea (Lam.), realizado para la selección de oligonucleótidos. Carriles con M: escalera de ADN (1con oligonucleótido (OPJ-
67
5 bandas por oligonucleótido con un total de 51 En base a este experimento se seleccionaron los oligonucleótidos:
16 y OPJ-18, por mostrar un mayor número de fragmentos amplificados que fueron reproducibles. En el caso específico del OPJseleccionado debido a que, presenta una sola banda lo cual permitió observar
ausencia o presencia del amplificado.
Los tamaños de los fragmentos amplificados oscilaron entre 100-1500 pb, lo cual coincide con lo reportado en la literatura (< 2000 pb) cuando esta técnica es utilizada en estudios similares con otras especies de plantas. Por ejemplo Zambrano
para verificar la calidad de los extractos de ADN realiza la amplificación de Saccharum spp., Musa sp. y Minihot esculenta
las tres especies productos de amplificación que oscilaron entre 200-1750 pb.
al 2 % teñido con BrEt a 0.5 µg mL-1
RAPDs de los productos de PCR amplificados del ADN de plántulas de realizado para la selección de oligonucleótidos. Carriles
escalera de ADN (100 pb plus DNA, Ladder, Invitrogen); - control negativo; 1-1 a OPJ-20).
5 bandas por oligonucleótido con un total de 51 bandas (Tabla : OPJ-4, OPJ-7,
, por mostrar un mayor número de fragmentos amplificados que fueron reproducibles. En el caso específico del OPJ-15 fue
permitió observar
1500 pb, lo cual coincide con lo reportado en la literatura (< 2000 pb) cuando esta técnica es utilizada
as. Por ejemplo Zambrano et al. para verificar la calidad de los extractos de ADN realiza la amplificación de
Minihot esculenta obteniendo en 1750 pb.
del ADN de plántulas de realizado para la selección de oligonucleótidos. Carriles marcados
control negativo; 1-20
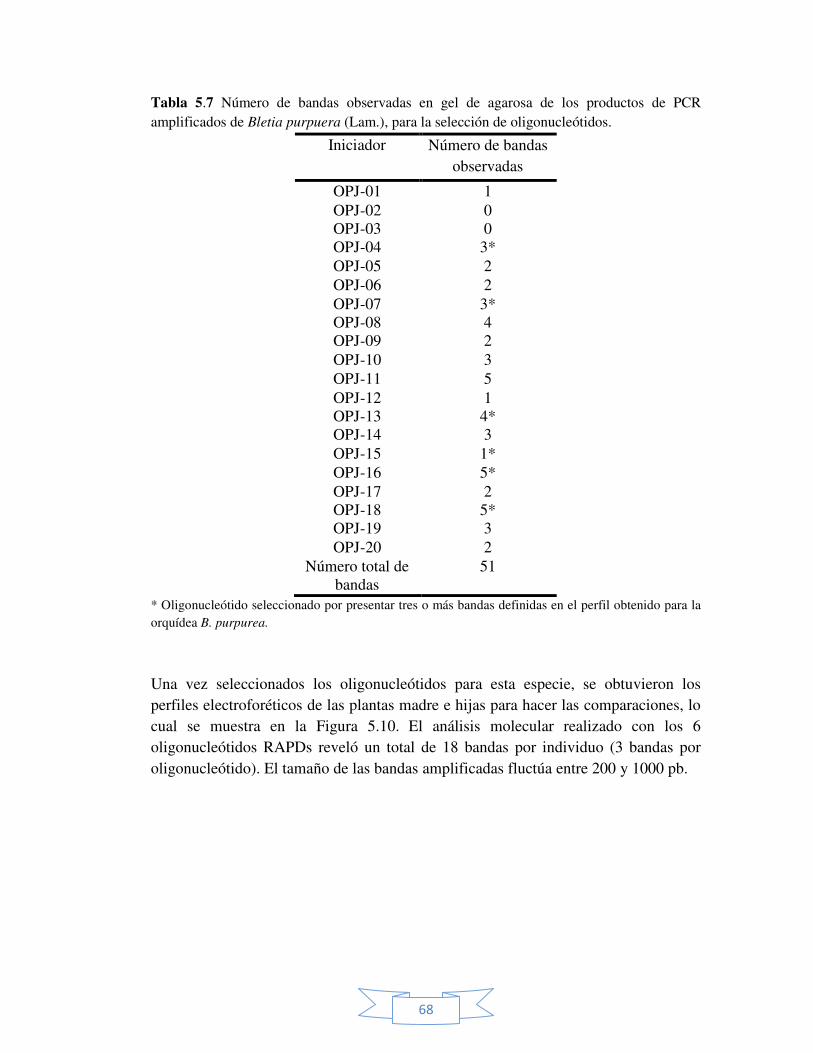
68
Tabla 5.7 Número de bandas observadas en gel de agarosa de los productos de PCR amplificados de Bletia purpuera (Lam.), para la selección de oligonucleótidos.
Iniciador Número de bandas observadas
OPJ-01 1 OPJ-02 0 OPJ-03 0 OPJ-04 3* OPJ-05 2 OPJ-06 2 OPJ-07 3* OPJ-08 4 OPJ-09 2 OPJ-10 3 OPJ-11 5 OPJ-12 1 OPJ-13 4* OPJ-14 3 OPJ-15 1* OPJ-16 5* OPJ-17 2 OPJ-18 5* OPJ-19 3 OPJ-20 2
Número total de bandas
51
* Oligonucleótido seleccionado por presentar tres o más bandas definidas en el perfil obtenido para la orquídea B. purpurea.
Una vez seleccionados los oligonucleótidos para esta especie, se obtuvieron los perfiles electroforéticos de las plantas madre e hijas para hacer las comparaciones, lo cual se muestra en la Figura 5.10. El análisis molecular realizado con los 6 oligonucleótidos RAPDs reveló un total de 18 bandas por individuo (3 bandas por oligonucleótido). El tamaño de las bandas amplificadas fluctúa entre 200 y 1000 pb.
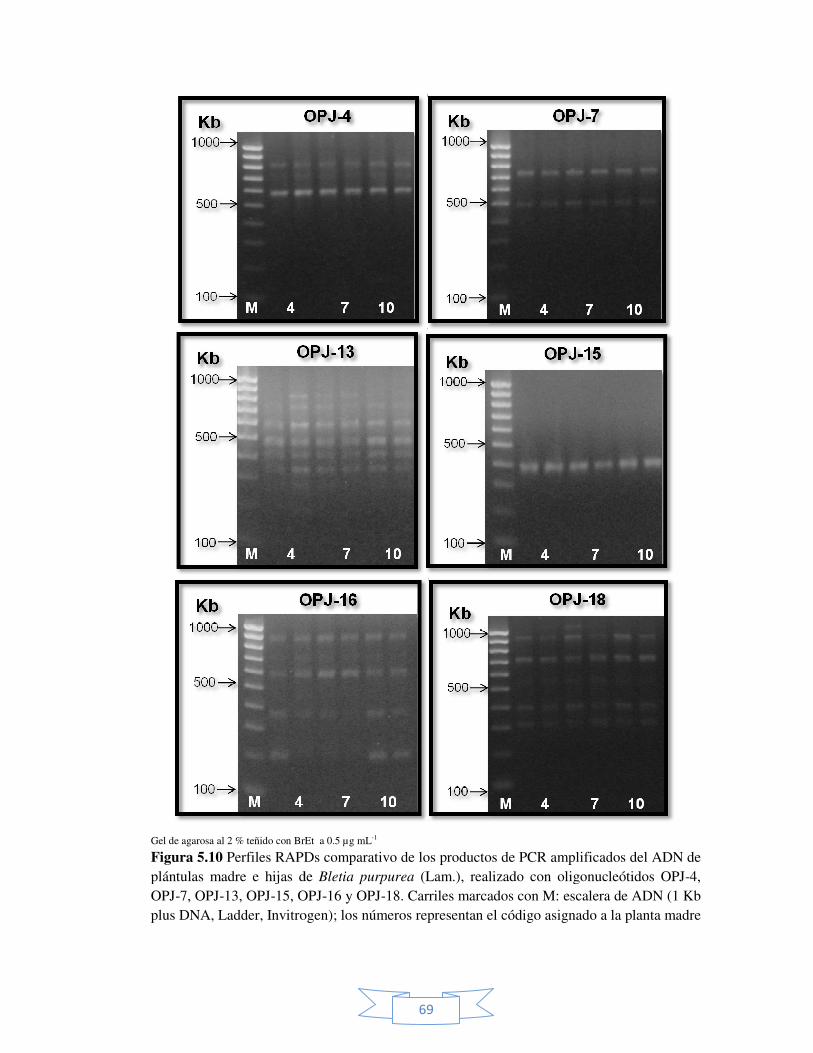
Gel de agarosa al 2 % teñido con BrEt a
Figura 5.10 Perfiles RAPDplántulas madre e hijas de OPJ-7, OPJ-13, OPJ-15, OPJplus DNA, Ladder, Invitrogen)
69
al 2 % teñido con BrEt a 0.5 µg mL-1
RAPDs comparativo de los productos de PCR amplificados del ADN plántulas madre e hijas de Bletia purpurea (Lam.), realizado con oligonucleótidos
15, OPJ-16 y OPJ-18. Carriles marcados con M: escalera de ADN (1 Kb nvitrogen); los números representan el código asignado a la planta madre
de los productos de PCR amplificados del ADN de oligonucleótidos OPJ-4, escalera de ADN (1 Kb
los números representan el código asignado a la planta madre

