essay ethics concept:
TRANSCRIPT

Report on the Risk Assessment of Bt Maize
Chapter 1 Introduction
Klaus Ammann, Delft, Istanbul and Neuchâtel, 16. 6. 2010

2
Fig. 1 Grenade (Haute-Garonne, France). left: Bt maize (MON810) variety PR33P67. right : non Bt variety PR33A46, with fungal infection, source Karine Affaton, EuropaBio, fall 2007, Fotos K.Ammann

3
1. Introduction: Bacillus thuringiensis and the make of Bt maize
CHAPTER 1 INTRODUCTION ............................................................................................................................. 1
Klaus Ammann, Delft and Istanbul, 16. 4. 2009 .............................................................................................. 1
1. INTRODUCTION: BACILLUS THURINGIENSIS AND THE MAKE OF BT MAIZE .............................. 3
1.1. BACILLUS THURINGIENSIS AND ITS USE, AN INTRODUCTION ...................................................................................... 8 1.1.1. Taxonomy, genetics and isolation of Bacillus thuringiensis ............................................................... 8
1.1.1.1. Comparison of Bacillus thuringiensis with its close relatives ..................................................................... 8 1.1.1.2. Discovery, natural occurrence and isolation of Bacillus thuringiensis ................................................ 10 1.1.1.3 Unfounded concerns about accumulation of Bt protoxins from GM crops in Water and soil ..... 13
1.1.2.Bacillus thuringiensis external applications.......................................................................................... 14
1.1.2.1. External Bt applications with a long tradition, but remaining marginal in use. ................................. 14 1.1.2.2. Bt sprays cause resistant insects, but not (yet) Bt crops ............................................................................ 15
1.1.3.Transgenic Bt maize ................................................................................................................................ 19
1.1.3.1. Conventional breeding of insect resistant maize not very successful .................................................. 19 1.1.3.2. How it all started, the (re)view of two initiators ................................................................................................ 19 1.1.3.3. First cloning of a Bt gene .......................................................................................................................................... 20 1.1.3.4. Genetically engineered insect resistant Bt maize ........................................................................................... 20
1.1.4. Chemistry, Biology and nomenclature of Bt toxins ............................................................................ 25
1.1.5. Mode of action of Bt proteins and selectivity of the toxins ................................................................ 34
1.1.6. Bt crops: present and future developments, synthetic Bt genes .................................................... 39
1.1.6.1. Present situation in Europe ....................................................................................................................................... 39 1.1.6.2. Recent and future developments of Bt maize breeding ................................................................................ 39
1.2. FUNDAMENTAL STATEMENTS ABOUT RISK ASSESSMENT, PARTICULARLY RELATED TO BT MAIZE. ............... 46 1.2.1.The situation in risk and benefit assessment of Bt maize worldwide and in Europe ..................... 46
1.2.2 The agricultural reality: ............................................................................................................................ 46
1.2.3. And what about Europe ? ...................................................................................................................... 47
1.2.3. There are three major reasons, why we should develop a critical view, strictly based on
scientific data, when we judge risk assessment research and the resulting scientific publications
related to Bt crops. ................................................................................................................................... 49
1.2.3.1..Assessment of the impact of GM crops should also take into account the benefits and balance
it against the risks. ................................................................................................................................................................ 49 1.2.3.2.Assessment of the impact of GM crops should also compare to non GM crops .............................. 50 1.2.3.3.Dealing with complex structures of parameters in risk assessment calls for scientific scrutiny
done by experts ....................................................................................................................................................................... 52 1.2.4.Example of a complex structure of risk assessment, an analysis related to the scientific
questions alone: The British Farm Scale Experiments ....................................................................... 53

4
1.2.6.Conclusion: Proposed procedure for future risk assessments .......................................................... 54
1.2.7. Cited Literature ........................................................................................................................................ 57
List of Figures
Fig. 1 Grenade (Haute-Garonne, France). left: Bt maize (MON810) variety PR33P67. right : non Bt
variety PR33A46, with fungal infection, source Karine Affaton, EuropaBio, fall 2007, Fotos
K.Ammann ........................................................................................................................................................... 2 Fig. 2 Schematic overview of the entomopathogenic spore-forming bacteria and their protein toxins.
Localization in crystals or outside the cell (secreted) is depicted as far as known from
literature. Mtx proteins contain signal sequences for secretion, but their localization has not
been experimentally determined. Similar colors for toxins identify members of the same
homology group (yellow for 3-domain Cry proteins; red-orange for Mtx2/3-like proteins; green
for Bin-like proteins). Fig.1 from (de Maagd et al., 2003) ...................................................................... 10 Fig. 3 Above: Primary and tertiary structure of Cry toxins. (a) Relative lengths of Cry protoxins and
position of the five conserved blocks, if present. More details on these conserved blocks, as
well as the identification of three more blocks in the C-terminal ends of the longer protoxins,
can be found in . The positions of the three domains of the activated toxin are indicated for
Cry1 and vary with the positions of blocks 2 and 3 for the other toxins. The remainder of the
protoxin, consisting of short N-terminal part (20–40 amino acids) preceding the first domain

5
and the C-terminal part following the third domain in the longer protoxins, is digested away by
gut proteases during the activation process. (b) Three-dimensional structure of an activated
toxin, Cry1Aa (Ref. 7). The toxin has three structural domains. Domain I (blue) is involved in
membrane insertion and pore formation. Domain II (green) and domain III (yellow-red) are
involved in receptor recognition and binding. Conserved block 1 is in the central helix of
domain I, block 2 is at the domain I–II interface, block 3 is at the boundary between domains II
and III, block 4 is in the central β-strand of domain III and block 5 is at the end of domain III.
From (de Maagd et al., 2001) ......................................................................................................................... 25 Fig. 4 Transmission electron micrograph of a sporulating Bacillus thuringiensis (Bt) cell. δ-
Endotoxins are produced as regularly shaped crystals (PB; protein body) – hence the name
crystal (Cry) proteins – next to a spore (SP). The vegetative cell wall will eventually break to
release the spore and crystal. The cell shown is approximately 2 μm long. From (de Maagd et
al., 2001) ............................................................................................................................................................. 26 Fig. 5 Spores and crystals of Bacillus thuringiensis serovar morrisoni strain T08025 Microscopy by
Jim Buckman from http://commons.wikimedia.org/wiki/File:Bacillus_thuringiensis.JPG .......... 26 Fig. 6 Scanning electron microscopy of spore (s) and crystals (c) of strains S285 (1); S447 (2); S479
3); S550 (4); S1255 (5); and Bti (6). From (Monnerat et al., 2005) Most of the Bt proteins toxic to
Mosquitos have a round shape, in contrast to the Bt proteins usually inserted in Bt maize. ..... 27 Fig. 7 Phylogenetic relationships of the separate domains. Unrooted phylogenetic trees of domains
I, II and III of 79 known subgroups of Cry proteins obtained by the parsimony method. Trees
were constructed basically as described earlier31, except that toxin alignments were made
using DbClustal45, and updated with Cry protein sequences that were released since 1997.
Cry6, Cry15, Cry22 and Cry23 sequences were not included because they do not show
similarities with the rest of the Cry protein family, see also Fig. 2(a). Shown are consensus
trees resulting from 100 analyses using the bootstrapping tool and the CONSENSE program.
Branches are color-coded according to the insect order specificity of the toxins, as far as is
known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta,
nematode specific; yellow, Hymenoptera specific. From de Maagd (de Maagd et al., 2001) ...... 29 Fig. 8 Phylogenetic relationships of the separate domains. Unrooted phylogenetic trees of domains
I, II and III of 79 known subgroups of Cry proteins obtained by the parsimony method. Trees
were constructed basically as described earlier31, except that toxin alignments were made
using DbClustal45, and updated with Cry protein sequences that were released since 1997.
Cry6, Cry15, Cry22 and Cry23 sequences were not included because they do not show
similarities with the rest of the Cry protein family, see also Fig. 2(a). Shown are consensus
trees resulting from 100 analyses using the bootstrapping tool and the CONSENSE program.
Branches are color-coded according to the insect order specificity of the toxins, as far as is
known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta,
nematode specific; yellow, Hymenoptera specific. From de Maagd (de Maagd et al., 2001) ...... 30 Fig. 9 Phylogenetic relationships of the separate domains. Unrooted phylogenetic trees of domains
I, II and III of 79 known subgroups of Cry proteins obtained by the parsimony method. Trees
were constructed basically as described earlier31, except that toxin alignments were made
using DbClustal45, and updated with Cry protein sequences that were released since 1997.
Cry6, Cry15, Cry22 and Cry23 sequences were not included because they do not show
similarities with the rest of the Cry protein family see also Fig. 2(a). Shown are consensus
trees resulting from 100 analyses using the bootstrapping tool and the CONSENSE program.
Branches are color-coded according to the insect order specificity of the toxins, as far as is
known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta,
nematode specific; yellow, Hymenoptera specific. From de Maagd (de Maagd et al., 2001) ..... 31 Fig. 10 Phylogenetic relationships between the entomocidal toxins. Four distinct homology groups
have been identified within the family of toxins (Crickmore, 2003), three of which are shown

6
here. The Cyt group is not shown. The branches are color coded according to insect order
specificity of the toxin where known: red, Coleoptera specific; green, Lepidoptera specific;
blue, Diptera specific; magenta, nematode specific; yellow, Hymenoptera specific. Solid and
dashed double-headed arrows indicate associations of proteins in binary toxins or suspected
binary toxins, respectively. Panel A: 3-domain Cry proteins. The C-terminal extension (if
present) was not included for the phylogenetic analysis. Hence, proteins that have the same
primary rank based mainly on sequence similarity between their C-terminal extensions may
sometimes end up on different branches. Panel B: Binary and Bin-like toxins; Panel C: Mtx2,
Mtx3 and Mtx2/3-related proteins. From de Maagd et al. (de Maagd et al., 2003) ........................... 32 Fig. 11 Mode of action of Bacillus thuringiensis crystal proteins. Fig 1 from (de Maagd, 2007) ......... 35 Fig. 12. Primary structure of Cry proteins indicating the variety in length of the protoxin and the
extent of the activated toxin after digestion of the protoxin by gut proteases, as well as the
position of the three structural domains. Bar indicates number of amino acids. B. Tertiary
structure of Cry1Aa toxin. Clearly recognizable are the three structural domains (Roman
numerals). Fig 2 from (de Maagd, 2007) .................................................................................................... 37 Fig. 13 Mode of action of Cry toxins. (a) After ingestion by the insect, crystals dissolve in the gut
juice. (b) Gut proteases subsequently clip off the C-terminal extension in the longer Cry
proteins (purple) as well as a small N-terminal fragment (yellow). (c) The resulting ‘activated’
toxin (i.e. the structure depicted in Fig. 2b) binds to receptors on the epithelial cell membrane,
a process in which both domain II and domain III are involved. (d) Structural rearrangement of
domain I might follow allowing a two-helix hairpin to insert into the membrane. (e) Inserted
toxins form pores probably as oligomers, but the architecture of the pore is still unknown.
From (de Maagd et al., 2001) ......................................................................................................................... 37 Fig. 14 Mechanism of Cry protein toxicity. A: Ingestion of spores or recombinant protein by
phytophagous larva. B: In the midgut, endotoxins are solubilized from Bt spores (s) and
inclusions of crystallized protein. (cp). C: Cry toxins are proteolytically processed to active
toxins in the midgut. Active toxin binds receptors on the surface of columnar epithelial cells.
Bound toxin inserts into the cellular membrane. D: Cry toxins aggregate to form pores in the
membrane. E: Pore formation leads to osmotic lysis. F: Heavy damage to midgut membranes
leads to starvation or septicemia. From (Whalon & Wingerd, 2003) ................................................. 38 Fig. 15 All the strains and plasmids used or constructed (Xue et al., 2008)are listed in Table fig. 15.
Escherichia coli JM110 was used for plasmid propagation, and SCS110 was used to produce
nonmethylated plasmid DNA for the transformation of B. thuringiensis. Escherichia coli
strains were grown in Luria–Bertani medium (LB) at 37 1C, while B. thuringiensis strains were
grown in peptone-beef extract (PB) medium (0.5% peptone, 0.3% beef extract) at 30 1C. Liquid
cultures were grown in a rotary shaker at 230 r.p.m. Antibiotics were added to autoclaved
media as follows: ampicillin, 100 mgmL-1
(for E. coli); erythromycin 10 mgmL-1
(for B.
thuringiensis), from (Xue et al., 2008) ........................................................................................................ 41 Fig. 16 Engineering specificity in a three-domain Cry toxin; mutagenesis of the toxin-receptor
interaction loop in domain II. Threedimensional structure of Cry3A (1dlc; RCSB) is shown in
ribbon format. Domain I (helices) is at top right, and domain II (sheet structure) is at bottom
left. Domain III (carbohydrate-binding domain; sheet structure) is behind the other domains,
central in this view. Residues mutated (Wu et al., 2000) to increase toxicity toward yellow
mealworm (Tenebrio molitor), Colorado potato beetle, and cottonwood leaf beetle (Chrysomela
scripta) are shown in ball-and stick representation. From (Gatehouse, 2008) ................................ 44 Fig. 17 Engineering specificity in a three-domain Cry toxin; mutagenesis to improve channel-
forming ability. Three-dimensional structure of Cry3Bb (1ji6; RCSB) is shown in ribbon format
in the same view as Figure 1. Residues mutated (English et al., 2003) to increase toxicity
toward corn rootworm are shown in ball-and-stick representation. Mutations are made in
helices of domain I and in the region linking domains I and II. The mutation sites shown are

7
taken from the most active toxin produced; a range of other sites for mutation were explored.
From (Gatehouse, 2008) ................................................................................................................................. 45 Fig. 18, Assessment continuum within a tiered scheme of ecological risk assessment. The decision
to reject the risk hypothesis includes consideration of residual uncertainties. With increasing
tiers, the assessment becomes more complex and realistic, with conclusions that are more
specific. The assessment can stop at any stage during the process as soon as sufficient
information has been compiled to address the risk hypothesis. Thus collection of data
irrelevant to the risk assessment is minimized. N, level of risk assessment tier; NTA, nontarget
arthropod. From (Romeis et al., 2008a) ..................................................................................................... 55 Fig. 19, Reconstruction of NTA risk assessment for Bt maize expressing Cry1Ab showing that
different risk hypotheses require different types of data and synthesis at different tiers. From
(US Environmental Protection Agency (USEPA), 1995) taken from (Romeis et al., 2008a) and
see also the fully cited accompanying text below: ................................................................................. 56

8
1.1. Bacillus thuringiensis and its use, an introduction
1.1.1. Taxonomy, genetics and isolation of Bacillus thuringiensis
Bacillus thuringiensis (Bt) is a bacterium of great agronomic and scientific interest.
Together, the many subspecies of this bacterium colonize and kill a large variety of
host insects and even nematodes, but each strain does so with a high degree of
specificity (de Maagd, 2007; de Maagd et al., 2003; de Maagd et al., 2001), which makes sense in the
complex web of the ecosystems.
The taxonomy of Bacillus thuringiensis has undergone revisions (Helgason et al., 2000; Lecadet et al.,
1999), which is important for any kind of communication related to risk assessment of Bt pesticide
strategies.
See also a summary of the wider Taxonomy in a recent review by (Swiecicka, 2008):
Bacillus cereus and Bacillus thuringiensis are treated there as members of the Bacillus cereus group, which
includes four additional species, Bacilius anthracis, Bacillus weihenstephanensis, Bacillus mycoides and
Bacillus pseudomycoides, they comprise a clearly distinguishable cluster among facultative anaerobic
endospore-forming bacteria of the genus Bacillus, following (Priest, 1993).
A major comparative genomic study has been published by (Han et al., 2006b). The complete genome
sequence of Bacillus thuringiensis has been uncovered subsequently by the same research group
(Challacombe et al., 2007).
In another recent review of the genetics and taxonomy of B. thuringiensis (Vilas-Boas et al., 2007) show
the full complexity of the matter: Since the first published genome sequence of the B. anthracis A2012
strain (Read et al., 2002) several complete-genome sequencings have been undertaken: 10 strains of B.
anthracis, 8 strains of B. cereus and 3 strains of B. thuringiensis are now available in GenBank, actually
making it the group of closely related bacteria with the highest number of fully sequenced genomes. This
offers an unprecedented opportunity for extensive comparative genomic studies (Vilas-Boas et al., 2007).
In essence it is concluded there that classic genomic comparison does not allow for direct conclusions
about horizontal gene flow, it will be necessary to include an ecological species concept (Weisse, 2007).
Analytical chemistry allows to identify and quantify the toxic proteins characterizing the various varieties
of Bacillus thuringiensis (Bt) (Hickle & Fitch, 1998). The large majority of known Bt toxins are classified as
3-domain toxins, according to their homology to a small number of toxins of which the 3-dimensional
structure has been experimentally determined.
1.1.1.1. Comparison of Bacillus thuringiensis with its close relatives
Although it has been suggested that Bacillus thuringiensis, B. cereus and B. anthrax are all members of the
same species (Helgason et al., 2000), and although there is ample evidence that B. thuringiensis and B.
cereus could be taken as members of the same species, this is certainly not the case for B. anthracis, since

9
the two plasmids that encode the toxins of B. anthracis do not occur naturally in the other two species,
and they do not have parasporal bodies containing Bt Cry proteins. This implies that there are probably
natural barriers not yet properly understood (Federici & Siegel, 2007).
It must also be stated clearly, that insecticidal crystals formed by Cry and Cyt proteins are the principal
characteristic that differentiates B. thuringiensis from B. cereus, as well as from other species of the B.
cereus group. As far as is known, most if not all Cry and Cyt proteins are encoded on plasmids present in
Bacillus thuringiensis, i.e. not on the bacterial chromosome (Crickmore et al., 1998). Just the other way
around this makes it also clear that any “Bt” lacking the Cry or Cyt plasmids, be it by natural loss or taken
out deliberately - the resulting strain would be identified as B. cereus (Baumann et al., 1984; Hill et al.,
2004; Rasko et al., 2005; Rasko et al., 2007). Despite this, B. thuringiensis is still considered a valid species
due to a combination of tradition and practical value, and this is, according to Federici et al. 2007 (Federici
& Siegel, 2007), unlikely to change, at least in the near future.
There has been a lot of scaremongering in the anti-biotech community about the close relationship of
Bacillus thuringiensis with B. anthracis, here just one example: (Mae, 2001). Such superficial comparisons
on a strict molecular level led to scientifically unfounded safety concerns about the close relationship
between Bacillus thuringiensis and the highly virulent B. anthracis (Heinemann & Traavik, 2004b), which
later had to be rectified (Heinemann & Traavik, 2004a) after having been contradicted (de Maagd et al.,
2005). However, horizontal gene flow seems to be possible between B. cereus and B. thuringiensis under
specific soil conditions: Experimental results (Vilas-Boas et al., 2002) suggest that the rate of gene flow is
higher between strains of the same species; but that exchanges between B. cereus and B. thuringiensis
are nonetheless possible. Overall, these data indicate that it is not important for risk assessment purposes
to determine whether B. cereus and B. thuringiensis belong to a single or to two species. Assessment of
the biosafety of pest control based on B. thuringiensis-toxins requires evaluation of the extent of genetic
exchange between strains in realistic natural conditions. This view has been recently confirmed and
extended (Vilas-Boas et al., 2007): A genomic comparison between B. cereus, B. anthracis and B.
thuringiensis should not barely include sequence data which obviously can lead to wrong superficial
conclusions, but it should follow a much more adequate ecological species concept (Godreuil et al., 2005;
Vanvalen, 1976; Vellai et al., 1999; Weisse, 2007).
A major comparative genomic study has been published by (Han et al., 2006a; Han et al., 2006b). (Hickle
& Fitch, 1998) present the analytical chemistry involved in identifying and quantifying the toxin proteins
present in various varieties of Bacillus thuringiensis.
An instructive figure on related species has been published by (de Maagd et al., 2003):

10
Fig. 2 Schematic overview of the entomopathogenic spore-forming bacteria and their protein toxins. Localization in crystals or outside the cell (secreted) is depicted as far as known from literature. Mtx proteins contain signal sequences for secretion, but their localization has not been experimentally determined. Similar colors for toxins identify members of the same homology group (yellow for 3-domain Cry proteins; red-orange for Mtx2/3-like proteins; green for Bin-like proteins). Fig.1 from (de Maagd et al., 2003)
1.1.1.2. Discovery, natural occurrence and isolation of Bacillus thuringiensis
Bacillus thuringiensis (Bt) was isolated from a flour moth collected in the German province of Thuringia
and described by (Berliner, 1915). Ernst Berliner isolated the type species of Bacillus thuringiensis from
the Mediterranean flour moth Ephestia kuehniella.
The same organism had already been described by (Ishiwata, 1901-1902) as Bacillus sotto from Japan
where it causes a wilt disease of silkworm caterpillars, but the description was not known to Berliner.
Bacillus thuringiensis is now the accepted name for a range of aerobic spore-forming bacteria which form
an insecticidal crystal during sporulation.
A still valid and classic account on bacterial insecticides has been published by (Heimpel & Angus, 1960).
Their account makes it clear, that there was ample knowledge on insect pathology of an unknown
bacterium with a very high toxicity, and the authors defined a process with ‘endotoxin’ produced by the
bacterium which is poisonous to many Lepidoptera. The endotoxins of this bacterium, later discovered by
Berliner as the Bt, killed the insects and then it reproduced by spores within the dead insect. This set of

11
processes and components evolved in grain-feeding and other pyralid insects, specifically in larvae of
species such as the southern European sunflower moth Homoeosoma nebulella, from which the type
species of Bacillus thuringiensis ssp thuringiensis was discovered by Berliner. So, after all, the toxicity
processes and life cycles of Bt are quite natural and existing in the insect world in abundance.
Since the pioneering work of Steinhaus in California in the early 1950s (Steinhaus, 1951), there has been
considerable commercial interest and products are now sold for external use in most countries of the
world for control of caterpillars (var. kurstaki, entomocidus, galleriae and aizawai), mosquito and blackfly
larvae (var. israelensis) and beetle larvae (var. tenebrionis and var. sandiego) (Milner, 1994).
Bacillus thuringiensis occurs in nature in many different substrates and environments.
(Damgaard et al., 1997) isolated Bacillus thuringiensis from the phylloplane of cabbage foliage. The same
authors have previously shown (Pedersen et al., 1995) that spores of B. thuringiensis serovar kurstaki can
readily be dispersed from soil to the lower leaves of cabbage plants. Therefore they could expect that the
population studied now would not differ from that normally found in soil. Natural occurrence of B.
thuringiensis has also been confirmed by (Kaur & Singh, 2000) in leguminous phylloplanes in the New
Delhi region of India.
The relatively high proportion of isolates from the phylloplane with lepidopteran activity both in the
studies above and in that of (Smith & Couche, 1991) is in contrast to the findings of most surveys on the
natural occurrence of B. thuringiensis in soil, which have shown ‘non-toxic’ strains to be the most
common types (Hastowo et al., 1992; Ohba & Aizawa, 1986a, b; Ohba & Aratake, 1994). The serotyping of
the isolates in (Damgaard et al., 1997) showed that the majority of the isolates belonged to serovar
kurstaki. Isolation of B. thuringiensis from soil has shown to contain a very diverse population of serovars,
but never with a frequency of the insecticidal serovar kurstaki above 50% (Delucca et al., 1981; Ohba &
Aratake, 1994; Rongsen et al., 1990).
The high frequency of lepidopteran-active serovar kurstaki isolates found on foliage in this study indicates
that the (natural!) population of B. thuringiensis on phylloplane is different from that normally found in
soil. It is therefore likely that the phylloplane population is not exclusively the result of transfer of soil
bacteria to the foliage. Apparently some kind of propagation and/or selection of the B. thuringiensis
population takes place on the phylloplane. Bacillus thuringiensis was also discovered on the surface of
clover and other phylloplanes (Bizzarri & Bishop, 2007).
In soils Bacillus thuringiensis is ubiquitous: In a selection approach, (Travers et al., 1987) using a high
acetate medium to isolate Bt semi-selectively from soil and obtained over 8000 isolates. They claimed
that these isolates represented some 73 new biochemically distinct varieties of Bt. (Martin & Travers,
1989) found the insect control agent Bacillus thuringiensis to be a ubiquitous soil microorganism. They
isolated B. thuringiensis in 785 of 1,115 soil samples. These samples were obtained in the United States
and 29 other countries. A total of 48% of the B. thuringiensis isolates (8,916 isolates) fit the biochemical
description of known varieties, while 52% represented undescribed B. thuringiensis types. Over 60%
(1,052 isolates) of the isolates tested for toxicity were toxic to insects in the orders Lepidoptera or
Diptera. This kind of ubiquitous occurrence was again confirmed by (Jouzani et al., 2008) and (Haddad et

12
al., 2005) who verified that 77%, 78% and 80.5% of the effective doses (viable spores) remained on the
leaf surface after the first day of external Bt treatment, respectively.
In a recent comprehensive review Swiecicka (Swiecicka, 2008) widens the picture of natural occurrence of
Bacillus thuringiensis and its close relatives: While much is known about the taxonomic properties and
molecular basis for virulence of Bacillus thuringiensis and Bacillus cereus, comparatively less is known
about their ecology in natural environments. Thus, there are limited data regarding their resilience, i.e.
recycling of vegetative and sporulated phases of growth in soil, ecolgical niches including symbiotic
interactions with other organisms, and the impact on ecosystems in which they proliferate. Nevertheless,
based on recent data, apicture is beginning to emerge that B. thuringiensis and B. cereus are capable of
establishing mutual and commensal relationships with both animals and plants. In this regard, these
bacilli can proliferate in the digestive tracts of animals, where upon defecation they form dormant spores
in the soil, and to a lesser extent on the phylloplane and rhizospheres of plants.
Bacillus thuringiensis has been found in many more and diverse habitats (Federici, 1999), such as animal
feces, sludge, etc. (Hwang et al., 1998; Lee et al., 2003a; Mizuki et al., 2001; Okumura et al., 2001; Yu et
al., 1991).
But it should also be mentioned that the widespread use of Bt toxin sprays is the cause of numerous test
results published by (Frederiksen et al., 2006): A total of 128 Bacillus cereus-like strains isolated from
fresh fruits and vegetables for sale in retail shops in Denmark were characterized. Of these strains, 39%
(50/128) were classified as Bacillus thuringiensis on the basis of their content of cry genes determined by
PCR or crystal proteins visualized by microscopy. Random amplified polymorphic DNA analysis and
plasmid profiling indicated that 23 of the 50 B. thuringiensis strains were of the same subtype as B.
thuringiensis strains used as commercial bioinsecticides. Fourteen isolates were indistinguishable from B.
thuringiensis subsp. kurstaki HD1 present in the products Dipel, Biobit, and Foray, and nine isolates
grouped with B. thuringiensis subsp. aizawai present in Turex. The commercial strains were
primarily isolated from samples of tomatoes, cucumbers, and peppers. A multiplex PCR method was
developed to simultaneously detect all three genes in the enterotoxin hemolysin BL (HBL) and the
nonhemolytic enterotoxin (NHE), respectively. This revealed that the frequency of these enterotoxin
genes was higher among the strains indistinguishable from the commercial strains than among the other
B. thuringiensis and B. cereus-like strains isolated from fruits and vegetables. The same was seen for a
third enterotoxin, CytK. In conclusion, the present study strongly indicates that residues of B.
thuringiensis-based insecticides can be found on fresh fruits and vegetables and that these are potentially
enterotoxigenic.
(Federici & Siegel, 2007) summarize the enormous complexity of more than 70 varieties and subspecies of
Bacillus thuringiensis, there are more than 100’000 isolates that occur among the plasmids and insecticide
protein complements detected in the Bt isolates. 120 different types of genes are encoding Cyt proteins,
and at least 12 different types of genes encode Cyt proteins having been cloned and sequenced up to
now.
Usually, each type of Cry protein has an extremely limited target spectrum (lepidopteran, dipteran,
coleopteran, nematodes), and each specific protein like Cry1Ac is always much narrower than the type as
a whole, and even within a target category such as Lepidoptera (Rosi-Marshall et al., 2007b) there can be
marked differences from species to species: Cry1Ac is highly toxic to Heliothis virescens, but non-sensitive
to Spodoptera exigua.

13
1.1.1.3 Unfounded concerns about accumulation of Bt protoxins from GM crops in Water and
soil
Related to the ubiquitous occurrence of Bt toxins and also due to the fact that widespread cultivation of
crops with the endotoxins of Bacillus thuringiensis there is an environmental debate on the impact of Bt
toxins, further chapters will deal with the impact of Bt endotoxins on non-target insects in the agricultural
and non agricultural terrestrial systems, here in advance, included in the general introduction, some
remarks about aquatic and soil systems:
Aquatic systems
(Douville et al., 2007) tested the short time persistence of Bt proteins in aquatic systems. The Cry1Ab
gene persisted for more than 21 and 40 days in surface water and sediment, respectively. The removal of
bacteria by filtration of surface water samples did not significantly increase the half-life of the transgene,
but the levels were fivefold more abundant than those in unfiltered water at the end of the exposure
period. In sediments, the Cry1Ab gene from Bt corn was still detected after 40 days in clay- and sand-rich
sediments. Field surveys revealed that the Cry1Ab gene from transgenic corn and from naturally occurring
Bt was more abundant in the sediment than in the surface water. The Cry1Ab transgene was detected as
far away as the Richelieu and St. Lawrence rivers (82 km downstream from the corn cultivation plot),
suggesting that there were multiple sources of this gene and/or that it undergoes transport by the water
column. Sediment-associated Cry1Ab gene from Bt corn tended to decrease with distance from the Bt
cornfield. Sediment concentrations of the Cry1Ab gene were significantly correlated with those of the
Cry1Ab gene in surface water (R = 0.83; P = 0.04). The data indicate that DNA from Bt corn and Bt were
persistent in aquatic environments and were detected in rivers draining farming areas.
However, the authors also refer to their own previous study (Douville et al., 2005), where the results
showed that Bt-corn endotoxin is degraded more rapidly in water than in soils (t1/2: 4 and 9 days,
respectively), while crystals appeared to be more resilient, as expected. The isotopic patterns of 13C and
15N in Bt-corn endotoxin differed markedly from Bt, making it possible to track the source of Cry1Ab in
the environment. Preliminary field surveys indicate that Cry1Ab is fairly uncommon in aquatic
environments, being found only at trace concentrations when it is detected. This will say that Bt protoxins
are highly unlikely to cause any environmental problems in aquatic systems. As a whole, the publications
of Douville et al. are anyway not convincing, because they lack an important scientific quality: the baseline
comparison is totally lacking. As an example: There are several publications from the same river system,
such as (Tall et al., 2008) and many others which clearly point to metal and phosphorus contamination of
the river sediments, causing negative effects to the fauna and flora.
Critical reference is given to the paper of (Rosi-Marshall et al., 2007a) on the occurrence of Bt protein in
headwater stream ecosystems, written in an unnecessary alarming style and not even confirmed with
hard field data in the chapter on non-target insects of this report. There is not even a hint on the nature
of the Bt toxin (it could well be at least partially of natural origin), and when you compare her own (!)
figure 3 B the graph with realistic concentrations, then you see that Bt shows a clearly lower mortality of
the scraping caddisflies experiment – so what?? And again it shows, like Douville, the deadly sin in science
of a lacking a proper baseline comparison.

14
These comments are extended in a full rebuttal in the ASK-FORCE blog 1 of the major conclusions of (Rosi-
Marshall et al., 2007b), see also the published controversy about the case: (Beachy et al., 2008; Parrott,
2008; Rosi-Marshall et al., 2008).
Soil systems
The whole question on persistence of Bt toxins in soil is treated in a separate chapter in this report – there
are again, after a first wistle blower phase (Saxena et al., 1999), enough long term studies to demonstrate
that accumulation does not take place to a degree that it could harm soil organisms, here just as an
example two papers: (Head et al., 2002; Saxena & Stotzky, 2001):
1.1.2.Bacillus thuringiensis external applications
1.1.2.1. External Bt applications with a long tradition, but remaining marginal in use.
Bacillus thuringiensis toxins have a long tradition in organic and conventional agriculture for over a 100
years as external sprays (Nester et al., 2002) and (Croft, 1990; Federici & Siegel, 2007; Flexner et al., 1986;
Glare & O'Callaghan, 2000; Hickle & Fitch, 1998; Krieg, 1968; Meher et al., 2002; Metz, 2004; Punja, 1997).
Bt toxins were also recommended as pesticide spray by Rachel Carson in her classic book The Silent Spring
(Carson, 1962 - 2002). It can be assumed that Rachel Carson would have welcomed the new Bt crops as a
more elegant solution to fight the European corn borer within the maize stems.
According to (Federici & Siegel, 2007) there are four major subspecies/serovarieties (kurstacki, aizawai,
morrisoni and israelensis with a total of 13 Bt proteins in use for bacterial insecticides.
In numerous risk assessments those sprays have revealed to be unproblematic related to environmental
impact, and provided the prescriptions are respected: The effects of the commercial Bt spray Dipel on
green lacewings and its prey herbivores (aphids, spider mites, and lepidopteran larvae) were compared to
those of transgenic Bt maize in a study (Dutton et al., 2003). The field studies reveal small differences, on
an agronomic scale they are negligible. However, another safety assessment of Bt spray applications of
Bishop et al. concludes with some critical remarks (Bishop et al., 1999):
“commercial insecticide containing B. thuringiensis was sprayed onto spinach leaves. After normal food
preparation regimes some leaves retained residual spore loads sufficient for a strongly enterotoxic strain to
cause food poisoning in humans. These findings suggest that the agricultural use of some, previously
unvalidated, strains of B. thuringiensis could give rise to cases of food poisoning and that rodents are
unsuitable for testing the safety to humans of oral exposure to this organism.”
The same authors dismiss for external Bt applications food safety concerns after experimenting with rats:
1 Ammann, K. ASK-FORCE blog on the impact of Bt toxins on aquatic organisms,
http://www.botanischergarten.ch/AF-3-Aquatic-Bt/AF-3-Aquatic-Bt-toxins-20100423-opensource.pdf

15
“Six strains of Bacillus thuringiensis were tested with two commercially available kits for their ability to produce
Bacillus cereus-type enterotoxin and by dipteran bioassay for the production of beta-exotoxin. All of the strains were
positive for enterotoxin production including three which have been used world-wide for many years to control pest
insects. Rats given oral doses totaling 1 x 10(12) spores ( +crystals), over three weeks, or a single subcutaneous dose
of 1 x 10(6) spores ( +crystals) showed no ill-effects in terms of their condition or in the pathology of their internal
organs: this was in spite of the strain of B. thuringiensis used (13B) being an active producer of both beta-exotoxin
and enterotoxin”
In a comparative study with 20 kinds of pesticides, the microbial formulations containing Bacillus
thuringiensis revealed to be harmless after having been tested against 19 beneficial arthropodes (Sterk et
al., 1999).
Overall, concerns about safety of Bt insecticides for humans can be dismissed. (Glare & O'Callaghan, 2000;
McClintock et al., 1995; Siegel, 2001). (Federici & Siegel, 2007) produce extensive results in table 3.6 p. 67,
results show, despite of the very large amounts of test materials used in these studies, which are 100- to
more than 1000-fold the amount of material used to control insect pests: as a result most assessments of
the safety of Bt to humans are based on a lack of reported effects, i.e., the overall lack of reported
infections or other documented cases of disease, especially in areas where human populations numbering
in the tens of thousands have been exposed to Bt applications during aerial spray programs to eliminate
lepidopteran forest pests (de Amorim et al., 2001; Pearce et al., 2002; Petrie et al., 2003; Teschke et al.,
2001).
1.1.2.2. Bt sprays cause resistant insects, but not (yet) Bt crops
There are several cases documented already in the early nineties and later, where insects developed
resistance against Bt formulations sprayed in the fields and in greenhouses: (Li et al., 2005; Tanaka &
Kimura, 1991). Already in 1994 and 1995 there was growing and well documented concern about evolving
pest resistance against Bt toxins used in sprays (Bauer, 1995; McGaughey, 1994). On the other hand,
despite massive use of Bt crops, resistance to the Bt endo-toxin remains rare up to now, although it can
be detected occasionally in the field with rather costly analysis, it does not pose a problem up to now
(Bates et al., 2005; Bourguet et al., 2005; Shelton et al., 1993; Tabashnik et al., 2006; Tabashnik et al.,
2008). The most recent paper of Tabashnik et al. 2008 (Tabashnik et al., 2008) is often cited as now having
provided the ‘ultimate proof’ with field data that Bt resistance for some Bt cotton pests has been
detected, which is fact, but only in exceptional cases and (not yet) causing any agronomic problems. Here
the authors own comments:
“Nonetheless, resistance of H. zea to Cry1Ac [endotoxin] has not caused widespread control failures for
several reasons. First, even in the few states with documented resistance, most populations tested were
not resistant to Cry1Ac. Second, insecticides have been used from the outset to augment control of H. zea
on Bt cotton because Cry1Ac alone is not sufficient to control high-density populations of the pest (EPA
Environmental Protection Agency, 1998; Jackson et al., 2004b). Insecticide sprays decrease any problems
associated with reduced control of H. zea by Bt cotton. Third, against strains with 44- to 100-fold
resistance to Cry1Ac, the Cry1Ac in Bt cotton still caused 48–60% larval mortality (Ali et al., 2006; Jackson
et al., 2004a; Luttrell et al., 2006),. Finally, ‘pyramided’ transgenic cotton producing Bt toxins Cry2Ab and

16
Cry1Ac was registered in December 2002 and planted on more than 1 million ha in the United States in
2006 and 2007 (Monsanto Co., 2002). Control of Cry1Acresistant H. zea by Cry2Ab also limits problems
associated with resistance to Cry1Ac19.”
This is also due to good agricultural practice with refuge areas, and, as (Jackson et al., 2008) reports, due
to ecological factors revealed in the latest field assessment:
“These data demonstrate that Helicoverpa. zea adults move extensively from their natal host origins. Therefore,
non-cotton crop hosts, and even relatively distant hosts, contribute significantly to effective refuge for H. zea on Bt
cotton. The results presented here demonstrate that substantial natural refuge is present for Bt-resistance
management of H. zea throughout the mid-South and Southeast portions of the US cotton belt.”
As (Moar et al., 2008) rightly mention, the two publications of (Ali et al., 2006; Luttrell et al., 1999) cannot
be used (as (Tabashnik et al., 2008) did), as an argument of raising resistance of cotton pests:
“We emphatically disagree with the conclusions of (Tabashnik et al., 2008) that the data published in
these two articles demonstrate field-evolved resistance in H. zea for four reasons: first, the definition of Bt
resistance used by Tabashnik et al. is purely laboratory based, whereas field efficacy and larval survival on
plant tissues are the ultimate criteria for contextualizing laboratory-based estimates of resistance, and no
change in Bt cotton efficacy has been documented during the past decade; second, larval samples should
not be collected from Bt crops because they will not be representative of the population as a whole,
especially for highly mobile insects such as H. zea; third, the data from Luttrell's laboratory on which
Tabashnik et al. base their conclusions have been evaluated using LC50 (median lethal dose; 50%) values to
measure resistance, which introduces artifacts into the analysis; and fourth, the baseline comparator used
to assess variability in these laboratory assays is not representative of field susceptibility; when a more
appropriate comparator colony is employed, results from Luttrell's laboratory bioassays indicate no
change in susceptibility. We discuss each of these aspects in turn below.”
In a recent paper, (Tabashnik et al., 2009b) points to the fact, that Asymmetrical cross-resistance between
Bacillus thuringiensis toxins Cry1Ac and Cry2Ab in pink bollworm has been detected, again not in the field,
but with laboratory experiments.
From the abstract:
“We show here, however, that laboratory selection of pink bollworm with Cry2Ab caused up to 420-fold
crossresistance to Cry1Ac as well as 240-fold resistance to Cry2Ab. Inheritance of resistance to high
concentrations of Cry2Ab was recessive. Larvae from a laboratory strain resistant to Cry1Ac and Cry2Ab in
diet bioassays survived on cotton bolls producing only Cry1Ac, but not on cotton bolls producing both
toxins. Thus, the asymmetrical cross-resistance seen here does not threaten the efficacy of pyramided Bt
cotton against pink bollworm. Nonetheless, the results here and previous evidence indicate that
crossresistance occurs between Cry1Ac and Cry2Ab in some key cotton pests. Incorporating the potential
effects of such cross-resistance in resistance management plans may help to sustain the efficacy of
pyramided Bt crops.”
In the most recent paper, Tabashnik et al. (Tabashnik et al., 2009a) come to the following conclusions,
summed up in a press release of the Entomological Society of America
http://www.entsoc.org/resources/press_releases/2009_btcrops.htm

17
“According to lead author Dr. Bruce E. Tabashnik, “Resistance is not something to be afraid of, but
something that we expect and can manage if we understand it. Dozens of studies monitoring how pests
have responded to Bt crops have created a treasure trove of data showing that resistance has emerged in
a few pest populations, but not in most others. By systematically analyzing the extensive data, we can
learn what accelerates resistance and what delays it. With this knowledge, we can more effectively predict
and thwart pest resistance.”
Among the authors’ conclusions are:
The refuge strategy (growing non-Bt crops near the Bt crops) can slow the evolution of insect
resistance by increasing the chances of resistant insects mating with non-resistant ones, resulting in non-
resistant offspring.
Crops that are “pyramided” to incorporate two or more Bt toxins are more effective at controlling
insect resistance when they are used independently from crops that contain only one Bt toxin.
Resistance monitoring can be especially effective when insects collected from the field include
survivors from Bt crops.
DNA screening can complement traditional methods for monitoring resistance, such as exposing
insects to toxins in the lab.
Despite a few documented cases of field-evolved resistance to the Bt toxins in transgenic crops, most
insect pest populations are still susceptible.
With Bt crop acreage increasing worldwide, incorporating enhanced understanding of observed patterns
of field-evolved resistance into future resistance management strategies can help to minimize the
drawbacks and maximize the benefits of current and future generations of transgenic crops.”
Consequently, again we will have a situation, where opponents to the technology will not read the
publication properly and herald loudly that the reistance against the pink bollworm and other major
cotton pests is now agricultural reality – which might well develop in the future, if the measures proposed
by Tabashnik in the same paper are not translated into reality (based on more field-research).
It is enigmatic to the author of this report why then certain proponents of organic farming, who use Bt
spray formulations – often not very wisely – are scared that Bt crops could make their own microbial
pesticides worthless by triggering Bt resistance to major pest insects (Greenpeace, 2002; Wallimann,
2000) – the facts show another picture. The reference list of literature on external Bt applications
collected counts already now over 300 items.
http://www.botanischergarten.ch/Bibliography/Bibliography-Bt-Sprays-20080316.pdf
In a final chapter the whole question about developing Bt resistance of target pests will be discussed in
extenso. Although up to now no resistance has developed of agronomic importance, this will almost
certainly happen in the future. The answer against will be stacked Bt genes or also artificially enhanced Bt
genes (Christou et al., 2006) – but read about the caveats in (Tabashnik et al., 2009b) commented above.

