enteric neural crest-derived cells: origin, identification, migration, and differentiation
TRANSCRIPT

Enteric Neural Crest-Derived Cells:Origin, Identification, Migration, and
DifferentiationH.M. YOUNG*1 AND D. NEWGREEN2
1Department of Anatomy and Cell Biology, University of Melbourne,3010, VIC, Australia
2The Murdoch Institute, Parkville, 3052, VIC, Australia
Neurons and glial cells forming the enteric nervoussystem (ENS) arise from neural crest cells that migrateaway from two different rostrocaudal levels of the neuralaxis, the vagal and sacral regions. The vagal region isdefined as the post-otic hindbrain level with somites 1–7(Le Douarin, 1982), and the sacral region is caudal tosomite 28 in chick embryos and caudal to somite 24 inembryonic mice.
CONTRIBUTION OF VAGAL LEVEL NEURALCREST CELLS TO THE ENS
Vagal level neural crest cells give rise to enteric neu-rons and glial cells throughout the entire gastrointesti-nal tract. Yntema and Hammond (1954) removed theneural crest and dorsal portion of the neural tube fromdefined rostrocaudal levels of the neural axis of chickembryos, and then approximately a week later, the em-bryos were fixed, sectioned, and silver stained to deter-mine if enteric ganglia were present in the gastrointes-tinal tract. They found that enteric ganglia were absentfrom the esophagus, stomach, and small and large in-testines if the lesions removed neural crest cells from aregion commencing 4 somites width rostral to the firstsomite and extended caudally to the predicted level ofthe 10th somite. ENS deficits were not observed whenany other levels of the neural crest were ablated (Yn-tema and Hammond, 1954, 1955; Hammond and Yn-tema, 1947). Thus, vagal level neural crest cells werededuced to be the only, or at least the major, source ofthe ENS throughout the entire gastrointestinal tract.These results were confirmed by Le Douarin and Teillet(1973, 1974) and Burns and Le Douarin (1998), whofound that following replacement of the vagal neuralaxis, between somites 1–7, of chick embryos with theequivalent region from quail embryos, quail neuronsand glial cells were found in ganglia throughout thegastrointestinal tract of the host embryo. More exactstudies have shown that most neural cells in the mid-and hindgut of chick embryos are derived from thelevels of somites 3– 6 (Peters-van der Sanden et al.,1993; Epstein et al., 1994). There may be some speciesdifferences, since in mice neural crest cells adjacent tosomites 1– 4 appear to give rise to enteric neuronsthroughout the gut, whereas neural crest adjacent tosomites 6 –7 gives rise to a sub-population of neurons inthe foregut, but no other gut region (Durbec et al., 1996;
note that Durbec and colleagues define neural crestadjacent to somites 6 –7 as “trunk” level neural crestsince it also gives rise to dorsal root ganglia).
CONTRIBUTION OF SACRAL LEVELNEURAL CREST CELLS TO THE ENTERIC
NERVOUS SYSTEMSince Yntema and Hammond (1954) observed no neu-
rons within the gut following ablation of the vagal-levelneural crest, it appeared that the vagal neural crest wasthe sole source of enteric neurons. However, using chick-quail grafts, Le Douarin and Teillet (1973) showed thatthe sacral-level neural crest contributes some cells to en-teric ganglia of the hindgut, although they did not ascer-tain with molecular markers if the sacral-derived cellswere neurons and/or glial cells. In the following 25 years,different studies reached different conclusions as to thecontribution of the sacral-level neural crest to the entericnervous system. In both chick and mouse embryos inwhich pre-migratory sacral level neural crest cells werelabelled with DiI or retroviruses, labelled cells were re-ported in the hindgut (Pomeranz et al., 1991; Serbedzija etal., 1991), at least 2 days prior to the arrival of vagalneural crest cells. However, variable results were ob-tained when segments of hindgut were removed prior tothe arrival of vagal level cells and grown in culture, orexplanted to the chorioallantoic membrane or kidney cap-sule; most studies reported a complete, or almost com-plete, absence of enteric neurons in the hindgut (Smith etal., 1977; Allan and Newgreen, 1980; Nishijima et al.,1990; Kapur et al., 1992; Lecoin et al., 1996; Young et al.,1996, 1998a), whereas other studies reported that neuronswere present (Rothman and Gershon, 1982; Pomeranz andGershon, 1990). Following transection of the midgut ofchick embryos prior to the arrival of vagal neural crestcells, no enteric neurons were observed in the hindgut
Contract grant sponsor: National Health and Medical ResearchCouncil of Australia (NHMRC); Contract grant sponsor: MurdochInstitute.
*Correspondence: Dr. H.M. Young, Department of Anatomyand Cell Biology, University of Melbourne, 3010, VIC, Australia.E-mail: [email protected]
Received 28 February 2000; Accepted 6 September 2000
THE ANATOMICAL RECORD 262:1–15 (2001)
© 2001 WILEY-LISS, INC.

(Meijers et al., 1989). Thus, most of these experiments inwhich the vagal source was absent suggested that sacralneural crest cells do not colonize the hindgut, or that theyrequire the presence of vagal cells in order to colonize thehindgut and form a recognizable ENS, or that they arrivesimultaneous to or later than the vagal cells (Allan andNewgreen, 1980).
The contribution of sacral level neural crest cells to theENS in birds was resolved by a detailed study utilizingchick-quail chimeric grafting by Burns and Le Douarin(1998), in which they showed definitively that sacral cellscontribute to the ENS in the hindgut. Importantly, theyexamined the time of arrival of sacral cells into the hind-gut, and the phenotypes of the derivatives of sacral neuralcrest cells. Sacral cells were observed to migrate ventrallyby E4.5 to form the pelvic plexus and nerve of Remak. Thelatter is an extension of the pelvic plexus in the form of aganglionated chain, which lies in the mesentery adjacentto the dorsal border of the post-umbilical intestine; it isunique to birds. However, the sacral neural crest cells didnot immediately continue their migration further ven-trally from the nerve of Remak to enter the hindgut;instead, they entered the hindgut approximately 3 dayslater, around the time that the vagal cells arrived in thehindgut, at E7.5. Sacral cells were found to give rise toneurons and glial cells. Cells derived from the sacral neu-ral crest gave rise to 17% of enteric neurons in the distalhindgut, with their contribution decreasing rostrally to0.3% in the rostral hindgut.
Several important questions remain unresolved aboutthe contribution of sacral neural crest cells to the ENS ofthe hindgut. The first question is why the sacral cellspause for approximately 3 days in the nerve of Remakprior to entering the hindgut. The gut has been postulatedto synthesize chemoattractive substances that attract mi-grating neural crest cells (Le Douarin and Teillet, 1974),and it is possible that the hindgut produces these sub-stances at a later stage than the fore- and midgut. Thepossibility has been raised that the presence of vagal levelneural crest cells within the hindgut may be necessary toinduce the sacral level neural crest cells to immigrate intothe hindgut (Allan and Newgreen, 1980; Gershon et al.,1993). However, a recent study has shown that followingablation of the vagal level neural tube, the migration ofthe sacral cells into the hindgut is unaffected (Burns et al.,2000), and combination grafts of aneural hindgut withsacral cells with and without vagal cells have shown thesame effect (Hearn and Newgreen, 2000). Thus, if there isa chemoattractive substance produced in the hindgut, itappears to be produced by the mesenchyme, and not vagallevel neural crest cells. Rather than an absence of che-moattractive substances to attract sacral cells into thehindgut, it is also possible that the delayed entry of sacralcells into the hindgut is due to a transient repellent effectof the hindgut. A recent study has shown that the mesen-chyme of the colo-rectum of chick embryos expresses col-lapsin-1, a secreted glycoprotein that is a member of thesemaphorin family (Shepherd and Raper, 1999). Studiesin other parts of the nervous system have established thatcollapsin-1 acts as axon repellent and neural crest guid-ance cue (Behar et al., 1996; Taniguchi et al., 1997; Eick-holt et al., 1999). Shepherd and Raper (1999) showed that,in vitro, extension of neurites from neurons in the nerve ofRemak is inhibited by explants of colo-rectum. Collapsin-1is initially (at E6) expressed throughout the mesenchyme
of the wall of the colo-rectum; at a later stage (E8), whichcoincides with the time at which the axons of the nerve ofRemak first enter the outer (muscle) layers of the hindgut,the expression of collapsin recedes to the inner submuco-sal and mucosal layers and is completely absent from themuscle layers (Shepherd and Raper, 1999). Since the sa-cral neural crest cells migrate into the hindgut alongnerve fibres from the nerve of Remak that are projectinginto the hindgut (Burns and Le Douarin, 1998), it is pos-sible that the expression of collapsin-1, either directly orindirectly, regulates the time of entry of the sacral cellsinto the hindgut.
The second unresolved question about the contributionof the sacral neural crest to the ENS in the hindgut is theidentity of labelled cells observed in the hindgut followingintroduction of DiI or a retrovirus into the sacral neuraltube prior to migration of neural crest cells (Pomeranz etal., 1991; Serbedzija et al., 1991), as these cells wereobserved within the hindgut 2–3 days prior to the arrivalof sacral neural crest cells as reported by Burns and LeDouarin (1998). It also remains to be determined whethercells derived from the sacral neural crest give rise tospecific functional classes of enteric neurons, and whethersacral cells have the potential to colonize other regions ofgut apart from the distal hindgut.
