equids of ÇatalhÖyÜk: an insight into theory and methodology in the identification of equidae
TRANSCRIPT
EQUIDS OF ÇATALHÖYÜK: AN INSIGHT INTO THEORY AND METHODOLOGY IN THE IDENTIFICATION OF EQUIDAE
IAN C. CAMERON
Dissertation submitted in partial fulfilment of the requirements for the degree of MSc in Palaeoecology of Human Societies of the University of London in 2005
UNIVERSITY COLLEGE LONDON INSTITUTE OF ARCHAEOLOGY
Note: This Dissertation is an unrevised examination copy for consultation only and it should not be quoted or cited without the permission of the Director of the Institute

ABSTRACT This dissertation is an attempt to further identify the equid remains from the Neolithic site of
Çatalhöyük in southwestern Anatolia and improve knowledge of equids in the region. Three
species of equids (E. caballus ferus, E. hemionus, and E. hydruntinus) will be discussed in
regards to the theory of their presence at Çatalhöyük. Furthermore, various methods of
identifying the equid remains will be discussed including morphology and morphometrics. The
success of these methods in the investigation of the equid remains will be discussed. A regional
comparison of the Çatalhöyük equid remains to equid remains from sites in Anatolia and the Near
East will also be presented. When the morphological and morphometric data are viewed together
for the equid first phalanges, a pattern can be identified that with further comparison to other
collections may possibly provide a basis for a method of differentiating between E. hydruntinus
and E. hemionus using this element.
ii

LIST OF CONTENTS
Chapter 1) Equids and the Environment of southwest Asia 1.1 Aims 1 1.2 The Holocene environment of Anatolia 1 1.3 Equid Species Neolithic Anatolia 2 1.4 Equid remains from sites in Anatolia and the Near East 5 1.5 Equids at Çatalhöyük – Potential problems 7 1.6 Summary 9 Chapter 2) Çatalhöyük and Potential Models for Equids 2.1 Background to Çatalhöyük 10 2.2 Opportunistic hunting 14 2.3 Logistical hunting 15 2.4 Ecosystem model 15 2.5 Effects of Taphonomy 17 2.6 Summary 18 Chapter 3) Methodology 3.1 Identifying the species of the equid remains from Çatalhöyük 19 3.2 Size of Element 19 3.3 Tooth Morphology 20 3.4 Tooth Morphometrics 20 3.5 First Phalanx Morphology 23 3.6 Ageing the equid remains from Çatalhöyük 26 3.7 Analysis of equid measurements 26 Chapter 4) Results 4.1 Representation of equid species 28 4.2 Metric analysis of equid teeth 28 4.3 Analysis of measurements 28 4.4 Morphological attributes of First Phalanx 36 4.5 Body-part representation 41 4.6 Age of equid population 41 4.7 Summary 42 Chapter 5) General Discussion 5.1 Equids at Çatalhöyük 43 Chapter 6) Conclusion 6.1 Concluding Remarks 46 6.2 Further research recommendations 46 References cited 48 Appendices 54
iii

LIST OF FIGURES Figure 1 - Equus hemionus 3 Figure 2 - Equus caballus ferus 5 Figure 3 - Pie Chart of Equid elements identified at Çatalhöyük 8 Figure 4 - Location map of Çatalhöyük in Turkey (Adapted from 10 http://catal.arch.cam.ac.uk/2005). Figure 5 - Plan of Çatalhöyük East Mound showing excavation areas 11 (Adapted from Richards et al. 2003: 68). Figure 6 - Artist’s depiction of Neolithic Çatalhöyük (Adapted from 13 http://catal.arch.cam.ac.uk/2005). Figure 7 - Measurements of equid teeth, upper teeth (top), lower teeth (bottom) 21 (Adapted from Buitenhuis 2003: 25). Figure 8 - Equid maxillary teeth from Çatalhöyük 22 Figure 9 - Equid mandibular teeth from Çatalhöyük 23 Figure 10 - Diagram illustrating equid foot (adapted from Sisson 1953:229) 23 Figure 11 - Attribute A - Depth of Sulcus (Id #s 7880 F242, 4465 F121). 24 Figure 12 - Attribute C - Roundness of Distal Articular Surface 25 (Id #s 1668 F325, 1505 F222). Figure 13 - Attribute F - Shape of Proximal Articulation (Id #s 4836 F1, 1505 F222). 25 Figure 14 - Attribute H - Presence of Posterior Proximal Sulcus Lipping 26 (Id#s 7880 F242, 3736 F127). Figure 15 - Scatter plot of Astragalus Measurement 7 versus Measurement 8 30 Figure 16 - Scatter plot of Astragalus Measurement 9 versus Measurement 10 31 Figure 17 - Scatter plot of Phalanx I Measurement 4 versus Measurement 5 32 Figure 18 - Scatter plot of Phalanx I Measurement 2 versus Measurement 7 32 Figure 19 - Scatter plot of Phalanx II Measurement 2 versus Measurement 5 33 Figure 20 - Scatter plot of Phalanx III Measurement 1 versus Measurement 6 34 Figure 21 - Scatter plot of Scapula Measurement 3 versus Measurement 4 34 Figure 22 - Scatter plot of Pelvis Measurement 1 versus Measurement 2 35 Figure 23 - Scatter plot of Tibia Measurement 3 versus Measurement 4 36 Figure 24 - Scatter plot of Phalanx I Measurements (SD versus GL) and 40 Morphological Attributes Figure 25 - Scatter plot of Phalanx I Measurements (Bp versus Dp) and 40 Morphological Attributes Figure 26 - Proportion of age determined equid elements at Çatalhöyük 41 Figure 27 - Age of equid population recovered from Çatalhöyük 42
iv

LIST OF TABLES
Table 1 - Scale of Stages for Measurement of Equid Teeth 22 Table 2 - Equid measurements from sites in Anatolia and the Near East 37 (measurements are means). Table 3 - List of Morphological Attributes applied to Equid First Phalanges 39 Table 4 - Two groups of Phalanx I attributes 39
LIST OF APPENDICES Appendix 1 - Metric analysis of maxillary equid teeth from Çatalhöyük 54 Appendix 2 - Metric analysis of mandibular equid teeth from Çatalhöyük 55 Appendix 3 - Measurements proposed by Payne (1991:135) for upper (above) 57 and lower equid teeth. Appendix 4 - Measurements of Postcranial Elements 58 Appendix 5 - Measurement statistics of postcranial equid elements 62 Appendix 6 - Morphological Attributes of First Phalanges 63
v

ACKNOWEDGEMENTS In the process of writing and researching this dissertation I had assistance from several people whose help was greatly appreciated. Thanks to Louise Martin and Ken Thomas for comments on drafted chapters. Thank you to Hiljke Buitenhuis, Louise Martin and Nerissa Russell for their insights. Thank you Lisa Yeomans for help with the database and for friendly advice. And of course thanks to all my family and friends who have helped me along the way.
vi

1
1) Equids and the environment of southwest Asia
1.1 Aims
Little is known about the different wild equid species utilized by people in Neolithic Anatolia.
This dissertation aims to analyse the equid remains from the site of Çatalhöyük in central
Anatolia to determine the species of the equids present using various morphological and metrical
analyses. Equids were most likely an important wild food resource utilized by the people of
Çatalhöyük in addition to agricultural and pastoral food sources (Hodder 1999:158). Equids were
recognized as a source of hunted food at Çatalhöyük during the 1960’s excavations by Mellart
(1975:99), though it is uncertain what equid species were recovered as they are listed only as
‘onager and half-ass’ (1967:223). The changes and fluctuations in time of the equid species
(Equus caballus ferus, Equus hemionus, and Equus hydruntinus) represented at Çatalhöyük are
expected to follow ecological models and economic models such as opportunistic and logistical
hunting models. Results from other sites in Anatolia and the Near East will be compared to the
Çatalhöyük remains in an effort to detect similarities between the equid populations.
1.2 The Neolithic environment of Anatolia
The environment of Anatolia was much different during the Neolithic than today with much
greater precipitation and cooler temperatures. Through analysis of pollen, charcoal, stable
isotope, and geochemical evidence in Anatolia, it is apparent that there was quite an increase in
precipitation at the onset of the Holocene which is visible in the palaeobotanical record by an
increase in Pistacia and non-arboreal vegetation (Wick et al. 2003: 673). This weather would
have slowed down the expansion of oak for the first three thousand years of the Holocene in the
Lake Van area (Ibid) and perhaps in other areas like the Konya Plain where steppe vegetation was
present. Through analysis of pollen cores taken from Akgol in south-western Anatolia, it has
been possible to reconstruct the Konya Plain environment (Bottema and Woldring 1984: 147)
which was apparently experiencing salinization to some extent around 9000 B.P., a condition that
was very good for grass land development. From 9000 B.P. there was a slight increase in
precipitation which created slightly saline meadows that would have been ideal for cattle grazing
(Ibid). In fact, maximum tree cover after the start of the Holocene was not achieved in central
and east Anatolia until ca. 6000 B.P. (Kuzucuoglu and Roberts 1998). In Cappadocia, pollen and
faunal evidence from the sites of Aşikli Höyük and Musular indicates that the Neolithic was

2
relatively wet and supporting open grasslands ideal for grazing animals (Woldring 2002: 64).
Basically, most of Anatolia was experiencing a wet period during the Neolithic that supported a
wide range of rich and varied natural habitats and encouraged both human and faunal activity in
the area (Ozdogan 1998:34).
The Konya Plain is surrounded by the Aradaglari Mountains to the east, the Taurus Mountains to
the south and southwest and the Sultandaglari Mountains to the west. Micromorphological
analysis of soil samples has provided evidence that the area around Çatalhöyük was a delta at the
edge of a Pleistocene lake and subject to seasonal flooding (Matthews 1999, Rosen and Roberts
2005). Today the Konya Plain exhibits several features of Central Anatolian continentality
including endoreism, plateau relief at 1000 m altitude, a continental climate with less than 400
mm precipitation yearly and steppe vegetation (Kuzucuoglu 2002: 33).
The Neolithic environment of Anatolia appears to have supported three species of equids, namely
E. caballus ferus (wild horse), E. hemionus (onager or Asiatic Wild Ass), and E. hydruntinus
(European Wild Ass) (Groves 1986). These species appear to have coexisted in Anatolia and
were hunted to varying degrees as is evident at archaeological sites such as Çatalhöyük, Can
Hasan III, Aşikli Höyük and others. These equid species will be described in the next section.
1.3 Equid Species of Neolithic Anatolia
There have been three equid species identified at Neolithic sites in Anatolia, and Çatalhöyük in
particular, including E. caballus ferus (wild horse), E. hemionus (hemione or Asiatic Wild Ass),
and E. hydruntinus (hydruntine or European Wild Ass). The following section is a brief
description of equid biology and the three equid species.
Equids are bulk feeders that require large amounts of low quality forage and fresh water due to
their large size (Saltz 2002:119). While their ability to survive off low quality forage allows
equids to adapt to poor habitat conditions and makes them fairly resilient to environmental
change, their need for water limits their range, movement patterns and survivability in times of
drought (Ibid:120). Due to the chances of mass mortality brought about by a period of drought,
disease, heavy frost, or through human predation, equid populations may experience boom/bust
cycles (Ibid:121) and these cycles may be reflected in the archaeological record. Coughenour
(2002:159) has suggested that dynamic ecosystem models could be used to predict fluctuations in

3
wild equid populations over time and these could perhaps be changes in abundance that could be
correlated with archaeological evidence. It has been suggested that human activity, the spread of
forests throughout Europe and environmental change caused enough disruption to force wild
equid populations to migrate eastwards (Clutton-Brock 1992:25). This could perhaps be seen as a
precursor to changes in equid populations (Equus caballus ferus, Equus hemionus and Equus
hydruntinus) on the Konya Plain and throughout the Near East.
Equus hemionus
The hemione (E. hemionus) or Asiatic Wild Ass is of a relative small size and gracile build
(Figure 1) when compared to the horse (E. caballus). This equid has no domestic descendants
and so far there is no evidence for its domestication in the ancient world (Clutton-Brock
1992:37). Hemiones are the only wild equids remaining today in the Middle East, and though
they once ranged from the Mediterranean into Central Asia (Uerpmann 1987:19). The natural
habitat of E. hemionus is ‘flat, desert country such as salt flats or gravel plains’ (Groves 1986:
38). The hemione is smaller than the wild horse and seems to be larger than E. hydruntinus on
average, though the size range between the hemione and hydruntine overlaps sufficiently to make
identification difficult (Uerpmann 1987). The hemione is regarded as one of the most important
hunted food resources for hunter-gatherer groups in the Near East and Middle East (Uerpmann
1987). E. hemionus remains have been identified throughout Anatolia at sites including
Çatalhöyük (Martin and Russell, in press), Cayonu (Meadow 1986), Can Hasan III (Payne 1991),
Demirci Huyuk (Boessneck and Von Den Driesch 1977) and in the Middle East at Shams ed-Din
(Uerpmann 1982) and possibly Mureybit (Uerpmann 1982, 1986, Meadow 1986).
Figure 1 - Equus hemionus

4
Equus hydruntinus
The hydruntine (Equus hydruntinus) or European Wild Ass is of a small size and gracile build
and thought to have been similar to zebras (Uerpmann 1987:32). This species is different from
horses, hemiones and asses as it is a palaeontological species or ‘an artificial unit put together on
the basis of morphological similarities’ (Ibid). Indeed it has been suggested that E. hydruntinus
was actually two or three different species separated by time intervals and geographic locations
(Uerpmann 1987). E. hydruntinus has been found to be present in Southern Europe from the
Middle Pleistocene into the Holocene and in the Levant until the Upper Pleistocene. Equid
remains from Can Hasan III in Holocene Anatolia have been identified by Sebastian Payne
(1991) as E. hydruntinus and combined with the identification of E. hydruntinus via tooth
morphology analysis at Çatalhöyük by Martin and Russell (in press), this lends evidence to the
presence of a Holocene E. hydruntinus population that was being utilized by Neolithic Anatolian
peoples at Can Hasan III and Çatalhöyük. In the Middle East, E. hydruntinus was apparently
abruptly replaced by asses or onagers at surprisingly late dates (Groves 1986: 47).
The disappearance of E. hydruntinus in Anatolia is interesting as the current environmental
conditions are similar to the Holocene conditions which supported the E. hydruntinus population.
This leads to questions of whether it was over exploitation by humans (not evident in the
archaeological remains), catastrophic extinction due to extreme environmental fluctuations
involving draught or harsh winters, epidemic disease, or perhaps competition with populations of
migrating groups of E. caballus and/or E. hemionus. The possibility of E. hydruntinus being
absorbed into other equid populations through breeding will be addressed in the next chapter.
Equus caballus ferus
The wild horse (E. caballus ferus) is the ancestor to the domestic horse (E. caballus) and believed
to be close to extinction today (Uerpmann 1987:13, Moehlman 2005) (Figure 2). Wild horse
remains have been recovered from various sites in Anatolia including Pulur Höyük in the Keban
Dam region (Deniz 1975) and Kara’in (Uerpmann 1987:15). It is believed that wild horses
entered the coldest parts of the Near East and Middle East after the rising temperatures of the
Holocene forced them to migrate southward (Uerpmann 1987:17). The presence of wild horse at
Çatalhöyük was not recognized until the excavations began again in the early 1990s and has been
recorded in reports ever since (Russell et al. 1996, Russell and Martin 2005). There has been no
evidence that the wild horse was domesticated at Çatalhöyük (Ibid). E. caballus ferus was most
likely hunted in much the same way the hemione and hydruntine were hunted.

5
Figure 2 - Equus caballus ferus
1.4 Equid remains from sites in Anatolia and Near East
The following section outlines some equid remains recovered from archaeological sites in
Anatolia and the Near East and the species designation assigned to each. This is designed to give
the reader a general idea of when and where certain equid species were being utilized by humans
as is evident through analysing the archaeological record. The most common problem evident
throughout the literature involves the identification of different equid species. Due to
morphological similarities between many of the equid species, it has been difficult for researches
to identify equid remains to species in some cases when observing postcranial remains and
sometimes cranial remains.
Equid remains retrieved from 4th millennium Arjoune in Syria have been identified as hemiones
(E. hemionus) or wild or domestic asses (E. africanus or E. asinus) and not as horses (E.
caballus). This identification is based on size and dental morphology (Grigson 2000:17).
Domestic asses (E. asinus) are thought to be possibly present due to evidence for the presence of
domestic donkeys at other sites of the same period in the region (Ibid).
At the small tell of Hayaz Höyük in south eastern Anatolia, one fragment of an equid mandible
was recovered from the Neolithic period (c. 6000 BC) and at the site of Nahr el Homr in northern
Syria (c. 8000 BC), 85 percent of the faunal assemblage (all hunted animals) are equids
(Buitenhuis 1991). The equids at Nahr el Homr appear to be E. hemionus as per the tooth
morphology evidence (Ibid: 41). Based on the small size of the equid mandible fragment
recovered from Hayaz Höyük, it is believed to be E. hemionus (Buitenhuis 1985: 67).
Richard Meadow studied 83 equid bones and teeth from the site of Cayonu in southeastern
Turkey and identified them as part of a population of Equus hemionus which was hunted in the
area during the late eighth and early seventh millennium b.c. (Meadow 1986:282). Meadow

6
(1986:274) relied primarily on tooth morphology and bone size to identify the equid remains as
hemione (E. hemionus) and not wild horse (E. caballus ferus) or hydruntine (E. hydruntinus). An
experimental approach using a size index of postcranial measurements compared against equid
measurements from Mureibit in Syria also provided confidence that the Cayonu equid bones
came from only one species of equid (Ibid: 281). The identification of equid species by their
relative size is based on the assumption that wild horses will be larger than hemiones which are,
in turn, larger than hydruntines (Uerpmann 1987).
At the Halafian site of Shams ed-Din Tannira in northern Syria, Uerpmann (1982:24) identified
equid teeth and discusses a possible E. hydruntinus tooth which was isolated in the equid remains
and may be a peculiarity that indicates the occasional presence of a deep external fold in E.
hemionus as well. It is suggested by Uerpmann that the presence of equid remains at Shams ed-
Din Tannira represents opportunistic hunting of equids that may have been feeding off of cereal
crops. Hunting of equids would have served two purposes including the protection of the cereal
crops and providing food resources for the community. Later, Uerpmann (1986) was concerned
with differentiating between E. hemionus and E. africanus and compared the results of metrical
and morphological analysis of postcranial remains from Shams ed-Din Tannira to Mureibit.
Mureibit is an early Holocene site (c. 8200 to c. 7600 B.C.) that has been studied by Ducos (1975,
1986) who identified the equid remains from the site as E. africanus. In the end, Uerpmann
(1986) decided that the remains from Shams ed-Din Tannira were E. hemionus due to differences
in size and morphology from the Mureibit equid remains.
At Tell Rubeidheh in eastern Iraq, Payne (1988) identified equid remains of moderate sized bones
and teeth which had no signs of butchery marks. While this site is from the fourth millennium, it
is a good example of attempting to differentiate between species using epigraphic and
osteological evidence. In this case the species in question were the onager (E. hemionus) and the
domestic ass (E. asinus).
Payne (1991) has identified likely candidates for E. hemionus and E. hydruntinus from equid
remains recovered from the early Holocene sites of Tall-i-Mushki (Iran) and Can Hasan III
(Turkey) respectively. Methods used by Payne include analysis of tooth morphology and the
robustness of the postcranial bones (Ibid). Payne believes that dental characteristics will be the
most useful factor in identification of equid species for several reasons; including the relative
paucity of postcranial remains as compared to teeth and that it is easier to identify remains to the

7
subgeneric and specific level using teeth while postcranial elements are more affected by
nutrition and environmental conditions (1991:134).
At Aşikili Hüyük, Payne (1985) identified four equid bones, a distal humerus and three phalanges
of moderate size and most likely to be E. hemionus or E. hydruntinus rather than horse (E.
caballus). Through identification of these equid bones and other grazing animals, Payne suggests
that the area was an open, steppe environment with little tree cover (Ibid:113). These equid
remains date to 6661 ± bc and 7008 ± 130 bc (Ibid:115). At Pulur Hüyük in the Kevan Dam
region of Turkey, Deniz (1975) undertook zooarchaeological studies of 596 bones including both
domestic and wild animals and human. One equid bone from the Neolithic was identified as
Equus ferus and no other bones were identified beyond being Equus species.
1.5 Equids at Çatalhöyük - Potential problems
It has not been suggested that the equid remains from Neolithic Çatalhöyük represent
domesticated populations and this has not been disputed by researchers at Çatalhöyük (Martin
and Russell, in press). The earliest domestic horse (E. caballus) and donkey (E. asinus) are not
known in Anatolia and the Near East until the fourth millennium bc (Grigson 2003). For the
purposes of this exercise, all equid remains will be considered to be representative of wild
Neolithic populations. As mentioned earlier, there have been three different species of equids
recovered during the excavation of Çatalhöyük. These species include E. caballus ferus, E.
hemionus, and E. hydruntinus. The initial identification of E. caballus ferus at Çatalhöyük was
considered to be unusual as the wild horse was not known to be in the area during the Neolithic
(Russell and Martin 2000). Even more unusual is the presence of the three different equid species
in the Çatalhöyük area, apparently at the same time.
There have been several types of problems associated with the equid remains from Çatalhöyük,
mainly identifying the equid remains and determining why the equids were at the site. As
mentioned in the previous section, it is quite difficult to differentiate between species of equids
that are in the same size range. It is simple to separate the larger wild horses (E. caballus ferus)
from the smaller wild asses (E. hydruntinus and E. hemionus) but it is much more difficult to
separate the hydruntines from the hemiones. Often, depending upon the degree of fragmentation,
it can be difficult to identify the equid element beyond Equus sp., as is evident in Figure 3. Of
course some elements will be more easily identified as equid than other elements, such as an

8
Proportion of identification of all equid remains
360, 33%
79, 7%
44, 4%
24, 2%
522, 47%
78, 7%
small-medium equidlarge equidEquus hemionusEquus caballusEquus speciesEquus hydruntinus
equid tooth fragment versus a long bone fragment. The identification of the cranial equid remains
at Çatalhöyük has been investigated successfully through tooth morphology (Martin and Russell,
In press, Russell and Martin 2005). The concept of ‘morphometry’ which implies that
morphology can be expressed through measurements is not yet widely used by all
zooarchaeologists (Eisenmann 1986: 68). An attempt to use morphometry at Çatalhöyük will be
discussed in the Methodology and Results chapters. Further attempts at identification of equid
species using cranial and postcranial remains are presented in the Methodology chapter and
Results chapter.
Figure 3 - Equid remains identified at Çatalhöyük
Explaining why the different equid species were at Çatalhöyük in the first place is a much more
difficult and hypothetical task but has been tackled via several models presented in the next
chapter. Why the representation of equid species appears to change over time at Çatalhöyük is
also an interesting question, necessitating further inquiry into the Neolithic environment and
human subsistence strategies on the Konya Plain. Identifying whether the change in equid
species representation at Çatalhöyük occurred because of environmental change or the effects of
culture is very difficult to ascertain (Klein and Cruz-Uribe 1984: 77) but should be queried to
further understand the Neolithic inhabitants of Çatalhöyük and their relationship with the equids
of the region. How much knowledge the inhabitants of Çatalhöyük had of equid biology and
behaviour is another query that will be discussed in the following chapters.