18
According to (de Maagd, 2007) applications of Bt sprays nowadays, consisting of some form of
spore/crystal-mixture, are found in three major areas:
1. Forest pest control. Particularly in North America, aerial Bt-sprays are used extensively for control of
forest pests (spruce budworm, gypsy moth).
2. Mosquito control. Particularly in the Middle East and Africa, but also in Europe (German Rhine
valley), sprays are used for control of mosquitoes, such as the vectors for malaria.
3. Organic agriculture. As Bt sprays are considered a natural pesticide, it is one of the few pesticides
that can be used in organic agriculture (particularly on horticultural crops).
Despite its attractiveness as a natural pesticide, Bt has never conquered a large share of the global
pesticide market. Although it is the most widely used biological pesticide, it takes up only about 1% of the
total insecticide market. Several reasons for this can be identified:
Low persistence. The crystal protein is rapidly inactivated by solar UV-radiation.
Limited activity spectrum. Each Bt strain is active only against a few pest species, so one
product is never sufficient for all pests encountered in the field.
Many important pest species are insensitive to all known Bt strains.
Bt sprays, as many chemical insecticides, are not very effective against insects that bore
into the crop tissue. There may be only a limited time window in which sprays can be
effective. This is particularly true for cotton bollworm and European corn borer. This
requires extensive monitoring by farmers to time spraying properly.
According to (Dutton et al., 2003) Dipel (a commercial Bt spray) had no effect on aphids;
however, negative effects on spider mites were observed. Spider mites reared on Bt-
sprayed plants had a significantly lower intrinsic rate of natural increase compared to
those reared on control plants. Similarly, S. littoralis larvae were significantly affected by
Dipel as the developmental time required by larvae which were fed Bt-sprayed plants was
prolonged when compared to larvae on untreated plants. Negative effects on C carnea
larvae were also shown through prey-mediated exposure to Dipel.
In contrast to those effects, the results of (Romeis et al., 2004) strongly suggest that C.
carnea larvae are not sensitive to Cry1Ab and that earlier reported negative effects of Bt-
maize were prey-quality mediated rather than direct toxic effects. These results, together
with the fact that lepidopteran larvae are not regarded as an important prey for C. carnea
in the field, led the authors to conclude that transgenic maize expressing Cry1Ab poses a
negligible risk for this predator, a clear advantage for the transgenic strategy of Bt
applications.

19
1.1.3.Transgenic Bt maize
1.1.3.1. Conventional breeding of insect resistant maize not very successful
Attempts about conventional breeding of insect resistant maize without using genetic engineering have
been often made, but so far they have not been successful (Bohn et al., 2001; Bohn et al., 1996; Bohn et
al., 1997; Bohn et al., 2003). The key parameter for conventional marker assisted breeding is The key
parameter for assessing the prospects of Marker Assisted Breeding (MAS) is a high number of selection
plants in large populationsIn order to obtain reliable and high values for quantitative traits, large
population sizes and (n>500) and a high number of test environments have to be employed. However,
mandatory large-scale experiments are not an option for most breeders due to financial and logistic
restrictions
1.1.3.2. How it all started, the (re)view of two initiators
In a review, Herman Höfte from Plant Genetic Systems N.V. in Gent, Belgium and H.R. Whiteley from the
University of Washington in Seattle (Hofte & Whiteley, 1989) present an comprehensive update of the
knowledge of the late eighties of B. thuringiensis crystal proteins C and their genes. It offers an excellent
insight in the pioneer times of Bt crop development. They also propose a nomenclature and classification
scheme for crystal proteins based on their structure (deduced from the deoxyribonucleic acid DNA
sequence as well as their host range which later has been thoroughly changed). Nevertheless, their review
let us understand how the Bt insect resistance as a major strategy of pest control in modern agriculture
was developed:
“Bacillus thuringiensis is a gram-positive soil bacterium characterized by its ability to produce crystalline
inclusions during sporulation. These inclusions consist of proteins exhibiting a highly specific insecticidal
activity (reviewed in references (Aronson et al., 1986; Whiteley & Schnepf, 1986). Many B. thuringiensis
strains with different insect host spectra have been identified (Burges, 1981)) They are classified into
different serotypes or subspecies based on their flagellar antigens. Most strains are active against larvae
of certain members of the Lepidoptera, but some show toxicity against dipterian (reviewed in (Federici et
al., 1991)) or coleopteran (Krieg et al., 1983) species. For several crystal-producing strains. no toxic
activity has yet been demonstrated. B. thuringiensis crystalline inclusions dissolve in the larval midgut,
releasing one or more insecticidal crystal proteins (also called delta-endotoxins) of 27 to 140 kilodaltons
(kDa). As described in the following section, most crystal proteins are protoxins that are proteolytically
converted into smaller toxic polypeptides in the insect midgut. The activated toxin interacts with the
midgut epithelium cells of susceptible insects. Electrophysiological (Harvey et al., 1983) and biochemical
(Knowles & Ellar, 1987) evidence suggests that the toxins generate pores in the cell membrane, thus
disturbing the osmotic balance. Consequently, the cells swell and lyse. The larva stops feeding and
eventually dies. For several B.thuringiensis toxins, specific high-affinity binding sites have been
demonstrated to exist on the midgut epithelium of susceptible insects (Hofmann & Luthy, 1986; Hofmann
et al., 1988a). This could, at least in part, explain the extreme specificity of these proteins.
Formulations of B. thuringiensis have been used for more than two decades as biological insecticides to
control agricultural pests and, more recently, insect vector-s of a variety of human and animal diseases.
Recently, the cloning of insecticidal crystal protein genes (Whiteley & Schnepf, 1986) and their expression

20
in plant-associated microorganisms (Obukowicz et al., 1986) or transgenic plants (Barton et al., 1987;
Fischhoff et al., 1987; Vaeck et al., 1987) has provided potentially powerful alternative strategies for the
protection of crops against insect damage. These applied aspects are to a large extent responsible for an
increased interest in this bacterium and its crystal proteins in recent years. Extensive screening programs
are being carried out by various groups to search for B. thuringiensis strains with new insecticidal spectra.
Numerous publications report the identification of crystal proteins and the cloning and sequencing of
crystal protein genes. One problem related to this is the lack of a uniform nomenclature for these genes
and their products, which makes the literature rather confusing.”
More molecular details can be found in a previous publication of the same author team: (Hofte et al.,
1986).
Some years later, the classification has been changed and based completely on the molecular structure of
the growing number of proteins, first on the sequence of amino acids after (Crickmore et al., 1998). Today
the classification is entirely built on phylogenetic relationships of the molecular structure, as shown below
in Fig. 4-6 from (de Maagd et al., 2001). For the latest views on taxonomy and nomenclature of the Bt
proteins see the website with a regularly updated database structure of (Crickmore et al., 2008)
http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/ .
1.1.3.3. First cloning of a Bt gene
The first gene coding for an insecticidal protein was cloned by Schnepf and Whiteley (Schnepf &
Whiteley, 1981), working with the HD 1 isolate of B. thuringiensis kurstaki, the strain used in the
commercial product Dipel. Following this, Whiteley, her co-workers, and others showed that there were
actually four endotoxin genes in this strain that coded for insecticidal proteins. According to Sussex
(Sussex, 2008), the following three research groups under Jeff Schell, Rob Horsch, and Mary-Dell Chilton
(Chilton et al., 1978; Chilton et al., 1977; Chilton et al., 1980; Drummond et al., 1977) all reported at a
Miami Winter Symposium in January 1983 success in producing chimeric genes that functioned in
transformed plant cells (Downey et al., 1983). All three groups used the nopaline synthetase (NOS)
promoter spliced to the bacterial NPT II (neomycin phosphotransferase) coding sequence as a dominant
selectable marker and NOS polyadenylation signals or a variation of this strategy. NOS was discovered by
(Van Montagu et al., 1980; Willmitzer et al., 1980; Willmitzer et al., 1983).
1.1.3.4. Genetically engineered insect resistant Bt maize
1.1.3.4.1. Transformation with Agrobacterium tumefaciens
Transformation with Agrobacterium was difficult in the beginning: The graminaceous monocots, including
the economically important cereals, seem to be refractory to infection by Agrobacterium tumefaciens, a
natural gene transfer system (De Cleene & Deley, 1976) that has been successfully exploited for
transferring foreign genes into higher plants (Caplan et al., 1983; De Block et al., 1984; Fraley et al., 1983;
Zambryski et al., 1984). Therefore, direct transfer techniques that are potentially applicable to all plant
species have been developed using a few dicot and monocot species as model systems (Fromm et al.,
1985; Fromm et al., 1986; Lorz et al., 1985; Potrykus et al., 1985). Only recently have we begun to
understand how Agrobacterium hijacks host factors and cellular processes during the transformation
process (Dafny-Yelin et al., 2008; Lacroix et al., 2006; Tzfira & Citovsky, 2006).

21
Attempts were successful in the Ghent school, first by systematically working out the procedures on how
to transfer plasmids of Agrobacterium tumefaciens to higher plants under the lead of Jeff Shell and Marc
van Montagu: (Deblock et al., 1985; Genetello et al., 1977; Holsters & Schell, 1975; Schell, 1977; Van
Montagu, 1997; Van Montagu et al., 1980; Van Montagu et al., 1983; Vanlarebeke et al., 1977). Later the
Gent research group also succeeded to transfer bacterial genes with the same methods into tobacco
plants. The first peer reviewed papers on a successful transformation were published by Herrera-Estrella
et al. including the senior authors Shell and van Montagu (Herrera-Estrella et al., 1983a; Herrera-Estrella
et al., 1983b). The genes, cloned from a Bacillus thuringiensis strain were expressed in transgenic
tobacco, which synthesized insecticidal proteins protecting the plants from feeding damage by larvae of
the tobacco hornworm (Fischhoff et al., 1987; Vaeck et al., 1987). Another early and successful attempt to
express Bt endotoxines Tobacco plants was published by (Barton & Umbeck, 1989; Barton et al., 1987).
1.1.3.4.2. Transformation with biolistic methods
The most successful and widely used transformation methods for maize and many other crops was
particle bombardment by the gene gun, developed by John Sanford and colleagues at Cornell University in
1984 (Klein et al., 1987; Sanford, 2000). Another success with electroporation was published by Fromm
(Fromm et al., 1986).
Also Rhodes achieved with electroporation transgenic protoplasts containing plasmid DNA with a
gene coding for neomycin phosphotransferase NPr II), from which the first transgenic maize
plants were grown (Rhodes et al., 1988). Two years later, it was the research group of Gordon-
Kamm (Gordon-Kamm et al., 1990) who created a reproducible system for the generation of
fertile, transgenic maize plants. Details from the recent historical account of Sussex (Sussex,
2008)
“These workers at DEKALB Plant Genetics (now Monsanto) bombarded cells from maize embryogenic suspension
cultures with tungsten particles coated with plasmids containing the selectable marker gene bar. This gene confers
resistance to the herbicide bialaphos, which was used to select transformed callus cells. Transformed calli were
shown to contain the integrated bar gene and to express the enzyme phosphinothricin acetyltransferase encoded by
bar. Fertile transformed plants were produced from the calli, and of 53 progeny tested, 29 had phosphinothricin
acetyltransferase activity. In other experiments, they cotransformed embryogenic suspension culture cells with a
mixture of two plasmids, one containing the bar gene and the other containing the gene encoding b-glucuronidase.
Regenerated plants expressed both genes. The authors concluded that ‘‘this system provides a new, powerful tool
for both the study of basic plant biology and the introduction of important agronomic traits into one of the world’s
major crops’’
See also Beegle, Shah and Songstad for further details and references (Beegle & Yamamoto, 1992; Shah et
al., 1987; Songstad et al., 1993).
In 1995 (Hill et al., 1995) followed with the first Bt maize, where the transgene has been inserted with
biolistic methods. Ever since then, the race to apply this technology and to make use of such genetically
improved pest resistant cultivars has been rapid and intense, it is summarized in a comprehensive way by
Moellenbeck and Sairam (Moellenbeck et al., 2001; Sairam et al., 2005).

22
A more specific view on Bt crops and their early development has been summarized by (Peferoen, 1997),
see also (Bohorova et al., 1999; Bohorova et al., 1995a; Bohorova et al., 1995b). Some early critical and
very detailed notes on the overall performance of Bt crops were given by (Hilder & Boulter, 1999; Obrycki
et al., 2001), they point to early developing difficulties and knowledge gaps of the eighties and early
nineties and offer thus indirectly the assurance, that performance and safety of the latest Bt crops is
nowadays considerably enhanced. Some more insight in the early development stories of the US is given
by (Koziel et al., 1993; Koziel et al., 1996).
New evidence shows, that the MON810 Yield Gard maize transgene situation is more complex than
anticipated:
Rosati, A., P. Bogani, A. Santarlasci and M. Buiatti (2008). "Characterisation of 3 ' transgene insertion site and
derived mRNAs in MON810 YieldGard (R) maize." Plant Molecular Biology 67(3): 271-281.<Go to
ISI>://WOS:000255414000006 AND http://www.botanischergarten.ch/Bt/Rosati-Characterisation-Mon810-publ-
2008.pdf
“The construct inserted in YieldGard (R) MON810 maize, produced by Monsanto, contains the CaMV 35S
promoter, the hsp70 intron of maize, the cryI(A)b gene for resistance to lepidopterans and the NOS terminator. In a
previous work a truncation event at the 3' end of the cryI(A)b gene leading to the complete loss of the NOS
terminator was demonstrated. The 3' maize genome junction region was isolated in the same experiment not
showing any homology with known sequences. The aim of the experiments here reported was therefore to isolate
and characterize a larger portion of the 3' integration junction from genomic DNA of two commercial MON810 maize
lines. Specific primers were designed on the 3' integration junction sequence for the amplification of a 476 bp
fragment downstream of the sequence previously detected. In silico analysis identified the whole isolated 3' genomic
region as a gene putatively coding for the HECT E3 ubiquitin ligase. RT-PCR performed in this region produced cDNA
variants of different length. In silico translation of these transcripts identified 2 and 18 putative additional
aminoacids in different variants, all derived from the adjacent host genomic sequences, added to the truncated
CRY1A protein. These putative recombinant proteins did not show homology with any known protein domains. Our
data gave new insights on the genomic organization of MON810 in the YieldGard (R) maize and confirmed the
previous suggestion that the integration in the genome of maize caused a complex recombination event without,
apparently, interfering with the activity of the partial CRY1A endotoxin and both the vigor and yield of the YieldGard
(R) maize.” (Rosati et al., 2008)
The absence of the NOS terminator in the MON810 transgene was already suggested by (Hernandez et
al., 2003)
1.1.3.4.3. The breakthrough to the widespread industrial production of Bt crops
Though with many of the earliest introduced transgenic traits the primary benefits go to growers, some of
those crops also provide secondary benefits to consumer health (Kershen, 2006). Bt corn is such an
example (there will be an extensive chapter dedicated to its proven low mycotoxin contents later in this
report). Transgenic maize is one of the most commonly grown transgenic crops in the world today. On a
commercial basis and regulated in numerous countries it contains genes from Bacillus thuringiensis,
encodes proteins toxic mainly to a very few members of the order Lepidoptera (Butterflies), Diptera (Flies)
and Coleoptera (Beetles). These include the common corn pests European corn borer Ostrinia nubilalis,
South Western corn borer Diatraea grandiosella, corn earworm Helicoverpa zea and corn rootworm
Diabrotica spp. More testing has revealed a limited toxicity to the following groups of insects: Orders of

23
Lepidoptera, Coleoptera, Hymenoptera, Homoptera, Dictyoptera, Orthoptera and Mallophaga, in addition
to nematodes (Strongylida, Tylenchida), protozoa (Diplomonadida) and Acari (mites) (Aronson & Shai,
2001; Ballester et al., 1999; Bravo et al., 2007; de Maagd et al., 1999; English & Slatin, 1992; Gahan et al.,
2005; Gomez et al., 2006; Gomez et al., 2003; Gomez et al., 2001; Gomez et al., 2007; Griffitts et al., 2005;
Griko et al., 2007; Heckel et al., 2007; Jurat-Fuentes & Adang, 2006; Munoz-Garay et al., 2006; Pardo-
Lopez et al., 2006; Potvin et al., 1998; Rang et al., 1999; Rausell et al., 2000b; Rausell et al., 2004b; Rausell
et al., 2004c).
The latest assessments come from ISAAA briefs (James, 2009) and a recent ENDS-Report (ENDS Report,
2008).
The benefits of Bt maize have been summarized several times in great detail by (Brookes, 2008b; Brookes
& Barfoot, 2008; Brooks et al., 2005b; Carpenter, 2001; Carpenter et al., 2001; Carpenter et al., 2004;
Sankula & Blumenthal, 2004; Silvers & Gianessi, 2001)
See also the chapter on the present and future development of new Bt maize traits, the research and
development within the companies and in public research is in full expansion, mainly for reasons to
overcome beginning Bt resistance to pests and also in order to enlarge the palette of target pests etc.
As a selected example on the situation of the industrial development a table is given here from the
comprehensive OECD report (OECD Consensus Documents, 2007): Consensus Document on Safety
Information on Transgenic Plants Expressing Bacillus thuringiensis - Derived Insect Control Protein. The
consensus document provides a lot of basic information about Bt maize toxins and their industrial use in
maize.
The table below demonstrates the various expression levels of bt crops across the industrially developed
traits:
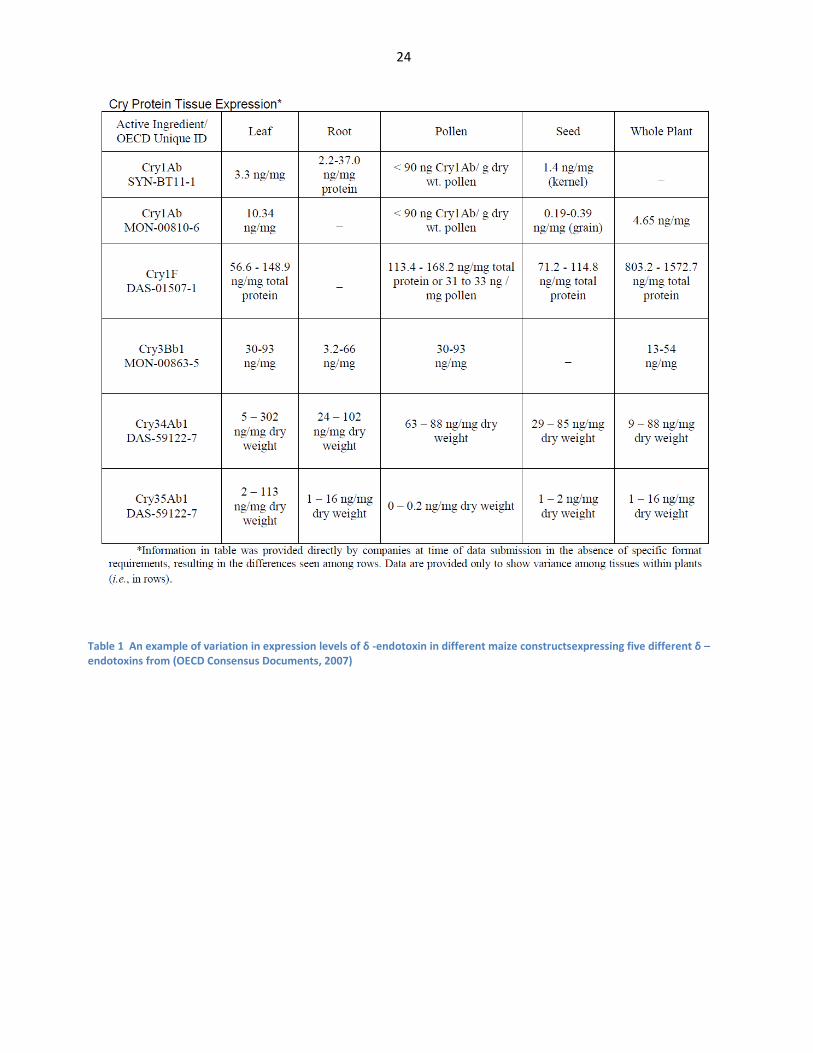
24
Table 1 An example of variation in expression levels of δ -endotoxin in different maize constructsexpressing five different δ –endotoxins from (OECD Consensus Documents, 2007)
25
1.1.4. Chemistry, Biology and nomenclature of Bt toxins
If we want to understand the full advantages of the Bt insect tolerance strategy, we will have to analyse
some of the mechanisms on how the Bt toxins work:
The mode of action of the Bt toxins on a restricted set of insect species is complex and varies from case to
case of the specific protein employed. Most studies on toxicity mechanisms are only available since a few
years (Broderick et al., 2006). It is interesting to note that Bacillus thuringiensis requires enterobacteria in
the midgut of the target insect to unfold its insecticidal activity described already in detail by (Broderick
et al., 2009; Broderick et al., 2003; Broderick et al., 2000; Broderick et al., 2004; Crickmore et al., 1998;
Schnepf et al., 1998). The extremely targeted and restricted toxic impact on a few lepidopteran species is
the major reason for the success of the pest management with Bt proteins. See for more details in 1.1.5.
below and also in the chapter on food safety.
Numerous papers have been published on the nomenclature and chemistry of Bt toxins, and still today
there is some confusion about classification. Some basic papers have been summarized by de Maagd 2001
(de Maagd et al., 2001), including some very helpful illustrations on the Bt toxins.
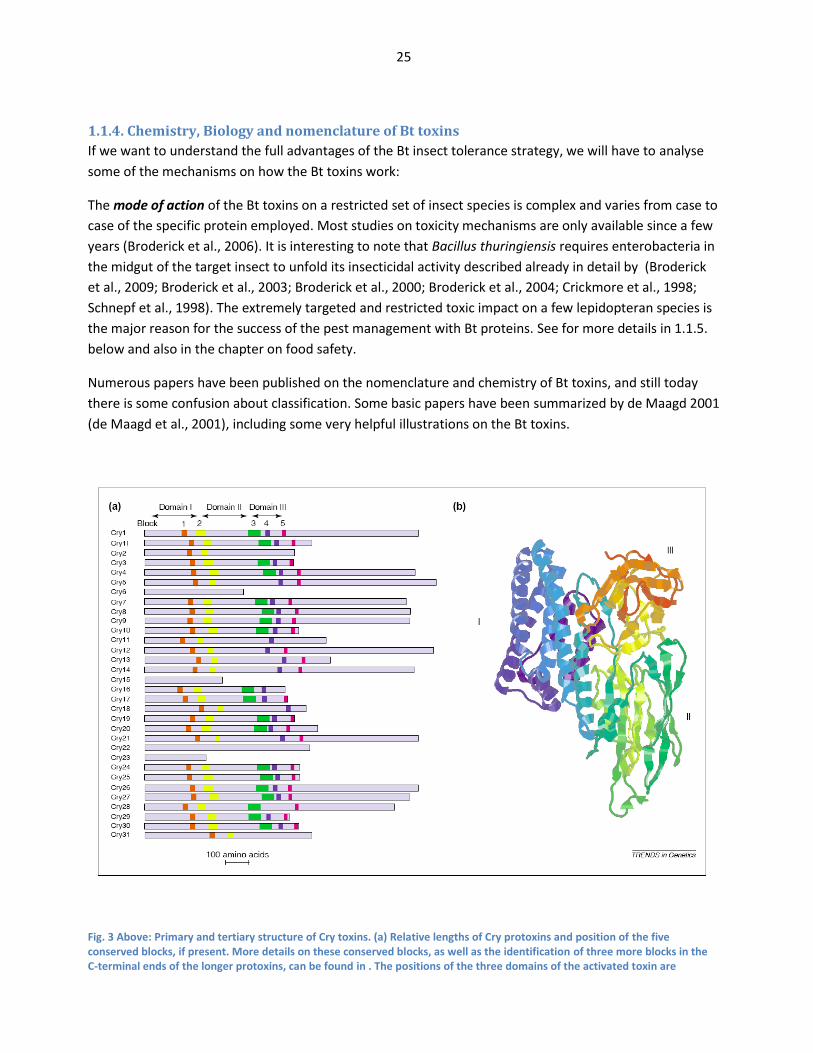
Fig. 3 Above: Primary and tertiary structure of Cry toxins. (a) Relative lengths of Cry protoxins and position of the five conserved blocks, if present. More details on these conserved blocks, as well as the identification of three more blocks in the C-terminal ends of the longer protoxins, can be found in . The positions of the three domains of the activated toxin are

26
indicated for Cry1 and vary with the positions of blocks 2 and 3 for the other toxins. The remainder of the protoxin, consisting of short N-terminal part (20–40 amino acids) preceding the first domain and the C-terminal part following the third domain in the longer protoxins, is digested away by gut proteases during the activation process. (b) Three-dimensional structure of an activated toxin, Cry1Aa (Ref. 7). The toxin has three structural domains. Domain I (blue) is involved in membrane insertion and pore formation. Domain II (green) and domain III (yellow-red) are involved in receptor recognition and binding. Conserved block 1 is in the central helix of domain I, block 2 is at the domain I–II interface, block 3 is at the boundary between domains II and III, block 4 is in the central β-strand of domain III and block 5 is at the end of domain III. From (de Maagd et al., 2001)
Fig. 4 Transmission electron micrograph of a sporulating Bacillus thuringiensis (Bt) cell. δ-Endotoxins are produced as regularly shaped crystals (PB; protein body) – hence the name crystal (Cry) proteins – next to a spore (SP). The vegetative cell wall will eventually break to release the spore and crystal. The cell shown is approximately 2 μm long. From (de Maagd et al., 2001)
Fig. 5 Spores and crystals of Bacillus thuringiensis serovar morrisoni strain T08025 Microscopy by Jim Buckman from
http://commons.wikimedia.org/wiki/File:Bacillus_thuringiensis.JPG

27
De Maagd et al. (de Maagd et al., 2001) point rightly so to the fact that there also exist other toxic
substances in the arsenal of Bacillus thuringiensis, they might also be used in the development of new
insect resistant crops:
“Besides the Cry proteins (PB in Fig. 2), which are often the only focus, cytolysins (Cyt toxins), which act by a different
mechanism, are also found within the crystal. Bt produces various virulence factors other than the crystal proteins,
including secreted insecticidal protein toxins, α-exotoxins, β-exotoxins, hemolysins, enterotoxins, chitinases and
phospholipasesa (Hansen & Hendriksen, 2001). The spore itself contributes to pathogenicity, often synergizing the
activity of the crystal proteins (Bernstein et al., 1999; Bernstein et al., 2003; Johnson et al., 1998). All of these factors
might have a role in insect pathogenesis under natural conditions, helping the bacterium to develop in the dead or
diseased insect larvae, but the exact contribution of each factor is often unknown. Although the Cry proteins are
commonly referred to as ‘Bt toxins’, a few Cry proteins were found in Bacillus popilliae and in Clostridium
bifermentans” (Schnepf et al., 1998)
Some years later, the classification has been changed, it is now based completely on the molecular
structure of the growing number of proteins, first on the sequence of amino acids (Crickmore et al., 1998).
Today the classification is entirely built on phyogenetic relationships of the molecular structure, as shown
below in Fig. 4-6 from de Maagd (de Maagd et al., 2001). For the latest views on taxonomy and
nomenclature of the Bt proteins see the website with an updated database structure of Crickmore
(Crickmore et al., 2008) http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/.
Fig. 6 Scanning electron microscopy of spore (s) and crystals (c) of strains S285 (1); S447 (2); S479 3); S550 (4); S1255 (5); and Bti (6). From (Monnerat et al., 2005) Most of the Bt proteins toxic to Mosquitos have a round shape, in contrast to the Bt proteins usually inserted in Bt maize.