70
de procedencia del explante, el primer carril extracto madre seguido del extracto plántulas hijas.
En este tipo de estudios, en los que la técnica RAPDs es aplicada cada banda nos indica un loci, aunque no se puede distinguir entre individuos heterocigotos y homocigóticos; si nos proporciona la información de si existe o no una similitud genética entre los individuos evaluados. Los perfiles obtenidos en este estudio muestran, que las plántulas micropropagadas son monomórficas y análogas a sus respectivas plántulas madre. El monitoreo de la variabilidad en las secuencias del ADN de las plantas ha sido reportado utilizando marcadores moleculares RAPD, SSR, ISSR y AFLP. La ausencia de la variación genética utilizando RAPD se ha reportado en varios casos, tal como en las plántulas regeneradas de Curcuma longa L. (Tyagi et al., 2007), en la micropropagación de dos variedades de caña de azúcar (Lal et al., 2008), en la micropropagación a partir de rizomas en tres genotipos de Vitis spp (Alizadeh y Singh, 2009). De Melo et al. (2006) no encontraron alteraciones genéticas en la propagación clonal de Dendrobium Second Love usando brotes axilares en presencia de tidiazuron. En contraste, la variación somaclonal fue reportada en las plántulas regeneradas mediante la inducción in vitro de brotes adventicios a partir de hojas de Piña (Soneji et al., 2002). Martin et al. (2002) detectaron pequeñas variaciones en plántulas de crisantemo, las cuales fueron sujetas a diferentes condiciones de proliferación, siguiendo una vía de regeneración mediada por la formación de callos. Chen et al. (1998), reportan variaciones genéticas en somaclones obtenidos a partir de cuerpos tipo protocormos en Phalaenopis True Lady. Por lo tanto, en el presente estudio utilizamos la técnica RAPD basado en la PCR para la identificación de la variación somaclonal en plántulas de B. purpurea obtenidas de la inducción morfogénica a partir de pseudobulbos debido a su simplicidad y bajo costo. De acuerdo, a los oligonucleótidos probados no hubo cambios en el patrón de bandas observado en las plántulas obtenidas del cultivo de tejidos en comparación con las plántulas madre, llegamos a la conclusión de que el protocolo de propagación podría llevarse a cabo sin mucho riesgo de inestabilidad genética en la regeneración de la primera generación.

71
VI. CONCLUSIONES

72

73
Se logro la optimización de la concentración del Acido Indol Acético y la Bencil Amino Purina sobre la morfogénesis in vitro en explantes de pseudobulbos de Bletia
purpurea. Se validaron las ecuaciones obtenidas para número de brotes, hojas y raíces. Siendo el tratamiento óptimo validado con la concentración de 1.8 mg L-1 de AIA, para la organogénesis directa en los pseudobulbos de Bletia purpurea. Se obtuvo un protocolo de propagación in vitro vía organogénica de Bletia purpurea (Lam.). a partir de pseudobulbos, los cuales mostraron formación directa de brotes a los 15 días de la inducción, desarrollando a plántulas completas a los 60 días del cultivo en medio MS con fuerza iónica completa suplementado con 1.8 mg L-1 de AIA e incubados a 23 ± 2°C en condiciones de fotoperíodo 16/8 horas luz. El análisis RAPDs con los oligonucleótidos seleccionados (OPJ-4, OPJ-7, OPJ-13, OPJ-15, OPJ-16 y OPJ-18), obtenidos mostraron perfiles similares entre la muestra analizada correspondiente a las plántulas madre e hijas. En base a lo anterior podemos asumir que no se encontró variación somaclonal, lo que sugiere que la concentración de 1.8 mg L-1 de AIA puede ser utilizada para la obtención de plántulas ya sea para la propagación masiva y en los casos cuando una propagación clonal sea requerida.