9
1.6 Summary
The environment of the Konya Plain where Çatalhöyük is located has been described in the
previous pages as fairly hospitable during the Neolithic where hunting of large herbivores
(including equids), pastoralism and agriculture all added to the economy. The various aspects of
the three equid species recovered from Neolithic Çatalhöyük have been discussed along with a
description of equid remains identified at other sites in the Near East and Anatolia. Problems
with the equid remains at Çatalhöyük have been discussed and it has been proposed that more
work must be done to identify the species of the equid remains at Çatalhöyük and determine the
different roles of equid species and possible changes through time of these roles as resulting from
a changing environment or economic practices.

10
2) Çatalhöyük and Potential Models for Equids
2.1 Background to Çatalhöyük
Çatalhöyük is a large village site located in central Anatolia on the edge of the Carsamba alluvial
fan on the Konya Plain (Figure 4). Çatalhöyük translates as ‘mound at a road-fork’ in Turkish
and is represented by two mounds, the east mound covering 13.6 ha (Cessford 2005: 323) with a
maximum height of about 20 m (Figure 5) (Mellart 1998:35). The site was occupied from the
mid 9th millennium bp to the early 8th millennium bp. These dates have recently been confirmed
by new Accelerator Mass Spectrometer (AMS) radiocarbon dates but it must be kept in mind that
the earliest occupation may not have yet been located due to the nature of such a large mounded
site (Cessford 2001: 724). The village consisted of tightly spaced mud brick houses containing
various features such as ovens, storage bins, sleeping platforms, burials and wall art (Mellart
1998, Hodder and Matthews 1998). It is estimated that the Neolithic population of Çatalhöyük
was around 3500 to 8000 individuals (Cessford 2005: 326).
Figure 4 - Location map of Çatalhöyük in Turkey (Adapted from
http://catal.arch.cam.ac.uk/2005).
A distributary of the Carsamba River flowed between the Çatalhöyük east and west mounds prior
to the construction of modern artificial drainage systems (Roberts et al. 1999:623). The relatively

11
moist environment of Neolithic Çatalhöyük was much different than today’s steppe environment.
The Konya plain is not believed to have been densely settled during the Neolithic and some see
the abandonment of Çatalhöyük East by the end of the 8th millennium as the result of movement
to Çatalhöyük West and not a result of a changing environment creating harsh living conditions
(Thiessen 2001:38).
Figure 5 - Plan of Çatalhöyük East Mound showing excavation areas (Adapted from
Richards et al. 2003: 68).
In this context of a seasonally flooded alluvial environment, the first sedentary and agricultural
communities settled and exploited the Konya Plain (Roberts et al. 1999:625, Baird 2002:151). It
has been argued that the Neolithic community of Çatalhöyük extensively exploited the territory of
the Konya Plain on a seasonal basis, incorporating resources from the marshes, flood plain, hills
and steppe into their subsistence system (Asouti and Fairbairn 2002: 188). The environment

12
surrounding Çatalhöyük, though always changing, provided pasture, food, fodder, groundstone
materials, and fuel for the community. Vegetation during the Neolithic appears to have consisted
of wild fruit trees amongst grassland and the maximum expansion of oak woodland around 8000
BP (Woldring and Bottema 2001:4). This environment would have been ideal grazing grounds
for many large herbivores including aurochs, sheep, goats, several species of equids and cervids.
Researchers have been able to reconstruct the ancient wood land of south central Anatolia
through analysis of charcoal recovered from Çatalhöyük and, Pinarbasi, a prehistoric campsite
(Asouti and Hather 2001, Asouti 2003). It is evident through several charcoal studies,
palynological work, and archaeobotanical analysis that the area surrounding Çatalhöyük would
have provided plenty of wild plant foods for human and animal diets and for fuel purposes
(Asouti and Hather 2001). The Konya Plain was plainly a very hospitable environment for
humans exploiting the area during the Neolithic.
While it has been previously argued that Çatalhöyük was primarily a hunter-gatherer/pastoralist
society with no reliance on agriculture, the archaeobotanical evidence so far indicates otherwise
(Asouti and Fairbairn 2002:181). It appears to be fairly evident that agriculture was providing a
portion of the food resources for the inhabitants of Çatalhöyük though the proportion is uncertain.
Perkins (1969) believed that cattle were domesticated at Çatalhöyük and provided a large portion
of the food resources for the inhabitants. This view has subsequently been dismissed as
systematic screening and constant sampling has revealed that sheep (Ovis sp.) and goat (Capra
sp.) were the most prevalent and likely domesticated animals at Çatalhöyük and cattle (Bos sp.)
were most likely wild and hunted in a similar vein to the other mammals found at the site
including boar (Sus scrofa), deer (Cervus elephus, Dama, and Capreolous capreolous) and equids
(E. caballus ferus, E. hydruntinus and E. hemionus) (Martin and Russell, in press, Russell and
Martin 2000, 2005).
Many archaeologists still argue that hunting was the main element of subsistence during the Pre-
Pottery Neolithic period at Çatalhöyük and at other Neolithic sites such as Aşikli Höyük (Esin
and Harmankaya 1999:119), and the upper horizons of Cayonu (Ozdogan 1997:9, 1999:39). The
chipped stone technology, primarily obsidian based, has been proposed to have developed over
time in response to changes in subsistence and ecology (Conolly 1999:799). The faunal data
appears to also reflect changes in subsistence strategies, perhaps in response to environmental
change, with a greater reliance on domestic sheep and goat over time and less utilization of
hunted wild animals such as equids, cervids and aurochs (Russell and Martin 1998, 2005).

13
Storage, social complexity and artwork are all present at Çatalhöyük and have been documented
extensively (Mellart 1966, 1967, 1975, Hodder 1999, 2004). Stable isotope evidence retrieved
from human bone collagen in burials at Çatalhöyük indicates that a mix of animal protein and
plants provided the bulk of the diet during the Neolithic period (Richards et al. 2003: 71). It is
proposed that the majority of animal protein in the diet comes from sheep (Ovis sp.) and goat
(Capra sp.) which correlates with the faunal remains at Çatalhöyük (Ibid). Using stable isotope
analysis it would be difficult to determine what proportion of the diet is from wild animals such
as equids (Equus sp.), cattle (Bos sp.) and boar (Sus scrofa).
Clearly the inhabitants at Çatalhöyük had a thriving economy with a multi-faceted subsistence
strategy involving hunting, gathering, pastoralism and agriculture. Understanding the climatic
sequence is important for understanding the Neolithic in the eastern Mediterranean region as it
coincided with a period of maximum moisture availability much different than the drier
environment experienced by early Bronze Age cultures (Figure 6) (Roberts et al 2001:735).
Changes in the environment would have had interesting effects on the inhabitants of Çatalhöyük
causing alterations in subsistence strategies that may be visible archaeologically. Perhaps
changes in equid utilization may be one of these alterations that are visible through analysing the
faunal remains retrieved from the site.
Figure 6 - Artist’s depiction of Neolithic Çatalhöyük (Adapted from
http://catal.arch.cam.ac.uk/2005).

14
In examining the distribution of equid remains from Çatalhöyük, Martin and Russell (in press)
propose that equids may have been processed in a different manner than other animals due to the
fact that equid remains are often recovered from midden contexts. Also they note that due to the
fact that equids are not usually installed as part of house structures at Çatalhöyük, this may also
account for their absence from house contexts. The majority of equid remains have been
recovered from the south area of Çatalhöyük and the equid proportion of the faunal assemblage
seems to taper off through time with more equid remains being recovered from the earlier levels
(Martin and Russell, in press). This may be due to sampling strategies as mentioned previously
and the smaller amount of midden sampling at Çatalhöyük over the time of its excavation. While
E. caballus ferus is consistent in small numbers at Çatalhöyük, E. hydruntinus appears to be the
most common equid recovered with an increase in the presence of E. hemionus through time
(Martin and Russell, in press, Russell and Martin 2005: 57).
It is uncertain as to what became of the equid populations at Çatalhöyük and on the Konya Plain
but one possible explanation may be sought in a following section titled ‘Ecosystem Model’. It is
generally thought that the environment changed and became inhospitable to wild equid species
prior to the introduction of domesticated equids such as E. caballus and E. asinus. While humans
most definitely had an effect upon the Neolithic environment through agriculture, pastoralism and
hunting, it is uncertain as to how much change for which they were responsible.
2.2 Opportunistic hunting
It would appear that many of the equid remains identified at Çatalhöyük arrived there as a result
of hunting. Due to the environment of the Konya Plain and its attractions to large herbivores and
humans, contact between species must have occurred on a fairly regular basis during the
acquisition of food and water. Though Çatalhöyük appears to have been somewhat reliant upon
hunted and gathered foods in the Neolithic, agriculture was also important to the economy as is
reflected in the archaeobotanical remains recovered in the lower levels (Asouti and Fairbairn
2002). It has been suggested that equids may have been hunted for two reasons, to protect cereal
crops, and of course as food (Uerpmann 1982:46). This could apply to other large herbivores that
would also compete with humans for the cereal crops including deer and aurochs. Hunting equids
in this manner would seem to follow an opportunistic model of food acquisition, relying upon the
equids taste for cereal crops to bring the animals within range of hunters or perhaps equids were
hunted when encountered during the gathering of other foods and resources (such as obsidian,

15
timber). It is proposed that most hunting techniques are opportunistic (Levine 1990: 728),
relating to either the hunted taxa being opportune or the condition of the animal (age and health).
The lack of iconography depicting equids at Çatalhöyük as compared to aurochs seems to indicate
that the equids were not regarded to be important symbolically (Martin and Russell: in press) so
this lends doubt that they were hunted for this purpose alone.
2.3 Logistical hunting
As suggested in the previous section, it is highly doubtful that equids were sought out for purely
iconographic or symbolic reasons due to the lack of artwork involving equids and absence of the
inclusion of equid remains in building structures or installations. There has been one painting
recovered that seems to include equids and several carvings (Mellart 1966) but the symbolic role
of large herbivores seems to have been largely assumed by aurochs as reflected by the numerous
horn core and buchranium installations found in the buildings at Çatalhöyük. As a food resource,
equids may have been specifically sought after by the inhabitants of Çatalhöyük, particularly
during periods when domestic animals could not be harvested for fresh meat (rutting or birthing
season) or periods of resource stress. Perhaps equids were hunted due to qualities of their flesh
such as taste or leanness or maybe equids were simply relatively easy to hunt as compared to
other non-herding herbivores such as isolated cervids. Assuming that equids were a preferred
food resource, perhaps hunting groups made specific forays to obtain equid meat. This would
indicate logistical hunting as planning would be needed for processing, transportation and the
organization of the hunting party (Binford 1978). Storage may also have been an issue if the
equids were obtained at some distance. Stallions have a tendency to defend their mare groups by
attacking potential threats, and consequently fall victim to hunters first thus leaving the mare
group open to further attacks (Uerpmann 1987). This protective tendency of equid groups could
have been an attraction to Neolithic hunters seeking a large quantity of animal protein.
2.4 Ecosystem Model
Through analysis of the Neolithic environment and palaeoecology, it may be possible to
determine what caused changes in equid populations and distributions throughout Neolithic
Anatolia. The effects of humans on the ecology of the Konya Plain in particular could perhaps
lead to a better understanding of fluctuations in equids and their utilization by people. Woldring
and Bottema (2001) have studied vegetation history in east-central Anatolia through pollen

16
evidence from Eski Acigöl and have found that although there was an apparent increase in
precipitation in the area ca. 8600 B.P., there was still an increase in xeric plant communities
which indicates increasing aridity. This is explained by several ideas including; stronger
continentality in central Anatolia; an increase in total evaporation as a result of the expanding
steppe forest; changes in vegetation due to interference from humans and domestic animals (Ibid:
28). Clearly, these changes in the environment of Neolithic central Anatolia would have had a
profound effect upon the equid populations on the Konya Plain. As mentioned earlier in the
section on equid biology, changes in the environment relating to water sources, competition for
food and increased predation would have all had significant effects upon the equid populations
(Saltz 2002) causing changes to the distribution and representation of the three species (E.
caballus ferus, E. hemionus and E. hydruntinus) on the Konya Plain. Unfortunately, at this point
in time, it is difficult to correlate the changes in the environment during the Neolithic with
changes in equid populations due to the relative coarseness of environmental data and small
sample size of equid remains. Perhaps as more environmental work is completed and the equid
sample size increases at Çatalhöyük and other sites in Anatolia, an ecosystem model as discussed
by Coughenour (2002), could be used to illustrate the changes in equid populations as related to
the environment.
The idea of species is a contested and often misunderstood concept with many believing that one
species may not breed with another species or if that this occurs, the offspring will be infertile
(Groves 2002: 95). Groves considers the Biological Species Concept (BSC) as put forward by
Mayr (1942) and notes the quote ‘that species do not interbreed under natural conditions’ which
implies that under unnatural conditions, species may interbreed and produce fertile offspring.
The true concept of species is that a taxon is 100 percent different than another taxon (but does
not imply that two taxa cannot interbreed). All equids can be interbred in captivity (Klingel
1974: 124) which can be considered to be an unnatural environment. This is a simplification of
the issue but presented in order to point out that in an environment that is being altered by
humans an ‘unnatural’ environment is created. In this ‘unnatural’ environment, perhaps,
hypothetically speaking, two taxa such as E. hydruntinus and E. hemionus could interbreed
causing the apparent extinction of one taxon, namely E. hydruntinus. While little is known about
the extinction of E. hydruntinus, it is often assumed that hunting and/or environmental change
caused its demise. Another option however, is to combine the ecosystem model (assuming
environmental conditions worsen) with the possibility that species will interbreed under unnatural
(or possibly harsh ?) conditions to explain the eventual disappearance of E. hydruntinus from

17
Anatolia and the Near East. Evidence supporting this idea could be sought in the general size
overlap of the two taxa (E. hydruntinus and E. hemionus) and morphological similarities in the
teeth (how many equid teeth cannot be identified beyond E. hemionus/E. hydruntinus?). A
genetic comparison of both taxa from the Pleistocene onwards could also shed light onto this
matter.
2.5 Effects of Taphonomy
Taphonomy is the ‘study of the transition, in all details, of organics from the biosphere into the
lithosphere or geological record’ (Lyman 1994:1). The taphonomy of the equid remains at
Çatalhöyük is important to consider when determining body-part distribution or the economic
utilization of the equids. How the equid remains were included in the formation of Çatalhöyük
can relay information concerning the utilization of equid remains and butchery or processing
techniques. The most obvious feature of equid remains at Çatalhöyük is the degree of bone
fragmentation. Most long bones have been fragmented prior to burial implying marrow
extraction. Another interesting feature is the lack of identified cut marks on the equid bones.
This may be the result of fragmentation masking the presence of cut marks but it is more likely
the butchery technique and use of extremely sharp chipped obsidian blades (Conolly 1999) that
has caused this lack of identified cut marks. Some equid bones have been burned but it is
uncertain whether this occurred during processing or after the event. The majority of the equid
remains appear to be moderately weathered. Although highly fragmented, it appears that the
entire equid skeleton is represented throughout the site with teeth and cranial fragments occurring
as often as post cranial fragments (Martin and Russell, in press). The length of fragments in the
equid size category (which also includes cattle (Bos sp.) and boar (Sus scrofa), size classes 7 and
5 respectively as according to the Çatalhöyük Faunal Database Codebook 2004) appears to fall
between 0 and 3 cm for the majority with most fragments belonging to the 2 cm length class.
This indicates that the larger animal remains were highly fragmented during processing, perhaps
for marrow extraction. The degree of weathering and bone modification is very similar among
the different equid species present at Çatalhöyük (E. caballus ferus, E. hemionus and E.
hydruntinus), though without identification of postcranial elements for the wild asses (E.
hemionus and E. hydruntinus), this is difficult to be certain.
Another issue that should be considered is that of the archaeological faunal assemblage and its
proximity to the true life assemblage of equid bones or the potential bone population that is

18
mostly affected by cultural practices (Meadow 1980). Ideally, one assumes that the
archaeological faunal assemblage is close to the potential bone population although it is well
known that this is rarely the situation. Beyond taphonomy and recovery practices affecting the
eventual archaeological assemblage, it should be noted that cultural practices involved in the
processing and transportation of the equid bones will be the most causative factors involved in the
eventual deposition and preservation of bones over time. While this is fairly intuitive, it must be
mentioned as this paper is concerned with only several taxa all belong to the Equidae family.
2.6 Summary
This chapter has sought to provide a background of the site of Çatalhöyük and potential models to
explain the presence of equid remains at the site. Discussion has also involved Çatalhöyük’s role
as a population centre with a complex economy and social structure as reflected by the tools,
artwork, buildings and food resources recovered through the archaeology that has taken place at
the site on and off since the 1960s. Models presented included opportunistic hunting and
logistical hunting. An ecosystem model to explain the changes in the presence/absence or
proportionate representation of the different equid species (E. caballus ferus, E. hemionus and E.
hydruntinus) at Çatalhöyük has also been explored. Finally, taphonomy was discussed and issues
addressed included how taphonomy has affected the equid assemblage at Çatalhöyük and the
interpretation of the assemblage.

19
3) Methodology
3.1 Identifying the Species of the Equid Remains from Çatalhöyük
When determining the species of the equid remains recovered from Çatalhöyük, several different
methods were utilized to provide more accuracy and consistency during analysis. The
combination of morphological comparative techniques and metric techniques allowed for a wider
examination of cranial and post-cranial variables to create a greater possibility of species
identification. Morphological comparative methods utilized included those used by Uerpmann
(1986) for post-cranial elements and Payne’s (1991) morphological criteria used for equid teeth.
Metric techniques included the measurement of post-cranial elements as completed by Uerpmann
(1986, 1991) using Von Den Driesch’s (1976) system of standard measurements and the metric
analysis of teeth as devised by Payne (1991). An experimental morphological technique to
determine species using the first phalanx in the wild-asses (E. hydruntinus and E. hemionus) was
also investigated.
The equid element measurements taken from Çatalhöyük have been compared to results from
several publications with attention to E. hydruntinus, E. hemionus and E. caballus ferus (Bonifay
1991, Turnbull 1986, Uerpmann 1986, Zeder 1986). Equid remains selected for analysis include
the astragalus, phalanx I, phalanx II, phalanx III, scapula, pelvis and tibia. These elements were
concentrated on as they are the most common equid post cranial elements to survive in
archaeological sites and have remained most complete due to their density thus offering more
comparable sets of measurements. Basic statistics such as mean and range were used to compare
the measurements of the elements that were analysed from Çatalhöyük to the elements measured
by the researchers listed above.
3.2 Size of Element
In sum, the most common method of determining the species of equids relied upon the
assumption that wild horses (E. caballus ferus) are bigger than wild-asses (E. hydruntinus and E.
hemionus) and there have been varying ideas of which of the wild asses were bigger than the
other. In the end it appears that on average, the hydruntine is smaller than the hemione though
the overlap in size makes determination of species difficult when using this method (Uerpmann
1987). While this method is still useful for postcranial remains to separate the wild horse (E.