28
Separate phylogenetic analysis of the three domains of the active toxin is more likely to yield interesting
insights into how the extensive variety in structure and specificity came into existence (Bravo, 1997;
Crickmore et al., 1998). Such an analysis is shown in the following 3 figures below, domains I – III, with the
insect-order specificity shown as color-coded branches, redrawn from (Bravo, 1997). The role of the
protoxin-specific part in the evolutionary development of insect specificity should not be discarded
however, as the presence of a C-terminal extension might affect activity as mentioned above for Cry1Ba
and Cry7Aa. A classic example is the co-evolution of Spodoptera frugiperda Smith (Lepidoptera :
Noctuidae) populations from Latin America, parallel to its susceptibility with various Bt Cry toxins
(Monnerat et al., 2006)
A brief comparison of the trees for domains I and II shows that their overall structure is very similar,
having identical main branches that correspond to the current classification of the protoxins; that is, the
nearest neighbors of Cry1Aa are the other Cry1A toxins, the nearest neighbor of Cry1Ca is Cry1Cb, and so
on. Domains I and II of Cry9Aa behave differently; they don’t cluster with the other Cry9 toxins. The
classification of Cry9Aa is therefore primarily based on its homology with Cry9Ea in domain III and with all
Cry9 proteins in the protoxinspecific part31,32, suggesting that Cry9Aa evolved independently from the
other Cry9 proteins but obtained a Cry9-specific C-terminal extension more recently, perhaps by a
recombination event. See also the section of Cry9Ab in the food safety chapter.
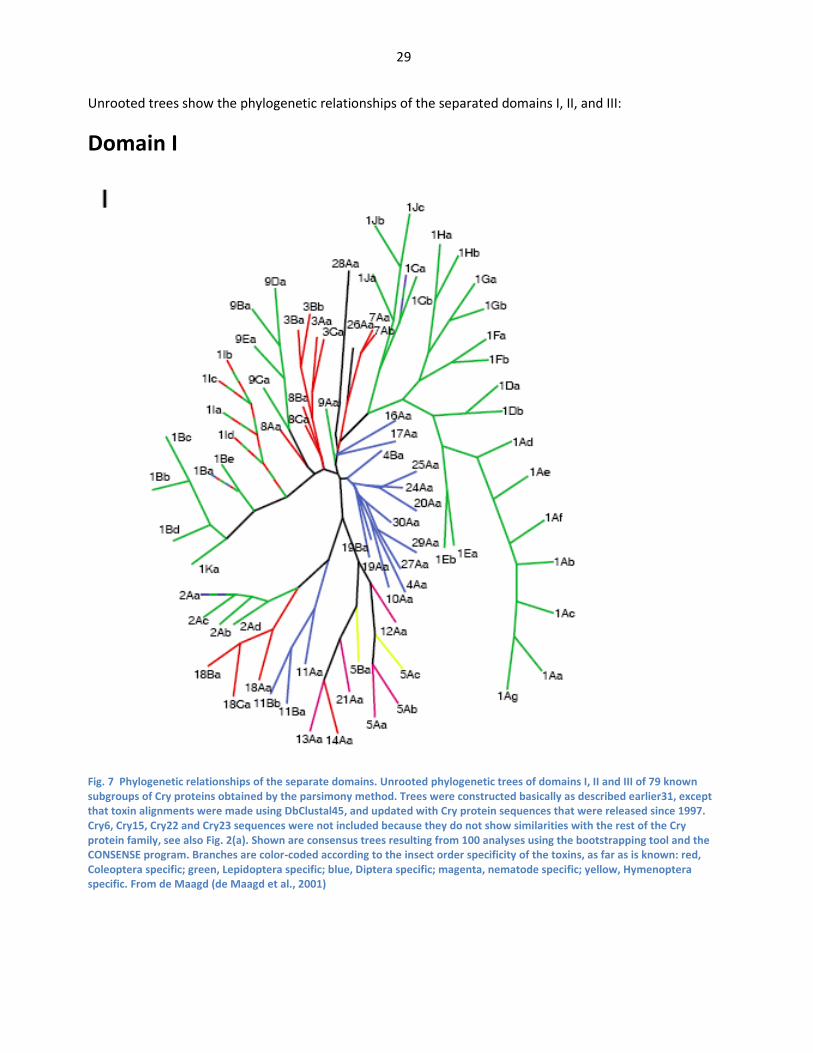
29
Unrooted trees show the phylogenetic relationships of the separated domains I, II, and III:
Domain I
Fig. 7 Phylogenetic relationships of the separate domains. Unrooted phylogenetic trees of domains I, II and III of 79 known subgroups of Cry proteins obtained by the parsimony method. Trees were constructed basically as described earlier31, except that toxin alignments were made using DbClustal45, and updated with Cry protein sequences that were released since 1997. Cry6, Cry15, Cry22 and Cry23 sequences were not included because they do not show similarities with the rest of the Cry protein family, see also Fig. 2(a). Shown are consensus trees resulting from 100 analyses using the bootstrapping tool and the CONSENSE program. Branches are color-coded according to the insect order specificity of the toxins, as far as is known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta, nematode specific; yellow, Hymenoptera specific. From de Maagd (de Maagd et al., 2001)
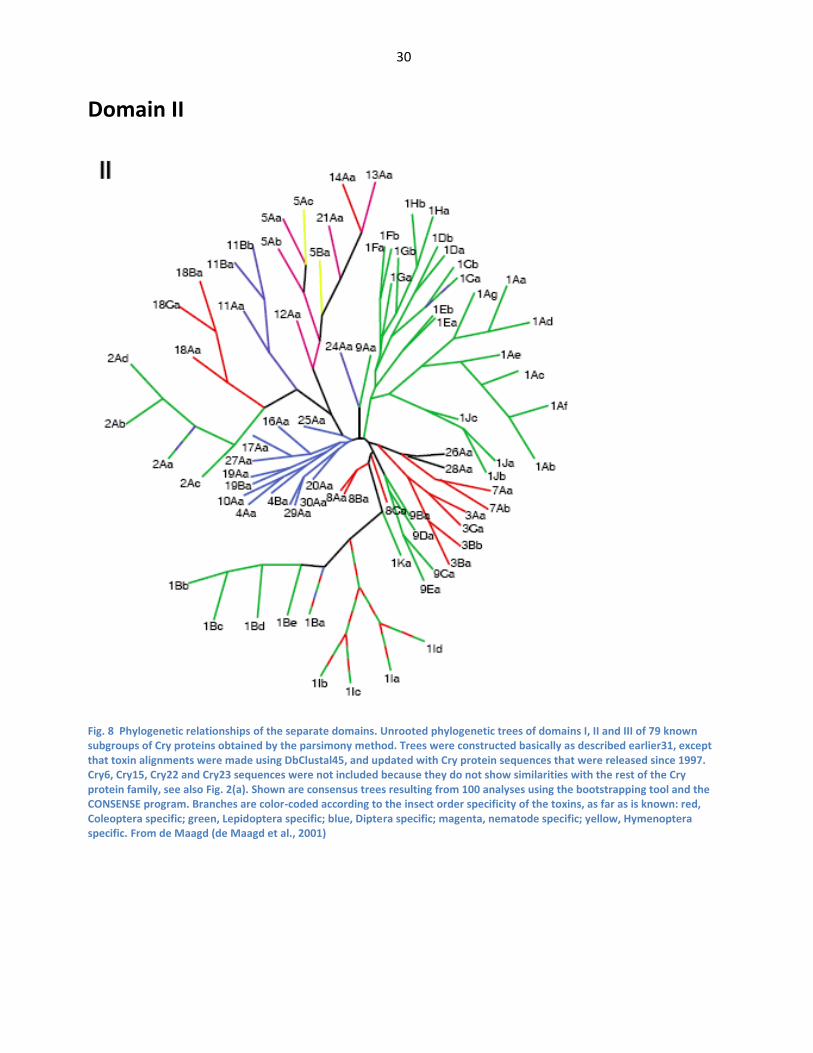
30
Domain II
Fig. 8 Phylogenetic relationships of the separate domains. Unrooted phylogenetic trees of domains I, II and III of 79 known subgroups of Cry proteins obtained by the parsimony method. Trees were constructed basically as described earlier31, except that toxin alignments were made using DbClustal45, and updated with Cry protein sequences that were released since 1997. Cry6, Cry15, Cry22 and Cry23 sequences were not included because they do not show similarities with the rest of the Cry protein family, see also Fig. 2(a). Shown are consensus trees resulting from 100 analyses using the bootstrapping tool and the CONSENSE program. Branches are color-coded according to the insect order specificity of the toxins, as far as is known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta, nematode specific; yellow, Hymenoptera specific. From de Maagd (de Maagd et al., 2001)

31
Domain III
Fig. 9 Phylogenetic relationships of the separate domains. Unrooted phylogenetic trees of domains I, II and III of 79 known subgroups of Cry proteins obtained by the parsimony method. Trees were constructed basically as described earlier31, except that toxin alignments were made using DbClustal45, and updated with Cry protein sequences that were released since 1997. Cry6, Cry15, Cry22 and Cry23 sequences were not included because they do not show similarities with the rest of the Cry protein family see also Fig. 2(a). Shown are consensus trees resulting from 100 analyses using the bootstrapping tool and the CONSENSE program. Branches are color-coded according to the insect order specificity of the toxins, as far as is known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta, nematode specific; yellow, Hymenoptera specific. From de Maagd (de Maagd et al., 2001)
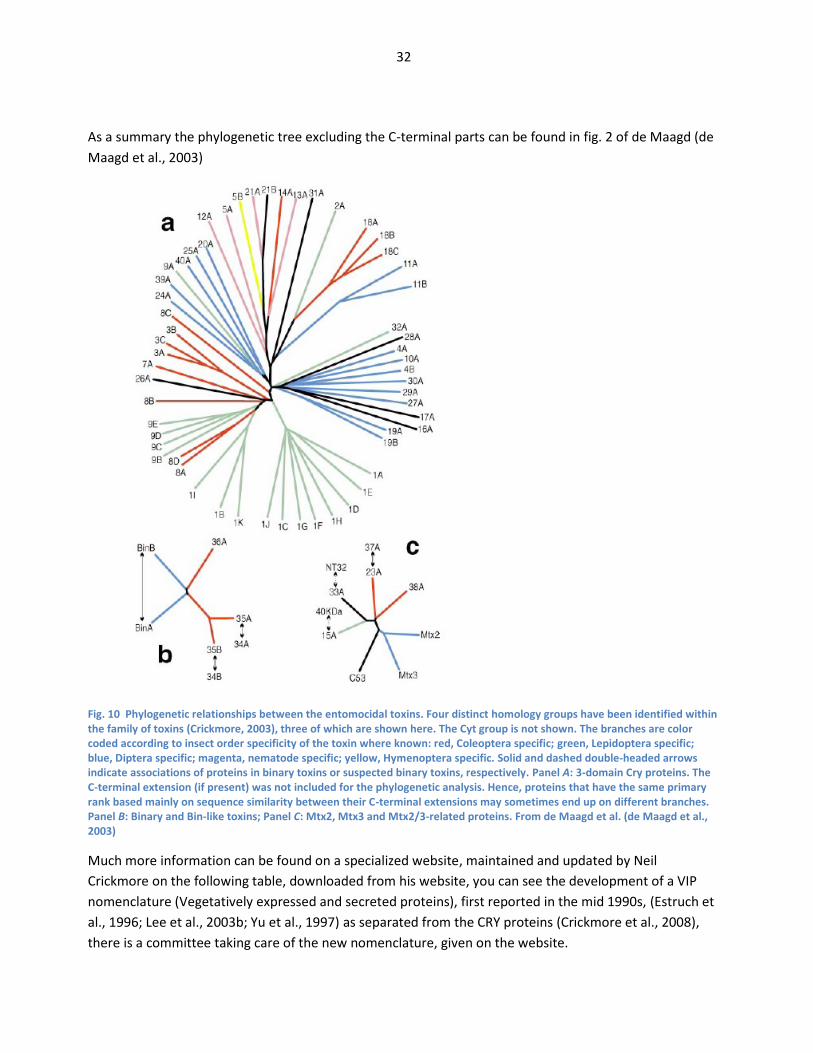
32
As a summary the phylogenetic tree excluding the C-terminal parts can be found in fig. 2 of de Maagd (de
Maagd et al., 2003)
Fig. 10 Phylogenetic relationships between the entomocidal toxins. Four distinct homology groups have been identified within the family of toxins (Crickmore, 2003), three of which are shown here. The Cyt group is not shown. The branches are color coded according to insect order specificity of the toxin where known: red, Coleoptera specific; green, Lepidoptera specific; blue, Diptera specific; magenta, nematode specific; yellow, Hymenoptera specific. Solid and dashed double-headed arrows indicate associations of proteins in binary toxins or suspected binary toxins, respectively. Panel A: 3-domain Cry proteins. The C-terminal extension (if present) was not included for the phylogenetic analysis. Hence, proteins that have the same primary rank based mainly on sequence similarity between their C-terminal extensions may sometimes end up on different branches. Panel B: Binary and Bin-like toxins; Panel C: Mtx2, Mtx3 and Mtx2/3-related proteins. From de Maagd et al. (de Maagd et al., 2003)
Much more information can be found on a specialized website, maintained and updated by Neil
Crickmore on the following table, downloaded from his website, you can see the development of a VIP
nomenclature (Vegetatively expressed and secreted proteins), first reported in the mid 1990s, (Estruch et
al., 1996; Lee et al., 2003b; Yu et al., 1997) as separated from the CRY proteins (Crickmore et al., 2008),
there is a committee taking care of the new nomenclature, given on the website.

33
Crickmore (Crickmore et al., 2008) comments on the Bt nomenclature:
http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/
“The current nomenclature, based solely on amino acid identity, allows closely related toxins to be ranked
together and removes the necessity for researchers to bioassay each new toxin against a growing series of
organisms. Biological specificity being a component of the orginal nomenclature. Roman numerals have
also been exchanged for Arabic numerals in the primary rank (eg CryIIIA became Cry3A). Each new toxin is
assigned a unique name incorporating four ranks. A completely new toxin might therefore be assigned the
name Cry50Aa1. For the sake of convenience we propose that the use of the quaternary rank (which
distinguishes between toxins that are more than 95% identical) is optional, only being used for the sake of
clarity. Note that quaternary ranks are assigned to each independently sequenced toxin gene, thus despite
the fact that some toxins have different quaternary ranks - they may in fact be identical.”
An updated list can bee downloaded at:
http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/toxins2.html
VIP proteins (Vegetative insecticidal proteins)
Since the vegetatively expressed, and secreted, VIP proteins were first reported in the mid 1990s (Estruch
et al., 1996) the rate of discovery of related toxins has increased in a similar fashion to that seen with the
Cry toxins a decade previously. Given that each discovering lab are adopting their own naming systems for
these toxins the idea was recently put forward that a nomenclature system, along the same lines used for
the Cry and Cyt toxins, be adopted. On this web page the results of applying the Bt Cry toxin
nomenclature analyses to the VIP and VIP-related toxins are presented. The following table shows the
proposed names for these toxins that result from such an analyses. A dendrogram showing the
relationship between each toxin can be found under
http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/viptree.pdf
The mode of action of Vip3A differs from that of Cry1Ab endotoxin
Vegetative insecticidal proteins (VIPs), produced during the vegetative stage of their growth in Bacillus
thuringiensis, are a group of insecticidal proteins and represent the second generation of insecticidal
trans-genes that will complement the novel δ-endotoxins in future (Wu et al., 2007).

34
1.1.5. Mode of action of Bt proteins and selectivity of the toxins
If we want to understand the full advantages of the Bt insect tolerance strategy, we will have to analyze
some of the mechanisms on how the Bt toxins work:
The mode of action of the Bt toxins on a restricted set of insect species is complex and varies from case to
case. Most studies on toxicity mechanisms are only available since a few years (Gomez et al., 2007; Heckel
et al., 2007; Soberon et al., 2007)
(Broderick et al., 2006). It is interesting to note that Bacillus thuringiensis requires enterobacteria in the
midgut of the target insect to unfold its insecticidal activity described already in detail by (Crickmore et
al., 1998; Schnepf et al., 1998). The extremely targetet and restricted toxic impact on a few lepidopteran
species is the major reason for the success of the pest management with Bt proteins.
In a summary, (de Maagd, 2007) writes: the large variety of natural Cry toxins, which each are active
against only a small number of species constitute an extensive arsenal of tools for insect control in
agriculture. An example of differences in specificity is shown in a table 2 below. It has been shown for
several insect species that the midgut epithelial cells contain receptors for Bt toxins. The presence of
specific receptors on the epithelial cells of different insect larvae, together with the proteolytic processing
under alkaline conditions, determines the specificity of Bt. Differences in sensitivity of insects for a
particular Bt toxin have been explained by differences in the concentrations of receptor molecules on the
epithelial cell membranes and by differences in affinity for particular Bt toxins.
Whereas previous binding studies have been done with fluorometric analysis which is not specific enough
(Hofmann & Luthy, 1986), using labelling with iodine-125 provides more precision (Hofmann et al., 1988a)
and specifity studies made progress (Hofmann et al., 1988b)
Some B. thuringiensis δ-endotoxins active against Manduca sexta (Tobacco Hornworm) compete for
binding of 125I-labeled Bt2-toxin to M. sexta vesicles, whereas toxins active against dipteran or
coleopteran larvae do not compete. Bt2-toxin and Bt4412-toxin bind to different sites on Plasmodiophora
brassicae vesicles. Together with other studies such as (Rie et al., 1989), these data provide evidence that
binding to a specific receptor on the membrane of gut epithelial cells is an important determinant with
respect to differences in insecticidal spectrum of B. thuringiensis insecticidal crystal proteins.
Earlier papers on the basics of Bt toxins have been published by (Crickmore et al., 1998; Schnepf et al.,
1998). According to (Federici & Siegel, 2007): As far as is known, Cyt proteins do not require a protein
receptor but, instead, bind directly to the nonglycosylated lipid portion of the microvillar membrane.
Once within the membrane, they appear to aggregate, forming lipind faults that cause an osmotic
imbalance that results in cell lysis (Butko, 2003; Nevels & Butko, 2007).

35
Following again (de Maagd, 2007): When ingested by a susceptible insect larva, Bt crystals are solubilized
by the high pH in the larval midgut and release proteins mostly varying in size between 70 and 130
kiloDaltons (kDa). These so-called protoxins (which themselves are non-toxic) are subsequently processed
by insect midgut proteases via stepwise degradation into true toxins of approximately 65 kDa (Figures 1
and 2 from (de Maagd, 2007)). The efficiency by which the crystals are solubilized and processed into the
corresponding toxins depends on the pH and the proteases present in the insect midgut. These protoxin
process control factors contribute to the specificity of Bt crystal proteins in addition to the receptor-toxin
interaction. The toxin proteins bind to the brush border membrane of midgut epithelial cells. After
binding the toxins presumably form pores in the cell membranes which disrupt the semi-permeability of
the membranes, thus leading to free ion transport. As a consequence the epithelial cells will swell and
eventually lyse. This part of the mode of action is still subject of debate, as there are also reports claiming
that binding by itself sets in motion a chain of intracellular events leading to cell death, here just a few
examples of the literature on protoxin binding from the last 3 years: (Abdullah et al., 2006; Chen et al.,
2007; de Barros Moreira Beltrao & Silva-Filha, 2007; Fabrick & Tabashnik, 2007; Fiorito et al., 2007; Griko
et al., 2007; Higuchi et al., 2007; Hossain et al., 2007; Ibiza-Palacios et al., 2008; Ikanovic et al., 2007;
Jurat-Fuentes & Adang, 2007; Krishnamoorthy et al., 2007; Luo et al., 2006; Oestergaard et al., 2007;
Shimada et al., 2006; Siqueira et al., 2006; Tang et al., 2007; Xu & Wu, 2008)
Fig. 11 Mode of action of Bacillus thuringiensis crystal proteins. Fig 1 from (de Maagd, 2007)

36
Finally, disintegration of the intestinal tract and death of the larva follows. Afterwards, germination of
spores and bacterial multiplication in the moribund or dead insect larvae can occur.
For the 130 kDa crystal proteins the toxic fragment roughly comprises the N-terminal half of the protoxin
molecule, whereas the C-terminal half is involved in crystal formation (Figure 2 A).
The N terminal half of the protoxin, when encoded by a truncated gene in heterologous systems such as
Escherichia coli or plants (see below), is as active as the toxin generated after cleavage of the crystal
protein by insect midgut juices in vivo.
Some of the 70 kDa crystal protein genes (e.g. cry3A), which occur naturally, resemble these engineered
truncated genes. This observation has been important for the engineering of other organisms, both
bacteria and plants.
The three-dimensional structure of the toxic fragment of several crystal proteins have been resolved, and
explained many of the biological features of this toxin. The N-terminal fragment consists of three domains
with specific roles in the toxin action (Figure 2 B).
The first domain consists of seven α-helices and is involved in pore formation. The second and third
domains contain ß-sheets and are involved in receptor binding.
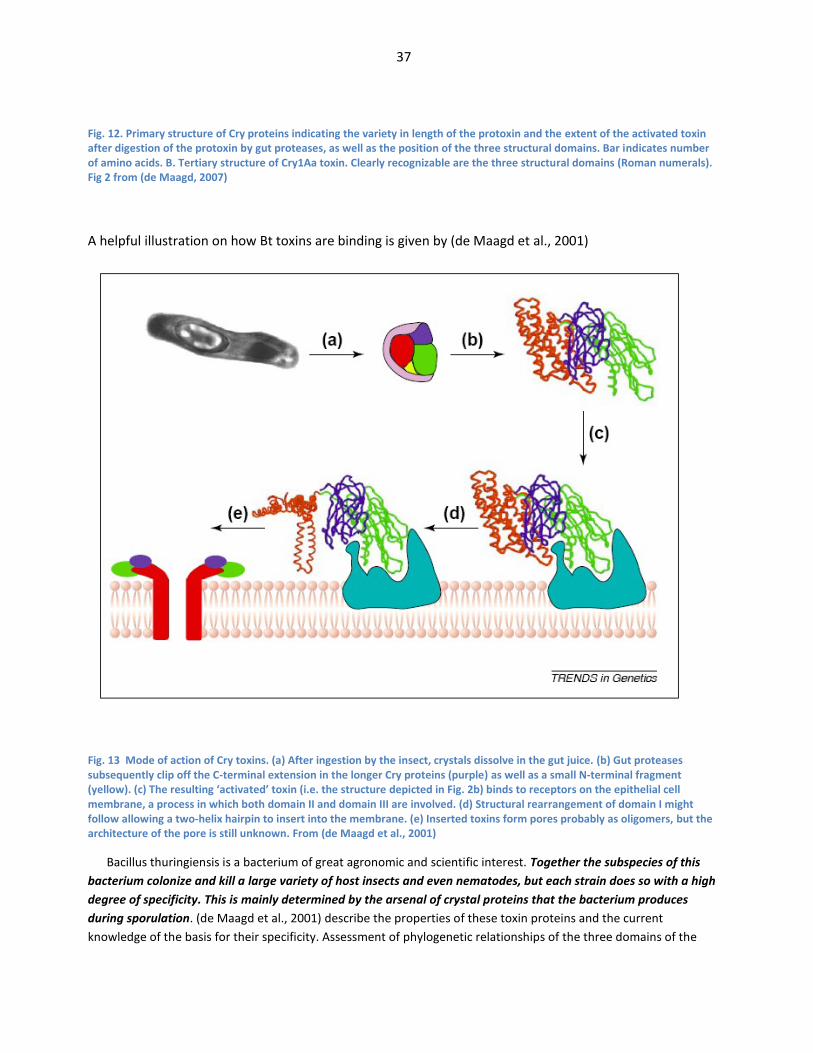
37
Fig. 12. Primary structure of Cry proteins indicating the variety in length of the protoxin and the extent of the activated toxin after digestion of the protoxin by gut proteases, as well as the position of the three structural domains. Bar indicates number of amino acids. B. Tertiary structure of Cry1Aa toxin. Clearly recognizable are the three structural domains (Roman numerals). Fig 2 from (de Maagd, 2007)
A helpful illustration on how Bt toxins are binding is given by (de Maagd et al., 2001)
Fig. 13 Mode of action of Cry toxins. (a) After ingestion by the insect, crystals dissolve in the gut juice. (b) Gut proteases subsequently clip off the C-terminal extension in the longer Cry proteins (purple) as well as a small N-terminal fragment (yellow). (c) The resulting ‘activated’ toxin (i.e. the structure depicted in Fig. 2b) binds to receptors on the epithelial cell membrane, a process in which both domain II and domain III are involved. (d) Structural rearrangement of domain I might follow allowing a two-helix hairpin to insert into the membrane. (e) Inserted toxins form pores probably as oligomers, but the architecture of the pore is still unknown. From (de Maagd et al., 2001)
Bacillus thuringiensis is a bacterium of great agronomic and scientific interest. Together the subspecies of this
bacterium colonize and kill a large variety of host insects and even nematodes, but each strain does so with a high
degree of specificity. This is mainly determined by the arsenal of crystal proteins that the bacterium produces
during sporulation. (de Maagd et al., 2001) describe the properties of these toxin proteins and the current
knowledge of the basis for their specificity. Assessment of phylogenetic relationships of the three domains of the
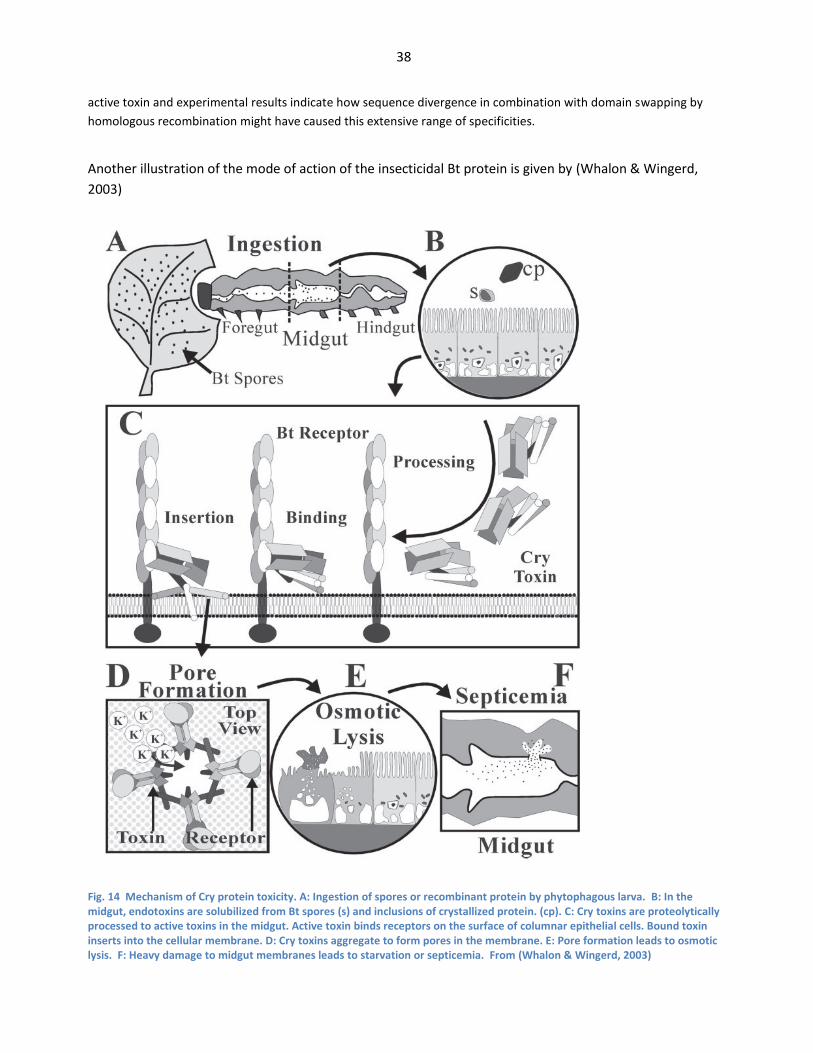
38
active toxin and experimental results indicate how sequence divergence in combination with domain swapping by
homologous recombination might have caused this extensive range of specificities.
Another illustration of the mode of action of the insecticidal Bt protein is given by (Whalon & Wingerd,
2003)
Fig. 14 Mechanism of Cry protein toxicity. A: Ingestion of spores or recombinant protein by phytophagous larva. B: In the midgut, endotoxins are solubilized from Bt spores (s) and inclusions of crystallized protein. (cp). C: Cry toxins are proteolytically processed to active toxins in the midgut. Active toxin binds receptors on the surface of columnar epithelial cells. Bound toxin inserts into the cellular membrane. D: Cry toxins aggregate to form pores in the membrane. E: Pore formation leads to osmotic lysis. F: Heavy damage to midgut membranes leads to starvation or septicemia. From (Whalon & Wingerd, 2003)

39
There is a lot more literature about the specificity of mode of action of Bt toxins existing, but this is not
the main topic of this review, and therefore a small selection of citations may be enough:
(Abdul-Rauf & Ellar, 1999; Aronson & Shai, 2001; Aronson et al., 1995; Bravo et al., 2007; de Maagd et al.,
1999; de Maagd et al., 2000; Garcia-Robles et al., 2001; Gomez et al., 2003; Gomez et al., 2007; Gonzalez-
Cabrera et al., 2006; Heckel et al., 2007; Herrero et al., 2004; Hofmann et al., 1988b; Ibiza-Palacios et al.,
2008; Jenkins & Dean, 2001; Jurat-Fuentes & Adang, 2000, 2001, 2006; Jurat-Fuentes et al., 2004;
Karumbaiah et al., 2007; Keeton & Bulla, 1997; Nakanishi et al., 2002; Oestergaard et al., 2007; Rausell et
al., 2000a; Rausell et al., 2004a; Rie et al., 1989; Soberon et al., 2007; Vanrie et al., 1989; Whalon &
Wingerd, 2003; Wu & Aronson, 1992).
1.1.6. Bt crops: present and future developments, synthetic Bt genes
1.1.6.1. Present situation in Europe
According to the latest report of Brookes (Brookes, 2008b) Only Bt 176 and MON 810 – resistant to the
Lepidopteran pests Ostrinia nubilalis European Corn Borer (ECB) and Sesamia nonagroides Mediteranean
Stem Borer (MSB) have been planted in Europe to date. Currently, only varieties of the event MON 810
are available for cultivation: 41 varieties in Spain, 6 in France, 5 in Germany and 36 have been registered
on the EU Common Variety Catalogue (status of December 2006). In total, the area planted to Bt maize in
the EU was just below a still rather modest 65,000 ha in 2006, equivalent to approximately 0.6% of total
‘EU 25’ maize plantings (including forage maize area) (James, 2007).
The pipeline is much more important in numbers and innovation, see the table 2 below.
1.1.6.2. Recent and future developments of Bt maize breeding
Some of the later developments are summarized in the following table2 below, collated by (de Maagd,
2007).
Table 2 below: Commercial Bt-crops registered by the EPA in the United States up to the year 2001, including pending registrations. An event name refers to the particular individual transformed plant from which all others originate by crossing and multiplication. Additional stacked events are not included. For a complete and updated overview, visit the Agbios GM database at http://www.agbios.com/dbase.php ECB= European corn borer (Ostrinia nubilalis), SWCB= Southwestern corn borer (Diatraea grandiosella); MCB= Mediterranean corn borer (Sesamia nonagroides); BCW=Black cutworm (Agrotis ipsilon); FAW=Fall Armyworm (Spodoptera frugiperda); CRW=Corn rootworm (Diabrotica virgifera virgifera); TBW= Tobacco budworm (Heliothis virescens), CBW=Cotton bollworm (Helicoverpa zea or H. armigera), PBW=Pink Bollworm (Pectinophora gossypiella); CPB= Colorado potato beetle (Leptinotarsa decimlineata). *These contain Bt genes for other proteins than the 3-domain toxins.
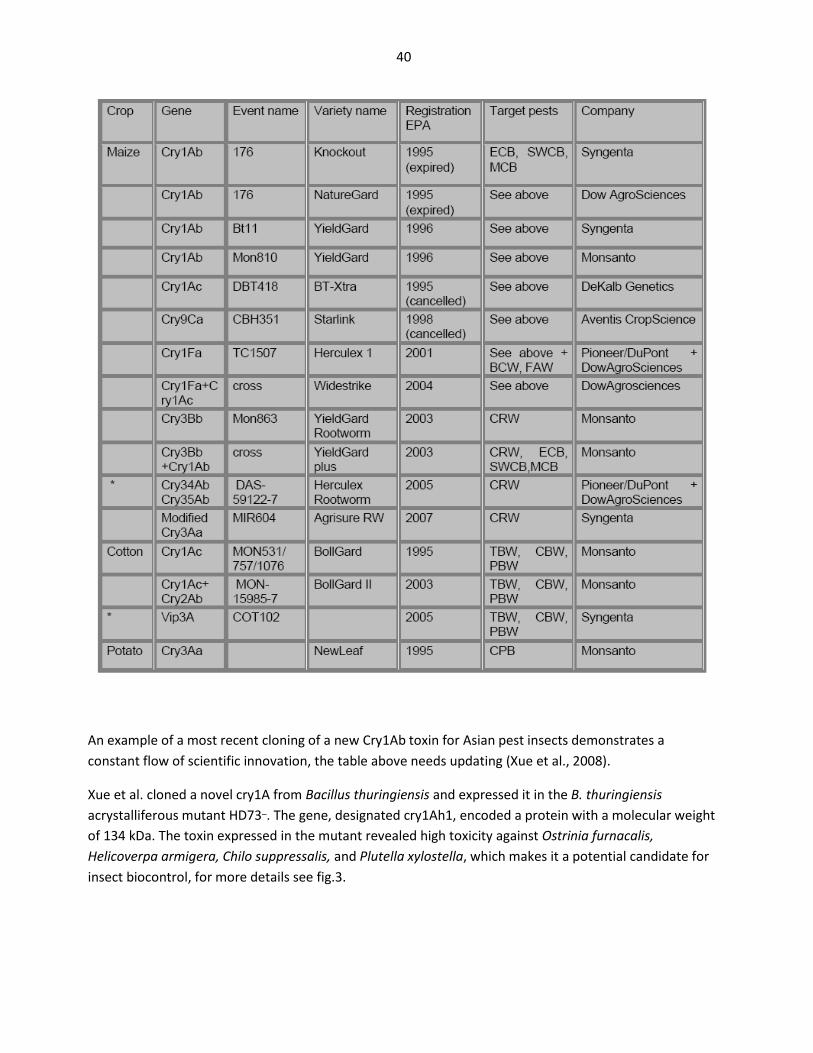
40
An example of a most recent cloning of a new Cry1Ab toxin for Asian pest insects demonstrates a
constant flow of scientific innovation, the table above needs updating (Xue et al., 2008).
Xue et al. cloned a novel cry1A from Bacillus thuringiensis and expressed it in the B. thuringiensis
acrystalliferous mutant HD73_. The gene, designated cry1Ah1, encoded a protein with a molecular weight
of 134 kDa. The toxin expressed in the mutant revealed high toxicity against Ostrinia furnacalis,
Helicoverpa armigera, Chilo suppressalis, and Plutella xylostella, which makes it a potential candidate for
insect biocontrol, for more details see fig.3.
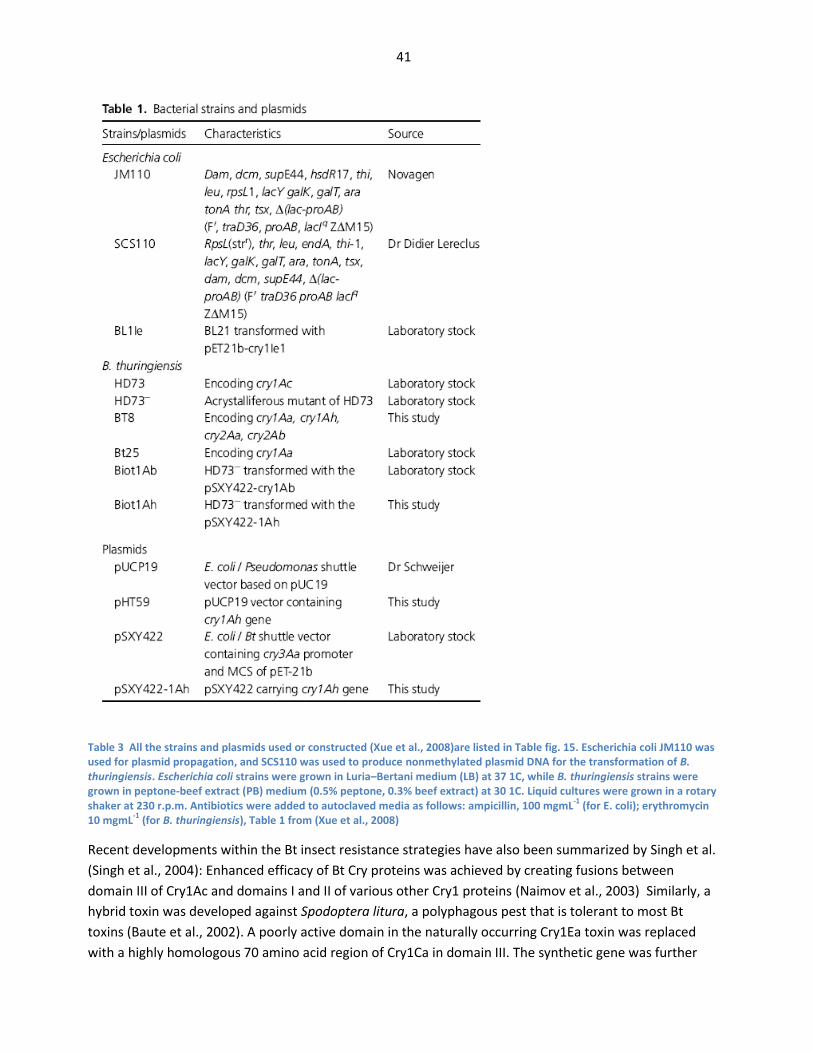
41
Table 3 All the strains and plasmids used or constructed (Xue et al., 2008)are listed in Table fig. 15. Escherichia coli JM110 was used for plasmid propagation, and SCS110 was used to produce nonmethylated plasmid DNA for the transformation of B. thuringiensis. Escherichia coli strains were grown in Luria–Bertani medium (LB) at 37 1C, while B. thuringiensis strains were grown in peptone-beef extract (PB) medium (0.5% peptone, 0.3% beef extract) at 30 1C. Liquid cultures were grown in a rotary shaker at 230 r.p.m. Antibiotics were added to autoclaved media as follows: ampicillin, 100 mgmL
-1 (for E. coli); erythromycin
10 mgmL-1
(for B. thuringiensis), Table 1 from (Xue et al., 2008)
Recent developments within the Bt insect resistance strategies have also been summarized by Singh et al.
(Singh et al., 2004): Enhanced efficacy of Bt Cry proteins was achieved by creating fusions between
domain III of Cry1Ac and domains I and II of various other Cry1 proteins (Naimov et al., 2003) Similarly, a
hybrid toxin was developed against Spodoptera litura, a polyphagous pest that is tolerant to most Bt
toxins (Baute et al., 2002). A poorly active domain in the naturally occurring Cry1Ea toxin was replaced
with a highly homologous 70 amino acid region of Cry1Ca in domain III. The synthetic gene was further

42
optimized for high-level expression in plants and was introduced into tobacco and cotton plants.
Resulting plants were found to be extremely toxic to Spodoptera litura at all stages of larval development.
A hybrid Bacillus thuringiensis gene was constructed using a synthetic and truncated cry1Ba gene as the
scaffold for inserting part of cryIIa gene encoding domain II (Baute et al., 2002). Transgenic potato plants
expressing this hybrid toxin were resistant to several insect pests, including both Coleoptera (Colorado
potato beetle) and Lepidoptera (potato tuber moth and European corn borer). As the target receptor
recognition of this hybrid protein is expected to be different from Cry proteins currently in use to control
these pests, this strategy provides new opportunities for resistance management studies involving
multiple transgenes in crops.
According to Cécile Rang et al. (Rang et al., 2004), who studied the binding and competition of five Bacillus
thuringiensis toxins - Cry1Ab, Cry1Ac, Cry1Ba, Cry1Ca, and Cry1E – it is possible to develop a tropical Bt
maize resistant to several stem borers. On Spodoptera frugiperda, Cry1Ab and Cry1Ac compete for the
same binding site; Cry1Ba and Cry1Ca compete for a second binding site. Cry1Ea recognizes a third
specific binding site in S. frugiperda and does not compete with any of the other toxins. On Diatraea
grandiosella and D. saccharalis, Cry1Ac competes with Cry1Ab and not with Cry1Ba and Cry1Ca.
Cry1Ba and Cry1Ca recognize each a specific binding site and do not compete with any of the other four
toxins. Cry1Ea does not recognize any binding site on Diatraea species. Combinations of toxins studied by
the authors are proposed to develop transgenic maize resistant to the three stemborers while allowing
resistance management. For an evolutionary view about those relationships see (Monnerat et al., 2006)
and comments in 1.1.4 below fig. 4.
A comprehensive study on future developments in insect resistant transgenic crops has been published by
Christou et al. 2006 (Christou et al., 2006). The authors concentrate on agronomically feasable traits for
the near future and distinguish (besides giving suggestions how to prevent resistance) on:
Second generation transgenic plants: transgenic plants containing, in addition to the selectable
marker, one or two transgenes encoding simple agronomic traits (such as pest and herbicide
resistance).
Third generation transgenic plants: transgenic plants that contain multiple transgenes targeting
multiple pests and diseases, often in a temporal or spatial manner. These might also express
additional value-added or agronomic traits.
The word is on targeting phloem feeding insects using the root phloem-specific promoter AAP3 (Okumoto
et al., 2004) and many other highly interesting examples, supported by an extensive list of references.
Future developments point to gene pyramiding, the combination of useful transgenes and management
systems, this is why Onishi et al (Onishi et al., 2005) propose novel multiplex polymerase chain reaction
(PCR) method for simultaneous detection of up to eight events of genetically modified (GM) maize within
a single reaction. Another possibility has been suggested by (Ammann, 2009 in press ): An appropriate mix
of seeds, each trait with a specific resistance against pest insects, in a combination adapted to local needs.

