enzyme polymorphism of an antioxidant system in chronically irradiated scots pine populations
TRANSCRIPT

ISSN 2079�0597, Russian Journal of Genetics: Applied Research, 2014, Vol. 4, No. 5, pp. 421–433. © Pleiades Publishing, Ltd., 2014.Original Russian Text © P.Yu. Volkova, S.A. Geraskin, 2013, published in Ekologicheskaya Genetika, 2013, Vol. 11, No. 3, pp. 48–62.
421
Forests play an important role in maintaining asteady state of the biosphere and are widely used foreconomic and recreational purposes. Violations of thegenetic structure of the tree species that are dominantin forest ecosystems and are characterized by a longlife cycle lead to a reduction in plasticity and adapt�ability in subsequent generations due to loss of allelicvariants and their combinations. In turn, the reducedresistance of the dominant species causes changes inthe cenosis structure, which may entail the loss ofsome dependent species that are part of the ecosystem(Whitham et al., 2006). Therefore, successful adapta�tion of forest ecosystems to changing environmentalconditions and to the increasing anthropogenic pres�sure requires preserving the genetic diversity of speciesthat constitute the ecosystem.
As a result of radiation accidents at PO Mayak(1957) and the Chernobyl Nuclear Power Plant(1986), forest ecosystems suffered the most becausecrowns of trees retain a significant portion of atmo�spheric depositions and self�cleaning takes a long time(Tikhomirov, 1972). Gymnosperms are the most radi�osensitive woody plants because they have large chro�mosomes (Sarapul’tsev et al., 1993). After the Cher�nobyl accident the area of lethal destruction of pineforests was 500–600 ha, and that of pine plantationswith strong and moderate damage was, respectively,3000 and 12000 ha (Kozubov et al., 1994). By the fallof 1959, pines completely died out over an area of2000 ha, which was part of the region contaminated asa result of the accident at PO Mayak (Alexakhin et al.,
2001). Much larger areas of woodlands experiencedradioactive contamination, but its levels were not suf�ficient for the mass death of trees. The long�termeffects of chronic radiation exposure for these forestecosystems remain relevant.
The ability of the population to adapt is related tothe magnitude of genetic polymorphism of featuresthat are subjected to selection (Bradshaw, 1991;Fisher, 1930). There are indications that the techno�logical impact can change the structure of intra�popu�lation variability (Altukhov, 2003; Shevchenko et al.,1992). However, to date the changes that occur in thegene pool of natural populations developing in condi�tions of chronic low�dose exposure, the intensity ofwhich is not sufficient for the manifestation of radia�tion effects in the phenotype of individuals, are stillnot entirely understood. In these conditions it is notclear how the increased frequencies of genetic andcytogenetic damage in somatic and germ cells affectthe reproductive capacity, adaptive differentiation,and the overall fate of populations inhabiting suchareas (Geras’kin et al., 2009; Geras’kin et al., 2011;Volkova et al., 2012).
Using the electrophoretic methods of analysis onecan identify point mutations that cannot be detectedat the higher level of organization and establish a rela�tionship of homozygotes and heterozygotes in thesample. Histochemical staining techniques have beendeveloped for many enzymes. The isozyme polymor�phism analysis allows estimating the frequency ofmutational events for specific loci and the population
Enzyme Polymorphism of an Antioxidant System in Chronically Irradiated Scots Pine Populations
P. Yu. Volkova and S. A. GeraskinLaboratory for Ecotoxicology and Radiobiology of Plants, Russian Institute of Agricultural Radiology and Agroecology,
sh. Kievskoe 109 km, Obninsk, 249030 Russiae�mail: [email protected], [email protected]
Received February 22, 2013; in final form, July 11, 2013
Abstract—This paper studies enzyme polymorphism of the antioxidant system in endosperms and embryosof Scots pine seeds from the areas of the Bryansk region that have been contaminated by the Chernobylnuclear disaster. The mutation frequency in isozyme loci, the effective number of alleles, and the heterozy�gosity increase with the increasing dose absorbed in reproductive organs of pine. The experimental popula�tions demonstrate the increased intrapopulation diversity and frequency of occurrence of rare alleles. Thegenetic differentiation of the studied populations is caused by the increased frequency of occurrence of rarealleles.
Keywords: Pinus sylvestris, Chernobyl accident, contamination, chronic exposure, antioxidant enzymes,polymorphism, rare alleles, null alleles
DOI: 10.1134/S2079059714050153

422
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
VOLKOVA, GERASKIN
as a whole and is traditionally one of the most impor�tant methods for the study of genetic processes in pop�ulations (Altukhov, 2003).
The exposure of living organisms to ionizing radia�tion is largely due to the radiolysis of water moleculeswith the formation of highly toxic reactive oxygen spe�cies (ROS) (Kudryashov, 2004). Their eliminationincludes participation of a number of antioxidantenzymes, the most important of which are superoxidedismutase, catalase, peroxidase, and enzymes involvedin the ascorbate�glutathione cycle. These enzymes aredistributed over the cell compartments where ROS areusually formed as a result of normal metabolism.Under stress the number of enzyme molecules andtheir polymorphism may increase (Polesskaya, 2007).
The aim of the work was to analyze polymorphismof the antioxidant system enzymes—superoxide dis�mutase (SOD, E.C. 1.15.1.1), glutathione reductase(GR, E.C. 1.6.4.2), and glutathione peroxidase (GPX,E.C. 1.11.4.2)—in Scots pine populations from areasin Bryansk region that have been contaminated as aresult of the Chernobyl accident.
MATERIALS AND METHODS
Object of Research
Scots pine, Pinus sylvestris L., is the main forest�forming species of Northern Eurasia. In the forestcommunities, pine is an edificator that largely affectsthe structure and functions of ecosystems. The data onhigh radiosensitivity of conifers were obtained in theearly 1960s at Brookhaven National Laboratory in the
United States (Sparrow et al., 1968) and confirmed inthe experiment “Ekos” in the Southern Urals (Kara�ban’ et al., 1979). The reproductive organs of conifers(Kozubov et al., 1994) are most sensitive to radiationdamage. If in the majority of angiosperm species thereproductive cycle lasts for several months, then inpine this process, starting from the moment of found�ing the generative organs until seed maturation, takesmore than two years (Kozubov et al., 1994). In thecontext of chronic exposure to anthropogenic factorssuch a long development cycle leads to the accumula�tion of sufficient seeds in the cells to indicate the exter�nal influences of the DNA damage. The pineendosperm is haploid, which makes it easy to electro�phoretically estimate the number and spectrum ofmutations in the early postgametic stages.
Study Area
The study was carried out in the most radioactivelycontaminated districts of Bryansk region—Novozy�bkovsky, Klintsovsky, and Krasnogorsky. The experi�mental sites of the All�Russian Research Institute ofFertilizers and Soil (VIUA), Starye Bobovichi (SB),Zaborye Pole (Z1), and Zaborye Kladbishche (Z2) arelocated approximately 200 km northeast of Chernobyl(Fig. 1). Immediately after the accident, the radioac�tive contamination of some sites in this territory wasmore than 1 MBq/m2 (Ramzaev et al., 2008). Thesites are homogeneous in terms of the climatic charac�teristics and physico�chemical properties of soils, theforest stand is homogeneous, and pine trees occupy a
Chernobyl NPP
Gomel
Z2Z1
VIUA
SB
Control
Bryansk
Fig. 1. Location of experimental sites in Bryansk region.

RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
ENZYME POLYMORPHISM OF AN ANTIOXIDANT SYSTEM 423
significant part of the phytocenosis. The control site(K) is located in the Vygonichsky district of Bryanskregion.
The experimental pine populations were chosen toensure an even step in terms of the exposure dose inthe places where they grow. The exposure dose wasmeasured using a dosimeter DRG�01 T at 1 m fromthe ground in 5 to 7 replicates under each tree fromwhich cones were selected and translated into units ofabsorbed dose (1 P = 8.76 × 10–3 Gy). The dose rate inthe air in the experimental sites ranged within 0.37–1.21 μGy/h (0.1 μGy/h in the control site). Thedetailed description of the experimental sites is pre�sented in (Geras’kin et al., 2011).