Although a contribution from the sacral neural crest tothe ENS in the hindgut has been shown conclusively inchick/quails (Burns and Le Douarin, 1998), the situationis still unclear in mammals. Mice in which the genesencoding either Ret or GFRa1 are inactivated lack entericneurons in most of the gastrointestinal tract caudal to thestomach, but some neurons are present in the distal rec-tum (Durbec et al., 1996; Cacalano et al., 1998). The mostparsimonious explanation for the existence of neurons inthe distal rectum, but not in the small intestine or colon,is that they arise from the sacral-level neural crest. How-ever, as in birds, it appears that if sacral level neural crestcells do give rise to enteric neurons in the hindgut, they donot enter the hindgut until after it has been fully colonizedby vagal level neural crest cells. A range of markers ofneural crest cells have been used to examine the coloniza-tion of the embryonic mouse gut (Kapur et al., 1992;Young et al., 1998a, 1999). These studies have shown asingle, unidirectional, rostral-to-caudal colonization of thegut by neural crest cells; labelled cells are not observed inthe hindgut prior to the arrival of the cells, which arededuced to be vagal neural crest cells (Fig. 1). Further-more, if the hindgut is removed prior to the arrival ofvagal crest cells and explanted or cultured, no neuronsdevelop in the vast majority of explants (Nishijima et al.,1990; Kapur et al., 1992; Young et al., 1996, 1998a). Mam-mals do not have a nerve of Remak, but the pelvic plexusis closely apposed to the developing distal hindgut inmouse embryos (see Young et al., 1998a), and it is possiblethat sacral neural crest cells migrate into the hindgut, butonly after pausing in the pelvic plexus for 3–4 days, asthey do in the nerve of Remak in birds. This is supportedby the observation that cells with a similar phenotype tovagal enteric neural crest cells are present in the pelvicplexus adjacent to the distal hindgut of embryonic micefrom E10.5 (see below; Young et al., 1996). The wavefrontof vagal-level neural crest cells starts to colonize the ros-tral hindgut of mice at E11.5 and reaches the distal hind-gut at around E14 (Kapur et al., 1992; Young et al.,1998a). Even at late E13.5, when the wavefront of the
2 YOUNG AND NEWGREEN
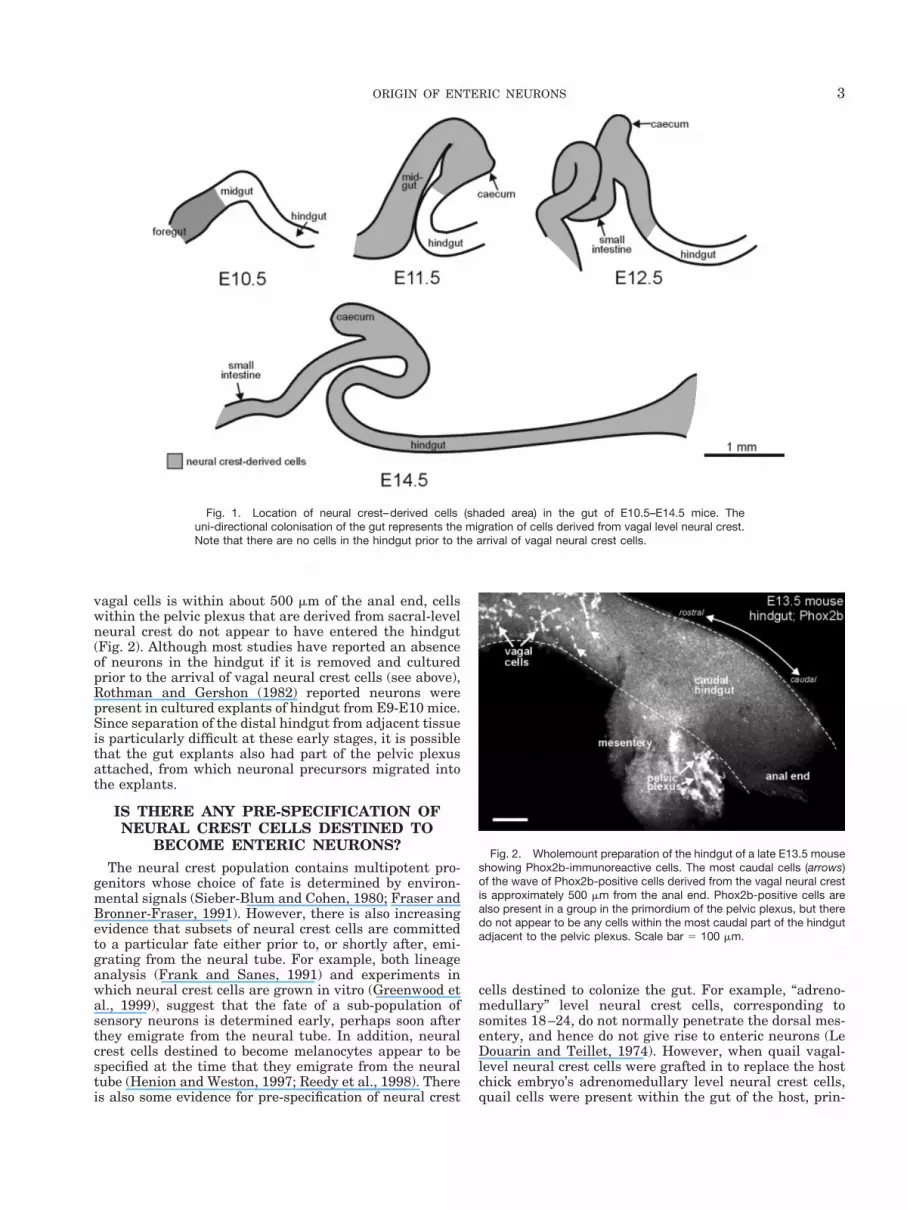
vagal cells is within about 500 mm of the anal end, cellswithin the pelvic plexus that are derived from sacral-levelneural crest do not appear to have entered the hindgut(Fig. 2). Although most studies have reported an absenceof neurons in the hindgut if it is removed and culturedprior to the arrival of vagal neural crest cells (see above),Rothman and Gershon (1982) reported neurons werepresent in cultured explants of hindgut from E9-E10 mice.Since separation of the distal hindgut from adjacent tissueis particularly difficult at these early stages, it is possiblethat the gut explants also had part of the pelvic plexusattached, from which neuronal precursors migrated intothe explants.
IS THERE ANY PRE-SPECIFICATION OFNEURAL CREST CELLS DESTINED TO
BECOME ENTERIC NEURONS?The neural crest population contains multipotent pro-
genitors whose choice of fate is determined by environ-mental signals (Sieber-Blum and Cohen, 1980; Fraser andBronner-Fraser, 1991). However, there is also increasingevidence that subsets of neural crest cells are committedto a particular fate either prior to, or shortly after, emi-grating from the neural tube. For example, both lineageanalysis (Frank and Sanes, 1991) and experiments inwhich neural crest cells are grown in vitro (Greenwood etal., 1999), suggest that the fate of a sub-population ofsensory neurons is determined early, perhaps soon afterthey emigrate from the neural tube. In addition, neuralcrest cells destined to become melanocytes appear to bespecified at the time that they emigrate from the neuraltube (Henion and Weston, 1997; Reedy et al., 1998). Thereis also some evidence for pre-specification of neural crest
cells destined to colonize the gut. For example, “adreno-medullary” level neural crest cells, corresponding tosomites 18–24, do not normally penetrate the dorsal mes-entery, and hence do not give rise to enteric neurons (LeDouarin and Teillet, 1974). However, when quail vagal-level neural crest cells were grafted in to replace the hostchick embryo’s adrenomedullary level neural crest cells,quail cells were present within the gut of the host, prin-
Fig. 2. Wholemount preparation of the hindgut of a late E13.5 mouseshowing Phox2b-immunoreactive cells. The most caudal cells (arrows)of the wave of Phox2b-positive cells derived from the vagal neural crestis approximately 500 mm from the anal end. Phox2b-positive cells arealso present in a group in the primordium of the pelvic plexus, but theredo not appear to be any cells within the most caudal part of the hindgutadjacent to the pelvic plexus. Scale bar 5 100 mm.
Fig. 1. Location of neural crest–derived cells (shaded area) in the gut of E10.5–E14.5 mice. Theuni-directional colonisation of the gut represents the migration of cells derived from vagal level neural crest.Note that there are no cells in the hindgut prior to the arrival of vagal neural crest cells.
3ORIGIN OF ENTERIC NEURONS

cipally in the post-umbilical small intestine (Le Douarinand Teillet, 1974; Smith et al., 1977). Le Douarin and hercolleagues suggested that vagal neural crest cells are pre-specified to form the ENS, and follow chemotactic cues toreach the gut. Endothelin (ET) (B) receptor (see Yanagi-sawa et al., 1998), Ret (Roberston and Mason, 1995), andCCK-LacZ (Lay et al., 1999) are expressed by pre-migra-tory cells in the neural tube. Unlike ET(B) receptor, whichis expressed by all neural crest lineages including pigmentcells, Ret appears to be expressed only by crest-derivedneural precursors including sensory, sympathetic and en-teric ganglia, and CCK-LacZ is expressed only by sympa-thetic and enteric ganglia. It is, therefore, possible thatthe Ret-positive and the CCK-LacZ-positive cells in theneural tube represent cells pre-specified to restricted lin-eages.
PHENOTYPE OF NEURAL CREST CELLSDESTINED TO FORM THE ENS
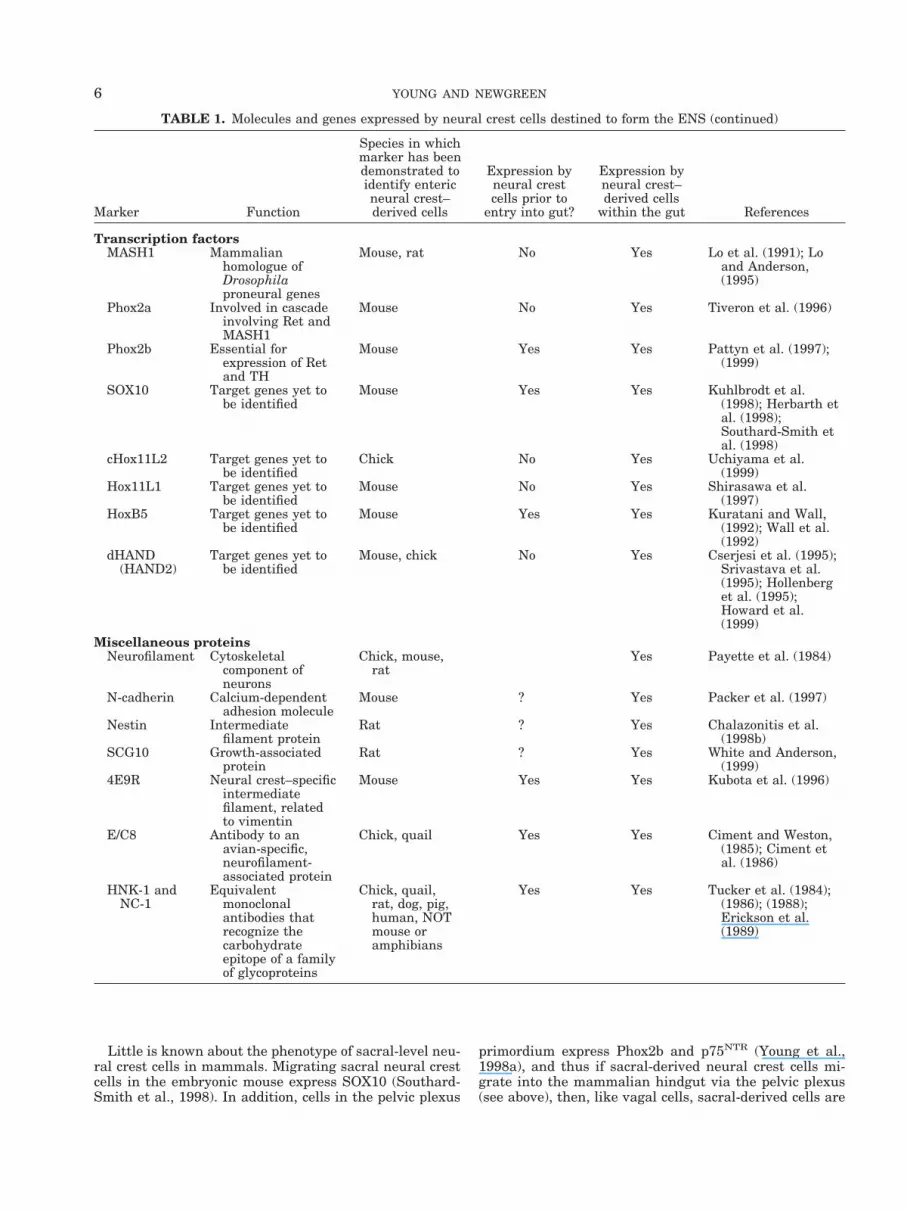
While they are migrating, both from the neural tube tothe gut, and then through the gut mesenchyme, neuralcrest cells are morphologically indistinguishable from themesenchymal cells through which they migrate. Hence,experiments examining the origin and migration of neuralcrest cells initially relied on observing the effect of abla-tions, or the use of chick-quail chimeras (see above). How-ever, a rapidly expanding number of markers has beendiscovered that identify undifferentiated neural crest cellsprior to their differentiation into enteric neurons and glialcells. These markers are shown in Table 1. Some of themarkers are expressed by neural crest cells prior to theirentry into the gut mesenchyme, whereas other markersare not detectable until after the cells have arrived withinthe gut mesenchyme.