43
Another trend is emerging: The search for novel toxic proteins with selective characters which could serve
in future as novel endotoxins, the search comprises toxins of organisms with known toxins which might
show potential: Sea anemones, spiders, scorpions etc. Although there are certain communication
problems and a lot of safety research to be done first, the work has started, as manifested in the following
publications: (Bosmans & Tytgat, 2007; Gordona et al., 2007; Gruber et al., 2007; Gurevitza et al., 2007;
King, 2007; Nicholson, 2007a; Nicholson, 2007b; Rohou et al., 2007; Whetstone & Hammock, 2007).
Karlova et al. (Karlova et al., 2005) investigated the role of domain III of Bacillus thuringiensis- endotoxin
Cry1Ac in determining toxicity against Heliothis virescens. Hybrid toxins, containing domain III of Cry1Ac
with domains I and II of Cry1Ba, Cry1Ca, Cry1Da, Cry1Ea, and Cry1Fb, respectively, were created. In this
way Cry1Ca, Cry1Fb, and to a lesser extent Cry1Ba were made considerably more toxic, promising
prospects for future commercialization and resistance management of Bt crops.
An outlook into the future risks related to new technologies has been given by (Karlova et al., 2005): in
the survey, involving more than 450 researchers, which has then been discussed properly, GM crops of
the first generation play only a minor role.
In a recent paper, Gatehouse et al. (Gatehouse, 2008) outline the future of Bt crop development: Since
not all pests are adequately targeted by the Bt toxins used at present, and there is still a need to develop
solutions to specific problems, such as resistance to sap-sucking pests and pests of stored products, some
developments to the basic Bt strategy and selected alternative methods for engineering insect resistance
are outlined: Basically, it is about:
Improving expression levels of Bt toxins
Gene stacking (multiple Bt toxins)
New Bt toxins
Here only the last possibility about new Bt toxins is given as an illustrated example: the aim is to change
the nature of the Bt toxin itself, in order to change its range of toxicity:
44
Mutagenesis of the toxin-receptor, example 1:
Residues mutated in Domain II
Fig. 15 Engineering specificity in a three-domain Cry toxin; mutagenesis of the toxin-receptor interaction loop in domain II. Threedimensional structure of Cry3A (1dlc; RCSB) is shown in ribbon format. Domain I (helices) is at top right, and domain II (sheet structure) is at bottom left. Domain III (carbohydrate-binding domain; sheet structure) is behind the other domains, central in this view. Residues mutated (Wu et al., 2000) to increase toxicity toward yellow mealworm (Tenebrio molitor), Colorado potato beetle, and cottonwood leaf beetle (Chrysomela scripta) are shown in ball-and stick representation. From (Gatehouse, 2008)
One of the current commercial transgenic maize variety with resistance to corn rootworm, MON863,
expresses a modified version of the Bt Cry3Bb1 toxin (Vaughn et al., 2005). Unmodified Cry3Bb1 is active
against a number of coleopteran species, but toxicity toward Western corn rootworm was not sufficient
to give adequate protection at levels of expression achievable in maize. A large number of variants of the
native Cry3Bb, incorporating a series of specific mutations that aimed to improve the channel-forming
ability of the toxin, were produced and screened for activity (English et al., 2003). Mutations (see Fig. 11)
were carried out to: (1) increase hydrophobicity of the protein in regions in domain I containing sheets of
bound water molecules and in loop regions; (2) increase the mobility of the channel-forming helices in
domain I by disrupting hydrogen-bond formation; (3) increase the mobility and flexibility of loop regions

45
in domain I; (4) alter potential ion-pair interactions and metal-binding sites; and (5) reduce or eliminate
binding to carbohydrates in the insect gut by mutation of a loop region between domains I and II. The
toxicity of the protein towards the corn rootworm was increased approximately 8-fold, giving a product
that could be expressed at levels sufficient for adequate protection against rootworm.
Mutagenesis of the toxin-receptor, example 1:
Residues mutated in Domain I and in region linking Domain I and II.
Fig. 16 Engineering specificity in a three-domain Cry toxin; mutagenesis to improve channel-forming ability. Three-dimensional structure of Cry3Bb (1ji6; RCSB) is shown in ribbon format in the same view as Figure 1. Residues mutated (English et al., 2003) to increase toxicity toward corn rootworm are shown in ball-and-stick representation. Mutations are made in helices of domain I and in the region linking domains I and II. The mutation sites shown are taken from the most active toxin produced; a range of other sites for mutation were explored. From (Gatehouse, 2008)

46
1.2. Fundamental statements about risk assessment,
particularly related to Bt maize.
1.2.1.The situation in risk and benefit assessment of Bt maize worldwide and in Europe
The Bt maize approvals of EPA have been contested by Greenpeace, but successfully rebutted by EPA, and
factually Greenpeace has withdrawn the petition subsequently, since chances were minimal to be
successful: Scientific arguments against the petition were overwhelming, for the full documentation see
Fox (Fox, 2000), with additional comments (Fox, 1995, 1997, 2003a, b), further comments under USEPA
(US Environmental Protection Agency (USEPA), 2001). In a reaction to two papers on possible hazards for
non-target arthropods (US Environmental Protection Agency (USEPA), 1995) subsequent field studies (all
cited in the chapter on the monarch butterfly), the EPA has released an extensive documentation by
stating that Bt crops are safe for commercialization (Wilson et al., 2005), thus confirming that the earlier
risk assessment (Burkness et al., 2002) of the same agency was correct.
The subsequent reality in politics and regulation looks less positive, the transatlantic rift is becoming over
the years more and more conspicuous. It is mainly based on a fundamental difference between the US
and Europe in regulation. Whereas the US insists on an assessment of the risks of GM crops in reality,
compared to all other crops, Europe concentrates – without giving proper scientific reasoning, on the
view that GM crops need separate and special attention, this will be discussed in the chapters on
environmental risk assessment, here just a few publications explaining this rift: (Bonny, 2003; Hodgson,
2008; Miller, 1994a, b; Miller, 2007; Nature-Editorial, 1992, 2007; Ramjoue, 2007a, b)
here just the controversy in some short impressions about the latest decision of Ilse Aigner, minister of
agriculture of Germany: A purely political negative decision related to the approval of Bt maize is
camouflaged with so called factual arguments. The author just got the official list of scientific publications
which are the fundament of the negative decision. ALL paper which were presented are questionable, of
low quality in expermimental planning and statistical interpretation. Since the political debate is still
ongoing, the remarks here may suffice. It should be made clear, that the Web of Science offers with a
short search related to keywords as MON810 and MON863 reveals more than two hundred publications
in scientific journals, not a single one with clear cut negative results which could provoke a negative
regulatory decision.
There are a lot of publications available, a short search in the Web of Science has revealed over 200
publications related to the two keywords MON81 and MON863.
http://www.botanischergarten.ch/Bt-Bibliography/Bibliography-MON810-MON863-WOS-20090416.pdf
1.2.2 The agricultural reality:
A recent survey by (Gatehouse et al., 2002) in one of the main cultivation areas of the American Midwest
produces a very positive picture of many years of agronomic practice. Pilcher et al. (Pilcher et al., 2002)
also demonstrated in the same region, that besides clear cut and persistent benefits in economy and

47
environmental impact, there is also a certain consistency in performance, regardless of the pest complex,
pest density, or geographic location in the Midwestern US. Further positive performance is also
summarized in numerous surveys and comments (Ammann, 2005; Betz et al., 2000; Chapman & Burke,
2006; Conner et al., 2003; Dale, 2002; de Maagd, 2002; Gray, 2004; Nap et al., 2003; Navon, 2000;
Noteborn et al., 1995; Obrycki et al., 2001; Romeis et al., 2007; Sanvido & Romeis, 2007; Sanvido et al.,
2006; Widmer, 2007) and in (Christou et al., 2006; Manyangarirwa et al., 2006; Mehlo et al., 2005) . It will
be important to consider seriously all those positive experiences from abroad also in Europe.
Overall, there are also many earlier literature reviews confirming that the use of Bt crops can be
considered safe (Bajaj, 1994; Freeling, 1994; Gay, 1999; Peterson & Bianchi, 1999).
In a reference manual, now published by the Springer Verlag, a whole range of experienced authors give
broad insight in the history of maize breeding: (Romeis et al., 2008b).
One review from Indian ICRISAT researchers deserves special mention, insofar as Sharma and his co-
authors (Sharma et al., 2004) strive to give more critical views about the downside of GM crops resistant
to pest insects. It is a plea to develop regional breeding strategies in order to avoid future problems with
Bt resistant pest insects. Also they call for more research to adapt Bt maize to tropical climates. It should
also be mentioned that the authors cannot come up with any concrete negative effect or event, and they
show nicely the great future potential of the whole pest resistance strategy with transgenic crops.
In dozens of papers researchers have dealt with future resistance problems, and solutions are available in
the case that Bt-resistant pests will develop. A separate report will deal with the issue. Detection methods
are summarized in a recent paper (Ferre et al., 2008), who itself offers the simultaneous detection of
eight events of Bt maize. In the recently published book, (Romeis et al., 2008c) their chapter presents a
broad and comprehensive survey on the relevant questions and management of upcoming Bt resistant
pests, see the chapter 1.1.2.2. on resistant management.
1.2.3. And what about Europe ?
There are considerable differences in the culture of risk assessment between the United States and
Europe, summarized by (Drobnik, 2007). A minority of European ecologists tries to re-invent the wheel in
risk assessment of crops and asks questions related to the safety of Bt crops which have long been
answered in science with dozens of papers published in peer reviewed journals and also in the agricultural
reality of the American Midwest. In addition, some ecologists apply the wrong principles to the ecological
risk assessment of GM crops: Used to work in pristine or slightly disturbed natural habitats to assess the
ecological science of ecosystems and food webs, they hardly realize that dealing with agriculture means to
cope up to a totally different dynamics of agricultural ecology with all its rapid changes and crop rotation
and succession from year to year (Ammann et al., 2004).
Delayed start of GM crop cultivation in Europe
In Europe the start of GM crop cultivation has been delayed also for other reasons, mainly political ones, a
recent account about the basically still negative regulatory situation in Europe has been given by
(Eurobarometer, 2006): Its time to relax on GM crop regulation, particularly on the first generation of GM

48
insect resistant crops: There is enough experience gained during 10 years of GM crop applications to
seriously evaluate the ratio of risk to benefit and reduce the existing regulation in Europe – but it is still
difficult to evaluate benefit and risk when GM crops are not used. The precautionary approach (= legal
name, not precautionary principle) is applied exclusively to GM crops, never or rarely to alternative
solutions of, e.g., pest biocontrol.
Europe risks in a dramatic way to lag behind, with serious consequences in agricultural production, trade
and welfare overall. The question is, whether it is still possible to avoid serious negative effects stemming
from this overall restrictive agricultural policy related to technological progress.
As early as 1992 many US authorities asked seriously whether there is enough reason to single out the
transformed crops: (Eurobarometer Special, 2008). The time will soon come when we will have to rethink
the process oriented regulation of the first generation GM crops which are rapidly spreading all over the
world.
As we will see below (2.2.), the biosafety research situation regarding to peer reviewed publications has
dramatically improved in the last few years – in particular with Bt crops – its time for a re-assessment. It is
also time to delve into future risk posed by biotechnology and other new technologies (see 1.1.4.)
The Eurobarometer 2005 shows how propaganda inseminates public opinion with shameful nonsense (Li
et al., 2004). However the situation in 2007 looks slightly better (Demont & Tollens, 2004; Farinos et al.,
2004; Lorch & Then, 2007), the risks coming from GM crops rank considerably below some of the other
environmental risks. Voices asking for change of the “old” politics of rejecting, based on claims of negative
consumer attitude, not on science, are getting stronger, they come from the parties within the European
Parliament, British ACRE, EuropaBio, the European Federation of Biotechnology, and from Commission,
scientists and other European and non-European sources, see also (Afolabi et al., 2007; Bohorova et al.,
1999; de Groote et al., 2004; Gouse et al., 2005; Gressel et al., 2004; McGeoch & Pringle, 2005; Thomson,
2002).
In Europe, particularly in Spain, the trends are recently more promising and this will continue in the same
manner (Sutherland et al., 2008). In Africa the prospects look positive as well, and as soon as regulatory
problems are solved outside South Africa, proliferation of Bt maize will find only a few obstacles (Christou
et al., 2006).
It is also time to throw a more detailed look at the difference of the risk assessment and risk perception
cultures between Europe and the Americas: A comparative case study of genetically modified corn in the
United States of America and European Union done by (Guehlstorf & Hallstrom, 2005) reveals some hard
facts: The way governments regulate modern biotechnology is not necessarily a reflection of how their
political culture perceives the new scientific technology, it is rather how their existing regulatory structure
can create a political culture of acceptance or rejection for contested technological advancements. This
comparative study casts doubt on interpreting agricultural biotechnology decisions solely on equations of
risk analysis, and offers a detailed cultural analysis of the regulatory differentiation of modern agricultural
policy between America and Europe. It is in this context laudable that the European Commission wants to

49
put more weight on the scientific risk assessment process, trying to clearly separate science and politics –
indeed a difficult process.
Apart from such subtle and difficult transatlantic thought it is important to realize, that on a global level,
the ongoing debate in science on GM crops suffers from a lack of awareness about the basic strategic
approaches in risk assessment as we will see below (1.2.)
1.2.3. There are three major reasons, why we should develop a critical view, strictly based on
scientific data, when we judge risk assessment research and the resulting scientific
publications related to Bt crops.
1.2.3.1..Assessment of the impact of GM crops should also take into account the benefits and
balance it against the risks.
Article 19 of the CBD http://www.cbd.int/convention/articles.shtml?a=cbd-19 which is the basis of all
biosafety assessment activity and which is legally also the root article in the CBD on which the Cartagena
Protocol is building upon, makes it crystal clear that benefits of biotechnology should be taken into
account – this is already obvious from the title of Article 19: Handling of Biotechnology and Distribution of
its Benefits. Unfortunately, the negotiations of the Cartagena Protocol and the drafted and later agreed
articles concentrate nearly exclusively on the risk side, thus clearly not following the original task given by
Article 19 of the CBD. Many opponents of the GM crop technology now call upon this fact and argue, that
in the negotiated part of the Cartagena Protocol there is no explicit mention of benefits. But this is a truly
tautological argument, since it is obvious that the negotiations did not follow the legal task given by the
CBD in Article 19.
Conclusion: there is no legal basis to exclude the benefits in the risk assessment prescribed by the
Cartagena Protocol and to balance it out with the risks of the new technology.
A typical example on how economic benefits can be linked directly to the damage of the European corn
borer has been given by:
Baute, T. S., M. K. Sears and A. W. Schaafsma (2002). "Use of transgenic Bacillus thuringiensis Berliner corn
hybrids to determine the direct economic impact of the, European corn borer (Lepidoptera : Crambidae) on field
corn in eastern Canada." Journal of Economic Entomology 95(1): 57-64.<Go to ISI>://WOS:000178371100008 AND
http://www.botanischergarten.ch/Bt/Baute-Direct-Economic-2002.pdf
“Transgenic corn expressing Bacillus thuringiensis Berliner (Bt corn) (Maximizer and Yieldgard hybrids, Novartis
Seeds), non-Bt isolines and high-performance.(check) hybrids were evaluated for European corn borer, Ostrinia
nubilalis. (Hubner), damage and grain yield in commercial strip plots across Ontario in 1996 and 1997. Bt corn
hybrids reduced stalk tunneling damage by 88-100%. In 1996, minimal damage was found in locations where only
one generation of European corn borer occurred per year. Bt corn proved its greatest potential for reducing the

50
number and length of, cavities below the primary ear in locations where two generations of European corn borer
were present. A yield response to using Bt hybrids only occurred when levels of tunneling damage exceeded 6 cm in
length. European corn borer infestations resulted in a 6 and 2.4% reduction in yield for 1996 and 1997, respectively,
when Bt hybrids were compared with their non-Bt isolines. A linear relationship was found between tunnel length per
plant in centimeters (x) and yield protection (%) obtained from using Bt corn (y) (y = 1.02 + 0.005x, r(2) = 0.7217). At a
premium of $34.58 Canadian (CDN) per hectare for Bt corn seed, an infestation of at least 6 ern of corn borer
tunneling per plant was required to break even at a market price for corn of $2.50 per bushel CDN. During the period
of study, low infestations (0-2 cm) of European corn borer occurred at 25% of the locations assessed, moderate
infestations (4-6 cm) occurred at 42% of the locations, and high infestations (>6 cm) occurred at 33% of the locations.
At a corn price of $3.00 per bushel CDN and seed premiums of $34.58 per hectare CDN, 5 ern of tunneling was
required for a return on investment in Bt seed, comprising only 55% of the growers in the study. With infestations of
more than 6 cm of tunneling occurring only 33% of the time, a return on seed investment would be realized in only
one of three growing seasons. At a seed premium of $24.70 per hectare CDN per year, at least $74 per hectare CDN
in the year of infestation would be required to make up for the two years of no. return. In this study, a $74 per
hectare CDN return at a corn price of $9.26 per hectare CDN with >16 cm of tunneling damage would have occurred
only 7.3% of the time”. (Baute et al., 2002)
It is beyond the scope of this report to add an extensive chapter on the benefits of Bt crops, there is only
room for one of the latest and most comprehensive studies which has been recently updated by Brookes:
(Brookes, 2007, 2008a; Brookes, 2008b).
1.2.3.2.Assessment of the impact of GM crops should also compare to non GM crops
An early and insightful comment has been given by Pimentel and Raven 2000: (Pimentel & Raven, 2000),
here the conclusions:
Pimentel, D. S. and P. H. Raven (2000). "Bt corn pollen impacts on nontarget Lepidoptera: Assessment of
effects in nature." Proceedings of the National Academy of Sciences of the United States of America 97(15): 8198-
8199.<Go to ISI>://WOS:000088273900003 AND http://www.botanischergarten.ch/Bt/Pimentel-Impacts-nontarget-
Lepidoptera-2000.pdf
In conclusion, the introduction of resistance factors in corn through the application of genetic engineering
technology is an effective strategy to reduce the extensive crop damage caused by corn rootworm and European
corn borer populations. Yet, the diverse reactions of the different Lepidoptera to corn genotypes with varying levels
of Bt endotoxin in their pollen signal that additional testing and development is required to limit or entirely prevent
damage to nontarget butterfly and other beneficial insect species.
Although Bt corn pollen under certain circumstances has the potential of adversely affecting the population levels of
Monarch butterflies and other nontarget Lepidoptera, we consider these impacts to be minimal when compared
with habitat loss and the widespread use of pesticides throughout the ecosystem. Broad agricultural investigations
should focus on improving pest-management strategies in the context of sustainability and productivity to reduce
the use of pesticides and make agriculture more environmentally and economically sound. In addition, broadly
based research should continue to be conducted on the reasons for the decline of populations of Monarch
butterflies and other species, and remedial steps should be directed to the more important of these factors as they
are identified (Pimentel & Raven, 2000)

51
It is a scientific imperative that risk assessment studies with GM crops should compare to non-GM crops,
including the agronomic practice linked to the traits. This is what it is all about in agriculture: Innovation in
crop breeding and agricultural management strives to improve production step by step, as it has always
been done, it would be ahistoric to overlook this aspect. But this includes also the view that improvement
steps cannot be judged along the scale of absolute certainty and absolute safety. This is seemingly a
modest and unquestionable principle, but strangely enough only a very few studies are meeting those
criteria in comparison and judging improvement.
An often neglected principle should be to compare properly GM crops with non-GM crops, in order to
evaluate natural variation in both cases. Only by doing this you can get a proper balance and judgement
about any kind of risk related to GM crops. In a well planned study this has been done by (Turlings et al.,
2005)
Turlings, T. C. J., P. M. Jeanbourquin, M. Held and T. Degen (2005). "Evaluating the induced-odour emission of a Bt
maize and its attractiveness to parasitic wasps." Transgenic Research 14(6): 807-816.<Go to ISI>://000233628500004
AND http://www.botanischergarten.ch/Bt/Turlings-Evaluating-Induced-Odor-2005.pdf
“The current discussion on the safety of transgenic crops includes their effects on beneficial insects, such as
parasitoids and predators of pest insects. One important plant trait to consider in this context is the emission of
volatiles in response to herbivory. Natural enemies use the odours that result from these emissions as cues to locate
their herbivorous prey and any significant change in these plant-provided signals may disrupt their search efficiency.
There is a need for practical and reliable methods to evaluate transgenic crops for this and other important plant
traits. Moreover, it is imperative that such evaluations are done in the context of variability for these traits among
conventional genotypes of a crop. For maize and the induction of volatile emissions by caterpillar feeding this
variability is known and realistic comparisons can therefore be made. Here we used a six-arm olfactometer that
permits the simultaneous collection of volatiles emitted by multiple plants and testing of their attractiveness to
insects. With this apparatus we measured the induced odour emissions of Bt maize (Bt11, N4640Bt) and its near-
isogenic line (N4640) and the attractiveness of these odours to Cotesia marginiventris and Microplitis rufiventris, two
important larval parasitoids of common lepidopteran pests. Both parasitoid species were strongly attracted to
induced maize odour and neither wasp distinguished between the odours of the transgenic and the isogenic line. Also
wasps that had previously experienced one of the odors during a successful oviposition divided their choices equally
between the two odours. However, chemical analyses of collected odours revealed significant quantitative
differences. The same 11 compounds dominated the blends of both genotypes, but the isogenic line released a
larger amount of most of these. These differences may be due to altered resource allocation in the transgenic line,
but it had no measurable effect on the wasps' behaviour. All compounds identified here had been previously
reported for maize and the differential quantities in which they were released fall well within the range of
variability observed for other maize genotype”s. (Turlings et al., 2005)
“Some GEOs are expected to provide environmental benefits, as outlined in Box 1. However, in this report, we focus
more on potential environmental risks of GEOs than on their benefits for two reasons: risks are a more immediate
concern for ecologists, regulatory agencies, and the public, and many environmental benefits have yet to be
developed or rigorously documented.”

52
The role of ecologists is not only to point (rightly so) to potential and real risks of using GMOs, it is also
likewise important to balance the risks out against the benefits – since otherwise we are just barring
progress from agriculture – and in times of growing famine and environmental pressure we have no
reason to stop such century old procedures from creating stepwise agricultural innovation. Logically we
should also evaluate the risk of delaying the cultivation of GM crops in Europe. But there is more to say
about the complexity of the task in the next paragraph:
1.2.3.3.Dealing with complex structures of parameters in risk assessment calls for scientific
scrutiny done by experts
The way risk assessment is handled in the United States is less controversial than in Europe, because it is
done by strong agencies with good funding and an old scientific tradition (EPA, FDA etc.). Risk assessment
gets ample support from independent governmental scientific bodies, and it is also good to see that US
politicians keep to the rule that scientific judgment should first be done by the experts, later politicians
have the task to implement risk/benefit assessment decisions in society. In Europe, this rule has been
broken several times on a national and European level.
Complex structures in risk assessment parameters should be carefully scrutinized by experts who know
their science, and there is no reason, why the complex debate should deteriorate into something like a
self-service supermarket of arguments, where everybody is allowed to shop along the lines of his own
world view.
We address in a separate chapter 1.2.4. specifically the false dynamics generated by the DG environment
of the EU, who in crucial questions ignores the scientific and independent judgment of the EFSA, a
process, which should be criticized and corrected.
Even members of the German government call lately for more respect for the science on GM crops,
before major political decisions are provoked. It has to be admitted, that it is often difficult to still keep
oversight of the complexity of the debate. Some papers deal with the complex structure of agricultural
biodiversity and the resulting food webs, without coming to workable conclusions (Brooks et al., 2005a;
Perry et al., 2004).
A reasonable way forward would be to come to a consensus of pre-selected nontarget organisms for
monitoring. Recently, those thoughts have been taken up and reinforced by the now published ECOGEN
studies: (Andersen et al., 2007; Birch et al., 2007; Bohanec et al., 2007; Cortet et al., 2007; de Vaufleury et
al., 2007; Debeljak et al., 2007; Griffiths et al., 2007; Krogh & Griffiths, 2007; Krogh et al., 2007; Wesseler
et al., 2007)..These studies demonstrate how it is possible to deal in a scientific way with agricultural
complexity, without loosing a pragmatic, solution oriented vision.
It is ironic to see that ecological complexity leaves a free room for argumentation in all directions, a
complex ecosystem can thus be compared with a supermarket of arguments of all prices.

53
1.2.4.Example of a complex structure of risk assessment, an analysis related to the scientific
questions alone: The British Farm Scale Experiments
To give an well known example, even the British Farm Scale Experiments (FSE) have become subject to an
ardent debate on the outcome, after having been much praised in the beginning, and although they have
been planned over years according to highest scientific field research standards and agreed upon by all
participating parties (Perry et al., 2003). Indeed, the committee decided on a layout which seemingly
compares systematically GM-crops with non-GM-crops (herbicide tolerance). But another important rule
is neglected: The rule to adjust to the agricultural management practice of the used transgenic and non-
transgenic varieties for the field experiments. Herbicide tolerant GM crops need to be treated differently
from the non-transgenic varieties, more details on future perspectives in agricultural management in
(Hails, 2002) . So, actually, the FSE are asking the wrong questions, i.e. comparison of non-GM-crops with
GM-crops, rather they give precise answers to the impact of different kinds of herbicides (Ammann,
2005; Chassy et al., 2003). Another obvious lacune is that there are no yield data available. And on top of
this, the results are often communicated incorrectly by the scientific community: The question was,
whether biodiversity would be harmed with GM crops, and in a first publication round it was summarized,
that only GM maize demonstrated better results related to biodiversity, Beet and Canola would do worse.
And maize would do only better, because broadband pesticides were used like Atrazine in the non-GM
crop controls, which is likely to be banned soon. But it was demonstrated through additional data
analysis within the FSE that this is the wrong conclusion and that maize will still do better with other
herbicides as well (Perry et al., 2004). Later, in a more thorough data analysis, it was demonstrated, that
also GM beet does better, related to biodiversity: (May et al., 2005). The case gives evidence on how
complex environmental questions can be related to the performance of GM crops. See also a similar
study published later: (Sanvido et al., 2006). Comparable conclusions in a recent and thorough analysis of
the FSE are drawn by Bohan (Bohan et al., 2005). The question remains as to how best to manage
biodiversity at a landscape scale, but there may well be opportunities to trade-off the convenience of
some GM systems with the more intensive habitat management required to deliver specific biodiversity
targets. Ecological research has provided much information on the relationship between habitat
management and biodiversity, and it is likely that specific biodiversity targets will be reached more
efficiently by the implementation of specific measures rather than the general promotion of a less
intensive agriculture (Dollaker & Rhodes, 2007)
It also should be mentioned that there is no doubt that it is imperative to involve knowledge and
conclusions from the social sciences and philosophy to fulfil the task of modern risk assessment, but this is
not the task of this review, on the contrary, the reviewer is concerned that science arguments are more
and more pushed into the background, when they should be intricately interwoven into decision making
processes taking up the notion of the ‘Symmetry of Ignorance’ (Fischer, 2000) and making sure, that a
professional discourse is initiated in the spirit of the ‘Systems Approach’ (Ammann & Papazova Ammann,
2004).

54
1.2.6.Conclusion: Proposed procedure for future risk assessments
In this report, those basic thoughts (about scientific data and risk assessment) will be kept in mind when
we review the papers on Bt crops. The study here builds on extensive literature searches, including more
than 5800 scientific papers and major reports and books, and through intensive search work and
networking, the author of the study has downloaded ca. 1300 full text documents, all linked to the
bibliographical references, in the widespread format of Endnote Version 10. The database Endnote 10 is
appended to the report in .enl format and can be used for specific searches for keywords in various
combinations, since most references are collected over the Web of Science, which provides with
professional data structure and well chosen keywords for most publications.
There are lots of papers written about the best way of assessing the risk for non-target arthropods related
to the cultivation of Bt-crops. A proposal by (Scholte & Dicke, 2005) seems at first sight logical, namely to
start with the “most important” ecological evaluation. But knowing about the nearly infinite complexity of
most ecosystems, this would be to start a risk assessment the wrong way around.
Time will come in the Cartagena Biosafety Protocol MOP5 negotiations in Japan 2010
http://www.cop10.jp/aichi-nagoya/english/committee/index.html, when the enhancement and streamlining of
the risk assessment procedures must be discussed without prejudice. This could include ideas and
reconsiderations of various origins (Anonymous, 1992; Borch & Rasmussen, 2000; Gruere & Rosegrant,
2008).
The probably best way of handling the complex matter of risk assessment and management is suggested
by a worldwide consortium of specialists on risk assessment related to non-target arthropods , starting
with a comprehensive study in the laboratory which builds on realistic feeding and toxicity levels: (Romeis
et al., 2008a).
The consortium proposes an assessment continuum within a tiered scheme of ecological risk assessment.
The decision to reject the risk hypothesis includes consideration of remaining uncertainties. With
increasing tiers, the assessment becomes more complex and realistic, but with conclusions that are more
specific. The assessment can stop at any stage during the process as soon as sufficient information has
been compiled to address the risk hypothesis. Thus collection of data irrelevant to the risk assessment is
minimized. It is crucial to focus first, before the risk assessment starts, on a clear-cut risk hypothesis.
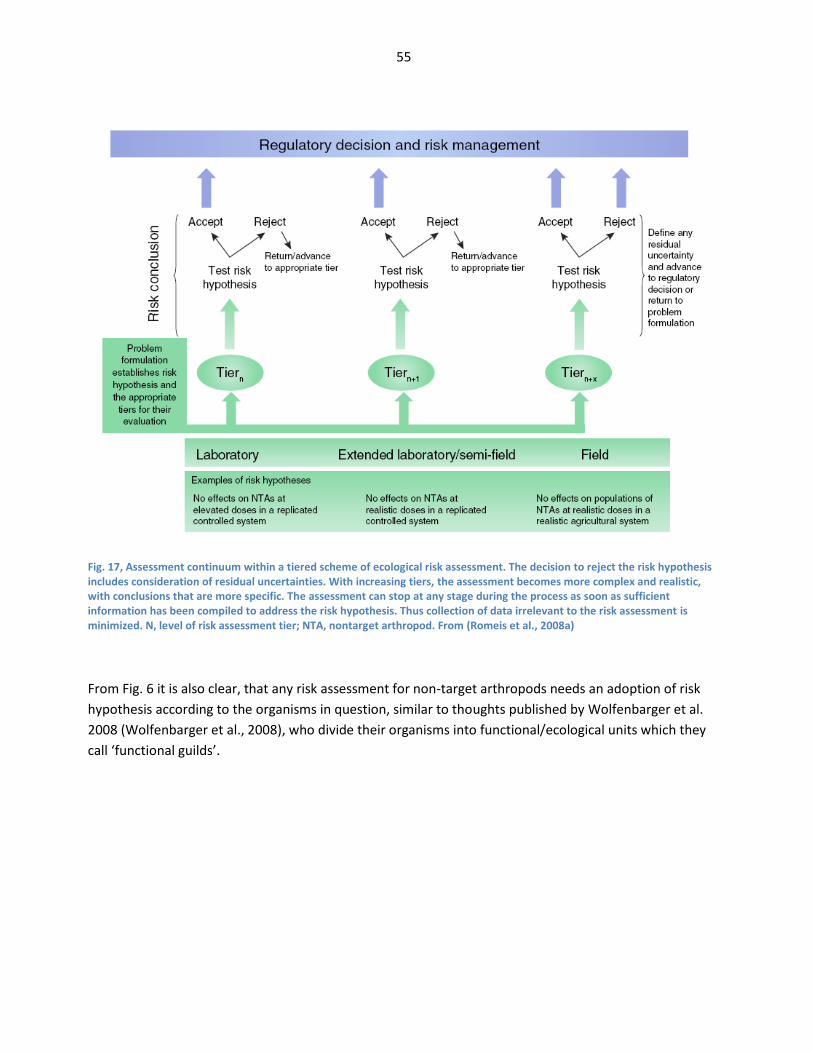
55
Fig. 17, Assessment continuum within a tiered scheme of ecological risk assessment. The decision to reject the risk hypothesis includes consideration of residual uncertainties. With increasing tiers, the assessment becomes more complex and realistic, with conclusions that are more specific. The assessment can stop at any stage during the process as soon as sufficient information has been compiled to address the risk hypothesis. Thus collection of data irrelevant to the risk assessment is minimized. N, level of risk assessment tier; NTA, nontarget arthropod. From (Romeis et al., 2008a)
From Fig. 6 it is also clear, that any risk assessment for non-target arthropods needs an adoption of risk
hypothesis according to the organisms in question, similar to thoughts published by Wolfenbarger et al.
2008 (Wolfenbarger et al., 2008), who divide their organisms into functional/ecological units which they
call ‘functional guilds’.

56
Fig. 18, Reconstruction of NTA risk assessment for Bt maize expressing Cry1Ab showing that different risk hypotheses require different types of data and synthesis at different tiers. From (US Environmental Protection Agency (USEPA), 1995) taken from (Romeis et al., 2008a) and see also the fully cited accompanying text below:
(Romeis et al., 2008a):
“In the evaluation of Bt maize expressing Cry1Ab, entities of concern included biological control organisms belonging
to, for example, the orders of Coleoptera (lady beetles), Neuroptera (lacewings) and Hymenoptera (parasitoid
wasps), as well as pollinators such as bees (also Hymenoptera), decomposers such as soil arthropods (for example,
springtails) and nontarget Lepidoptera. The problem formulation identified several risk hypotheses that were
subsequently addressed in the analytical phase of the risk assessment (Fig. 6).
Because analysis of the available precursor information revealed with sufficient certainty that the only meaningful
difference between Bt maize and its nontransformed comparators was the expression of the Cry1Ab protein, early
tier (worst-case) studies were conducted using elevated doses of purified protein or plant tissue. These studies
confirmed existing knowledge (precursor information) that these proteins are not likely to affect nonlepidopteran
insects (risk hypotheses 1–3) , . Testing could thus be terminated at this early tier.
The potential hazard to nontarget Lepidoptera (risk hypothesis 4) was recognized initially but it was concluded that
the risk is negligible . Additional studies under more realistic exposure conditions were triggered once a note and a
more comprehensive study had revealed a hazard of Cry1Ab to larvae of the monarch butterfly. Consecutive studies

57
were conducted under semi-field conditions. These studies concluded that the risk of Cry1Ab maize to monarch
populations is negligible because larval exposure to Cry1Ab toxin under field conditions is low confirming the initial
risk assessment .”
It is time to delve into future risks and benefits posed by biotechnology and leave the issues of the first
Bt crop generation behind, it would be simply a waste of time and money to continue with extensive
biosafety research related to the widespread and vastly commercialized crops, which will in most cases
deal with very interesting agro-ecological questions, but they are not anymore relevant for the risks of
agricultural production compared to conventional methods. This are questions which are situated in the
realms of the “nice-to-knows”.
1.2.7. Cited Literature
Abdul-Rauf, M. & Ellar, D.J. (1999)
Mutations of loop 2 and loop 3 residues in domain II of Bacillus thuringiensis Cry1C delta-endotoxin affect insecticidal specificity and initial binding to Spodoptera littoralis and Aedes aegypti midgut membranes. Current Microbiology, 39, 2, pp 94-98
<Go to ISI>://WOS:000081486100008 Abdullah, M.A.F., Valaitis, A.P., & Dean, D.H. (2006)
Identification of a Bacillus thuringiensis Cry11Ba toxin-binding aminopeptidase from the mosquito, Anopheles quadrimaculatus. BMC Biochemistry, 7, pp
<Go to ISI>://BIOSIS:PREV200700260492 Afolabi, C.G., Ojiambo, P.S., Ekpo, E.J.A., Menkir, A., & Bandyopadhyay, R. (2007)
Evaluation of maize inbred lines for resistance to Fusarium ear rot and fumonisin accumulation in grain in tropical Africa. Plant Disease, 91, 3, pp 279-286
<Go to ISI>://000244263500009 AND http://www.botanischergarten.ch/Bt/Alfolabi-Resistance-Fumonisin-2007.pdf
Ali, M.I., Luttrell, R.G., & Young, S.Y. (2006)
Susceptibilities of Helicoverpa zea and Heliothis virescens (Lepidoptera : Noctuidae) populations to Cry1Ac insecticidal protein. Journal of Economic Entomology, 99, 1, pp 164-175
<Go to ISI>://WOS:000235342200022 AND http://www.botanischergarten.ch/Bt/Ali-Susceptibilities-2006.pdf Ammann, K. (2005)
Effects of biotechnology on biodiversity: herbicide-tolerant and insect-resistant GM crops. Trends in Biotechnology, 23, 8, pp 388-394
<Go to ISI>://000231342700005 and http://www.sciencedirect.com/science/article/B6TCW-4GG2HJM-2/2/b23d0cc8c6846b9f6625162f3351b0ae and http://www.botanischergarten.ch/TIBTECH/Ammann-TIBTECH-Biodiversity-2005.pdf
Ammann, K. (2009 in press )
Why farming with high tech methods should integrate elements of organic agriculture New Biotechnology, 4, pp
http://www.botanischergarten.ch/NewBiotech/Integrated-Farming-Biotech-Org-20090410.pdf