74

75
VII. RECOMENDACIONES

76

77
El presente trabajo representa una importante contribución para la obtención de nuevas plántulas mediante la vía organogénica a partir de pseudobulbos y puede servir como base para futuras aplicaciones para la regeneración in vitro de Bletia
purpurea (Lam.). Se propone realizar estudios de optimización en las etapas de multiplicación y enraizamiento, para su posterior adaptación al medio ambiente y al mismo tiempo considerar su estudio molecular en cada una de estas etapas. En nuestro estudio sobre variación somaclonal mediante la técnica RAPD, recomendamos una evaluación con otras técnicas moleculares tales como AFLPs ó ISSRs, con las cuales podríamos corroborar los resultados del presente trabajo; o bien realizar estudios sobre posibles cambios epigenéticos en la vía de regeneración in
vitro.

78

79
VII. REFERENCIAS BIBLIOGRÁFICAS

80

81
Ahloowalia, B. S. (1998). In-vitro techniques and mutagenesis for the improvement of vegetatively propagated plants. In: Jain, SM; Brar, DS. Ahloowalia, BS. eds. Somaclonal variation and induced mutations in crop improvement. Dordrecht, Kluwer. p. 293-309. Alizadeh, M. y Singh, S. K. (2009). Molecular assessment of clonal fidelity in micropropagated (Vitis spp.) rootstock genotypes using RAPD and ISSR markers. Iranian journal of biotechnology. 7(1): 37-44. Allen, G. C., Flores, V. M. A., Krasnyanski, S., Kumar, S. y Thompson W. F. (2006). A modified protocol for rapic DNA isolation from plant tissue using cetyltrimethylammonium bromide. Nature protocols. 1(5): 2320-2325. Alonso, G. M. (2002). Biotecnología aplicada a la mejora de Pelargonium.
Universidad Complutense de Madrid. Facultad de CC. Biológicas. Departamento de Genética. Tesis doctoral. p. 23-25, 35, 84-86. Al-Zahim, M. A., Ford-Lloyd, B. V., Newbury, H. J. (1999). Detection of somaclonal variation in garlic (Allium sativum L.) using RAPD and cytological analysis. Plant Cell Reports. 18: 473-477. Anu, A., Babu, K. N. y Peter, K .V. (2004). Variations among somaclones and its see dling progeny in Capsicum annuum. Plant Ce ll, Tissue and Organ Culture 76: 261–267. Araújo, L. G., Prabhu, A. S., Filippi, M. C., Chaves, L. J. (2001). RAPD analysis of blast resistant somaclones from upland rice cultivar IAC 47 for genetic divergence. Plant Cell, Tissue and Organ Culture. 67: 165-172. Agarwal, M., Shrivastava, N. y Padh, H. (2008). Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep. 27: 617-631. Ávila, J. T., De Vera, A. M., Hernández, C. y Vasallo, M. P. (1995). Biología molecular en nefrología, Análisis del DNA. II: Clonar, secuenciar y PCR. Nefrología. Vol. XV. Suplemento 2. Azofeifa-Delgado, A. (2006). Uso de marcadores moleculares en plantas; aplicaciones en frutales del trópico. Agronomía Mesoamericana. 17(2): 221-242.

82
Bajaj, Y. P. S. (1991). Biotechnology in Agriculture and Forestry 17 High-Tech and Micropropagation I. Springer-Verlag, Berlin Heidelberg New York. p. 53. Basker, S. y Narmatha, B. V. (2006).Micropropagation of Coelogyne stricta (D. Don) Schltr. via pseudobulbo segment cultures. Tropical and sub tropical agroecosystems. 6: 31-35. Bellido, O. (2007). Orquídeas. Naturart, S. A. Editado por Blume jardinería. Barcelona. p. 6. Brar, S. M., Brar, D. S. (1998). Somaclonal variation: mechanism and applications in crop improvement. In Jain S. M. Brar, DS. eds. Somaclonal variation and induced mutations in crop improvement. Dordrecht, Kluwer. p. 15-37. Bridg, H. (2001). Estabilidad genética en Annona cherimola y A. muricata (en línea). Berlín, Alemania. Disponible en: http://arc.cs.odu.edu:8080/dp9/getrecord/oaidc/oai:HUBerlin:dissertationen/bridg-hannia 2000-03-24. Cardone, S., Olmos, S. y Echenique, V. (2004). Variacion somaclonal In: Echenique V., Rubinstein C., y Mroginski L. (eds). Biotecnologia y mejormiento vegetal. Ediciones INTA III (1): 83-96. Carnevali, F., Tapia, M., Jiménez, M., Sánchez, S., Ibarra, G., Ramírez, I. y Gómez, M. (2001). Notes on the flora of the Yucatán peninsula II: a synopsis of the orchid flora of the Mexican Yucatan Peninsula and a tentative checklist of the orchidaceae of the Yucatán peninsula biotic province. Harvard Papers in Botany. 5(2): 1-84. Cassells, C. A., Curry, R. F. (2001). Oxidative stress and physiological, epigenetic and genetic variability in plant tissue culture: implications for micropropagators and genetic engineers. Plant Cell, Tissue and Organ Culture. 64: 145-157. Castro, A.; Rosas, J.C.; Aranda, L. (2001). Manual del Módulo de Biotecnología. Escuela Agrícola Panamericana, Zamorano. p. 19. Chen, L.; Pan, R. y Chen, R. (1999). Effects of media, growth regulators and dividing on the growth of Cymbidium sinense protocorm cultured in vitro. J. Trop. and Subtrop. Bot. 7: 59-64. Chen, T-Y, Chen, J-T y Chang, W-C. (2002a). Multiple shoot formation and plant regeneration from stem nodal explants of Paphiopedilum orchid. In vitro cell. Dev. Boil. Plant. 38: 595-597.