Research Sampling
In December 2009–2010, the soil and biologicalmaterial (cones) were sampled to determine the levelof contamination with radionuclides and chemicalelements, as well as to carry out the isozyme analysis.The soil samples were collected at each site in layers of0–5, 5–10, and 10–15 cm under the tree where themaximum dose on a site was recorded. On each of thefive sites the cones were collected at altitudes rangingfrom 1.5 to 2 meters above the ground from 20–29trees aged 30–50 years within a homogeneous treestand, 30–50 pieces per tree. For maturation andstratification, the cones were kept outdoors under nat�ural background radiation until the end of February.Then the cones were delivered to the laboratory andstored at room temperature and low humidity untildisclosure and emptying; the seeds were dewingedmanually. For the electrophoretic analysis, we usedonly freely spilled out, well�formed seeds.
Determination of the Physico�Chemical Characteristics of the Soil and the Content of Heavy Metals
in the Soil and Cones
In the soil samples, we estimated the pH value,content of humus and main mineral components, cat�ion exchange capacity, and hydrolytic acidity.
To evaluate the degree of contamination by heavymetals, the soil samples and cones were analyzed forconcentrations of Cd, Cu, Co, Cr, Mn, Ni, Pb, andZn. The heavy metal concentrations were determinedusing a plasma optical emission spectrometer accord�ing to the ISO 11047 standard.
Determination of Radionuclidesin Soil Samples and Biological Material
The contents of 40K, 137Cs, 226Ra, and 232Th in thesoil samples were determined by γ�spectrometry on amultichannel analyzer IN�1200 (France) with a ger�manium detector GEM�1200 (United States).
To estimate the content of radionuclides, one ran�domly selected cone from each tree in the experimen�
tal site was pooled into one sample. The content of137Cs in the cones was determined by γ�spectrometry,while the content of 90Sr was determined by the radio�chemical method in accordance with the Rosleskhozprocedure (Guidelines for the control of radionuclidesin forest resources, 2009) for the radiochemical deter�mination of strontium�90 in soil samples and plantmaterials. The selected samples were mineralized bydry ashing. 90Sr was isolated by the oxalate method byprocessing the ash with the HCl solution at a concen�tration of 6 mol/dm3 when heated to boiling. The con�tent of 90Sr in the analyzed specimens was evaluatedaccording to the formula
A90Sr =
where A90Sr is the specific activity of 90Sr in the ana�lyzed sample, Bq/kg; A is the activity of 90Y in thecounting sample, Bq; WSr is the chemical yield of stron�tium; WY is the chemical yield of yttrium; and m is themass of the sample analyzed in the air�dry state, kg.
Isozyme Analysis
For the study of isozyme polymorphism, threeantioxidant enzymes were selected from the experi�mental populations: superoxide dismutase, glu�tathione reductase, and glutathione peroxidase. Theenzymes were extracted from the endosperm andScots pine seed embryos collected from each tree indi�vidually. Each endosperm and embryo were homoge�nized in 100 μL of extraction buffer: 1% solution ofTriton X�100 and 0.2% solution of β�mercaptoetha�nol. The extracts were separated in vertical plates of7.5% polyacrylamide gel in Tris�HCl buffer withpH 8.0 using the device Hoefer SE 600 Chroma. Thegel was polymerized for 30 min. A Tris�glycine bufferwith a pH of 8.9 was used as an electrode buffer. Theelectrophoresis time was 1.5–2 h at a current of 60–80 mA. Histochemical staining was carried outaccording to (Manchenko, 1994) with modifications.On average, 15 endosperms and embryos per tree wereanalyzed. The seeds of the trees from different siteswere mixed and encrypted; and the samples were iden�tified at the end of the experiment, after decipheringand documenting the zymograms. The samples of2009 were analyzed for polymorphism of superoxidedismutase in the endosperms and pine seed embryos,and the samples of 2010 were analyzed for polymor�phism of glutathione reductase and glutathione perox�idase in the endosperms. The pine seed embryos in thesamples of 2010 were not analyzed, since due to thelow concentration of enzymes in the embryo extract, itwas difficult to identify the zymograms. In total, morethan 9000 locus tests were analyzed.
The allozymes were identified by their relative elec�trophoretic mobility. The most common allozyme wasdesignated as 1.00, the rest of the allozymes of thislocus were designated according to their electro�
A90Sr
WSrWYm������������������,

424
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
VOLKOVA, GERASKIN
phoretic mobility relative to 1.00. Three types ofmutational events in the isozyme loci were assessed(Fig. 2). Null mutations occur as a result of deletionsor gene mutations that lead to either suppression of thefunctional activity of the enzyme or termination of itsbiosynthesis. Other types of detected mutationalevents are mutations, which alter the structure andcharge of the isoenzyme and can be identified by thechange in the electrophoretic mobility of the locusproduct, and duplications, which may arise as a resultof unequal crossing�over during oogenesis, when twoalleles of a single locus are combined in a single chro�mosome.
In the zymograms, null mutations are defined asthe absence of an enzyme activity band. When analyz�ing some seeds, null mutations were identified in sev�eral loci on one zymogram. These seeds were consid�ered dead and were not taken into account in the anal�ysis. Duplications are characterized by the appearanceof two zones of activity in the zymogram that arelocated one above the other. Changes in the electro�phoretic mobility of the isoenzyme were recognizedwhen the locus product was detected outside the pre�viously identified areas of activity of different alleles.Thus, if the locus revealed two 1.00 and 1.10 alleles,the single appearance of a band at the level of 0.90 wasconsidered to be a mutational event.
Statistical Analysis of the Experimental Data
For each population, allele frequencies and param�eters describing the genetic variability were calculated.To assess frequencies of alleles, an unbiased frequencyestimator was used (Zhivotovsky, 1991) according tothe formula
where n is the number of detections of the test allele inthe sample and N is the total sample size. In the case ofa haploid genotype such an assessment is justifiedsince we are not dealing with combinations of alleles.
p nN���,=
The statistical error was determined by the formula
To assess the phenotypic diversity of populations,we used the Zhivotovsky index (μ), which varies from1 to m, where m is the number of alleles and is inter�preted as the average number of phenotypes in thesample. The Zhivotovsky index was calculated by theformula
where p1, …, pm are allele frequencies in the popula�tion.
The statistical error was determined by the formula
To specify the allozyme structure of populations,we assessed the share of rare morphs:
The statistical error was determined by the formula
The sum of the alleles of a given locus is often usedto assess the allelic diversity of populations. However,their number is more informative not in absoluteterms, but rather when taking into account the fre�quency of occurrence: the smaller the allele frequency,the smaller the contribution it makes to the allelicdiversity of the locus. One of the measures of allelicdiversity is the effective number of alleles ne, which isdetermined by the formula
The significance of differences in terms of allelefrequencies between the control and experimentalpopulation was estimated using a modified χ2�test rec�ommended by (Zhivotovsky, 1991) for the case ofsmall sample sizes and the presence of rare phenotypesin the population. The statistics of the test and the
number of degrees of freedom were calculated as =
cχ2; νG = cν, where c is the correction that takes intoaccount the smallness of the expected numbers ofsome alleles. To determine differences between themean values, the Student’s test was used.
For estimating the expected heterozygosity, weused the following formula (Zhivotovsky, 1991):
The statistical error was determined by the formula
spp 1 p–( )
N����������������.=
μ p1 p2 … pm+ + +( )2,=
sμμ m μ–( )
N������������������.=
hμ 1 μ/m( ).–=
shhμ 1 hμ–( )
N��������������������.=
ne 1/Σp12.=
χG2
He 1 pi2.
m
∑–=
1
2
3
4
Fig. 2. Three types of mutations detected on a zymogram.1 norm; 2 duplication; 3 null mutation; 4 change in elec�trophoretic mobility.

RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
ENZYME POLYMORPHISM OF AN ANTIOXIDANT SYSTEM 425
sp2 n 1–( )
n3���������������� 3 2n–( ) 1 He–( )
2 2 n 2–( ) pi3
i 1=
m
∑ 1 He–( )++ ,=
where n = 2N is the number of alleles, and for calcu�lating the observed heterozygosity, we used (Zhivot�ovsky, 1991)
where N is the total number of individuals and Nij is thenumber of individuals classified as heterozygotes.
The statistical error was determined by the formula
Nei’s genetic distance (D), which was calculatedby the formula (Nei, 1972),
was used as metrics in the hierarchical cluster analysisby the method of unweighted arithmetic averages,
where is the averaged theoretical homozygosity over
all loci in the first sample, and is the averaged the�oretical homozygosity over all loci in the second sam�
ple, and is the mutual identity of the two samplesover all loci. Based on the matrix of genetic distancesusing the Statistica program, a dendrogram showingthe hierarchy of the populations under study was built.
