essential fatty acids as determinants of lipid requirements in infants, children and adults
TRANSCRIPT

Essential fatty acids as determinants of lipid requirements in
infants, children and adults
R Uauy1,2*, P Mena1 and A Valenzuela1
1Institute of Nutrition and Food Technology (INTA), University of Chile, Casilla 138-11, Santiago, Chile; and 2Retina Foundation of theSouthwest, Dallas, Texas
Essential fatty acids (EFA) are the indispensable component of the lipid supply beyond the provision of energyas a fuel for oxidation. They serve as dietary precursors for the formation of prostanoids and other eicosanoidsthus are of great signi®cance in health and modulation of disease conditions. Eicosanoids are powerful autocrineand paracrine regulators of cell and tissue functions: thrombocyte aggregation, in¯ammatory reactions andleukocyte functions, vasoconstriction and vasodilatation, blood pressure, bronchial constriction, and uterinecontraction. Recent attention has focused on the effect of n-3 and n-6 long chain EFAs in normal fetaldevelopment. Results from human infant studies suggest that n-3 fatty acids are needed for optimal developmentof visual and brain function. Human milk is the best and only time proven source of fat and dietary essential fattyacids for infant feeding. International recommendations for n-3 and n-6 EFA dietary intake are reviewed andsuggested intakes for long chain EFAs are provided.
Basis for essentiality of EFAs, dietary de®cit, structuraland biochemical functional correlations and diseaseprevention
Dietary essential fatty acids (EFA) have long been con-sidered part of the lipid supply necessary for energy,growth, cellular metabolism, and muscle activity. Thefact that some EFAs serve as indispensable dietary pre-cursors for eicosanoid formation has provided greatersigni®cance to the study of their role in health and disease.During the past decade increasing attention has been placedon the effect of n-3 and n-6 EFAs in normal fetal and infantdevelopment (Sprecher, 1981; Bazan, 1989; Willis, 1984;Uauy & Hoffman, 1991; Simopoulos, 1991; Innis, 1991;Uauy et al, 1989). Dietary fatty acid intake also affectscholesterol metabolism throughout the life cycle and isassociated with cardiovascular morbidity and mortality.
Basis for nutritional essentiality of EFAs and derivativesThe concept that speci®c components of fat may benecessary for the proper growth and development ofanimals and possibly humans was introduced in the 1930s(Burr & Burr, 1929). Yet EFAs were considered of mar-ginal importance until the 1960s when signs of clinicalde®ciency became apparent in infants fed skim-milk-basedformula and in those given lipid-free parenteral nutrition.The essentiality of n-6 and n-3 FAs for humans is bestexplained by the inability of animal tissue to introducedouble bonds in positions prior to carbon 9.
Biochemical pathways for n-3 and n-6 FA desaturationare only present in chloroplasts. Terrestrial and marineplants and phytoplankton are the primary source of EFAs
in the food chain; ®sh and other marine animals are able toelongate and desaturate the parent EFAs forming the longchain polyunsaturated fatty acids (LCPUFAs) (Sprecher,1981; Willis, 1984; Simopoulos, 1991). Animal tissues,especially the liver, are capable of further elongating anddesaturating the parent EFAs generating families of com-pounds as shown in Figure 1. As depicted in the ®gure,arachidonic acid [AA (20:4, n-6)] can be formed fromlinoleic acid [LA (18:2, n-6)]; thus it becomes essentialonly if the capacity for elongation and desaturation oflinoleic acid is limited. This is the case in the cat andother felines. Further details on EFA metabolism can befound in recent reviews (Sprecher, 1981; Willis, 1984;Innis, 1991). The competitive desaturation of the n-3, n-6and n-9 series by delta-6 desaturase is of major signi®cancesince this step is controlled by the interaction of hormonesand diet including the amount of LA in the diet. Sprecher'sgroup has further demonstrated that the formation ofdocosahexaenoic acid [DHA (22:6, n-3)], initially thoughtto be catalyzed by a delta-4-desaturase, is indeed a 3-stepprocess as depicted in Figure 1. The initial step is anelongation, followed by delta-6 desaturation, and then,through a peroxisomal beta-oxidation, the chain is shor-tened to a 22 carbon PUFA. This latter step has beentermed retroconversion (Voss et al, 1991). The Sprecherpathway has now been veri®ed for both DHA and docosa-pentaenoic acid [DPA (22:5, n-6)] formation. The delta-6desaturase in the Sprecher pathway is probably differentfrom the enzyme responsible for the initial step of theparent EFA metabolism. If n-3 FAs are absent or de®cientin the diet, the elongation=desaturation of the n-6 com-pounds generates a signi®cant elevation of DPA; if bothEFAs are lacking eicosatrienoic acid [ETA (20:3, n-9)]accumulates (Sprecher, 1981; Uauy & Hoffman, 1991;Innis, 1991). The triene=tetraene ratio may be used as an
*Correspondence: Dr R Uauy, Insitute of Nutrition and Food Technology(INTA), University of Chile, Casilla 138-11, Santiago, Chile.
European Journal of Clinical Nutrition (1999) 53, Suppl 1, S66±S77ß 1999 Stockton Press. All rights reserved 0954±3007/99 $12.00
http://www.stockton-press.co.uk/ejcn
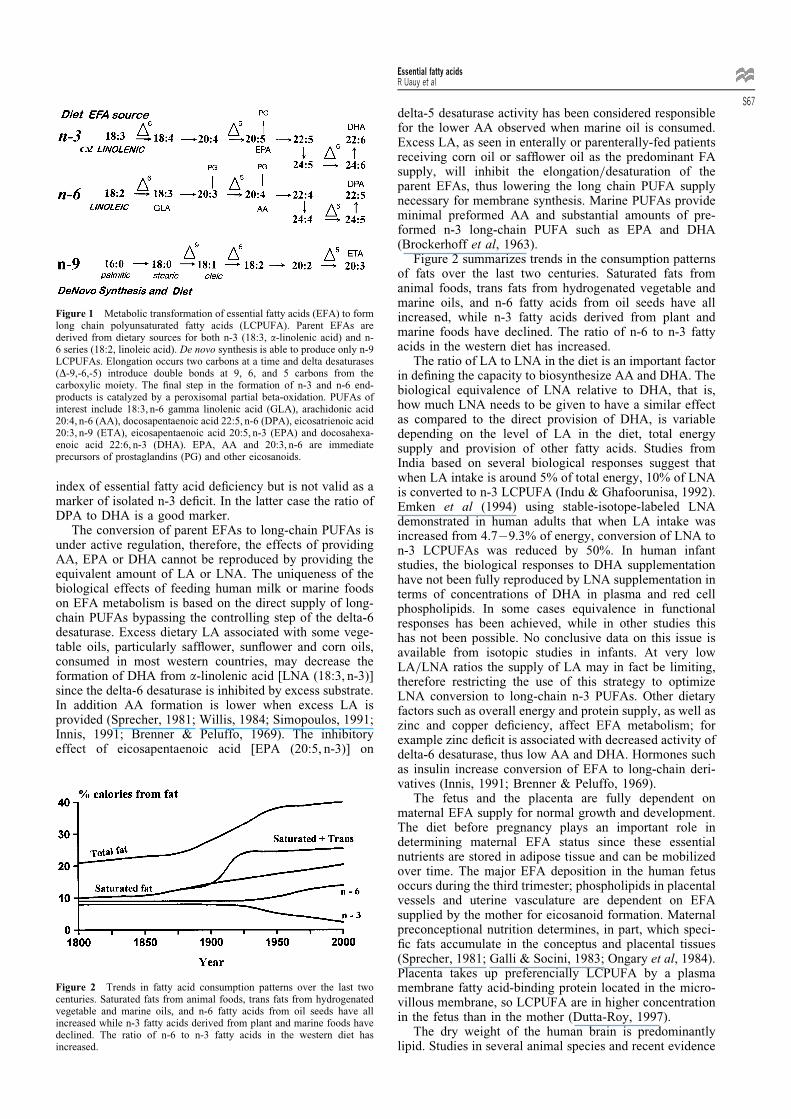
index of essential fatty acid de®ciency but is not valid as amarker of isolated n-3 de®cit. In the latter case the ratio ofDPA to DHA is a good marker.
The conversion of parent EFAs to long-chain PUFAs isunder active regulation, therefore, the effects of providingAA, EPA or DHA cannot be reproduced by providing theequivalent amount of LA or LNA. The uniqueness of thebiological effects of feeding human milk or marine foodson EFA metabolism is based on the direct supply of long-chain PUFAs bypassing the controlling step of the delta-6desaturase. Excess dietary LA associated with some vege-table oils, particularly saf¯ower, sun¯ower and corn oils,consumed in most western countries, may decrease theformation of DHA from a-linolenic acid [LNA (18:3, n-3)]since the delta-6 desaturase is inhibited by excess substrate.In addition AA formation is lower when excess LA isprovided (Sprecher, 1981; Willis, 1984; Simopoulos, 1991;Innis, 1991; Brenner & Peluffo, 1969). The inhibitoryeffect of eicosapentaenoic acid [EPA (20:5, n-3)] on
delta-5 desaturase activity has been considered responsiblefor the lower AA observed when marine oil is consumed.Excess LA, as seen in enterally or parenterally-fed patientsreceiving corn oil or saf¯ower oil as the predominant FAsupply, will inhibit the elongation=desaturation of theparent EFAs, thus lowering the long chain PUFA supplynecessary for membrane synthesis. Marine PUFAs provideminimal preformed AA and substantial amounts of pre-formed n-3 long-chain PUFA such as EPA and DHA(Brockerhoff et al, 1963).
Figure 2 summarizes trends in the consumption patternsof fats over the last two centuries. Saturated fats fromanimal foods, trans fats from hydrogenated vegetable andmarine oils, and n-6 fatty acids from oil seeds have allincreased, while n-3 fatty acids derived from plant andmarine foods have declined. The ratio of n-6 to n-3 fattyacids in the western diet has increased.
The ratio of LA to LNA in the diet is an important factorin de®ning the capacity to biosynthesize AA and DHA. Thebiological equivalence of LNA relative to DHA, that is,how much LNA needs to be given to have a similar effectas compared to the direct provision of DHA, is variabledepending on the level of LA in the diet, total energysupply and provision of other fatty acids. Studies fromIndia based on several biological responses suggest thatwhen LA intake is around 5% of total energy, 10% of LNAis converted to n-3 LCPUFA (Indu & Ghafoorunisa, 1992).Emken et al (1994) using stable-isotope-labeled LNAdemonstrated in human adults that when LA intake wasincreased from 4.7 ± 9.3% of energy, conversion of LNA ton-3 LCPUFAs was reduced by 50%. In human infantstudies, the biological responses to DHA supplementationhave not been fully reproduced by LNA supplementation interms of concentrations of DHA in plasma and red cellphospholipids. In some cases equivalence in functionalresponses has been achieved, while in other studies thishas not been possible. No conclusive data on this issue isavailable from isotopic studies in infants. At very lowLA=LNA ratios the supply of LA may in fact be limiting,therefore restricting the use of this strategy to optimizeLNA conversion to long-chain n-3 PUFAs. Other dietaryfactors such as overall energy and protein supply, as well aszinc and copper de®ciency, affect EFA metabolism; forexample zinc de®cit is associated with decreased activity ofdelta-6 desaturase, thus low AA and DHA. Hormones suchas insulin increase conversion of EFA to long-chain deri-vatives (Innis, 1991; Brenner & Peluffo, 1969).