58
Ammann, K. & Papazova Ammann, B. (2004) Factors Influencing Public Policy Development in Agricultural Biotechnology. In Risk Assessment of
Transgenic Crops (ed S. Shantaram), Vol. 9, pp. 1552. Wiley and Sons, Hoboken, NJ, USA.P. Christou & H. Klee Handbook of Plant Biotechnology
http://www.botanischergarten.ch/Wiley/Factors-Discourse-Wiley.pdf Ammann, K.i., Wolfenbarger, L., Andow DA. and Hilbeck, A., Nickson, T., Wu, F., Thompson, B., & Ammann, K. (2004)
Electronic Source: Biosafety in agriculture: is it justified to compare directly with natural habitats ? (ed ESA Ecological Society of America), Frontiers in Ecology, Forum: GM crops: balancing predictions of promise and peril
published by: Ecological Society of America www.frontiersinecology.org and http://www.botanischergarten.ch/Frontiers-Ecology/Ammann-Forum-def1.pdf Andersen, M.N., Sausse, C., Lacroix, B., Caul, S., & Messean, A. (2007)
Agricultural studies of GM maize and the field experimental infrastructure of ECOGEN. Pedobiologia, 51, 3, pp 175-184
<Go to ISI>://000249273500002 AND http://www.botanischergarten.ch/Bt/Andersen-Agricutural-Studies-ECOGEN-2007.pdf
Anonymous (1992)
Products pose no special risks just because of the processes used to make them. Nature, 356, 6364, pp 1-2 http://dx.doi.org/10.1038/356001b0 AND http://www.botanischergarten.ch/Regulation/Anonymous-US-
Regulation-Nature-1992.pdf Aronson, A.I., Beckman, W., & Dunn, P. (1986)
Bacillus-Thuringiensis and Related Insect Pathogens. Microbiological Reviews, 50, 1, pp 1-24 <Go to ISI>://WOS:A1986A289000001 AND http://www.botanischergarten.ch/Bt/Aronson-Bt-Related-Insect-
Pathogens-1986.pdf Aronson, A.I. & Shai, Y. (2001)
Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action. FEMS Microbiology Letters, 195, 1 %R doi:10.1111/j.1574-6968.2001.tb10489.x, pp 1-8
http://www.blackwell-synergy.com/doi/abs/10.1111/j.1574-6968.2001.tb10489.x AND http://www.botanischergarten.ch/Bt/Aronson-Why-Bt-Insecticidal-2001.pdf
Aronson, A.I., Wu, D., & Zhang, C.L. (1995)
Mutagenesis of Specificity and Toxicity Regions of a Bacillus-Thuringiensis Protoxin Gene. Journal of Bacteriology, 177, 14, pp 4059-4065
<Go to ISI>://A1995RH81000023 Bajaj, S. (1994)
Maize (Biotechnology in Agriculture and Forestry) (Hardcover) Springer; 1 edition (September 29, 1994) IS: ISBN-10: 354056392X ISBN-13: 978-3540563921, pp 632
Ballester, V., Granero, F., de Maagd, R.A., Bosch, D., Mensua, J.L., & Ferre, J. (1999)
Role of Bacillus thuringiensis toxin domains in toxicity and receptor binding in the diamondback moth. Applied and Environmental Microbiology, 65, 5, pp 1900-1903
<Go to ISI>://000080034500013 Barton, K.A. & Umbeck, P.F. (1989)
Expression of Bacillus-Thuringiensis Delta-Endotoxin in Commercial Crop Plants. Abstracts of Papers of the American Chemical Society, 198, pp 133-AGRO
<Go to ISI>://WOS:A1989DG64200204 Barton, K.A., Whiteley, H.R., & Yang, N.S. (1987)
Bacillus-Thuringiensis Delta-Endotoxin Expressed in Transgenic Nicotiana-Tabacum Provides Resistance to Lepidopteran Insects. Plant Physiology, 85, 4, pp 1103-1109
<Go to ISI>://WOS:A1987L542800044 AND http://www.botanischergarten.ch/Bt/Barton-Bt-Endotoxin-Nicotiana-1987.pdf
Bates, S.L., Zhao, J.Z., Roush, R.T., & Shelton, A.M. (2005)
Insect resistance management in GM crops: past, present and future. Nature Biotechnology, 23, 1, pp 57-62

59
<Go to ISI>://000226195700029 AND http://www.botanischergarten.ch/Bt/Bates-resistance-management-2000.pdf
Bauer, L.S. (1995)
Resistance - a Threat to the Insecticidal Crystal Proteins of Bacillus-Thuringiensis. Florida Entomologist, 78, 3, pp 414-443
<Go to ISI>://A1995RX01300004 AND http://www.botanischergarten.ch/Bt/Bauer-Resistance-1995.pdf AND http://www.botanischergarten.ch/Bt/Bauer-Resistance-Erratum-1995.pdf
Baumann, L., Okamoto, K., Unterman, B.M., Lynch, M.J., & Baumann, P. (1984)
Phenotypic Characterization of Bacillus-Thuringiensis and Bacillus-Cereus. Journal of Invertebrate Pathology, 44, 3, pp 329-341
<Go to ISI>://WOS:A1984TS43900013 NOT IN EBZ, NOT IN NEBIS Baute, T.S., Sears, M.K., & Schaafsma, A.W. (2002)
Use of transgenic Bacillus thuringiensis Berliner corn hybrids to determine the direct economic impact of the, European corn borer (Lepidoptera : Crambidae) on field corn in eastern Canada. Journal of Economic Entomology, 95, 1, pp 57-64
<Go to ISI>://WOS:000178371100008 AND http://www.botanischergarten.ch/Bt/Baute-Direct-Economic-2002.pdf
Beachy, R.N., Fedoroff, N.V., Goldberg, R.B., & McHughen, A. (2008)
The burden of proof: A response to Rosi-Marshall et al. Proceedings of the National Academy of Sciences, pp --
http://www.pnas.org AND http://www.botanischergarten.ch/Bt/Beachy-Rosi-Marshall-Burden-2008.pdf Beegle, C.C. & Yamamoto, T. (1992)
Invitation Paper (Cp-Alexander-Fund) - History of Bacillus-Thuringiensis Berliner Research-and-Development. Canadian Entomologist, 124, 4, pp 587-616
<Go to ISI>://WOS:A1992JK19900003 AND NEBIS 20080906 Berliner, E. (1915)
Ueber die Schlaffsucht der Ephestia kuhniella und Bac. thuringiensis n. sp. Zeitschrift Fur angewandte Entomologie, 2, pp 21-56
Bernstein, I.L., Bernstein, J.A., Miller, M., Tierzieva, S., Bernstein, D.I., Lummus, Z., Selgrade, M.K., Doerfler, D.L., & Seligy, V.L. (1999)
Immune responses in farm workers after exposure to Bacillus thuringiensis pesticides. Environmental Health Perspectives, 107, 7, pp 575-582
<Go to ISI>://000081426900028 AND http://www.botanischergarten.ch/Bt/Bernstein-Immune-Responses-1999.pdf
Bernstein, J.A., Bernstein, I.L., Bucchini, L., Goldman, L.R., Hamilton, R.G., Lehrer, S., Rubin, C., & Sampson, H.A. (2003)
Clinical and laboratory investigation of allergy to genetically modified foods. Environmental Health Perspectives, 111, 8, pp 1114-1121
<Go to ISI>://000184122200012 AND http://www.botanischergarten.ch/Bt/Bernstein-Clinical-Laboratory-2003.pdf
Betz, F.S., Hammond, B.G., & Fuchs, R.L. (2000)
Safety and advantages of Bacillus thuringiensis-protected plants to control insect pests. Regulatory Toxicology and Pharmacology, 32, 2, pp 156-173
<Go to ISI>://000165626300003 AND http://www.botanischergarten.ch/Bt/Betz-Safety-Advantages-2000.pdf Birch, A.N.E., Griffiths, B., Caul, S., Thompson, J., Heckmann, L.H., Krogh, P.H., & Cortet, J. (2007)
The role of laboratory, glasshouse and field scale experiments in understanding the interactions between genetically modified crops and soil ecosystems: A review of the ECOGEN project SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 251-260
http://www.sciencedirect.com/science/article/B7CW5-4NTHKYM-1/2/540aa6409068a9bb49d8b0b8953b9aca AND http://www.botanischergarten.ch/Bt/Birch-Laboratory-Field-ECOGEN-2007.pdf
Bishop, A.H., Johnson, C., & Perani, M. (1999)

60
The safety of Bacillus thuringiensis to mammals investigated by oral and subcutaneous dosage. World Journal of Microbiology & Biotechnology, 15, 3, pp 375-380
<Go to ISI>://000082676300014 AND http://www.botanischergarten.ch/Bt/Bishop-safety-Bt-Mammals-1999.pdf
Bizzarri, M.F. & Bishop, A.H. (2007)
Recovery of Bacillus thuringiensis in vegetative form from the phylloplane of clover (Trifolium hybridum) during a growing season. Journal of Invertebrate Pathology, 94, 1, pp 38-47
<Go to ISI>://WOS:000243419300005 AND http://www.botanischergarten.ch/Bt/Bizzarri-Recovery-Phylloplane-2007.pdf
Bohan, D.A., Boffey, C.W.H., Brooks, D.R., Clark, S.J., Dewar, A.M., Firbank, L.G., Haughton, A.J., Hawes, C., Heard, M.S., May, M.J., Osborne, J.L., Perry, J.N., Rothery, P., Roy, D.B., Scott, R.J., Squire, G.R., Woiwod, I.P., & Champion, G.T. (2005)
Effects on weed and invertebrate abundance and diversity of herbicide management in genetically modified herbicide-tolerant winter-sown oilseed rape. Proceedings of the Royal Society B-Biological Sciences, 272, 1562, pp 463-474
<Go to ISI>://000228535600001 AND http://www.botanischergarten.ch/Farmscale/Bohan-Effects-Weed-2005.pdf
Bohanec, M., Cortet, J., Griffiths, B., Znidarsik, M., Debeljak, M., Caul, S., Thompson, J., & Krogh, P.H. (2007)
A qualitative multi-attribute model for assessing the impact of cropping systems on soil quality SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 239-250
http://www.sciencedirect.com/science/article/B7CW5-4NNYD85-3/2/45f91364c722057d717681d7e6af3f12 AND http://www.botanischergarten.ch/Bt/Bohanek-Qualaitative-Multiattribute-ECOGEN-2007.pdf
Bohn, M., Groh, S., Khairallah, M.M., Hoisington, D.A., Utz, H.F., & Melchinger, A.E. (2001)
Re-evaluation of the prospects of marker-assisted selection for improving insect resistance against Diatraea spp. in tropical maize by cross validation and independent validation. Theoretical and Applied Genetics, 103, 6-7, pp 1059-1067
<Go to ISI>://000172530400028 Bohn, M., Khairallah, M.M., GonzalezdeLeon, D., Hoisington, D.A., Utz, H.F., Deutsch, J.A., Jewell, D.C., Mihm, J.A., & Melchinger, A.E. (1996)
QTL mapping in tropical maize .1. Genomic regions affecting leaf feeding resistance to sugarcane borer and other traits. Crop Science, 36, 5, pp 1352-1361
<Go to ISI>://A1996VK30900045 Bohn, M., Khairallah, M.M., Jiang, C., GonzalezdeLeon, D., Hoisington, D.A., Utz, H.F., Deutsch, J.A., Jewell, D.C., Mihm, J.A., & Melchinger, A.E. (1997)
QTL mapping in tropical maize .2. Comparison of genomic regions for resistance to Diatraea spp. Crop Science, 37, 6, pp 1892-1902
<Go to ISI>://A1997YL42000038 Bohn, M., Magg, T., Klein, D., & Melchinger, A.E. (2003)
Breeding early maturing European dent maize (Zea mays L.) for improved agronomic performance and resistance against the European corn borer (Ostrinia nubilalis HB.). Maydica, 48, 3, pp 239-247
<Go to ISI>://000187456900008 AND http://www.botanischergarten.ch/Bt/Bohn-Breeding-2003.pdf Bohorova, N., Zhang, W., Julstrum, P., McLean, S., Luna, B., Brito, R.M., Diaz, L., Ramos, M.E., Estanol, P., Pacheco, M., Salgado, M., & Hoisington, D. (1999)
Production of transgenic tropical maize with cryIAb and cryIAc genes via microprojectile bombardment of immature embryos. Theoretical and Applied Genetics, 99, 3-4, pp 437-444
<Go to ISI>://000082570500008 Bohorova, N.E., Luna, B., Brito, R.M., Huerta, L.D., & Hoisington, D.A. (1995a)
Regeneration Potential of Tropical, Subtropical, Midaltitude, and Highland Maize Inbreds. Maydica, 40, 3, pp 275-281
<Go to ISI>://A1995TG29100010 Bohorova, N.E., Vanginkel, M., Rajaram, S., & Hoisington, D.A. (1995b)
Tissue-Culture Response of Cimmyt Elite Bread Wheat Cultivars and Evaluation of Regenerated Plants. Cereal Research Communications, 23, 3, pp 243-249

61
<Go to ISI>://A1995TE18200006 Bonny, S. (2003)
Why are most Europeans opposed to GMOs? Factors explaining rejection in France and Europe. Electronic Journal of Biotechnology, 6, 1, pp 50-71
<Go to ISI>://WOS:000183582900008 AND http://www.botanischergarten.ch/Discourse/Bonny-Why-Europeans-Opposed-2003.pdf
Borch, K. & Rasmussen, B. (2000)
An analytical approach to the implementation of genetically modified crops. Trends in Biotechnology, 18, 12, pp 484-486
<Go to ISI>://WOS:000166909500003 AND http://www.botanischergarten.ch/Bt/Borch-Analytical-Approach-2000.pdf
Bosmans, F. & Tytgat, J. (2007)
Sea anemone venom as a source of insecticidal peptides acting on voltage-gated Na+ channels. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 550-560
http://www.sciencedirect.com/science/article/B6TCS-4MH2991-3/2/69040cba44e3da230f60ab178f1f80ba AND http://www.botanischergarten.ch/Bt/Bosmans-Sea-Anemone-Venom-2007.pdf
Bourguet, D., Desquilbet, M., & Lemarie, S. (2005)
Regulating insect resistance management: the case of non-Bt corn refuges in the US. Journal of Environmental Management, 76, 3, pp 210-220
<Go to ISI>://000230714000003 AND http://www.botanischergarten.ch/Bt/Bourguet-Regulating-Resistance-2005.pdf
Bravo, A. (1997)
Phylogenetic relationships of Bacillus thuringiensis delta-endotoxin family proteins and their functional domains. Journal of Bacteriology, 179, 9, pp 2793-2801
<Go to ISI>://WOS:A1997WX57400001 AND http://www.botanischergarten.ch/Bt/Bravo-Phylogenetic-Relationships-Bt-1997.pdf
Bravo, A., Gill, S.S., & Soberon, M. (2007)
Mode of action of Bacillus thuringiensis Cry and Cyt toxins and their potential for insect control. Toxicon, 49, 4, pp 423-435
<Go to ISI>://000245526000002 AND http://www.botanischergarten.ch/Bt/Bravo-Mode-Action-2007.pdf Broderick, N., Robinson, C., McMahon, M., Holt, J., Handelsman, J., & Raffa, K. (2009)
Contributions of gut bacteria to Bacillus thuringiensis-induced mortality vary across a range of Lepidoptera. BMC Biology, 7, 1, pp 11
http://www.biomedcentral.com/1741-7007/7/11 AND http://www.botanischergarten.ch/Bt/Broderick-Contributions-Gut-Bacteria-2009.pdf
Broderick, N.A., Goodman, R.M., Handelsman, J., & Raffa, K.F. (2003)
Effect of host diet and insect source on synergy of gypsy moth (Lepidoptera : Lymantriidae) mortality to Bacillus thuringiensis subsp kurstaki by zwittermicin A. Environmental Entomology, 32, 2, pp 387-391
<Go to ISI>://WOS:000182354100019 NEBIS TO ORDER 20080325 Broderick, N.A., Goodman, R.M., Raffa, K.F., & Handelsman, J. (2000)
Synergy between zwittermicin A and Bacillus thuringiensis subsp kurstaki against gypsy moth (Lepidoptera : Lymantriidae). Environmental Entomology, 29, 1, pp 101-107
<Go to ISI>://WOS:000089623000015 Broderick, N.A., Raffa, K.F., Goodman, R.M., & Handelsman, J. (2004)
Census of the bacterial community of the gypsy moth larval midgut by using culturing and culture-independent methods. Applied and Environmental Microbiology, 70, 1, pp 293-300
<Go to ISI>://WOS:000188115300037 Broderick, N.A., Raffa, K.F., & Handelsman, J. (2006)
Midgut bacteria required for Bacillus thuringiensis insecticidal activity. Proceedings of the National Academy of Sciences of the United States of America, 103, 41, pp 15196-15199
<Go to ISI>://WOS:000241247300039 AND http://www.botanischergarten.ch/Bt/Broderick-Midgut-Bacteria-Bt-2006.pdf

62
Brookes, G. (2007)
The benefits of adopting genetically modified, insect resistant (Bt) maize in the European Union (EU): first results from 1998-2006 plantings, PG Economics Ltd pp 39 (Report)
www.pgeconomics.co.uk Brookes, G. (2008a)
The impact of using GM insect resistant maize in Europe since 1998. International Journal of Biotechnology, 10, 2/3, pp 148-166
http://www.botanischergarten.ch/Bt/Brookes-btmaizeineuropeiIJBTApr2008.pdf Brookes, G. (2008b)
The impact of using GM insect resistant maize in Europe since 1998. International Journal of Biotechnology, 10, 2-3, pp 148-166
<Go to ISI>://BIOSIS:PREV200800387918 AND http://www.botanischergarten.ch/Bt/Brookes-btmaizeineuropeiIJBTApr2008.pdf
Brookes, G. & Barfoot, P. (2008)
GM crops: global socio-economic and environmental impacts 1996-2006, PG economics Ltd. UK. pp 118 Dorchester, UK (Report)
http://www.pgeconomics.co.uk/pdf/globalimpactstudyjune2008PGEconomics.pdf Brooks, D.R., Clark, S.J., Perry, J.N., Bohan, D.A., Champion, G.T., Firbank, L.G., Haughton, A.J., Hawes, C., Heard, M.S., & Woiwod, I.P. (2005a)
Invertebrate biodiversity in maize following withdrawal of triazine herbicides. Proceedings of the Royal Society B-Biological Sciences, 272, 1571, pp 1497-1502
<Go to ISI>://000231268700011 Brooks, T.D., Williams, W.P., Windham, G.L., Willcox, M.C., & Abbas, H.K. (2005b)
Quantitative trait loci contributing resistance to aflatoxin accumulation in the maize inbred Mp313E. Crop Science, 45, 1, pp 171-174
<Go to ISI>://WOS:000226435300021 Burges, H.D. (1981)
Microbiol control of pests and plant diseases 1970-1980 Academic Press Ltd., London, IS: ISBN-10: 0121433609 ISBN-13: 978-0121433604 pp 949
Burkness, E.C., Hutchison, W.D., Weinzierl, R.A., Wedberg, J.L., Wold, S.J., & Shaw, J.T. (2002)
Efficacy and risk efficiency of sweet corn hybrids expressing a Bacillus thuringiensis toxin for Lepidopteran pest management in the Midwestern US. Crop Protection, 21, 2, pp 157-169
<Go to ISI>://000174435900009 AND http://www.botanischergarten.ch/Burkness-Consistent-Benefits-2002.pdf
Butko, P. (2003)
Cytolytic toxin Cyt1A and its mechanism of membrane damage: Data and hypotheses. Applied and Environmental Microbiology, 69, 5, pp 2415-2422
<Go to ISI>://WOS:000182808300001 Caplan, A., Herreraestrella, L., Inze, D., Vanhaute, E., Vanmontagu, M., Schell, J., & Zambryski, P. (1983)
Introduction of Genetic Material into Plant-Cells. Science, 222, 4625, pp 815-821 <Go to ISI>://WOS:A1983RQ47300022 AND http://www.botanischergarten.ch/Bt/Caplan-Introduction-Genetic-
Material-1983.pdf Carpenter, J.E. (2001)
Electronic Source: Case study in benefits and risks of agricultural biotechnology: roundup ready soybeans and Bt corn,
published by: National Center for Food and Agricultural Policy http://www.ncfap.org/reports/biotech/benefitsandrisks.pdf Carpenter, J.E., Gianessi, L.P., & Silvers, C.S. (2001)
Insecticidal Bt plants versus chemical insecticides. Abstracts of Papers of the American Chemical Society, 222, pp U69-U69
<Go to ISI>://000170690000265 Carpenter, J.E., Sankula, S., Silvers, C.S., & Gianessi, L.P. (2004)

63
Insecticidal Bacillus thurgingiensis plants versus chemical insecticides. In Agricultural Biotechnology: Challenges and Prospects, pp. 37-51. AMER CHEMICAL SOC, WASHINGTON Acs Symposium Series
866 <Go to ISI>://000189476000004 Carson, R. (1962 - 2002)
Silent Spring Houghton Mifflin Company, Boston, IS: ISBN-10: 0395683297 ISBN-13: 978-0395683293, pp 368
http://www.amazon.com/exec/obidos/ASIN/0395683297/rachelcarson/104-0439964-0595154 Challacombe, J.F., Altherr, M.R., Xie, G., Bhotika, S.S., Brown, N., Bruce, D., Campbell, C.S., Campbell, M.L., Chen, J., Chertkov, O., Cleland, C., Dimitrijevic, M., Doggett, N.A., Fawcett, J.J., Glavina, T., Goodwin, L.A., Green, L.D., Han, C.S., Hill, K.K., Hitchcock, P., Jackson, P.J., Keim, P., Kewalramani, A.R., Longmire, J., Lucas, S., Malfatti, S., Martinez, D., McMurry, K., Meincke, L.J., Misra, M., Moseman, B.L., Mundt, M., Munk, A.C., Okinaka, R.T., Parson-Quintana, B., Reilly, L.P., Richardson, P., Robinson, D.L., Saunders, E., Tapia, R., Tesmer, J.G., Thayer, N., Thompson, L.S., Tice, H., Ticknor, L.O., Wills, P.L., Gilna, P., & Brettin, T.S. (2007)
The complete genome sequence of Bacillus thuringiensis Al Hakam. Journal of Bacteriology, 189, 9, pp 3680-3681
<Go to ISI>://000246028400041 AND http://www.botanischergarten.ch/Bt/Challacombe-Genome-Sequence-Bt-2007.pdf
Chapman, M.A. & Burke, J.M. (2006)
Letting the gene out of the bottle: the population genetics of genetically modified crops. New Phytologist, 170, 3, pp 429-443
<Go to ISI>://000236738600004 AND http://www.botanischergarten.ch/Bt/Chapman-gene-outof-Bottle-2006.pdf
Chassy, B., Carter, C., McGloughlin, M., McHughen, A., Parrott, W., Preston, C., Roush, R., Shelton, A., & Strauss, S.H. (2003)
UK field-scale evaluations answer wrong questions. Nature Biotechnology, 21, 12, pp 1429-1430 <Go to ISI>://000186845200011 AND http://www.botanischergarten.ch/Farmscale/Chassy-wrongquestions-
2003.pdf Chen, J., Hua, G., Jurat-Fuentes, J.L., Abdullah, M.A., & Adang, M.J. (2007)
Synergism of Bacillus thuringiensis toxins by a fragment of a toxin-binding cadherin. Proceedings of the National Academy of Sciences of the United States of America, 104, 35, pp 13901-13906
<Go to ISI>://000249187500011 Chilton, M.D., Drummond, M.H., Merlo, D.J., & Sciaky, D. (1978)
Highly Conserved DNA of Ti Plasmids Overlaps T-DNA Maintained in Plant Tumors. Nature, 275, 5676, pp 147-149
<Go to ISI>://WOS:A1978FP06300045 AND http://www.botanischergarten.ch/Bt/Chilton-Highly-Conserved-DNA-1978.pdf
Chilton, M.D., Drummond, M.H., Merlo, D.J., Sciaky, D., Montoya, A.L., Gordon, M.P., & Nester, E.W. (1977)
Stable Incorporation of Plasmid DNA into Higher Plant-Cells - Molecular-Basis of Crown Gall Tumorigenesis. Cell, 11, 2, pp 263-271
<Go to ISI>://WOS:A1977DL69200003 AND http://www.botanischergarten.ch/Bt/Chilton-Stable-Incorporation-1977.pdf
Chilton, M.D., Saiki, R.K., Yadav, N., Gordon, M.P., & Quetier, F. (1980)
T-DNA from Agrobacterium Ti Plasmid Is in the Nuclear-DNA Fraction of Crown Gall Tumor-Cells. Proceedings of the National Academy of Sciences of the United States of America-Biological Sciences, 77, 7, pp 4060-4064
<Go to ISI>://WOS:A1980KB88900064 AND http://www.botanischergarten.ch/Bt/Chilton-T-DNA-Plasmid-1980.pdf
Christou, P., Capell, T., Kohli, A., Gatehouse, J.A., & Gatehouse, A.M.R. (2006)
Recent developments and future prospects in insect pest control in transgenic crops. Trends in Plant Science, 11, 6, pp 302-308
<Go to ISI>://000239064600008 AND http://www.botanischergarten.ch/Bt/Christou-Recent-Developments-2006.pdf
Conner, A.J., Glare, T.R., & Nap, J.-P. (2003)

64
The release of genetically modified crops into the environment. Part II. Overview of ecological risk assessment. The Plant Journal, 33, 1 %R doi:10.1046/j.0960-7412.2002.001607.x, pp 19-46
http://www.blackwell-synergy.com/doi/abs/10.1046/j.0960-7412.2002.001607.x AND http://www.botanischergarten.ch/Bt/Conner-Release-Review-2-2003.pdf
Cortet, J., Griffiths, B.S., Bohanec, M., Demsar, D., Andersen, M.N., Caul, S., Birch, A.N.E., Pemin, C., Tabone, E., de Vaufleury, A., Ke, X., & Krogh, P.H. (2007)
Evaluation of effects of transgenic Bt maize on microarthropods in a European multi-site experiment SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 207-218
http://www.sciencedirect.com/science/article/B7CW5-4NNYD85-1/2/5e21fd4b0b272b4eb569ade3131a1718 AND http://www.botanischergarten.ch/Bt/Cortet-Evaluation-Microarthropods-ECOGEN-2007.pdf
Crickmore, N. (2003)
Electronic Source: INSECTICIDAL PROTEINS FROM BACILLUS THURINGIENSIS, keywords Crickmore, CA Patent CA2395897 Wiki Patents, Community patent review
published by: WIKIPEDIA http://www.wikipatents.com/ca/2395897.html Crickmore, N., Zeigler, D.R., Feitelson, J., Schnepf, E., Van Rie, J., Lereclus, D., Baum, J., & Dean, D.H. (1998)
Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins. Microbiology and Molecular Biology Reviews, 62, 3, pp 807-+
<Go to ISI>://WOS:000075782400011 and http://www.botanischergarten.ch/Bt/Crickmore-Revision-Nomenclature-Bt-1998.pdf
Crickmore, N., Zeigler, D.R., Schnepf, E., Van Rie, J., Lereclus, D., Baum, J., Bravo, A., & Dean, D.H. (2008)
Electronic Source: Bacillus thuringiensis toxin nomenclature (ed N. Crickmore), http://www.lifesci.sussex.ac.uk/Home/Neil_Crickmore/Bt/ Croft, B.A. (1990)
Arthropod Biological Control Agents and Pesticides. Wiley and Sons, New York, pp
Dafny-Yelin, M., Levy, A., & Tzfira, T. (2008)
The ongoing saga of Agrobacterium-host interactions. Trends in Plant Science, 13, 3, pp 102-105 <Go to ISI>://WOS:000254900000002 AND http://www.botanischergarten.ch/Bt/Dafny-Yelin-Ongoing-Saga-
2008.pdf Dale, P.J. (2002)
The environmental impact of genetically modified (GM) crops: a review. Journal of Agricultural Science, 138, pp 245-248
http://www.botanischergarten.ch/biodiversity/Dale-Pot-Imp-nbt0602-567.pdf and http://www.botanischergarten.ch/biodiversity/Dale-Erratum-nbt0802-843b.pdf
Damgaard, P.H., Hansen, B.M., Pedersen, J.C., & Eilenberg, J. (1997)
Natural occurrence of Bacillus thuringiensis on cabbage foliage and in insects associated with cabbage crops. Journal of Applied Microbiology, 82, 2, pp 253-258
<Go to ISI>://WOS:A1997WM84400016 AND http://www.botanischergarten.ch/Bt/Damgaard-Natural-Occurrence-1997.pdf
de Amorim, G.V., Whittome, B., Shore, B., & Levin, D.B. (2001)
Identification of Bacillus thuringiensis subsp kurstaki strain HD1-like bacteria from environmental and human samples after aerial spraying of Victoria, British Columbia, Canada, with Foray 48B. Applied and Environmental Microbiology, 67, 3, pp 1035-1043
<Go to ISI>://WOS:000167266200003 AND http://www.botanischergarten.ch/Bt/Valadares-de-Amorim-Identification-Btk-2001.pdf
de Barros Moreira Beltrao, H. & Silva-Filha, M.H.N.L. (2007)
Interaction of Bacillus thuringiensis svar. israelensis Cry toxins with binding sites from Aedes aegypti (Diptera: Culicidae) larvae midgut. FEMS Microbiology Letters, 266, 2 %R doi:10.1111/j.1574-6968.2006.00527.x, pp 163-169
http://www.blackwell-synergy.com/doi/abs/10.1111/j.1574-6968.2006.00527.x AND http://www.botanischergarten.ch/Bt/deBarros-Interaction-Aedes-2007.pdf
De Block, M., Herreraestrella, L., Vanmontagu, M., Schell, J., & Zambryski, P. (1984)

65
Expression of Foreign Genes in Regenerated Plants and in Their Progeny. Embo Journal, 3, 8, pp 1681-1689 <Go to ISI>://A1984TC78000003 AND http://www.botanischergarten.ch/Bt/De-Block-Expression-Foreign-
1984.pdf De Cleene, M. & Deley, J. (1976)
Host Range of Crown Gall. Botanical Review, 42, 4, pp 389-466 <Go to ISI>://A1976DR88600001 AND http://www.botanischergarten.ch/Bt/DeCleene-Host-Range-Crown-
Gall-.pdf de Groote, H., Mugo, S., Bergvinson, D., & Odhiambo, B. (2004)
Debunking the myths of GM crops for Africa: The case of Bt maize in Kenya pp 36 Paper presented at the Annual Meetings of the American Agricultural Economics Association, August 4, Denver, Colorado (Report)
http://www.botanischergarten.ch/Bt/De-Groote-Debunking-Myths-2004.pdf de Maagd, R.A. (2002)
Environmental Impact of insect-resistant crop plants expressing a Bt-toxin, II Non-target effects (updated) and Bt-resistance in target insects, Note 218, Wageningen University Plant Research International pp 62 Wageningen (Report)
http://www.botanischergarten.ch/Bt/de-Maagd-Environmental-Impact-Nontarget-2002.pdf de Maagd, R.A. (2007)
Chapter 9: The use of Bacillus thuringiensis crystal proteins for insect control pp 14 (Manuscript) http://www.botanischergarten.ch/Bt/de-Maagdt-Bt-Script-Chapter-9-2007.pdf de Maagd, R.A., Bakker, P., Staykov, N., Dukiandjiev, S., Stiekema, W., & Bosch, D. (1999)
Identification of Bacillus thuringiensis delta-endotoxin Cry1C domain III amino acid residues involved in insect specificity. Applied and Environmental Microbiology, 65, 10, pp 4369-4374
<Go to ISI>://000082959100012 AND http://www.botanischergarten.ch/Bt/de-Maagd-Identification-Bt-1999.pdf de Maagd, R.A., Bravo, A., Berry, C., Crickmore, N., & Schnepf, H.E. (2003)
Structure, diversity, and evolution of protein toxins from spore-forming entomopathogenic bacteria. Annual Review of Genetics, 37, pp 409-433
<Go to ISI>://000187792500016 AND http://www.botanischergarten.ch/Bt/de-Maagd-Structure-Diversity-2003.pdf
de Maagd, R.A., Bravo, A., & Crickmore, N. (2001)
How Bacillus thuringiensis has evolved specific toxins to colonize the insect world. Trends in Genetics, 17, 4, pp 193-199
<Go to ISI>://000168718300020 AND http://www.botanischergarten.ch/Bt/de-Maagd-Bac-thuring-colonize-2001.pdf
de Maagd, R.A., Bravo, A., & Crickmore, N. (2005)
Bt toxin not guilty by association. Nature Biotechnology, 23, 7, pp 791-791 <Go to ISI>://000230427700016 AND http://www.botanischergarten.ch/Bt/deMaagd-Not-guilty-2005.pdf de Maagd, R.A., Weemen-Hendriks, M., Stiekema, W., & Bosch, D. (2000)
Bacillus thuringiensis delta-endotoxin Cry1C domain III can function as a specificity determinant for Spodoptera exigua in different, but not all, Cry1-Cry1C hybrids. Applied and Environmental Microbiology, 66, 4, pp 1559-1563
<Go to ISI>://000086284700047 NOCH NEBIS de Vaufleury, A., Kramarz, P.E., Binet, P., Cortet, J., Caul, S., Andersen, M.N., Plumey, E., Coerdassier, M., & Krogh, P.H. (2007)
Exposure and effects assessments of Bt-maize on non-target organisms (gastropods, microarthropods, mycorrhizal fungi) in microcosms, SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 185-194
http://www.sciencedirect.com/science/article/B7CW5-4NNPB70-3/2/5f608d81279243437505a067fdac57b1 AND http://www.botanischergarten.ch/Bt/Vaufleury-Exposure-Microcosmos-ECOGEN-2007.pdf
Debeljak, M., Cortet, J., Demsar, D., Krogh, P.H., & Dzeroski, S. (2007)
Hierarchical classification of environmental factors and agricultural practices affecting soil fauna under cropping systems using Bt maize. SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN, 51, 3, pp 229-238

66
http://www.sciencedirect.com/science/article/B7CW5-4NWMY85-1/2/d40c9d25c340c200f03168be3e388efd AND http://www.botanischergarten.ch/Bt/Debeljak-Hierarchical-Class-ECOGEN-2007.pdf
Deblock, M., Schell, J., & Vanmontagu, M. (1985)
Chloroplast Transformation by Agrobacterium-Tumefaciens. Embo Journal, 4, 6, pp 1367-1372 <Go to ISI>://A1985ALC7400002 Delucca, A.J., Simonson, J.G., & Larson, A.D. (1981)
Bacillus-Thuringiensis Distribution in Soils of the United-States. Canadian Journal of Microbiology, 27, 9, pp 865-870
<Go to ISI>://WOS:A1981MH20800001 AND NEBIS 20081003 Demont, M. & Tollens, E. (2004)
First impact of biotechnology in the EU: Bt maize adoption in Spain. Annals of Applied Biology, 145, 2, pp 197-207
<Go to ISI>://000224481800008 AND http://www.botanischergarten.ch/Bt/Demont-First-Impact-Spain-2004.pdf
Dollaker, A. & Rhodes, C. (2007)
Integrating crop productivity and biodiversity conservation pilot initiatives developed by Bayer CropScience, in Weed Science in Time of Transition. Crop Science, 26, 3, pp 408-416
http://www.sciencedirect.com/science/article/B6T5T-4MV1NS0-1/2/45ef7df1e5582e65fbde44fa32651b26 Douville, M., Gagne, F., Blaise, C., & Andre, C. (2007)
Occurrence and persistence of Bacillus thuringiensis (Bt) and transgenic Bt corn cry1Ab gene from an aquatic environment. Ecotoxicology and Environmental Safety, 66, 2, pp 195-203
<Go to ISI>://000243187000009 AND http://www.botanischergarten.ch/Bt/Douville-Occurrence-Persistence-2007.pdf
Douville, M., Gagne, F., Masson, L., McKay, J., & Blaise, C. (2005)
Tracking the source of Bacillus thuringiensis Cry1Ab endotoxin in the environment. Biochemical Systematics and Ecology, 33, 3, pp 219-232
<Go to ISI>://000227414300001 AND http://www.botanischergarten.ch/Bt/Douville-Tracking-Source-2005.pdf Downey, K., Voellmy, R., Ahmad, F., & Schultz, J., eds. (1983)
Advances in Gene Technology: Molecular Genetics of Plants and Animals., Academic Press, New York,
Drobnik, J. (2007)
Time to relax GMO regulation in Europe. Plant Cell, Tissue and Organ Culture, pp http://dx.doi.org/10.1007/s11240-007-9331-3 AND http://www.botanischergarten.ch/Regulation/Drobnik-Time-
Relax-2007.pdf Drummond, M.H., Gordon, M.P., Nester, E.W., & Chilton, M.D. (1977)
Foreign DNA of Bacterial Plasmid Origin Is Transcribed in Crown Gall Tumors. Nature, 269, 5628, pp 535-536
<Go to ISI>://WOS:A1977DW65000050 AND http://www.botanischergarten.ch/Bt/Drummond-Foreign-DNA-1977.pdf
Dutton, A., Klein, H., Romeis, J., & Bigler, F. (2003)
Prey-mediated effects of Bacillus thuringiensis spray on the predator Chrysoperla carnea in maize. Biological Control, 26, 2, pp 209-215
<Go to ISI>://000181149000014 ENDS Report (2008)
Ministers and scientists urge rethink on GM crops. ENDS Report, July, pp 25-17 http://www.botanischergarten.ch/Bt/ENDS-Article-GM-crops-2008.pdf English, L., Brussock, S., Malvar, T., Bryson, J., Kulesza, C., Walters, F.S., Slatin, S., Von Tersch, M., & Romano, C., Inventors (2003)
Coleopteran-resistant transgenic plants and methods of their production., U.S.A.US Patent No. 7,227,056
English, L. & Slatin, S.L. (1992)
Mode of Action of Delta-Endotoxins from Bacillus-Thuringiensis - a Comparison with Other Bacterial Toxins. Insect Biochemistry and Molecular Biology, 22, 1, pp 1-7