To analyze the results obtained using the above for�mulas, Microsoft Office Excel software was used. Theformulas were prescribed independently.
RESULTS
Anthropogenic Contamination of the Experimental Sites And the Doses Absorbed in the Generative Organs of Pine
The soil in all the experimental sites are close interms of physico�chemical properties, and the contentof heavy metals and their mobile forms available to
Ho1N��� Nij,
i j≠
∑=
spHo 1 Ho–( )
N���������������������� .=
D rln– Jpqln 12�� Jpln Jqln+( ),+–= =
Jp
Jq
Jpq
plants does not exceed the permissible levels(Geras’kin et al., 2011; Geras’kin et al., 2008). A qual�itatively different picture is observed in relation to theradioactive contamination of the experimental sites, themain contribution to which is made by 137Cs—the lead�ing dose�forming radionuclide in the areas contami�nated by the Chernobyl disaster. The specific activity ofthis radionuclide in a 5 cm layer of soil varies dependingon the site in the range of 1570–96900 Bq/kg (Table 1).Thus, the specific activity of 137Cs in the soils of theexperimental sites is by 20–1320 times higher thanthat in the control site. The radionuclide content inthe cones of trees from the radioactively contaminatedsites is significantly higher than this parameter for thecontrol populations. Moreover, the content of 137Cs inthe cones of all the studied sites is higher than the con�tent of 90Sr by at least an order of magnitude, althoughit is known (Ipatyev, 1999) that trees absorb 90Sr 5–10 times more from the soil than 137Cs. The observedratio of radionuclides in the cones is due to the lowcontent of 90Sr in the studied soils. The maximumcontent of 137Cs and 90Sr in the cones of the experi�mental sites exceeds this parameter for the controlsites by factors of 315 and 78, respectively (Geras’kinet al., 2011).
To estimate the dose loads on the generative organsof pine trees, a dosimetric model was developed (Spir�idonov et al., 2008; Geras’kin et al., 2011). The resultsof calculating the doses absorbed by the pine genera�tive organs are given in Table 1. The main contributionto the formation of the dose received by the pine gen�erative organs, according to calculations using thedosimetric model, is made by the 137Cs contained inthe upper 10 cm layer of soil. The contribution ofβ�radiation, depending on the site, is 0.4–9.3%. Thecalculation results indicate that the dose absorbed ingenerative organs from the radionuclides distributed inthe forest canopy is formed mainly due to β�radiation.
Table 1. Radioactive contamination of experimental sites and doses absorbed in generative organs of pine
Site
137Cs, 0–5 cm,
Bq/kg
Dose rate in air, µGy/h
Radionuclide concentrations in cones, Bq/kg Dose rate, mGy/year
137Cs 90Sr Dγ, mGy/year
Dβ, mGy/year
Dtotal, mGy/year
K 73.2 0.10 ± 0.01 30.0 ± 3.7 1.32 ± 0.65 0.12 0.01 0.14
VIUA 1570 0.37 ± 0.03 948 ± 13 16.79 ± 1.93 6.6 0.3 6.9
SB 16900 0.49 ± 0.16 342 ± 8 20.81 ± 1.31 22.7 0.2 22.9
Z1 39190 0.73 ± 0.18 3246 ± 32 64.25 ± 2.20 90.2 1.2 91.4
Z2 96900 1.21 ± 0.23 1008 ± 18 83.00 ± 2.19 129.4 0.5 129.9

426
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
VOLKOVA, GERASKIN
Isozyme Polymorphism GPX and GRin Populations of Scots Pine
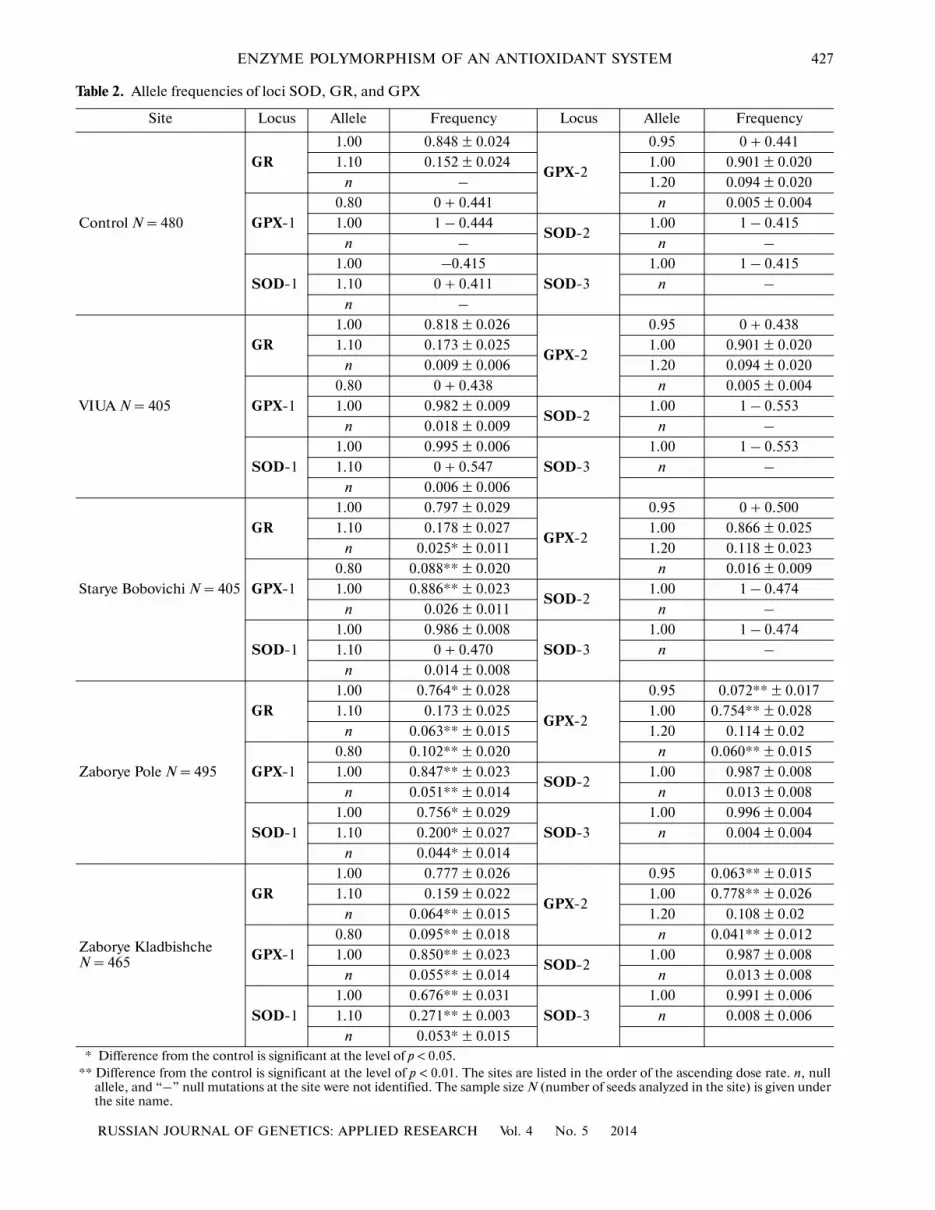
Polymorphism of the studied populations in termsof the SOD enzyme was described in detail previously(Volkova et al., 2012). One polymorphic locus wasidentified in glutathione reductase, three—in glu�tathione peroxidase, but only two of them, having bet�ter resolution, were used for the analysis. The analysisof loci GR and GPX�1 revealed two alleles, while locusGPX�2 revealed three. The obtained data (Table 2) sug�gest that the increase in the dose absorbed by genera�tive organs of plants is accompanied by a growingnumber and frequency of rare alleles. The correlationcoefficient is 85% for (GPX�1, 0.80); 86% for (GPX�2,0.95); 62% for (GPX�2, 1.20); and 63% for (SOD�1,1.10).