83
Chen, T-Y, Chen, J-T y Chang, W-C. (2002b). Efficient production of protocorm-like bodies and plant regeneration from flower stalk explants of the sympodial orchid Epidendrun radicans. In vitro cell. Dev. Boil. Plant. 38: 441-445. Chen. W. H., Chen, T. M., Fu, Y. M., Hsieh, R. M. y Chen W. S. (1998). Studies on somaclonal variation in Phalaenopsis Plant Cell Reports. 18:7-13. Chen, Y. H., Tsai, Y. J., Huang, J. Z. y Chen, F. C. (2005). Transcription analysis of peloric mutants of Phalaenopsis orchids derived from tissue cultura. Cell Research, 15(8): 639-657. Condemarín M. C. E., Chico R. J. y Vargas A. C. (2007). Efecto del acido indolbutirico (iba) y 6-bencilaminopurina (BAP) en el desarrollo in vitro de yemas axilares de Encyclia microtos (RCHB.F.) hoehne (Orchidaceae). Lankesteriana 7(1-2): 247-254. Davies, P. J. (1995). The plant Hormones: Their Nature, occurrence and functions. In: Davies, P. J. (Ed). Plant hormones physiology, biochemistry and molecular biology. Kluwer Academic Publisher, Dordrecht. p. 1-11. De Melo, F. W., Barbante, K. G. y Pimentel, C. A. P. (2006). Micropropagation and genetic stability of a Dendrobium hybrid (Orchidaceae). In Vitro Cell. Dev. Biol. Plant. 42: 568–571. Dutra, D., Johnson, T. R., Kauth, J. P., Stewart, S. L. Richardson, L. y Kane, M. E. (2008). Asymbiotic germination, In Vitro seedling development, and greenhouse acclimatization of the threatened terrestrial orchid Bletia purpurea. Plant Cell Tiss Organ Cult. 94: 11-21. Echeverri, S. A. P. Jaramillo, E. J. R. y Villegas, V. F. (2001). Manual de cultivo de orquídeas. Sociedad Colombiana de Orquideología. George, E. F., Hall, M. A. y De Klerk, G-J. (2008). Plant propagation by tissue culture. Springer, 3rd edition, vol. 1 the background. p. 185. Goh, M. W. K., Kumar, P. P., Lim S. H. y Tan, H. T. W. (2005). Random amplified polymorphic DNA analysis of the moth orchids, Phalaenopsis (Epidendroideae:
Orchidaceae) Euphytica. 141: 11–22.

84
Gow, W-P., Chen, J-T. y Chang, W-C. (2008). Influence of growth on direct embryo formation from leaf explants of Phalaenopsis orchid. Acta physiol. Plant. 30: 507-512. Gupta, P. K. y Varshney, R. K. (1999). Molecular markers for genetic fidelity during micropropagation and germplasm conservation. Current Science. 76: 1308–1310. Grima i, C., P., Prat, B. A. y Posueta F. L. (2004). Métodos estadísticos, control y mejora de la calidad. Ediciones UPC, S.L. Segunda edición: ISBN: 8483017865 ISBN-13: 9788483017869. Halušková, J. y Košuth, J. (2003). RAPD analysis of somaclonal and natural DNA variation in Hypericum perforatum L. Acta Biologica Cracoviensia Series Botánica. 45: 101-104. Hammerschlag, F., Garcés, S., Koch-Dean, M., Ray, S., Lewers, K., Maas, J. y Smith, B. J. (2006). In vitro response of strawberry cultivars and regenerants to Colletotrichum acutatum. Plant Cell, Tissue and Organ Culture 84: 255-261. Hassan, N. A. y Bekheet, S. A. (2008). Mid-term Storage and Genetic Stability of Strawberry Tissue Cultures. Journal of Agriculture and Biological Sciences, 4(5): 505-511. Hew, C, S. y Clifford, P. E. (1993). Plant growth regulators and the orchid cut-flower industry. http://www.scielo.org.ve/scielo.php. Hew, C. S. y Yong, J. W. H. (2004). The physiology of tropical orchid in relation to the industry. World Scientific. Second edition. Cap. 2 p. 11-36. Hong, P-I., Chen, J-T y Chang, W-C. (2010). Shoot development and plant regeneration from protocorm-like bodies of Zygopetalum mackayi. In Vitro Cell. Dev. Biol. Plant. 46: 306–311. Hurtado, D; Merino, M. (1994). Cultivo de Tejidos Vegetales. Trillas. México, D.F. México. p. 232. Jain, S. M. (2001). Tissue culture-derived variation in crop improvement. Euphytica. 118: 153-166. Janarthanam, B. y Seshadri, S. (2008). Plantlet regeneration from leaf derived callus of Vanilla planifolia Andr. In Vitro Cell. Dev. Biol. Plant. 44: 84-89.

85
Jankiewicz, L. S. (2003). Desarrollo de semillas y la presencia de latencia en yemas y semillas; el papel de los reguladores de plantas. En: Jankiewicz, L.S. (ed), p. 193-240. Reguladores del crecimiento, desarrollo y resistencia en plantas. Vol. I. Propiedades y acción. Ediciones Mundi- Prensa, México. p. 487. Johnson, T. R. y Kane, M. E. (2007). Asymbiotic germination of ornamental Vanda: In Vitro germination and development of three hybrids. Plant Cell Tiss Organ Cult. 91: 251-261. Kaeppler, S. M.; Kaeppler, H. y Rhee, Y. (2000). Epigenetic aspects of somaclonal variation in plants. Plant Mol. Biol. 43: 179-188. Kubis, S. E., Castilho, A. M. M. F., Vershinin, A. V. y Heslop-Harrison, J. S. (2003). Retroelements, transposons and methylation status in the genome of oil palm (Elaeis
guineensis) and the relationship to somaclonal variation. Plant Molecular Biology. 52: 69–79. Kuo, H-L., Chen, J-T. y Chang, W-C (2005). Efficient plant regeneration through direct somatic embrigenesis from leaf explants of Phalaenopsis Little Steve. In Vitro Cell. Dev. Biol. Plant. 41: 453-456. Lakshmanan, V., Venkataramareddy, R. S. y Neelwarne, (2007). Molecular analysis of genetic stability in long-term micropropagated shoots of banana using RAPD and ISSR markers. Electronic Journal of Biotechnology B. by Pontificia Universidad Católica de Valparaíso Chile. 10(1): 106-113. Lara, A. J. P. (2005). Obtención de la huella genética de agaves mezcaleros de San Luis Potosí por RAPD y AFLP. Instituto Potosino de investigación científica y tecnológica, A. C. Tesis de maestría en Biología Molecular. p. 12, 13, 19-22, 26, 27. Larkin, P. (2004). Somaclonal Variation: Origins and Causes. Encyclopedia of Plant and Crop Science by Marcel Dekker, Inc. All rights reserved. p. 1158-1161. Larkin, P. y Scowcroft. W. (1981). Somaclonal variation a novel source of variability from cells cultures from plant improvement. Theor. Appl. Genet. 60: 197-214. Lee, E. H. E., Laguna, C. A., Murguía, G. J., Elorza, M. P., Iglesias, A., García, R. B., Barredo, P. F. A. y Santana, B. N. (2007). Regeneración In vitro de Laelia anceps ssp. Dawsonii. Revista UDO Agrícola. 7(1): 58-67.