20
caballus ferus) from the wild-asses (E. hemionus and E. hydruntinus), this appears to be the limit
of its usefulness, though this will be tested in this thesis. Another problem with some postcranial
elements involves the case of the phalanges as it is difficult to determine whether one is
examining posterior or anterior phalanges with a small sample of elements. This compounds the
problem of identifying species using size as an indicator as there are differences in size and shape
between the anterior and posterior phalanges, particularly in the first phalanx. Unfortunately, no
fully articulated hydruntine skeletons have been recovered of yet to allow even a basic
comparison with hemione skeletons in collections. Therefore, any determination of species using
post-cranial elements will be speculative at this point, even with identification of hydruntine teeth
at a site.
3.3 Tooth Morphology
The most commonly used method of identifying the hydruntine (E. hydruntinus) is examination
of the tooth morphology. The most distinctive features include the ‘deep penetration of the
external fold in the lower molars, and the triangular or ‘shoe’-shaped protocone in the upper
teeth’ (Davis 1980:295). The hemione (E. hemionus) has typical tooth morphology
characteristics which include an ‘oval protocone with concave interior surface, and a non-
penetrating external fold in the lower molars’ (Ibid: 297). The wild horse (E. caballus ferus) is
usually identified by the mandibular teeth through a u-shaped lingual fold and the presence of an
extra small caballine fold on the buccal fold while the maxillary teeth are identified as E. caballus
ferus by an elongated an narrow protocone and presence of a caballine fold at the base of the
protocone (Martin and Russell, in press). This method was used to identify the presence of E.
hydruntinus, E. caballus ferus and E. hemionus at Çatalhöyük (Martin and Russell, in press,
Russell and Martin 2005).
3.4 Tooth Morphometrics
The results of the tooth morphology investigation were expanded further by conducting a metric
analysis of the teeth that were identified to species by Martin and Russell (in press). The
measurements were selected from those proposed by Payne (1991) and Buitenhuis (2005) and are
presented in a table as Appendix 1 and Appendix 2. The measurements selected for the
mandibular teeth include crown height and from Payne (1991:135, Appendix 3); OL (#1), B3
(#2), B4 (#3), Lnd (#4), LF (#5) and from Buitenhuis (2005b:26, Figure 7); Bei (#6) has been

21
modified to a rated scale 1 through 4 (Table 1) due to the difficulty in measuring this particular
feature using callipers, Measurement #7 is the Pli hypoconid which has been given a stage of
development on scale 1 through 4, and Measurement #8 is the Pli ptychostylid which has been
given a stage of development on a scale 1 through 4. The measurements selected for the
maxillary teeth include crown height, and from Payne (1991:135); OL (#1), Be (#2), LP (#3) and
from Buitenhuis (2003), the caballine fold has been given a scale from 1 through 4.
Figure 7 - Measurements of equid teeth, upper teeth (top), lower teeth (bottom) (Adapted
from Buitenhuis 2003: 25).
22
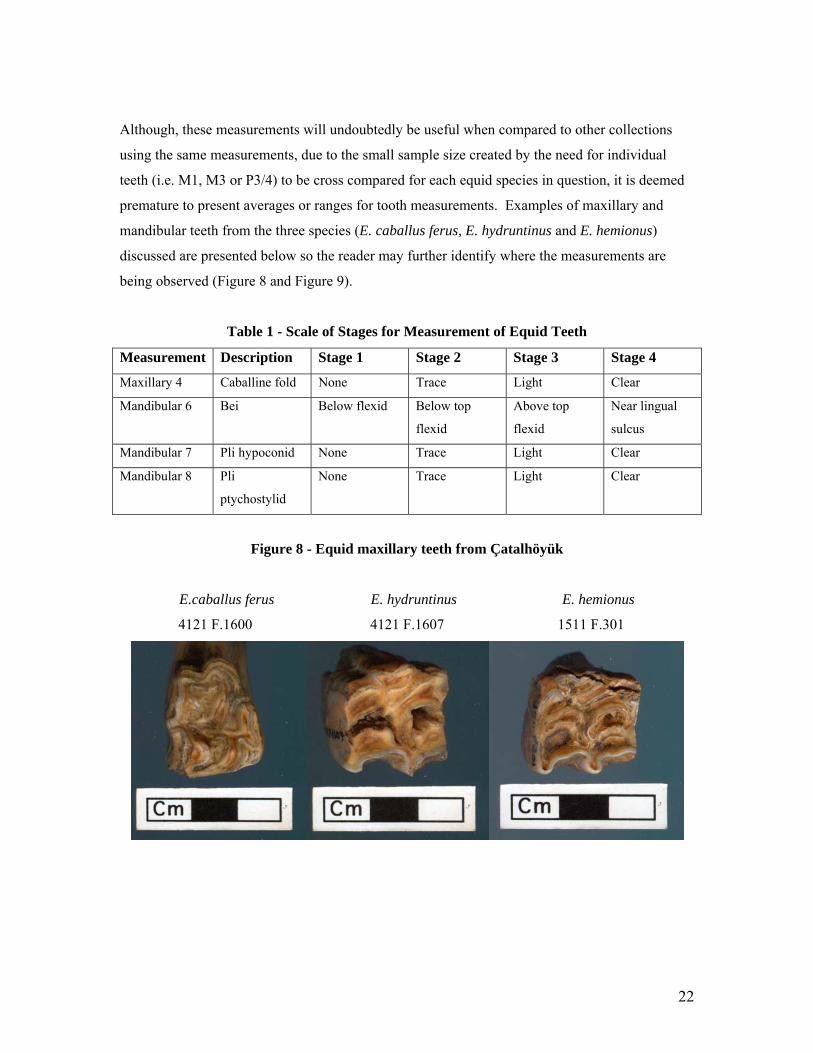
Although, these measurements will undoubtedly be useful when compared to other collections
using the same measurements, due to the small sample size created by the need for individual
teeth (i.e. M1, M3 or P3/4) to be cross compared for each equid species in question, it is deemed
premature to present averages or ranges for tooth measurements. Examples of maxillary and
mandibular teeth from the three species (E. caballus ferus, E. hydruntinus and E. hemionus)
discussed are presented below so the reader may further identify where the measurements are
being observed (Figure 8 and Figure 9).
Table 1 - Scale of Stages for Measurement of Equid Teeth
Measurement Description Stage 1 Stage 2 Stage 3 Stage 4
Maxillary 4 Caballine fold None Trace Light Clear
Mandibular 6 Bei Below flexid Below top
flexid
Above top
flexid
Near lingual
sulcus
Mandibular 7 Pli hypoconid None Trace Light Clear
Mandibular 8 Pli
ptychostylid
None Trace Light Clear
Figure 8 - Equid maxillary teeth from Çatalhöyük
E.caballus ferus E. hydruntinus E. hemionus
4121 F.1600 4121 F.1607 1511 F.301
23
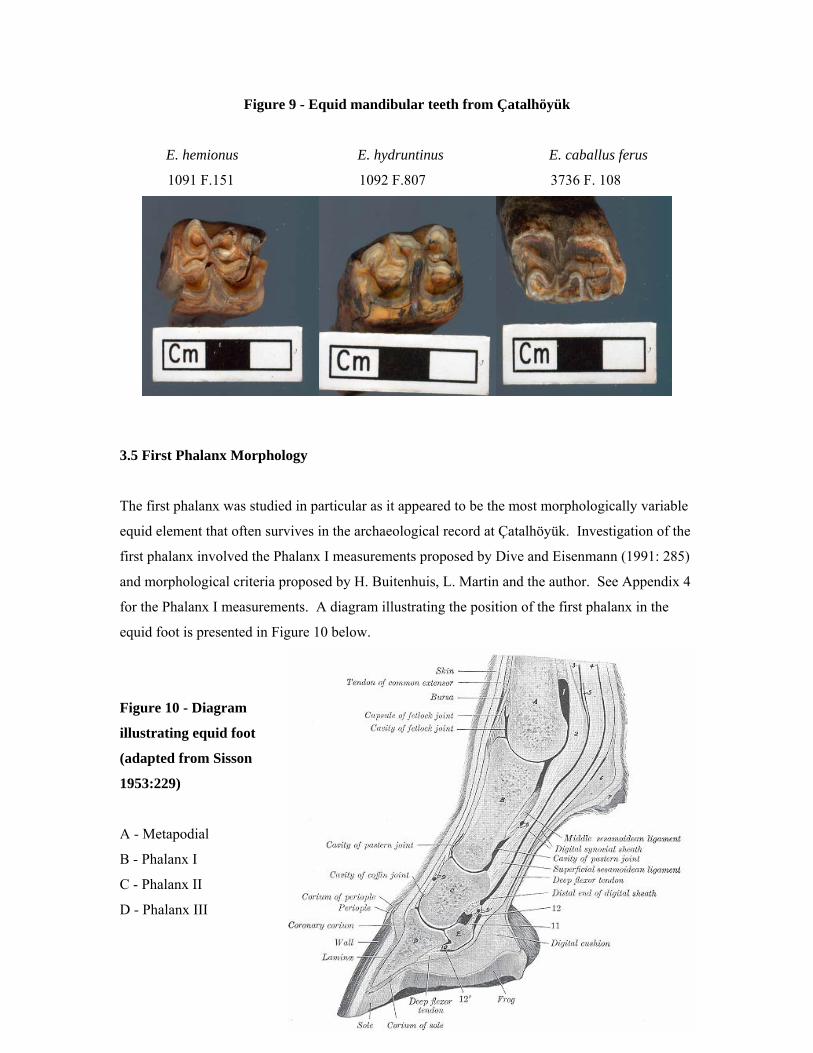
Figure 9 - Equid mandibular teeth from Çatalhöyük
E. hemionus E. hydruntinus E. caballus ferus
1091 F.151 1092 F.807 3736 F. 108
3.5 First Phalanx Morphology
The first phalanx was studied in particular as it appeared to be the most morphologically variable
equid element that often survives in the archaeological record at Çatalhöyük. Investigation of the
first phalanx involved the Phalanx I measurements proposed by Dive and Eisenmann (1991: 285)
and morphological criteria proposed by H. Buitenhuis, L. Martin and the author. See Appendix 4
for the Phalanx I measurements. A diagram illustrating the position of the first phalanx in the
equid foot is presented in Figure 10 below.
Figure 10 - Diagram
illustrating equid foot
(adapted from Sisson
1953:229)
A - Metapodial
B - Phalanx I
C - Phalanx II
D - Phalanx III

24
The morphological criteria involve the application of attributes which appear to separate the wild
ass (E. hemionus and E. hydruntinus) first phalanges into two groups. The attributes selected are
identifiable visually and occur to some degree on all the first phalanges studied. The attributes
selected for comparison between the two wild asses (E. hemionus and E. hydruntinus) will be
discussed below.
Each attribute was given a designation, namely A through H. Attribute A concerns the depth of
the sulcus which is present on the posterior side of the proximal articulation of the Phalanx I.
While not always definitive, this attribute ranges from ‘deep’ in some elements to ‘shallow’ in
others. The photo below (Figure 11) illustrates two first phalanges examined during this study
with the first phalanx on the left exhibiting a shallow sulcus and the first phalange on the right
exhibiting a deep sulcus.
Figure 11 - Attribute A - Depth of Sulcus (Id #s 7880 F242, 4465 F121).
Attribute B concerns the length of the trigonum phalangis as this feature on the posterior of first
phalanges appears to differ in length between the elements investigated. Attribute C is concerned
with the roundness of the distal articular surface, whether it appears to be rounder or flatter. An
example of Attribute C is illustrated in the photograph below (Figure 12). The first phalanx on
the left is an example of a rounder distal articulation while the one on the right is an example of a
flatter distal articulation.

25
Figure 12 - Attribute C - Roundness of Distal Articular Surface (Id #s 1668 F325, 1505
F222).
Attribute D concerns the position of the supratuberosity on the lateral/medial side of the first
phalanges. The supratuberosity appears to be more anterior or more posterior depending upon the
first phalanx examined. Attribute E is the shape of the distal trigonum phalangis and whether it is
raised or smooth. Attribute F is the shape of the proximal articulation and ranges from a squarish
shape to a rounded half-moon shape. Attribute F is illustrated in the photograph below (Figure
13). The first phalanx on the left is an example of the rounded half-moon shape while the one on
the right is an example of the squared shape.
Figure 13 - Attribute F - Shape of Proximal Articulation (Id #s 4836 F1, 1505 F222).
26
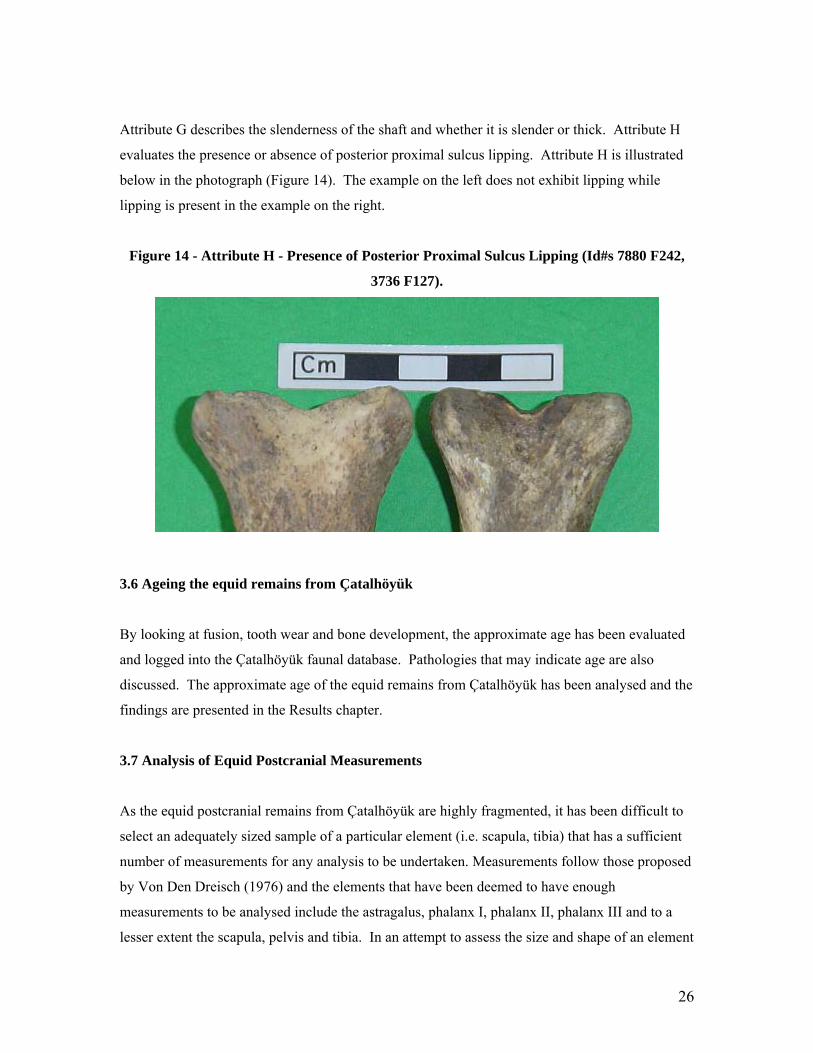
Attribute G describes the slenderness of the shaft and whether it is slender or thick. Attribute H
evaluates the presence or absence of posterior proximal sulcus lipping. Attribute H is illustrated
below in the photograph (Figure 14). The example on the left does not exhibit lipping while
lipping is present in the example on the right.
Figure 14 - Attribute H - Presence of Posterior Proximal Sulcus Lipping (Id#s 7880 F242,
3736 F127).
3.6 Ageing the equid remains from Çatalhöyük
By looking at fusion, tooth wear and bone development, the approximate age has been evaluated
and logged into the Çatalhöyük faunal database. Pathologies that may indicate age are also
discussed. The approximate age of the equid remains from Çatalhöyük has been analysed and the
findings are presented in the Results chapter.
3.7 Analysis of Equid Postcranial Measurements
As the equid postcranial remains from Çatalhöyük are highly fragmented, it has been difficult to
select an adequately sized sample of a particular element (i.e. scapula, tibia) that has a sufficient
number of measurements for any analysis to be undertaken. Measurements follow those proposed
by Von Den Dreisch (1976) and the elements that have been deemed to have enough
measurements to be analysed include the astragalus, phalanx I, phalanx II, phalanx III and to a
lesser extent the scapula, pelvis and tibia. In an attempt to assess the size and shape of an element

27
or feature of an element for the purpose of analysis, measurements that attempt to record these
were selected (e.g. width versus length). These measurements were then plotted against each
other in scatter plots. These scatter plots and their analysis will be presented in the Results
chapter. Any extreme outliers that appeared on the scatter plots were investigated and
remeasured. Any bone that was burnt or very weathered was also excluded from this analysis.
Young, unfused or fusing bones were also again excluded from analysis. For the purposes of
analysis, it was vital that factors such as age, weathering and modification did not skew the
results of the investigation. The data resulting from this analysis are presented as Appendix 4 and
Appendix 5.

28
4) RESULTS
4.1 Representation of Equid Species
Using the morphological criteria of teeth for identifying equid species as proposed by Payne
(1991), it was possible to identify three equid species at Çatalhöyük including E. caballus ferus,
E. hydruntinus, and E. hemionus (Martin and Russell in press, Russell and Martin 2005). Metric
analysis of the equid teeth as proposed by Payne (1991) and Buitenhuis (2005) was also
conducted and these results are presented as Appendix 1 and Appendix 2. These metric analyses
are not conclusive in assigning teeth to species but perhaps may be used for future comparison of
equid teeth.
4.2 Metric Analysis of Equid teeth
Equid teeth that had been identified to species on the basis of morphology or that were potentially
identifiable to species were measured by Hijlke Buitenhuis and the author. The measurements
selected have been described in the Methodology chapter. Upon examination of the
measurements, it appears to be unfeasible, at this point, to identify patterns between the three
equid species as there are no identified teeth (i.e. maxillary M3, mandibular P3/P4) that are
represented by all three species of equid (E. caballus ferus, E. hemionus, and E. hydruntinus) at
Çatalhöyük. These measurements may, however, be useful for comparison with other equid tooth
measurements taken at other sites in the future.
4.3 Analysis of Measurements
Postcranial elements were also investigated at Çatalhöyük for purposes of equid species
identification. The postcranial elements selected for study included the astragalus, phalanges I,
II, and III, scapula, pelvis, and tibia. Other elements were not utilized in this study due to small
sample size or lack of measurable areas on the elements. For most elements the measurable areas
follow Von Den Dreisch’s standard measurements (1976). All elements that had suitable
measurable areas for analysis were selected from the Çatalhöyük faunal database and re-measured
using a pair of digital callipers. All measurements are in millimetres and any measurements
differing by 0.5 mm or more and/or any abnormalities were corrected in the database. Scatter
plots were created by plotting applicable measurements against one another to create a range of