The fetus and the placenta are fully dependent onmaternal EFA supply for normal growth and development.The diet before pregnancy plays an important role indetermining maternal EFA status since these essentialnutrients are stored in adipose tissue and can be mobilizedover time. The major EFA deposition in the human fetusoccurs during the third trimester; phospholipids in placentalvessels and uterine vasculature are dependent on EFAsupplied by the mother for eicosanoid formation. Maternalpreconceptional nutrition determines, in part, which speci-®c fats accumulate in the conceptus and placental tissues(Sprecher, 1981; Galli & Socini, 1983; Ongary et al, 1984).Placenta takes up preferencially LCPUFA by a plasmamembrane fatty acid-binding protein located in the micro-villous membrane, so LCPUFA are in higher concentrationin the fetus than in the mother (Dutta-Roy, 1997).
The dry weight of the human brain is predominantlylipid. Studies in several animal species and recent evidence
Figure 2 Trends in fatty acid consumption patterns over the last twocenturies. Saturated fats from animal foods, trans fats from hydrogenatedvegetable and marine oils, and n-6 fatty acids from oil seeds have allincreased while n-3 fatty acids derived from plant and marine foods havedeclined. The ratio of n-6 to n-3 fatty acids in the western diet hasincreased.
Figure 1 Metabolic transformation of essential fatty acids (EFA) to formlong chain polyunsaturated fatty acids (LCPUFA). Parent EFAs arederived from dietary sources for both n-3 (18:3, a-linolenic acid) and n-6 series (18:2, linoleic acid). De novo synthesis is able to produce only n-9LCPUFAs. Elongation occurs two carbons at a time and delta desaturases(D-9,-6,-5) introduce double bonds at 9, 6, and 5 carbons from thecarboxylic moiety. The ®nal step in the formation of n-3 and n-6 end-products is catalyzed by a peroxisomal partial beta-oxidation. PUFAs ofinterest include 18:3, n-6 gamma linolenic acid (GLA), arachidonic acid20:4, n-6 (AA), docosapentaenoic acid 22:5, n-6 (DPA), eicosatrienoic acid20:3, n-9 (ETA), eicosapentaenoic acid 20:5, n-3 (EPA) and docosahexa-enoic acid 22:6, n-3 (DHA). EPA, AA and 20:3, n-6 are immediateprecursors of prostaglandins (PG) and other eicosanoids.
Essential fatty acidsR Uauy et al
S67

from humans have established that brain phospholipidDHA decreases while n-9 and n-7 mono- and polyunsatu-rated fatty acids increase when LA and LNA or only n-3fatty acids are de®cient in the diet (Galli et al, 1971; Menon& Dhopeshwarkar, 1982; Bourre et al, 1989a). Typically,n-3 fatty acid de®cient cells have decreased DHA andincreased levels of the end products of n-6 metabolism,docosapentaenoic acid (DPA). Within the subcellular orga-nelles, synaptosomes and mitochondria seem to be moresensitive to a low dietary n-3 supply (Menon & Dhopesh-warkar, 1982; Bourre et al, 1989b).
The high concentrations of long-chain derivativesof EFAs in cerebral cortex and retina suggest theirparticipation in neural and visual function. The n-3 end-product DHA reaches levels of up to 40% of total FAs in thephospholipids of these tissues (Bazan, 1989; Widdowson &Dickerson, 1981; Clandinin et al, 1980). In the human,Clandinin et al (1980) have shown signi®cant increases inthe AA and DHA content of brain tissue during the lasttrimester of gestation. More recently Martinez (1992), usingimproved methodology, has provided complementary dataon retinal and brain accretion of LCEFA.
The potential for dietary EFA de®ciency has become animportant issue in the nutrition of preterm infants sincethese do not receive the third trimester intrauterine supplyof DHA and AA. Even full term infants are potentially atrisk of DHA de®ciency since most formulas are devoid ofthis critical EFA. Recent investigations have demonstratedthat term infants who died suddenly, without a clearexplanation, present a diet-dependent fatty acid composi-tion of their brain cortex. Infants who were breastfed hadhigher DHA and lower DPA and AA in their cortexphospholipids than infants having received cow's milk-based formulas (Farquharson et al, 1992; Makrides et al,1994). DHA content increases with advancing age inbreast-fed infants and is proportional to the duration ofhuman milk feeding, while AA content remains stable.Furthermore the dietary ratio of n-6 to n-3 fatty acidsappears relevant as the lowest DHA and highest DPAcontents in brain cortex phospholipids were observed inpreterm infants fed formulas with a high LA to LNA ratio(Makrides et al, 1994).
Studies in rodents and non-human primates have char-acterized the biochemical and functional effects of dietaryn-3 de®ciency (Wheeler et al, 1975; Watanabe et al, 1987;Neuringer et al, 1984, 1991). The animal models usedpuri®ed LA as the only dietary fat or saf¯ower or sun¯oweroil in which the n-6=n-3 FA ratio (approximately 250:1) isvery high (Neuringer et al, 1984, 1991). High n-6=n-3 FAratios are found in powdered infant formulas currently inuse in some parts of the world (Koletzko & Bremer, 1989;Hansen, 1993). If these diets are fed from conceptionand=or during early life, signi®cant declines in DHAlevels and elevations of DPA content of brain, retina andof other tissues occur in animals. Similar data have beenobtained from plasma, red blood cell (RBC) lipids inpreterm human infants fed corn-oil-based formula with ahigh LA=LNA ratio (Hoffman & Uauy, 1992).
The measurement of the fatty acid composition of RBClipids as a potential index of EFA status in retinal mem-branes is based on the observed correlations between fattyacid pro®les in RBCs and neural=retinal tissue from rats,non-human primates, and human infants (Makrides et al,1994; Carlson et al, 1986; Connor et al, 1993). Yet thesecorrelations are not always signi®cant.
Structural role of EFAs and derived compoundsThe implications of compositional changes in structurallipids have been extensively studied (Wheeler et al, 1975;Bourre et al, 1989c; Stubbs & Smith, 1984). Membranephysiologists have suggested that the fatty acid composi-tion of structural membrane lipids can affect membranefunction by modifying overall membrane ¯uidity, byaffecting membrane thickness, by changing lipid phaseproperties, by speci®c changes in the membrane microen-vironment, or by interaction of fatty acids with membraneproteins (Wheeler et al, 1975; Stubbs & Smith, 1984; Leeet al, 1986). The changes in neural membranes that are ofgreatest potential signi®cance to humans, are changesaffecting physical properties and membrane excitability(Wheeler et al, 1975; Bourre et al, 1989c; Stubbs &Smith, 1984; Lee et al, 1986).
Multiple studies have shown that a de®ciency of essen-tial fatty acids can change physical properties of mem-branes including membrane-bound enzymes, receptoractivity, antigenic recognition, and signal transduction(Stubbs & Smith, 1984; Lee et al, 1986; Salem et al,1988). Also, n-3 fatty acid de®ciency affects rotationalmobility, as measured by ¯uorescent diphenylhexatrieneprobes, less than lateral mobility, as measured by pyrenedimer formation within the lipid bilayer (Stubbs & Smith,1984; Lee et al, 1986; Salem et al, 1988; Treen et al, 1992).We have found that excited dimer (eximer) pyrene forma-tion in human retinal cells, which occurs when two pyrenemonomers collide, is increased by DHA supplementation ofthe culture media (Treen et al, 1992). Membrane composi-tion changes in human retinoblastoma and neuroblastomacell lines also affect membrane transport mechanisms(Treen et al, 1992; Yorek et al, 1984). A higher DHAcontent increases the af®nity and transport rates for cholineand taurine (Treen et al, 1992; Yorek et al, 1984). Further-more, preliminary studies suggest that the dietary n-3 fattyacid supply may affect nucleotide cyclase activity andrybosylation of guanine nucleotide binding proteins(Ahmad et al, 1990).
Most dietary n-3 fatty acid-induced membrane changesare not re¯ected by an overall change in membrane ¯uiditybut rather result in selective changes in membrane micro-environment affecting speci®c domains. The replacementof DHA by DPA usually results in the same overall lipidunsaturation level. Thus ¯uidity on average would remainunchanged. Furthermore, the main changes in the physicalstate, induced by changes in the fatty acid composition oflipid bilayers, occur after the ®rst and second double bondsare introduced; namely when a saturated fatty acid such asstearic acid (18:0) is replaced by oleic acid (18:1, n-9) or bylinoleic acid (18:2, n-6) (Treen et al, 1992; Yorek et al,1984). Others have suggested that DHA supply modi®esthe phospholipid molecular species present in neural tissuesand also gene expression, therefore possibly affecting over-all function (Lin et al, 1990; Clarke & Jump, 1993).Recently Litman & Mitchell (1996) have reported thatLCPUFAs present in membrane phospholipid molecularspecies have profound effects on G protein activationand related structural modi®cations. The rhodopsinMI$MII equilibrium constant is six times higher withdi DHA acylated phosphatidyl choline (PC) than withdi myristic PC, the di DHA PC having an equilibriumconstant that is almost identical to that of native rod disks.The effect is mostly explained by the increase in membranefree volume. This greater mobility within the rhodopsin
Essential fatty acidsR Uauy et al
S68

microenvironment most likely explains the change in Gprotein activation and the corresponding enhanced signaltransduction to photon stimuli (Litman & Mitchell, 1996).
Diet-induced changes in structural lipids affect thefunctional characteristics of excitable membranes in severalanimal species and in human neural cell lines (Wheeler etal, 1975; Neuringer et al, 1984; Bourre et al, 1989b; Loveet al, 1985; Holh & Rosen, 1987). Electrocardiographicabnormalities, such as a notching in the QRS complex,indicating impaired electrical conduction, occur in linoleicand a-linolenic acid de®ciency before clinical signs appear(Caster & Ahn, 1963). Either linoleic or a-linolenic acidcorrected these abnormalities. More recently, it has beenshown that dietary fat in¯uences the susceptibility tocardiac arrhythmias, their incidence and severity (Char-nock, 1991). Furthermore, studies with myocardial prepara-tions have indicated that the vulnerability tocatecholamine-induced arrhythmia is reduced in animalsfed either n-6 or n-3 PUFA-enriched diets (Abeywardena etal, 1987). Feeding ®sh oil from blue®n tuna rather thansun¯ower oil and saturated fat resulted in a marked reduc-tion in induced arrhythmias in two animal species and inisolated papillary muscle (Charnock, 1991). Changes incardiac electrophysiologic responses to beta-mimetics andreduced excitability of cardiac myocytes and in the sus-ceptibility to arrhythmias have also been noted (Hallaq etal, 1992; Kang et al, 1995). Myocytes form minimalamounts of cycloxygenase products and no lipoxygenaseproducts, thus the changes in excitability and conductionare probably related to structural lipid composition, andre¯ect changes in the function of ion channels (Holh &Rosen, 1987). N-3 fatty acid supplementation amelioratesthe ¯uidifying effect of ethanol on neural membranes whilelinoleic and a-linolenic acid de®ciency potentiates volatileanesthetic action in rats; linoleic acid supplementationspeci®cally reverses this effect (Evers et al, 1986).