Phenotype of Enteric Neuron Precursors Priorto Entering the Gut
Chick and quail. One of the first markers of undif-ferentiated neural crest-derived cells in the chick embryowas the E/C8 antibody, which recognizes an avian-spe-cific, 73-kD neurofilament-associated protein (Ciment andWeston, 1985; Ciment et al., 1986). Around the same timethat the E/C8 antibody was first utilized to label neuralcrest cells, the equivalent monoclonal antibodies, HNK-1and NC-1, were discovered to recognise a carbohydrateepitope on the cell surface of neural crest cells (Tucker etal., 1984). HNK-1 and NC-1 stain virtually the same pop-ulation of cells that is stained with E/C8 (Tucker et al.,1986). Since then, HNK-1 and NC-1 have been frequentlyused to identify neural crest-derived cells within the em-bryonic gut, primarily of chick and quail embryos (Figs.3A,4; Tucker et al., 1986; Pomeranz and Gershon, 1990;Epstein et al., 1991; Newgreen et al., 1996, 1997). HNK-1and NC-1 also bind to neural crest-derived cells in a rangeof species including rat (Fig. 5A), but not mice or amphib-ians (Tucker et al., 1988; Erickson et al., 1989; Newgreenand Hartley, 1995). However, the HNK-1 epitope is alsofound on a number of cells not derived from the neuralcrest, and also on extracellular matrix molecules (Fig. 3A;Newgreen et al., 1990), and so results must be interpretedwith caution. E/C8, HNK-1, and NC-1-positive cells areobserved within the somites adjacent to the vagal levelneural tube, and thus the antigens to which the antibodiesbind appear to be first expressed shortly after the neuralcrest cells leave the neural tube (Tucker et al., 1986).
ET(B) receptor is expressed by the precursors of mela-nocytes, the adrenal medulla and virtually all componentsof the peripheral nervous system, including dorsal rootganglia and autonomic (including enteric) ganglia. Neuralcrest cells at all levels of the neural axis of chick and quailembryos express ET(B) receptor both before, and during,their emigration from the neural tube (Nataf et al., 1996).Ret is also expressed in the neural tube prior to the emi-gration of neural crest cells, but is restricted to vagal levelneural tube, between rhombomeres 6 and somite 7 (Rob-ertson and Mason, 1995). Following migration, Ret is ex-pressed by some cranial ganglia, dorsal root ganglia, sym-pathetic ganglia, the ENS, and some cranial ganglia of thechick (Robertson and Mason, 1995).
The only detailed study that examined the migration ofsacral-level neural crest cells into the hindgut used quail-chick chimeric grafting to detect sacral-derived neural crestcells (Burns and Le Douarin, 1998, see above), rather thanany of the markers shown in Table 1. Hence, little is knownabout the phenotype of undifferentiated sacral-derived neu-ral crest cells in the chick, either prior to, or following, theirentry into the hindgut. Although Ret is expressed in thevagal neural tube of the chick prior to emigration of theneural crest cells, Ret is not expressed by the sacral levelneural tube (Robertson and Mason, 1995). Sacral cells des-tined to populate the chick hindgut appear to pause in thenerve of Remak for about 3 days prior to entering the hind-gut (Burns and Le Douarin, 1998; see above). HNK-1 stain-ing, and Ret- and GFRa1-immunoreactivity is shown bymany cells in the nerve of Remak (Tucker et al., 1986; Ep-stein et al., 1991; Schiltz et al., 1999), but it is unknown ifany of these cells emigrate into the hindgut.
Mouse and rat. ET(B) receptor, tyrosine hydroxylase(TH), p75NTR, Phox2b, Ret, MASH1, CCK-lacZ, andSOX10 are all expressed by vagal neural crest cells priorto their entry into the gut mesenchyme (for references seeTable 1). ET(B) receptor (see Yanagisawa et al., 1998) andCCK-lacZ (Lay et al., 1999) are expressed in the neuraltube before the cells commence migration, but the othermolecules are not detectable until after the cells have leftthe neural tube. Although Ret is expressed by pre-migra-tory crest cells of chick embryos (see above), it is firstdetected in migratory phase neural crest cells in miceembryos (Pachnis et al., 1993). Some molecules or genes,such as TH (Cochard et al., 1978; Teitelman et al., 1979),Phox2b (Pattyn et al., 1997, 1999), and CCK-lacZ (Lay etal., 1999), are expressed by enteric and sympathetic neu-ron precursors, but not dorsal root ganglion precursors,whereas other molecules such as ET(B) receptor(Southard-Smith et al., 1998), Ret (Pachnis et al., 1993;Tsuzuki et al., 1995; Nosrat et al., 1997), p75NTR (Britschet al., 1998), ErbB3 receptor (Britsch et al., 1998), andSOX10 (Herbarth et al., 1998; Southard-Smith et al.,1998), are expressed by the precursors of enteric, sympa-thetic, and sensory neurons. It is unknown whether allneural crest cells destined to enter the gut express all ofthe molecules, or whether different phenotypic classes ofcells exist, based on the expression of different markers.However, after entering the mouse gut, undifferentiatedneural crest cells are of only two phenotypes (Young et al.,1999; see phenotype of neural crest-derived cells withinthe gut wall below). It, therefore, seems likely that prior toentering the gut, the neural crest cells will show a similar,limited range of phenotypes.
4 YOUNG AND NEWGREEN

TABLE 1. Molecules and genes expressed by neural crest cells destined to form the ENS
Marker Function
Species in whichmarker has beendemonstrated toidentify entericneural crest–derived cells
Expression byneural crestcells prior to
entry into gut?
Expression byneural crest–derived cells
within the gut References
TransgenesDBH-lacZ Expression of lacZ is
driven by thepromoter of thecatecholaminesynthetic enzyme,DBH
Mouse No Yes Kapur et al. (1992)
CCK-lacZ Expression of lacZ isdriven by thepromoter of theregulatorypeptide, CCK
Mouse Yes Yes Lay et al. (1999)
P0-Cre Expression of lacZoccurs is cellsexpressing theglycoprotein, P0, acell adhesionmolecule
Mouse Yes Yes Yamauchi et al.(1999)
Receptors for neurotrophic factorsRet Receptor tyrosine
kinase; forms partof the receptorcomplex for GDNF
Mouse, chick,rat
Yes Yes Pachnis et al. (1993);Robertson andMason, (1995);Watanabe et al.(1997); Schiltz et al.(1999)
GFRa1 Along with Ret,forms receptor forGDNF
Mouse, rat,chick
? Yes Cacalano et al. (1998);Enomoto et al.(1998); Naveilhan etal. (1998); Schiltz etal. (1999)
GFRa2 Along with Ret,forms receptor forneurturin
Mouse Naveilhan et al.(1998)
ErbB3 Receptor tyrosinekinase; receptorfor neuregulin-1
Mouse Yes Yes Britsch et al. (1998)
p75NTR Low-affinityneurotrophinreceptor
Rat, mouse Yes Yes Baetge et al. (1990)
TrkC Receptor for NT-3 Rat ? Yes Chalazonitis et al.(1998b)
Endothelin-Breceptor
(ET(B)receptor)
Receptor forendothelin 3
Mouse, human,chick, quail
Yes Yes Gariepy et al. (1998);Brand et al. (1998);Nataf et al. (1996)
CNTFRa a component ofneuropoieticcytokine receptor
Rat ? Yes Chalazonitis et al.(1998a)
gp130 andLIFRb
b components ofneuropoieticcytokine receptor
Rat ? Yes Chalazonitis et al.(1998a)
Neurotransmitter-related molecules5-HT
transporterSpecific transporter
for 5-HT(serotonin)
Rat ? Yes Hansson et al. (1999)
5-HT2Breceptor
Receptor for 5-HT(serotonin)
Mouse ? Yes Fiorica-Howells et al.(2000)
Tyrosinehydroxylase(TH)
Catecholaminesynthetic enzyme
Mouse, rat, NOTchick
Yes Yes; sub-populationonly andtransiently
Cochard et al. (1978);Teitelman et al.(1979)
5ORIGIN OF ENTERIC NEURONS
Little is known about the phenotype of sacral-level neu-ral crest cells in mammals. Migrating sacral neural crestcells in the embryonic mouse express SOX10 (Southard-Smith et al., 1998). In addition, cells in the pelvic plexus
primordium express Phox2b and p75NTR (Young et al.,1998a), and thus if sacral-derived neural crest cells mi-grate into the mammalian hindgut via the pelvic plexus(see above), then, like vagal cells, sacral-derived cells are
TABLE 1. Molecules and genes expressed by neural crest cells destined to form the ENS (continued)
Marker Function
Species in whichmarker has beendemonstrated toidentify entericneural crest–derived cells
Expression byneural crestcells prior to
entry into gut?
Expression byneural crest–derived cells
within the gut References
Transcription factorsMASH1 Mammalian
homologue ofDrosophilaproneural genes
Mouse, rat No Yes Lo et al. (1991); Loand Anderson,(1995)
Phox2a Involved in cascadeinvolving Ret andMASH1
Mouse No Yes Tiveron et al. (1996)
Phox2b Essential forexpression of Retand TH
Mouse Yes Yes Pattyn et al. (1997);(1999)
SOX10 Target genes yet tobe identified
Mouse Yes Yes Kuhlbrodt et al.(1998); Herbarth etal. (1998);Southard-Smith etal. (1998)
cHox11L2 Target genes yet tobe identified
Chick No Yes Uchiyama et al.(1999)
Hox11L1 Target genes yet tobe identified
Mouse No Yes Shirasawa et al.(1997)
HoxB5 Target genes yet tobe identified
Mouse Yes Yes Kuratani and Wall,(1992); Wall et al.(1992)
dHAND(HAND2)
Target genes yet tobe identified
Mouse, chick No Yes Cserjesi et al. (1995);Srivastava et al.(1995); Hollenberget al. (1995);Howard et al.(1999)
Miscellaneous proteinsNeurofilament Cytoskeletal
component ofneurons
Chick, mouse,rat
Yes Payette et al. (1984)
N-cadherin Calcium-dependentadhesion molecule
Mouse ? Yes Packer et al. (1997)
Nestin Intermediatefilament protein
Rat ? Yes Chalazonitis et al.(1998b)
SCG10 Growth-associatedprotein
Rat ? Yes White and Anderson,(1999)
4E9R Neural crest–specificintermediatefilament, relatedto vimentin
Mouse Yes Yes Kubota et al. (1996)
E/C8 Antibody to anavian-specific,neurofilament-associated protein
Chick, quail Yes Yes Ciment and Weston,(1985); Ciment etal. (1986)
HNK-1 andNC-1
Equivalentmonoclonalantibodies thatrecognize thecarbohydrateepitope of a familyof glycoproteins
Chick, quail,rat, dog, pig,human, NOTmouse oramphibians
Yes Yes Tucker et al. (1984);(1986); (1988);Erickson et al.(1989)
6 YOUNG AND NEWGREEN

also likely to express both of these molecules prior toentering the gut.