29
sizes for each element that could be analysed for patterns or irregularities. Any element that
appeared to be outside of the normal distribution was examined carefully and was either included
or excluded from the study due to various reasons. Elements could be excluded for reasons such
as taphonomy (burning, extreme weathering), age (unfused or fusing elements were not included)
and errors in database recording (wrong number entered in database). Re-examining the elements
being studied from the database was necessary to control for outliers and increase accuracy which
was deemed to be necessary for the task at hand. Factors such as overlap in size ranges between
the hemione and hydruntine and lack of knowledge of posterior or anterior phalanges necessitated
that all measurements and prior analysis of equid elements be accurate. Problems associated with
the metrical analysis included a small sample size, a lack of a comparative equid collection for
interpreting trends and the necessity of keeping the anterior and posterior phalanges in the same
analytical category due to the presence of similarly sized E. hydruntinus and E. hemionus.
It was hoped that the size of elements via metrics could be used to separate the larger horses (E.
caballus ferus) from the smaller wild asses (E. hydruntinus and E. hemionus). This method only
applied to separating the wild horses (E. caballus ferus) from the wild-asses (E. hydruntinus and
E. hemionus), as the size difference between the hydruntine and hemione is negligible. This
division between species (i.e. between E. caballus ferus and E. hydruntinus and E. hemionus) can
be easily identified in the graphs presented for all elements investigated. While it is currently
impossible to separate the E. hydruntinus from the E. hemionus in these scatter plots, it would
perhaps be beneficial to compare these measurements with known E. hemionus collections
(something that the author was unable to undertake).
Astragalus
Measurements selected for analysis of the astragalus include Measurement 8 (Greatest Breadth)
and Measurement 7 (Greatest Height) as presented in the scatter plot below (Figure 15). These
measurements are described in the Çatalhöyük Faunal Database Codebook 2004 and the
corresponding measurements described by Von Den Dreisch (1976) are presented in the
parentheses. It would appear in the scatter plot below that there is one element at the far top right
corner much larger than the other elements. The rest of the elements seem to be grouped in the
smaller size range on the left side of the chart with no obvious separation appearing in this group.
This group which does not appear to divide any further most likely belongs to the wild asses (E.
hemionus and E. hydruntinus) due to their relative size and homogeneity. Though there are a few
outliers in this group, this could be due to age or size range within the group. The isolated
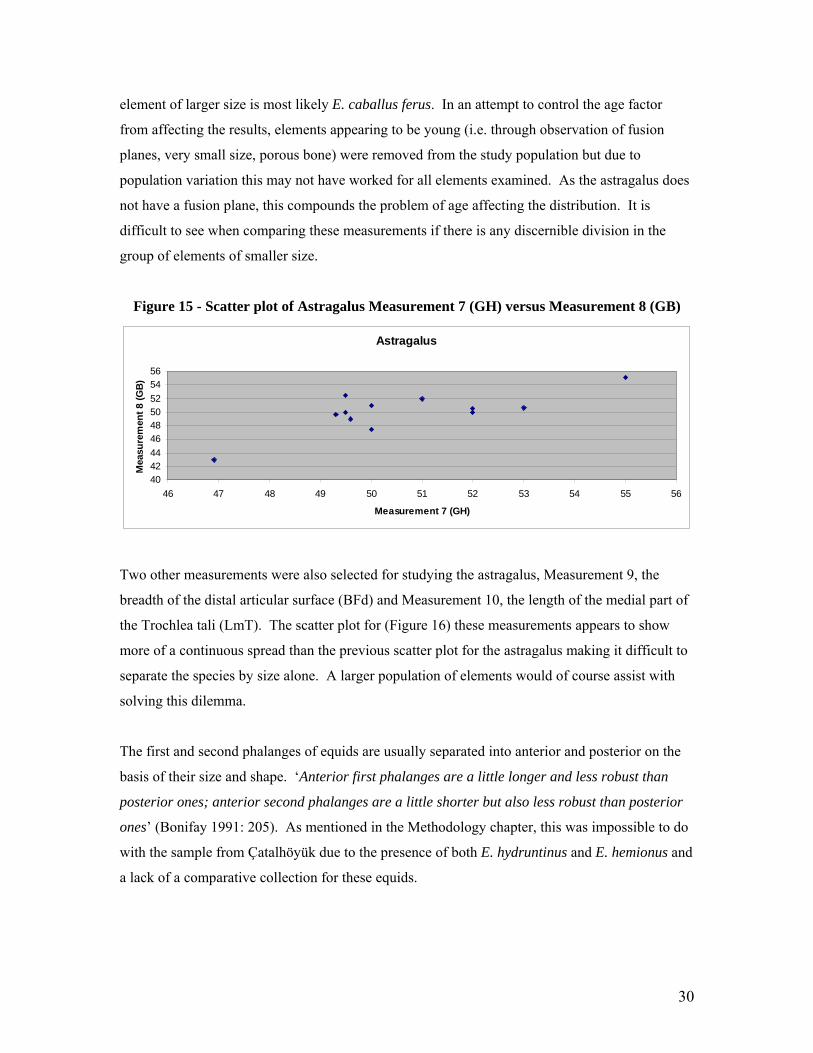
30
element of larger size is most likely E. caballus ferus. In an attempt to control the age factor
from affecting the results, elements appearing to be young (i.e. through observation of fusion
planes, very small size, porous bone) were removed from the study population but due to
population variation this may not have worked for all elements examined. As the astragalus does
not have a fusion plane, this compounds the problem of age affecting the distribution. It is
difficult to see when comparing these measurements if there is any discernible division in the
group of elements of smaller size.
Figure 15 - Scatter plot of Astragalus Measurement 7 (GH) versus Measurement 8 (GB)
Astragalus
404244464850525456
46 47 48 49 50 51 52 53 54 55 56
Measurement 7 (GH)
Mea
sure
men
t 8 (G
B)
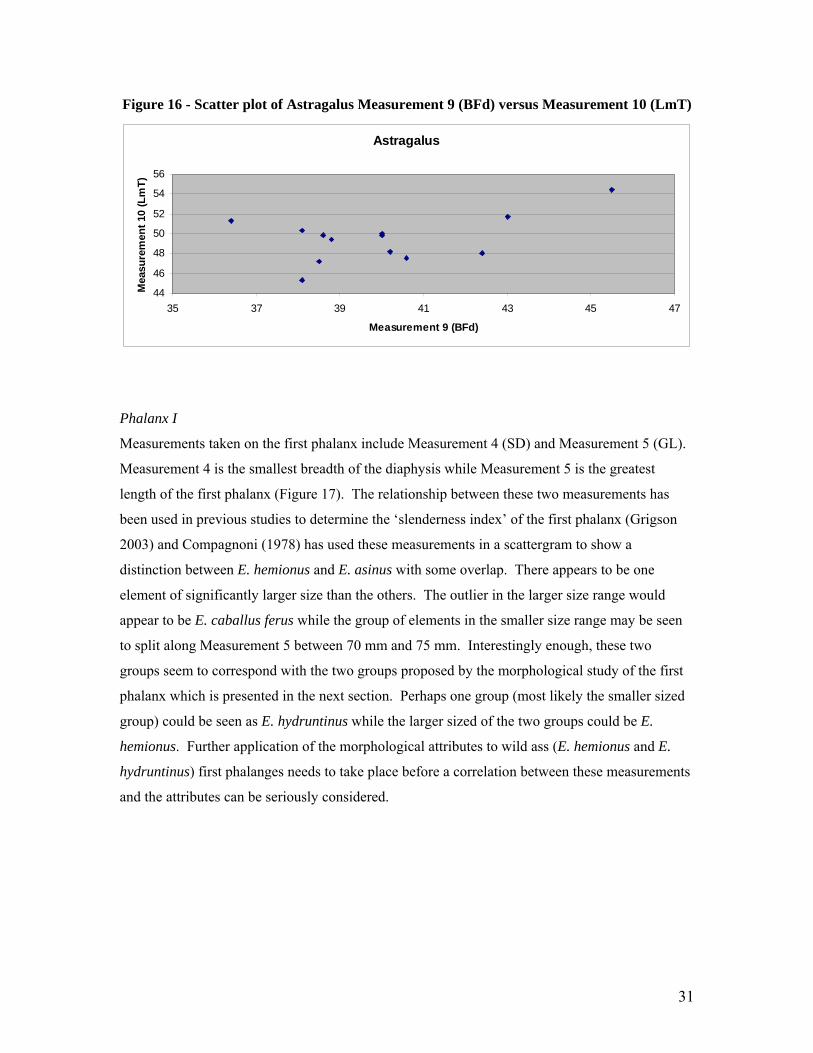
Two other measurements were also selected for studying the astragalus, Measurement 9, the
breadth of the distal articular surface (BFd) and Measurement 10, the length of the medial part of
the Trochlea tali (LmT). The scatter plot for (Figure 16) these measurements appears to show
more of a continuous spread than the previous scatter plot for the astragalus making it difficult to
separate the species by size alone. A larger population of elements would of course assist with
solving this dilemma.
The first and second phalanges of equids are usually separated into anterior and posterior on the
basis of their size and shape. ‘Anterior first phalanges are a little longer and less robust than
posterior ones; anterior second phalanges are a little shorter but also less robust than posterior
ones’ (Bonifay 1991: 205). As mentioned in the Methodology chapter, this was impossible to do
with the sample from Çatalhöyük due to the presence of both E. hydruntinus and E. hemionus and
a lack of a comparative collection for these equids.
31
Figure 16 - Scatter plot of Astragalus Measurement 9 (BFd) versus Measurement 10 (LmT)
Astragalus
44
46
48
50
52
54
56
35 37 39 41 43 45 47
Measurement 9 (BFd)
Mea
sure
men
t 10
(Lm
T)
Phalanx I
Measurements taken on the first phalanx include Measurement 4 (SD) and Measurement 5 (GL).
Measurement 4 is the smallest breadth of the diaphysis while Measurement 5 is the greatest
length of the first phalanx (Figure 17). The relationship between these two measurements has
been used in previous studies to determine the ‘slenderness index’ of the first phalanx (Grigson
2003) and Compagnoni (1978) has used these measurements in a scattergram to show a
distinction between E. hemionus and E. asinus with some overlap. There appears to be one
element of significantly larger size than the others. The outlier in the larger size range would
appear to be E. caballus ferus while the group of elements in the smaller size range may be seen
to split along Measurement 5 between 70 mm and 75 mm. Interestingly enough, these two
groups seem to correspond with the two groups proposed by the morphological study of the first
phalanx which is presented in the next section. Perhaps one group (most likely the smaller sized
group) could be seen as E. hydruntinus while the larger sized of the two groups could be E.
hemionus. Further application of the morphological attributes to wild ass (E. hemionus and E.
hydruntinus) first phalanges needs to take place before a correlation between these measurements
and the attributes can be seriously considered.

32
Phalanx I
60
65
70
75
80
85
90
20 22 24 26 28 30 32 34 36
Measurement 4 (SD)
Mea
sure
men
t 5 (G
L)
Figure 17 - Scatter plot of Phalanx I Measurement 4 (SD) versus Measurement 5 (GL)
Other measurements selected for the first phalanx include Measurement 2 (Bp) and Measurement
7 (Dp). Measurement 2 is the greatest breadth of the proximal end and Measurement 7 is the
depth of the proximal end (Figure 18). The elements represented in this scatter plot appear to
follow the standard distribution that has appeared so far in the division between wild horses (E.
caballus ferus) and the wild assess (E. hemionus and E. hydruntinus). The two elements skewed
towards the larger size spectrum are most likely those of E. caballus ferus while the cluster of
elements at the smaller end of the size spectrum most probably belong to E. hemionus and E.
hydruntinus.
Figure 18 - Scatter plot of Phalanx I Measurement 2 (Bp) versus Measurement 7 (Dp)
Phalanx I
27
29
31
33
35
37
35 40 45 50 55 60
Measurement 2 (Bp)
Mea
sure
men
t 7 (D
p)
33
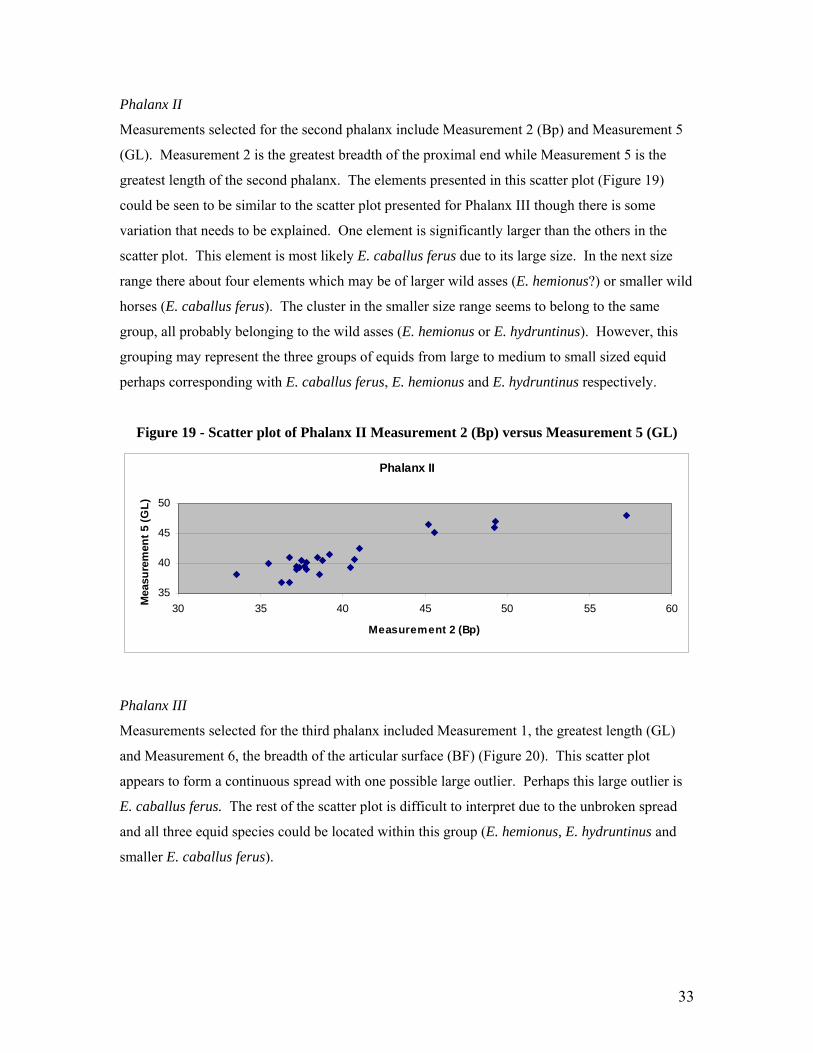
Phalanx II
35
40
45
50
30 35 40 45 50 55 60
Measurement 2 (Bp)
Mea
sure
men
t 5 (G
L)
Phalanx II
Measurements selected for the second phalanx include Measurement 2 (Bp) and Measurement 5
(GL). Measurement 2 is the greatest breadth of the proximal end while Measurement 5 is the
greatest length of the second phalanx. The elements presented in this scatter plot (Figure 19)
could be seen to be similar to the scatter plot presented for Phalanx III though there is some
variation that needs to be explained. One element is significantly larger than the others in the
scatter plot. This element is most likely E. caballus ferus due to its large size. In the next size
range there about four elements which may be of larger wild asses (E. hemionus?) or smaller wild
horses (E. caballus ferus). The cluster in the smaller size range seems to belong to the same
group, all probably belonging to the wild asses (E. hemionus or E. hydruntinus). However, this
grouping may represent the three groups of equids from large to medium to small sized equid
perhaps corresponding with E. caballus ferus, E. hemionus and E. hydruntinus respectively.
Figure 19 - Scatter plot of Phalanx II Measurement 2 (Bp) versus Measurement 5 (GL)
Phalanx III
Measurements selected for the third phalanx included Measurement 1, the greatest length (GL)
and Measurement 6, the breadth of the articular surface (BF) (Figure 20). This scatter plot
appears to form a continuous spread with one possible large outlier. Perhaps this large outlier is
E. caballus ferus. The rest of the scatter plot is difficult to interpret due to the unbroken spread
and all three equid species could be located within this group (E. hemionus, E. hydruntinus and
smaller E. caballus ferus).
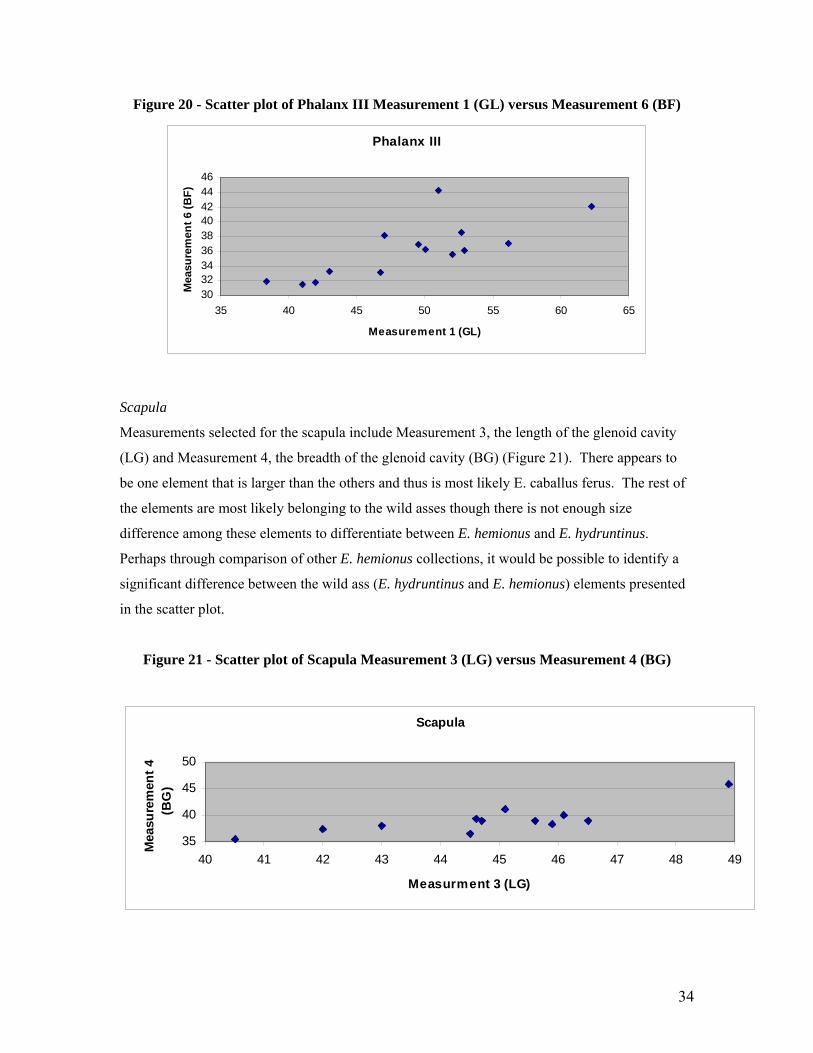
34
Phalanx III
303234363840424446
35 40 45 50 55 60 65
Measurement 1 (GL)
Mea
sure
men
t 6 (B
F)
Figure 20 - Scatter plot of Phalanx III Measurement 1 (GL) versus Measurement 6 (BF)
Scapula
Measurements selected for the scapula include Measurement 3, the length of the glenoid cavity
(LG) and Measurement 4, the breadth of the glenoid cavity (BG) (Figure 21). There appears to
be one element that is larger than the others and thus is most likely E. caballus ferus. The rest of
the elements are most likely belonging to the wild asses though there is not enough size
difference among these elements to differentiate between E. hemionus and E. hydruntinus.
Perhaps through comparison of other E. hemionus collections, it would be possible to identify a
significant difference between the wild ass (E. hydruntinus and E. hemionus) elements presented
in the scatter plot.
Figure 21 - Scatter plot of Scapula Measurement 3 (LG) versus Measurement 4 (BG)
Scapula
35
40
45
50
40 41 42 43 44 45 46 47 48 49
Measurment 3 (LG)
Mea
sure
men
t 4
(BG
)
35
Pelvis
40
45
50
55
60
45 50 55 60 65 70
Measurement 1 (LA)
Mea
sure
men
t 2 (B
read
th)
Pelvis
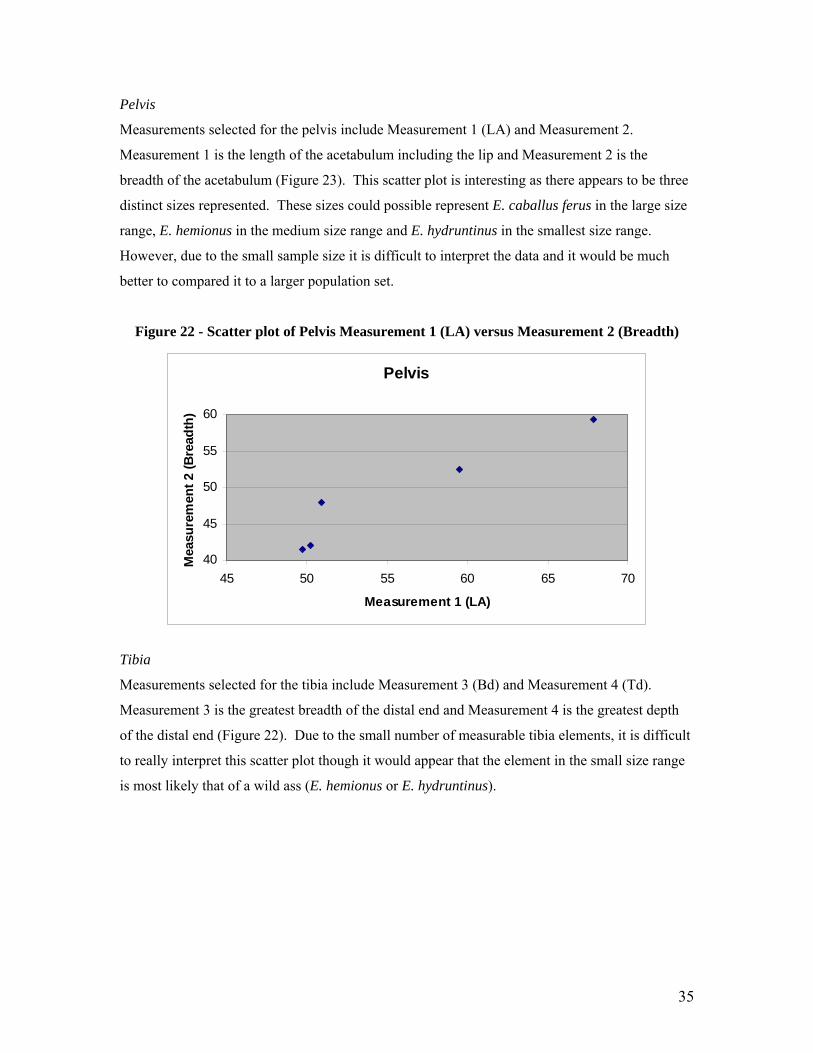
Measurements selected for the pelvis include Measurement 1 (LA) and Measurement 2.
Measurement 1 is the length of the acetabulum including the lip and Measurement 2 is the
breadth of the acetabulum (Figure 23). This scatter plot is interesting as there appears to be three
distinct sizes represented. These sizes could possible represent E. caballus ferus in the large size
range, E. hemionus in the medium size range and E. hydruntinus in the smallest size range.
However, due to the small sample size it is difficult to interpret the data and it would be much
better to compared it to a larger population set.
Figure 22 - Scatter plot of Pelvis Measurement 1 (LA) versus Measurement 2 (Breadth)
Tibia
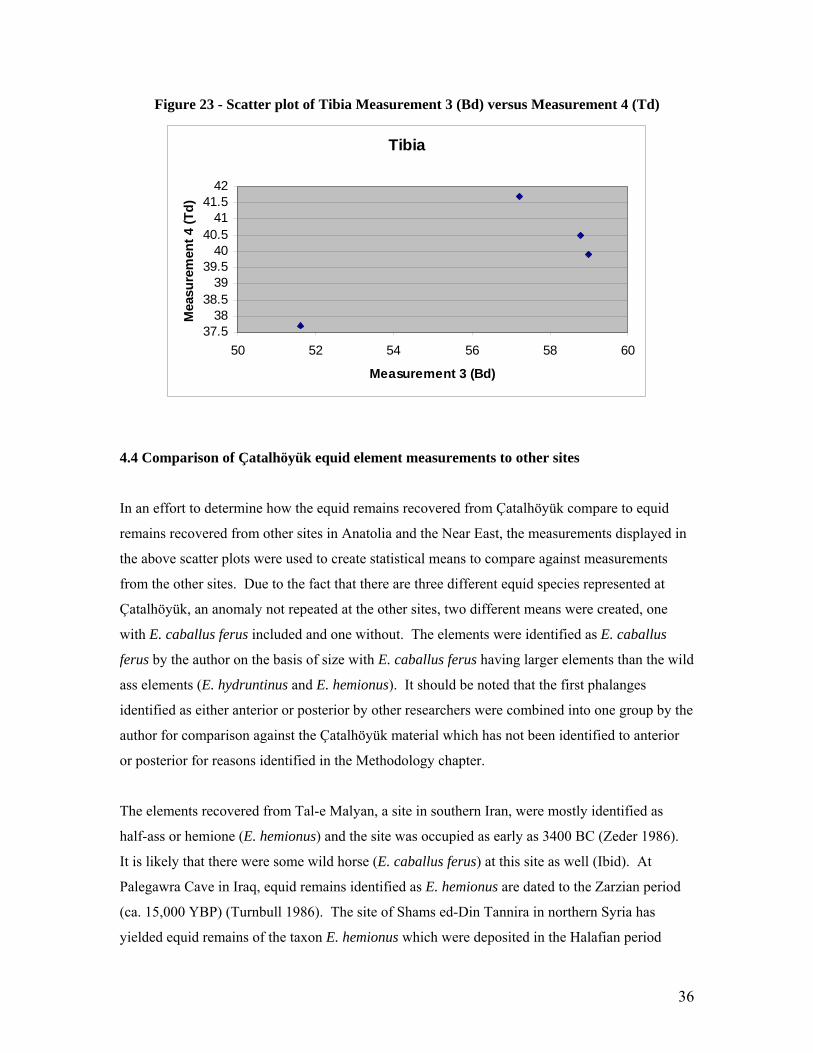
Measurements selected for the tibia include Measurement 3 (Bd) and Measurement 4 (Td).
Measurement 3 is the greatest breadth of the distal end and Measurement 4 is the greatest depth
of the distal end (Figure 22). Due to the small number of measurable tibia elements, it is difficult
to really interpret this scatter plot though it would appear that the element in the small size range
is most likely that of a wild ass (E. hemionus or E. hydruntinus).
36
Tibia
37.538
38.539
39.540
40.541
41.542
50 52 54 56 58 60
Measurement 3 (Bd)
Mea
sure
men
t 4 (T
d)
Figure 23 - Scatter plot of Tibia Measurement 3 (Bd) versus Measurement 4 (Td)
4.4 Comparison of Çatalhöyük equid element measurements to other sites
In an effort to determine how the equid remains recovered from Çatalhöyük compare to equid
remains recovered from other sites in Anatolia and the Near East, the measurements displayed in
the above scatter plots were used to create statistical means to compare against measurements
from the other sites. Due to the fact that there are three different equid species represented at
Çatalhöyük, an anomaly not repeated at the other sites, two different means were created, one
with E. caballus ferus included and one without. The elements were identified as E. caballus
ferus by the author on the basis of size with E. caballus ferus having larger elements than the wild
ass elements (E. hydruntinus and E. hemionus). It should be noted that the first phalanges
identified as either anterior or posterior by other researchers were combined into one group by the
author for comparison against the Çatalhöyük material which has not been identified to anterior
or posterior for reasons identified in the Methodology chapter.
The elements recovered from Tal-e Malyan, a site in southern Iran, were mostly identified as
half-ass or hemione (E. hemionus) and the site was occupied as early as 3400 BC (Zeder 1986).
It is likely that there were some wild horse (E. caballus ferus) at this site as well (Ibid). At
Palegawra Cave in Iraq, equid remains identified as E. hemionus are dated to the Zarzian period
(ca. 15,000 YBP) (Turnbull 1986). The site of Shams ed-Din Tannira in northern Syria has
yielded equid remains of the taxon E. hemionus which were deposited in the Halafian period
37
(Uerpmann 1986). At the cave site of Lunel-Viel in Herault, France, the Pleistocene remains of
E. hydruntinus were recovered (Bonifay 1991). The means of the measurements applicable to the
sample studied from Catalhoyuk are presented below (Table 2)
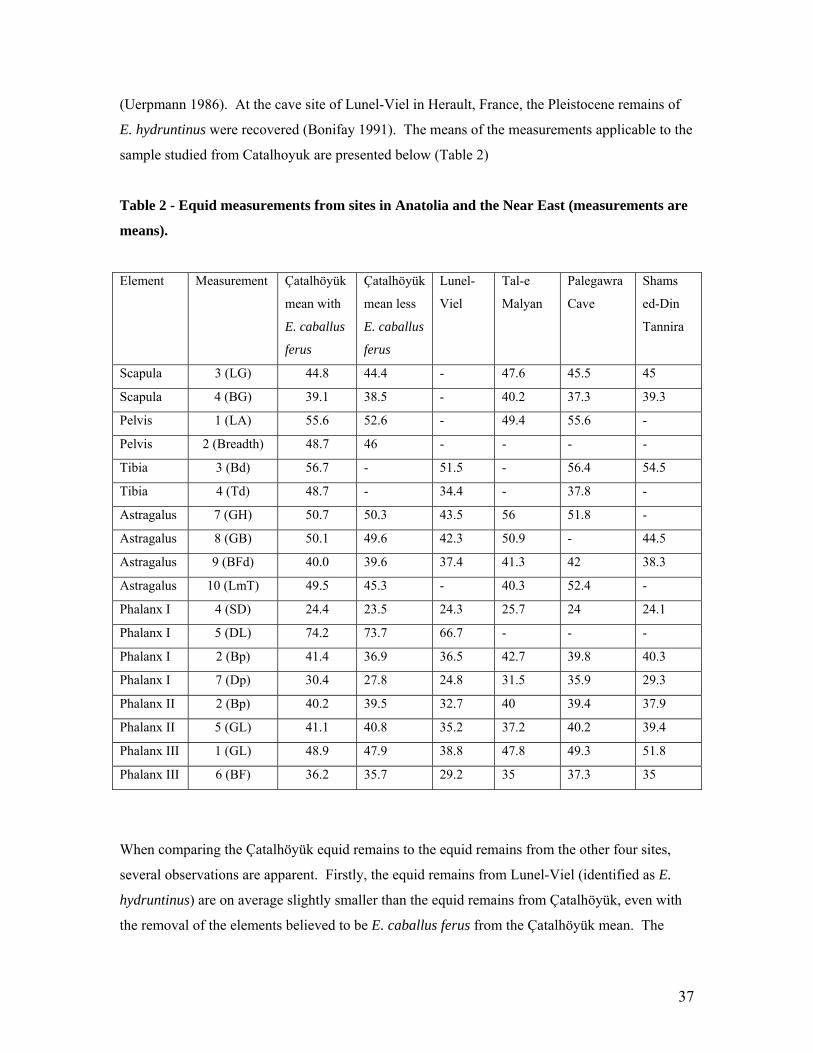
Table 2 - Equid measurements from sites in Anatolia and the Near East (measurements are
means).
Element Measurement Çatalhöyük
mean with
E. caballus
ferus
Çatalhöyük
mean less
E. caballus
ferus
Lunel-
Viel
Tal-e
Malyan
Palegawra
Cave
Shams
ed-Din
Tannira
Scapula 3 (LG) 44.8 44.4 - 47.6 45.5 45
Scapula 4 (BG) 39.1 38.5 - 40.2 37.3 39.3
Pelvis 1 (LA) 55.6 52.6 - 49.4 55.6 -
Pelvis 2 (Breadth) 48.7 46 - - - -
Tibia 3 (Bd) 56.7 - 51.5 - 56.4 54.5
Tibia 4 (Td) 48.7 - 34.4 - 37.8 -
Astragalus 7 (GH) 50.7 50.3 43.5 56 51.8 -
Astragalus 8 (GB) 50.1 49.6 42.3 50.9 - 44.5
Astragalus 9 (BFd) 40.0 39.6 37.4 41.3 42 38.3
Astragalus 10 (LmT) 49.5 45.3 - 40.3 52.4 -
Phalanx I 4 (SD) 24.4 23.5 24.3 25.7 24 24.1
Phalanx I 5 (DL) 74.2 73.7 66.7 - - -
Phalanx I 2 (Bp) 41.4 36.9 36.5 42.7 39.8 40.3
Phalanx I 7 (Dp) 30.4 27.8 24.8 31.5 35.9 29.3
Phalanx II 2 (Bp) 40.2 39.5 32.7 40 39.4 37.9
Phalanx II 5 (GL) 41.1 40.8 35.2 37.2 40.2 39.4
Phalanx III 1 (GL) 48.9 47.9 38.8 47.8 49.3 51.8
Phalanx III 6 (BF) 36.2 35.7 29.2 35 37.3 35
When comparing the Çatalhöyük equid remains to the equid remains from the other four sites,
several observations are apparent. Firstly, the equid remains from Lunel-Viel (identified as E.
hydruntinus) are on average slightly smaller than the equid remains from Çatalhöyük, even with
the removal of the elements believed to be E. caballus ferus from the Çatalhöyük mean. The