The role of membrane lipid composition in determiningthe electrical properties of cultured neuronal cells exposedto exogenous fatty acids has been investigated (Lin et al,1990; Love et al, 1985). Both n-3 and n-6 fatty acidsinduced slower rates of rise, and to a lesser extent a loweramplitude, of Na� action potentials. The opposite effectswere observed when saturated or trans monoenoic fattyacids were added. It seems likely that these effects weremediated by a change in the number of active Na�
channels. A change in membrane composition or alteredfatty acid availability to the cells could have caused thisevent (Love et al, 1985). Eicosanoid production is anothermechanism by which the effect of n-3 fatty acid supple-mentation on neural function could be explained. Phos-pholipases liberate AA, EPA, and DHA from membranelipids and make them available to cycloxygenase orlipoxygenase action for production of eicosanoids (Holh& Rosen, 1987).
Models to study the effect of EFAs in `healthy' humans
Fat-free parenteral nutrition as a model to study the effectof EFA de®ciencyThe fact that the use of TPN initially did not include lipids,permitted to con®rm the characteristic acute effects on theskin (Caldwell et al, 1972) and, of greater interest, the long-term effects of EFA de®ciency on sleep organization(Fagioli et al, 1989). Studies in children maintained onfat-free TPN for 2 ± 6 months served to demonstrate
consistently reduced duration of slow-wave sleep comparedwith children who received TPN plus essential lipids. Themain difference was a lower cumulative amount of slow-wave sleep time from the second hour of night-sleeponwards. These results suggest that early lipid nutritionmay modulate sleep regulation and may have implicationsfor whole body energy expenditure during sleep, sinceslow-wave sleep is associated with a lower metabolic ratethan other stages of sleep (Stother & Warner, 1977).Interestingly, no signi®cant difference was found ineither total sleep time or intra-sleep waking in the EFA-de®cient group. Alterations in sleep-wake cycle develop-ment may become evident only if the methods used includeevaluations that are more subtle than total sleep or wakingtime measures. The possibility that fat-free TPN affectedstructural brain lipids is suggested by these results. Inaddition the excess LA relative to LNA in lipids used inparenteral nutrition has also been reported to affect brainfatty acid composition of infants receiving intravenous lipidemulsions in early life. Higher linoleic and lower DHAcontent in brain phospholipid fractions have been reported.The functional effect of this has not been studied (Martinez,1992).
Studies in preterm infants with controlled EFA intakeVery-low-birth-weight (VLBW) infants are particularlyvulnerable to de®ciency given the virtual absence ofadipose tissue stores at birth. This de®ciency may beattributable to a possible immaturity of the FA elonga-tion=desaturation pathways and the inadequate LNA andDHA intake provided by formula. We and others conductedstudies to evaluate the effect of n-3 FA in VLBW infantsexamining the effects of LNA or LNA plus DHA supple-mentation on plasma and tissue lipid composition, retinalelectrophysiologic function, maturation of the visual cortexand measures of infant growth and development (Koletzkoet al, 1989; Uauy et al, 1990; Carlson et al, 1991; Hoffman& Uauy, 1992; Carlson et al, 1993; Birch et al, 1992a,1992b; Werkman & Carlson, 1996).
Infants fed the n-3 FA-de®cient formula, in all studies,had signi®cant reductions in DHA of plasma and red celllipids compared to the groups on other formula (Koletzkoet al, 1989; Uauy et al, 1990; Carlson et al, 1991; Hoffman& Uauy, 1992). DHA-supplemented infants presentedhigher DHA concentrations than those provided LNA butno DHA. In most studies where a human milk fed group hasbeen used as a reference, the DHA�AA-supplementedgroup has matched the LCPUFA blood levels of humanmilk fed infants. Initial studies used marine oil, whichcontains high EPA, as a source of LCPUFA; this lead todrops in AA, which in one study was signi®cantly asso-ciated to poor growth (Carlson et al, 1993).
The functional measures of neurodevelopment haveconsistently shown delayed retinal maturation in theDHA-de®cient groups up to 40 weeks post-conceptionalage; early differences in rod photoreceptor function dis-appeared after this time (Uauy et al, 1990; Birch et al,1992a). Retinal function responses demonstrated signi®-cantly higher threshold values in rod photoreceptors in then-3 de®cient group indicating that the sensitivity andmaturity of the rod photoreceptors in n-3 FA-de®cientinfants were reduced signi®cantly compared to infants fedDHA-supplemented formula or human milk. Small differ-ences in indices of inner retinal function (oscillatorypotentials) persist for up to four months postnatal age
Essential fatty acidsR Uauy et al
S69
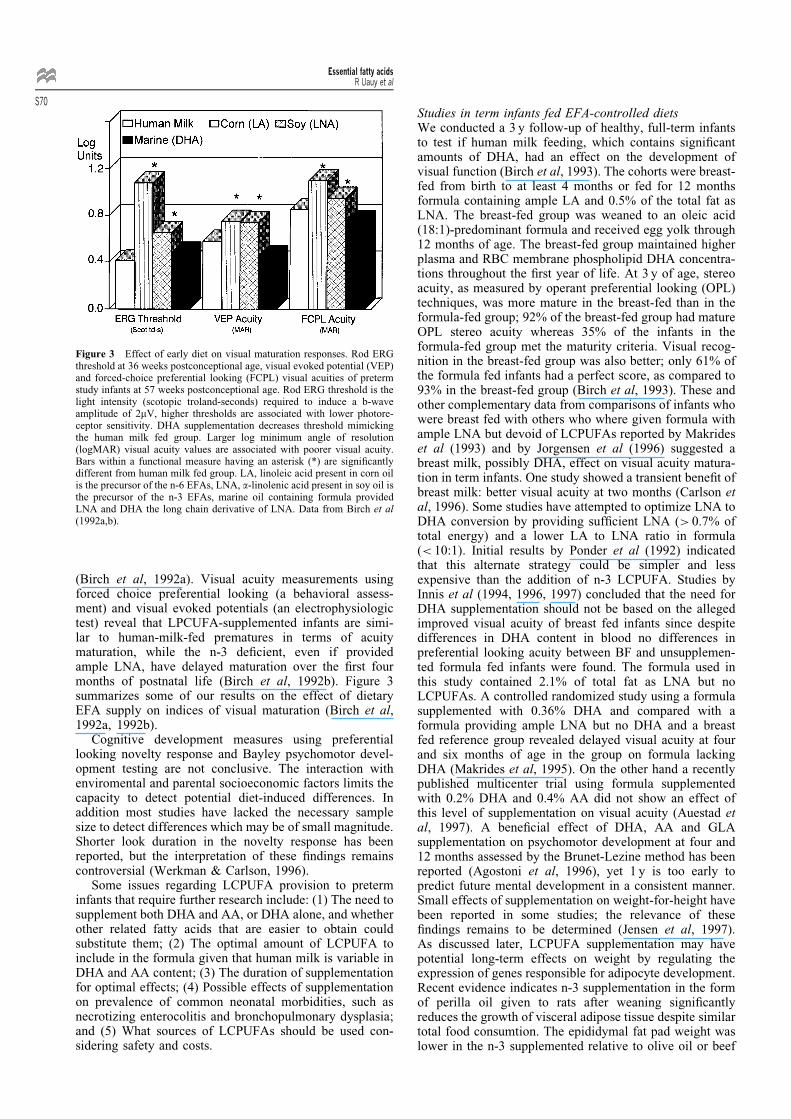
(Birch et al, 1992a). Visual acuity measurements usingforced choice preferential looking (a behavioral assess-ment) and visual evoked potentials (an electrophysiologictest) reveal that LPCUFA-supplemented infants are simi-lar to human-milk-fed prematures in terms of acuitymaturation, while the n-3 de®cient, even if providedample LNA, have delayed maturation over the ®rst fourmonths of postnatal life (Birch et al, 1992b). Figure 3summarizes some of our results on the effect of dietaryEFA supply on indices of visual maturation (Birch et al,1992a, 1992b).
Cognitive development measures using preferentiallooking novelty response and Bayley psychomotor devel-opment testing are not conclusive. The interaction withenviromental and parental socioeconomic factors limits thecapacity to detect potential diet-induced differences. Inaddition most studies have lacked the necessary samplesize to detect differences which may be of small magnitude.Shorter look duration in the novelty response has beenreported, but the interpretation of these ®ndings remainscontroversial (Werkman & Carlson, 1996).
Some issues regarding LCPUFA provision to preterminfants that require further research include: (1) The need tosupplement both DHA and AA, or DHA alone, and whetherother related fatty acids that are easier to obtain couldsubstitute them; (2) The optimal amount of LCPUFA toinclude in the formula given that human milk is variable inDHA and AA content; (3) The duration of supplementationfor optimal effects; (4) Possible effects of supplementationon prevalence of common neonatal morbidities, such asnecrotizing enterocolitis and bronchopulmonary dysplasia;and (5) What sources of LCPUFAs should be used con-sidering safety and costs.