86
Li, A. y Ge, S. (2006). Genetic variation and conservation of Changnienia amoena, an endangered orchid endemic to China Pl. Syst. Evol. 258: 251–260. Li, J. T., Yang, J. Chen, D. C., Zhang, X. L. y Tang, Z. S. (2007). An optimized mini-preparation method to obtain high-quality genomic DNA from mature leaves of sunflower. Genetics and molecular Research. 6(4): 1064-1071. Luan, V. Q., Thien, N. Q., Khiem, D. V. y Nhut, D. T. (2006). In vitro germination capacity and plant recovery of some native and rare orchids. Proceedings of International Workshop on Biotechnology in Agriculture, Nong Lam University Ho Chi Minh City. October, 20-21. Margara, J. (1988). Multiplicación vegetativa y cultivo in vitro: los meristemos y la organogénesis. Madrid: Mundi-Prensa. Martín, C., Uberhuaga, E. y Pérez, C. (2002). Application of RAPD markers in the characterization of Chrysanthemum varieties and the assessment of somaclonal variation. Euphytica. 127: 247-253. Martins, M., Sarmento, D. y Oliveira, M. M. (2004). Genetic stability of micropropagated almond plantlets, as assessed by RAPD and ISSR markers. Plant Cell Rep. 23: 492–496. Marulanda, M. L. y Márquez, M del P (2002). Evaluación de la estabilidad genética de vitroplantas de Rubus glaucus mediante marcadores moleculares (RAPD). Actual boil. 24(76): 31-36. Mata, R. M., Baltazar, G. R. J., Moon, P., Hietz P. y Luna, M. V. E. (2010). In vitro regeneration of Lycaste aromatica (Graham ex Hook) Lindl. (Orchidaceae) from pseudobulb sections. Plant Biotechnol. Rep 4: 157–163. Mohamed, A. E-S. (2007). Somaclonal Variation in Micro-propagated Strawberry Detected at the Molecular Level. Int. J. Agri. Biol. 9(5): 721-725. Montgomery, D. (2007). Diseño y análisis de experimentos. Segunda edición, México, Limusa Wiley, Capítulo 11 p. 456. Mujib, A. (2004). In vitro variability in tissue culture: a fresh look. In Mujib, A; Myeong-je, C; Predieri, S; Banerjee, S. eds. In vitro application in crop improvement. Enfield, EUA, Science Publishers. p. 261-271.

87
Murashige, T. y Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 15: 473-497. Murthy, H. N. (2005). In vitro multiplication and ecorehabilitation of rare orchid Aerides crispum. The role of biotechnology. p. 191-192. Narváez, C. Valenzuela, B. J., Muñoz, Sch. C. y Hinrichsen, R. P. (2000). Comparación de RAPD y AFLP como métodos de identificación genética de vid basados en el estudio de fragmentos genómicos anónimos. Agric. Téc. 60(4): 320-340. Nayak, N. R., Patnaik, S. y Rath, S. P. (1997). Direc shoot regeneration from foliar explants of an epiphytic orchid, Acampe praemorsa (Roxb.) Blatter and McCann. Plant cell reports. 16: 583-586. Nayak, N. R., Chand, P. K., Rath, S. P. y Patnaik, S. N. (1998). Influence of some plant growth regulators on the growth and organogenesis of Cymbidium aloifolium (L.) sw. seed-derived rhizomes in vitro. In Vitro Cell. Dev. Biol. Plant. 34: 185-188. Noro, Y., Takano-Shimizu, T., Syono, S., Kishima, Y. y Sano, Y. (2007). Genetic variations in rice in vitro cultures at the EPSPs-RPS20 region. Theoretical and Applied Genetics. 114: 705-711. Nuñez, V. M. L. (1995). Optimización de las condiciones de operación de un reactor de lecho fluidizado para la producción de alcohol empleando células inmovilizadas de Sacharomyces cerevisiae. Tesis de licenciatura en biología. Universidad de Guadalajara. p. 21 y 22
Osipova, E. S, Koveza O. V, Troitskii A. V, Dolgikh lul, Shamina, Z. B y Gostimskii, S. A. (2003). Analysis of specific RAPD and ISSR fragments in somaclonal maize (Zea mays L.) and development of SCAR markers based on them. Genet. 39: 1664-1672. Pedroza, M. J. A (2009). Efecto del carbón activado, ácido indolacético (AIA) y bencil amino purina (BAP) en el desarrollo de protocormos de Epidemdrum
elongatum Jacq bajo condiciones in vitro. Rev. Colomb. Biotecnol. Vol. XI (1): 17-32. Pérez, E., Ramírez, R., Gordon, H. y Ochoa, N. (1999). Introducción al cultivo de tejidos vegetales. Universidad de Aguascalientes. Primera edición, México. p. 71-80.