Frequency and Spectrum of Mutationsat the Loci of the Studied Isozymes
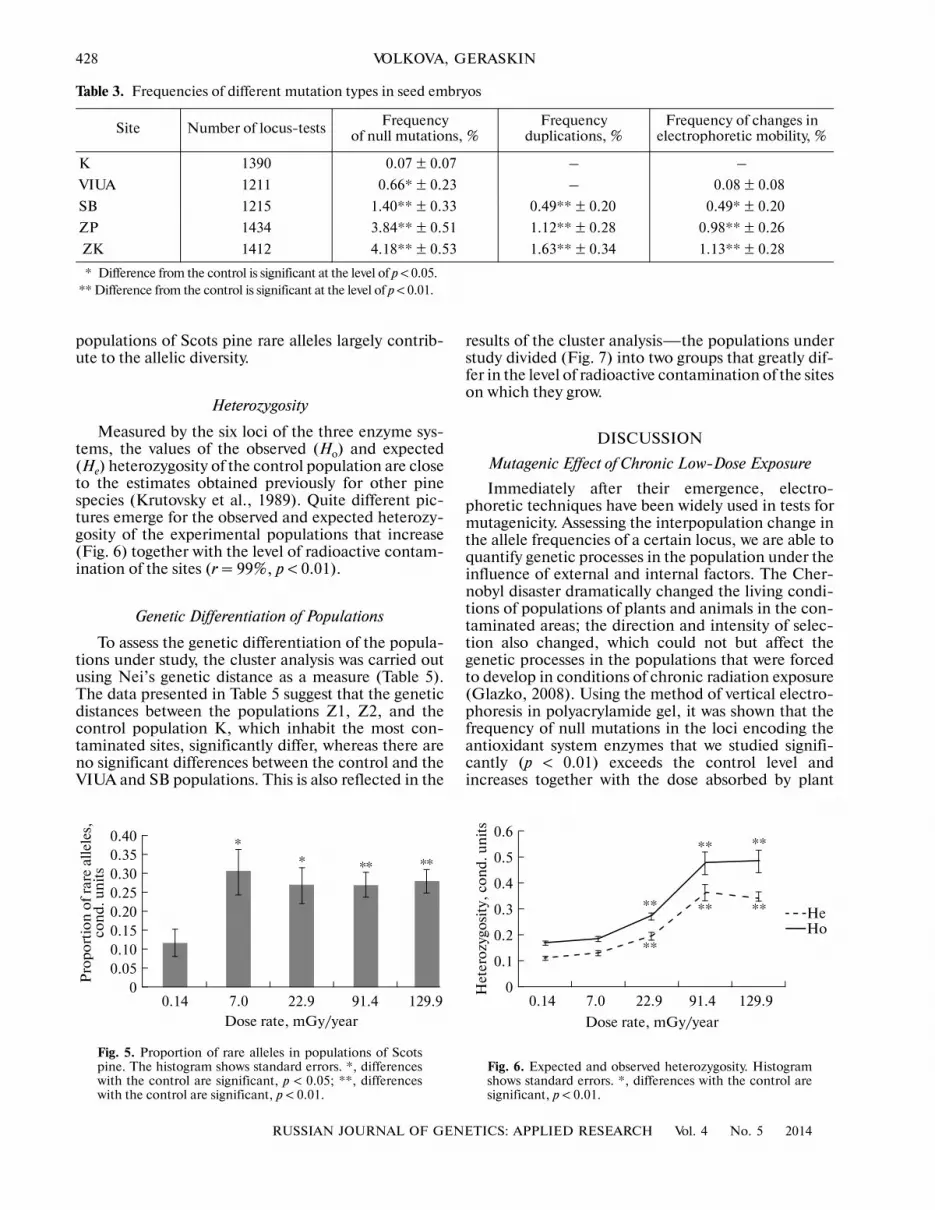
Three types of mutational events in isozyme lociare estimated. The largest contribution to the overallfrequency of mutations is made by null mutations.Table 3 shows the frequency of the mutations of all theconsidered types by six enzyme loci that encode thebiosynthesis of three antioxidant enzymes. Theobtained data are in good agreement with other studies(Altukhov et al., 1983; Fedotov et al., 2006), whichshow that changes in electrophoretic mobility andduplication are relatively rare types of mutations com�pared to null mutations. Mutations in the control pop�ulation are extremely rare and their frequency corre�sponds to the level of spontaneous mutagenesis ofisozyme loci in Scots pine (Altukhov et al., 1983). Theaverage frequency of mutations (Fig. 3) over threeenzymes significantly (r = 99%, p < 0.01) increasestogether with the growth of the dose absorbed by plantgenerative organs.
Intrapopulation Diversity
In terms of the value of the Zhivotovsky index (theaverage number of alleles in the population with regardto their frequency), the studied loci can be divided(Zhivotovsky, 1991) into three groups: monomorphic,with average (1 < μ < 3) and high (μ > 3) levels of vari�ability. All the loci that we studied fall into the groupwith an average level of variability. The indices ofgenotypic diversity in the populations studied are sig�nificantly different from the control (Fig. 4). The thusobtained variability underestimates the one actuallyexisting in the population, since some of single aminoacid replacements (which are synonymous) are notdetected by electrophoresis (Altukhov, 2003).
Proportion of Rare Alleles and the Effective Number of Alleles
The proportion of rare alleles characterizes the popu�lation genotypic structure by estimating the ratiobetween the frequencies of the rarest and the mostcommon genotypes in the sample. This parametersharply increases (Fig. 5) in the populations inhabitingthe areas contaminated with radionuclides, which isassociated with the emergence of new alleles in severalenzyme systems (null alleles, allele 0.80 in the systemGPX�1, and allele 0.95 in the system GPX�2). Withincreasing levels of radioactive contamination, onlythe frequency of occurrence of rare alleles changes,rather than their number, so that the values of thisparameter for different experimental sites are not sig�nificantly different.
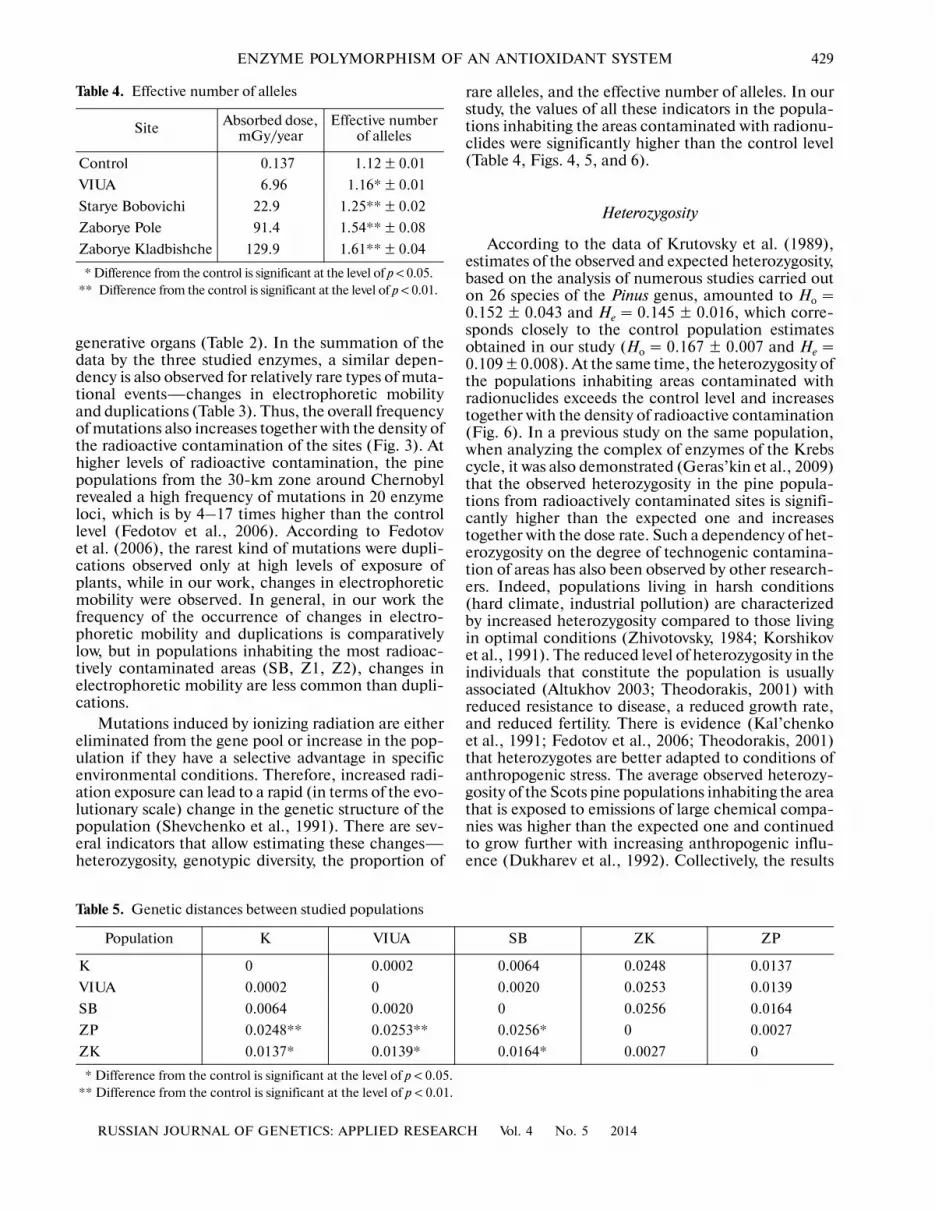
The effective number of alleles characterizes theircontribution to the allelic diversity of loci. Thisparameter increases (Table 4) together with the level ofradiation exposure, since in chronically irradiated
0.08
0.07
0.06
0.05
0.04
0.03
0.02
0.01
00.14 7.0 22.9 91.4 129.9
****
*
*Fre
quen
cy o
f mut
atio
ns
Dose rate, mGy/year
Fig. 3. Overall frequency of mutations in seed embryos bythree enzymes. Histogram shows standard errors. *, differ�ences with the control are significant, p < 0.05; **, differ�ences with the control are significant, p < 0.01.