67
<Go to ISI>://WOS:A1992HH82300001 EPA Environmental Protection Agency (1998)
The Environmental Protection Agency’s White Paper on Bt Plant-pesticide Resistance Management, EPA pp 79 Biopesticides and Pollution Prevention Division Office of Pesticide Programs Office of the Assistant Administrator for Prevention,Pesticides and Toxic Substances Washington (Report)
www.epa.gov/EPAPEST/1998/January/Day-14/paper.pdf AND http://www.botanischergarten.ch/Bt/EPA-Resistance-Management-1998.pdf
Estruch, J.J., Warren, G.W., Mullins, M.A., Nye, G.J., Craig, J.A., & Koziel, M.G. (1996)
Vip3A, a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects. Proceedings of the National Academy of Sciences of the United States of America, 93, 11, pp 5389-5394
<Go to ISI>://WOS:A1996UN25300040 AND http://www.botanischergarten.ch/Bt/Estruch-Vip3A-Novel-Bt-vegetative-1996.pdf
Eurobarometer (2006)
Europeans and Biotechnology in 2005:Patterns and Trends, TNS Opinion & Social network, European Commission pp 88 Final report on Eurobarometer 64.3 Brussels (Report)
http://ec.europa.eu/public_opinion/archives/ebs/ebs_244b_en.pdf AND http://www.botanischergarten.ch/Eurobaro/Eurobaro-2005-ebs_244b_en.pdf
Eurobarometer Special (2008)
Attitudes of European citizens towards the environment 295, TNS Opinion & Social network, European Commission pp 127 Wave 68.2 Brussels (Report)
http://ec.europa.eu/public_opinion/archives/ebs/ebs_295_en.pdf AND http://www.botanischergarten.ch/Eurobaro/Eurobaro-2007-ebs_295_en.pdf AND slides http://www.botanischergarten.ch/Eurobaro/Eurobarometer-Gaskell-2007.ppt
Fabrick, J.A. & Tabashnik, B.E. (2007)
Binding of Bacillus thuringiensis toxin Cry1Ac to multiple sites of cadherin in pink bollworm. Insect Biochemistry and Molecular Biology, 37, 2, pp 97-106
<Go to ISI>://WOS:000244239700001 Farinos, G.P., de la Poza, M., Hernandez-Crespo, P., Ortego, F., & Castanera, P. (2004)
Resistance monitoring of field populations of the corn borers Sesamia nonagrioides and Ostrinia nubilalis after 5 years of Bt maize cultivation in Spain. Entomologia Experimentalis Et Applicata, 110, 1, pp 23-30
<Go to ISI>://000187405100003 AND http://www.botanischergarten.ch/Bt/Farinos-Resistance-Spain-2004.pdf Federici, B. (1999) Bacillus thuringiensis in Biological Control, Chapter 21. In Handbook of Biological Control (eds T.S. Bellows,
G. Gorth & T.W. Fischer), Vol. Chapter 21. Academic Press, San Diego Federici, B.A., Luthy, P., & lbarra, J.E. (1991) The parasporal bodv of Bacillus thuringiensis subsp. israelensis: structure. protein composition and toxicity. In
Bacterial control of mosquitoes and blackflies: biochemistry. genetics and applications of Bacillus thuringiensis and Bacillus sphaericus. (eds H. de Barjac & S. Sutherland), pp. 336pp. Rudgers University Press, New Brunswick, N.J. U.S.A.
Federici, B.A. & Siegel, J.P. (2007)
3. Safety Assessment of Bacillus thuringiensis and Bt Crops Used in Insect Control. In Food Safety of Proteins in Agricultural Biotechnology (Food Science and Technology) (ed B.G. Hammond), pp. 45-102. Taylor and Francis, CRC; 1 edition (November 28, 2007), Bota Raton, Florida, U.S.A.
Ferre, J., Van Rie, J., & MacIntosh, S. (2008) Insecticidal Genetically Modified Crops and Insect Resistance Management (IRM). In Integration of Insect-
Resistant Genetically Modified Crops within IPM Programs (eds J. Romeis, A.M. Shelton & G.G. Kennedy), Vol. Progress in Biological Control 5, pp. 45. Springer, Heidelberg
http://www.amazon.com/gp/reader/1402083726/ref=sib_dp_pt#reader-link AND http://www.botanischergarten.ch/Bt/Ferre-Insecticidal-GM-IRM-2008.pdf
Fiorito, T.M., Icoza, I., & Stotzky, G. (2007)

68
Adsorption and binding of the transgenic plant proteins, human serum albumin, [beta]-glucuronidase, and Cry3Bb1, on montmorillonite and kaolinite: Microbial utilization and enzymatic activity of free and clay-bound proteins. Applied Clay Science In Press, Corrected Proof, online, pp xxx
http://www.sciencedirect.com/science/article/B6V8Y-4PB6VPW-1/2/487fcf6b24b8088c723b11f099509867 AND doi:10.1016/j.clay.2007.07.006 AND http://www.botanischergarten.ch/Bt/Fiorito-Adsorption-Binding-2007.pdf
Fischer, G. (2000)
Symmetry of ignorance, social creativity, and meta-design. Knowledge-Based Systems, 13, 7-8, pp 527-537 <Go to ISI>://000165855800011 AND http://www.botanischergarten.ch/Discourse/Fischer-Symmetry-
Ignorance-2000.pdf Fischhoff, D.A., Bowdish, K.S., Perlak, F.J., Marrone, P.G., McCormick, S.M., Niedermeyer, J.G., Dean, D.A., Kusanokretzmer, K., Mayer, E.J., Rochester, D.E., Rogers, S.G., & Fraley, R.T. (1987)
Insect Tolerant Transgenic Tomato Plants. Bio-Technology, 5, 8, pp 807-813 <Go to ISI>://WOS:A1987J380000017 AND http://www.botanischergarten.ch/Bt/Fischhoff-Insect-Tolerant-
Tomato-1987.pdf Flexner, J.L., Lighthart, B., & Croft, B.A. (1986)
The Effects of Microbial Pesticides on Nontarget, Beneficial Arthropods. Agriculture Ecosystems & Environment, 16, 3-4, pp 203-254
<Go to ISI>://A1986D869700005 AND NEBIS 20080101 Fox, J.L. (1995)
EPA okays Bt corn; USDA eases plant testing. Bio-Technology, 13, October, pp 1035 <Go to ISI>://A1991HA47200009 AND http://www.botanischergarten.ch/Bt/Fox-EPA-Okays-1995.pdf Fox, J.L. (1997)
EPA seeks refuge from Bt resistance. Nature Biotechnology, 15, 5, pp 409-409 <Go to ISI>://A1997WX23700013 AND http://www.botanischergarten.ch/Bt/Fox-EPA-Seeks-Refuge-1997.pdf Fox, J.L. (2000)
Withdrawn Greenpeace Bt suit enters spin cycle. Nature Biotechnology, 18, 9, pp 920-920 <Go to ISI>://WOS:000089078300009 AND http://www.botanischergarten.ch/Bt/Fox-GP-Spin-Cycle-2000.pdf Fox, J.L. (2003a)
Resistance to Bt toxin surprisingly absent from pests. Nature Biotechnology, 21, 9, pp 958-959 <Go to ISI>://000185051000006 AND http://www.botanischergarten.ch/Bt/Fox-Resistance-2006.pdf Fox, J.L. (2003b)
Widespread commercial use not triggering expected Bt resistance. Asm News, 69, 10, pp 484-485 <Go to ISI>://000187558200007 AND http://www.botanischergarten.ch/Bt/Fox-Widespread-Resistance-
2003.pdf Fraley, R.T., Rogers, S.G., Horsch, R.B., Sanders, P.R., Flick, J.S., Adams, S.P., Bittner, M.L., Brand, L.A., Fink, C.L., Fry, J.S., Galluppi, G.R., Goldberg, S.B., Hoffmann, N.L., & Woo, S.C. (1983)
Expression of Bacterial Genes in Plant-Cells. Proceedings of the National Academy of Sciences of the United States of America-Biological Sciences, 80, 15, pp 4803-4807
<Go to ISI>://WOS:A1983RB47600043 AND http://www.botanischergarten.ch/Bt/Fraley-Expression-bacterial-1983.pdf
Frederiksen, K., Rosenquist, H., Jorgensen, K., & Wilcks, A. (2006)
Occurrence of natural Bacillus thuringiensis contaminants and residues of Bacillus thuringiensis-based insecticides on fresh fruits and vegetables. Applied and Environmental Microbiology, 72, 5, pp 3435-3440
<Go to ISI>://000237491200044 AND http://www.botanischergarten.ch/Bt/Frederiksen-Occurence-natural-Bt-2006.pdf
Freeling, M.E. (1994)
Maize Handbook Springer; 1 edition (1994) IS: ISBN-10: 0387978267 ISBN-13: 978-0387978260, pp 759
Fromm, M., Taylor, L.P., & Walbot, V. (1985)
Expression of genes transferred into monocot and dicot plant cells by electroporation. Proceedings of the National Academy of Sciences of the United States of America, 82, 17, pp 5824-5828 %R
http://www.pnas.org/content/82/17/5824.abstract AND http://www.pnas.org/content/82/17/5824.full.pdf+html

69
Fromm, M.E., Taylor, L.P., & Walbot, V. (1986)
Stable Transformation of Maize after Gene-Transfer by Electroporation. Nature, 319, 6056, pp 791-793 <Go to ISI>://A1986A232100065 AND NEBIS 20080723 Gahan, L.J., Ma, Y.T., Coble, M.L.M., Gould, F., Moar, W.J., & Heckel, D.G. (2005)
Genetic basis of resistance to Cry1Ac and Cry2Aa in Heliothis virescens (Lepidoptera : Noctuidae). Journal of Economic Entomology, 98, 4, pp 1357-1368
<Go to ISI>://000231056100037 AND http://www.botanischergarten.ch/Bt/Gahan-Basis-Resistance-2005.pdf Garcia-Robles, I., Sanchez, J., Gruppe, A., Martinez-Ramirez, A.C., Rausell, C., Real, M.D., & Bravo, A. (2001)
Mode of action of Bacillus thuringiensis PS86Q3 strain in hymenopteran forest pests. Insect Biochemistry and Molecular Biology, 31, 9, pp 849-856
<Go to ISI>://WOS:000170013900002 AND http://www.ask-force.org/web/Bt1/Garcia-Robles-Mode-Action-2001.pdf
Gatehouse, A.M.R., Ferry, N., & Raemaekers, R.J.M. (2002)
The case of the monarch butterfly: a verdict is returned. Trends in Genetics, 18, 5, pp 249-251 http://www.botanischergarten.ch/Bt/GatehouseMonarch.pdf Gatehouse, J.A. (2008)
Biotechnological prospects for engineering insect-resistant plants. Plant Physiology, 146, 3, pp 881-887 <Go to ISI>://WOS:000256412200013 AND http://www.botanischergarten.ch/Bt/Gatehouse-Biotechnological-
Prospects-2008.pdf Gay, J. (1999)
Mais, mythe et realite Atlantica, Biarritz, IS: 2-84394128-8, pp 619
Genetello, C., Vanlarebeke, N., Holsters, M., Depicker, A., Vanmontagu, M., & Schell, J. (1977)
Ti Plasmids of Agrobacterium as Conjugative Plasmids. Nature, 265, 5594, pp 561-563 <Go to ISI>://WOS:A1977CU78200048 AND http://www.botanischergarten.ch/Bt/Genetello-Plasmids-
Agrobacterium-Nature-1977.pdf Glare, T.R. & O'Callaghan, M. (2000)
Bacillus Thuringiensis - Biology, Ecology & Safety (Hardcover), 1. Edition edn. Wiley; 1 edition (September 7, 2000) New York, pp 368
See also the NZ-Report: http://www.botanischergarten.ch/Bt/Glare-OCallaghan-Biocontrol-Biodiversity-1998.pdf
Godreuil, S., Cohan, F., Shah, H., & Tibayrenc, M. (2005)
Which species concept for pathogenic bacteria? An E-Debate. Infection Genetics and Evolution, 5, 4, pp 375-387
<Go to ISI>://BIOSIS:PREV200600072568 AND http://www.botanischergarten.ch/Bt/Godreuil-Which-Species-Concept-2005.pdf
Gomez, I., Arenas, I., Benitez, I., Miranda-Rios, J., Becerril, B., Grande, R., Almagro, J.C., Bravo, A., & Soberon, M. (2006)
Specific epitopes of domains II and III of Bacillus thuringiensis Cry1Ab toxin involved in the sequential interaction with cadherin and aminopeptidase-N receptors in Manduca sexta. Journal of Biological Chemistry, 281, 45, pp 34032-34039
<Go to ISI>://000241767600023 AND http://www.botanischergarten.ch/Bt/Gomez-Epitopes-2006.pdf Gomez, I., Dean, D.H., Bravo, A., & Soberon, M. (2003)
Molecular basis for Bacillus thuringiensis Cry1Ab toxin specificity: Two structural determinants in the Manduca sexta Bt-R-1 receptor interact with loops alpha-8 and 2 in domain II of Cy1Ab toxin. Biochemistry, 42, 35, pp 10482-10489
<Go to ISI>://000185170300016 AND http://www.botanischergarten.ch/Gomez-Basis-Specifity-2003.pdf Gomez, I., Oltean, D.I., Gill, S.S., Bravo, A., & Soberon, M. (2001)
Mapping the epitope in cadherin-like receptors involved in Bacillus thuringiensis Cry1A toxin interaction using phage display. Journal of Biological Chemistry, 276, 31, pp 28906-28912
<Go to ISI>://000170346000037

70
Gomez, I., Pardo-Lopez, L., Munoz-Garay, C., Fernandez, L.E., Perez, C., Sanchez, J., Soberon, M., & Bravo, A. (2007)
Role of receptor interaction in the mode of action of insecticidal Cry and Cyt toxins produced by Bacillus thuringiensis. Peptides, 28, 1, pp 169-173
<Go to ISI>://000243802500023 AND http://www.botanischergarten.ch/Bt/Gomez-Receptor-Interaction-2006.pdf
Gonzalez-Cabrera, J., Farinos, G.P., Caccia, S., Diaz-Mendoza, M., Castanera, P., Leonardi, M.G., Giordana, B., & Ferre, J. (2006)
Toxicity and mode of action of Bacillus thuringiensis cry proteins in the Mediterranean corn borer, Sesamia nonagrioides (Lefebvre). Applied and Environmental Microbiology, 72, 4, pp 2594-2600
<Go to ISI>://WOS:000236749400041 AND http://www.botanischergarten.ch/Bt/Gonzales-Cabrera-Toxicity-Mode-Bt-2006.pdf
Gordon-Kamm, W.J., Spencer, T.M., Mangano, M.L., Adams, T.R., Daines, R.J., Start, W.G., Obrien, J.V., Chambers, S.A., Adams, W.R., Willetts, N.G., Rice, T.B., Mackey, C.J., Krueger, R.W., Kausch, A.P., & Lemaux, P.G. (1990)
Transformation of Maize Cells and Regeneration of Fertile Transgenic Plants. Plant Cell, 2, 7, pp 603-618 <Go to ISI>://WOS:A1990DP61000003 AND http://www.botanischergarten.ch/Bt/Gordon-Kamm-
Transformation-Maize-1990.pdf Gordona, D., Karbat, I., Ilana, N., Cohena, L., Kahna, R., Giles, N., Dong, K., Stuehmer, W., Tytgate, J., & Gurevitza, M. (2007)
The differential preference of scorpion [alpha]-toxins for insect or mammalian sodium channels: Implications for improved insect control. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 452-472
http://www.sciencedirect.com/science/article/B6TCS-4MFJ2GX-3/2/27400b70afc9d8c1592b32ad4d617789 AND http://www.botanischergarten.ch/Bt/Gordon-Differencial-Preference-Scorpion-2007.pdf
Gouse, M., Pray, C.E., Kirsten, J., & Schimmelpfennig, D. (2005)
A GM subsistence crop in Africa: the case of Bt white maize in South Africa. International Journal of Biotechnology, 7, 1-3, pp 84-94
DOI 10.1504/..006447 Gray, A.J. (2004)
Ecology and government policies: the GM crop debate. J Appl Ecology, 41, 1, pp 1-10 http://www.blackwell-synergy.com/links/doi/10.1111/j.1365-2664.2004.00873.x/abs Greenpeace (2002)
Environmental Dangers of Insect Resistant Bt Crops pp 4 Genetic Engineering Briefing Pack (Report) http://www.greenpeace.org/raw/content/chile/press/reports/environmental-danger-of-insect.pdf Gressel, J., Hanafi, A., Head, G., Marasas, W., Obilana, B., Ochanda, J., Souissi, T., & Tzotzos, G. (2004)
Major heretofore intractable biotic constraints to African food security that may be amenable to novel biotechnological solutions. Crop Protection, 23, 8, pp 661-689
http://www.botanischergarten.ch/Mycotoxins/Gressel-African-bioconstraints.pdf Griffiths, B.S., Caul, S., Thompson, J., Birch, A.N.E., Cortet, J., Andersen, M.N., & Krogh, P.H. (2007)
Microbial and microfaunal community structure in cropping systems with genetically modified plants SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 195-206
http://www.sciencedirect.com/science/article/B7CW5-4NNPB70-1/2/2ff9d77eeb929f2df2857021f6998604 AND http://www.botanischergarten.ch/Bt/Griffiths-Microbial-Community-ECOGEN-2007.pdf
Griffitts, J.S., Haslam, S.M., Yang, T.L., Garczynski, S.F., Mulloy, B., Morris, H., Cremer, P.S., Dell, A., Adang, M.J., & Aroian, R.V. (2005)
Glycolipids as receptors for Bacillus thuringiensis crystal toxin. Science, 307, 5711, pp 922-925 <Go to ISI>://000226985100053 AND http://www.botanischergarten.ch/Bt/Griffitts-Glycolipids-Receptors-
2005.pdf Griko, N.B., Rose-Young, L., Zhang, X., Carpenter, L., Candas, M., Ibrahim, M.A., Junker, M., & Bulla, L.A. (2007)
Univalent binding of the Cry1Ab toxin of Bacillus thuringiensis to a conserved structural motif in the cadherin receptor BT-R-1. Biochemistry, 46, 35, pp 10001-10007
<Go to ISI>://000249021100014 AND http://www.botanischergarten.ch/Bt/Griko-Univalent-Binding-2007.pdf

71
Gruber, C.W., M., C., Anderson, M.A., & Craik, D.J. (2007)
Insecticidal plant cyclotides and related cystine knot toxins. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 561-575
http://www.sciencedirect.com/science/article/B6TCS-4MG0641-3/2/44de411aa4f995a033d8805bfab62e67 AND http://www.botanischergarten.ch/Bt/Gruber-Insecticidal-Cyclotides-2007.pdf
Gruere, G.P. & Rosegrant, M.W. (2008)
Assessing the implementation effects of the biosafety protocol's proposed stringent information requirements for genetically modified commodities in countries of the Asia Pacific Economic Cooperation. Review of Agricultural Economics, 30, 2, pp 214-232
<Go to ISI>://WOS:000256102200002 AND http://www.botanischergarten.ch/Bt/Gruere-Assessing-Implementation-Effects-2008.pdf
Guehlstorf, N.P. & Hallstrom, L.K. (2005)
The role of culture in risk regulations: a comparative case study of genetically modified corn in the United States of America and European Union. Environmental Science & Policy, 8, 4, pp 327-342
<Go to ISI>://000231352300001 and http://www.botanischergarten.ch/Discourse/Guehlsdorf-Culture-Regulation-2005.pdf
Gurevitza, M., Karbat, I., Cohen, L., Ilan, N., Kahn, R., Turkov, M., Stankiewicz, M., Stuehmer, W., Dong, K., & Gordon, D. (2007)
The insecticidal potential of scorpion [beta]-toxins. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 473-489
http://www.sciencedirect.com/science/article/B6TCS-4MFJ2GX-2/2/7472be2cba3780081202ebd29b67c300 Haddad, M.D.L., Polanczyk, R.A., Alves, S.B., & Garcia, M.D.O. (2005)
Field persistence of Bacillus thuringiensis on maize leaves (Zea mays L.). Brazilian Journal of Microbiology, 36, 4, pp 309-314
<Go to ISI>://000238508900001 AND http://www.botanischergarten.ch/Bt/Haddad-Field-Persistence-Bt-2005.pdf
Hails, R.S. (2002)
Assessing the risks associated with new agricultural practices. Nature, 418, 6898, pp 685-688 http://dx.doi.org/10.1038/nature01016 AND http://www.botanischergarten.ch/Bt/Hails-Assessing-2002.pdf Han, C.S., Xie, G., Challacombe, J.F., Altherr, M.R., Bhotika, S.S., Brown, N., Bruce, D., Campbell, C.S., Campbell, M.L., Chen, J., Chertkov, O., Cleland, C., Dimitrijevic, M., Doggett, N.A., Fawcett, J.J., Glavina, T., Goodwin, L.A., Green, L.D., Hill, K.K., Hitchcock, P., Jackson, P.J., Keim, P., Kewalramani, A.R., Longmire, J., Lucas, S., Malfatti, S., McMurry, K., Meincke, L.J., Misra, M., Moseman, B.L., Mundt, M., Munk, A.C., Okinaka, R.T., Parson-Quintana, B., Reilly, L.P., Richardson, P., Robinson, D.L., Rubin, E., Saunders, E., Tapia, R., Tesmer, J.G., Thayer, N., Thompson, L.S., Tice, H., O Ticknor, L., Wills, P.L., Brettin, T.S., & Gilna, P. (2006a)
Pathogenomic sequence analysis of Bacillus cereus and Bacillus thuringiensis isolates closely related to Bacillus anthracis (vol 188, pg 3382, 2006). Journal of Bacteriology, 188, 21, pp 7711-7711
<Go to ISI>://WOS:000241471600046 Han, C.S., Xie, G., Challacombe, J.F., Altherr, M.R., Bhotika, S.S., Bruce, D., Campbell, C.S., Campbell, M.L., Chen, J., Chertkov, O., Cleland, C., Dimitrijevic, M., Doggett, N.A., Fawcett, J.J., Glavina, T., Goodwin, L.A., Hill, K.K., Hitchcock, P., Jackson, P.J., Keim, P., Kewalramani, A.R., Longmire, J., Lucas, S., Malfatti, S., McMurry, K., Meincke, L.J., Misra, M., Moseman, B.L., Mundt, M., Munk, A.C., Okinaka, R.T., Parson-Quintana, B., Reilly, L.P., Richardson, P., Robinson, D.L., Rubin, E., Saunders, E., Tapia, R., Tesmer, J.G., Thayer, N., Thompson, L.S., Tice, H., Ticknor, L.O., Wills, P.L., Brettin, T.S., & Gilna, P. (2006b)
Pathogenomic Sequence Analysis of Bacillus cereus and Bacillus thuringiensis Isolates Closely Related to Bacillus anthracis. Journal of Bacteriology, 188, 9, pp 3382-3390
http://jb.asm.org/cgi/content/abstract/188/9/3382 AND http://www.botanischergarten.ch/Bt/Han-Pathogenomic-Sequence-2006.pdf
Hansen, B.M. & Hendriksen, N.B. (2001)
Detection of enterotoxic Bacillus cereus and Bacillus thuringiensis strains by PCR analysis. Applied and Environmental Microbiology, 67, 1, pp 185-189
<Go to ISI>://WOS:000166151300024 AND http://www.botanischergarten.ch/Bt/Hansen-Detection-Enterotoxic-2001.pdf
Harvey, W.R., Cioffi, M., Dow, J.A.T., & Wolfersberger, M.G. (1983)
Potassium-Ion Transport ATPase in Insect Epithelia. Journal of Experimental Biology, 106, SEP, pp 91-&

72
<Go to ISI>://WOS:A1983RU29900008 AND http://www.botanischergarten.ch/Bt/Harvey-Potassium-Transport-1983.pdf
Hastowo, S., Lay, B.W., & Ohba, M. (1992)
Naturally-Occurring Bacillus-Thuringiensis in Indonesia. Journal of Applied Bacteriology, 73, 2, pp 108-113 <Go to ISI>://WOS:A1992JG80500002 Head, G., Surber, J.B., Watson, J.A., Martin, J.W., & Duan, J.J. (2002)
No detection of Cry1Ac protein in soil after multiple years of transgenic Bt cotton (Bollgard) use. Environmental Entomology, 31, 1, pp 30-36
http://www.botanischergarten.ch/Bt/Head-Monitoring-Bt-v31n1p30.pdf Heckel, D.G., Gahan, L.J., Baxter, S.W., Zhao, J.Z., Shelton, A.M., Gould, F., & Tabashnik, B.E. (2007)
The diversity of Bt resistance genes in species of Lepidoptera. Journal of Invertebrate Pathology, 95, 3, pp 192-197
<Go to ISI>://000247850700008 AND http://www.botanischergarten.ch/Bt/Heckel-Diversity-Bt-resistance-2007.pdf
Heimpel, A.M. & Angus, T.A. (1960)
Bacterial Insecticides. Bacteriological Reviews, 24, 3, pp 266-288 <Go to ISI>://WOS:A1960WA15000002 AND http://www.botanischergarten.ch/Bt/Heimpel-Bacterial-
Insecticides-1960.pdf Heinemann, J.A. & Traavik, T. (2004a)
Monitoring horizontal gene transfer - Reply. Nature Biotechnology, 22, 11, pp 1349-1350 <Go to ISI>://000224960600015 AND http://www.botanischergarten.ch/HorizontalGT/Heinemann-Traavik-
Corrigendum-nbt0405-488d-2005.pdf Heinemann, J.A. & Traavik, T. (2004b)
Problems in monitoring horizontal gene transfer in field trials of transgenic plants. Nature Biotechnology, 22, 9, pp 1105-1109
<Go to ISI>://000223653400028 http://www.botanischergarten.ch/HorizontalGT/Heinemann-Problems-NB-2004.pdf
Helgason, E., Okstad, O.A., Caugant, D.A., Johansen, H.A., Fouet, A., Mock, M., Hegna, I., & Kolsto, A.-B. (2000)
Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis---One Species on the Basis of Genetic Evidence. Appl. Environ. Microbiol. %R 10.1128/AEM.66.6.2627-2630.2000, 66, 6, pp 2627-2630
http://aem.asm.org/cgi/content/abstract/66/6/2627 AND http://www.botanischergarten.ch/Bt/Helgason-Cereus-Group-HT-2000.pdf
Hernandez, M., Pla, M., Esteve, T., Prat, S., Puigdomenech, P., & Ferrando, A. (2003)
A specific real-time quantitative PCR detection system for event MON810 in maize YieldGard (R) based on the 3 '-transgene integration sequence. Transgenic Research, 12, 2, pp 179-189
<Go to ISI>://WOS:000181852600005 AND http://www.botanischergarten.ch/Bt/Hernandez-Specific-Real-Time-PCR-2003.pdf
Herrera-Estrella, L., Deblock, M., Messens, E., Hernalsteens, J.P., Vanmontagu, M., & Schell, J. (1983a)
Chimeric Genes as Dominant Selectable Markers in Plant-Cells. Embo Journal, 2, pp 987-995 <Go to ISI>://WOS:A1983QU57500027 AND http://www.botanischergarten.ch/Bt/Herrera-Estrella-Chimeric-
Genes-1983.pdf Herrera-Estrella, L., Depicker, A., Vanmontagu, M., & Schell, J. (1983b)
Expression of Chimaeric Genes Transferred into Plant-Cells Using a Ti-Plasmid-Derived Vector. Nature, 303, 5914, pp 209-213
<Go to ISI>://WOS:A1983QQ80500028 AND http://www.botanischergarten.ch/Bt/Herrera-Estrella-Expression-Chimeric-1983.pdf
Herrero, S., Gonzalez-Cabrera, J., Ferre, J., Bakker, P.L., & De Maagd, R.A. (2004)
Mutations in the Bacillus thuringiensis Cry1Ca toxin demonstrate the role of domains II and III in specificity towards Spodoptera exigua larvae. Biochemical Journal, 384, pp 507-513
<Go to ISI>://000226084000007 Hickle, L.A. & Fitch, W.L. (1998)

73
Analytical Chemistry of Bacillus thuringiensis (Acs Symposium Series) An American Chemical Society Publication (March 10, 1998) IS: ISBN-10: 0841218153, ISBN-13: 978-0841218154 pp 149
Higuchi, M., Haginoya, K., Yamazaki, T., Miyamoto, K., Katagiri, T., Tomimoto, K., Shitomi, Y., Hayakawa, T., Sato, R., & Hori, H. (2007)
Binding of Bacillus thuringiensis Cry1A toxins to brush border membrane vesicles of midgut from Cry1Ac susceptible and resistant Plutella xylostella. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology, 147, 4, pp 716-724
<Go to ISI>://WOS:000248110600015 Hilder, V.A. & Boulter, D. (1999)
Genetic engineering of crop plants for insect resistance - a critical review. Crop Protection, 18, 3, pp 177-191 http://www.botanischergarten.ch/Bt/Hilder-Insect-Resistance-critical-1999.pdf Hill, K.K., Ticknor, L.O., Okinaka, R.T., Asay, M., Blair, H., Bliss, K.A., Laker, M., Pardington, P.E., Richardson, A.P., Tonks, M., Beecher, D.J., Kemp, J.D., Kolsto, A.B., Wong, A.C.L., Keim, P., & Jackson, P.J. (2004)
Fluorescent amplified fragment length polymorphism analysis of Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis isolates. Applied and Environmental Microbiology, 70, 2, pp 1068-1080
<Go to ISI>://WOS:000188854900054 Hill, M., Launis, K., Bowman, C., McPherson, K., Dawson, J., Watkins, J., Koziel, M., & Wright, M.S. (1995)
Biolistic Introduction of a Synthetic Bt Gene into Elite Maize. Euphytica, 85, 1-3, pp 119-123 <Go to ISI>://A1995TF37600016 AND http://www.botanischergarten.ch/Bt/Hill-Biolistic-Bt-Maize-1995.pdf Hodgson, E. (2008)
Scientists cry foul as Europe plays politics with GM crops. Nature Biotechnology, 26, 2, pp 139-140 http://dx.doi.org/10.1038/nbt0208-139 AND http://www.botanischergarten.ch/Bt/Hodgson-Cry-Foul-2008.pdf Hofmann, C. & Luthy, P. (1986)
Binding and Activity of Bacillus-Thuringiensis Delta-Endotoxin to Invertebrate Cells. Archives of Microbiology, 146, 1, pp 7-11
<Go to ISI>://WOS:A1986F014200002 AND http://www.botanischergarten.ch/Bt/Hofmann-Binding-Activity-1986.pdf
Hofmann, C., Luthy, P., Hutter, R., & Pliska, V. (1988a)
Binding of the Delta Endotoxin from Bacillus-Thuringiensis to Brush-Border Membrane-Vesicles of the Cabbage Butterfly (Pieris-Brassicae). European Journal of Biochemistry, 173, 1, pp 85-91
<Go to ISI>://WOS:A1988M765300012 AND http://www.botanischergarten.ch/Bt/Hofmann-Binding-Endotoxin-Bt-1988.pdf
Hofmann, C., Vanderbruggen, H., Hofte, H., Vanrie, J., Jansens, S., & Vanmellaert, H. (1988b)
Specificity of Bacillus-Thuringiensis Sigma-Endotoxins Is Correlated with the Presence of High-Affinity Binding-Sites in the Brush-Border Membrane of Target Insect Midguts. Proceedings of the National Academy of Sciences of the United States of America, 85, 21, pp 7844-7848
<Go to ISI>://WOS:A1988Q834100004 AND http://www.botanischergarten.ch/Bt/Hofmann-Specifity-Bt-Endotoxins-Correlation-1988.pdf
Hofte, H., Degreve, H., Seurinck, J., Jansens, S., Mahillon, J., Ampe, C., Vandekerckhove, J., Vanderbruggen, H., Vanmontagu, M., Zabeau, M., & Vaeck, M. (1986)
Structural and Functional-Analysis of a Cloned Delta Endotoxin of Bacillus-Thuringiensis-Berliner 1715. European Journal of Biochemistry, 161, 2, pp 273-280
<Go to ISI>://WOS:A1986F108400003 AND http://www.botanischergarten.ch/Bt/Hofte-Structural-Functional-Bt-1986.pdf
Hofte, H. & Whiteley, H.R. (1989)
Insecticidal Crystal Proteins of Bacillus-Thuringiensis. Microbiological Reviews, 53, 2, pp 242-255 <Go to ISI>://WOS:A1989AA13900005 AND http://www.botanischergarten.ch/Bt/Hofte-Insecticidal-Crystal-Bt-
1989.pdf Holsters, M. & Schell, J. (1975)
Agrobacterium-Tumefaciens Bacteriophages and Crown Gall. Annales De Microbiologie, B126, 4, pp 517-518 <Go to ISI>://WOS:A1975BB62300013 AND http://www.botanischergarten.ch/Bt/Holsters-Bacteriophages-
Agrobacterium-1975.pdf

74
Hossain, D.M., Hayakawa, T., Shitomi, Y., Itoh, K., Mitsui, T., Sato, R., & Hori, H. (2007)
Histochemical analysis of Bacillus thuringiensis Cry1A toxin binding to midgut epithelial cells of Bombyx mori. Pesticide Biochemistry and Physiology, 87, 1, pp 30-38
<Go to ISI>://WOS:000242741800005 Hwang, S.H., Saitoh, H., Mizuki, E., Higuchi, K., & Ohba, M. (1998)
A novel class of mosquitocidal delta-endotoxin, Cry19B, encoded by a Bacillus thuringiensis serovar higo gene. Systematic and Applied Microbiology, 21, 2, pp 179-184
<Go to ISI>://WOS:000074998800002 AND http://www.botanischergarten.ch/Bt/Hwang-Novel-Class-Endotoxin-1998.pdf
Ibiza-Palacios, M.S., Ferre, J., Higurashi, S., Miyamoto, K., Sato, R., & Escriche, B. (2008)
Selective inhibition of binding of Bacillus thuringiensis Cry1Ab toxin to cadherin-like and aminopeptidase proteins in brush-border membranes and dissociated epithelial cells from Bombyx mori. Biochemical Journal, 409, pp 215-221
<Go to ISI>://WOS:000252414300022 AND http://www.botanischergarten.ch/Bt/Ibiza-Palacios-Seledtive-Inhibition-2008.pdf
Ikanovic, M., Rudzinski, W.E., Bruno, J.G., Allman, A., Carrillo, M.P., Dwarakanath, S., Bhahdigadi, S., Rao, P., Kiel, J.L., & Andrews, C.J. (2007)
Fluorescence assay based on aptamer-quantum dot binding to Bacillus thuringiensis spores. Journal of Fluorescence, 17, 2, pp 193-199
<Go to ISI>://000244682300010 Ishiwata, S. (1901-1902)
On a type of severe Xacherie (sotto disease).(Original in Japanese). Dainihon Sanshi Kaiho, 114, pp 1-5
Jackson, R.E., Bradley, J.R., Van Duyn, J., Leonard, B.R., Allen, K.C., Luttrell, R., Ruberson, J., Adamczyk, J., Gore, J., Hardee, D.D., Voth, R., Sivasupramaniam, S., Mullins, J.W., & Head, G. (2008)
Regional assessment of Helicoverpa zea populations on cotton and non-cotton crop hosts. Entomologia Experimentalis Et Applicata, 126, pp 89-106
<Go to ISI>://WOS:000252442900001 AND http://www.botanischergarten.ch/Bt/Jackson-Regional-Assessment-2008.pdf
Jackson, R.E., Bradley, J.R., & Van Duyn, J.W. (2004a)
Performance of feral and Cry1Ac-selected Helicoverpa zea (Lepidoptera : Noctuidae) strains on transgenic cottons expressing one or two Bacillus thuringiensis ssp kurstaki proteins under greenhouse conditions. Journal of Entomological Science, 39, 1, pp 46-55
<Go to ISI>://WOS:000220233100005 AND NEBIS 20080426 Jackson, R.E., Bradley, J.R., Van Duyn, J.W., & Gould, F. (2004b)
Comparative production of Helicoverpa zea (Lepidoptera : Noctuidae) from transgenic cotton expressing either one or two Bacillus thuringiensis proteins with and without insecticide oversprays. Journal of Economic Entomology, 97, 5, pp 1719-1725
<Go to ISI>://WOS:000224653200031 AND http://www.botanischergarten.ch/Bt/Jackson-Comparative-Production-2004.pdf
James, C. (2007)
Global Status of Commercialized Biotech/GM Crops: ISAAA Brief 37-2007: Executive Summary ISAAA pp 16 (Report)
http://www.isaaa.org/resources/publications/briefs/37/executivesummary/default.html James, C. (2009)
Global Status of Commercialized Biotech/GM Crops: 2008, Brief 39, Executive Summary, Vol. Brief 39, pp. 20. ISAAA
http://www.botanischergarten.ch/ISAAA/ISAAA-Briefs-39-Executive-Summary.pdf AND http://www.botanischergarten.ch/ISAAA/Brief39Slides-2008.pdf
Jenkins, J.L. & Dean, D.H. (2001)
Binding specificity of Bacillus thuringiensis Cry1Aa for purified, native Bombyx mori aminopeptidase N and cadherin-like receptors. BMC Biochemistry, 2, 12 Cited April 23, 2002, pp 1-8
<Go to ISI>://BIOSIS:PREV200200410108 Johnson, D.E., Oppert, B., & McGaughey, W.H. (1998)