88
Pérez, B. M., Delgado, M., Hernández, C. A. y Armas, R. (2008). Organogénesis indirecta a partir de meristemos apicales caulinares de la variedad cubana de arroz reforma. Cultivos tropicales. 29(1): 23-28. Phillips, R. L., Kaeppler, S. M. y Olhoft, P. (1994). Genetic instability of plant tissue cultures: breakdown of normal controls. Proc. Natl. Acad. Sci USA. 91: 5222-5226. Phillips, W.; Rodríguez, H. y Fritz, P. (1995). Marcadores de ADN: Teoría, aplicaciones y protocolos de trabajo con ejemplos de investigaciones en cacao (Theobroma cacao). Serie técnica. Informe técnico # 252. CATIE. Turrialba, Costa Rica. p. 183. Picca, A., Helguera, M., Salomón, N., Carrera, A. (2004). Marcadores moleculares. In Echenique, V. Rubinstein, C. Mroginski, L. eds. Biotecnología y mejoramiento vegetal. Consejo Argentino para la Información y el Desarrollo de la Biotecnología. Argentina, Ediciones INTA. p. 61-68 (11-marzo-2009) Pierik, R. L. M. (1988). Cultivo in vitro de las plantas superiores, 3rd Ed. Ediciones Mundi-Prensa, Madrid. p. 326. Pierik, R. L. M. (1997). In vitro culture of higher plants. Dordrecht, Kluwer. p. 364. Pierik, R. L. M. (1990). Producción de plantas libres de enfermedades. En: Cultivo in vitro de las plantas superiores. 3rd Ed. Ediciones Mundi-Prensa. Madrid, España. p. 169-179. Podwyszyńska, M. (2005). Somaclonal variation in micropropagated tulips based on phenotype observation. Journal of Fruit and Ornamental Plant Research. 13: 109-122. Polanco, C. y Ruiz, M. L. (2002). AFLP analysis of somaclonal variation in Arabidopsis thaliana regenerated plants. Plant Science 162: 817-824. Qureshi, J. A, y Saxena, P. K. (1992). Adventitious shoot induction and somatic embryogenesis with intact seedlings of several hybrid seed geranium (Pelargonium x hortorum Bailey) varieties. Plant Cell Reports. 11(9): 443-448. Ramírez, R., González, L. M., González, M. C., Chávez, L., Fernández, A y Camejo, Y. (2008). Estudio bioquímico-molecular de la estabilidad genética en plantas de tomate procedentes de semillas tratadas con bajas dosis de rayos x. Cultivos Tropicales. 29(3): 39-46.

89
Rocha, S. P. J. (2002) Teoría y práctica para la extracción y purificación del ADN de palma de aceite. Palmas. 23(3): 9-17. Romero, T. R., Luna, R. B. S. y Barba, A. A. (2007). Uso de complejos comerciales como sustitutos de componentes del medio de cultivo en la propagación In vitro de Laelia anceps. Lankesteriana 7(1-2): 353-356. Rout, G. R., Das, P., Goel, s. y Raina, S. N. (1998). Determination of genetic stability of micropropagated plants of ginger using Ramdom Amplified Polymorphic DNA (RAPD) markers. Bot. Bull. Acad. Sin. 39: 23-27. Saghai-Maroof, M. A, Soliman, K. M., Jorgensen, R. A y Allard, R. W. (1984). Ribosomal DNA spacer-Iength polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proceedings of the National Academy of Sciences, USA. 81: 8014-8019. Sahijram, L., Soneyi J. R. y Bollamma, K. T. (2003). Analyzing somaclonal variation in micropropagated bananas (Musa spp.). In Vitro Cellular and Developmental Biology Plant. 39:551-556. Salisbury, F. y C. Ross. (2000). Fisiología de las plantas. Thomson Edited Spain Paraninfo. Madrid. Sánchez, M. A., Sarmiento, M. y Andrews, J. M. (1998). Orquídeas de Campeche. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP produce), Centro de investigación regional del sureste campo experimental “Ing. Eduardo Sangri Serrano”. Sánchez, M. A., Sarmiento, M. y Andrews, J. M. (2002). Orquídeas silvestres de Campeche con alto potencial económico. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). p. 15-17, 28-30, 77, 120-122. Sánchez, M., Sarmiento, M. y Andrews, J. (2002). Orquídeas de Campeche. Primera edición. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAB). Campeche, México. p. 21-24 Sánchez, T. L. F., Quiróz, F. F., Loyola, V. V. e Infante, D. (2003). Culture-induced variation in plants of Coffea arabica cv. Caturra rojo, regenerated by direct and indirect somatic embryogenesis. Molecular Biotechnology. 23: 107-115.

90
Sánchez-Chiang, N., Jiménez, V. M. (2009). Técnicas moleculares para la detección de variantes somaclonales. Agronomía Mesoamericana. 20(1): 135-151. Sansinforiano, M. E., Rabasco, A., Martínez-Trancón, M., Parejo, J. C., Hermoso de Mendoza M. y Padilla, J. Á. (2001). Optimización de las condiciones RAPD-PCR en Candida spp. Y Cryptococcus spp. Rev Iberoam Micol. 18: 65-69. Scovel, G., Ben-Meir, H., Ovadis, M., Itzhaki, H. y Vainstein, A. (1998). RAPD y RFLP markers tightly linked to the locus controlling carnation (Dianthus caryophyllus) flower type. Theor. Appl. Genet. 96: 117-122. Sharma, U., Rama, R. V., Mohan, J. S. S. y Reddy, A. S. (2007). In vitro propagation of Dendrobium microbulbon A. Rich A rare ethnomedicinal herb. Indian Journal of Biotechnology. 6: 381-384. Sheelavantmath, S. S., Murthy, H. N. , Pyati, A. N., Kumar, Ashok, H. G. y Ravishankar (2000). In vitro propagation of the endangered orchid, Geodorum
densiflorum (Lam) Schltr. through rhizome section culture. Plant cell tissue and organ culture. 60: 151-154. Sheidai, M., Aminpoor, H., Noormohammadi, Z. y Farahani, F. (2008). RAPD analysis of somaclonal variation in banana (Musa acuminate L.) cultivar Valery. Acta Biologica Szegediensis. (2): 307-311. Smulders, M. J. M. (2005). Are there adequate methods for assessing somaclonal variation in tissue culture propagated plants? In: Libiaková, G; Gaidosová, A. eds. Final conference/Cost 843 and Cost 851 (June 28-July 3, 2005, Slovakia). p. 201-203. Soneji, J. R., Rao, P. S y Mhatre, M. (2002). Suitability of RAPD for analyzing Spined and Spineless variant regenerants of Pineapple (Ananas comosus L., Merr.). Plant Molecular Biology Reporter. 20: 307a-307i. Sosa, P., Batista, F. J., Gonzales, M. A y Bouza, N. (2004). VI. La conservación genética de las especies vegetales amenazadas. Universidad de las Palmas de gran canaria. Biblioteca digital. p. 134-160. Stewart, C. N. Jr., y Via, L. E. (1993). A Rapid CTAB DNA Isolation Technique Useful for RAPD Fingerprinting and Other PCR Applications. BioTechniques. 14(5): 748-749.