Studies in term infants fed EFA-controlled dietsWe conducted a 3 y follow-up of healthy, full-term infantsto test if human milk feeding, which contains signi®cantamounts of DHA, had an effect on the development ofvisual function (Birch et al, 1993). The cohorts were breast-fed from birth to at least 4 months or fed for 12 monthsformula containing ample LA and 0.5% of the total fat asLNA. The breast-fed group was weaned to an oleic acid(18:1)-predominant formula and received egg yolk through12 months of age. The breast-fed group maintained higherplasma and RBC membrane phospholipid DHA concentra-tions throughout the ®rst year of life. At 3 y of age, stereoacuity, as measured by operant preferential looking (OPL)techniques, was more mature in the breast-fed than in theformula-fed group; 92% of the breast-fed group had matureOPL stereo acuity whereas 35% of the infants in theformula-fed group met the maturity criteria. Visual recog-nition in the breast-fed group was also better; only 61% ofthe formula fed infants had a perfect score, as compared to93% in the breast-fed group (Birch et al, 1993). These andother complementary data from comparisons of infants whowere breast fed with others who where given formula withample LNA but devoid of LCPUFAs reported by Makrideset al (1993) and by Jorgensen et al (1996) suggested abreast milk, possibly DHA, effect on visual acuity matura-tion in term infants. One study showed a transient bene®t ofbreast milk: better visual acuity at two months (Carlson etal, 1996). Some studies have attempted to optimize LNA toDHA conversion by providing suf®cient LNA (> 0.7% oftotal energy) and a lower LA to LNA ratio in formula(< 10:1). Initial results by Ponder et al (1992) indicatedthat this alternate strategy could be simpler and lessexpensive than the addition of n-3 LCPUFA. Studies byInnis et al (1994, 1996, 1997) concluded that the need forDHA supplementation should not be based on the allegedimproved visual acuity of breast fed infants since despitedifferences in DHA content in blood no differences inpreferential looking acuity between BF and unsupplemen-ted formula fed infants were found. The formula used inthis study contained 2.1% of total fat as LNA but noLCPUFAs. A controlled randomized study using a formulasupplemented with 0.36% DHA and compared with aformula providing ample LNA but no DHA and a breastfed reference group revealed delayed visual acuity at fourand six months of age in the group on formula lackingDHA (Makrides et al, 1995). On the other hand a recentlypublished multicenter trial using formula supplementedwith 0.2% DHA and 0.4% AA did not show an effect ofthis level of supplementation on visual acuity (Auestad etal, 1997). A bene®cial effect of DHA, AA and GLAsupplementation on psychomotor development at four and12 months assessed by the Brunet-Lezine method has beenreported (Agostoni et al, 1996), yet 1 y is too early topredict future mental development in a consistent manner.Small effects of supplementation on weight-for-height havebeen reported in some studies; the relevance of these®ndings remains to be determined (Jensen et al, 1997).As discussed later, LCPUFA supplementation may havepotential long-term effects on weight by regulating theexpression of genes responsible for adipocyte development.Recent evidence indicates n-3 supplementation in the formof perilla oil given to rats after weaning signi®cantlyreduces the growth of visceral adipose tissue despite similartotal food consumtion. The epididymal fat pad weight waslower in the n-3 supplemented relative to olive oil or beef
Figure 3 Effect of early diet on visual maturation responses. Rod ERGthreshold at 36 weeks postconceptional age, visual evoked potential (VEP)and forced-choice preferential looking (FCPL) visual acuities of pretermstudy infants at 57 weeks postconceptional age. Rod ERG threshold is thelight intensity (scotopic troland-seconds) required to induce a b-waveamplitude of 2mV, higher thresholds are associated with lower photore-ceptor sensitivity. DHA supplementation decreases threshold mimickingthe human milk fed group. Larger log minimum angle of resolution(logMAR) visual acuity values are associated with poorer visual acuity.Bars within a functional measure having an asterisk (*) are signi®cantlydifferent from human milk fed group. LA, linoleic acid present in corn oilis the precursor of the n-6 EFAs, LNA, a-linolenic acid present in soy oil isthe precursor of the n-3 EFAs, marine oil containing formula providedLNA and DHA the long chain derivative of LNA. Data from Birch et al(1992a,b).
Essential fatty acidsR Uauy et al
S70

tallow fed groups. The expression of genes responsible forlate adipocyte differentiation, adipsin, adipocyte P2 andperoxisome-activated receptor alpha where all down-regu-lated in the perilla oil fed group (Masataka et al, 1997).
This summary of results must lead to a re¯ection onseveral issues: the biological signi®cance of the effects, theselection of outcome measures, the duration and reversi-bility of the change and ®nally the long-term consequencesof the effect. The presence of an effect does not in itselfmake it clinically signi®cant. For example, diet-inducedchanges in triglyceride composition are of little conse-quence except to validate compliance with a particulardiet. Tissue fatty acid pools, particularly those requiredfor retinal and CNS neural membrane formation, may notbe re¯ected by plasma or even red cell fatty acid composi-tion. Biochemical effects can not by themselves be used assurrogates for functional measures.
Another relevant issue is the selected outcome measure.Ideally it should be clinically relevant, but the sensitivity ofclinical responses is usually low, therefore functionalresponses are considered valid to de®ne biologically sig-ni®cant outcomes. In the case of LCPUFA supplementa-tion, growth is affected only in extreme n-6 fatty acidde®ciency therefore is not considered a sensitive measureof n-6 suf®ciency. The effect of n-3 fatty acids on sensorymaturation and cognitive development are the outcomes ofgreater interest in studies of n-3 supplementation.
The duration and reversibility of diet-induced effects isanother important issue. There may be transient effects thatre¯ect the acceleration or the slowing of a maturationalprocess with a fully normal ®nal outcome. This is of specialrelevance during the ®rst few months of life when visualmaturation is progressing rapidly. Several studies havedemonstrated signi®cant effects of the dietary LCPUFAson visual maturation in the ®rst four months of life, but inmost cases the delayed response becomes normal at sixmonths or at most by 1 y of age. Should we dismiss thisphenomenon as transitory and of limited signi®cance orassume that we failed to detect a signi®cant change at 1 ybecause our tools were not sensitive enough, or that in factother related function are indeed affected? Using this sameexample, we failed to detect differences in visual acuity atsix months but depth=spatial perception assessed bystereoacuity responses were different at 3 y of age, asshown by our studies comparing breast fed to routineformula fed infants. These examples illustrate that unlesssensitive outcome measures are used and suf®cient follow-up time is provided, it is impossible to fully discard thepossible long-term consequences of early developmentaleffects.
Evidence of potential bene®cial long-term effects ofLCPUFA supplementation on brain development of terminfants is suggestive, proof is lacking. The resolution of thisissue should be forthcoming, the follow-up of term infantsincluded in the controlled clinical trials of DHA�AAsupplementation into the school age period should help toaddress the question of long-term effects (Dobbing 1997).
Studies of essential fatty acid metabolism using labeledprecursorsWe and others have also conducted studies using labeledEFA precursors (Salem et al, 1996; Sauerwald et al, 1996).Our stable isotope studies have used as a label 5 deuteratedatoms distributed in the ®rst (n-1) and second (n-2) carbonatoms of the fatty acid chain (d5-LA and d5-LNA) to
evaluate the conversion of these into LCPUFA (Salem etal, 1996). Concentration of deuterated precursors andproducts were measured in plasma using negative ionmass spectrometry of penta¯uorobenzyl derivatives. Peakconcentrations of labeled precursor EFAs were reachedduring the ®rst day after dosing, deuterated productsincreased concentrations with time reaching peak levelsfor the n-6 metabolites by 48 h in the term infants andcloser to 96 h in the preterm SGA or AGA infants. For then-3 series, concentrations of elongated=desaturated pro-ducts increase with time reaching highest levels in theterm infants by 48 h; for preterm infants peak levels arereached by 96 h. The rise in labeled products for SGAinfants is slower than that for AGA preterm infants. Otherstudies using uniformly labeled 13C con®rm that pretermand term neonates are able to elongate and desaturateparent EFAs (Sauerwald et al, 1996; Carnielli et al,1996). Estimation of pool size, relevant precursor poolenrichment, knowledge of homogeneity in distribution inone or more pools and information on rates of oxidation ofprecursors and products are lacking in these studies. There-fore, they have not been able to provide quantitativeinformation on synthetic rates in order to orient dietaryrecommendations. Earlier studies in adults have also docu-mented n-3 LCPUFA formation from labeled LNA but noconversion of LA to arachidonic was documented (Emkenet al, 1992). El Boustani et al (1989) reported delta 5desaturation and arachidonic acid formation in four humanadults using deuterated 20:3, n-6. More recently Emken etal (1997) evaluated the in¯uence of AA on delta 5desaturation concluding that AA supplementation inadults increased Dihommo GLA (20:3, n-6) turnover anddesaturation into AA.
Studies in malnourished infantsLow-fat diets are a frequent cause of primary malnutritionin infants and children in developing countries. In additionthese diets are of low energy density and therefore limit thetotal energy a child is able to consume in a single meal.There is very limited data on EFA composition of localdiets used to feed infants as well as on EFA status ofmalnourished infants. If children are not breast fed mostlikely they will receive insuf®cient EFAs unless they aregiven appropriate complementary foods. Cereals are low infat and most oil seeds will provide only parent EFAs LArich and LNA poor, unless soy or rapeseed oil are available.Starchy roots, tubers and starchy fruits used as traditionalcomplementary foods are low in fat and low in EFAs.Powdered cow milk used as a source of high quality proteinis also low in EFA and provides virtually no n-3 fatty acids;therefore malnourished children are at risk for EFA de®-ciency and specially n-3 de®cit. The risk for n-3 de®cit isenhanced if during the recovery period infants are givenenergy-dense diets high n-6 fatty acids. These diets areusually formulated with vegetable oils that are high in n-6and low in n-3 fatty acids (corn, sun¯ower, saf¯ower)(Beare-Rogers et al, 1998).
The breast fed infant will have suf®cient n-6 fatty acidsup to 8 and probably up to 12 months of age. If we considerthat the n-3 fatty acid supply should be at least 0.5% ofenergy, the breast fed infant will have suf®cient n-3 EFAfrom human milk for the ®rst six months of life if the milkvolume is 800 ± 1000 mL=d and the fat content is 3 ±4 g=dL. If the mother is malnourished, breast milk fat,EFA content and total volume may be reduced; therefore
Essential fatty acidsR Uauy et al
S71

n-3 may be below accepted levels at an earlier age (Beare-Rogers et al, 1998).
Prevention of chronic disease in relation to EFAs supplyPublished experimental work indicates that consumption ofterrestial and marine foods with n-3 PUFA has a bene®cialeffect on primary and secondary prevention of athero-sclerosis, thrombosis and embolism, hypertriglyceridemia,hypertension, autoimmune disease and possibly allergicproblems. Bleeding time in the human can be prolongedby 30 ± 50% by supplementing diets with marine oils;platelet aggregation can also be reduced. These effectscan be explained by changes in eicosanoid metabolism,shifting it from the arachidonate-derived series2 metabo-lites to the EPA-derived series3 metabolites. Prostaglan-dins, thromboxanes, leukotrienes, hydroxy fatty acids andlipoxins formed from EPA decrease blood viscosity, plate-let aggregation, increase bleeding time, promote vasodila-tion and insulin sensitivity; in addition they inhibit VLDLformation, cell proliferation, the in¯ammatory and allergicresponse. Effects are evident with doses ranging from 5 ±30 g of marine oils or 2 ± 10 g of EPA per day (Uauy &Hoffman, 1991; Innis, 1991; Uauy et al, 1989; Glomset,1985; Simopoulos, 1991; Galli et al, 1994; Katan, 1995;Bang & Dyerberg, 1972, 1986). Figure 4 summarizes theeffects of n-3 polyunsaturated fatty acids inhibiting keysteps of the arachidonic acid cascade. Biological effects aredependent on n-6=n-3 balance present in membrane phos-pholipids.
Recent reviews of the vast body of literature accumu-lated over the past decade are available. The conclusionsreached by the reviewers differ based on their biases and onthe selection of evidence presented. Simopoulos concludesthat `research reports make it increasingly evident thateating o-3 fatty acids can have bene®cial effects on chronicin¯ammatory and cardiovascular diseases' (Simopoulos,1991; Galli et al, 1994). A more recent review by Okuyamaet al, 1997 from Japan (491 refs) concludes: `It appears tous that an excess of linoleic acid and elevated n-6=n-3ratios in tissues result in over- and unbalanced production
of lipid mediators related to arachidonic acid, and that thissituation leads to enhanced ischemic=in¯ammatory ten-dencies thereby increasing the incidence of chronicdiseases . . . '. It goes on to say: `We suggest that a relativen-3 de®ciency as evidenced by the very high n-6=n-3 ratiosof plasma lipids might be affecting the behavioral patternsof a signi®cant part of the young generations in industria-lized countries.' (Okuyama et al, 1997). In the opposingcamp, the FDA, after an extensive review of this subject aspart of its effort to de®ne health claims for food labelingpurposes, concluded that most of the documented bene®tswere attributable to the consumption of ®sh as a food andnot of marine oil supplementation (FDA=HHS USA, 1991).The verdict is being contended by several groups, amongstthem the International Society for Study of Fatty Acids andLipids (ISSFAL) (Galli et al, 1994). The issue of whetherthe bene®cial effects are secondary to n-3 fatty acids orother phenomena associated with the consumption of ®shwill only be settled by prospective randomized controlledclinical trials of speci®c fatty acids, alone or in combina-tion.