75
Spore coat protein synergizes Bacillus thuringiensis crystal toxicity for the indianmeal moth (Plodia interpunctella). Current Microbiology, 36, 5, pp 278-282
<Go to ISI>://WOS:000073040700006 AND http://www.botanischergarten.ch/Bt/Johnson-Spore-Coat-Protein-1998.pdf
Jouzani, G.S., Abad, A.P., Seifinejad, A., Marzban, R., Kariman, K., & Maleki, B. (2008)
Distribution and diversity of Dipteran-specific cry and cyt genes in native Bacillus thuringiensis strains obtained from different ecosystems of Iran. Journal of Industrial Microbiology & Biotechnology, 35, 2, pp 83-94
<Go to ISI>://WOS:000252192100002 AND http://www.botanischergarten.ch/Bt/Jouzani-Distribution-Diversity-2008.pdf
Jurat-Fuentes, J.L. & Adang, M.J. (2000)
Importance of Cry 1 delta-endotoxins Domain II loops for binding specificity of heliothis virescens l. Abstracts of Papers of the American Chemical Society, 219, pp U67-U67
<Go to ISI>://000087246100242 Jurat-Fuentes, J.L. & Adang, M.J. (2001)
Importance of Cry1 delta-endotoxin domain II loops for binding specificity in Heliothis virescens (L.). Applied and Environmental Microbiology, 67, 1, pp 323-329
<Go to ISI>://000166151300042 Jurat-Fuentes, J.L. & Adang, M.J. (2006)
Cry toxin mode of action in susceptible and resistant Heliothis virescens larvae. Journal of Invertebrate Pathology, 92, 3, pp 166-171
<Go to ISI>://000239198300009 AND http://www.botanischergarten.ch/Bt/Jurat-Fuentes-Mode-of-Action-2006.pdf
Jurat-Fuentes, J.L. & Adang, M.J. (2007)
A proteomic approach to study Cry1Ac binding proteins and their alterations in resistant Heliothis virescens larvae. Journal of Invertebrate Pathology, 95, 3, pp 187-191
<Go to ISI>://000247850700007 AND http://www.botanischergarten.ch/Bt/Jurat-Fuentes-Proteomic-Resistance-2007.pdf
Jurat-Fuentes, J.L., Gahan, L.J., Gould, F.L., Heckel, D.G., & Adang, M.J. (2004)
The HevCaLP protein mediates binding specificity of the Cry1A class of Bacillus thuringiensis toxins in Heliothis virescens. Biochemistry, 43, 44, pp 14299-14305
<Go to ISI>://000224939400046 AND http://www.botanischergarten.ch/Bt/Jurat-Fuentes-HevCaLP-Binding-2004.pdf
Karlova, R., Weemen-Hendriks, M., Naimov, S., Ceron, J., Dukiandjiev, S., & de Maagd, R.A. (2005)
Bacillus thuringiensis delta-endotoxin Cry1Ac domain III enhances activity against Heliothis virescens in some, but not all Cry1-Cry1Ac hybrids. Journal of Invertebrate Pathology, 88, 2, pp 169-172
<Go to ISI>://WOS:000227921600011 AND http://www.botanischergarten.ch/Bt/Karlova-Bacillus-thuringiensis-2005.pdf
Karumbaiah, L., Oppert, B., Jurat-Fuentes, J.L., & Adang, M.J. (2007)
Analysis of midgut proteinases from Bacillus thuringiensis-susceptible and -resistant Heliothis virescens (Lepidoptera : Noctuidae). Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology, 146, 1, pp 139-146
<Go to ISI>://000244623600018 AND http://www.botanischergarten.ch/Bt/Karumbajah-Analysis-Midgut-Proteinases-2006.pdf
Kaur, S. & Singh, A. (2000)
Natural occurrence of Bacillus thuringiensis in leguminous phylloplanes in the New Delhi region of India. World Journal of Microbiology & Biotechnology, 16, 7, pp 679-682
<Go to ISI>://WOS:000166367500018 AND http://www.botanischergarten.ch/Bt/Kaur-Natural-Occurrence-Bt-India-2001.pdf
Keeton, T.P. & Bulla, L.A. (1997)
Ligand specificity and affinity of BT-R-1, the Bacillus thuringiensis Cry1A toxin receptor from Manduca sexta, expressed in mammalian and insect cell cultures. Applied and Environmental Microbiology, 63, 9, pp 3419-3425
<Go to ISI>://WOS:A1997XV51000014

76
Kershen, D.L. (2006)
Health and food safety: The benefits of Bt-corn. Food and Drug Law Journal, 61, 2, pp 197-235 <Go to ISI>://WOS:000238696000002 AND
http://jay.law.ou.edu/faculty/kershen/articles/healthandfoodsafety.pdf AND http://www.botanischergarten.ch/Bt/Kershen-Bt-Food-2006.pdf AND Italian translation http://www.botanischergarten.ch/Bt/Kershen-Bt-Food-italiano-2007.pdf
King, G.F. (2007)
Modulation of insect Cav channels by peptidic spider toxins. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 513-530
http://www.sciencedirect.com/science/article/B6TCS-4MFJ2GX-4/2/3d936f0bfa0be6360b2a22cadf7485df AND http://www.botanischergarten.ch/Bt/King-Modulation-Spider-Toxins-2007.pdf
Klein, T.M., Wolf, E.D., Wu, R., & Sanford, C. (1987)
High-velocity microprojectiles for delivering nucleic acids into living cells. Nature, 327, 6117, pp 70-73 http://dx.doi.org/10.1038/327070a0 AND http://www.botanischergarten.ch/Bt/Klein-High-velocity-
Micropojectiles-1987.pdf Knowles, B.H. & Ellar, D.J. (1987)
Colloid-Osmotic Lysis Is a General Feature of the Mechanism of Action of Bacillus-Thuringiensis Delta-Endotoxins with Different Insect Specificity. Biochimica Et Biophysica Acta, 924, 3, pp 509-518
<Go to ISI>://WOS:A1987J062400017 AND http://www.botanischergarten.ch/Bt/Knowles-Colloid-Osmotic-Lysis-1987.pdf
Koziel, M.G., Beland, G.L., Bowman, C., Carozzi, N.B., Crenshaw, R., Crossland, L., Dawson, J., Desai, N., Hill, M., Kadwell, S., Launis, K., Lewis, K., Maddox, D., McPherson, K., Meghji, M.R., Merlin, E., Rhodes, R., Warren, G.W., Wright, M., & Evola, S.V. (1993)
Field Performance of Elite Transgenic Maize Plants Expressing an Insecticidal Protein Derived from Bacillus-Thuringiensis. Bio-Technology, 11, 2, pp 194-200
<Go to ISI>://WOS:A1993KV06200024 AND http://www.botanischergarten.ch/Bt/Koziel-Field-Performnce-Bt-1993.pdf
Koziel, M.G., Carozzi, N.B., & Desai, N. (1996)
Optimizing expression of transgenes with an emphasis on post-transcriptional events. Plant Molecular Biology, 32, 1-2, pp 393-405
<Go to ISI>://WOS:A1996VY81900019 Krieg, A. (1968)
A Taxonomic Study of Bacillus Thuringiensis Berliner. Journal of Invertebrate Pathology, 12, 3, pp 366-& <Go to ISI>://A1968C654500019 NOT IN EBZ, NOT IN NEBIS Krieg, A., Huger, A.M., Langenbruch, G.A., & Schnetter, W. (1983)
Bacillus-Thuringiensis Var Tenebrionis, a New Pathotype Effective against Larvae of Coleoptera. Zeitschrift Fur Angewandte Entomologie-Journal of Applied Entomology, 96, 5, pp 500-508
<Go to ISI>://WOS:A1983RW83900013 NOT IN EBZ, NOT IN NEBIS Krishnamoorthy, M., Jurat-Fuentes, J.L., McNall, R.J., Andacht, T., & Adang, M.J. (2007)
Identification of novel CrylAc binding proteins in midgut membranes from Heliothis virescens using proteomic analyses. Insect Biochemistry and Molecular Biology, 37, 3, pp 189-201
<Go to ISI>://000244810200001 Krogh, P.H. & Griffiths, B. (2007)
ECOGEN-Soil ecological and economic evaluation of genetically modified crops, SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 171-173
http://www.sciencedirect.com/science/article/B7CW5-4NTHKYM-2/2/e0f4e52a6128e3b2d70f15a8bf816276 AND http://www.botanischergarten.ch/Bt/Krogh-ECOGEN-intro-2007.pdf
Krogh, P.H., Griffiths, B., Demsar, D., Bohanec, M., Debeljak, M., Andersen, M.N., Sausse, C., Birch, A.N.E., Caul, S., Holmstrup, M., & Heckmann, L.H. (2007)
Responses by earthworms to reduced tillage in herbicide tolerant maize and Bt maize cropping systems SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 219-227

77
http://www.sciencedirect.com/science/article/B7CW5-4NNPB70-4/2/25971e5aa6055a8cc566f7621076b1eb AND http://www.botanischergarten.ch/Bt/Krogh-Earthworms-ECOGEN-2007.pdf
Lacroix, B., Tzfira, T., Vainstein, A., & Citovsky, V. (2006)
A case of promiscuity: Agrobacterium's endless hunt for new partners. Trends in Genetics, 22, 1, pp 29-37 <Go to ISI>://WOS:000234909900006 AND http://www.botanischergarten.ch/Bt/Lacroix-Case-of-Promiscuity-
2006.pdf Lecadet, M.M., Frachon, E., Dumanoir, V.C., Ripouteau, H., Hamon, S., Laurent, P., & Thiery, I. (1999)
Updating the H-antigen classification of Bacillus thuringiensis. Journal of Applied Microbiology, 86, 4 %R doi:10.1046/j.1365-2672.1999.00710.x, pp 660-672
http://www.blackwell-synergy.com/doi/abs/10.1046/j.1365-2672.1999.00710.x AND http://www.botanischergarten.ch/Bt/Lecadet-Classification-Bt-1999.pdf
Lee, D.H., Cha, I.H., Woo, D.S., & Ohba, M. (2003a)
Microbial ecology of Bacillus thuringiensis: Fecal populations recovered from wildlife in Korea. Canadian Journal of Microbiology, 49, 7, pp 465-471
<Go to ISI>://WOS:000186118600007 AND http://www.botanischergarten.ch/Bt/Lee-Microbial-Ecology-Korea-2003.pdf
Lee, M.K., Walters, F.S., Hart, H., Palekar, N., & Chen, J.S. (2003b)
Mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab delta-endotoxin. Applied and Environmental Microbiology, 69, 8, pp 4648-4657
<Go to ISI>://WOS:000184672500040 Li, G.P., Wu, K.M., Gould, F., Feng, H.Q., He, Y.Z., & Guo, Y.Y. (2004)
Frequency of Bt resistance genes in Helicoverpa armigera populations from the Yellow River cotton-farming region of China. Entomologia Experimentalis Et Applicata, 112, 2, pp 135-143
<Go to ISI>://000223268800008 AND http://www.botanischergarten.ch/Bt/Li-Frequency-Resistance-Helicoverpa-2004.pdf
Li, H.R., Oppert, B., Higgins, R.A., Huang, F.N., Buschman, L.L., & Zhu, K.Y. (2005)
Susceptibility of Dipel-resistant and -susceptible Ostrinia nubilalis (Lepidoptera : Crambidae) to individual Bacillus thuringiensis protoxins. Journal of Economic Entomology, 98, 4, pp 1333-1340
<Go to ISI>://000231056100034 AND http://www.botanischergarten.ch/Bt/Li-Susceptibility-Dipel-Resistant-2005.pdf
Lorch, A. & Then, C. (2007)
How much Bt toxin genetically engineered MON810 maize plants actually produce? Bt concentration in field plants from Germany and Spain pp 27 (Report)
http://www.botanischergarten.ch/Bt/Lorch-Bt-toxin-MON806-2007.pdf Lorz, H., Baker, B., & Schell, J. (1985)
Gene transfer to cereal cells mediated by protoplast transformation. Molecular and General Genetics MGG, 199, 2, pp 178-182
http://dx.doi.org/10.1007/BF00330256 AND http://www.botanischergarten.ch/Bt/Lorz-Gene-Transfer-Cereal-1986.pdf
Luo, S.D., Wang, G.R., Liang, G.M., Wu, K.M., Bai, L.Y., Ren, X.G., & Guo, Y.Y. (2006)
Binding of three Cry1A toxins in resistant and susceptible strains of cotton bollworm (Helicoverpa armigera). Pesticide Biochemistry and Physiology, 85, 2, pp 104-109
<Go to ISI>://WOS:000238008900007 Luttrell, K.C., Luttrell, R.G., Ali, M.I., & Allen, K.C. (2006)
Variation in susceptibility of Arkansas populations of Helicoverpa zea to cypermethrin. Beltwide Cotton Conferences|Beltwide Cotton Conferences, pp 9 pp.
<Go to ISI>://INSPEC:9584726 Luttrell, R.G., Wan, L., & Knighten, K. (1999)
Variation in susceptibility of noctuid (Lepidoptera) larvae attacking cotton and soybean to purified endotoxin proteins and commercial formulations of Bacillus thuringiensis. Journal of Economic Entomology, 92, 1, pp 21-32
<Go to ISI>://000078588400003

78
Mae, W. (2001)
Electronic Source: Witness Statement for Defence of José Bove and Dominique Soulier, Institute of Science and Society Website
published by: ISIS http://www.i-sis.org.uk/Montpellierbrief.php AND http://www.botanischergarten.ch/Bt/Ho-Anthracis-Bove-
2001.pdf Manyangarirwa, W., Turnbull, M., McCutcheon, G.S., & Smith, J. (2006)
Gene pyramiding as a Bt resistance management strategy: How sustainable is this strategy? African Journal of Biotechnology, 5, 10, pp 781-785
<Go to ISI>://000237675000001 AND http://www.botanischergarten.ch/Bt/Manyangarirwa-Pyramiding-2006.pdf
Martin, P.A.W. & Travers, R.S. (1989)
Worldwide Abundance and Distribution of Bacillus-Thuringiensis Isolates. Applied and Environmental Microbiology, 55, 10, pp 2437-2442
<Go to ISI>://WOS:A1989AT48300002 AND http://www.botanischergarten.ch/Bt/Martin-Worldwide-Abundance-Bt-1989.pdf
May, M.J., Champion, G.T., Dewar, A.M., Qi, A.M., & Pidgeon, J.D. (2005)
Management of genetically modified herbicide-tolerant sugar beet for spring and autumn environmental benefit. Proceedings of the Royal Society B-Biological Sciences, 272, 1559, pp 111-119
<Go to ISI>://000226980800001 AND http://www.botanischergarten.ch/Farmscale/May-Beet-FSE-2005.pdf McClintock, J.T., Schaffer, C.R., & Sjoblad, R.D. (1995)
A Comparative Review of the Mammalian Toxicity of Bacillus Thuringiensis-Based Pesticides. Pesticide Science, 45, 2, pp 95-105
<Go to ISI>://A1995RZ98500001 McGaughey, W.H. (1994)
Problems of Insect Resistance to Bacillus-Thuringiensis. Agriculture Ecosystems & Environment, 49, 1, pp 95-102
<Go to ISI>://A1994NZ67100015 AND http://www.botanischergarten.ch/Bt/McGaughey-Problems-Resistance-1994.pdf
McGeoch, M.A. & Pringle, K.L. (2005)
Science and advocacy: the GM debate in South Africa. South African Journal of Science, 101, 1-2, pp 7-9 <Go to ISI>://000229423000004 AND http://www.botanischergarten.ch/Bt/McGeoch-Science-Avocacy-
2005.pdf Meher, S.M., Bodhankar, S.L., Arunkumar, Dhuley, J.N., Khodape, D.J., & Naik, S.R. (2002)
Toxicity studies of microbial insecticide Bacillus thuringiensis var. kenyae in rats, rabbits, and fish. International Journal of Toxicology, 21, 2, pp 99-105
<Go to ISI>://000174941600004 AND http://www.botanischergarten.ch/Bt/Meher-Toxicity-Studies-Bt-kenyae-2002.pdf
Mehlo, L., Gahakwa, D., Nghia, P.T., Loc, N.T., Capell, T., Gatehouse, J.A., Gatehouse, A.M.R., & Christou, P. (2005)
An alternative strategy for sustainable pest resistance in genetically enhanced crops. PNAS, 102, 22, pp 7812-7816
http://www.pnas.org/cgi/content/abstract/102/22/7812 and http://www.botanischergarten.ch/Bt/Mehlo-Atern-Bt-PNAS-2005.pdf
Metz, M., ed. (2004)
Bacillus Thuringiensis: A Cornerstone of Modern Agriculture (Hardcover), pp 242, Haworth Press; 1 edition (January 2004), IS: ISBN-10: 1560221089 ISBN-13: 978-1560221081
Miller, H. (1994a)
CLINTONS MALIGN NEGLECT BATTERS BIOTECHNOLOGY. Bio-Technology, 12, 6, pp 571-573 <Go to ISI>://WOS:A1994NP63700015 AND http://www.botanischergarten.ch/Regulation/Miller-Clintons-
Malign-Neglect-1994.pdf Miller, H. (1994b)
Us Must Rationalize Biotech Regulation. Bio-Technology, 12, 5, pp 441-442

79
<Go to ISI>://A1994NJ86300010 AND http://www.botanischergarten.ch/Regulation/Miller-US-Must-Rationalize-1994.pdf
Miller, H.I. (2007)
Biotech's defining moments. Trends in Biotechnology, 25, 2, pp 56-59 http://www.sciencedirect.com/science/article/B6TCW-4MK0HGK-1/2/409c558b4630f54cdc689f843e290a6d
AND http://www.botanischergarten.ch/Regulation/Miller-Regulation-2007.pdf Milner, R.J. (1994)
History of Bacillus-Thuringiensis. Agriculture Ecosystems & Environment, 49, 1, pp 9-13 <Go to ISI>://A1994NZ67100002 Mizuki, E., Maeda, M., Tanaka, R., Lee, D.W., Hara, M., Akao, T., Yamashita, S., Kim, H.S., Ichimatsu, T., & Ohba, M. (2001)
Bacillus thuringiensis: A common member of microflora in activated sludges of a sewage treatment plant. Current Microbiology, 42, 6, pp 422-425
<Go to ISI>://000168466100010 AND http://www.botanischergarten.ch/Bt/Mizuki-Bt-Microflora-Sludges-2001.pdf
Moar, W., Roush, R., Shelton, A., Ferré, J., MacIntosh, S., Lennoard, R., & Abel, C. (2008)
Field-evolved resistance to Bt toxins. Nature Biotechnology, 26, 10, pp 1072-1074 http://dx.doi.org/10.1038/nbt1008-1072 AND http://www.nature.com/nbt/journal/v26/n10/pdf/nbt1008-1072.pdf
AND http://www.botanischergarten.ch/Bt/Moar-Field-Evolved-Resistance-2008.pdf Moellenbeck, D.J., Peters, M.L., Bing, J.W., Rouse, J.R., Higgins, L.S., Sims, L., Nevshemal, T., Marshall, L., Ellis, R.T., Bystrak, P.G., Lang, B.A., Stewart, J.L., Kouba, K., Sondag, V., Gustafson, V., Nour, K., Xu, D.P., Swenson, J., Zhang, J., Czapla, T., Schwab, G., Jayne, S., Stockhoff, B.A., Narva, K., Schnepf, H.E., Stelman, S.J., Poutre, C., Koziel, M., & Duck, N. (2001)
Insecticidal proteins from Bacillus thuringiensis protect corn from corn rootworms. Nature Biotechnology, 19, 7, pp 668-672
<Go to ISI>://WOS:000169677200024 AND http://www.botanischergarten.ch/Bt/Moellenbeck-Insecticidal-Proteins-Rootworms-2001.pdf
Monnerat, R., Martins, E., Queiroz, P., Orduz, S., Jaramillo, G., Benintende, G., Cozzi, J., Real, M.D., Martinez-Ramirez, A., Rausell, C., Ceron, J., Ibarra, J.E., Del Rincon-Castro, M.C., Espinoza, A.M., Meza-Basso, L., Cabrera, L., Sanchez, J., Soberon, M., & Bravo, A. (2006)
Genetic variability of Spodoptera frugiperda Smith (Lepidoptera : Noctuidae) populations from Latin America is associated with variations in susceptibility to Bacillus thuringiensis cry toxins. Applied and Environmental Microbiology, 72, 11, pp 7029-7035
<Go to ISI>://WOS:000242003800021 AND http://www.botanischergarten.ch/Bt/Monnerat-Genetic-Variability-2006.pdf
Monnerat, R.G., Dias, D.G.S., da Silva, S.F., Martins, E.S., Berry, C., Falcao, R., Gomes, A., Praca, L.B., & Soares, C.M.S. (2005)
Screening of Bacillus thuringiensis strains effective against mosquitoes. Pesquisa Agropecuaria Brasileira, 40, 2, pp 103-106
<Go to ISI>://WOS:000228389700001 AND http://www.botanischergarten.ch/Bt/Gomes-Monnerat-Screening-Bt-2005.pdf
Monsanto Co. (2002)
Electronic Source: Monsanto biotechnology trait acreage. Fiscal years 1996 to 2007, published by: (Monsanto, St. Louis, updated October 10, 2007) http://www.monsanto.com/investors/ presentations.asp; scroll down to Biotech Acres under Fourth-Quarter
2007 Monsanto Company Earnings Conference Call). Munoz-Garay, C., Sanchez, J., Darszon, A., de Maagd, R.A., Bakker, P., Soberon, M., & Bravo, A. (2006)
Permeability changes of Manduca sexta midgut brush border membranes induced by oligomeric structures of different cry toxins. Journal of Membrane Biology, 212, 1, pp 61-68
<Go to ISI>://000243465600006 Naimov, S., Dukiandjiev, S., & de Maagd, R.A. (2003)
A hybrid Bacillus thuringiensis delta-endotoxin gives resistance against a coleopteran and a lepidopteran pest in transigenic potato. Plant Biotechnology Journal, 1, 1, pp 51-57
<Go to ISI>://WOS:000188993900006 IN EZB RED, NOT IN NEBIS

80
Nakanishi, K., Yaoi, K., Nagino, Y., Hara, H., Kitami, M., Atsumi, S., Miura, N., & Sato, R. (2002)
Aminopeptidase N isoforms from the midgut of Bombyx mori and Plutella xylostella - their classification and the factors that determine their binding specificity to Bacillus thuringiensis Cry1A toxin. Febs Letters, 519, 1-3, pp 215-220
<Go to ISI>://WOS:000175822100041 Nap, J.P., Metz, P.L.J., Escaler, M., & Conner, A.J. (2003)
The release of genetically modified crops into the environment - Part I. Overview of current status and regulations. Plant Journal, 33, 1, pp 1-18
<Go to ISI>://000180276400001 AND http://www.botanischergarten.ch/Bt/Nap-Release-Review-1-2003.pdf Nature-Editorial (1992)
Products pose no special risks just because of the processes used to make them. Nature, 356, 6364, pp 1-2 http://dx.doi.org/10.1038/356001b0 AND http://www.botanischergarten.ch/Regulation/Anonymous-US-
Regulation-Nature-1992.pdf Nature-Editorial (2007)
Directive Action Required. Editorial, Europe's handling of applications to grow genetically modified crops amounts to bad governance. Nature, 450, 7172, pp 921-921
http://dx.doi.org/10.1038/450921a AND http://www.botanischergarten.ch/Regulation/Nature-Editorial-Directive-action-required-2007.pdf
Navon, A. (2000)
Bacillus thuringiensis insecticides in crop protection - reality and prospects. Crop Protection, 19, 8-10, pp 669-676
<Go to ISI>://000165301100022 Nester, E., Thomashow, L.S., Metz, M., & Gordon, M. (2002)
100 YEARS OF Bacillus thuringiensis: A Critical Scientific Assessment, a Report for the American Academy of Microbiology, American Academy of Microbiology pp 22 This report is based on a colloquium, “100 Years of Bacillis thuringiensis, a Paradigm for Producing Transgenic Organisms: A Critical Scientific Assessment,” sponsored by the American Academy of Microbiology and held November 16-18, in Ithaca, New York Washington (Report)
http://www.asm.org/ASM/files/CCPAGECONTENT/docfilename/0000003782/Btreport[1].pdf Nevels, K. & Butko, P. (2007)
Lipid specificity of the Bacillus thuringiensis toxin/membrane interactions. Faseb Journal, 21, 6, pp A982-A982
<Go to ISI>://WOS:000245708702057 Nicholson, G. (2007a)
Fighting the global pest problem: Preface to the special Toxicon issue on insecticidal toxins and their potential for insect pest control. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 413-422
http://www.sciencedirect.com/science/article/B6TCS-4MH2991-2/2/342619491f36e9b7073b037ce45f8434 AND http://www.botanischergarten.ch/Bt/Nicholson-Fighting-Global-Pest-Problem-2007.pdf
Nicholson, G.M. (2007b)
Insect-selective spider toxins targeting voltage-gated sodium channels. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 490-512
http://www.sciencedirect.com/science/article/B6TCS-4MH2991-1/2/6c2c892cea0c2cbc59fd0ba4af98f584 AND http://www.botanischergarten.ch/Bt/Nicholson-Insect-selective-Spider-2007.pdf
Noteborn, H., BienenmannPloum, M.E., vandenBerg, J.H.J., Alink, G.M., Zolla, L., Reynaerts, A., Pensa, M., & Kuiper, H.A. (1995)
Safety assessment of the Bacillus thuringiensis insecticidal crystal protein CRYIA(b) expressed in transgenic tomatoes. In Genetically Modified Foods, pp. 134-147. AMER CHEMICAL SOC, WASHINGTON Acs Symposium Series
605 <Go to ISI>://A1995BF66M00012 Obrycki, J.J., Losey, J.E., Taylor, O.R., & Jesse, L.C.H. (2001)
Transgenic insecticidal corn: Beyond insecticidal toxicity to ecological complexity. Bioscience, 51, 5, pp 353-361

81
<Go to ISI>://000169293900008 AND http://www.botanischergarten.ch/Bt/Obrycki-Transgenic-Beyond-2001.pdf
Obukowicz, M.G., Perlak, F.J., Kusanokretzmer, K., Mayer, E.J., Bolten, S.L., & Watrud, L.S. (1986)
Tn5-Mediated Integration of the Delta-Endotoxin Gene from Bacillus-Thuringiensis into the Chromosome of Root-Colonizing Pseudomonads. Journal of Bacteriology, 168, 2, pp 982-989
<Go to ISI>://WOS:A1986E687900075 AND http://www.botanischergarten.ch/Bt/Obukowicz-Integration-EndotoxinBt-1986.pdf
OECD Consensus Documents (2007)
Consensus Document on Safety Information on Transgenic Plants Expressing Bacillus thuringiensis - Derived Insect Control Protein, OECD pp 109 OECD Environment, Health and Safety Publications Paris, France (Report)
http://www.olis.oecd.org/olis/2007doc.nsf/LinkTo/NT00002DF6/$FILE/JT03230592.PDF AND http://www.botanischergarten.ch/Bt/OECD-Consensus-Bt-crops-2007.pdf
Oestergaard, J., Ehlers, R.U., Martinez-Ramirez, A.C., & Real, M.D. (2007)
Binding of Cyt1Aa and Cry11Aa toxins of Bacillus thuringiensis serovar israelensis to brush border membrane vesicles of Tipula paludosa (Diptera : Nematocera) and subsequent pore formation. Applied and Environmental Microbiology, 73, 11, pp 3623-3629
<Go to ISI>://WOS:000247016600021 AND http://www.botanischergarten.ch/Bt/Oestergaard-Binding-Cyt1Aa-Bt-Pore-2007.pdf
Ohba, M. & Aizawa, K. (1986a)
Distribution of Bacillus-Thuringiensis in Soils of Japan. Journal of Invertebrate Pathology, 47, 3, pp 277-282 <Go to ISI>://WOS:A1986C223800005 Ohba, M. & Aizawa, K. (1986b)
Insect Toxicity of Bacillus-Thuringiensis Isolated from Soils of Japan. Journal of Invertebrate Pathology, 47, 1, pp 12-20
<Go to ISI>://WOS:A1986AZK5400003 Ohba, M. & Aratake, Y. (1994)
Comparative-Study of the Frequency and Flagellar Serotype Flora of Bacillus-Thuringiensis in Soils and Silkworm-Breeding Environments. Journal of Applied Bacteriology, 76, 3, pp 203-209
<Go to ISI>://WOS:A1994MY79300001 Okumoto, S., Koch, W., Tegeder, M., Fischer, W.N., Biehl, A., Leister, D., Stierhof, Y.D., & Frommer, W.B. (2004)
Root phloem-specific expression of the plasma membrane amino acid proton co-transporter AAP3. Journal of Experimental Botany, 55, 406, pp 2155-2168
<Go to ISI>://WOS:000224081500003 AND http://www.botanischergarten.ch/Bt/Okumoto-Root-Phloem-2004.pdf AND supplements http://jxb.oxfordjournals.org/cgi/content/full/erh233/DC1
Okumura, S., Akao, T., Mizuki, E., Ohba, M., & Inouye, K. (2001)
Screening of the Bacillus thuringiensis Cry1Ac delta-endotoxin on the artificial phospholipid monolayer incorporated with brush border membrane vesicles of Plutella xylostella by optical biosensor technology. Journal of Biochemical and Biophysical Methods, 47, 3, pp 177-188
<Go to ISI>://WOS:000167681700002 AND http://www.botanischergarten.ch/Bt/Okumura-Screening-Bt-2001.pdf
Onishi, M., Matsuoka, T., Kodama, T., Kashiwaba, K., Futo, S., Akiyama, H., Maitani, T., Furui, S., Oguchi, T., & Hino, A. (2005)
Development of a multiplex polymerase chain reaction method for simultaneous detection of eight events of genetically modified maize. Journal of Agricultural and Food Chemistry, 53, 25, pp 9713-9721
<Go to ISI>://WOS:000233891700011 AND http://www.botanischergarten.ch/Bt/Onishi-Simultaneous-Detection-2005.pdf
Pardo-Lopez, L., Gomez, I., Munoz-Garay, C., Jimenez-Juarez, N., Soberon, M., & Bravo, A. (2006)
Structural and functional analysis of the pre-pore and membrane-inserted pore of Cry1Ab toxin. Journal of Invertebrate Pathology, 92, 3, pp 172-177
<Go to ISI>://000239198300010 Parrott, W. (2008)

82
Study of Bt impact on caddisflies overstates its conclusions: Response to Rosi-Marshall et al. Proceedings of the National Academy of Sciences, pp E10
http://www.pnas.org AND http://www.botanischergarten.ch/Bt/Parrott-Rosi-Marshall-2008.pdf Pearce, M., Habbick, B., Williams, J., Eastman, M., & Newman, M. (2002)
The effects of aerial spraying with Bacillus thuringiensis Kurstaki on children with asthma. Canadian Journal of Public Health-Revue Canadienne De Sante Publique, 93, 1, pp 21-25
<Go to ISI>://WOS:000174508500006 Pedersen, J.C., Damgaard, P.H., Eilenberg, J., & Hansen, B.M. (1995)
Dispersal of Bacillus-Thuringiensis Var Kurstaki in an Experimental Cabbage Field. Canadian Journal of Microbiology, 41, 2, pp 118-125
<Go to ISI>://WOS:A1995QM34600002 AND http://www.botanischergarten.ch/Bt/Pedersen-Dispersal-Bt-Cabbage-1995.pdf
Peferoen, M. (1997)
Progress and prospects for field use of Bt genes in crops. Trends in Biotechnology, 15, 5, pp 173-177 <Go to ISI>://A1997WY03200006 AND http://www.botanischergarten.ch/Bt/Peferoen-Progress-and-Prospect-
1997.pdf Perry, J.N., Firbank, L.G., Champion, G.T., Clark, S.J., Heard, M.S., May, M.J., Hawes, C., Squire, G.R., Rothery, P., Wolwod, I.P., & Pidgeon, J.D. (2004)
Ban on triazine herbicides likely to reduce but not negate relative benefits of GMHT maize cropping. Nature, 428, 6980, pp 313-316
<Go to ISI>://000220250200042 and http://www.botanischergarten.ch/Farmscale/Perry-et-al-Nature-0404.pdf Perry, J.N., Rothery, P., Clark, S.J., Heard, M.S., & Hawes, C. (2003)
Design, analysis and statistical power of the Farm-Scale Evaluations of genetically modified herbicide-tolerant crops. Journal of Applied Ecology, 40, 1, pp 17-31
<Go to ISI>://000180852600002 and http://www.botanischergarten.ch/Farmscale/Perry-Design-Farm-Scale-2003.pdf
Peterson, P.A. & Bianchi, A., eds. (1999)
Maize Genetics and Breeding in the 20th Century, pp 379, IS: ISBN-10: 981022866X ISBN-13: 978-9810228668
Petrie, K., Thomas, M., & Broadbent, E. (2003)
Symptom complaints following aerial spraying with biological insecticide Foray 48B. New Zealand Medical Journal, 116, 1170 Cited March 14, 2003, pp
<Go to ISI>://BIOSIS:PREV200300250079 AND http://www.botanischergarten.ch/Bt/Petrie-Symptom-Complaints-2003.pdf
Pilcher, C.D., Rice, M.E., Higgins, R.A., Steffey, K.L., Hellmich, R.L., Witkowski, J., Calvin, D., Ostlie, K.R., & Gray, M. (2002)
Biotechnology and the European corn borer: Measuring historical farmer perceptions and adoption of transgenic Bt corn as a pest management strategy. Journal of Economic Entomology, 95, 5, pp 878-892
<Go to ISI>://000178550000002 AND http://www.botanischergarten.ch/Bt/Pilcher-Bt-Corn-Perception-2002.pdf
Pimentel, D.S. & Raven, P.H. (2000)
Bt corn pollen impacts on nontarget Lepidoptera: Assessment of effects in nature. Proceedings of the National Academy of Sciences of the United States of America, 97, 15, pp 8198-8199
<Go to ISI>://WOS:000088273900003 AND http://www.botanischergarten.ch/Bt/Pimentel-Impacts-nontarget-Lepidoptera-2000.pdf
Potrykus, I., Paszkowski, J., Saul, M.W., Petruska, J., & Shillito, R.D. (1985)
Molecular and General Genetics of a Hybrid Foreign Gene Introduced into Tobacco by Direct Gene-Transfer. Molecular & General Genetics, 199, 2, pp 169-177
<Go to ISI>://WOS:A1985AJE6900002 AND http://www.botanischergarten.ch/Bt/Potrykus-Molecular-General-Genetics-1985.pdf
Potvin, L., Laprade, R., & Schwartz, J.L. (1998)

83
Cry1Ac, a Bacillus thuringiensis toxin, triggers extracellular Ca2+ influx and Ca2+ release from intracellular stores in Cf1 cells (Choristoneura fumiferana, Lepidoptera). Journal of Experimental Biology, 201, 12, pp 1851-1858
<Go to ISI>://WOS:000074602300001 AND http://www.botanischergarten.ch/Bt/Potvin-Extracellular-Influx-1998.pdf
Priest, F.G. (1993) Systematics and Ecology of Bacillus and other Gram positive Bacteria. In Biochemistry, Physiology, and
Molecular Genetics, (eds A.L. Sonenshein, J.A. Hoch & R. Losick), pp. 3-16. ASM, Washington DC.
Punja, Z.K. (1997)
Comparative efficacy of bacteria, fungi, and yeasts as biological control agents for diseases of vegetable crops. Canadian Journal of Plant Pathology-Revue Canadienne De Phytopathologie, 19, 3, pp 315-323
<Go to ISI>://A1997YJ03300014 Ramjoue, C. (2007a)
The transatlantic rift in genetically modified food policy. Doctoral Thesis, University of Zurich, Zurich Thesis, pp 263
http://www.botanischergarten.ch/Regulation/Ramjoue-Thesis-Transatlantic-Rift-2007.pdf Ramjoue, C. (2007b)
The transatlantic rift in genetically modified food policy. Journal of Agricultural & Environmental Ethics, 20, 5, pp 419-436
<Go to ISI>://WOS:000248855000003 AND http://www.botanischergarten.ch/Regulation/Ramjoue-Transatlantic-Rift-2007.pdf
Rang, C., Bergvingson, D., Bohorova, N., Hoisington, D., & Frutos, R. (2004)
Competition of Bacillus thuringiensis Cry1 toxins for midgut binding sites: A basis for the development and management of transgenic tropical maize resistant to several stemborers. Current Microbiology, 49, 1, pp 22-27
<Go to ISI>://WOS:000222607500005 AND http://www.botanischergarten.ch/Bt/Rang-Tropical-Maize-2004.pdf
Rang, C., Vachon, V., de Maagd, R.A., Villalon, M., Schwartz, J.L., Bosch, D., Frutos, R., & Laprade, R. (1999)
Interaction between functional domains of Bacillus thuringiensis insecticidal crystal proteins. Applied and Environmental Microbiology, 65, 7, pp 2918-2925
<Go to ISI>://000081199000017 AND http://www.botanischergarten.ch/Bt/Rang-Interactions-Functional-1999.pdf
Rasko, D.A., Altherr, M.R., Han, C.S., & Ravel, J. (2005)
Genomics of the Bacillus cereus group of organisms. Fems Microbiology Reviews, 29, 2, pp 303-329 <Go to ISI>://WOS:000228506000009 AND http://www.botanischergarten.ch/Bt/Rasko-Genomics-Bacillus-
Cereus-2005.pdf Rasko, D.A., Rosovitz, M.J., Okstad, O.A., Fouts, D.E., Jiang, L.X., Cer, R.Z., Kolsto, A.B., Gill, S.R., & Ravel, J. (2007)
Complete sequence analysis of novel plasmids from emetic and periodontal Bacillus cereus isolates reveals a common evolutionary history among the B-cereus-group plasmids, including Bacillus anthracis pXO1. Journal of Bacteriology, 189, 1, pp 52-64
<Go to ISI>://WOS:000243182500004 AND http://www.botanischergarten.ch/Bt/Rasko-Complete-Sequence-B-Cereus-2007.pdf
Rausell, C., De Decker, N., Garcia-Robles, I., Escriche, B., Van Kerkhove, E., Real, M.D., & Martinez-Ramirez, A.C. (2000a)
Effect of Bacillus thuringiensis toxins on the midgut of the nun moth Lymantria monacha. Journal of Invertebrate Pathology, 75, 4, pp 288-291
<Go to ISI>://WOS:000087759000007 AND http://www.ask-force.org/web/Bt1/Rausell-Effect-Bacillus-Toxins-2000.pdf
Rausell, C., Garcia-Robles, I., Sanchez, J., Munoz-Garay, C., Martinez-Ramirez, A.C., Real, M.D., & Bravo, A. (2004a)
Role of toxin activation on binding and pore formation activity of the Bacillus thuringiensis Cry3 toxins in membranes of Leptinotarsa decemlineata (Say). Biochimica Et Biophysica Acta-Biomembranes, 1660, 1-2, pp 99-105