91
Su, Y. J., Chen, J. T. y Chang, W. C. (2006). Efficient and repetitive production of leaf-derived somatic embryos of Oncidium. Biol. Plantarum. 50(1): 107-110. Sunitibala, H. y Kishor, R. (2009). Micropropagation of Dendrobium transparents L. from axenic pseudobulb segments. Indian Journal of Biotechnology. 8: 448-452. Takashi, H. (1998). Utilization of molecular markers for ornamental plants. Jounal of the Japanese society for horticultural science. 67(6): 1197-1199. Taniguchi, H., Katsumi, M., Yamamoto, Y., Tatsumi, Y., Sano, C. M., Choi, Y-E. y Sano, H. (2008). In vitro proliferation and genetic diversity of Cypripedium
macranthos var. rebunense. Plant Biotechnology. 25: 341–346. Tokuhara, K. y Masahiro, M. (2001). Induccion of embryogenic callus and cell suspension culture from shoot tips excised from flower stalk buds of Phalaenopsis (Orchidaceae). In vitro cell dev. Boil. Plant. 37: 157-161. Tregear, J. W., Morcillo, F., Richaud, F., Berger, A., Singh, R., Cheah, S. C., Hartmann, C., Rival, A. y Duval, Y. (2002). Characterization of a defensin gene expressed in oil palm inflorescences: induction during tissue culture and possible association with epigenetic somaclonal variation events. Journal of Experimental Botany. 53: 1381-1396. Tyagi, R. K., Agrawal, A., Mahalakshmi, C. Hussain, Z. y Tyagi, H. (2007). Low-cost media for in vitro conservation of turmeric (Curcuma longa L.) and genetic stability assessment using RAPD markers. In Vitro Cell.Dev.Biol. Plant. 43: 51–58. Valadez, M. E. y Kahl, G. (2000). Huellas de ADN en genomas de plantas. MP, Mundi-Prensa México, S. A. de C. V. Universidad de Chapingo. Cap. 2 p. 39, 41, 45-53. Vargas, T. E., Xena, N., Vidal, M. C., Oropeza, M. y García, E. (2008). Genetic stability of Solanum tuberosum L. CV. Désirée plantlets obtained from embryogenic cell suspension cultures. 33(3): 213-218. Velasco, M. R. (2005). Marcadores moleculares y la extracción de ADN. Facultad de Ciencias Agropecuarias. 3(1): 14-18. Vij, S. P., Kondo, K. y Pathak, P. (1994). Regeneration potential of Cymbidium pendulum (Roxb) Sw. Nodal explants: a study in vitro. J. Orch Soc India. 8: 19-23.

92
Williams, J. G. K., Kubelik, A. R., Livak, K. J., Rafalski, J. A. y Tingey, S. V. (1990). DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucl Acids Res. 18: 6531-6535. Wilson, D. P. M., Sullivan, J. A., Marsolais, A. A., Tsujita, M. J. y Senaratna, T. (1996). Improvement of somatic embryogenesis in zonal geranium. Plant Cell, Tissue and Organ Culture. 47(1): 27-32. Wu, I-F., Chen, J-T. y Chang, W-C. (2004). Effects of auxins and cytokinins on embryo formation from root-derived callus of Oncidum `Gower Ramsey`. Plant cell tissue and Organ Culture. 77: 107-109. Zambrano, Y. A., Demey, J. R., Martínez, G, Fuenmayor, F., Gutiérrez Z., Saldaña, G. y Torrealba, M. (2002). Método rápido, económico y confiable de, mini preparación de ADN para amplificaciones por RAPD en bancos de germoplasma. Agronomía Tropical. 52(2): 235-243. Zhou, H., Li, S., Qian, X. H. y Gong, H. F. 1995. Effects of some factors on plantlet growth of Dendrobium in tissue culture. Journal of Zhejiang Agricultural University. 21: 622-624.