38
presence of E. hemionus remains which are thought to be larger than E. hydruntinus (Uerpmann
1987) is most likely the cause of this size difference represented by the means Secondly, the
remains from the other three sites, Tal-e Malyan, Palegawra Cave, and Shams ed-Din Tannira are
all bigger on average than the remains from Çatalhöyük. As the remains from these sites are
mostly E. hemionus with some E. caballus ferus, it would seem that it may be E. hydruntinus
remains that are lowering the Çatalhöyük mean. The equid remains from Çatalhöyük appear to
be unique in their average size, most likely due to the presence of the three species of equids.
While the size difference does not appear to be very great (usually only 1-2 mm), it is fairly
consistent for all elements from Çatalhöyük.
4.4 Morphological Attributes of First Phalanx
In an attempt to differentiate between the hemione and hydruntine, an experiment using
morphological attributes applied to the first phalanx was conducted. The first phalanx was
selected as it was a larger sample than other elements available and also appeared to have
morphological variety among the specimens examined. Wild horse first phalanges were
separated on the basis of size and a list of attributes was applied to the remaining ten first
phalanges that had been recorded in the Çatalhöyük database. This method relies entirely on
observing morphological variation within the archaeological sample. The attributes examined for
these ten elements were determined through consultation with Louise Martin and Hijlke
Buitenhuis. These phalanges were rated by the qualitative attribute list which is discussed in the
Methodology chapter. Attributes are listed in the following Table 3. A more detailed description
of the attributes being examined is included in the Methodology chapter.
Each of the first phalanges was examined for each attribute and scored as having either the
morphology described as Quality 1 or Quality 2. Two of the first phalanges were dissimilar in
every attribute except for Attribute B in which they were both exhibiting Quality 2. Thus
Attribute B was not considered when comparing the remaining first phalanges. The remaining
eight phalanges appeared to separate into two groups, each based off the attributes of one of the
dissimilar first phalanges. These results are displayed in Appendix 6. Due to the qualitative
nature of this experiment and the possibility of error (as the author was the only one to conduct
this experiment), observer bias must be kept in mind. Based on the two dissimilar first phalanges,
each group seems to have the following attributes as displayed in the following Table 4.
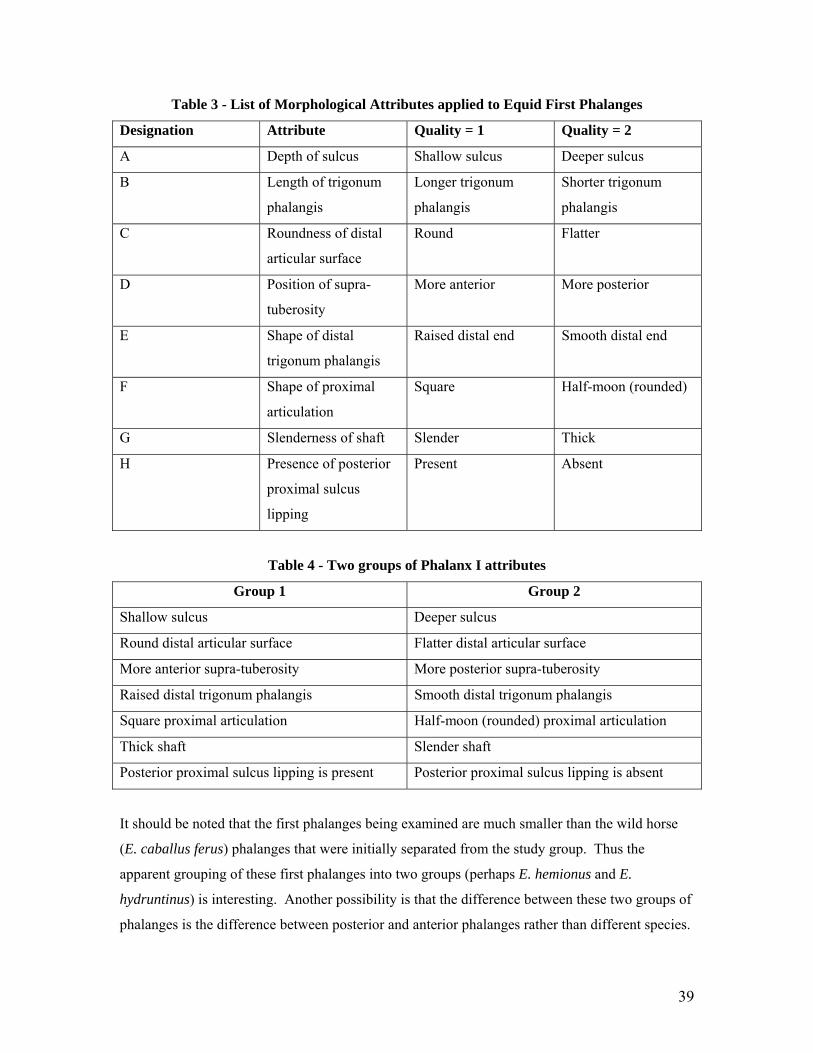
39
Table 3 - List of Morphological Attributes applied to Equid First Phalanges
Designation Attribute Quality = 1 Quality = 2
A Depth of sulcus Shallow sulcus Deeper sulcus
B Length of trigonum
phalangis
Longer trigonum
phalangis
Shorter trigonum
phalangis
C Roundness of distal
articular surface
Round Flatter
D Position of supra-
tuberosity
More anterior More posterior
E Shape of distal
trigonum phalangis
Raised distal end Smooth distal end
F Shape of proximal
articulation
Square Half-moon (rounded)
G Slenderness of shaft Slender Thick
H Presence of posterior
proximal sulcus
lipping
Present Absent
Table 4 - Two groups of Phalanx I attributes
Group 1 Group 2
Shallow sulcus Deeper sulcus
Round distal articular surface Flatter distal articular surface
More anterior supra-tuberosity More posterior supra-tuberosity
Raised distal trigonum phalangis Smooth distal trigonum phalangis
Square proximal articulation Half-moon (rounded) proximal articulation
Thick shaft Slender shaft
Posterior proximal sulcus lipping is present Posterior proximal sulcus lipping is absent
It should be noted that the first phalanges being examined are much smaller than the wild horse
(E. caballus ferus) phalanges that were initially separated from the study group. Thus the
apparent grouping of these first phalanges into two groups (perhaps E. hemionus and E.
hydruntinus) is interesting. Another possibility is that the difference between these two groups of
phalanges is the difference between posterior and anterior phalanges rather than different species.

40
Phalanx I
60
6570
75
80
85
90
20 22 24 26 28 30 32 34 36
Measurement 4 (SD)
Mea
sure
men
t 5 (G
L)
Phalanx I
27
32
37
35 40 45 50 55 60
Measurement 2 (Bp)
Mea
sure
men
t 7
(Dp)
Due to the lack of comparative material and small sample size, it is difficult to say either way at
the present time.
In the scatter plots below (Figure 24 and Figure 25), the two groups of first phalanges are plotted.
The light blue squares represent one group while the red circles represent the other. The dark
blue diamonds are first phalanges that were either thought to be E. caballus ferus phalanges or
could not be located for reanalysis. In both scatter plots it appears that there is some size
difference between the two morphological groups though it is not large. A larger sample size
could perhaps make this difference in size clearer. Again it must be reiterated that the difference
in morphology and size may be due to either the difference between the two taxa (E. hemionus
and E. hydruntinus) or it may be due to the difference between anterior and posterior phalanges.
Figure 24 - Scatter plot of Phalanx I Measurements (SD versus GL) and Morphological
Attributes
Figure 25 - Scatter plot of Phalanx I Measurements (Bp versus Dp) and Morphological
Attributes

41
Proportion of age determined equid elements
218, 64%
123, 36%
CRANIALPOSTCRANIAL
4.5 Body-part Representation
The equid body-parts represented at Çatalhöyük seems to include the whole body with many
elements from both the cranial (teeth and skull fragments) and postcranial (phalanges, tibia,
scapula, pelvis) being recovered from the site (Martin and Russell, in press, Russell and Martin
2005). This section is covered quite sufficiently in the references mentioned but is important to
note as it helps provide some evidence towards the economic models of equid acquisition, the
opportunistic hunting model and the logistical hunting model. The fact that the entire skeleton is
present at Çatalhöyük could perhaps be seen to favour the opportunistic hunting model as the
whole carcass has been brought back to site with little apparent field processing that seems to be
inherent with logistical hunting (Binford 1978).
4.6 Age of Equid Population
The majority of elements that have had their age determined at Çatalhöyük are cranial, consisting
of loose teeth and/or skull and mandible fragments (Figure 26). The equid remains retrieved from
Çatalhöyük appear to be adult for the majority with relatively few younger elements identified
(Figure 27). Age has been determined through tooth wear, bone fusion and bone
appearance/porosity. Taphonomy may of course have affected the preservation of unfused
postcranial elements thus creating an under representation of young animals in the Çatalhöyük
equid population. Of the total equid bones recorded up until May 2004, there have been 11 bones
exhibiting pathologies which is about 1 percent of the total equid bones. As displayed in Russell
and Martin (2005: 58), most of the pathologies seem to be related to age or injury. These results
would appear to support the idea that the equid remains recovered from Çatalhöyük represent
wild, hunted populations.
Figure 27 - Proportion of age determined equid elements at Çatalhöyük

42
Age of equid population
18%
0%
11%
8%50%
13%
YOUNGFETAL/NEONATEINFANTILE/JUVENILEJUVENILE/SUBADULTSUBADULT/ADULTOLD
Figure 27 - Age of equid population recovered from Çatalhöyük
4.7 Summary
Various methods of analysis have been used to investigate the equid population at Çatalhöyük in
an attempt to identify the equid species (E. caballus ferus, E. hemionus, and E. hydruntinus). In
an attempt to expand and verify the identification of the equid teeth, a metric analysis was
conducted. Unfortunately, the population size of the teeth at Çatalhöyük does not yet allow for a
more detailed analysis of these measurements. Postcranial elements, in particular, were studied
for the purpose of identifying the three equid species present at Çatalhöyük. Measurements of
several post cranial elements were gathered from the Çatalhöyük faunal database, carefully
examined and presented in scatter plots for analysis. The scatter plots present size differences
between the wild horse (E. caballus ferus) elements and wild ass (E. hydruntinus and E.
hemionus) elements. The measurements from Çatalhöyük were then compared as means to equid
remains from other sites in the region.
In an attempt to identify morphological differences between the first phalanges of E. hydruntinus
and E. hemionus, a list of morphological attributes was selected and applied to ten first phalanges.
Two groups of morphologically different first phalanges appear to be present in the population
studied. The grouping of the first phalanges may be due to the phalanges belonging to two
different taxa (E. hemionus and E. hydruntinus) or may indicate the presence of anterior and
posterior phalanges. Finally the body-part representation and age distribution of the equid
population at Çatalhöyük was presented in this chapter and it appears that most of the equid body
was brought back to Çatalhöyük and the population seems to be mostly adult.