2.0
1.5
1.0
0.5
00.14 7.0 22.9 91.4 129.9
****
*
2.5
**
Inde
x of
gen
otyp
ic
Dose rate, mGy/year
dive
rsit
y, c
ond.
un
its
Fig. 4. Genotypic diversity of Scots pine populations bythree enzymes. Histogram shows standard errors. *, differ�ences with the control are significant, p < 0.05; **, differ�ences with the control are significant, p < 0.01.
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
ENZYME POLYMORPHISM OF AN ANTIOXIDANT SYSTEM 427
Table 2. Allele frequencies of loci SOD, GR, and GPX
Site Locus Allele Frequency Locus Allele Frequency
Control N = 480
GR1.00 0.848 ± 0.024
GPX�2
0.95 0 + 0.441
1.10 0.152 ± 0.024 1.00 0.901 ± 0.020
n – 1.20 0.094 ± 0.020
GPX�1
0.80 0 + 0.441 n 0.005 ± 0.004
1.00 1 – 0.444SOD�2
1.00 1 – 0.415
n – n –
SOD�1
1.00 –0.415
SOD�3
1.00 1 – 0.415
1.10 0 + 0.411 n –
n –
VIUA N = 405
GR1.00 0.818 ± 0.026
GPX�2
0.95 0 + 0.438
1.10 0.173 ± 0.025 1.00 0.901 ± 0.020
n 0.009 ± 0.006 1.20 0.094 ± 0.020
GPX�1
0.80 0 + 0.438 n 0.005 ± 0.004
1.00 0.982 ± 0.009SOD�2
1.00 1 – 0.553
n 0.018 ± 0.009 n –
SOD�1
1.00 0.995 ± 0.006
SOD�3
1.00 1 – 0.553
1.10 0 + 0.547 n –
n 0.006 ± 0.006
Starye Bobovichi N = 405
GR1.00 0.797 ± 0.029
GPX�2
0.95 0 + 0.500
1.10 0.178 ± 0.027 1.00 0.866 ± 0.025
n 0.025* ± 0.011 1.20 0.118 ± 0.023
GPX�1
0.80 0.088** ± 0.020 n 0.016 ± 0.009
1.00 0.886** ± 0.023SOD�2
1.00 1 – 0.474
n 0.026 ± 0.011 n –
SOD�1
1.00 0.986 ± 0.008
SOD�3
1.00 1 – 0.474
1.10 0 + 0.470 n –
n 0.014 ± 0.008
Zaborye Pole N = 495
GR1.00 0.764* ± 0.028
GPX�2
0.95 0.072** ± 0.017
1.10 0.173 ± 0.025 1.00 0.754** ± 0.028
n 0.063** ± 0.015 1.20 0.114 ± 0.02
GPX�1
0.80 0.102** ± 0.020 n 0.060** ± 0.015
1.00 0.847** ± 0.023SOD�2
1.00 0.987 ± 0.008
n 0.051** ± 0.014 n 0.013 ± 0.008
SOD�1
1.00 0.756* ± 0.029
SOD�3
1.00 0.996 ± 0.004
1.10 0.200* ± 0.027 n 0.004 ± 0.004
n 0.044* ± 0.014
Zaborye Kladbishche N = 465
GR1.00 0.777 ± 0.026
GPX�2
0.95 0.063** ± 0.015
1.10 0.159 ± 0.022 1.00 0.778** ± 0.026
n 0.064** ± 0.015 1.20 0.108 ± 0.02
GPX�1
0.80 0.095** ± 0.018 n 0.041** ± 0.012
1.00 0.850** ± 0.023SOD�2
1.00 0.987 ± 0.008
n 0.055** ± 0.014 n 0.013 ± 0.008
SOD�1
1.00 0.676** ± 0.031
SOD�3
1.00 0.991 ± 0.006
1.10 0.271** ± 0.003 n 0.008 ± 0.006
n 0.053* ± 0.015 * Difference from the control is significant at the level of p < 0.05. ** Difference from the control is significant at the level of p < 0.01. The sites are listed in the order of the ascending dose rate. n, null
allele, and “–” null mutations at the site were not identified. The sample size N (number of seeds analyzed in the site) is given underthe site name.
428
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
VOLKOVA, GERASKIN
populations of Scots pine rare alleles largely contrib�ute to the allelic diversity.
Heterozygosity
Measured by the six loci of the three enzyme sys�tems, the values of the observed (Ho) and expected(He) heterozygosity of the control population are closeto the estimates obtained previously for other pinespecies (Krutovsky et al., 1989). Quite different pic�tures emerge for the observed and expected heterozy�gosity of the experimental populations that increase(Fig. 6) together with the level of radioactive contam�ination of the sites (r = 99%, p < 0.01).
Genetic Differentiation of Populations
To assess the genetic differentiation of the popula�tions under study, the cluster analysis was carried outusing Nei’s genetic distance as a measure (Table 5).The data presented in Table 5 suggest that the geneticdistances between the populations Z1, Z2, and thecontrol population K, which inhabit the most con�taminated sites, significantly differ, whereas there areno significant differences between the control and theVIUA and SB populations. This is also reflected in the
results of the cluster analysis—the populations understudy divided (Fig. 7) into two groups that greatly dif�fer in the level of radioactive contamination of the siteson which they grow.
DISCUSSION
Mutagenic Effect of Chronic Low�Dose Exposure
Immediately after their emergence, electro�phoretic techniques have been widely used in tests formutagenicity. Assessing the interpopulation change inthe allele frequencies of a certain locus, we are able toquantify genetic processes in the population under theinfluence of external and internal factors. The Cher�nobyl disaster dramatically changed the living condi�tions of populations of plants and animals in the con�taminated areas; the direction and intensity of selec�tion also changed, which could not but affect thegenetic processes in the populations that were forcedto develop in conditions of chronic radiation exposure(Glazko, 2008). Using the method of vertical electro�phoresis in polyacrylamide gel, it was shown that thefrequency of null mutations in the loci encoding theantioxidant system enzymes that we studied signifi�cantly (p < 0.01) exceeds the control level andincreases together with the dose absorbed by plant
Table 3. Frequencies of different mutation types in seed embryos
Site Number of locus�tests Frequency of null mutations, %
Frequency duplications, %
Frequency of changes in electrophoretic mobility, %
K 1390 0.07 ± 0.07 – –
VIUA 1211 0.66* ± 0.23 – 0.08 ± 0.08
SB 1215 1.40** ± 0.33 0.49** ± 0.20 0.49* ± 0.20
ZP 1434 3.84** ± 0.51 1.12** ± 0.28 0.98** ± 0.26
ZK 1412 4.18** ± 0.53 1.63** ± 0.34 1.13** ± 0.28
* Difference from the control is significant at the level of p < 0.05. ** Difference from the control is significant at the level of p < 0.01.