93
XI. ANEXOS

94
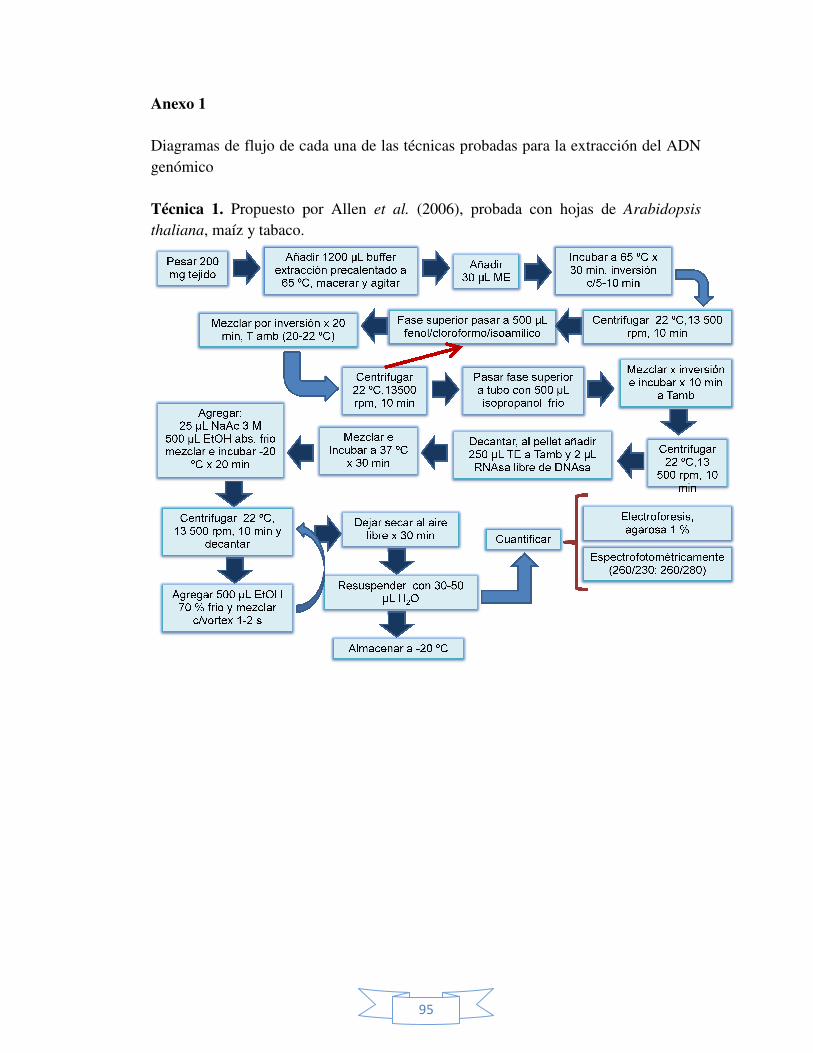
Anexo 1 Diagramas de flujo de cada una de las técnicas probadas para la extracción del ADN genómico Técnica 1. Propuesto por Allen thaliana, maíz y tabaco.
95
Diagramas de flujo de cada una de las técnicas probadas para la extracción del ADN
Propuesto por Allen et al. (2006), probada con hojas de , maíz y tabaco.
Diagramas de flujo de cada una de las técnicas probadas para la extracción del ADN
probada con hojas de Arabidopsis
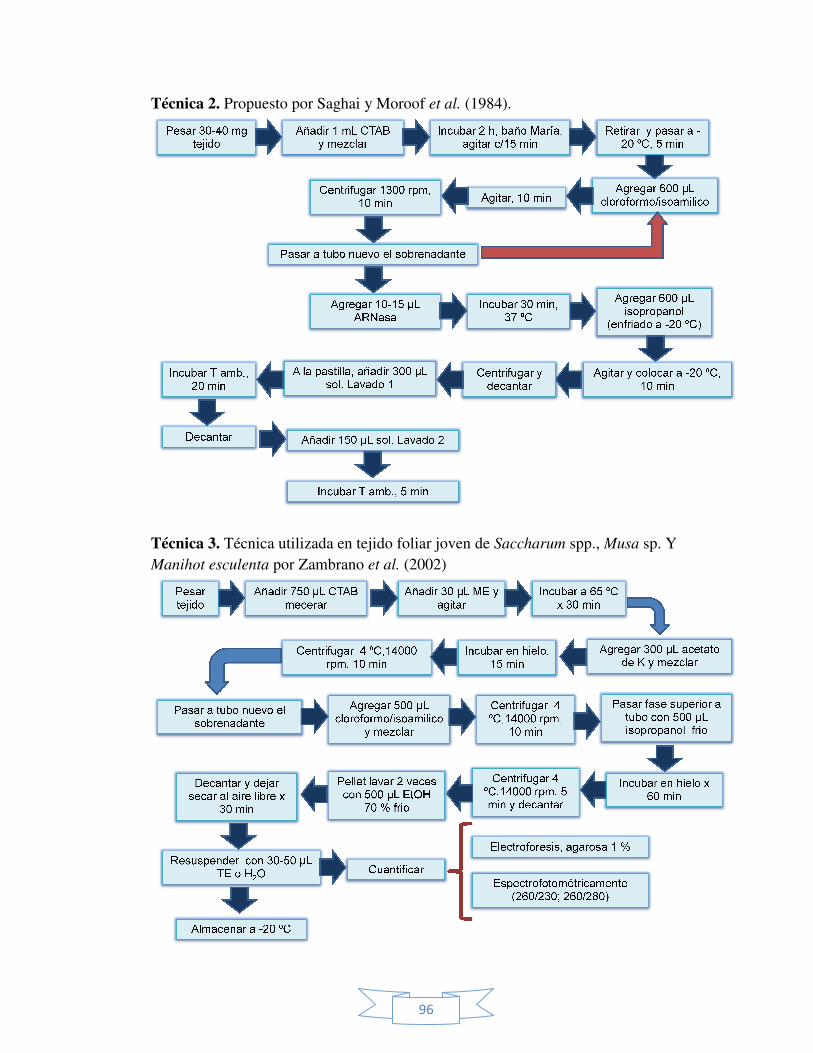
Técnica 2. Propuesto por
Técnica 3. Técnica utilizada en tejido foliar joven de Manihot esculenta por Zambrano
96
Propuesto por Saghai y Moroof et al. (1984).
Técnica utilizada en tejido foliar joven de Saccharum spp., Musa
por Zambrano et al. (2002)
Musa sp. Y
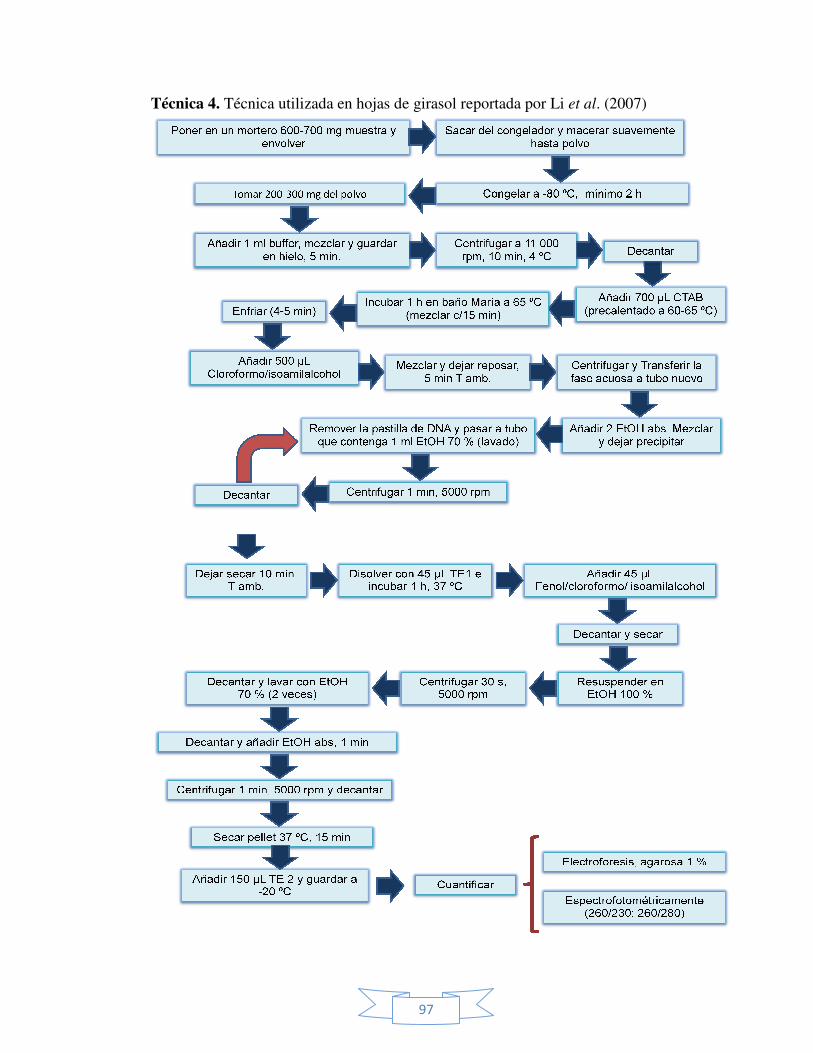
Técnica 4. Técnica utilizada en hojas de girasol reportada
97
Técnica utilizada en hojas de girasol reportada por Li et al. (2007)(2007)

98