43
5) General Discussion
5.1 Equids at Çatalhöyük
‘Animals and plants are important elements of the cultural landscape, since, by inhabiting the
landscape, they imbue the landscape with meaning; they do not simply exist as entities in the
landscape, they are constitute of the landscape.
(Jones 1998)
As this quote implies, it is most probable that equids would have been important to the
inhabitants of Çatalhöyük on some level and aspects of the equid biology and behaviour would
have been firmly entrenched in the Çatalhöyük cultural world view. Whether this was to orient
hunting strategies or to protect agricultural land, knowledge of equid behaviour would have been
imperative while living on the Neolithic central Anatolian landscape. How the knowledge of the
equid populations in central Anatolia would be passed on to each generation of inhabitants at
Çatalhöyük is an interesting question as it leads to ideas of social memory proposed by Hodder
and Cessford (2004) and modes of passing on knowledge. Art and architecture can be seen as
examples of these modes. While equid remains are not often installed in houses or even very
often used as tools, there are several cases in which the semblances of equids are depicted
through painting or carvings (Martin and Russell, in press). Perhaps changes in equid
representation in art will correlate with changes in equid representation on the landscape or
changes in the Neolithic environment.
The issue of equid species distribution throughout Anatolia and the Near East is constantly being
revised and re-examined. Difficulties with the identification of the different equid species (E.
caballus ferus, E. hemionus and E. hydruntinus) have hampered the understanding of equid
distribution and species representation. In some cases further identification of equid species
using Payne’s (1991) proposed morphological standards has increased the known distribution of
E. hydruntinus in Anatolia during the Neolithic (Martin and Russell, in press, Russell and Martin
2005). Application of Payne’s morphological standards to other sites will further this process.
Unfortunately there has been no reliable method of identification of equid species using just
postcranial elements. One of the purposes of this exercise was to propose a new method using
postcranial elements and the first phalanx in particular. While there is no clear demarcation
between species evident when examining the scatter plots of the postcranial elements, it is most

44
likely that future comparison of these measurements with equid remains from other sites in the
region will yield some significant results. As an experiment in this dissertation, the equid
remains from Çatalhöyük that were used in this study were given statistical means and then
compared to equid measurement means from other sites in the region. It would appear that there
are some differences between the sites and that Çatalhöyük has quite a distinctive equid
population. Further comparison to collections of postcranial remains in museum collections and
equid remains recovered from sites in Anatolia and the Near East could all be used to create
standards that delineate the difference between equid species such as E. hydruntinus and E.
hemionus.
Morphological study of equid postcranial elements may yield some interesting results.
Particularly when attempting to differentiate between E. hemionus and E. hydruntinus. In this
dissertation, the first phalanx was examined in an attempt to provide morphological criteria or
attributes that may be visible when examining wild ass size first phalanges. Though the sample
size used for the establishment of the morphological criteria is very small, it did provide some
interesting results that could possibly be correlated with some of the first phalanges
measurements. At this point, the significance of this correlation is weak but with a larger sample
size it could be proven to be quite useful in differentiating between the two taxa (E. hemionus and
E. hydruntinus). The morphological data also appear to slightly separate into two groups when
plotted along with the metrics on the scatter plots. Though there is some mixing of the two
groups, the sample size is quite small and this will affect accuracy. As mentioned in the Results
chapter, this apparent grouping may be due to the presence of anterior and posterior phalanges
rather than the difference between species.
Several models for equid utilization at Çatalhöyük that have been discussed include opportunistic
hunting, logistical hunting and an ecosystem model. Logistical hunting as proposed by Binford
(1978) involves quite a bit of planning and a known, desirable prey. This model most likely
works for the acquisition of wild cattle (Bos sp.) as they seem to have had a symbolic importance
at Çatalhöyük that may have necessitated the constant presence of wild cattle remains on site and
made the formation of specific wild cattle hunting parties an economically viable and
symbolically important practice. As equids do not seem to be symbolically or spiritually
important at Çatalhöyük (Martin and Russell, in press), it is more probable that equids were
opportunistically hunted, most likely when other tasks were being completed (Levine 1990).
Perhaps equids were hunted to protect crops (Uerpmann 1982: 46) or were hunted while after

45
bigger and more desirable prey (i.e. wild cattle). Another model that has been discussed is the
ecosystem model which may be used to explain why the equids at Çatalhöyük appear to change in
their proportion and composition of taxa over time. Basically the ecosystem model assumes that
changes in the environment affect equid populations which can be identified in the archaeological
record. Russell and Martin (2005: 59) suggest that the equids recovered from Çatalhöyük may
have occupied different ecological zones with E. hemionus on the hot, dry plain in the summer
and E. hydruntinus in the foothills and on the edge of the plain. E. caballus ferus may have
grazed throughout the region and as none of these equids are thought to have been territorial
(unknown for E. hydruntinus) (Klingel 1974), all three taxa could have occupied the same area
with relative ease. As E. hydruntinus seems to be slowly replaced by E. hemionus at Çatalhöyük,
external factors must have been affecting the equid populations (Russell and Martin 2005).
Whether these changes occur because of natural environmental fluctuations or were brought
about by human interventions such as pastoralism or agriculture is a question difficult to answer
via the faunal remains alone and will take a combined human palaeoecology effort of
zooarchaeology palynology, geoarchaeology and archaeobotany to formulate a likely ecosystem
model. An ecosystem model could be important for a variety of reasons including the possibility
of comparing archaeological equid populations to modern equid populations to search for patterns
in population fluctuations and also to use in assisting with wild equid management practices
(Coughenour 2002: 161).

46
Chapter 6) Conclusion
6.1 Concluding Remarks
Several avenues of investigation have been utilized to further identify the equid species at
Çatalhöyük. While the results are fairly insignificant at this point, they are an important step
necessary for the comparison of equid species in Anatolia and the Near East. A morphological
technique using the first phalanx has been presented to further identify between the equid species
E. hemionus and E. hydruntinus. Models of equid utilization and procurement have also been
discussed with the author favouring the opportunistic model due to the small sample size of equid
remains at Çatalhöyük compared to the domesticated taxa (Ovis sp. and Capra sp.) and other wild
taxa such as cattle (Bos sp.). The effects of environmental change on equids at Çatalhöyük have
been discussed and further environmental research and refinement of equid species identification
must take place before a definitive relationship can be postulated. It is difficult at this point to
separate the effects of human culture from environmental change as related to equid populations
in Anatolia and the Near East. Further metric and morphological comparison of equid remains
from sites in the region will greatly assist in determining the roles of the three equid species at
Çatalhöyük and at other Neolithic sites in the area.
6.2 Recommendations for further research
In order to further refine the differences in the postcranial elements of E. hemionus and E.
hydruntinus, further measurements of hemione and hydruntine postcranial elements in museum
collections could be taken and compared to those from Çatalhöyük. Also, it would be useful to
compare the morphological attributes proposed for the first phalanges of E. hemionus and E.
hydruntinus at Çatalhöyük to other populations to investigate possible congruencies. Reanalysis
of some collections of equid remains from archaeological sites in Anatolia and the Near East may
yield new results and further identification of equid species. It has been possible in some
circumstances to further identify or correctly identify equid remains previously examined when
using updated techniques (Forsten 2000).
Furthermore, it may be beneficial to the study of equid utilization and subsistence strategies at
Çatalhöyük to investigate the season of death of the equid remains via dental thin-sections.
Human subsistence strategies and life ways are often significantly affected by seasonal changes

47
and can be reflected through analysis of archaeological evidence (Monks 1981). Any sources of
evidence, such as dental thin sections, that can be brought to investigate the seasonality of a past
culture are quite useful. The dental thin section technique relies upon the seasonal deposition of
cementum on mammal teeth (Bourque and Morris 1978), and in particular, equid teeth (McKinley
and Burke 2000). Basically, the layers of cementum deposited throughout the year differ in
thickness and colour by the season in which they were deposited. By studying the season of
death through dental thin section of teeth from each equid species (E. caballus ferus, E. hemionus
and E. hydruntinus), it may be possible to identify when and where equids were being obtained
by the inhabitants of Çatalhöyük. Ethnographically, most cultures that eat equid flesh seem to eat
it when fresh (Levine 1998). This could possibly be used to indicate that the season of death of
an equid was also when the animal was consumed and suggests that the meat was not usually
stored. This information could lead to interesting insights into subsistence strategies and perhaps
if compared to equid remains from other sites in central Anatolia, lead to further developments in
understanding equids and the Neolithic environment.
Stable isotope work on human and faunal remains at Çatalhöyük has yielded some interesting
results regarding the diet of humans and the fauna they exploited (Richards et al. 2003). Stable
isotope analysis of equid teeth that have been identified to species (E. caballus ferus, E. hemionus
and E. hydruntinus) could perhaps be compared to postcranial elements that are thought to
represent the three species and any correlation could perhaps be identified. If there are significant
differences in the stable isotope signature of the equid species, this could be used to indicate that
the equids are utilizing different ecological zones. By examining equid remains from different
levels, a change in the stable isotope signature may also indicate changes in diet as caused by
human culture or environmental change over time.

48
References Asouti, Eleni (2003). Woodland vegetation and fuel exploitation at the prehistoric campsite of Pinarbasi, south-central Anatolia, Turkey: the evidence from the wood charcoal macro-remains. Journal of Archaeological Science 30: 1185-1201. Asouti, Eleni and Jon Hather (2001). Charcoal analysis and the reconstruction of ancient woodland vegetation in the Konya Basin, south-central Anatolia, Turkey: results from the Neolithic site of Çatalhöyük East. Vegetation History and Archaeobotany 10: 23-32. Asouti, Eleni and Andrew Fairbairn (2002). ‘Subsistence economy in Central Anatolia during the Neolithic: the archaeobotanical evidence’ in The Neolithic of Central Anatolia, internal developments and external relations during the 9th-6th millennia cal BC, Proceedings of the International CANeW Round Table, Istanbul 23-24 November 2001, L. Thiessen & F. Gérard (eds.). Istanbul: Ege Yayınları, Pp. 181-192. Baird, D. (2002). ‘Early Holocene settlement in Central Anatolia: problems and prospects as seen from the Konya Plain’ in The Neolithic of Central Anatolia, internal developments and external relations during the 9th-6th millennia cal BC, Proceedings of the International CANeW Round Table, Istanbul 23-24 November 2001, L. Thissen & F. Gérard (eds.). Istanbul: Ege Yayınları, Pp. 139-52. Binford, L.R. (1978). Nunamiut Ethnoarchaeology. Academic Press, New York. Bonifay, Marie-Francoise (1991). ‘Equus hydruntinus Regalia minor n. ssp. from the Caves of Lunel-Viel (Herault, France)’ in Equids in the Ancient World: Volume II. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 178-216. Bottema, S. and H. Woldring (1984). Late Quaternary Vegetation and Climate of Southwestern Turkey Part II. Palaeohistoria 26: 123-149. Bourque, B.J. and K. Morris (1978). Determining Season of Death of Mammal Teeth from Archaeological Sites : A New Sectioning Technique. Science 199: 530-531. Buitenhuis, H. (1985). Preliminary Report on the Faunal Remains of Hayaz Höyük from the 1979-1983 Seasons. Anatolica XII: 61-74. Buitenhuis, H. (1991). ‘Some Equid Remains from South Turkey, North Syria, and Jordan’ in Equids in the Ancient World: Volume II. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 34-74. Buitenhuis, H. (2003). Code List: For Coding to Computer Use By Programs of H. Buitenhuis. ARC-BV, Groningen. Buitenhuis, H. (2005). Personal Communication. Cessford, C. (2001). A new dating sequence for Çatalhöyük. Antiquity 75: 717-25. Cessford, C. (2005). ‘Estimating the Neolithic Population at Çatalhöyük’ in Inhabiting Çatalhöyük: reports from the 1995-1999 seasons, Volume 4. I. Hodder (ed.), McDonald Institute for Archaeological Research, Cambridge. Pp. 323-326. Clutton-Brock, Juliet (1992). Horse Power: A History of the Horse and the Donkey in Human Societies. Natural History Museum Publications, London. Conolly, J. (1999). Technical strategies and technical change at Neolithic Çatalhöyük, Turkey. Antiquity 73: 791-800.

49
Coughenour, Michael B. (2002). ‘Ecosystem Modelling in Support of the Conservation of Wild Equids - the Example of the Pryor Mountain Wild Horse Range’ in Equids: Zebras, Asses and Horses: Status Survey and Conservation Action Plan. P.D. Moehlman (ed.), IUCN-The World Conservation Union, Gland, Switzerland. Pp. 154-162. Davis, Simon J. (1980). Late Pleistocene Holocene equid remains from Israel. Zoological Journal of the Linnaean Society 70: 289-312. Deniz, E. (1975). ‘Neolithic, Chalcolithic and Early Bronze Age faunal remains from Pulur Huyuk (Keban Dam Region), Turkey’ in Archaeozoological Studies. A.T. Clason (ed.), North-Holland Publishing Company, Amsterdam. Dive, Jerome and Vera Eisenmann (1991). ‘Identification and Discrimination of First Phalanges from Pleistocene and Modern Equus, Wild and Domestic’ in Equids in the Ancient World: Volume II. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 278-333. Ducos, Pierre (1975). A New Find of an Equid Metatarsal Bone from Tell Mureibet in Syria and its Relevance to the Identification of Equids from the Early Holocene of the Levant. Journal of Archaeological Science 2: 71-73. Ducos, Pierre (1986). ‘The Equid of Tell Muraibit, Syria’ in Equids in the Ancient World. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 237-245. Eisenmann, Vera (1986). ‘Comparative Osteology of Modern and Fossil Horses, Half-asses, and Asses’ in Equids in the Ancient World. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 67-116. Eisenmann, Vera and Sophie Beckouche (1986). ‘Identification and Discrimination of Metapodials from Pleistocene and Modern Equus, Wild and Domestic’ in Equids in the Ancient World. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 117-163. Esin, Ufuk, Savas Harmankaya (1999). ‘Aşikli’ in Neolithic in Turkey: The Cradle of Civilization. M. Ozdogan and N. Basgelen (eds.), Arkeoloji Ve Sanat Yayinlari, Istanbul. Pp. 115-132. Forsten, Ann (2000). ‘A Note on the Equid from Anau, Turkestan, “Equus Caballus Pumpellii” Duerst’ in Archaeology of the Near East IVB: Proceedings of the fourth international symposium on the archaeozoology of southwestern Asia and adjacent areas. M. Mashkour, A.M. Choyke, H. Buitenhuis and F. Poplin (eds.), ARC - Publicatie 32, Groningen, The Netherlands. Pp. 153-155. Grigson, Caroline (1993). The Earliest Domestic Horses in the Levant? - New Finds from the Fourth Millennium of the Negev. Journal of Archaeological Science 20: 645-655. Grigson, Caroline (2000). ‘The Secondary Products Revolution? Changes in Animal Management from the Fourth to the Fifth Millennium, at Arjoune, Syria’ in Archaeology of the Near East IVB: Proceedings of the fourth international symposium on the archaeozoology of southwestern Asia and adjacent areas. M. Mashkour, A.M. Choyke, H. Buitenhuis and F. Poplin (eds.), ARC - Publicatie 32, Groningen, The Netherlands. Pp. 12-28. Grigson, Caroline (2003). ‘Animal husbandry in the Late Neolithic and Chalcolithic at Arjoune: the secondary products revolution revisited’ in Excavations at Arjoune, Syria. P.J. Parr (ed.), BAR International Series 1134. Pp. 187-240. Groves, Colin P. (1986). ‘The Taxonomy, Distribution, and Adaptations of Recent Equids’ in Equids in the Ancient World. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 11-66.

50
Groves, Colin P. (2002). ‘Chapter 8: Taxonomy of Living Equidae’ in Equids: Zebras, Asses and Horses: Status Survey and Conservation Action Plan. P.D. Moehlman (ed.), IUCN-The World Conservation Union, Gland, Switzerland. Pp. 94-107. Hillson, Simon (1996). Mammal Bones and Teeth: An Introductory Guide to Methods of Identification. Institute of Archaeology, University College London, London. Hodder, Ian (1999). ‘Renewed Work at Çatalhöyük’ in Neolithic in Turkey: The Cradle of Civilization. M. Ozdogan and N. Basgelen (eds.), Arkeoloji Ve Sanat Yayinlari, Istanbul. Pp. 157-164. Hodder, Ian and Craig Cessford (2004). Daily Practice and Social Memory at Catalhoyuk. American Antiquity 69(1): 17-40. Hodder, Ian and Roger Matthews (1998). ‘Çatalhöyük: the 1990s Seasons’ in Ancient Anatolia: Fifty Years Work by the British Institute of Archaeology of Ankara. R. Matthews (ed.), The British Institute of Archaeology at Ankara. Jones, A. (1998). Where eagles dare: landscape, animals and the Neolithic of Orkney. Journal of Material Culture 3: 301-24. Klein, Richard G. and Kathryn Cruz-Uribe (1984). The Analysis of Animal Bones from Archaeological Sites. The University of Chicago Press, Chicago. Klingel, Hans (1974). ‘A Comparison of the Social Behaviour of the Equidae’ in The Behaviour of Ungulates and its relation to management, Volume 1. V. Geist and F. Wather (eds.), International Union for Conservation of Nature and Natural Resources, Morges, Switzerland. Pp. 124-132. Kuzucuoglu, C. and N. Roberts (1998). Evolution de l’Environment en Anatolie de 20,000 a 6000 BP. Paleorient 23(2): 7-24. Kuzucuoglu, Catherine (2002). ‘The environmental frame in Central Anatolia from the 9th to the 6th millennia cal BC.’ in The Neolithic of Central Anatolia, internal developments and external relations during the 9th-6th millennia cal BC, Proceedings of the International CANeW Round Table, Istanbul 23-24 November 2001, L. Thissen & F. Gérard (eds.). Istanbul: Ege Yayınları, Pp. 33-58. Levine, Marsha A. (1990). Dereivka and the problem of horse domestication. Antiquity 64: 727-40. Levine, Marsha A. (1998). Eating horses: the evolutionary significance of hippophagy. Antiquity 72: 90-100. Lyman, R. (1994). Vertebrate Taphonomy. Cambridge University Press, Cambridge. Martin, Louise and Nerissa Russell (In Press). The Equid Remains from Neolithic Çatalhöyük, Central Anatolia: A Preliminary Report. Matthews, Wendy (1999). ‘Micromorphology Archive Report’ in Çatalhöyük 1999 Archive Report. http://101.111.34.58/catal/Archive_rep99/matthews99.html. Mayr, E. (1947). Systematics and the Origin of Species. Columbia University Press, New York. McKinley, V. and A. Burke (2000). A new control sample for season of death estimates for Equus caballus from dental thin-sections. Archaeozoologia XI: 33-42. Meadow, R.H. (1980). Animal bones: Problems for the archaeologist together with some possible solutions. Paléorient 6: 65-77.

51
Meadow, Richard H. (1986). ‘Some Equid Remains from Çayönü, Southeastern Turkey’ in Equids in the Ancient World.. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 266-287. Mellart, James (1966). Excavations at Çatal Hüyük, 1965: Fourth preliminary report. Anatolian Studies 16:165-191. Mellart, James (1967). Catal Huyuk: A Neolithic Town in Anatolia. Thames and Hudson, London. Mellart, James (1975). The Neolithic of the Near East. Charles Scribner’s Sons, New York. Mellart, James (1998). ‘Catal Huyuk: the 1960s Seasons’ in Ancient Anatolia: Fifty Years Work by the British Institute of Archaeology of Ankara. R. Matthews (ed.), The British Institute of Archaeology at Ankara. Pp. 35-41. Moehlman, Patricia D. (2005). Endangered Wild Equids. Scientific American 292 (3): 86-93. Monks, Gregory G. (1981). Seasonality Studies. Advances in Archaeological Method and Theory 4: 177-240. Ozdogan, Mehmet (1997). The Beginning of Neolithic Economies in Southeastern Europe: An Anatolian Perspective. Journal of European Archaeology 5(2): 1-33. Ozdogan, M. (1998). Anatolia from the Last Glacial Maximum to the Holocene Climatic Optimum: Cultural Formations and the Impact of the Environmental Setting. Paleorient 23(2): 25-38. Ozdogan, Mehmet, (1999). ‘Cayonu’ in Neolithic in Turkey: The Cradle of Civilization. M. Ozdogan and N. Basgelen (eds.), Arkeoloji Ve Sanat Yayinlari, Istanbul. Pp. 35-64. Payne, Sebastian (1985). Animal Bones from Aşikli Hüyük. Anatolian Studies 35: 109-122. Payne, Sebastian (1988). ‘Chapter 5: Animal Bones from Tell Rubeidheh’ in Tell Rubeidheh: An Uruk Village in the Jebel Hamrin. R.G. Killick (ed.), Iraq Archaeological Reports - 2, Ais and Phillips Ltd., Warminster. Payne, Sebastian (1991). ‘Early Holocene Equids from Tall-i-Mushki (Iran) and Can Hasan III (Turkey)’ in Equids in the Ancient World: Volume II. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 132-177. Perkins, Dexter Jr. (1969). Fauna of Catal Huyuk: Evidence for Early Cattle Domestication in Anatolia. Science 164: 177-179. Richards, M.P., J.A. Pearson, T.I. Molleson, N. Russell, L. Martin (2003). Stable Isotope Evidence of Diet at Neolithic Çatalhöyük, Turkey. Journal of Archaeological Science 30: 67-76. Roberts, N., S. Black, P. Boyer, W.J. Eastwood, H.I. Griffiths, H.F. Lamb, M.J. Leng, R. Parish, J.M. Reed, D. Twigg, H. Yigitbasioglu (1999). Chronology and stratigraphy of Late Quaternary sediments in the Konya Basin, Turkey: Results from the KOPAL Project. Quaternary Science Reviews 18: 611-630. Roberts, N., J.M. Reed, M.J. Leng, C. Kuzucuoglu, M. Fontugue, J. Bertaux, H. Woldring, S. Bottema, S. Black, E. Hunt and M. Karabiyikoglu (2001). The tempo of Holocene climatic change in the eastern Mediterranean region: new high-resolution crater-lake sediment data from central Turkey. The Holocene 11(6): 721-736.