Omega-3 PUFA ingestion can have an effect on cardi-ovascular disease through a variety of mechanisms. Theexperimental evidence does not permit deciding which ofthe many putative mechanisms are crucial in determiningthe effects. Marine oils have antithrombotic effects viamodi®cation of platelet aggregation, decreasing vascularadhesiveness, enhancing small vessel relaxation mechan-isms, promoting endothelial repair and minimizing in¯am-matory responses in vessel walls (Simopoulos, 1991; Galliet al, 1994; Shimokawa & Vanhoutte, 1988; Leaf & Weber,1988). In addition, the risk of heart disease can be indir-ectly modi®ed by effects on plasma lipoproteins, choles-terol uptake by macrophages, foam cell formation and cellproliferation in vessel walls, blood pressure regulation,changes in cardiac electrophysiologic responses to beta-mimetics, in the susceptibility to arrhythmias and reducedexcitability of cardiac myocytes (Charnock, 1991; Connor& Connor, 1990; Dyerberg et al, 1978; Harris et al, 1988;Knapp & FitzGerald, 1989; Kang et al, 1995). In aprospective randomized controlled trial of ®sh oil as anantiarrythmic agent, patients with spontaneous ventricularpremature complexes (VPC) were given ®sh oil(0.9 g EPA� 1.5 g DHA) or n-6-rich sun¯ower oil(5 g LA) for 16 weeks. VPCs were signi®cantly reducedin 44% of patients of the ®sh oil group and only in 15% ofthe sun¯ower oil group (Sellmayer et al, 1995). The otherintervention trial used the Mediterranean diet, rich in LNAsources (2 g=d) as a model. Over 600 French patients whosurvived their ®rst infarcts where randomly assigned to thisor the control diet providing only a third of the LNA. Thegroup on the LNA-rich diet had 73% fewer cardiac deathsand non fatal heart attacks than controls (De Lorgeril &Renaud, 1994). A recent report from the Chicago WesternElectric Study to examine the possible bene®t of eating ®shon risk of dying from coronary heart disease showed inover 1800 `healthy' middle aged men that consumption of35 g or more ®sh per day signi®cantly reduced the cumu-lative 30 y risk of death from coronary heart disease by38% and that of myocardial infarction by 44% (Daviglus etal, 1997).
The mechanism for the antiarrythmic effect has beenstudied by Leaf's group who has demonstrated that myo-cites become less excitable and more resistant to betaadrenergic stimuli due to a modulation of Na channels.
Figure 4 Effect of n-3 polyunsaturated fatty acids (n-3 PUFA), inhibitingkey steps of the arachidonic acid (n-6) cascade. Phospholipases releasearachidonate from membranes which serves to synthesize eicosanoids; n-6derived eicosanoids formed by cycloxigenase pathway (prostaglandins,prostacyclins, thromboxanes) or lipoxigenase pathway (leukotrienes) arecounterbalanced by n-3 PUFA derived eicosanoids. Biological effects aredependent on n-6=n-3 balance present in membrane phospholipids.
Essential fatty acidsR Uauy et al
S72

The effect is observed with n-6 and n-3 PUFA, but is moreprominent in the latter (Kang & Leaf, 1995).
Initially it was thought that marine oils had an effect onplasma cholesterol comparable to that of n-6 predominantvegetable oils, however subsequent studies have demon-strated that only n-3 long-chain PUFA lower triglyceridesin normal or hyperlipemic subjects (Harris, 1989). Patientswith type V hyperlipidemia have shown dramatic drops inVLDL and triglyceride levels in response to marine oilsupplemented diets (Harris, 1989; Nestel et al, 1984). Incontrast, n-6 fatty acids from oil seeds caused a signi®cantrise in triglycerides in this group. The effect on triglycer-ides appears to be mediated by decreased production andincreased fractional catabolic rate of the triglyceride-richVLDL lipoprotein particle (Nestel et al, 1984). Small butsigni®cant reductions in plasma cholesterol have also beennoted after ®sh oil administration in patients with type IIheterozygous hypercholesterolemia, while in those withtype IV a rise has been noted (Simopoulos, 1991; Nestelet al, 1984; Massarei et al, 1989). The evidence does notsupport an LDL cholesterol lowering effect in response tomarine oil supplementation in patients with hypercholes-terolemia. The response of HDL-cholesterol and apoproteinA in supplementation trials has been variable but thechanges, if any, are of minor magnitude. The variabilityin lipoprotein responses to supplementation in the varioustrials can be attributed to the differences in overall design,duration of study, patient populations, differences in base-line diets tested, and different oils used. Some studies used®sh or marine oils while others used concentratedEPA�DHA (Simopoulos, 1991; FDA=HHS USA, 1991;Leaf & Weber, 1988; Harris, 1989; Nestel et al, 1984).Overall these studies suggest a bene®cial effect of ®sh andn-3 LCPUFAs on coronary disease mortality.
Results of studies on the use of marine oil in theprevention of restenosis after coronary angioplasty orbypass surgery were suggestive of bene®cial effects of acombined use of ®sh oil and anticoagulant drugs in redu-cing thrombotic occlusion of coronary vessels. Furthermoredouble-blind controlled studies using angiographic evi-dence for presence of restenosis after coronary angioplastyhave failed to disclose the putative bene®ts (Leaf et al,1994). A more recent randomized controlled study on graftpatency after bypass surgery using 4 g=d of ®sh oil inaddition to anticoagulants demonstrated a clear bene®t interms of graft patency 1 y after surgery. The ®sh oil grouphad only 27% graft occlusion vs 33% in the control group,an inverse relationship between increase in n-3 content ofplasma phospholipids and vein graft occlusion was noted(Eritsland et al, 1996).
EPA and DHA prolong bleeding time and decreasethromboxane A2 production. These effects seem to bedose dependent but to date no major clinically signi®cantbleeding has been demonstrated in relation to the ingestionof ®sh oils (Lox, 1990), although one study showedincreased incidence of epistaxis when high dose ®sh oilwas administered to hypercholesterolemic adolescents(Clarke et al, 1990). Platelet counts may be affected bylarge doses of marine oil and platelet half-life may beshortened. The effect on endothelial walls is receivingincreasing attention. The production of platelet-derivedgrowth factor is decreased while the formation of endothe-lium derived relaxing factor is increased (Fox & Dicor-leto,1988). These effects may be important in mediating theantiatherogenic effect of ®sh oil.
Fish oil has also been noted to signi®cantly reducesystolic blood pressure in double-blind crossover controlledstudies. Increase in sodium excretion and a decrease inprostaglandin metabolites derived from AA were alsofound. The lowering of diastolic blood pressure, of greaterclinical signi®cance, has been the exception. The interac-tion of marine oil supplementation and need for antihyper-tensive drugs has been less explored. A meta analysisconducted by Morris et al (1993) involving a total ofover 1300 patients concluded that dietary n-3 fatty acidsinduce a small but signi®cant reduction in blood pressure inhypertensive but not in normal subjects. This area requiresfurther research before the use of marine oils can beadvocated for this purpose (Levinson et al, 1990).
In adult onset diabetes the effects are mixed since thepotentially bene®cial effect on microvessel occlusion andproliferation is offset by a potential increase in glucoseoutput and higher serum glucose levels observed in somestudies. Additionally total cholesterol, LDL and apo Bserum levels are increased by ®sh oil supplementation indiabetics. On the other hand recent evidence suggests thatinsulin resistance of adipose tissue may be ameliorated byn-3 and by n-9 fatty acids (Ranaian et al, 1996). In additiontriglyceride levels will commonly decrease with n-3LCPUFA supplements. A major problem in the interpreta-tion of metabolic effects of PUFAs is the genetic hetero-geneity of patients included in these studies; the responsemay in fact be different within a group that appears to be ofa similar phenotype. This may be due to genetic factorswhich are only beginning to be unraveled. A study inpatients with coronary heart disease and normal volunteersdemonstrated that hyperinsulinemia, thus insulin resistance,is inversely correlated with long-chain PUFA content ofmuscle cell membrane phospholipid. The lower the AA andDHA content of membranes the higher the insulin level.This association could be related to dietary EFA content butalso both conditions could be determined by a commoncause since LCPUFA biosynthesis is affected by insulinactivity (Storlien et al, 1996). No prospective controlledstudy has addressed this point, thus for now we can onlyspeculate. In terms of mechanisms, it is well known thatPUFAs promote fatty acid oxidation and in the case of n-3LCPUFAs decrease VLDL secretion from the liver. Thepossibility that speci®c fatty acids promote or inhibitperoxisomal proliferation can be raised given the discoveryof peroxisomal proliferation activator receptor (PPAR) andthe modulation of its level of expression by speci®c fattyacids (Masataka et al, 1997; Schoonjans et al, 1996).Increase in saturated fat and=or trans fatty acids relativeto n-3 LCPUFA affect PPAR activity thus could potentiallyadversely affect fatty acid oxidation and prevent the bio-synthesis of LCPUFAs by affecting peroxisomes, key sitefor LCPUFA synthesis from parent EFAs (Martinez, 1996).The main regulatory actions of PPAR on lipid metabolismare summarized in Table 1. The putative regulatory effects
Table 1 Regulatory actions of the peroxisome proliferator-activatedreceptor (PPAR) in intracellular and extracellular lipid metabolism
Activation of peroxisomal and mitochondrial b-oxidationRegulatory action on transcription of HDL apolipoproteinsControl of adipocyte differentiation and adipogenesisStimulation of genes that control lipoprotein lipase activityIncreased expression of fatty acid transport proteinDecreased production of VLDL and fatty acid synthesis
Essential fatty acidsR Uauy et al
S73

of dietary fatty acids on mitochondrial and peroxisomalfunction has led to the proposed hypothesis summarized inFigure 5. This could be central in explaining the effect ofdietary fat on the epidemic of non communicable chronicdisease observed in industrialized countries and in somesegments of developing societies (Simopoulos, 1994).
The use of omega-3 marine oils as a natural immuno-suppressant has been proposed based on their inhibitoryrole on arachidonic acid derived products such as prosta-glandins and leukotrienes. Several studies on experimentalanimals have shown that ®sh oil increased survival ofanimals genetically predisposed to lupus erythematosusand collagen-induced arthritis (Simopoulos, 1991; Galli etal, 1994). Controlled clinical studies in humans haveprovided encouraging preliminary results; the effect of®sh oil are probably mediated by a suppression of LTB4
and increased LTB5 formation. IL-1 production is loweredby o-3 LCPUFAs while IL-2 is increased. The role of ®shoil alone or in combination with antirheumatic medicationremains to be determined. Another potential use of marinePUFA being explored is the induction of an immunosup-pressed state to prevent the development or progression oftype I insulin dependent diabetes in susceptible individuals.Other uses documented by anecdotal evidence or uncon-trolled trials include bene®ts in the treatment of migraine,asthma and other allergic conditions and in the control ofpsoriasis (Simopoulos, 1991; Galli et al, 1994).