84
<Go to ISI>://WOS:000188707400012 AND http://www.ask-force.org/web/Bt1/Rausell-Role-Toxin-Activation-2004.pdf
Rausell, C., Martinez-Ramirez, A.C., Garcia-Robles, I., & Real, M.D. (2000b)
A binding site for Bacillus thuringiensis Cry1Ab toxin is lost during larval development in two forest pests. Applied and Environmental Microbiology, 66, 4, pp 1553-1558
<Go to ISI>://WOS:000086284700046 Rausell, C., Munoz-Garay, C., Miranda-CassoLuengo, R., Gomez, I., Rudino-Pinera, E., Soberon, M., & Bravo, A. (2004b)
Tryptophan spectroscopy studies and black lipid bilayer analysis indicate that the oligomeric structure of Cry1Ab toxin from Bacillus thuringiensis is the membrane-insertion intermediate. Biochemistry, 43, 1, pp 166-174
<Go to ISI>://WOS:000187894800018 Rausell, C., Pardo-Lopez, L., Sanchez, J., Munoz-Garay, C., Morera, C., Soberon, M., & Bravo, A. (2004c)
Unfolding events in the water-soluble monomeric Cry1Ab toxin during transition to oligomeric pre-pore and membrane-inserted pore channel. Journal of Biological Chemistry, 279, 53, pp 55168-55175
<Go to ISI>://000225960800024 Read, T.D., Salzberg, S.L., Pop, M., Shumway, M., Umayam, L., Jiang, L.X., Holtzapple, E., Busch, J.D., Smith, K.L., Schupp, J.M., Solomon, D., Keim, P., & Fraser, C.M. (2002)
Comparative genome sequencing for discovery of novel polymorphisms in Bacillus anthracis. Science, 296, 5575, pp 2028-2033
<Go to ISI>://WOS:000176273300054 AND http://www.botanischergarten.ch/Bt/Read-Genome-Antracis-2002.pdf
Rhodes, C.A., Pierce, D.A., Mettler, I.J., Mascarenhas, D., & Detmer, J.J. (1988)
Genetically transformed maize plants from protoplasts. Science, 240, 4849, pp 204-207 http://www.sciencemag.org/cgi/content/abstract/240/4849/204 AND
http://www.botanischergarten.ch/Bt/Rhodes-Genetically-Modified-Maize-1988.pdf Rie, v.J., Jansens, S., Hofte, H., Degheele, D., & Mellaert, H. (1989)
Specificity of Bacillus thuringiensis δ-endotoxins. Importance of specific receptors on the brush border membrane of the mid-gut of target insects. European Journal of Biochemistry, 186, 1-2 %R doi:10.1111/j.1432-1033.1989.tb15201.x, pp 239-247
http://www.blackwell-synergy.com/doi/abs/10.1111/j.1432-1033.1989.tb15201.x AND http://www.botanischergarten.ch/Bt/VanRie-SpecifityBt-Endotoxins-1989.pdf
Rohou, A., Nield, J., & Usharyov, Y.A. (2007)
Insecticidal toxins from black widow spider venom. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 531-549
http://www.sciencedirect.com/science/article/B6TCS-4MG0641-2/2/3c961fd8a9044167c4713780e7123a7a AND http://www.botanischergarten.ch/Bt/Rohou-Insecticidal-Black-Widow-2007.pdf
Romeis, J., Bartsch, D., Bigler, F., Candolfi, M., Gielkens, M.C., Hartley, S.E., Hellmich, R., Huesing, J.E., Jepson, P.C., Layton, R.J., Quemada, H., Raybould, A., Rose, R., Schiemann, J., Sears, M.K., Shelton, M., Sweet, J., Vaituzis, Z., & Wolt, J.D. (2008a)
Assessment of risk of insect-resistant transgenic crops to nontarget arthropods. Nature Biotechnology, 26, 2, pp 203-208
http://dx.doi.org/10.1038/nbt1381 AND http://www.nature.com/nbt/journal/v26/n2/suppinfo/nbt1381_S1.html AND http://www.botanischergarten.ch/Bt/Romeis-Nontarget-2008.pdf
Romeis, J., Detlef Bartsch2, Franz Bigler1, Marco P. Candolfi3, Marco M.C. Gielkens4, Susan E. Hartley5, Richard L. Hellmich6, Joseph E. Huesing7, Paul C. Jepson8,, Raymond Layton9, H.Q., Alan Raybould11, Robyn I. Rose12,, Joachim Schiemann13, M.K.S., Anthony M. Shelton15, Jeremy Sweet16,, & Zigfridas Vaituzis17, J.D.W. (2007)
Moving Through the Tiered and Methodological Framework for Non-Target Arthropod Risk Assessment of Transgenic Insecticidal Crops, Jeju Island, Korea, Proceedings of the 9th International Symposium on the Biosafety of Genetically Modified Organisms, September 24-29, 2006 Ed. pp 62-67
http://www.botanischergarten.ch/Nuetzlinge/Romeis-NontargetISBGMO-2006.pdf Romeis, J., Dutton, A., & Bigler, F. (2004)

85
Bacillus thuringiensis toxin (Cry1Ab) has no direct effect on larvae of the green lacewing Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). Journal of Insect Physiology, 50, 2-3, pp 175-183
http://www.sciencedirect.com/science/article/B6T3F-4B84T3T-1/2/041f2a2f6f5410dd5ec2748a70707ec6 or http://www.botanischergarten.ch/Bt/Romeis-et-al-04-Chrysoperla.pdf and evaluation F1000: http://www.facultyof1000.com/article/15019519/evaluation
Romeis, J., Shelton, A., & Kennedy, G.G., eds. (2008b)
Integration of Insect-Resistant Genetically Modified Crops within IPM Programs, pp ca. 400, Springer.com, Heidelberg, IS: 978-1-4020-8372-3
Romeis, J., Shelton, A., & Kennedy, G.G., eds. (2008c)
Integration of Insect-Resistant Genetically Modified Crops within IPM Programs, pp 441, Springer, Hamburg, IS: ISBN-10: 1402083726 ISBN-13: 978-1402083723
http://www.amazon.com/gp/reader/1402083726/ref=sib_dp_pt#reader-link Rongsen, L., Shun!-ing, D., Xiaogang, L., Cheng, L., Zhumei, S., & \ling, S. (1990)
Survey of Bacillus turingiensis and Bacillus sphaericus from soils of four provinces of China and their principal biological properties. Acta Microbiologica Sinica, 30, pp 380-388
http://www.cqvip.com/QK/94144X/1990005/378316.html Rosati, A., Bogani, P., Santarlasci, A., & Buiatti, M. (2008)
Characterisation of 3 ' transgene insertion site and derived mRNAs in MON810 YieldGard (R) maize. Plant Molecular Biology, 67, 3, pp 271-281
<Go to ISI>://WOS:000255414000006 AND http://www.botanischergarten.ch/Bt/Rosati-Characterisation-Mon810-publ-2008.pdf
Rosi-Marshall, E.J., Tank, J.L., Royer, T.V., & Whiles, M.R. (2008)
Reply to Beachy et al. and Parrott: Study indicates Bt corn may affect caddisflies. Proceedings of the National Academy of Sciences, 105, 7, pp E11-E11
http://www.pnas.org/content/105/7/E11.short AND http://www.botanischergarten.ch/Bt/Rosi-Marschall-Bt-Aquatic-reply-2008.pdf
Rosi-Marshall, E.J., Tank, J.L., Royer, T.V., Whiles, M.R., Evans-White, M., Chambers, C., Griffiths, N.A., Pokelsek, J., & Stephen, M.L. (2007a)
Toxins in transgenic crop byproducts may affect headwater stream ecosystems. Proceedings of the National Academy of Sciences, electronic prepublication, pp --- ---
http://www.pnas.org/cgi/content/abstract/0707177104v2 AND http://www.botanischergarten.ch/Bt/Rosi-Marschall-Bt-Aquatic-2007.pdf
Rosi-Marshall, E.J., Tank, J.L., Royer, T.V., Whiles, M.R., Evans-White, M., Chambers, C., Griffiths, N.A., Pokelsek, J., & Stephen, M.L. (2007b)
Toxins in transgenic crop byproducts may affect headwater stream ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 104, pp 16204-16208
<Go to ISI>://WOS:000250128800043 10.1073/pnas.0707177104 AND http://www.pnas.org/cgi/content/abstract/0707177104v2 AND http://www.botanischergarten.ch/Bt/Rosi-Marschall-Bt-Aquatic-2007.pdf
Sairam, R., Chennareddy, S., Parani, M., Zhang, S.L., Al-Abed, D., Abou-Alaiw, W., & Goldman, S. (2005)
OBPC Symposium: Maize 2004 & Beyond - Plant regeneration, gene discovery, and genetic engineering of plants for crop improvement. In Vitro Cellular & Developmental Biology-Plant, 41, 4, pp 411-423
<Go to ISI>://000231708100009 AND http://www.botanischergarten.ch/Bt/Sairam-OBPC-Symposium-Discovery-2005.pdf
Sanford, J.C. (2000)
The development of the biolistic process. In Vitro Cellular & Developmental Biology - Plant, 36, 5, pp 303-308 http://dx.doi.org/10.1007/s11627-000-0056-9 AND http://www.botanischergarten.ch/Bt/Sanford-Biolistic-
2000.pdf Sankula, S. & Blumenthal, E. (2004)
Impacts on US Agriculture of Biotechnology-Derived Crops Planted in 2003– An Update of Eleven Case Studies pp 92 (Report)
www.ncfap.org and http://www.botanischergarten.ch/Pesticides/Sankula-2004finalreport.pdf Sanvido, O. & Romeis, J. (2007)

86
Ecological Impacts of Genetically Modified Crops: Ten Years of Field Research and Commercial Cultivation. Adv Biochem Engin/Biotechnol, 107, pp 235–278
http://www.botanischergarten.ch/Environment/Sanvido-Ecological-Impacts-2007.pdf Sanvido, O., Stark, M., Romeis, J., & Bigler, F. (2006)
Ecological impacts of genetically modified crops, Experiences from ten years of experimental field research and commercial cultivation, Agroscope Reckenholz-Tänikon Research Station ART, Reckenholzstrasse 191, CH-8046 Zurich, Phone +41 (0)44 377 71 11, Fax +41 (0)44 377 72 01, [email protected], www.art.admin.ch. 1 pp 108 ART-Schriftenreihe 1 Zürich Reckenholz (Report)
http://www.botanischergarten.ch/Environment/Sanvido-Agroscope-2006.pdf Saxena, D., Flores, S., & Stotzky, G. (1999)
Transgenic plants - Insecticidal toxin in root exudates from Bt corn. Nature, 402, 6761, pp 480-480 http://www.botanischergarten.ch/Bt/Saxena-Stotzky-Nature-1999.pdf Saxena, D. & Stotzky, G. (2001)
Bacillus thuringiensis (Bt) toxin released from root exudates and biomass of Bt corn has no apparent effect on earthworms, nematodes, protozoa, bacteria, and fungi in soil. Soil Biology & Biochemistry, 33, 9, pp 1225-1230
www.elsevier.com/locate/soilbio and http://www.botanischergarten.ch/Bt/Saxena-Stotzky-2001.pdf Schell, J. (1977)
Transfer and Expression of Procaryotic DNA in Plant-Cells Transformed by Agrobacterium Tumefaciens. Hoppe-Seylers Zeitschrift Fur Physiologische Chemie, 358, 4, pp 422-423
<Go to ISI>://WOS:A1977DD74000016 AND NEBIS 20080603 Schnepf, E., Crickmore, N., Van Rie, J., Lereclus, D., Baum, J., Feitelson, J., Zeigler, D.R., & Dean, D.H. (1998)
Bacillus thuringiensis and its pesticidal crystal proteins. Microbiology and Molecular Biology Reviews, 62, 3, pp 775-+
<Go to ISI>://WOS:000075782400010 AND http://www.botanischergarten.ch/Bt/Schnepf-Crystal-Proteins-1998.pdf
Schnepf, H.E. & Whiteley, H.R. (1981)
Cloning and Expression of the Bacillus-Thuringiensis Crystal Protein Gene in Escherichia-Coli. Proceedings of the National Academy of Sciences of the United States of America Biological Sciences, 78, 5, pp 2893-2897
<Go to ISI>://WOS:A1981LS91000048 AND http://www.botanischergarten.ch/Bt/Schnepf-Cloning-Expression-1981.pdf
Scholte, E.J. & Dicke, M. (2005)
Effects of insect-resistant transgenic crops on non-target arthropods: first step in pre-market risk assessment studies, A literature-based study, proposing an ecologically
based first step to select non-target organisms, COGEM pp 59 (Report) http://www.botanischergarten.ch/Bt/Scholte-Effects-Select-2005.pdf Shah, D.M., Tumer, N.E., Fischhoff, D.A., Horsch, R.B., Rogers, S.G., Fraley, R.T., & Jaworski, E.G. (1987)
The Introduction and Expression of Foreign Genes in Plants. Biotechnology & Genetic Engineering Reviews, 5, pp 81-106
<Go to ISI>://WOS:A1987M496900003 AND http://www.botanischergarten.ch/Bt/Shah-Introducion-Expression-Foreign-1987.pdf
Sharma, H.C., Sharma, K.K., & Crouch, J.H. (2004)
Genetic transformation of crops for insect resistance: Potential and limitations. Critical Reviews in Plant Sciences, 23, 1, pp 47-72
<Go to ISI>://000220026200004 AND http://www.botanischergarten.ch/Bt/Sharma-Genetic-Tranformation-Potential-Limitations-2004.pdf
Shelton, A.M., Robertson, J.L., Tang, J.D., Perez, C., Eigenbrode, S.D., Preisler, H.K., Wilsey, W.T., & Cooley, R.J. (1993)
Resistance of Diamondback Moth (Lepidoptera, Plutellidae) to Bacillus-Thuringiensis Subspecies in the Field. Journal of Economic Entomology, 86, 3, pp 697-705
<Go to ISI>://A1993MA89500009 AND http://www.botanischergarten.ch/Bt/Shelton-Risistance-Diamondback-1993.pdf
Shimada, N., Miyamoto, K., Kanda, K., & Murata, H. (2006)

87
Binding of Cry1Ab toxin, a Bacillus thuringiensis insecticidal toxin, to proteins of the bovine intestinal epithelial cell: An in vitro study. Applied Entomology and Zoology, 41, 2, pp 295-301
<Go to ISI>://000238903300013 Siegel, J.P. (2001)
The mammalian safety of Bacillus thuringiensis-based insecticides. Journal of Invertebrate Pathology, 77, 1, pp 13-21
<Go to ISI>://000167001600002 AND Silvers, C.S. & Gianessi, L.P. (2001)
Disease resistance through biotechnology: Assessing benefits to U.S. growers. Phytopathology, 91, 6 Supplement, pp S82
<Go to ISI>://BIOSIS:PREV200100416475 Singh, P.K., Kumar, M., Chaturvedi, C.P., Yadav, D., & Tuli, R. (2004)
Development of a hybrid delta-endotoxin and its expression in tobacco and cotton for control of a polyphagous pest Spodoptera litura. Transgenic Research, 13, 5, pp 397-410
<Go to ISI>://WOS:000224724600001 AND http://www.botanischergarten.ch/Bt/Singh-Development-hybrid-2004.pdf
Siqueira, H.A.A., Gonzalez-Cabrera, J., Ferre, J., Flannagan, R., & Siegfried, B.D. (2006)
Analyses of Cry1Ab binding in resistant and susceptible strains of the European corn borer, Ostrinia nubilalis (Hubner) (Lepidoptera : Crambidae). Applied and Environmental Microbiology, 72, 8, pp 5318-5324
<Go to ISI>://WOS:000239780400025 AND http://www.botanischergarten.ch/Bt/Siqueira-Analysis-Cry1Ab-2006.pdf
Smith, R.A. & Couche, G.A. (1991)
The Phylloplane as a Source of Bacillus-Thuringiensis Variants. Applied and Environmental Microbiology, 57, 1, pp 311-315
<Go to ISI>://WOS:A1991ER20600049 AND http://www.botanischergarten.ch/Bt/Smith-Phylloplane-Bt-1991.pdf
Soberon, M., Fernandez, L.E., Perez, C., Gill, S.S., & Bravo, A. (2007)
Mode of action of mosquitocidal Bacillus thuringiensis toxins. Toxicon, 49, 5, pp 597-600 <Go to ISI>://000245866100001 AND http://www.botanischergarten.ch/Bt/Soberon-Mode-Action-Bt-2007.pdf Songstad, D.D., Halaka, F.G., Deboer, D.L., Armstrong, C.L., Hinchee, M.A.W., Fordsantino, C.G., Brown, S.M., Fromm, M.F., & Horsch, R.B. (1993)
Transient Expression of Gus and Anthocyanin Constructs in Intact Maize Immature Embryos Following Electroporation. Plant Cell Tissue and Organ Culture, 33, 2, pp 195-201
<Go to ISI>://WOS:A1993KZ21200012 AND http://www.botanischergarten.ch/Bt/Songstad-Transient-Expression-GUS-1993.pdf
Steinhaus, E.A. (1951)
Possible use of Bacillus thuringiensis Berliner as an aid in the biological control of the alfalfa cate. Hilgardia, 20, pp 359-381
Sterk, G., Hassan, S.A., Baillod, M., Bakker, F., Bigler, F., Blumel, S., Bogenschutz, H., Boller, E., Bromand, B., Brun, J., Calis, J.N.M., Coremans-Pelseneer, J., Duso, C., Garrido, A., Grove, A., Heimbach, U., Hokkanen, H., Jacas, J., Lewis, G., Moreth, L., Polgar, L., Roversti, L., Samsoe-Peterson, L., Sauphanor, B., Schaub, L., Staubli, A., Tuset, J.J., Vainio, A., Van de Veire, M., Viggiani, G., Vinuela, E., & Vogt, H. (1999)
Results of the seventh joint pesticide testing programme carried out by the IOBC/WPRS-Working Group 'Pesticides and Beneficial Organisms'. Biocontrol, 44, 1, pp 99-117
<Go to ISI>://WOS:000082271600008 AND http://www.botanischergarten.ch/Bt/Sterk-Pesticides-1999.pdf Sussex, I.M. (2008)
The Scientific Roots of Modern Plant Biotechnology. Plant Cell %R 10.1105/tpc.108.058735, 20, 5, pp 1189-1198
http://www.plantcell.org AND http://www.botanischergarten.ch/Bt/Sussex-Historical-Perspective-2008.pdf Sutherland, W.J., Bailey, M.J., Bainbridge, I.P., Brereton, T., Dick, J.T.A., Drewitt, J., Dulvy, N.K., Dusic, N.R., Freckleton, R.P., Gaston, K.J., Gilder, P.M., Green, R.E., Heathwaite, A.L., Johnson, S.M., Macdonald, D.W., Mitchell, R., Osborn, D., Owen, R.P., Pretty, J., Prior, S.V., Prosser, H., Pullin, A.S., Rose, P., Stott, A., Tew, T.,

88
Thomas, C.D., Thompson, D.B.A., Vickery, J.A., Walker, M., Walmsley, C., Warrington, S., Watkinson, A.R., Williams, R.J., Woodroffe, R., & Woodroof, H.J. (2008)
Future novel threats and opportunities facing UK biodiversity identified by horizon scanning. Journal of Applied Ecology, 0, 0 %R doi:10.1111/j.1365-2664.2008.01474.x, pp???-???
http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-2664.2008.01474.x AND http://www.botanischergarten.ch/Bt/Sutherland-Future-Novel-Threats-2008.pdf
Swiecicka, I. (2008)
Natural occurrence of Bacillus thuringiensis and Bacillus cereus in eukaryotic organisms: a case for symbiosis. Biocontrol Science and Technology, 18, 3, pp 221-239
<Go to ISI>://WOS:000255367200001 AND http://www.botanischergarten.ch/Bt/Swiecicka-Natural-Occurrence-Symbiosis-2008.pdf
Tabashnik, B., Van Rensburg, J.B.J., & Carriere, Y. (2009a)
Field-evolved resistance to Bt crops: Definition, Theory and Data. Journal of Economic Entomology, 102, 6, pp 2011-2025
http://www.botanischergarten.ch/Bt/Tabashnik-Field-Evolved-Insect-Resistance-2009.pdf Tabashnik, B.E., Fabrick, J.A., Henderson, S., Biggs, R.W., Yafuso, C.M., Nyboer, M.E., Manhardt, N.M., Coughlin, L.A., Sollome, J., Carriere, Y., Dennehy, T.J., & Morin, S. (2006)
DNA screening reveals pink bollworm resistance to Bt cotton remains rare after a decade of exposure. Journal of Economic Entomology, 99, 5, pp 1525-1530
<Go to ISI>://000241240400003 AND http://www.botanischergarten.ch/Bt/Tabashnik-DNA-Screening-2006.pdf
Tabashnik, B.E., Gassmann, A.J., Crowder, D.W., & Carriére, Y. (2008)
Insect resistance to Bt crops: evidence versus theory. Nature Biotechnology, 26, 2, pp 199-202 http://dx.doi.org/10.1038/nbt1382 AND http://www.nature.com/nbt/journal/v26/n2/suppinfo/nbt1382_S1.html
AND http://www.botanischergarten.ch/Bt/Tabashnik-Resistance-Evidence-2008.pdf Tabashnik, B.E., Unnithan, G.C., Masson, L., Crowder, D.W., Li, X., & Carriere, Y. (2009b)
Asymmetrical cross-resistance between Bacillus thuringiensis toxins Cry1Ac and Cry2Ab in pink bollworm. Proceedings of the National Academy of Sciences, pp 11889-11894
http://www.pnas.org/content/early/2009/07/02/0901351106.abstract AND http://www.botanischergarten.ch/Bt/Tabshnik-X-resistance-Bt-2009.pdf
Tall, L., Methot, G., Armellin, A., & Pinel-Alloul, B. (2008)
Bioassessment of Benthic Macroinvertebrates in Wetland Habitats of Lake Saint-Pierre (St. Lawrence River). Journal of Great Lakes Research, 34, 4, pp 599-614
<Go to ISI>://WOS:000262663900004 AND http://www.botanischergarten.ch/Bt/Tall-Bioassessment-Benthic-2008.pdf
Tanaka, H. & Kimura, Y. (1991)
Resistance to Bt Formulation in Diamondback Moth, Plutella- Xylostella L, on Watercress. Japanese Journal of Applied Entomology and Zoology, 35, 3, pp 253-255
<Go to ISI>://A1991GD11600010 Tang, M., Bideshi, D.K., Park, H.W., & Federici, B.A. (2007)
Iteron-binding ORF157 and FtsZ-Like ORF156 proteins encoded by pBtoxis play a role in its replication in Bacillus thuringiensis subsp. israelensis. Journal of Bacteriology, 189, pp 8053-8058
<Go to ISI>://WOS:000250991300014 Teschke, K., Chow, Y., Bartlett, K., Ross, A., & van Netten, C. (2001)
Spatial and temporal distribution of airborne Bacillus thuringiensis var. kurstaki during an aerial spray program for gypsy moth eradication. Environmental Health Perspectives, 109, 1, pp 47-54
<Go to ISI>://WOS:000166391400023 AND http://www.botanischergarten.ch/Bt/Teschke-Spatial-Temporal-Btk-2001.pdf
Thomson, J.A. (2002)
Research needs to improve agricultural productivity and food quality, with emphasis on biotechnology. Journal of Nutrition, 132, 11, pp 3441S-3442S
<Go to ISI>://000179109300034 Travers, R.S., Martin, P.A.W., & Reichelderfer, C.F. (1987)

89
Selective Process for Efficient Isolation of Soil Bacillus Spp. Applied and Environmental Microbiology, 53, 6, pp 1263-1266
<Go to ISI>://WOS:A1987H528700011 AND http://www.botanischergarten.ch/Bt/Travers-Selective-Process-Bt-1987.pdf
Turlings, T.C.J., Jeanbourquin, P.M., Held, M., & Degen, T. (2005)
Evaluating the induced-odour emission of a Bt maize and its attractiveness to parasitic wasps. Transgenic Research, 14, 6, pp 807-816
<Go to ISI>://000233628500004 AND http://www.botanischergarten.ch/Bt/Turlings-Evaluating-Induced-Odor-2005.pdf
Tzfira, T. & Citovsky, V. (2006)
Agrobacterium-mediated genetic transformation of plants: biology and biotechnology. Current Opinion in Biotechnology, 17, 2, pp 147-154
<Go to ISI>://WOS:000237135800007 AND http://www.botanischergarten.ch/Bt/Tzfira-Agrobacterium-mediated-2006.pdf
US Environmental Protection Agency (USEPA) (1995)
Pesticide fact sheet for Bacillus thuringiensis ssp. kurstaki Cry1(A)b delta-endotoxin and the genetic material necessary for the production (plasmid vector pCIB4431) in corn., Vol. EPA publication no. EPA 731-F-95–004 (Environmental Protection Agency, Washington, DC, USA,
US Environmental Protection Agency (USEPA) (2001)
Biopesticide registration action document.Bacillus thuringiensis (Bt) plant-incorporated protectants. (eds U. EPA). U.S. Environmental Protection Agency U.S. EPA
http://www.epa.gov/oppbppd1/biopesticides/pips/bt_brad.htm Vaeck, M., Reynaerts, A., Hofte, H., Jansens, S., Debeuckeleer, M., Dean, C., Zabeau, M., Vanmontagu, M., & Leemans, J. (1987)
Transgenic Plants Protected from Insect Attack. Nature, 328, 6125, pp 33-37 <Go to ISI>://WOS:A1987H995400044 AND http://www.botanischergarten.ch/Bt/Vaeck-Transgenic-Insect-
Nature-1987.pdf Van Montagu, M. (1997)
Once upon a time ... the Belgian epic of transgenesis. Biofutur, 172, pp 9-11 <Go to ISI>://000071078300002 Van Montagu, M., Holsters, M., Zambryski, P., Hernalsteens, J.P., Depicker, A., Debeuckeleer, M., Engler, G., Lemmers, M., Willmitzer, L., & Schell, J. (1980)
The Interaction of Agrobacterium Ti-Plasmid DNA and Plant-Cells. Proceedings of the Royal Society of London Series B-Biological Sciences, 210, 1180, pp 351-365
<Go to ISI>://A1980KR46300004 AND http://www.botanischergarten.ch/Bt/VanMontagu-Interaction-Agrobacterium-1980.pdf
Van Montagu, M., Inze, D., Deblaere, R., Joos, H., Deblock, M., Hernalsteens, J.P., Caplan, A., Messens, E., Zambryski, P., & Schell, J. (1983)
Plant Tumor-Induction by Agrobacterium-Tumefaciens - Structure and Function of the T-DNA-Encoded Oncogenic Proteins. European Journal of Cancer & Clinical Oncology, 19, 9, pp 1322-1322
<Go to ISI>://A1983RH72900175 NOT IN EZB, NEBIS from 1993 ff Vanlarebeke, N., Genetello, C., Hernalsteens, J.P., Depicker, A., Zaenen, I., Messens, E., Vanmontagu, M., & Schell, J. (1977)
Transfer of Ti Plasmids between Agrobacterium Strains by Mobilization with Conjugative Plasmid Rp4. Molecular & General Genetics, 152, 2, pp 119-124
<Go to ISI>://WOS:A1977DD80100002 AND http://www.botanischergarten.ch/Bt/VanLarebeke-Transfer-Plasmids-1977.pdf
Vanrie, J., Jansens, S., Hofte, H., Degheele, D., & Vanmellaert, H. (1989)
Specificity of Bacillus-Thuringiensis Delta-Endotoxins - Importance of Specific Receptors on the Brush-Border Membrane of the Mid-Gut of Target Insects. European Journal of Biochemistry, 186, 1-2, pp 239-247
<Go to ISI>://WOS:A1989CE48300032 AND http://www.botanischergarten.ch/Bt/VanRie-SpecifityBt-Endotoxins-1989.pdf
Vanvalen, L. (1976)

90
Ecological Species, Multispecies, and Oaks. Taxon, 25, 2-3, pp 233-239 <Go to ISI>://WOS:A1976BW85700002 AND http://links.jstor.org/sici?sici=0040-
0262%28197605%2925%3A2%2F3%3C233%3AESMAO%3E2.0.CO%3B2-H AND http://www.botanischergarten.ch/Bt/vanValen-Ecological-Concept-1976.pdf
Vaughn, T., Cavato, T., Brar, G., Coombe, T., DeGooyer, T., Ford, S., Groth, M., Howe, A., Johnson, S., Kolacz, K., Pilcher, C., Purcell, J., Romano, C., English, L., & Pershing, J. (2005)
A method of controlling corn rootworm feeding using a Bacillus thuringiensis protein expressed in transgenic maize. Crop Science, 45, 3, pp 931-938
<Go to ISI>://000229105700018 AND http://www.botanischergarten.ch/Bt/Vaughn-Controlling-Rootworm-2005.pdf
Vellai, T., Kovacs, A.L., Kovacs, G., Ortutay, C., & Vida, G. (1999)
Genome economization and a new approach to the species concept in bacteria. Proceedings of the Royal Society of London Series B-Biological Sciences, 266, 1432, pp 1953-1958
<Go to ISI>://WOS:000083173100004 AND http://www.botanischergarten.ch/Bt/Vellai-Genome-Economization-1999.pdf
Vilas-Boas, G., Sanchis, V., Lereclus, D., Lemos, M.V.F., & Bourguet, D. (2002)
Genetic Differentiation between Sympatric Populations of Bacillus cereus and Bacillus thuringiensis. Appl. Environ. Microbiol. %R 10.1128/AEM.68.3.1414-1424.2002, 68, 3, pp 1414-1424
http://aem.asm.org/cgi/content/abstract/68/3/1414 AND http://www.botanischergarten.ch/Bt/Vilas-Boas-Genetics-cereus-thuring-2002.pdf
Vilas-Boas, G.T., Peruca, A.P.S., & Arantes, O.M.N. (2007)
Biology and taxonomy of Bacillus cereus, Bacillus anthracis, and Bacillus thuringiensis. Canadian Journal of Microbiology, 53, 6, pp 673-687
<Go to ISI>://WOS:000249202500001 AND http://www.botanischergarten.ch/Bt/Vilas-Boas-Biology-Taxonomy-Bacillus-2007.pdf
Wallimann, T. (2000)
Bt Toxin: Assessing GM Strategies. Science %R 10.1126/science.287.5450.41c, 287, 5450, pp 41c- http://www.sciencemag.org AND http://www.botanischergarten.ch/Bt/Wallimann-Assessing-Reistance-
2000.pdf Weisse, T. (2007)
The species concept of bacteria, protists and metazoa. Denisia, 20, pp 149-154 <Go to ISI>://BIOSIS:PREV200800037130 AND http://www.botanischergarten.ch/Bt/Weisse-Artbegriff-
2007.pdf AND http://www.botanischergarten.ch/Bt/Weisse-Artbegriff-2007-alpha.pdf Wesseler, J., Scatasta, S., & Nillesen, E. (2007)
The Maximum Incremental Social Tolerable Irreversible Costs (MISTICs) and other benefits and costs of introducing transgenic maize in the EU-15, SPECIAL ISSUE SOIL ECOLOGICAL AND ECONOMIC EVALUATION OF GENETICALLY MODIFIED CROPS - ECOGEN. Pedobiologia, 51, 3, pp 261-269
http://www.sciencedirect.com/science/article/B7CW5-4NNYD85-2/2/de0c1947c9225141970eed546bc92fb0 AND http://www.botanischergarten.ch/Bt/Wesseler-MISTICs-ECOGEN-2007.pdf
Whalon, M.E. & Wingerd, B.A. (2003)
Bt: Mode of action and use. Archives of Insect Biochemistry and Physiology, 54, 4, pp 200-211 <Go to ISI>://WOS:000186961000007 AND http://www.botanischergarten.ch/Bt/Whalon-Mode-of-Action-
2003.pdf Whetstone, P.A. & Hammock, B.D. (2007)
Delivery methods for peptide and protein toxins in insect control. Insecticidal Toxins and their Potential for Insect Pest Control, 49, 4, pp 576-596
http://www.sciencedirect.com/science/article/B6TCS-4MD2T9R-6/2/198ef6259760dd5508973d2c0e955dea AND http://www.botanischergarten.ch/Bt/Whetstone-Delivery-Methods-2007.pdf
Whiteley, H.R. & Schnepf, H.E. (1986)
The Molecular-Biology of Parasporal Crystal Body Formation in Bacillus-Thuringiensis. Annual Review of Microbiology, 40, pp 549-576
<Go to ISI>://WOS:A1986E243900022 AND http://www.botanischergarten.ch/Bt/Whiteley-Molecular-Biotlogy-Bt-1986.pdf

91
Widmer, F. (2007)
Assessing effects of Transgenic crops on soil microbial communities. In Green Gene Technology: Research in an Area of Social Conflict, pp. 207-234. SPRINGER-VERLAG BERLIN, BERLIN Advances in Biochemical Engineering / Biotechnology-107,
<Go to ISI>://000248483300011 AND http://www.botanischergarten.ch/Bt/Widmer-Assessing-Effects-2007.pdf
Willmitzer, L., Debeuckeleer, M., Lemmers, M., Vanmontagu, M., & Schell, J. (1980)
DNA from Ti Plasmid Present in Nucleus and Absent from Plastids of Crown Gall Plant-Cells. Nature, 287, 5780, pp 359-361
<Go to ISI>://WOS:A1980KJ23500058 AND http://www.botanischergarten.ch/Bt/Willmitzer-DNA-Ti-Plasmid-1980.pdf
Willmitzer, L., Dhaese, P., Schreier, P.H., Schmalenbach, W., Vanmontagu, M., & Schell, J. (1983)
Size, Location and Polarity of T-DNA-Encoded Transcripts in Nopaline Crown Gall Tumors - Common Transcripts in Octopine and Nopaline Tumors. Cell, 32, 4, pp 1045-1056
<Go to ISI>://WOS:A1983QN11800010 Wilson, T.A., Rice, M.E., Tollefson, J.J., & Pilcher, C.D. (2005)
Transgenic corn for control of the European corn borer and corn rootworms: a survey of Midwestern farmers' practices and perceptions. Journal of Economic Entomology, 98, 2, pp 237-247
<Go to ISI>://000228259000001 AND http://www.botanischergarten.ch/Bt/Wilson-Survey-Midwestern-2006.pdf
Wolfenbarger, L.L., Naranjo, S.E., Lundgren, J.G., Bitzer, R.J., & Watrud, L.S. (2008)
Bt Crop Effects on Functional Guilds of Non-Target Arthropods: A Meta-Analysis. PLoS ONE, 3, 5, pp e2118 http://dx.doi.org/10.1371%2Fjournal.pone.0002118 AND http://www.botanischergarten.ch/Bt/LaReesa-Bt-
crop-Meta-Analysis-2008.pdf AND http://www.botanischergarten.ch/Bt/LaReesa-Meta-Analysis-Powerpoints-2008.ppt
Wu, D. & Aronson, A.I. (1992)
Localized Mutagenesis Defines Regions of the Bacillus-Thuringiensis Delta-Endotoxin Involved in Toxicity and Specificity. Journal of Biological Chemistry, 267, 4, pp 2311-2317
<Go to ISI>://A1992HB53200032 Wu, J., Zhao, F., Bai, J., Deng, G., Qin, S., & Bao, Q. (2007)
Evidence for positive Darwinian selection of Vip gene in Bacillus thuringiensis. Journal of Genetics and Genomics, 34, 7, pp 649-660
<Go to ISI>://BIOSIS:PREV200700493441 AND http://www.botanischergarten.ch/Bt/Wu-Evidence-Darwinian-2007.pdf
Wu, S.J., Koller, C.N., Miller, D.L., Bauer, L.S., & Dean, D.H. (2000)
Enhanced toxicity of Bacillus thuringiensis Cry3A delta-endotoxin in coleopterans by mutagenesis in a receptor binding loop. Febs Letters, 473, 2, pp 227-232
<Go to ISI>://WOS:000087126800023 AND http://www.botanischergarten.ch/Bt/Wu-Enhanced-Toxicity-Bt-2000.pdf
Xu, X.J. & Wu, Y.D. (2008)
Disruption of Ha_BtR alters binding of Bacillus thuringiensis delta-endotoxin Cry1Ac to midgut BBMVs of Helicoverpa armigera. Journal of Invertebrate Pathology, 97, pp 27-32
<Go to ISI>://WOS:000252228100005 Xue, J., Liang, G., Crickmore, N., Li, H., He, K., Song, F., Feng, X., Huang, D., & Zhang, J. (2008)
Cloning and characterization of a novel Cry1A toxin from Bacillus thuringiensis with high toxicity to the Asian corn borer and other lepidopteran insects. Fems Microbiology Letters, 280, 1, pp 95-101
<Go to ISI>://WOS:000252929800014 AND http://www.botanischergarten.ch/Bt/Xue-Cloning-Novel-Cry1A-2008.pdf
Yu, C.G., Mullins, M.A., Warren, G.W., Koziel, M.G., & Estruch, J.J. (1997)
The Bacillus thuringiensis vegetative insecticidal protein Vip3A lyses midgut epithelium cells of susceptible insects. Applied and Environmental Microbiology, 63, 2, pp 532-536
<Go to ISI>://WOS:A1997WF49600026 AND http://www.botanischergarten.ch/Bt/Yu-Bt-Vegetative-Vip3A-1997.pdf

92
Yu, Y.M., Ohba, M., & Gill, S.S. (1991)
Characterization of Mosquitocidal Activity of Bacillus-Thuringiensis Subsp Fukuokaensis Crystal Proteins. Applied and Environmental Microbiology, 57, 4, pp 1075-1081
<Go to ISI>://WOS:A1991FF03900030 AND http://www.botanischergarten.ch/Bt/Yu-Bt-ssp.-fukuokaensis-1991.pdf
Zambryski, P., Herrera-Estrella, L., De Block, M., Van Montagu, M., & Schell, J. (1984)
The use of the Ti plasmid of Agrobacterium to study the transfer and expression of foreign DNA in plant cells: new vectors and methods Genetic Engineering, 6, pp 253-78 CODEN: GENGDC