52
Rosen, Arlene and Neil Roberts (2005). ‘The nature of Çatalhöyük: people and their changing environments on the Konya Plain’ in Çatalhöyük Perspectives: themes from the 1995-1999 seasons, Volume 6. I. Hodder (ed.), McDonald Institute for Archaeological Research, Cambridge. Russell, N. and L. Martin (2000). ‘Neolithic Çatalhöyük: Preliminary zooarchaeological results from the Renewed excavations’ in Archaeozoology of the Near East IV: Proceedings of the Fourth International Symposium on the Archaeozoology of Southwestern Asia and Adjacent Areas, M. Mashkour and H. Buitenhuis (eds.), Groningen Institute for Archaeology, Groningen. Pp. 164-170. Russell, Nerissa and Louise Martin (2005). ‘Çatalhöyük Mammal Remains’ in Inhabiting Çatalhöyük: reports from the 1995-1999 seasons, Volume 4. I. Hodder (ed.), McDonald Institute for Archaeological Research, Cambridge. Pp. 33-98. Russell, N., L. Martin, & L. LeBlanc (1996). Animal bone report, Çatalhöyük 1996 Archive Report, http://catal.arch.cam.ac.uk/catal/Archive_rep96/russmartin96.html Russell, N., L. Martin, & S. Frame (2000). Faunal remains, Çatalhöyük 2000 Archive Report, http://catal.arch.cam.ac.uk/catal/Archive_rep00/martin00.html Saltz, David (2002). ‘The Dynamics of Equid Populations’ in Equids: Zebras, Asses and Horses: Status Survey and Conservation Action Plan. P.D. Moehlman (ed.), IUCN-The World Conservation Union, Gland, Switzerland. Pp. 118-123. Sisson, Septimus (1953). The Anatomy of the Domestic Animals. W.B. Saunders Company, London. Thiessen, Laurens (2001). Trajectories towards the neolithisation of NW Turkey. Documenta Praehistorica XXVI: 29-39. Turnbull, Priscilla F. (1986). ‘Measurements of Equus hemionus from Palegawra Cave (Zarzian, Iraq)’ in Equids in the Ancient World. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 319-365. Uerpmann, Hans-Peter (1982). Faunal Remains from Shams Ed-Din Tannira. Berytus 30:3-48. Uerpmann, Hans-Peter (1986). ‘Halafian Equid Remains from Shams ed-Din Tannira in Northern Syria’ in Equids in the Ancient World. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 246-265. Uerpmann, Hans-Peter (1987). The Ancient Distribution of Ungulate Mammals in the Middle East. Dr. Ludwig Reichert Verlag, Wiebaden. Pp. 13-39. Uerpmann, Hans-Peter (1991). ‘Equus africanus in Arabia’ in Equids in the Ancient World: Volume II. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 12-33. Von Den Driesch, Angela (1976). A Guide to the Measurement of Animal Bones from Archaeological Sites. Peabody Museum Bulletin 1, Harvard University. Wick, Lucia, Gerry Lemcke, and Michael Sturm (2003). Evidence of Lateglacial and Holocene climatic change and human impact in Eastern Anatolia: high-resolution pollen, charcoal, isotopic and geochemical records from the laminated sediments of Lake Van, Turkey. The Holocene 13(5): 665-675. Woldring, Henk (2002). ‘Climate change and the onset of sedentism in Cappadocia’ in The Neolithic of Central Anatolia, internal developments and external relations during the 9th-6th millennia cal BC, Proceedings of the International CANeW Round Table, Istanbul 23-24 November 2001, L. Thissen & F. Gérard (eds.). Istanbul: Ege Yayınları, Pp. 59-66.

53
Woldring, H. and S. Bottema (2001). The Vegetation History of East-Central Anatolia in Relation to Archaeology: The Eski Acigol Pollen Evidence Compared with the Near Eastern Environment. Palaeohistoria 43: 1-31. Zeder, Melina A. (1986). ‘The Equid Remains from Tal-e Malyan, Southern Iran’ in Equids in the Ancient World.. R. H. Meadow and H. Uerpmann (eds.), Dr. Ludwig Reichert Verlag, Wiesbaden. Pp. 366-412.

54
Appendix 1 - Metric analysis of maxillary equid teeth from Çatalhöyük
Unit F-nr Species Psp element side M1 M2 M3 M4 M5 wear MAXILLARY 4121 1605 ferus ferus P3/4 R 25.1 22.4 10.9 2 59.2 medium 4121 1603 ferus ferus P3/4 L 27.3 23.8 10.9 2 49.4 medium 4121 1612 hydruntine hydruntine M1/2 R 23.7 22.1 6.3 1 17.8 heavy 4121 1600 ferus ferus M3 L 24.4 20.3 11.0 1 35.3 medium++ 4121 1607 equus hydruntine M1/2 R 22.9 23.9 8.7 2 heavy 4874 217 ferus ferus P3/4 L 25.9 25.1 12.7 4 heavy 4824 141 hydruntine hydruntine P3/4 L 25.4 22.7 7.2 3 48.7 heavy 4121 882 equus hemione M1/2 L 25.9 22.0 9.9 1 62.3 medium 5290 2731 equus hemione M1/2 R 24.2 22.4 11.1 2 54.8 medium 4846 231 ferus ferus P3/4 R 25.7 24.5 11.5 4 63.9 medium++ 4836 239 equus hemione M1/2 L 20.6 20.2 9.2 1 34.9 medium++ 4190 52 equus he/fe P3/4 L 24.7 22.4 9.0 2 45.9 heavy 4121 1616 ferus ferus M3 R 21.0 18.8 9.8 1 43.2 M+++ 4121 1604 ferus ferus M3 L 21.6 11.4 1 50.0 M+++ 4121 1610 equus ferus M3 R 23.5 19.6 10.5 1 H++ 4121 1599 ferus ferus P3/4 R 24.5 10.2 1 45.2 M+++ 4121 1608 hydruntine hydruntine PD L 32.0 16.0 5.8 1 M++ 4121 1080 ferus ferus PD R 30.8 18.4 4 21.0 H 4121 1606 hy/he hy/he M1 R 20.4 21.1 8.3 2 31.0 m+++ 4121 1606 hy/he hy/he M2 R 21.5 22.1 8.8 1 36.0 m+++ 4121 1604 hydruntine hydruntine pd2 L 29.2 13.9 12.7 1 10.8 m+++ 4121 1604 hydruntine hydruntine pd3 L 27.1 14.3 11.7 2 12.3 m+++ 1092 797 hydruntine hydruntine P2 L 31.0 20.6 5.4 1 m++ 1092 800 hemione hemione M3 R 23.4 18.1 10.2 1 20.0 H++ 1092 801 equus equus P3/4` L 24.5 3 43.7 m++ 1092 803 hy/he hy/he P3/4 R 25.5 22.0 9.2 4 m++ 1092 799 ferus ferus pd L 28.2 15.9 8.8 1 M++ 1612 244 equus equus P2 L 30.9 20.4 2 M+++ 4518 314 ferus ferus M1/2 R 26.6 20.1 8.0 4 M+++ 1523 47 he/hy he/hy P3/4 R 26.3 24.4 8.2 4 51.7 H++ 1297 38? ferus ferus P3/4 L 4 H++ 6948 71 ferus ferus P3/4 R 28.2 18.3 8.7 3 H+++ 1511 301 hemione hemione M1/2 R 21.2 21.8 10.6 2 H++ 1668 875 ferus ferus M3 L 22.1 19.8 9.9 1 40.0 M+ 1506 449 hy/he hy/he P3/4 R 26.2 8.9 3 59.0 M+++ 1506 448 equus equus P2 L 31.2 7.1 1 M+++ 1506 447 equus equus pd2 R 35.1 17.2 6.9 2 M+ 1506 447 equus equus pd3 R 27.8 19.2 7.7 3 M+ 1506 447 equus equus pd4 R 29.8 17.0
Legend (Adapted from Buitenhuis 2003: 25).
Maxilar Molar Measurements
1 OL - L - occlusal length 2 B - Be - occlusal width 3 LP length protocone 4 Caballine fold (stage) 1 = none, 2 = trace, 3 = light, 4 = clear 5 H - crown height
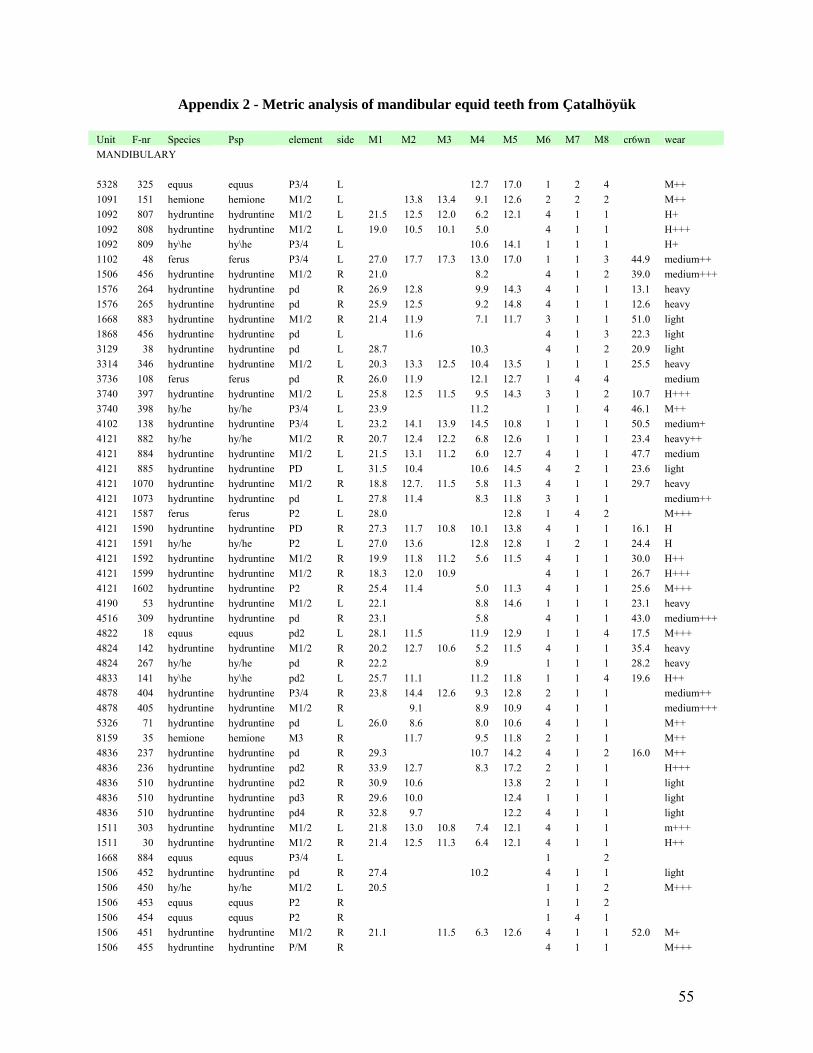
55
Appendix 2 - Metric analysis of mandibular equid teeth from Çatalhöyük
Unit F-nr Species Psp element side M1 M2 M3 M4 M5 M6 M7 M8 cr6wn wear MANDIBULARY 5328 325 equus equus P3/4 L 12.7 17.0 1 2 4 M++ 1091 151 hemione hemione M1/2 L 13.8 13.4 9.1 12.6 2 2 2 M++ 1092 807 hydruntine hydruntine M1/2 L 21.5 12.5 12.0 6.2 12.1 4 1 1 H+ 1092 808 hydruntine hydruntine M1/2 L 19.0 10.5 10.1 5.0 4 1 1 H+++ 1092 809 hy\he hy\he P3/4 L 10.6 14.1 1 1 1 H+ 1102 48 ferus ferus P3/4 L 27.0 17.7 17.3 13.0 17.0 1 1 3 44.9 medium++ 1506 456 hydruntine hydruntine M1/2 R 21.0 8.2 4 1 2 39.0 medium+++ 1576 264 hydruntine hydruntine pd R 26.9 12.8 9.9 14.3 4 1 1 13.1 heavy 1576 265 hydruntine hydruntine pd R 25.9 12.5 9.2 14.8 4 1 1 12.6 heavy 1668 883 hydruntine hydruntine M1/2 R 21.4 11.9 7.1 11.7 3 1 1 51.0 light 1868 456 hydruntine hydruntine pd L 11.6 4 1 3 22.3 light 3129 38 hydruntine hydruntine pd L 28.7 10.3 4 1 2 20.9 light 3314 346 hydruntine hydruntine M1/2 L 20.3 13.3 12.5 10.4 13.5 1 1 1 25.5 heavy 3736 108 ferus ferus pd R 26.0 11.9 12.1 12.7 1 4 4 medium 3740 397 hydruntine hydruntine M1/2 L 25.8 12.5 11.5 9.5 14.3 3 1 2 10.7 H+++ 3740 398 hy/he hy/he P3/4 L 23.9 11.2 1 1 4 46.1 M++ 4102 138 hydruntine hydruntine P3/4 L 23.2 14.1 13.9 14.5 10.8 1 1 1 50.5 medium+ 4121 882 hy/he hy/he M1/2 R 20.7 12.4 12.2 6.8 12.6 1 1 1 23.4 heavy++ 4121 884 hydruntine hydruntine M1/2 L 21.5 13.1 11.2 6.0 12.7 4 1 1 47.7 medium 4121 885 hydruntine hydruntine PD L 31.5 10.4 10.6 14.5 4 2 1 23.6 light 4121 1070 hydruntine hydruntine M1/2 R 18.8 12.7. 11.5 5.8 11.3 4 1 1 29.7 heavy 4121 1073 hydruntine hydruntine pd L 27.8 11.4 8.3 11.8 3 1 1 medium++ 4121 1587 ferus ferus P2 L 28.0 12.8 1 4 2 M+++ 4121 1590 hydruntine hydruntine PD R 27.3 11.7 10.8 10.1 13.8 4 1 1 16.1 H 4121 1591 hy/he hy/he P2 L 27.0 13.6 12.8 12.8 1 2 1 24.4 H 4121 1592 hydruntine hydruntine M1/2 R 19.9 11.8 11.2 5.6 11.5 4 1 1 30.0 H++ 4121 1599 hydruntine hydruntine M1/2 R 18.3 12.0 10.9 4 1 1 26.7 H+++ 4121 1602 hydruntine hydruntine P2 R 25.4 11.4 5.0 11.3 4 1 1 25.6 M+++ 4190 53 hydruntine hydruntine M1/2 L 22.1 8.8 14.6 1 1 1 23.1 heavy 4516 309 hydruntine hydruntine pd R 23.1 5.8 4 1 1 43.0 medium+++ 4822 18 equus equus pd2 L 28.1 11.5 11.9 12.9 1 1 4 17.5 M+++ 4824 142 hydruntine hydruntine M1/2 R 20.2 12.7 10.6 5.2 11.5 4 1 1 35.4 heavy 4824 267 hy/he hy/he pd R 22.2 8.9 1 1 1 28.2 heavy 4833 141 hy\he hy\he pd2 L 25.7 11.1 11.2 11.8 1 1 4 19.6 H++ 4878 404 hydruntine hydruntine P3/4 R 23.8 14.4 12.6 9.3 12.8 2 1 1 medium++ 4878 405 hydruntine hydruntine M1/2 R 9.1 8.9 10.9 4 1 1 medium+++ 5326 71 hydruntine hydruntine pd L 26.0 8.6 8.0 10.6 4 1 1 M++ 8159 35 hemione hemione M3 R 11.7 9.5 11.8 2 1 1 M++ 4836 237 hydruntine hydruntine pd R 29.3 10.7 14.2 4 1 2 16.0 M++ 4836 236 hydruntine hydruntine pd2 R 33.9 12.7 8.3 17.2 2 1 1 H+++ 4836 510 hydruntine hydruntine pd2 R 30.9 10.6 13.8 2 1 1 light 4836 510 hydruntine hydruntine pd3 R 29.6 10.0 12.4 1 1 1 light 4836 510 hydruntine hydruntine pd4 R 32.8 9.7 12.2 4 1 1 light 1511 303 hydruntine hydruntine M1/2 L 21.8 13.0 10.8 7.4 12.1 4 1 1 m+++ 1511 30 hydruntine hydruntine M1/2 R 21.4 12.5 11.3 6.4 12.1 4 1 1 H++ 1668 884 equus equus P3/4 L 1 2 1506 452 hydruntine hydruntine pd R 27.4 10.2 4 1 1 light 1506 450 hy/he hy/he M1/2 L 20.5 1 1 2 M+++ 1506 453 equus equus P2 R 1 1 2 1506 454 equus equus P2 R 1 4 1 1506 451 hydruntine hydruntine M1/2 R 21.1 11.5 6.3 12.6 4 1 1 52.0 M+ 1506 455 hydruntine hydruntine P/M R 4 1 1 M+++

56
Legend (Adapted from Buitenhuis 2003: 25)
Mandibular Molar Measurements 1 OL - L - occlusal length 2 B - B3 - occlusal width 3 B3 - hypoconid - metastylid width 4 Lnd - LDk length metaconid - metastylid 5 LF - LPF - length postflexid 6 Bei(stage) 1=below flexid, 2=below top flexid, 3=above top flexid, 4=near lingual sulcus 7 Pli hypoconid (stage) 1=none, 2=trace, 3=light, 4=clear 8 Pli ptychostylid (stage) 1=none, 2=trace, 3=light, 4=clear