Phenotype of Neural Crest-Derived Cells Withinthe Gut Wall
Chick and quail. All of the markers expressed byneural crest-derived cells en route to the gut are alsoexpressed by the crest-derived cells within the gut wall,for varying periods of time. Vagally-derived HNK-1/NC-1-positive cells colonize the embryonic chick and quail gut ina rostral-to-caudal wave (Fig. 4; Tucker et al., 1986; Ep-stein et al., 1991; Newgreen et al., 1997; Schiltz et al.,1999), and it is assumed that all undifferentiated neuralcrest-derived cells in the chick gut are HNK-1-postive.Little is known about the overlap of expression of otherneural crest markers by the HNK-1-positive cells. How-
ever, the relationship between HNK-1 staining and theexpression of the GDNF receptors, Ret and GFRa1, byneural crest cells as they colonize the gut has recentlybeen examined (Schiltz et al., 1999). Unlike the situationin the mouse embryo (Young et al., 1999; see mouse andrat below), not all of the undifferentiated neural crest–derived cells in the embryonic chick gut appear to expressRet or GFRa1 as they are colonizing the gut, since HNK-1-positive cells at the migratory wavefront do not showRet- or GFRa1-immunoreactivity.
Although TH is expressed by vagal neural crest cellsboth prior to their entry into the gut, and for several daysafter they have entered the gut mesenchyme of embryonicmouse and rat (see below), it is not expressed by neuralcrest cells in chick embryos either prior to or after theyhave entered the gut mesenchyme (Smith et al., 1977).
Fig. 3. A: Transverse slice through the level of somites 5–6 of anE2.75 quail embryo. HNK-1-positive cells are present in the position ofthe dorsal root ganglia (DRG) and sympathetic chain (asterisk), and insome cells (arrows) that have migrated further ventrally and are enteringthe developing gut. Note that HNK-1 staining is also observed in extra-cellular matrix around the notochord. Scale bar 5 50 mm. B: Confocalmicroscope image of neural crest derived cells in the gut of an E10.5mouse after processing for Phox2b (red) and p75NTR (green) immunore-activity. Phox2b-immunoreactivity is confined to the nuclei and p75NTR
immunoreactivity is predominantly on the cell membranes. All of thePhox2b-positive nuclei are surrounded by p75NTR-positive membranes.Nucleated blood cells (asterisks) show non-specific fluorescence. Scalebar 5 10 mm. C: Transverse section of resin-embedded small intestinefrom an E15 mouse stained with toluidine blue. Phox2b-positive cells(arrows) are only present in the outer part of the intestine, just under theserosa. There are no cells at the inner side of the circular muscle, wherethe submucosal plexus will later form. Scale bar 5 10 mm. D: Whole-mount preparation of a myenteric ganglion from an adult mosaic mouse
in which 50% of the cells express the reporter gene, lacZ. The ganglionis composed of both b-gal-positive cells (asterisks) and b-gal-negativecells (arrows), and hence does not arise from a single progenitor cell.Scale bar 5 20 mm. E: Confocal microscope image of a wholemountpreparation of the hindgut from an E14.5 mouse. A sub-population(arrows) of the Phox2b cells (stained red) has differentiated into NOSneurons (stained green), but most Phox2b-positive cells (asterisks) donot express NOS. Scale bar 5 10 mm. F: Wholemount preparation of gutfrom an E12.5 mouse stained with an antibody to p75NTR (red) and thenucleic acid stain, SYTO 16. The two p75NTR-positive neural crest cellsin this field of view (arrows) are both undergoing mitosis, as is evidentfrom their chromosomal arrangement. The SYTO 16-stained cells thatare not neural crest–derived cells are mesenchymal cells. Scale bar 5 10mm. G: High-magnification image of a neural crest–derived cell under-going mitosis (telophase) that was immunostained for p75NTR (green)and Phox2b (red). At the late stages of mitosis, Phox2b immunostainingis associated with the chromosomes. Scale bar 5 5 mm.
7ORIGIN OF ENTERIC NEURONS

However, when neural crest cells are immunoselectedfrom dissociated segments of embryonic chick gut usingHNK-1, and then grown in culture, some cells do expressTH (Pomeranz et al., 1993; Sextier-Sainte-Claire Devilleet al., 1994). Sacral neural crest-derived cells differentiateinto neurons (including nitric oxide synthase-containingneurons) and glial cells in the hindgut (Burns and Le
Douarin, 1998). However, to our knowledge, nothing isknown about the phenotype of sacral-derived neural crestcells within the chick hindgut prior to their differentiationinto neurons and glial cells.
Mouse and rat. A large number of molecules areexpressed by vagal neural crest–derived cells in the em-bryonic mouse and rat gut (see Table 1; Fig. 5B,C). Thecolonization of the gut by neural crest–derived cells inmice was first examined in detail in transgenic mice inwhich the promotor of the gene encoding the catechol-amine synthetic enzyme, dopamine-b-hydroxylase (DbH),was used to direct expression of the reporter gene, lacZ(Kapur et al., 1992). DbH-lacZ appears to be expressed bymany, if not all, vagally-derived neural crest cells withinthe gut, and the expression of the transgene by entericneurons persists in adult mice, even though there are nocatecholaminergic neurons in the adult ENS. AlthoughDbH-lacZ-expressing cells almost certainly include thecells that transiently express TH (see below), DbH-lacZ isexpressed by many more cells, and for a far longer periodof time, than TH. By breeding DbH-lacZ mice with micecarrying mutations that result in ENS defects, the role ofparticular genes in the migration of neural crest–derivedcells into the gut has been examined (see Kapur et al.,1993, 1996).
Vagal crest cells colonise the mouse and rat gut in arostral-to-caudal wave (Fig. 1; Kapur et al., 1992;Newgreen and Hartley, 1995; Young et al., 1998a), andthus during the colonization, the most caudal cells areundifferentiated, whereas cells rostral to the wavefrontare at various stages of differentiation. The overlap in theexpression of some of these molecules was examined re-cently in the embryonic mouse gut (Young et al., 1999).Between E10.5 and E13.5, undifferentiated (the most cau-dal) neural crest cells were all found to co-express Ret,p75NTR and Phox2b (Fig. 3B), but none was Phox2a-posi-tive. TH is also expressed by neural crest–derived cellswithin the early embryonic rat and mouse gut (Cochard etal., 1978; Teitelman et al., 1979). At E10.5, TH is ex-pressed by about 20% of the Ret/p75NTR/Phox2b cells atthe wavefront (Young et al., 1999). In the E10.5 mousegut, although we have never observed the most caudal cellto be TH-positive, we have observed preparations wherethe second and third most caudal cells are TH-positive(Fig. 6A,A’). However, at subsequent stages, TH-positivecells become progressively further (more rostral) from thewavefront, and, in fact, TH cells are never observed in thehindgut. It is unclear whether the absence of TH cells fromthe hindgut is due to the environment of the hindgut, or tothe poor migratory ability of TH cells. Since TH cells showmany features of neurons, including the expression ofneuron-specific proteins such as peripherin, neurofila-ment, and PGP9.5 (Fig. 6B,B’,C,C’; Baetge et al., 1990), itis possible that TH cells fail to reach the hindgut becausethey are early differentiated neurons and hence have poormigratory abilities. Although all undifferentiated neuralcrest–derived cells within the gut appear to express Ret,p75NTR and Phox2b, it remains to be determined for mostof the other markers shown in Table 1, whether they areexpressed by all or a sub-population of undifferentiatedneural crest cells, or whether they are expressed by dif-ferentiating neurons or glial cells.
The DNA binding protein, HoxB5, is expressed by en-teric neural crest–derived cells in the stomach and small
Fig. 4. HNK-1 labeled whole-mount of E5 (HH27-) quail mid andhindgut. Vagal neural crest–derived HNK-1-labelled enteric neural cellsextend through the pre-umbilical (Pre-U) and post-umbilical (Po-U) in-testine, to the rostral border of the caecum (arrow), but no HNK-1-stained cells are seen distally at this stage, for example in the colo-rectum. Sacral neural crest–derived HNK-1-labelled cells form the nerveof Remak in the dorsal mesentery directly adjacent to the intestine butdo not at this stage extend into the intestine at any level. Scale bar 5200 mm.
Fig. 5. A: High-magnification image of an HNK-1-positive cell in themidgut of an E16 rat. Scale bar 5 5 mm. B: p75NTR-positive cells in themidgut of an E11 mouse. Scale bar 5 20 mm. C: Phox2b-positive nucleiin the esophagus of an E13.5 mouse. Scale bar 5 50 mm.
8 YOUNG AND NEWGREEN

and large intestines of embryonic mice, but not the esoph-agus (Wall et al., 1992). Since sacral-level neural crestcells do not appear to express HoxB5 (Kuratani and Wall,1992) and since neurons in the esophagus arise from neu-ral crest cells adjacent to somites 6–7, whereas neurons inthe rest of the gastrointestinal tract arise from neuralcrest cells adjacent to somites 1–4 (Durbec et al., 1996; seeabove), HoxB5 may be expressed exclusively by the mostanterior vagal level neural crest cells.