Studies of requirements under disease conditions
Metabolic=genetic diseaseThere is yet no conclusive evidence of an inherited meta-bolic defect of DHA and=or AA synthesis causing alteredbrain or retinal function. However, there is strong evidencethat Zellweger and other peroxisomal diseases may beassociated with defective formation of C 22 PUFAs.Martinez (1996) and others have provided data fromseveral patients, indicating that in peroxisomal disorders,brain DHA content is extremely low, that plasma and RBC
DHA levels are below normal and that the biochemicalresponse of these patients to ®sh oil test material is onlypartial. Sprecher (1981) has reported that the peroxisomalbeta oxidation step necesary for C22 PUFA biosynthesis isrestricted in Zellweger patients, thus they can not convert C24:6, n-3 to C 22:6, n-3. Preliminary results on a limitednumber of patients studied by Martinez indicate thatsupplementation with pure DHA is capable of preventingand also of reverting the abnormal motor and mentalfunction associated with these conditions. Concomitant®broblast studies of these patients provide supportiveevidence as to the relative importance of peroxisomalbeta oxidation in determining the biochemical and func-tional consequences of this disease (Martinez, 1996).
Other potentially adverse health effectsThe vulnerability of lipid membranes to damage by oxygenfree-radicals is directly related to their degree of unsaturation.The double bonds in the carbon chain of the fatty acids are thepoint of attack for oxygen free radicals. The number ofpentadienic units present in a polyunsaturated fatty acid isdirectly related to its rate of oxidation (Willis, 1984). There-fore membranes enriched with AA, EPA and DHA are mostsusceptible to develop oxidative damage. The oxidationprocess is further catalyzed by the products of oxidation.This self-propagating process may severely affect membranestructure and function unless quenched by proper antioxidantsin the membrane. The protection of LCPUFAs is dependenton enzymatic and non-enzymatic systems. Ingestion of highdoses ofo-3 fatty acids in adults has raised the question of riskfor tissue oxidative damage. This effect has been observed inexperimental animals by several authors. Moreover, agingappears to be an important factor in the development of thissusceptibility. Aged rats, fed ®sh oil show a higher suscept-ibility to develop tissue lipid peroxidation than young ones(Garrido et al, 1989, 1993).
Evidence from human studies in young infants suggeststhat excess dietary PUFAs will increase red cell hemolysisunless appropriate tocopherol supplementation is provided(Oski & Barness, 1967). Experimental evidence indicatesthat hyperoxic-induced lung damage may be affected bydietary fat, although the results from mice exposed tooxygen immediately after birth indicate that PUFAs andspeci®cally ®sh oil feeding protect from oxygen-induceddamage. The proposed explanation for this apparent para-dox is that PUFAs in plasma triglycerides may trap freeradicals, thus protecting membrane phospholipid PUFAs(Sosenko et al, 1989). The role of o-3 LCPUFAs indetermining the increased susceptibility of the retina tolight-induced oxidative damage has been recently studieddemonstrating that lower retinal DHA protects rod outersegments from light-induced damage. Tocopherol has beenshown to be protective (Bush et al, 1991). The role ofPUFAs and immature antioxidant systems in determiningoxygen-induced lung, eye and red cell damage in prematureinfants is an area of great interest. Increased risk ofbronchopulmonary dysplasia and retinopathy of prematur-ity in preterm infants of Alaskan eskimo background havebeen observed (Arnold, 1992). The high o-3 diets and lowconsumption of green vegetables or other sources oftocopherol may play a role in this phenomenon (Wo &Draper, 1975).
Increased bleeding is a potentially adverse effect offeeding o-3 fatty acids. The clinical implications of thesehave not been demonstrated except for anecdotal evidence
Figure 5 Speculative mechanisms to explain the effect of dietary fattyacids on b-oxidation and LCPUFA biosynthesis and insulin resistance.These effects could explain the effect of fat on diet related non commu-nicable chronic diseases, obesity, hypertension, non insulin dependentdiabetes and hyperlipidemia.
Essential fatty acidsR Uauy et al
S74

of frequent bruising occurring among ®sh oil eating Green-land eskimos (Dyerberg & Bang, 1979). Clinical studieshave demonstrated increased epistaxis in adolescents givenhigh doses of ®sh oil. On the other hand there is evidencefor small non-signi®cant changes in bleeding time in infantsgiven a marine oil supplemented formula (Clarke et al,1990; Uauy et al, 1994). The use of marine oils has somepotential toxicological risks that can be circumvented bycareful processing, storing and preservation of the unsatu-rated fatty acids.
Conclusions
Infants should be breast fed if at all possible. The fatty acidcomposition of infant formula should correspond to theamount and proportion of fatty acids contained in maturebreast milk from healthy omnivorous women.
During weaning and at least until 2 y of age, a child'sdiet should contain at lest 30 ± 40% of energy from fat andprovide similar levels of essential fatty acids as are found inhuman milk (FAO=WHO 1994). The n-6 and n-3 fatty acidshave critical roles in the membrane structure and as pre-cursors of eicosanoids, which are potent and highly reactivecompounds. Since they compete for the same enzymes andhave different biological roles, balance between n-6 and n-3fatty acids in the diet can be of considerable importance.
Desirable intakes of linoleic acid should provide between4 and 10% of energy (FAO=WHO 1994). Intakes at theupper end of this range are recommended when intakes ofsaturated fatty acids and cholesterol are relatively high.
Particular attention must be paid to promoting adequatematernal intakes of essential fatty acids throughout preg-nancy and lactation to meet the requirements of fetal andinfant development.
ISSFAL suggests an intake of 0.8 ± 1.1 g=d of LNA,which is roughly 0.5% of energy; it also suggests 0.3 ±0.4 g=d of n-3 LCPUFAs. To prevent cardiovascular andother non-communicable chronic diseases the recommenda-tion is to eat ®sh 2 ± 3 times a week. Alternatively marineoil 3 ± 4 g per day or n-3 LCPUFAs (EPA�DHA) 1.2 g perday or 0.5% of energy have been recommended by theBritish Nutrition Foundation.
Acknowledgements ÐWork by the authors of this publication is supported
in part by Fondecyt grant 1961001 and a CONICYT Presidential Award in
Biological Sciences to RU.
References
Abeywardena MY, McLennan PL & Charnock JS (1987): Diet and cardiacarrhythmia: involvement of eicosanoids. In Proceedings of the AOCSShort Course of Polyunsaturated Fatty Acids and Eicosanoids, ed.WEM Lands. Illinois: American Oil Chemist's Society, pp 62 ± 64.
Agostoni C, TrojaÂn S, Bellu R et al (1996): LCPUFA status and develop-mental quotient in term infants fed different dietary sources of lipids inthe ®rst months of life. In Recent Developments in Infant Nutrition, eds.JC Bindels, AC Goedhart & HKA Visser, pp 212 ± 217, Kluwer Aca-demic Publishers.
Ahmad SN, Alam SQ & Alam BA (1990): In¯uence of dietary omega-3fatty acids on transmembrane signalling in rat submandibular salivarygland. Cell. Signal. 2, 29 ± 41.
Arnold RW (1992): Retinopathy of prematurity in Alaska. Proceedings ofthe 19th Annual Providence Hospital Perinatal Symposium. AnchorageAlaska, March 1992.
Auestad N, Montalto MB, Hall RT, Fitzgerald KM, Wheeler RE, ConnorWE, Neuringer M, Connor SL, Taylor JA & Hartmann EE (1997):Visual acuity, erythrocytes fatty acid composition, and growth in terminfants fed formulas with and without long chain polyunsaturated fattyacids for one year. Pediatr. Res. 41, 1 ± 10.
Bang HO & Dyerberg J (1972): Plasma lipids and lipoproteins in Green-landic West-coast Eskimos. Acta Med. Scand. 192, 85 ± 94.
Bang HO & Dyerberg J (1986): Lipid metabolism and ischemic heartdisease in Greenland eskimos. Adv. Nutr. Res. 3, 1 ± 21.
Bazan NG (1989): The metabolism of omega-3 polyunsaturated fatty acids intheeye: thepossible roleofdocosahexaenoicacid and docosanoids in retinalphysiology and ocular pathology. Prog. Clin. Biol. Res. 312, 95 ± 112.
Beare-Rogers J, Ghafoorunissa, Korver O, Rocquelin G, Sundram K &Uauy R (1998): Dietary fats in developing countries. Food and Nutr.Bull. 19, 251 ± 267.
Birch D, Birch E, Hoffman D & Uauy R (1992a): Retinal development invery low birth weight infants fed diets differing in omega-3 fatty acids.Invest. Ophtal. Vis. Sci. 33, 2365 ± 2376.
Birch EE, Birch DG, Hoffman DR & Uauy R (1992b): Dietary essentialfatty acid supply and visual acuity development. Invest. Ophthal.Vis.Sci. 33, 3242 ± 3253.
Birch E, Birch D, Hoffman D, Hale L & Uauy R (1993): Breast-feedingand optimal visual development. J. Pediatr. Ophtalmol. Strabismus 30,33 ± 38.
Bourre JM, Durand G, Pascal G & Youyou A (1989a): Brain cell and tissuerecovery in rats made de®cient in n-3 fatty acids by alteration of dietaryfat. J. Nutr. 119, 15 ± 22.
Bourre JM, Francois M & Youyou A (1989b): The effects of dietary alpha-linolenic acid on the composition of nerve membranes, enzymaticactivity, amplitude of electrophysiological parameters, resistance topoisons and performance of learning tasks in rats. J. Nutr. 119,1880 ± 1892.
Brenner RR & Peluffo RO (1969): Regulation of unsaturated fatty acidbiosynthesis. Biochim. Biophys. Acta 176, 471 ± 479.
Brockerhoff H, Ackman RG & Hoyle RJ (1963): Speci®c distribution offatty acids in marine lipids. Arch. Biochem. Biophys. 100, 93 ± 100.
Burr GO & Burr MM (1929): A new de®ciency disease produced by rigidexclusion of fat from the diet. J. Biol. Chem. 82, 345 ± 367.
Bush RA, Reme CE & Malnoe A (1991): Light damage in the rat retina: theeffect of dietary deprivation of n-3 fatty acids on acute structuralalterations. Exp. Eye Res. 53, 741 ± 752.
Caldwell MD, Johnson HT & Othersen HB (1972): Essential fatty acidde®ciency in an infant receiving prolonged parenteral alimentation. J.Pediatr. 81, 894 ± 898.
Carlson SE, Carver JD & House SG (1986): High fat diets varying in ratiosof polyunsaturated to saturated fatty acid and linoleic to linolenic acid: acomparison of rat neural and red cell membrane phospholipids. J. Nutr.116, 718 ± 726.
Carlson SE, Cooke RS & Rhodes PG (1991): Effect of vegetable andmarine oils in preterm infant formulas on blood arachidonic anddocosahexaenoic acids. J. Pediatr. 120, S159 ± S167.