0.35
0.250.20
0.10
00.14 7.0 22.9 91.4 129.9
****
*0.40
*
0.05
0.15
0.30
Pro
port
ion
of r
are
alle
les,
Dose rate, mGy/year
con
d. u
nit
s
Fig. 5. Proportion of rare alleles in populations of Scotspine. The histogram shows standard errors. *, differenceswith the control are significant, p < 0.05; **, differenceswith the control are significant, p < 0.01.
0.5
0.4
0.3
0.2
0.1
0129.922.97.00.14
0.6
91.4
****
** ****
**
HeHo
Het
eroz
ygos
ity,
con
d. u
nit
s
Dose rate, mGy/year
Fig. 6. Expected and observed heterozygosity. Histogramshows standard errors. *, differences with the control aresignificant, p < 0.01.
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
ENZYME POLYMORPHISM OF AN ANTIOXIDANT SYSTEM 429
generative organs (Table 2). In the summation of thedata by the three studied enzymes, a similar depen�dency is also observed for relatively rare types of muta�tional events—changes in electrophoretic mobilityand duplications (Table 3). Thus, the overall frequencyof mutations also increases together with the density ofthe radioactive contamination of the sites (Fig. 3). Athigher levels of radioactive contamination, the pinepopulations from the 30�km zone around Chernobylrevealed a high frequency of mutations in 20 enzymeloci, which is by 4–17 times higher than the controllevel (Fedotov et al., 2006). According to Fedotovet al. (2006), the rarest kind of mutations were dupli�cations observed only at high levels of exposure ofplants, while in our work, changes in electrophoreticmobility were observed. In general, in our work thefrequency of the occurrence of changes in electro�phoretic mobility and duplications is comparativelylow, but in populations inhabiting the most radioac�tively contaminated areas (SB, Z1, Z2), changes inelectrophoretic mobility are less common than dupli�cations.
Mutations induced by ionizing radiation are eithereliminated from the gene pool or increase in the pop�ulation if they have a selective advantage in specificenvironmental conditions. Therefore, increased radi�ation exposure can lead to a rapid (in terms of the evo�lutionary scale) change in the genetic structure of thepopulation (Shevchenko et al., 1991). There are sev�eral indicators that allow estimating these changes—heterozygosity, genotypic diversity, the proportion of
rare alleles, and the effective number of alleles. In ourstudy, the values of all these indicators in the popula�tions inhabiting the areas contaminated with radionu�clides were significantly higher than the control level(Table 4, Figs. 4, 5, and 6).
Heterozygosity
According to the data of Krutovsky et al. (1989),estimates of the observed and expected heterozygosity,based on the analysis of numerous studies carried outon 26 species of the Pinus genus, amounted to Ho =0.152 ± 0.043 and He = 0.145 ± 0.016, which corre�sponds closely to the control population estimatesobtained in our study (Ho = 0.167 ± 0.007 and He =0.109 ± 0.008). At the same time, the heterozygosity ofthe populations inhabiting areas contaminated withradionuclides exceeds the control level and increasestogether with the density of radioactive contamination(Fig. 6). In a previous study on the same population,when analyzing the complex of enzymes of the Krebscycle, it was also demonstrated (Geras’kin et al., 2009)that the observed heterozygosity in the pine popula�tions from radioactively contaminated sites is signifi�cantly higher than the expected one and increasestogether with the dose rate. Such a dependency of het�erozygosity on the degree of technogenic contamina�tion of areas has also been observed by other research�ers. Indeed, populations living in harsh conditions(hard climate, industrial pollution) are characterizedby increased heterozygosity compared to those livingin optimal conditions (Zhivotovsky, 1984; Korshikovet al., 1991). The reduced level of heterozygosity in theindividuals that constitute the population is usuallyassociated (Altukhov 2003; Theodorakis, 2001) withreduced resistance to disease, a reduced growth rate,and reduced fertility. There is evidence (Kal’chenkoet al., 1991; Fedotov et al., 2006; Theodorakis, 2001)that heterozygotes are better adapted to conditions ofanthropogenic stress. The average observed heterozy�gosity of the Scots pine populations inhabiting the areathat is exposed to emissions of large chemical compa�nies was higher than the expected one and continuedto grow further with increasing anthropogenic influ�ence (Dukharev et al., 1992). Collectively, the results
Table 4. Effective number of alleles
Site Absorbed dose, mGy/year
Effective number of alleles
Control 0.137 1.12 ± 0.01
VIUA 6.96 1.16* ± 0.01
Starye Bobovichi 22.9 1.25** ± 0.02
Zaborye Pole 91.4 1.54** ± 0.08
Zaborye Kladbishche 129.9 1.61** ± 0.04
* Difference from the control is significant at the level of p < 0.05.** Difference from the control is significant at the level of p < 0.01.
Table 5. Genetic distances between studied populations
Population K VIUA SB ZK ZP
K 0 0.0002 0.0064 0.0248 0.0137
VIUA 0.0002 0 0.0020 0.0253 0.0139
SB 0.0064 0.0020 0 0.0256 0.0164
ZP 0.0248** 0.0253** 0.0256* 0 0.0027
ZK 0.0137* 0.0139* 0.0164* 0.0027 0
* Difference from the control is significant at the level of p < 0.05.** Difference from the control is significant at the level of p < 0.01.

430
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
VOLKOVA, GERASKIN
of these studies show a selective advantage of het�erozygotes in conditions of chronic anthropogenicexposure.
Genotypic Diversity
The data obtained for the three enzymes allows usto suggest that genetic processes in populations areaimed at increasing the genotypic diversity, which canbe estimated using the Zhivotovsky index. The geno�typic diversity over all the studied loci is significantly(p < 0.01) higher than the control level (Fig. 4), whichis associated (Geras’kin et al., 2010) with the reactionof the population to stress: under the pressure ofanthropogenic factors, the genetic and phenotypicvariability increases; however, under excessive levels ofstress the most sensitive individuals are eliminated,and the intrapopulation diversity declines (Dukharevet al., 1992; Shuiskaya et al., 2012). The level of intrap�opulation diversity in the plant populations inhabitingthe contaminated territories is defined by the species’sensitivity, intensity, and the nature of the anthropo�genic impact. This thesis is confirmed on the exampleof investigating the allozyme structure of the commondandelion populations that inhabit banks of the TechaRiver in the Southern Urals, in which the indices ofthe intrapopulation diversity of the background andexperimental populations did not differ significantly(Ul’yanova et al., 2004) despite the doses absorbed byplants which are comparable to our study. The radiore�sistance of gymnosperms is much lower than inangiosperms (Sarapul’tsev et al., 1993). Therefore, toachieve the same effect in populations of angiosperms,much larger doses are required. Indeed, the sameauthors carried out the study of the allozyme structurein dandelion populations from the EURT territory(East Ural Radioactive Trace), where plants receivedmuch higher doses, which showed (Antonov et al.,2007) that the average population values of the Zhivot�ovsky index exceed those of the background values.
Proportion of Rare Alleles and Effective Number of Alleles
The mutation process is the main source of newalleles, which in turn leads to an increase in the geneticdiversity of the population. By the proportion of rarealleles, all the contaminated sites are significantly dif�ferent from the control (Fig. 5), but not from eachother. Such an effect is due to the sharp increase in thelevel of chronic radiation exposure (the power of theannual dose at the control and least contaminatedexperimental site VIUA differs by more than50 times), the mutagenic effect of which is manifestedin the increasing number of rare allelic variants (Table 2).A further increase in the density of radioactive con�tamination leads to an increase in the frequency ofthese variants rather than their number. An increase inthe frequency of rare alleles in populations of greaterknapweed (Centaurea scabiosa L.), which for a fewdecades have been living under conditions of chronicirradiation in the East Ural Radioactive Trace, wasshown in Kal’chenko et al. (1991, 1996).
Genetic Differentiation of Experimental Populations
In the cluster analysis, the populations under studywere divided into two groups (Fig. 7), based either ontheir geographical location or chronic radiation expo�sure. Thus, industrial contamination caused anincrease in polymorphism and genetic diversity of theexperimental populations of Viola tricolor, which ledto their separation into a single cluster (Slomka et al.,2011). However, heavy metal contamination did notaffect (Müller et al., 2007) the genetic subdivision ofpopulations of Suillus luteus. Mengoni et al. (2000)showed that the genetic variability of populations ofSilene affects both the geographical location and heavymetal contamination of soils. In this study, the popu�lations resistant to heavy metals’ contamination weredivided into a separate cluster despite their geographicproximity to less stable populations. Thus, the geneticdifferentiation of populations is affected by the geo�graphic location, species affiliation, type and intensityof anthropogenic impact, and other environmentalfactors. We note that the Z1 and Z2 populations aregeographically much closer to each other than to therest of the populations, so their isolation into a singlecluster may be due to their location and constantexchange of genes, rather than to radiation exposure.The pollen and seed exchange between the K, VIUA,and SB populations is impossible because of the con�siderable distance between them. Nevertheless, theyare combined into one cluster due to the equally lowfrequency of the occurrence of rare alleles.