MOLECULES NECESSARY FOR THEMIGRATION, SURVIVAL, PROLIFERATION,
AND DIFFERENTIATION OF ENTERICNEURON PRECURSORS
The role of particular signaling pathways and transcrip-tion factors in the development of the ENS has been re-viewed recently (Gershon, 1999; Taraviras and Pachnis,1999), and, thus, this topic will only be covered brieflyhere. Inactivation of the genes encoding a number of dif-ferent transcription factors, SOX10, Phox2b, Hox11L1,and MASH1, and the components of three signaling path-ways result in defects in the ENS (see Fig. 7). The signal-ing pathways currently known to be essential for ENSdevelopment are ET-3, its receptor ET(B) receptor and itsactivator ECE-1, GDNF, and its co-receptors GFRa1 andRet, and neurturin and its co-receptors GFRa2 and Ret.The most dramatic of knockout mice in terms of ENSphenotype are mice in which the genes encoding the tran-scription factors Phox2b or SOX10 are inactivated.Phox2b-/- and SOX10 -/- mice have no ENS in any regionof the gastrointestinal tract (Pattyn et al., 1999; Herbarthet al., 1998; Southard-Smith et al., 1998; Kapur, 1999).Thus, Phox2b and SOX10 are essential for the develop-ment of all enteric neurons and glial cells. However, inother transgenic mice in which the ENS is affected, thedefect is regional. Mash1-/- mice lack enteric neurons inthe esophagus (Guillemot et al., 1993), Gdnf-/-, Gfra1-/-,
and Ret-/- mice lack enteric neurons caudal to the stom-ach (Schuchardt et al., 1994; Moore et al., 1996; Pichel etal., 1996; Sanchez et al., 1996; Cacalano et al., 1998;Enomoto et al., 1998), and Et-3-/-, ET(B) receptor-/- andEce-1-/- mice lack enteric neurons in the colon and rectum(Baynash et al., 1994; Hosoda et al., 1994; Yanagisawa etal., 1998) (Fig. 7). Finally, in Neurturin-/- and Gfra2-/-mice, specific sub-populations of neurons are absent fromthe intestine (Heuckeroth et al., 1999; Rossi et al., 1999),whereas in Hox11L1-/- mice, there are increased numbersof enteric neurons in the colon (Hatano et al., 1997; Shira-sawa et al., 1997).
The mechanisms of action of some of these gene prod-ucts have recently been studied. The GDNF-GFRa1/Retsignalling pathway appears to be essential for the prolif-eration and differentiation of enteric neuron precursors(Chalazonitis et al., 1998b; Hearn et al., 1998; Heuckerothet al., 1998; Natarajan et al., 1999; Taraviras et al., 1999;Wu et al., 1999), whereas the main role of the ET-3-ET(B)receptor signalling pathway is to inhibit differentiation, sothat a sufficient pool of undifferentiated precursors existsto proliferate and populate the entire gastrointestinaltract (Hearn et al., 1998; Wu et al., 1999).
SOX10 is essential for the survival of migratory neuralcrest cells, since in Sox10 -/- mice, crest-derived cellsundergo apoptosis before they reach the gut (Southard-Smith et al., 1998; Kapur, 1999). However, the genesregulated by SOX10 are unknown. Phox2b is essential forthe expression of Ret and TH, since in Phox2b-/- mice,there is no Ret or TH expression in enteric or sympatheticneural crest-derived cells (Pattyn et al., 1999). The rela-tionship between Phox2b and MASH1 expression is morecomplex. In E10.5 Phox2b-/- mice, MASH1 is expressedby sympathetic precursors, but not by cells in the mostrostral sympathetic ganglia or in the gut, but by E11.5,MASH1 expression is also lost from the more caudal sym-pathetic ganglia (Pattyn et al., 1999). Since the expression
Fig. 6. Paired micrographs of wholemount preparations of the E10.5mouse gut. The TH-positive cells (A’–C’) are also positive for Phox2b (A),PGP9.5 (B), and neurofilament (C). The TH-positive cells form only asub-population of the Phox2b cells. Although the most caudal Phox2b-
positive cell (arrow in A) is not TH-positive, the second most caudalPhox2b cell (asterisk) is TH-positive. Scale bars 5 10 mm (A,B),20 mm (C).
9ORIGIN OF ENTERIC NEURONS

of Phox2a is dramatically reduced in Mash1-/- mice (Hir-sch et al., 1998; Lo et al., 1998), and as there is a reductionin the expression of Ret in some Phox2a-/- mice (Morin etal., 1997), it has been proposed that Phox2a is also in-volved in a signalling cascade involving MASH1 and Ret(Lo et al., 1998). However, Phox2a-/- mice do not have anydetectable defects in their ENS (Morin et al., 1997).MASH1 appears to be essential for promoting the differ-entiation of committed neural precursors (Sommer et al.,1995).
With the exception of Phox2b-/- and SOX10 -/- mice(see above), the effect of inactivation of particular genesoften has differential effects on the superior cervical gan-glion and the ENS caudal to the stomach on the one hand,and the rest of the sympathetic ganglia and the ENS inthe esophagus on the other. For example, Ret knockoutmice lack the superior cervical ganglion and an ENS cau-dal to the stomach, but the ENS is present in the esoph-agus (see Fig. 7) and more caudal sympathetic ganglia arealso present. However, a complementary phenotype re-sults when Mash1 is inactivated. There is no ENS in theesophagus, but enteric neurons are present in the stomachand the small and large intestines; there are also dramat-ically reduced numbers of neurons in the sympatheticganglia (Guillemot et al., 1993). Enteric neuron precursorsin all regions of the gastrointestinal tract, including theesophagus, and in all sympathetic ganglia, appear to ex-press Ret, GFRa1, and MASH1 (Pachnis et al., 1993; Lo etal., 1994; Young et al., 1999), and thus the regional spec-ificity seen in the knockouts is not due to different regionalexpression patterns of the molecules. As suggested byDurbec et al. (1996), the phenotype of the ENS and thesympathetic nervous system of Ret-/-, Gfra1-/-, Gdnf-/-,and Mash1-/- mice appears to be correlated with therostrocaudal level of the neural axis from which the dif-ferent structures originate. Derivatives of the neural crestadjacent to somites 1–4 give rise to the ENS caudal to thestomach and to the superior cervical ganglion, and these
lineages are dependent on the GDNF-GFRa1/Ret signal-ing pathway, whereas the ENS in the esophagus arisesfrom neural crest adjacent to somites 6–7 and is depen-dent on MASH1. The different phenotypes of Mash1 -/-and Ret -/- mice also suggest that Ret and MASH1 musthave additional interactions, in addition to the signalingcascade involving MASH1, Phox2, and Ret proposed by Loet al. (1998).
FORMATION OF GANGLIA ANDDIFFERENTIATION OF ENTERIC NEURONSGershon et al. (1993) estimated that there are more
neurons in the gut than in the spinal cord. In the smalland large intestine of the adult mouse, there are 400–900neurons/mm2 of gut wall; these neurons, plus an unknownnumber of glial cells, develop from an initial population ofless than 1,000 neural crest–derived cells at E10.5 (HMY,personal observation). Thus, massive proliferation mustoccur in the developing ENS. Dividing neural crest–de-rived cells within the gut can be detected using nucleicacid stains combined with neural crest cell-specific mark-ers, or by the pattern of immunoreactivity for the tran-scription factor, Phox2b (Fig. 3F,G). At E10.5–E14.5, atany one time point, 1–2% of the neural crest–derived cellsare undergoing mitosis. Crest cells undergoing mitosisappear to be distributed randomly along the gut, and thusthere does not seem to be any region that is a proliferativezone for neural crest cells.
In the midgut of chick and mice embryos, neural crest–derived cells migrate through the outer part of the mes-enchyme, just under the serosa, where the myentericplexus will form (Fig. 3C; Kapur et al., 1992; Newgreen etal., 1996; Young et al., 1996; Burns and Le Douarin, 1998).At this stage, the midgut mesenchyme is undifferentiated,and circular and longitudinal muscle layers have notformed (Allan and Newgreen, 1980; Torihashi et al.,1999). Neural crest–derived cells are not seen in the loca-tion of the submucosal plexus until 5–6 days later, and the
Fig. 7. Presence of neurons in the gastrointestinal tract of some knockout mice. Gdnf, glial derivedneurotrophic factor; Et-3, endothelin 3; Et(B) receptor, endothelin (B) receptor; Ece-1, endothelin convertingenzyme-1. Note the presence of enteric neurons in the distal hindgut (asterisk) of Ret -/- and GFRa1 -/- mice.
10 YOUNG AND NEWGREEN

submucosal plexus is thought to arise from a secondarymigration from the myenteric plexus (Gershon et al., 1980;Payette et al., 1987). In the hindgut of embryonic mice, thelocation of the migratory pathway of neural crest cellswithin the hindgut mesenchyme, and the development ofthe submucosal and myenteric plexuses, have not beeninvestigated. However, in the colo-rectum of chick em-bryos, vagal neural crest cells follow a migratory routealong the inner (mucosal) side of the mesenchyme, wherethe submucosal plexus will form, and some cells then latermigrate to the outer side of the circular muscle, to theregion of the myenteric plexus (Burns and Le Douarin,1998). It is likely that crest-derived cells in the colo-rec-tum of the chick embryo migrate along a route closer to themucosa than they do in the small intestine because, by thetime that the cells arrive in the large intestine, the circu-lar muscle layer has differentiated.
For several days after neural crest–derived cells havecolonized a particular region of the embryonic mouse gut,the cells are evenly distributed, with no indication of cellclustering (Fig. 8A), except in the caecum. Later, as thegut increases in length and diameter, the cells form gan-glionic groupings (Fig. 8B,C). The factors inducing cells tocluster into ganglia are unknown. Cells forming a gan-glion do not arise from a single progenitor cell. This hasbeen demonstrated by Rothman et al. (1993) who foundthat following back-transplantation of quail gut contain-ing post-migratory crest cells into chick embryos contain-ing migratory-stage neural crest–derived cells, many en-teric ganglia formed within the host (chick) embryo thatcontained both quail and chick cells. A multiple progenitororigin of neurons in a single ganglion can also be demon-strated using an in situ cell lineage marker in female miceheterozygous for an X-linked transgene. This line of trans-genic mice has multiple copies of lacZ inserted into the Xchromosome (Tam and Tan, 1992). Following random Xinactivation in heterozygous female offspring aroundE9.5, b-galactosidase is expressed in about 50% of cells(Tan and Breen, 1993). As most gastric glands are usuallyderived from a single precursor stem cell, the cells ofmature individual gastric glands are either predomi-nantly b-gal positive or b-gal negative in these mosaicmice (Nomura et al., 1998). However, enteric ganglia arealmost always composed of a mixture of b-gal positive andb-gal negative cells (Fig. 3D), indicating that a gangliondoes not represent the offspring of a single progenitor.Moreover, neurons of a particular type within a ganglionare also not clonal, since within a single ganglion, neuronsof a particular type often include both b-gal positive andb-gal negative cells.
The mature ENS is composed of many different func-tional types of neurons that differ in the combinations ofneurotransmitters that they contain and their projectionpatterns (Costa et al., 1996; Sang and Young, 1996; Sanget al., 1997). Little is known about the factors influencingan undifferentiated neural crest–derived cell to differen-tiate into a glial cell or a particular type of neuron. Theneurturin-GFRa2/Ret signalling pathway appears to playsome role in the development of cholinergic neurons, notonly in the gut, but also in parasympathetic ganglia, sinceneurturin -/- and GFRa2 -/- mice have reduced numbersof cholinergic neurons in cranial and enteric ganglia(Heuckeroth et al., 1999; Rossi et al., 1999). MASH1 seemsto be essential for the development of some types of neu-rons since Mash1 -/- mice lack 5-HT neurons in the small
intestine (Blaugrund et al., 1996). Different types of en-teric neurons develop at different developmental stages.In embryonic mice, nitric oxide synthase-containing neu-
Fig. 8. Formation of ganglia in the small intestine of the mouse, inwhich all enteric neurons and their precursors are labelled with antibod-ies to Phox2b (A, B) and differentiated neurons with antibodies toPGP9.5 (C). At E12.5 (A), Phox2b-positive cells are densely, but relativelyevenly distributed; at E16.5 (B) the cells have clustered into recognizableganglia (arrows), which resemble adult ganglia (C) in terms of the numberof cells per ganglion. Scale bar 5 50 mm (A), 100 mm (B, C).