Carlson SE, Ford AJ, Werkman SH, Peeples JM & Koo WW (1996):Visual acuity and fatty acid status of term infants fed human milk andformulas with or without docosahexaenoate and arachidonate from eggyolk lecithin. Pediatr. Res. 39, 882 ± 888.
Carlson SE, Werkman SH, Peeples JM, Cooke RJ & Tolley EA (1993):Arachidonic acid status correlates with ®rst year growth in preterminfants. Proc. Natl. Acad. Sci. USA 90, 1073 ± 1077.
Carnielli V, Wattimena DJL, Luijendijk IHT, Boerlage A, Degenhart HJ &Sauer PJJ (1996): The very low birth weight premature infant is capableof synthesizing arachidonic and docosahexaenoic acids from linoleic andlinolenic acids. Pediatr. Res. 40, 169 ± 174.
Caster WO & Ahn P (1963): Electrocardiographic notching in rats de®cientin EFA. Science 139, 1213.
Charnock JS (1991): Antiarrhythmic effects of ®sh oils. In Health Effects ofw-3 Polyunsaturated Fatty Acids in Seafoods, eds. AP Simopoulos, RRKifer, RE MartõÂn & SM Barlow. World Rev. Nutr. Diet. 66,278 ± 291.
Clandinin MT, Chappell JE, Leong S, Heim T, Swyer PR & Chance GW(1980): Intrauterine fatty acid accretion rates in human brain: implica-tion for fatty acid requirements. Early Hum. Dev. 4, 121 ± 130.
Clarke JTR, Cullen-Dean G, Regelink E, Chan L & Rose V(1990): Increased incidence of epistaxis in adolescents withfamilial hypercholesterolemia treated with ®sh oil. J. Pediatr. 116,139 ± 141.
Clarke SD & Jump DB (1993): Fatty acid regulation of gene expression: aunique role for polyunsaturated fats. In Nutrition and Gene Expression,eds. C Berdanier & JL Hargrove. pp 227 ± 246, Boca Raton, Florida:CRC Press.
Essential fatty acidsR Uauy et al
S75

Connor WE & Connor SL (1990): Diet, atherosclerosis, and ®sh oil. Adv.Intern. Med. 35, 139 ± 171.
Connor WE, Lin DS & Neuringer M (1993): Is the docosahexaenoic acid(DHA, 22:6n-3) content of erythrocytes a marker for the DHA contentof brain phospholipids? FASEB J. 7, A152.
Daviglus M, Stamler J, Orencia AJ, Dyer AR, Kiang L, Greenland PD,Walsh MK, Morris DA & Shekelle RB (1997): Fish consumption andthe 30-year risk of fatal myocardial infarction. N. Engl. J. Med. 336,1046 ± 1053.
De Lorgeril M & Renaud S (1994): Mediterranean alpha-linolenic acid-rich diet in secondary prevention of coronary heart disease. Lancet 343,1454 ± 1459.
Dobbing J (1997): Developing Brain and Behaviour: The Role of Lipids inInfant Formula. London: Academic Press.
Dutta-Roy AK (1997): Fatty acid transport and metabolism in the feto-placental unit and the role of fatty acid-binding protein. J. Nutr.Biochem. 8, 548 ± 557.
Dyerberg J & Bang HO (1979): Haemostatic function and plateletpolyunsaturated fatty acids in Eskimos. Lancet 2, 433 ± 435.
Dyerberg J, Bang HO & Stofferson E (1978): Eicosapentaenoic acid andprevention of thrombosis and atherosclerosis. Lancet 2, 117 ± 119.
El Boustani S, Causse J, Descomps B, Monnier L, Mendy F & Crastes dePaulet A (1989): Direct in vivo characterization of delta 5 desaturaseactivity in human by deuterium labeling: effect of insulin. Metabolism38, 315 ± 321.
Emken EA, Adlof RO & Gulley RM (1994): Dietary linoleic acidin¯uences desaturation and acylation of deuterium-labeled linoleicand linolenic acids in young adult males. Biochem. Biophys. Acta1213, 277 ± 288.
Emken EA, Adlof RO, Duval SM & Nelson GJ (1997): In¯uence ofdietary arachidonic acid on metabolism in vivo of 8 cis, 11 cis,14-eicosatrienoic acid in humans. Lipids 32, 441 ± 448.
Emken EA, Adlof RO, Rakoff H, Rohwedder WK & Gulley M (1992):Human metabolic studies with deuterated alpha linolenic acid. Nutrition8, 213 ± 214.
Eritsland J, Arnesen H, Gronseth K, Fjeld NB & Abdelnoor M (1996):Effect of dietary supplementation with n-3 fatty acids on coronaryartery bypass graft patency. Am. J. Cardiol. 77, 31 ± 36.
Evers AS, Elliott WJ, Lefkowith JB & Needleman P (1986): Manipulationof rat brain fatty acid composition alters volatile anesthetic potency. J.Clin. Invest. 77, 1028 ± 1033.
Fagioli I, Baroncini P, Ricour C & Salzarulo P (1989): Decrease of slow-wave sleep in children with prolonged absence of essential fatty acidsintake. Sleep 12, 495 ± 499.
FAO=WHO Report of a Joint Expert Consultation (1994): Fats and Oils inHuman Nutrition. FAO Food and Nutrition Paper, # 57, pp 49 ± 55.Rome: Food and Agricultural Organization.
Farquharson J, Cockburn F & Ainslie PW (1992): Infant cerebralcortexphospholipidfatty-acidcompositionanddiet.Lancet340, 810 ± 813.
FDA=HHS USA (1991): Food Labeling: Health Claims and Label State-ments: Omega-3 Fatty Acids and Coronary Heart Disease. ProposedRule. Federal Register; 56 (229), 60663 ± 60689.
Fox PL & Dicorleto PE (1988): Fish oils inhibit endothelial cell productionof a platelet-derived growth factor-like protein. Science 241, 453 ± 456.
Galli C, Simopoulos AP & Tremoli E, eds (1994): Effects of Fatty Acidsand Lipids in Health and Disease. Proceedings of the 1st InternationalCongress of the International Society for the Study of Fatty Acids andLipids. World Rev. Nutr. Diet. 76, 1 ± 149.
Galli C & Socini A (1983): Dietary lipids in pre- and post-natal develop-ment. In Dietary Fats and Health, eds. EG Perkins & WJ Visek.Proceeding of American Oil Chemist Society Conference, Chicago,IL, pp 278 ± 301.
Galli C, Trzeciak HI & Paoletti R (1971): Effects of dietary fatty acids onthe fatty acid composition of brain athanolamine phosphoglyceride:reciprocal replacement of n-6 and n-3 polyunsaturated fatty acids.Biochim. Biophys. Acta 248, 449 ± 454.
Garrido A, Garate M, Campos R, Villa A, Nieto S & Valenzuela A (1993):Increased susceptibility of cellular membranes to the induction ofoxidative stress is observed after ingestion of high doses of ®sh oil:effect of aging and protective action of dl-a tocopherol supplementa-tion. Nutr. Biochem. 4, 118 ± 122.
Garrido A, Garrido F, Guerra R & Valenzuela A (1989): Ingestion of highdoses of ®sh oil increases the susceptibility of cellular membranes to theinduction of oxidative stress. Lipids 24, 833 ± 835.
Glomset JA (1985): Fish, fatty acids and human health. N. Engl. J. Med.312, 1253 ± 1254.
Hallaq H,Smith TW & Leal A (1992): Modulation of dihydropyridine-sensitive calcium channels in hearts cells by ®sh oil fatty acids. Proc.Natl. Acad. Sci. USA 89, 1760 ± 1764.
Hansen J (1993): Appendix: Commercial formulas for preterm infants1992. In Nutritional Needs of Preterm Infants: Scienti®c Basis andPractical Guidelines, eds. RC Tsang, A Lucas, R Uauy & S Zlotkin. pp297 ± 301, Pawling New York: Williams and Wilkins by CaduceusMedical Publishers Inc.
Harris WS (1989): Fish oils and plasma lipid and lipoprotein metabolismin humans: A critical review. J. Lipid Res. 30, 785 ± 807.
Harris WS, Connor WE, Alam D & Illingworth DR (1988): Reduction ofpostprandial triglyceridemia in humans by dietary N-3 fatty acids.J. Lipid Res. 29, 1451 ± 1460.
Hoffman D & Uauy R (1992): Essentiality of dietary omega-3 fatty acidsfor premature infants: plasma and red blood cell fatty acid composition.Lipids 27, 886 ± 895.
Holh CM & Rosen P (1987): The role of arachidonic acid in rat heart cellmetabolism. Biochem. Biophys. Acta 921, 356 ± 363.
Indu M & Ghafoorunisa MI (1992): N-3 fatty acids in Indian diets.Comparison of the effect of precursosr (LNA) vs. Product (LCPUFAn-3). Nutr. Res. 12, 569 ± 582.
Innis SM (1991): Essential fatty acids in growth and development. Prog.Lipid. Res. 30, 39 ± 103.
Innis SM, Akrabawi SS, Diersen Schade DA, Dobson MV & Guy DG(1997): Visual acuity and blood lipids in term infants fed human milk orformulae. Lipids 32, 63 ± 72.
Innis SM, Nelson CM, Lwanga D, Rioux FM & Waslen P (1996): Feedingformula without arachidonic acid and docosahexaenoic acid has noeffect on preferential looking acuity or recognition in healthy full terminfants at 9 mo of age. Am. J. Clin. Nutr. 64, 40 ± 46.
Innis SM, Nelson CM, Rioux MF & King J (1994): Development of visualacuity in relation to plasma and erythrocyte n-6 and n-3 fatty acids inhealthy term gestation infants. Am. J. Clin. Nutr. 60, 347 ± 352.
Jensen CL, Prager TC, Fraley JK, Chen H, Anderson R & Heird WC(1997): Effect of dietary linoleic = alpha-linolenic acid ratio on growthand visual function of term infants. J. Pediatr. 131, 200 ± 209.
Jorgensen MH, Hernell O, Lund P, Holmer G & Michaelsen KF (1996):Visual acuity and erythrocyte docosahexaenoic acid status in breast-fedand formula-fed term infants during the ®rst 4 months of life. Lipids 31,99 ± 105.
Kang JX & Leaf A (1995): Prevention and termination of the beta-adrenergic agonist-induced arrhythmias by free polyunsaturated fattyacids in neonatal rat cardiac myocytes. Biochem. Biophys. Res.Commun. 208, 629 ± 636.
Kang JX, Xiao YF & Leaf A (1995): Free, long-chain polyunsaturatedfatty acids reduce membrane electrical excitability in neonatal ratcardiac myocytes. Proc. Natl. Acad. Sci. USA 92, 3997 ± 4001.
Katan MB (1995): Fish and heart disease. NEJM 332, 1024 ± 1025.Knapp HR & FitzGerald GA (1989): The antihypertensive effects of ®sh
oil. A controlled study of polyunsaturated fatty acid supplements inessential hypertension. N. Engl. J. Med. 320, 1037 ± 1043.
Koletzko B, Schmidt E & Bremer HJ (1989): Effects of dietary long-chainpolyunsaturated fatty acids on the essential fatty acid status of pre-mature infants. Eur. J. Pediatr. 148, 669 ± 675.