Role of the Studied Enzymesin the Modification of Radiation Exposure
The metabolism of plant cells under stress condi�tions is characterized by the increased production of
0.020
0.015
0.010
0.005
0
Gen
etic
dis
tan
ceDendrogram based on distance matrix
Z2 Z1 SB VIUA K
Fig. 7. Genetic differentiation of populations studied.

RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
ENZYME POLYMORPHISM OF AN ANTIOXIDANT SYSTEM 431
reactive oxygen species (ROS) (Elstner et al., 1994;Foyer et al., 2000). Radioactive exposure is alsoaccompanied by oxidative stress (Inze, 1995) due tothe radiolysis of water molecules in the extracellularand intracellular environment. In the radiolysis ofwater, many ROS are formed, exclusively reactivemolecules, to neutralize which the cell uses the anti�oxidant defense system. Therefore, under conditionsof chronic radiation exposure, the role of the antioxi�dant enzymes becomes more important. Researchersof isozyme polymorphism in populations underchronic exposure often unjustifiably ignore the mostimportant antioxidant system—glutathione. Con�temporary authors (Tausz et al., 2004) note the centralrole of glutathione metabolism in the response ofplants to stress. We have investigated the key enzymesof this system—glutathione reductase (GR) and glu�tathione peroxidase (GPX). GR restores oxidized glu�tathione using NADPH as a reducing agent, and GPXis involved in the detoxification of hydrogen peroxideand lipid hydroperoxides using GSH as a reducingagent (Gechev et al., 2006). Glutathione itself is a sub�strate for various peroxidases, glutathione peroxidase,and glutathione reductase; it provides detoxifieshydrogen peroxide and other hydroperoxides andtoxic substances. The polymorphism of enzymesSOD, GR, and GPX in the investigated populationsincreases (Table 2, Volkova et al., 2012), together withthe level of radioactive contamination. In the condi�tions of the increased production of reactive oxygenspecies, the role of antioxidant enzymes significantlyincreases, which explains the biochemical polymor�phism that we observed. The occurrence of rare allelesof antioxidant enzymes in a population and theincrease in their frequency can be interpreted as a pro�cess aimed at obtaining an evolutionary advantage inconditions of radiation�induced chronic oxidativestress.
Polymorphism of Antioxidant Systemas a Response to Chronic Radiation Stress
The reserve of ecological plasticity and geneticdiversity of populations is largely determined by thenumber of polymorphic traits in its constituent indi�viduals (Sozinov, 1985). In our study, we identified thata high level of mutagenesis in the polymorphic enzymeloci creates preconditions for the acceleration ofmicro�evolutionary processes in chronically irradiatedpopulations. Since germ cells in the ontogeny of plantsare formed much later than in the ontogeny of ani�mals, radiation�induced genetic changes occurring insomatic cells can be transmitted to progeny (Maletskiiet al., 2007). This fundamental feature lies in the basisof the genotypic and phenotypic plasticity of plantsand their ability to adapt quickly to changing environ�mental conditions.
It remains unclear whether the mutational variabil�ity identified in our study contributes to the adaptation
of populations to chronic radiation exposure. Earlier,on the same populations, it has been shown (Geras’kinet al., 2011) that chronic exposure, significantlyincreasing the frequency of cytogenetic damage, hasno significant effect on the reproductive potential ofpopulations and does not affect the stability of seeds toadditional γ�irradiation. Isozyme polymorphism innatural populations usually is not neutral and is asso�ciated with the role of enzymes in the mechanisms ofneutralization of pollutants or toxic products of theirindirect effect (Altukhov, 2003); that is, different alle�les of polymorphic loci may have a different impact onthe fitness to certain living conditions. Indeed, thepopulations of greater knapweed, which for 12 yearswere developing in conditions of chronic exposure inthe EURT territory, revealed (Kal’chenko et al., 1996)a significant increase in the frequency of allele S thatencodes isozyme SOD, which with increased effi�ciency eliminates reactive oxygen species. The sameauthors found (Fedotov et al., 2006) a strong selectionpressure against the F allele in the populations of Scotspine growing within the 30�km zone of the Chernobyldisaster.
The data obtained show that even relatively lowlevels of chronic exposure (7–130 mGy/year) can leadto an increase in the frequency of mutations inendosperm and seed embryos and alter the geneticstructure of Scots pine populations. The chronicallyexposed pine populations produce progeny with a highlevel of mutational variability which significantlyexceeds the control figures. An increase in radioactivecontamination of the sites up to 39 kBq/kg leads to asignificant increase in the frequency of the mutationsof isozyme loci, the effective number of alleles, intrap�opulation diversity, and the frequency of rare morphs.Thus, despite their low values, the observed dose ratesmay be considered as a factor capable of modifying thegenetic structure of populations.
ACKNOWLEDGMENTS
This work was supported by the Russian Founda�tion for Basic Research (Project nos. 11�04�00670 and11�04�97524) and Russian Science Foundation(Project nos. 14�14�00666).
REFERENCES
Aleksakhin, R.M., Buldakov, L.A., Gubanov, V.A., et al.,Krupnye radiatsionnye avarii: posledstviya i zashchitnyemery (Large Radiation Accidents: Consequences andProtective Measures), Moscow: IzdAT, 2001.
Altukhov, Yu.P., Dukharev, V.A., and Zhivotovskii, L.A.,Selection against rare electrophoretic protein variantsand the rate of spontaneous mutation process in popu�lations, Genetika, 1983, vol. 19, no. 2, pp. 264–275.
Altukhov, Yu.P., Geneticheskie protsessy v populyatsiyakh(Genetic Processes in Populations), Moscow: Aka�demkniga, 2003.

432
RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
VOLKOVA, GERASKIN
Antonova, E.V. and Pozolotina, V.N., Specific features ofthe allozyme structure of dandelion populations underconditions of radionuclide and chemical contamina�tion, Russ. J. Ecol., 2007, vol. 38, no. 5, pp. 327–333.
Bradshaw, A.D., Genostasis and the limits to evolution,Phil. Trans. R. Soc. Lond. B, 1991, vol. 33, pp. 289–305.
Dukharev, V.A., Korshikov, I.I., Ryabokon’, S.M., et al.,Genetic differentiation of subpopulations of Scots pinein the conditions of technogenic pollution, Tsitol.Genet., 1992, vol. 26, no. 3, pp. 7–11.
Elstner, E.F. and Osswald, W., Mechanisms of oxygen acti�vation during plant stress, Proc. Roy. Soc. EdinburghBiol., 1994, vol. 102B, pp. 131–154.
Fedotov, I.S. and Kal’chenko, V.A., Radiation and geneticconsequences of irradiation of Scots pine populationsin the Chernobyl accident area, Radiats. Biol. Radi�oekol., 2006, vol. 46, no. 3, pp. 268–278.
Fisher, R.A., The Genetic Theory of Natural Selection,Oxford: Clarendon Press, 1930.
Foyer, C.H. and Noctor, G., Oxygen processing in photo�synthesis: a molecular approach, New Phytol., 2000,vol. 146, pp. 359–388.
Gechev, S.T., Breusegem, F., Stone, J., et al., Reactive oxy�gen species as signals that modulate plant stressresponses and programmed cell death, BioEssays, 2006,vol. 28, pp. 1091–1101.
Geras’kin, S.A., Dikareva, N.S., Udalova, A.A., et al.,Cytogenetic effects in Scots pine populations from theBryansk region contaminated by radioactive pollutantsas a result of the Chernobyl NPP accident, Radiats.Biol. Radioekol., 2008, vol. 48, no. 5, pp. 584–595.
Geras’kin, S.A., Vanina, Yu.S., Dikarev, V.G., et al.,Genetic variability in Scots pine populations from theBryansk region contaminated by radioactive pollutantsas a result of the Chernobyl NPP accident, Radiats.Biol.: Radioekol., 2009, vol. 49, no. 2, pp. 136–146.