11ORIGIN OF ENTERIC NEURONS

rons develop early, at E12.5 in the mid-gut andE14.5 inthe hindgut (Fig. 3E), whereas cholinergic neurons cannotbe detected until around birth (Branchek and Gershon,1989; Sang and Young, 1997; Young et al., 1998b), and itis likely that different types of enteric neurons are gener-ated in response to signals that change over time. As inother parts of the nervous system (Bredt and Snyder,1994; Ward et al., 1994), nitric oxide synthase is tran-siently expressed by some enteric neurons during devel-opment (Young and Ciampoli, 1998).
The gut environment seems to influence the phenotypeof neurons derived from the neural crest. For example, thegut environment appears to inhibit the expression of TH.Over 55% of post-migratory neural crest derived cells im-munoselected from E14.5 rat gut express TH when cul-tured for 4 days compared to 16% of freshly isolated cells(Pisano and Birren, 1999). Moreover, following exposureto the gut environment, neural crest cells appear to beable to generate only a limited range of neuronal and glialphenotypes. When post-migratory enteric neural crest–derived cells from either rat or quail embryos are back-transplanted into E3-E4 chick embryos, some grafted cellscolonize sensory, sympathetic, and parasympathetic gan-glia and the nerve of Remak if they are grafted into theappropriate axial level (Rothman et al., 1993; White andAnderson, 1999). Although the gut-derived cells that con-tribute to the pelvic ganglia or nerve of Remak exhibit aphenotype that is typical of neurons in these locations, thegut-derived cells that contribute to sensory or sympatheticganglia do not express an appropriate sensory or sympa-thetic phenotype respectively (White and Anderson,1999). Hence, the gut environment is likely to play a vitalrole in the generation of different neuronal phenotypes.Interestingly, crest-derived cells able to provide sensoryand autonomic neurons and peripheral glia appear tomake a lineage decision to the parasympathetic/entericfate before the decision to be neurons or glial cells (Whiteand Anderson, 1999).
CONCLUSIONSIn the past 5 years, rapid advances in our understand-
ing of ENS development have been made largely because(1) gene knockout studies have identified transcriptionfactors and signaling pathways that are essential for thedevelopment of the ENS in different regions of the gut,and (2) many markers have been identified that recognizeneural crest–derived cells within the gut prior to theirdifferentiation into neurons and glial cells, and thus thecolonization of the gut by neural crest–derived cells inwild-type and mutant animals can be investigated. How-ever, there are still many unanswered questions, particu-larly regarding the generation of neuronal diversitywithin the gut.
ACKNOWLEDGMENTSH.M.Y. and D.F.N. are supported by the National
Health and Medical Research Council of Australia(NHMRC) and the Murdoch Institute. We thank AlisonCanty and Kate Atherton for assistance with obtainingsome of the confocal microscope images, preparing thefigures, and compiling references. We also thank Jean-Francois Brunet for the Phox2b antiserum, and SeongSeng Tan for providing tissue from the mosaic mice.
LITERATURE CITEDAllan IJ, Newgreen DF. 1980. The origin and differentiation of enteric
neurons of the intestine of the fowl embryo. Am J Anat 157:137–154.
Baetge G, Pintar JE, Gershon MD. 1990. Transiently catecholamin-ergic (TC) cells in the bowel of the fetal rat: precursors of noncat-echolaminergic enteric neurons. Dev Biol 141:353–380.
Baynash AG, Hosoda K, Giaid A, Richardson JA, Emoto N, HammerRE, Yanagisawa M. 1994. Interaction of endothelin-3 with endothe-lin-B receptor is essential for development of epidermal melano-cytes and enteric neurons. Cell 79:1277–1285.
Behar O, Golden JA, Mashimo H, Schoen FJ, Fishman MC. 1996.Semaphorin III is needed for normal patterning and growth ofnerves, bones and heart. Nature 383:525–528.
Blaugrund E, Pham TD, Tennyson VM, Lo L, Sommer L, AndersonDJ, Gershon MD. 1996. Distinct subpopulations of enteric neuronalprogenitors defined by time of development, sympathoadrenal lin-eage markers and Mash-1-dependence. Development 122:309–320.
Branchek TA, Gershon MD. 1989. Time course of expression of neu-ropeptide Y, calcitonin gene-related peptide, and NADPH diapho-rase activity in neurons of the developing murine bowel and theappearance of 5-hydroxytryptamine in mucosal enterochromaffincells. J Comp Neurol 285:262–273.
Brand M, Le Moullec JM, Corvol P, Gasc JM. 1998. Ontogeny ofendothelins-1 and -3, their receptors, and endothelin convertingenzyme-1 in the early human embryo. J Clin Invest 101:549–559.
Bredt DS, Snyder SH. 1994. Transient nitric oxide synthase neuronsin embryonic cerebral cortical plate, sensory ganglia, and olfactoryepithelium. Neuron 13:301–313.
Britsch S, Li L, Kirchhoff S, Theuring F, Brinkmann V, Birchmeier C,Riethmacher D. 1998. The ErbB2 and ErbB3 receptors and theirligand, neuregulin-1, are essential for development of the sympa-thetic nervous system. Genes Dev 12:1825–1836.
Burns AJ, Le Douarin NM. 1998. The sacral neural crest contributesneurons and glia to the post- umbilical gut: spatiotemporal analysisof the development of the enteric nervous system. Development125:4335–4347.
Burns AJ, Champeval D, Le Douarin NM. 2000. Scaral neural crestcells colonise aganglionic hindgut in vivo but fail to compensate forlack of enteric ganglia. Dev Biol 219:30–43.
Cacalano G, Farinas I, Wang LC, Hagler K, Forgie A, Moore M,Armanini M, Phillips H, Ryan AM, Reichardt LF, Hynes M, DaviesA, Rosenthal A. 1998. GFRalpha1 is an essential receptor compo-nent for GDNF in the developing nervous system and kidney. Neu-ron 21:53–62.
Chalazonitis A, Rothman TP, Chen J, Vinson EN, MacLennan AJ,Gershon MD. 1998a. Promotion of the development of enteric neu-rons and glia by neuropoietic cytokines: interactions with neurotro-phin-3. Dev Biol 198:343–365.
Chalazonitis A, Rothman TP, Chen J, Gershon MD. 1998b. Age-dependent differences in the effects of GDNF and NT-3 on thedevelopment of neurons and glia from neural crest–derived precur-sors immunoselected from the fetal rat gut: expression of GFRal-pha-1 in vitro and in vivo. Dev Biol 204:385–406.
Ciment G, Weston JA. 1985. Segregation of developmental abilities inneural-crest-derived cells: identification of partially restricted in-termediate cell types in the branchial arches of avian embryos. DevBiol 111:73–83.
Ciment G, Ressler A, Letourneau PC, Weston JA. 1986. A novelintermediate filament-associated protein, NAPA-73, that binds todifferent filament types at different stages of nervous system devel-opment. J Cell Biol 102:246–251.
Cochard P, Goldstein M, Black IB. 1978. Ontogenetic appearance anddisappearance of tyrosine hydroxylase and catecholamines in therat embryo. Proc Natl Acad Sci USA 75:2986–2990.
Costa M, Brookes SJ, Steele PA, Gibbins I, Burcher E, Kandiah CJ.1996. Neurochemical classification of myenteric neurons in theguinea-pig ileum. Neuroscience 75:949–967.
Cserjesi P, Brown D, Lyons G, Olson E. 1995. Expression of the novelbasic helix-loop-helix gene eHAND in neural crest derivatives and
12 YOUNG AND NEWGREEN

extraembryonic membranes during mouse development. Dev Biol170:664–678.
Durbec PL, Larsson-Blomberg LB, Schuchardt A, Costantini F, Pach-nis V. 1996. Common origin and developmental dependence on c-retof subsets of enteric and sympathetic neuroblasts. Development122:349–358.
Eickholt BJ, Mackenzie SL, Graham A, Walsh FS, Doherty P. 1999.Evidence for collapsin-1 functioning in the control of neural crestmigration in both trunk and hindbrain regions. Development 126:2181–2189.
Enomoto H, Araki T, Jackman A, Heuckeroth RO, Snider WD, John-son EM, Jr., Milbrandt J. 1998. GFR alpha1-deficient mice havedeficits in the enteric nervous system and kidneys. Neuron 21:317–324.
Epstein ML, Poulsen KT, Thiboldeaux R. 1991. Formation of gangliain the gut of the chick embryo. J Comp Neurol 307:189–199.
Epstein ML, Mikawa T, Brown AM, McFarlin DR. 1994. Mapping theorigin of the avian enteric nervous system with a retroviral marker.Dev Dyn 201:236–244.
Erickson CA, Loring JF, Lester SM. 1989. Migratory pathways ofHNK-1-immunoreactive neural crest cells in the rat embryo. DevBiol 134:112–118.
Fiorica-Howells E, Maroteaux L, Gershon MD. 2000. Serotonin andthe 5-HT2B receptor in the development of enteric neurons. J Neu-rosci 20:294–305.
Frank E, Sanes JR. 1991. Lineage of neurons and glia in chick dorsalroot ganglia: analysis in vivo with a recombinant retrovirus. Devel-opment 111:895–908.
Fraser SE, Bronner-Fraser M. 1991. Migrating neural crest cells inthe trunk of the avian embryo are multipotent. Development 112:913–920.
Gariepy CE, Williams SC, Richardson JA, Hammer RE, YanagisawaM. 1998. Transgenic expression of the endothelin-B receptor pre-vents congenital intestinal aganglionosis in a rat model of Hir-schprung disease. J Clin Invest 102:1092–1101.
Gershon MD. 1999. II. Disorders of enteric neuronal development:insights from transgenic mice. Am J Physiol 277:G262–267.
Gershon MD, Epstein ML, Hegstrand L. 1980. Colonization of thechick gut by progenitors of enteric serotonergic neurons: distribu-tion, differentiation, and maturation within the gut. Dev Biol 77:41–51.
Gershon MD, Chalazonitis A, Rothman TP. 1993. From neural crest tobowel: development of the enteric nervous system. J Neurobiol24:199–214.
Greenwood AL, Turner EE, Anderson DJ. 1999. Identification of di-viding, determined sensory neuron precursors in the mammalianneural crest. Development 126:3545–3559.
Guillemot F, Lo LC, Johnson JE, Auerbach A, Anderson DJ, JoynerAL. 1993. Mammalian achaete-scute homolog 1 is required for theearly development of olfactory and autonomic neurons. Cell 75:463–476.
Hammond WS, Yntema CL. 1947. Depletions in the thoracolumbarsympathetic system follwoing removal of neural crest in the chick.J Comp Neurol 86:237–266.