Koletzko B & Bremer HJ (1989): Fat content and fatty acid composition ofinfant formulas. Acta Paediatr. Scand. 78, 513 ± 521.
Leaf A, Jorgensen MB, Jacobs AK, Cote G, Schoenfeld DA, Scheer J,Weiner BH, Slack JD, Kellett MA, Raizner AE et al (1994): Do ®shoils prevent restenosis after coronary angioplasty? Circulation 90,2248 ± 2257.
Leaf A & Weber PC (1988): Cardiovascular effects of n-3 fatty acids.N. Engl. J. Med. 318, 549 ± 557.
Lee AG, East JM & Froud RJ (1986): Are essential fatty acids essential formembrane function? Prog. Lipid Res. 25, 41 ± 46.
Levinson PD, Iosiphidis AH, Saritelli AL, Herbert PN & Steiner M (1990):Effects of n-3 fatty acids in essential hypertension. Am. J. Hypertens. 3,754 ± 760.
Lin DS, Connor WE, Anderson GJ & Neuringer M (1990): Effects ofdietary n-3 fatty acids on the phospholipid molecular species of monkeybrain. J. Neurochem. 55, 1200 ± 1207.
Litman BJ & Mitchell DC (1996): A role for phospholipids polyun-saturation in modulating membrane protein function. Lipids 31, S13 ±S197.
Love JA, Saurn WR & McGee R (1985): The effects of exposure toexogenous fatty acids and membrane fatty acid modi®cation on theelectrical properties of NG108-15 cells. Cell. Molec. Neurobiol. 5,333 ± 352.
Lox CD (1990): The effects of dietary marine ®sh oils (Omega-3 fattyacids) on coagulation pro®les in men. Gen. Pharmacol. 21, 241 ± 246.
Makrides M, Neumann M, Simmer K, Pater J & Gibson R (1995): Arelong-chain polyunsaturated fatty acids essential nutrients in infancy?Lancet 345, 1463 ± 1468.
Essential fatty acidsR Uauy et al
S76

Makrides M, Simmer K, Goggin M & Gibson RA (1993): Erythrocytedocosahexaenoic acid correlates with the visual response of healthy,term infants. Pediatr. Res. 34, 425 ± 427.
Makrides M, Neumann MA, Byard RW & Gibson RA (1994): Fatty acidcomposition of brain retina and erythrocytes in breast and formula fedinfants. Am. J. Clin. Nutr. 60, 189 ± 194.
Martinez M (1992): Tissue levels of polyunsaturated fatty acids duringearly human development. J. Pediatr. 120, S129 ± S138.
Martinez M (1996): Docosahexaenoic acid therapy in docosahexaenoicacid-de®cient patients with disorders of peroxisomal biogenesis. Lipids31, Suppl, S145 ± S152.
Masataka O, Kajiwara K, Imai S, Kobayashi T, Honma N, Maki T, SurugaK, Goda T, Takase S, Muto Y & Moriwaki H (1997): Perilla oilprevents the excessive growth of visceral adipose tissue in rats by downregulating adipocyte differentiation. J. Nutr. 127, 1752 ± 1757.
Massarei K, Stanton G & Dunbar D (1989): Dietary ®sh oils increaseserum lipids in insulin-dependent diabetics compared with healthycontrols. Metabolism 38, 404 ± 409.
Menon NK & Dhopeshwarkar GA (1982): Essential fatty acid de®ciencyand brain development. Prog. Lipid Res. 21, 309 ± 326.
Morris MC, Sacks F & Rosner B (1993): Does ®sh oil lower bloodpressure? A meta-analysis of controlled trials. Circulation 88,523 ± 533.
Nestel PJ, Connor WE, Reardon MR, Connor S, Wong S &Boston R (1984): Suppression by diets rich in ®sh oil of verylow density lipoprotein production in man. J. Clin. Invest. 74,72 ± 89.
Neuringer M, Connor WE, Lin DS, Anderson GJ & Barstad L (1991):Dietary o-3 fatty acids: Effect on retinal lipid composition and functionin primates. In Retinal Degenerations, eds. RE Anderson, JG Holly®eld& MM LaVail. pp 1 ± 13, New York: CRC Press.
Neuringer M, Connor WE, Van Petten C & Barstad L (1984): Dietaryomega-3 fatty acid de®ciency and visual loss in infant rhesus monkeys.J. Clin. Invest. 73, 272 ± 276.
Okuyama H, Kobasyashi & Watanabe S (1997): Dietary fatty acids. The n-6=n-3 balance and chronic elderly diseases. Excess linoleic acid andrelative n-3 de®ciency syndrome seen in Japan. Prog. Lipid Res. 35,409 ± 457.
Ongari MA, Ritter JM, Orchard MA, Waddell KA, Blair IA & Lewis PJ(1984): Correlation of prostacyclin synthesis by human umbilical arterywith status of essential fatty acid. Am. J. Obstet. Gynecol. 149, 455 ± 460.
Oski FA & Barness LA (1967): Vitamin E de®ciency: a previouslyunrecognized cause of hemolytic anemia in the premature infant.J. Pediatr. 70, 211 ± 220.
Ponder DL, Innis SM, Benson JD, & Siegman JS (1992): Docosahexaenoicacid status of term infants fed breast milk, or soy oil or corn oilcontaining infants formula. Pediatr. Res. 32, 638 ± 688.
Ranaian M, Szilasi J, Storlien L & Calvert GD (1996): The effect ofmodi®ed fat diet on insulin resistance and metabolic parameters in typeII diabetes. Diabetologia 39, A7.
Salem N Jr, Shingu T, Kim HY, Hullin F, Bougnoux P & Karanian JW(1988): Specialization in membrane structure and metabolism withrespect to polyunsaturated lipids. In Biological Membranes: Aberra-tions in Membrane Structure and Function, eds. ML Karnovsky, A Leaf& LC Bollis. pp 319 ± 333, New York: Alan R. Liss, Inc.
Salem N, Wegher B, Mena P & Uauy R (1996): Arachidonic anddocosahexaenoic acids are biosynthesized from their 18-carbon pre-cursors in human infants. Proc. Natl. Acad. Sci. USA 93, 49 ± 54.
Sauerwald TU, Hachey DL, Jensen CL, Chen H, Anderson RE & HeirdWC (1996): Effect of dietary alpha-linolenic acid intake on incorpora-tion of docosahexaenoic and arachidonic acids into plasma phospho-lipids of term infants. Lipids 31, Suppl, S131 ± S135.
Schoonjans K, Staels B & Auwerx J (1996): Role of the peroxi-some proliferator-activated receptor (PPAR) in mediating theeffects of ®brates and fatty acids on gene expression. J. Lipid Res.37, 907 ± 925.
Sellmayer A, Witzgall H, Lorenz RL & Weber PC (1995): Effects ofdietary ®sh oil on ventricular premature complexes. Am. J. Cardiol. 76,974 ± 977.
Shimokawa H & Vanhoutte PM (1988): Dietary omega-3 fatty acids andendothelium-dependent relaxations in porcine coronary arteries. Am. J.Physiol. 256, H968 ± H973.
Simopoulos AP (1991): Omega-3 fatty acids in health and disease and ingrowth and development. Am. J. Clin. Nutr. 54, 438 ± 463.
Simopoulos AP (1994): Fatty acid composition of skeletal muscle mem-brane phospholipids, insulin resistance and obesity. Nutr Today 2,12 ± 16.
Sosenko IR, Innis SM & Frank L (1989): Menhaden ®sh oil, n-3polyunsaturated fatty acids, and protection of newborn rats fromoxygen toxicity. Pediatr. Res. 25, 399 ± 404.
Sprecher H (1981): Biochemistry of essential fatty acids. Prog. Lipid Res.20, 13 ± 22.
Storlien LH, Pan DA, Kriketos AD, O'Connor J, Caterson ID, Cooney GJ,Jenkins AB & Baur LA (1996): Skeletal muscle membrane lipids andinsulin resistance. Lipids 31, S261 ± S265.
Stother JK & Warner RM (1977): Oxygen consumption and sleep state inthe newborn. J. Physiol. 269, 57 ± 58.
Stubbs CD & Smith AD (1984): The modi®cation of mammalianmembrane polyunsaturated fatty acid composition in relation tomembrane ¯uidity and function. Biochim. Biophys. Acta 779,89 ± 137.
Treen M, Uauy RD, Jameson D, Thomas V & Hoffman DR (1992): Effectof docosahexaenoic acid on membrane ¯uidity and function inintact cultured Y-79 retinoblastoma cells. Arch. Biochem. Biophys.294, 564 ± 570.
Uauy R & Hoffman DR (1991): Essential fatty acid requirementsfor normal eye and brain development. Semin. Perinatol. 15,449 ± 455.
Uauy R, Hoffman DR, Birch EE, Birch DG, Jameson DM & Tyson J(1994): Safety and ef®cacy of omega-3 fatty acids in the nutrition ofvery low birth weight infants: soy oil and marine oil supplementation offormula. J. Pediatr. 124, 612 ± 620.
Uauy R, Treen M & Hoffman D (1989): Essential fatty acidmetabolism and requirements during development. Sem. Perinatol.13, 118 ± 130.
Uauy RD, Birch DG, Birch EE, Tyson JE & Hoffman DR (1990): Effect ofdietary omega-3 fatty acids on retinal function of very low birth weightneonates. Pediatr. Res. 28, 484 ± 492.
Voss A, Reinhart M, Shankarappa S & Sprecher H (1991): The metabo-lism of 7,10,13,16,19-docosapentaenoic acid to 4,7,10,13,16,19-doco-sahexaenoic acid in the rat liver is independent of a 4-desaturase.J. Biol. Chem. 266, 19995 ± 20000.
Watanabe I, Kato M & Aonuma H (1987): Effect of dietary alpha-linolenate=linoleate balance on the lipid composition and electroretino-graphic responses in rats. Adv. Biosci. 62, 563 ± 568.
Werkman SH & Carlson SE (1996): A randomized trial of visual attentionof preterm infants fed docosahexaenoic acid until nine months. Lipids31, 91 ± 97.
Wheeler TG, Benolken RM & Anderson RE (1975): Visual membranes:speci®city of fatty acid precursors for the electrical response toillumination. Science 188, 1312 ± 1314.
Widdowson EM & Dickerson JW (1981): Composition of the body. InGeigy Scienti®c Tables, ed. C Letner. West Caldwell, NJ: Ciba-Geigy,pp 217 ± 225.
Willis AL (1984): Essential fatty acids, prostaglandins, and relatedeicosanoids. In Present Knowledge in Nutrition, ed. RE Olson. pp90 ± 113. Washington, D.C.: The Nutrition Foundation Inc.
Wo CKW & Draper HH (1975): Vitamin E status of Alaskan Eskimos.Am. J. Clin. Nutr. 28, 808 ± 812.
Yorek MA, Bohnker RR, Dudley DT & Spector AA (1984): Comparativeutilization of N-3 polyunsaturated fatty acids by cultured human Y-79retinoblastoma cells. Biochim. Biophys. Acta 795, 277 ± 285.
Essential fatty acidsR Uauy et al
S77