57
Appendix 3 - Measurements proposed by Payne (1991:135) for upper (above) and lower equid teeth.
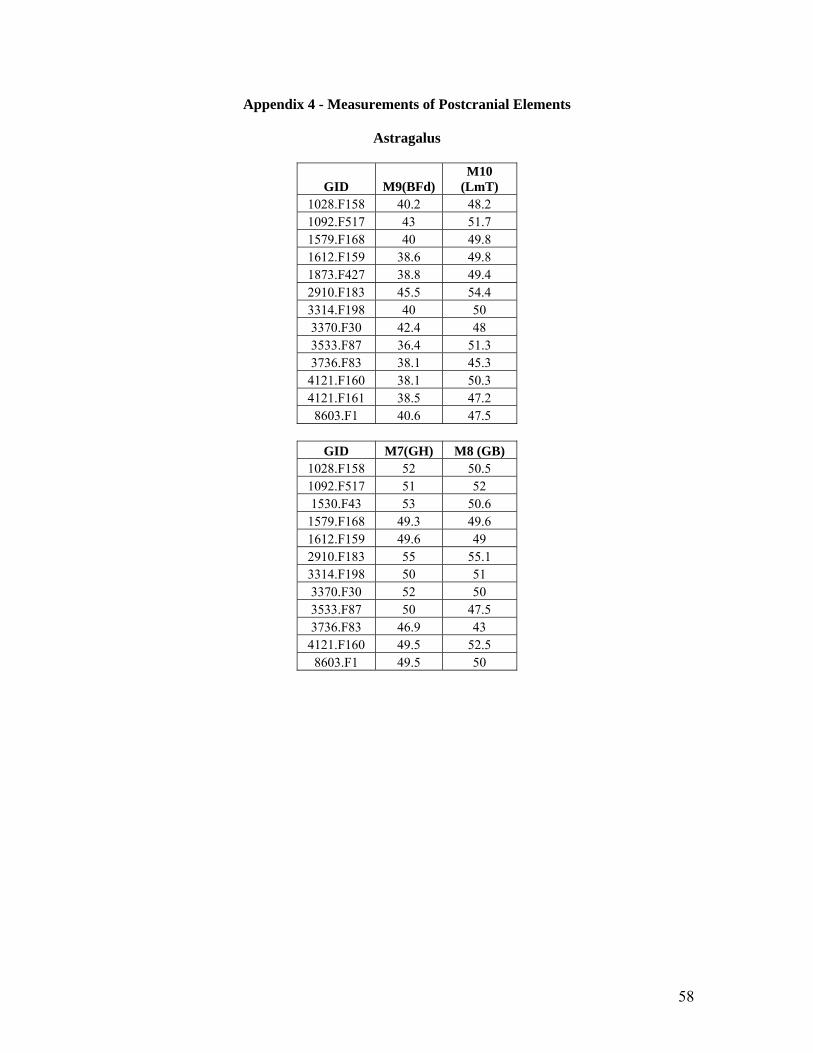
58
Appendix 4 - Measurements of Postcranial Elements
Astragalus
GID M9(BFd) M10
(LmT) 1028.F158 40.2 48.2 1092.F517 43 51.7 1579.F168 40 49.8 1612.F159 38.6 49.8 1873.F427 38.8 49.4 2910.F183 45.5 54.4 3314.F198 40 50 3370.F30 42.4 48 3533.F87 36.4 51.3 3736.F83 38.1 45.3
4121.F160 38.1 50.3 4121.F161 38.5 47.2 8603.F1 40.6 47.5
GID M7(GH) M8 (GB)
1028.F158 52 50.5 1092.F517 51 52 1530.F43 53 50.6
1579.F168 49.3 49.6 1612.F159 49.6 49 2910.F183 55 55.1 3314.F198 50 51 3370.F30 52 50 3533.F87 50 47.5 3736.F83 46.9 43
4121.F160 49.5 52.5 8603.F1 49.5 50
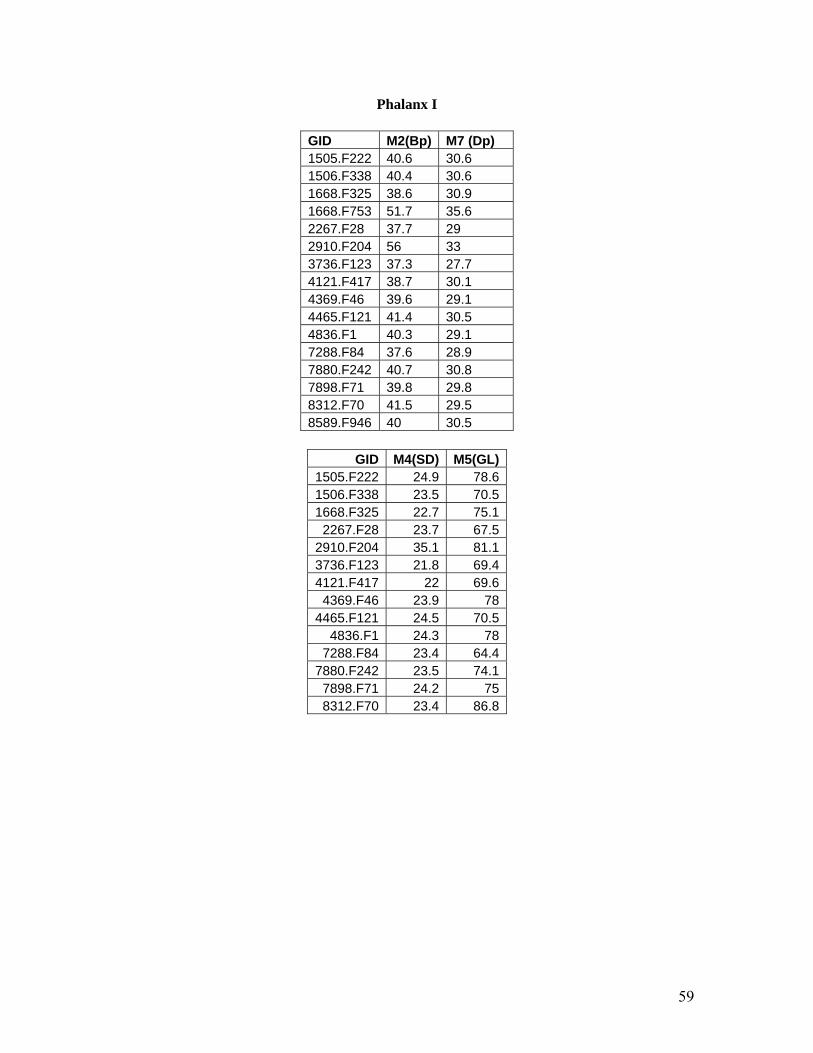
59
Phalanx I
GID M2(Bp) M7 (Dp) 1505.F222 40.6 30.6 1506.F338 40.4 30.6 1668.F325 38.6 30.9 1668.F753 51.7 35.6 2267.F28 37.7 29 2910.F204 56 33 3736.F123 37.3 27.7 4121.F417 38.7 30.1 4369.F46 39.6 29.1 4465.F121 41.4 30.5 4836.F1 40.3 29.1 7288.F84 37.6 28.9 7880.F242 40.7 30.8 7898.F71 39.8 29.8 8312.F70 41.5 29.5 8589.F946 40 30.5
GID M4(SD) M5(GL)
1505.F222 24.9 78.61506.F338 23.5 70.51668.F325 22.7 75.1
2267.F28 23.7 67.52910.F204 35.1 81.13736.F123 21.8 69.44121.F417 22 69.6
4369.F46 23.9 784465.F121 24.5 70.5
4836.F1 24.3 787288.F84 23.4 64.4
7880.F242 23.5 74.17898.F71 24.2 758312.F70 23.4 86.8
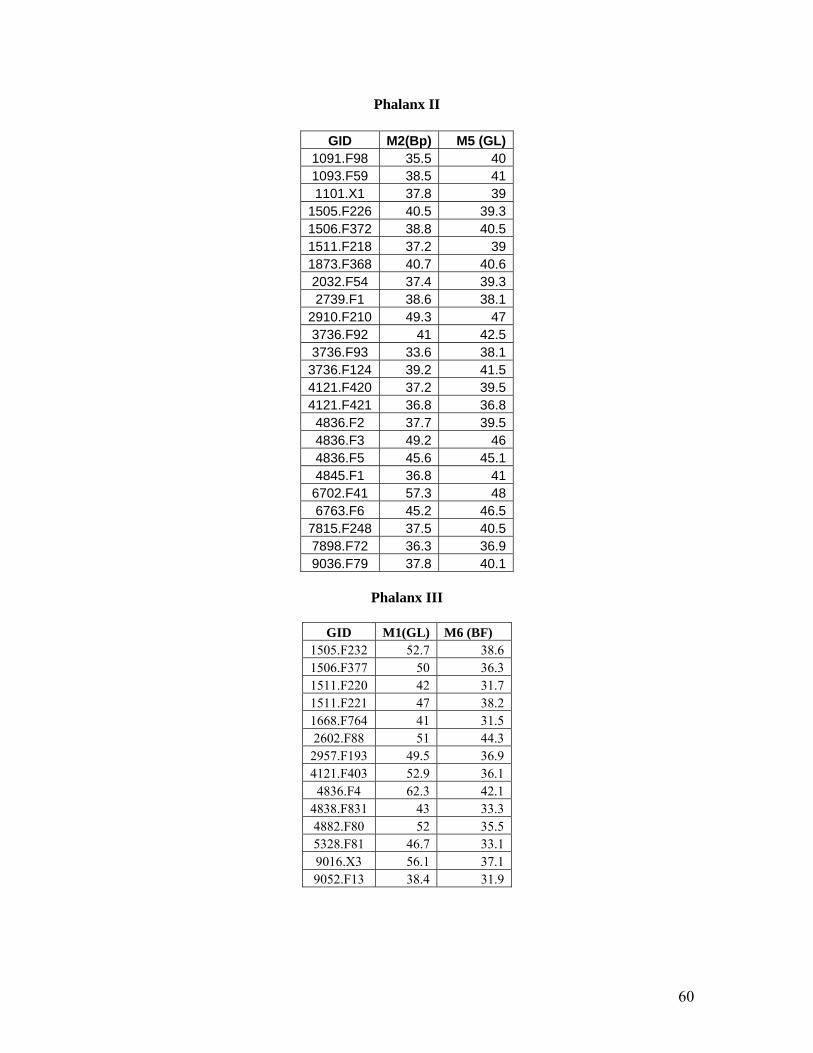
60
Phalanx II
GID M2(Bp) M5 (GL)1091.F98 35.5 401093.F59 38.5 411101.X1 37.8 39
1505.F226 40.5 39.31506.F372 38.8 40.51511.F218 37.2 391873.F368 40.7 40.62032.F54 37.4 39.32739.F1 38.6 38.1
2910.F210 49.3 473736.F92 41 42.53736.F93 33.6 38.1
3736.F124 39.2 41.54121.F420 37.2 39.54121.F421 36.8 36.8
4836.F2 37.7 39.54836.F3 49.2 464836.F5 45.6 45.14845.F1 36.8 416702.F41 57.3 486763.F6 45.2 46.5
7815.F248 37.5 40.57898.F72 36.3 36.99036.F79 37.8 40.1
Phalanx III
GID M1(GL) M6 (BF)
1505.F232 52.7 38.6 1506.F377 50 36.3 1511.F220 42 31.7 1511.F221 47 38.2 1668.F764 41 31.5 2602.F88 51 44.3
2957.F193 49.5 36.9 4121.F403 52.9 36.1 4836.F4 62.3 42.1
4838.F831 43 33.3 4882.F80 52 35.5 5328.F81 46.7 33.1 9016.X3 56.1 37.1 9052.F13 38.4 31.9
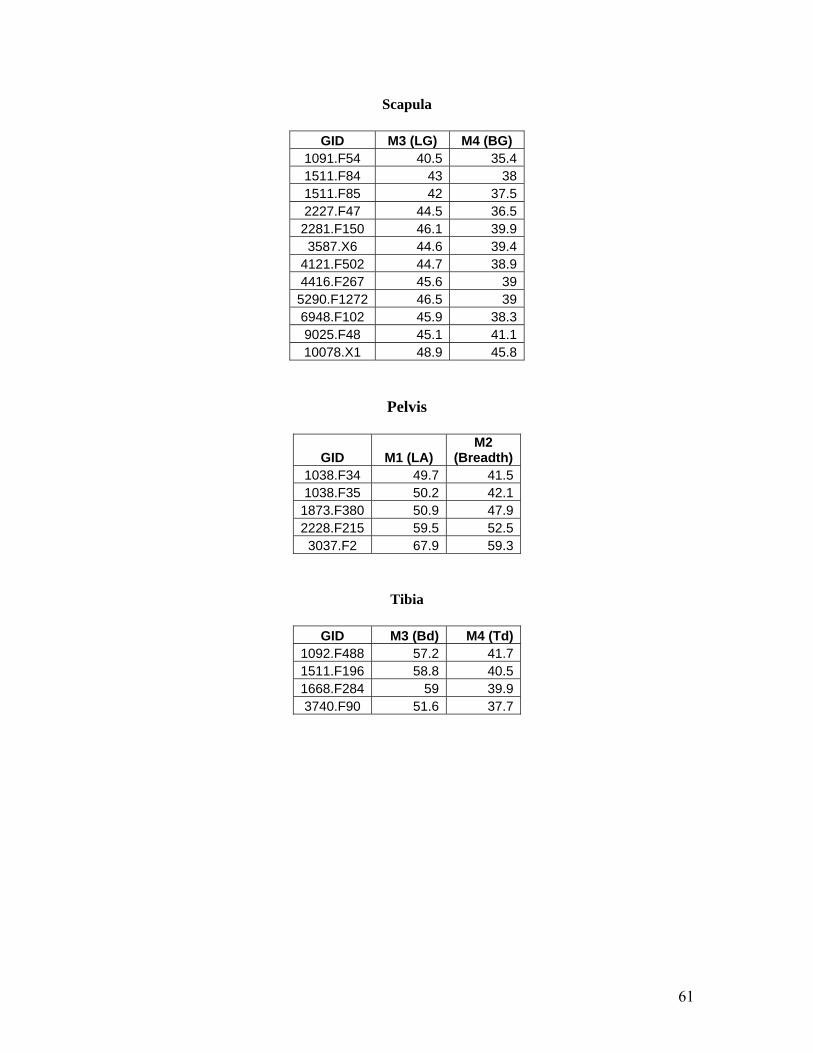
61
Scapula
GID M3 (LG) M4 (BG) 1091.F54 40.5 35.41511.F84 43 381511.F85 42 37.52227.F47 44.5 36.52281.F150 46.1 39.93587.X6 44.6 39.4
4121.F502 44.7 38.94416.F267 45.6 395290.F1272 46.5 396948.F102 45.9 38.39025.F48 45.1 41.110078.X1 48.9 45.8
Pelvis
GID M1 (LA) M2
(Breadth) 1038.F34 49.7 41.51038.F35 50.2 42.1
1873.F380 50.9 47.92228.F215 59.5 52.5
3037.F2 67.9 59.3
Tibia
GID M3 (Bd) M4 (Td)1092.F488 57.2 41.71511.F196 58.8 40.51668.F284 59 39.93740.F90 51.6 37.7
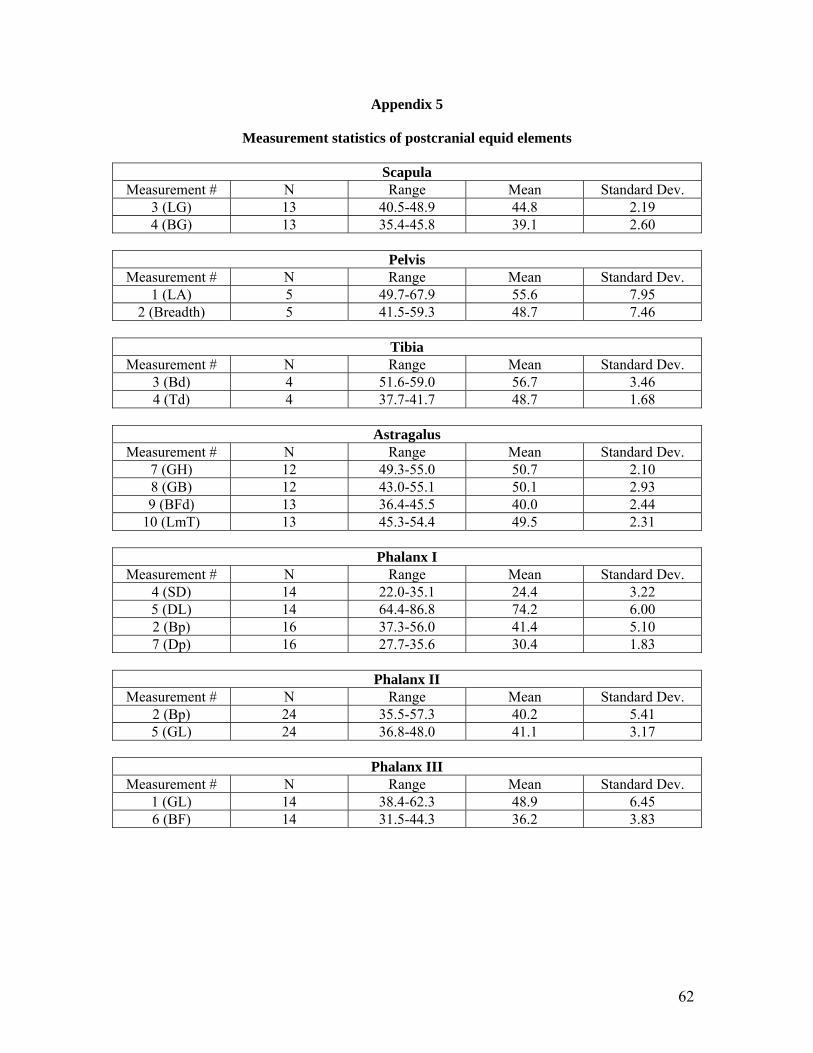
62
Appendix 5
Measurement statistics of postcranial equid elements
Scapula Measurement # N Range Mean Standard Dev.
3 (LG) 13 40.5-48.9 44.8 2.19 4 (BG) 13 35.4-45.8 39.1 2.60
Pelvis
Measurement # N Range Mean Standard Dev. 1 (LA) 5 49.7-67.9 55.6 7.95
2 (Breadth) 5 41.5-59.3 48.7 7.46
Tibia Measurement # N Range Mean Standard Dev.
3 (Bd) 4 51.6-59.0 56.7 3.46 4 (Td) 4 37.7-41.7 48.7 1.68
Astragalus
Measurement # N Range Mean Standard Dev. 7 (GH) 12 49.3-55.0 50.7 2.10 8 (GB) 12 43.0-55.1 50.1 2.93 9 (BFd) 13 36.4-45.5 40.0 2.44
10 (LmT) 13 45.3-54.4 49.5 2.31
Phalanx I Measurement # N Range Mean Standard Dev.
4 (SD) 14 22.0-35.1 24.4 3.22 5 (DL) 14 64.4-86.8 74.2 6.00 2 (Bp) 16 37.3-56.0 41.4 5.10 7 (Dp) 16 27.7-35.6 30.4 1.83
Phalanx II
Measurement # N Range Mean Standard Dev. 2 (Bp) 24 35.5-57.3 40.2 5.41 5 (GL) 24 36.8-48.0 41.1 3.17
Phalanx III
Measurement # N Range Mean Standard Dev. 1 (GL) 14 38.4-62.3 48.9 6.45 6 (BF) 14 31.5-44.3 36.2 3.83
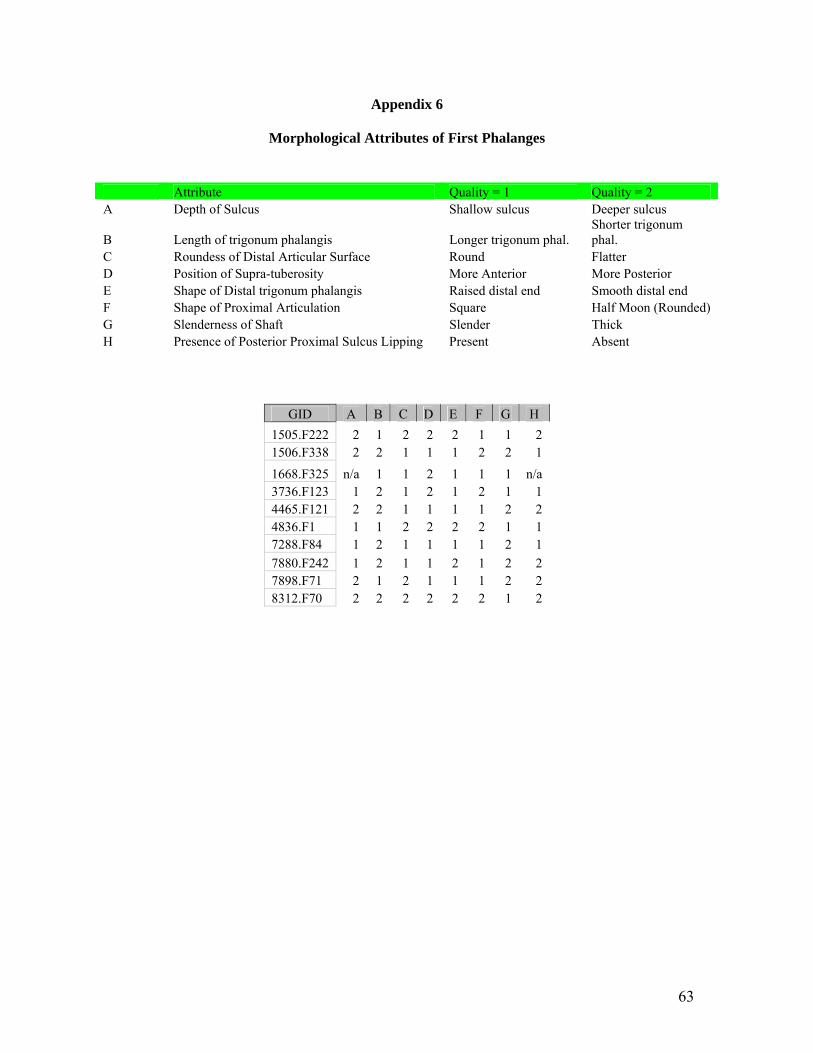
63
Appendix 6
Morphological Attributes of First Phalanges
Attribute Quality = 1 Quality = 2 A Depth of Sulcus Shallow sulcus Deeper sulcus
B Length of trigonum phalangis Longer trigonum phal. Shorter trigonum phal.
C Roundess of Distal Articular Surface Round Flatter D Position of Supra-tuberosity More Anterior More Posterior E Shape of Distal trigonum phalangis Raised distal end Smooth distal end F Shape of Proximal Articulation Square Half Moon (Rounded) G Slenderness of Shaft Slender Thick H Presence of Posterior Proximal Sulcus Lipping Present Absent
GID A B C D E F G H 1505.F222 2 1 2 2 2 1 1 2 1506.F338 2 2 1 1 1 2 2 1 1668.F325 n/a 1 1 2 1 1 1 n/a 3736.F123 1 2 1 2 1 2 1 1 4465.F121 2 2 1 1 1 1 2 2 4836.F1 1 1 2 2 2 2 1 1 7288.F84 1 2 1 1 1 1 2 1 7880.F242 1 2 1 1 2 1 2 2 7898.F71 2 1 2 1 1 1 2 2 8312.F70 2 2 2 2 2 2 1 2