Hansson SR, Mezey E, Hoffman BJ. 1999. Serotonin transportermessenger RNA expression in neural crest–derived structures andsensory pathways of the developing rat embryo. Neuroscience 89:243–265.
Hatano M, Aoki T, Dezawa M, Yusa S, Iitsuka Y, Koseki H, TaniguchiM, Tokuhisa T. 1997. A novel pathogenesis of megacolon in Ncx/Hox11L.1 deficient mice. J Clin Invest 100:795–801.
Hearn C, Newgreen D. 2000. The sacral neural crest contribution tothe avian enteric nervous system is independent of vagal neuralcrest cells. Dev Dyn 218:525–530.
Hearn CJ, Murphy M, Newgreen D. 1998. GDNF and ET-3 differen-tially modulate the numbers of avian enteric neural crest cells andenteric neurons in vitro. Dev Biol 197:93–105.
Henion PD, Weston JA. 1997. Timing and pattern of cell fate restric-tions in the neural crest lineage. Development 124:4351–4359.
Herbarth B, Pingault V, Bondurand N, Kuhlbrodt K, Hermans-Borg-meyer I, Puliti A, Lemort N, Goossens M, Wegner M. 1998. Muta-tion of the Sry-related Sox10 gene in Dominant megacolon, a mouse
model for human Hirschsprung disease. Proc Natl Acad Sci USA95:5161–5165.
Heuckeroth RO, Lampe PA, Johnson EM, Milbrandt J. 1998. Neur-turin and GDNF promote proliferation and survival of enteric neu-ron and glial progenitors in vitro. Dev Biol 200:116–129.
Heuckeroth RO, Enomoto H, Grider JR, Golden JP, Hanke JA, Jack-man A, Molliver DC, Bardgett ME, Snider WD, Johnson EM Jr,Milbrandt J. 1999. Gene targeting reveals a critical role for neur-turin in the development and maintenance of enteric, sensory, andparasympathetic neurons [see comments]. Neuron 22:253–263.
Hirsch MR, Tiveron MC, Guillemot F, Brunet JF, Goridis C. 1998.Control of noradrenergic differentiation and Phox2a expression byMASH1 in the central and peripheral nervous system. Development125:599–608.
Hollenberg SM, Sternglanz R, Cheng PF, Weintraub H. 1995. Identi-fication of a new family of tissue-specific basic helix-loop- helixproteins with a two-hybrid system. Mol Cell Biol 15:3813–3822.
Hosoda K, Hammer RE, Richardson JA, Baynash AG, Cheung JC,Giaid A, Yanagisawa M. 1994. Targeted and natural (piebald-le-thal) mutations of endothelin-B receptor gene produce megacolonassociated with spotted coat color in mice. Cell 79:1267–1276.
Howard M, Foster DN, Cserjesi P. 1999. Expression of HAND geneproducts may Be sufficient for the differentiation of avian neuralcrest–derived cells into catecholaminergic neurons in culture. DevBiol 215:62–77.
Kapur RP. 1999. Early death of neural crest cells is responsible for thetotal enteric aganglionosis in Sox10(Dom)/Sox10(Dom) mouse em-bryos. Pediatr Dev Pathol 2:559–569.
Kapur RP, Yost C, Palmiter RD. 1992. A transgenic model for study-ing development of the enteric nervous system in normal and agan-glionic mice. Development 116:167–175.
Kapur RP, Yost C, Palmiter RD. 1993. Aggregation chimeras demon-strate that the primary defect responsible for aganglionic megaco-lon in lethal spotted mice is not neuroblast autonomous. Develop-ment 117:993–999.
Kapur RP, Livingston R, Doggett B, Sweetser DA, Siebert JR,Palmiter RD. 1996. Abnormal microenvironmental signals underlieintestinal aganglionosis in Dominant megacolon mutant mice. DevBiol 174:360–369.
Kubota Y, Morita T, Ito K. 1996. New monoclonal antibody (4E9R)identifies mouse neural crest cells. Dev Dyn 206:368–378.
Kuhlbrodt K, Herbarth B, Sock E, Hermans-Borgmeyer I, Wegner M.1998. Sox10, a novel transcriptional modulator in glial cells. J Neu-rosci 18:237–250.
Kuratani SC, Wall NA. 1992. Expression of Hox 2.1 protein in re-stricted populations of neural crest cells and pharyngeal ectoderm.Dev Dyn 195:15–28.
Lay JM, Gillespie, P. J., Samuelson, L. C. 1999. Murine prenatalexpression of cholecystokinin in neural crest, enteric neurons, andenterochromaffin cells. Dev Dyn 216:190–200.
Lecoin L, Gabella G, Le Douarin N. 1996. Origin of the c-kit-positiveinterstitial cells in the avian bowel. Development 122:725–733.
Le Douarin N. 1982. The neural crest. Cambridge: Cambridge Uni-versity Press.
Le Douarin NM, Teillet MA. 1973. The migration of neural crest cellsto the wall of the digestive tract in avian embryo. J Embryol ExpMorphol 30:31–48.
Le Douarin NM, Teillet MA. 1974. Experimental analysis of themigration and differentiation of neuroblasts of the autonomic ner-vous system and of neurectodermal mesenchymal derivatives, usinga biological cell marking technique. Dev Biol 41:162–184.
Lo L, Anderson DJ. 1995. Postmigratory neural crest cells expressingc-RET display restricted developmental and proliferative capaci-ties. Neuron 15:527–539.
Lo LC, Johnson JE, Wuenschell CW, Saito T, Anderson DJ. 1991.Mammalian achaete-scute homolog 1 is transiently expressed byspatially restricted subsets of early neuroepithelial and neural crestcells. Genes Dev 5:1524–1537.
Lo L, Guillemot F, Joyner AL, Anderson DJ. 1994. MASH-1: a markerand a mutation for mammalian neural crest development. PerspectDev Neurobiol 2:191–201.
13ORIGIN OF ENTERIC NEURONS

Lo L, Tiveron MC, Anderson DJ. 1998. MASH1 activates expression ofthe paired homeodomain transcription factor Phox2a, and couplespan-neuronal and subtype-specific components of autonomic neuro-nal identity. Development 125:609–620.
Meijers JH, Tibboel D, van der Kamp AW, van Haperen-Heuts IC,Molenaar JC. 1989. A model for aganglionosis in the chicken em-bryo. J Pediatr Surg 24:557–561.
Moore MW, Klein RD, Farinas I, Sauer H, Armanini M, Phillips H,Reichardt LF, Ryan AM, Carver-Moore K, Rosenthal A. 1996. Renaland neuronal abnormalities in mice lacking GDNF. Nature 382:76–79.
Morin X, Cremer H, Hirsch MR, Kapur RP, Goridis C, Brunet JF.1997. Defects in sensory and autonomic ganglia and absence oflocus coeruleus in mice deficient for the homeobox gene Phox2a.Neuron 18:411–423.
Nataf V, Lecoin L, Eichmann A, Le Douarin NM. 1996. Endothelin-Breceptor is expressed by neural crest cells in the avian embryo. ProcNatl Acad Sci USA 93:9645–9650.
Natarajan D, Grigoriou M, Marcos-Gutierrez CV, Atkins C, PachnisV. 1999. Multipotential progenitors of the mammalian enteric ner-vous system capable of colonising aganglionic bowel in organ cul-ture. Development 126:157–168.
Naveilhan P, Baudet C, Mikaels A, Shen L, Westphal H, Ernfors P.1998. Expression and regulation of GFRalpha3, a glial cell line-derived neurotrophic factor family receptor. Neurobiology 95:1295–1300.
Newgreen DF, Hartley L. 1995. Extracellular matrix and adhesivemolecules in the early development of the gut and its innervation innormal and spotting lethal rat embryos. Acta Anat (Basel) 154:243–260.
Newgreen DF, Powell ME, Moser B. 1990. Spatiotemporal changes inHNK-1/L2 glycoconjugates on avian embryo somite and neural crestcells. Dev Biol 139:100–120.
Newgreen DF, Southwell B, Hartley L, Allan IJ. 1996. Migration ofenteric neural crest cells in relation to growth of the gut in avianembryos. Acta Anat (Basel) 157:105–115.
Newgreen DF, Kerr RS, Minichiello J, Warren N. 1997. Changes incell adhesion and extracellular matrix molecules in spontaneousspinal neural tube defects in avian embryos. Teratology 55:195–207.
Nishijima E, Meijers JH, Tibboel D, Luider TM, Peters-van derSanden MM, van der Kamp AW, Molenaar JC. 1990. Formation andmalformation of the enteric nervous system in mice: an organ cul-ture study. J Pediatr Surg 25:627–631.
Nomura S, Esumi H, Job C, Tan SS. 1998. Lineage and clonal devel-opment of gastric glands. Dev Biol 204:124–135.
Nosrat CA, Tomac A, Hoffer BJ, Olson L. 1997. Cellular and devel-opmental patterns of expression of Ret and glial cell line-derivedneurotrophic factor receptor alpha mRNAs. Exp Brain Res 115:410–422.
Pachnis V, Mankoo B, Costantini F. 1993. Expression of the c-retproto-oncogene during mouse embryogenesis. Development 119:1005–1017.
Packer AI, Elwell VA, Parnass JD, Knudsen KA, Wolgemuth DJ.1997. N-cadherin protein distribution in normal embryos and inembryos carrying mutations in the homeobox gene Hoxa-4. Int JDev Biol 41:459–468.
Pattyn A, Morin X, Cremer H, Goridis C, Brunet JF. 1997. Expressionand interactions of the two closely related homeobox genes Phox2aand phox2b during neurogenesis. Development 124:4065–4075.
Pattyn A, Morin X, Cremer H, Goridis C, Brunet JF. 1999. Thehomeobox gene Phox2b is essential for the development of auto-nomic neural crest derivatives. Nature 399:366–370.
Payette RF, Bennett GS, Gershon MD. 1984. Neurofilament expres-sion in vagal neural crest–derived precursors of enteric neurons.Dev Biol 105:273–287.
Payette RF, Tennyson VM, Pham TD, Mawe GM, Pomeranz HD,Rothman TP, Gershon MD. 1987. Origin and morphology of nervefibers in the aganglionic colon of the lethal spotted (ls/ls) mutantmouse. J Comp Neurol 257:237–252.
Peters-van der Sanden MJ, Kirby ML, Gittenberger-de Groot A, Tib-boel D, Mulder MP, Meijers C. 1993. Ablation of various regions
within the avian vagal neural crest has differential effects on gan-glion formation in the fore-, mid- and hindgut. Dev Dyn 196:183–194.
Pichel JG, Shen L, Sheng HZ, Granholm AC, Drago J, Grinberg A, LeeEJ, Huang SP, Saarma M, Hoffer BJ, Sariola H, Westphal H. 1996.Defects in enteric innervation and kidney development in micelacking GDNF. Nature 382:73–76.
Pisano JM, Birren SJ. 1999. Restriction of developmental potentialduring divergence of the enteric and sympathetic neuronal lineages.Development 126:2855–2868.