Geras’kin, S.A., Udalova, A.A., Dikareva, N.S., et al., Bio�logical effects of chronic radiation exposure on plantpopulations, Radiats. Biol. Radioekol., 2010, vol. 50,no. 4, pp. 374–382.
Geraskin, S.A., Oudalova, A.A., Dikareva, N.S., et al.,Cytogenetic damage and reproductive effects in Scotspine populations affected by the Chernobyl accident,Ecotoxicology, 2011, vol. 20, pp. 1195–1208.
Glazko, T.T., Arkhipov, N.P., and Glazko, V.I., Populyat�sionno�geneticheskie posledstviya ekologicheskikh katas�trof na primere Chernobyl’skoi avarii (PopulationGenetic Consequences of Environmental Disasters: ACase Study of the Chernobyl Accident), Moscow:MSKhA im. K.A. Timiryazeva, 2008.
Inze, D. and Van Montagu, M., Oxidative stress in plants,Curr. Opin. Biotechnol., 1995, vol. 6, pp. 153–158.
Ipatyev, V., Bulavik, I., Braginsky, V., et al., Forest andChernobyl: forest ecosystems after the Chernobylnuclear power plant accident: 1986–1994, J. Environ.Radioactiv., 1999, vol. 42, pp. 9–38.
Kal’chenko, V.A., Kalabushkin, V.A., and Rubanovich, A.V.,Chronic irradiation as an environmental factor affect�ing the genetic structure of populations, Genetika,1991, vol. 27, no. 4, pp. 676–684.
Kal’chenko, V.A., Rubanovich, A.V., and Shevchenko, V.A.,Adaptive nature of the polymorphism for the superox�ide dismutase locus in chronically irradiated naturalpopulations of Centaurea scabiosa L., Russ. J. Genet.,1996, vol. 32, no. 11, pp. 1307–1310.
Karaban’, R.T., Mishenkov, N.N., Prister, B.S., et al., Theaction of acute γ�radiation on a forest plant community,in Problemy lesnoi radioekologii. Trudy IPG (Problemsof Forest Radioecology. IPG Transactions), Moscow:Gidrometeoizdat, 1979, vol. 38, pp. 27–52.
Korshikov, I.I., Dukharev, V.A., Kotova, A.A., et al., Alloz�ymic polymorphism of GOT, GDH, and SOD loci inScots pine in conditions of technogenic environmentalpollution, Tsitol. Genet., 1991, vol. 25, no. 6, pp. 60–64.
Kozubov, G.M. and Taskaev, A.I., Radiobiologicheskie iradioekologicheskie issledovaniya drevesnykh rastenii(Radiobiological and Radioecological Studies of Arbo�real Plants), St. Petersburg: Nauka, 1994.
Krutovskii, K.V., Politov, D.V., Altukhov, Yu.P., et al.,Genetic variability of Siberian cedar pine Pinus sibiricaDu Tour. IV. Genetic diversity and the degree of geneticdifferentiation between populations, Genetika, 1989,vol. 25, no. 11, pp. 2009–2032.
Kudryashov, Yu.B., Radiatsionnaya biofizika (ioniziruyush�chie izlucheniya) (Radiation Biophysics (IonizingRadiation)), Moscow: FIZMATLIT, 2004.
Maletskii, S.I. and Yudanova, S.S., Germ pathways andstem cells in higher plants, Tsitol. Genet., 2007, no. 5,pp. 67–80.
Manchenko, G.P., Handbook of Detection of Enzymes onElectrophoretic Gels, CRC Press, 1994.
Mengoni, A., Gonnelli, C., Galardi, F., et al., Geneticdiversity and heavy metal tolerance in populations ofSilene paradoxa L.: a random amplified DNA analysis,Mol. Ecol., 2000, vol. 9, pp. 1319–1324.
Muller, L., Vangronsveld, J., and Colpaert, J., Geneticstructure of Suillus leteus populations in heavy metalpolluted and nonpolluted habitats, Mol. Ecol., 2007,vol. 16, pp. 4728–4737.
Nei, M., Genetic distance between populations, Am. Nat.,1972, vol. 106, no. 949, pp. 283–292.
Polesskaya, O.G., Rastitel’naya kletka i aktivnye formykisloroda (Plant Cell and Reactive Oxygen species),Moscow: KDU, 2007.
Ramzaev, V., Botter�Jensen, L., Thompsen, K.J., et al., Anassessment of cumulative external doses from Cherno�byl fallout for a forested area in Russia using opticallystimulated luminiscence from quartz inclusions inbricks, J. Environ. Radioactiv., 2008, vol. 99, pp. 1154–1164.
Sarapul’tsev, B.I. and Geras’kin, S.A., Geneticheskie osnovyradiorezistentnosti i evolyutsiya (Genetic Basics ofRadioresistance and Evolution), Moscow: Energoat�omizdat, 1993.
Shevchenko, V.A., Pechkurenkov, V.L., and Abramov, V.I.,Radiatsionnaya genetika prirodnykh populyatsii (Radia�tion Genetics of Natural Populations), Moscow:Nauka, 1992.
Shuiskaya, E.V., Gismatullina, L.G., Toderich, K.N., et al.,Genetic differentiation of black saxaul, Haloxylon aph�yllum (Chenopodiaceae), along a soil salinity gradient

RUSSIAN JOURNAL OF GENETICS: APPLIED RESEARCH Vol. 4 No. 5 2014
ENZYME POLYMORPHISM OF AN ANTIOXIDANT SYSTEM 433
in the Kyzylkum desert, Russ. J. Ecol., 2012, vol. 43,no. 4, pp. 302–306.
Slomka, A., Sutkowska, A., Szczepaniak, M., et al.,Increased genetic diversity of viola tricolor l. in metal�polluted environments, Chemosphere, 2011, vol. 83,pp. 435–442.
Sozinov, A.A., Polimorfizm belkov i ego znachenie v genetikei selektsii (Polymorphism of Proteins and Its Impor�tance in Genetics and Breeding), Moscow: Nauka,1985.
Sparrow, A.H., Rogers, A.F., and Schwemmer, S.S., Radi�osensitivity studies with woody plants, Radiat. Bot.,1968, vol. 8, pp. 149–186.
Spiridonov, S.I., Fesenko, S.V., Geras’kin, S.A., et al.,Evaluation of irradiation doses of woody plants in theremote period after the Chernobyl accident, Radiats.Biol. Radioekol., 2008, vol. 48, no. 4, pp. 432–438.
Tausz, M., Sircelj, H., and Grill, D., The glutathione sys�tem as a stress market in plant ecophysiology: is a stress�response concept valid, J. Exp. Bot., 2004, vol. 55,no. 404, pp. 1955–1962.
Theodorakis, C.W., Integration of genotoxic and popula�tion genetic endpoints in biomonitoring and risk assess�ment, Ecotoxicology, 2001, vol. 10, pp. 245–256.
Tikhomirov, F.A., Deistvie ioniziruyushchikh izluchenii naekologicheskie sistemy (Effects of Ionizing Radiation onEcological Systems), Moscow: Atomizdat, 1972.
Ul’yanova, E.V., Pozolotina, V.N., and Sarapul’tsev, I.E.,Ecogenetic characteristics of dandelion (Taraxacumofficinale s.l.) populations from the Techa River flood�plain ecosystems, Russ. J. Ecol., 2004, vol. 35, no. 5,pp. 308–315.
Volkova, P.Yu. and Geras’kin, S.A., Analysis of superoxidedismutase polymorphism in chronically irradiatedScots pine populations, Radiats. Biol. Radioekol., 2012,vol. 52, no. 4, pp. 370–380.
Whitham, T.G., Bailey, J.K., Schweitzer, J.A., et al., Aframework for community and ecosystem genetics:from genes to ecosystems, Nature Rev. Genet., 2006,vol. 7, pp. 510–523.
Wooley, S.C. and Wimp, G.M., Community and ecosystemgenetics: a framework for integrating from genes to eco�systems, Nature Rev. Genet., 2006, vol. 7, pp. 510–523.
Zhivotovskii, L.A., Integratsiya poligennykh sistem v popu�lyatsiyakh (Integration of Polygenic Systems in Popula�tions), Moscow: Nauka, 1984.
Zhivotovskii, L.A., Populyatsionnaya biometriya (Popula�tion Biometrics), Moscow: Nauka.
Translated by K. Lazarev