Pomeranz HD, Gershon MD. 1990. Colonization of the avian hindgutby cells derived from the sacral neural crest. Dev Biol 137:378–394.
Pomeranz HD, Rothman TP, Gershon MD. 1991. Colonization of thepost-umbilical bowel by cells derived from the sacral neural crest:direct tracing of cell migration using an intercalating probe and areplication-deficient retrovirus. Development 111:647–655.
Pomeranz HD, Rothman TP, Chalazonitis A, Tennyson VM, GershonMD. 1993. Neural crest–derived cells isolated from the gut byimmunoselection develop neuronal and glial phenotypes when cul-tured on laminin. Dev Biol 156:341–361.
Reedy MV, Faraco CD, Erickson CA. 1998. The delayed entry ofthoracic neural crest cells into the dorsolateral path is a conse-quence of the late emigration of melanogenic neural crest cells fromthe neural tube. Dev Biol 200:234–246.
Robertson K, Mason I. 1995. Expression of ret in the chicken embryosuggests roles in regionalisation of the vagal neural tube andsomites and in development of multiple neural crest and placodallineages. Mech Dev 53:329–344.
Rossi J, Lukko K, Poteryaev D, Laurikainen A, Sun YF, Laakso T,Eerikainen S, Tuominen R, Lakso M, Rauvala H, Arumae U, Pas-ternack M, Saarma M, Airaksinen MS. 1999. Retarded growth anddeficits in the enteric and parasympathetic nervous system in micelacking GFR alpha2, a functional neurturin receptor. Neuron 22:243–252.
Rothman TP, Gershon MD. 1982. Phenotypic expression in the devel-oping murine enteric nervous system. J Neurosci 2:381–393.
Rothman TP, Le Douarin NM, Fontaine-Perus JC, Gershon MD.1993. Colonization of the bowel by neural crest–derived cells re-migrating from foregut backtransplanted to vagal or sacral regionsof host embryos. Dev Dyn 196:217–233.
Sanchez MP, Silos-Santiago I, Frisen J, He B, Lira SA, Barbacid M.1996. Renal agenesis and the absence of enteric neurons in micelacking GDNF. Nature 382:70–73.
Sang Q, Young HM. 1996. Chemical coding of neurons in the myen-teric plexus and external muscle of the small and large intestine ofthe mouse. Cell Tissue Res 284:39–53.
Sang Q, Young HM. 1997. Development of nicotinic receptor clustersand innervation accompanying the change in muscle phenotype inthe mouse esophagus. J Comp Neurol 386:119–136.
Sang Q, Williamson S, Young HM. 1997. Projections of chemicallyidentified myenteric neurons of the small and large intestine of themouse. J Anat 190:209–222.
Schiltz CA, Benjamin J, Epstein ML. 1999. Expression of the GDNFreceptors Ret and GFRalpha1 in the developing avian enteric ner-vous system. J Comp Neurol 414:193–211.
Schuchardt A, D’Agati V, Larsson-Blomberg L, Costantini F, PachnisV. 1994. Defects in the kidney and enteric nervous system of micelacking the tyrosine kinase receptor Ret [see comments]. Nature367:380–383.
Serbedzija GN, Burgan S, Fraser SE, Bronner-Fraser M. 1991. Vitaldye labelling demonstrates a sacral neural crest contribution to theenteric nervous system of chick and mouse embryos. Development111:857–866.
Sextier-Sainte-Claire Deville F, Ziller C, Le Douarin NM. 1994. De-velopmental potentials of enteric neural crest–derived cells inclonal and mass cultures. Dev Biol 163:141–151.
Shepherd IT, Raper JA. 1999. Collapsin-1/semaphorin D is a repellentfor chick ganglion of Remak axons. Dev Biol 212:42–53.
Shirasawa S, Yunker AM, Roth KA, Brown GA, Horning S, Kors-meyer SJ. 1997. Enx (Hox11L1)-deficient mice develop myentericneuronal hyperplasia and megacolon. Nat Med 3:646–650.
14 YOUNG AND NEWGREEN

Sieber-Blum M, Cohen AM. 1980. Clonal analysis of quail neural crestcells: they are pluripotent and differentiate in vitro in the absenceof noncrest cells. Dev Biol 80:96–106.
Smith J, Cochard P, Le Dourain NM. 1977. Development of cholineacetyltransferase and cholinesterase activites in enteric gangliaderived from presumptive adrenergic and cholinergic levels of theneural crest. Cell Differ 6:199–216.
Sommer L, Shah N, Rao M, Anderson DJ. 1995. The cellular functionof MASH1 in autonomic neurogenesis. Neuron 15:1245–1258.
Southard-Smith EM, Kos L, Pavan WJ. 1998. Sox10 mutation dis-rupts neural crest development in Dom Hirschsprung mouse model.Nat Genet 18:60–64.
Srivastava D, Cserjesi P, Olsen EN. 1995. A subclass of bHLHproteins required for cardiac morphogenesis. Science 270:1995–1999.
Tam PP, Tan SS. 1992. The somitogenetic potential of cells in theprimitive streak and the tail bud of the organogenesis-stage mouseembryo. Development 115:703–715.
Tan SS, Breen S. 1993. Radial mosaicism and tangential cell disper-sion both contribute to mouse neocortical development [see com-ments]. Nature 362:638–640.
Taniguchi M, Yuasa S, Fujisawa H, Naruse I, Saga S, Mishina M,Yagi T. 1997. Disruption of semaphorin III/D gene causes severeabnormality in peripheral nerve projection. Neuron 19:519–530.
Taraviras S, Pachnis V. 1999. Development of the mammalian entericnervous system. Curr Opin Genet Dev 9:321–327.
Taraviras S, Marcos-Gutierrez CV, Durbec P, Jani H, Grigoriou M,Sukumaran M, Wang L-C, Hynes M, Raisman G, Pachnis V. 1999.Signalling by the RET receptor kinase and its role in the develop-ment of the mammalian enteric nervous system. Development 126:2785–2797.
Teitelman G, Baker H, Joh TH, Reis DJ. 1979. Appearance of cate-cholamine-synthesising enzymes during development of rat sympa-thetic nervous system: possible role of tissue environment. ProcNatl Acad Sci USA 76:509–513.
Tiveron MC, Hirsch MR, Brunet JF. 1996. The expression pattern ofthe transcription factor Phox2 delineates synaptic pathways of theautonomic nervous system. J Neurosci 16:7649–7660.
Torihashi S, Nishi K, Tokutomi Y, Nishi T, Ward S, Sanders KM.1999. Blockade of kit signaling induces transdifferentiation of in-terstitial cells of cajal to a smooth muscle phenotype. Gastroenter-ology 117:140–148.
Tsuzuki T, Takahashi M, Asai N, Iwashita T, Matsuyama M, Asai J.1995. Spatial and temporal expression of the ret proto-oncogeneproduct in embryonic, infant and adult rat tissues. Oncogene 10:191–198.
Tucker GC, Aoyama H, Lipinski M, Tursz T, Thiery JP. 1984. Iden-tical reactivity of monoclonal antibodies HNK-1 and NC-1: conser-vation in vertebrates on cells derived from the neural primordiumand on some leukocytes. Cell Differ 14:223–230.
Tucker GC, Ciment G, Thiery JP. 1986. Pathways of avian neuralcrest cell migration in the developing gut. Dev Biol 116:439–450.
Tucker GC, Delarue M, Zada S, Boucaut JC, Thiery JP. 1988. Expres-sion of the HNK-1/NC-1 epitope in early vertebrate neurogenesis.Cell Tissue Res 251:457–465.
Uchiyama K, Otsuka R, Hanaoka K. 1999. cHox11L2, a Hox11 relatedgene, is expressed in the peripheral nervous system and subpopu-lation of the spinal cord during chick development. Neurosci Lett273:97–100.
Wall NA, Jones CM, Hogan BL, Wright CV. 1992. Expression andmodification of Hox 2.1 protein in mouse embryos. Mech Dev 37:111–120.
Ward SM, Shuttleworth CW, Kenyon JL. 1994. Dorsal root ganglionneurons of embryonic chicks contain nitric oxide synthase andrespond to nitric oxide. Brain Res 648:249–258.
Watanabe Y, Harada T, Ito T, Ishiguro Y, Ando H, Seo T, KobayashiS, Takahashi M, Nimura Y. 1997. ret Proto-oncogene product is auseful marker of lineage determination in the development of theenteric nervous system in rats. J Pediatr Surg 32:28–33.
White PM, Anderson DJ. 1999. In vivo transplantation of mammalinaneural crest cells into chick host reveals a new autonomic sublin-eage restrcition. Development 126:4351–4363.
Wu JJ, Chen JX, Rothman TP, Gershon MD. 1999. Inhibition of invitro enteric neuronal development by endothelin-3: mediation byendothelin B receptors [In Process Citation]. Development 126:1161–1173.
Yamauchi Y, Abe K, Mantani A, Hitoshi Y, Suzuki M, Osuzu F,Kuratani S, Yamamura K. 1999. A novel transgenic technique thatallows specific marking of the neural crest cell lineage in mice. DevBiol 212:191–203.
Yanagisawa H, Yanagisawa M, Kapur RP, Richardson JA, WilliamsSC, Clouthier DE, de Wit D, Emoto N, Hammer RE. 1998. Dualgenetic pathways of endothelin-mediated intercellular signaling re-vealed by targeted disruption of endothelin converting enzyme-1gene. Development 125:825–836.
Yntema CL, Hammond WS. 1954. The origin of intrinsic ganglia oftrunk viscera from vagal neural crest in the chick embryo. J CompNeurol 101:515 – 541.
Yntema CL, Hammond WS. 1955. Experiments on the origin anddevelopment of the sacral autonomic nerves in the chick embryo. JExp Zool 129:375–413.
Young HM, Ciampoli D. 1998. Transient expression of neuronal nitricoxide synthase by neurons of the submucous plexus of the mousesmall intestine. Cell Tissue Res 291:395–401.
Young HM, Ciampoli D, Southwell BR, Newgreen DF. 1996. Origin ofinterstitial cells of Cajal in the mouse intestine. Dev Biol 180:97–107.
Young HM, Hearn CJ, Ciampoli D, Southwell BR, Brunet JF,Newgreen DF. 1998a. A single rostrocaudal colonization of therodent intestine by enteric neuron precursors is revealed by theexpression of Phox2b, Ret, and p75 and by explants grown underthe kidney capsule or in organ culture. Dev Biol 202:67–84.
Young HM, Torihashi S, Ciampoli D, Sanders KM. 1998b. Identifica-tion of neurons that express stem cell factor in the mouse smallintestine. Gastroenterology 115:898–908.
Young HM, Ciampoli D, Hsuan J, Canty AJ. 1999. Expression of ret-,p75(NTR)-, Phox2a-, Phox2b-, and tyrosine hydroxylase-immunore-activity by undifferentiated neural crest–derived cells and differentclasses of enteric neurons in the embryonic mouse gut. Dev Dyn216:137–152.
15ORIGIN OF ENTERIC NEURONS