endothelia of term human placentae display diminished expression of tight junction proteins during...
TRANSCRIPT

Cell Tissue Res (2006) 324: 433–448DOI 10.1007/s00441-005-0135-7
REGULAR ARTICLE
Samuel Liévano . Lourdes Alarcón .Bibiana Chávez–Munguía .Lorenza González–Mariscal
Endothelia of term human placentae display diminished expressionof tight junction proteins during preeclampsia
Received: 23 May 2005 / Accepted: 22 November 2005 / Published online: 1 March 2006# Springer-Verlag 2006
Abstract This study explores the molecular compositionof the tight junction (TJ) in human term placenta fromnormal women and from patients with preeclampsia, ahypertensive disorder of pregnancy. Maternal endothelialdysfunction is a critical characteristic of preeclampsia;hence, we have analyzed its impact on placental vessels.The study concentrates on the TJ because this structureregulates the sealing of the paracellular route. We havefound that, in placental endothelial vessels, TJ componentsinclude the peripheral protein ZO–1 and the integralproteins occludin and claudins 1, 3, and 5. Duringpreeclampsia, the amounts of occludin and ZO–1 exhibitno significant variation, whereas those of claudins 1, 3, and5 diminish, suggesting the presence of leakier TJs in theendothelia of the preeclamptic placenta, possibly inresponse to the decreased perfusion of this organ duringpreeclampsia. We have unexpectedly found that, in normalplacentae, the multinucleated syncytiotrophoblast layerdisplays claudin 4 at the basal surface of the plasmamembrane, and claudin 16 along the apical and basolateralsurfaces. The presence of membrane-lined channels thatcross the syncytiotrophoblast constituting a paracellular
pathway has been determined by transmission electronmicroscopy and by the co-immunolocalization of claudin16 with the plasma membrane proteins Na+K+-ATPase andGP135. Since claudin 16 functions as a paracellularchannel for Mg2+, its diffuse pattern in preeclampticplacentae suggests the altered paracellular transport of Mg2+
between the maternal blood and the placental tissue.
Keywords Placenta . Preeclampsia . Endothelia .Tight junction . Claudins . Mg2+ . Human
Introduction
Preeclampsia is a disorder exclusive to human pregnancyand is diagnosed when hypertension and proteinuria occurafter 20 weeks of gestation. Although the cause ofpreeclamspia is still unknown, recent evidence suggeststhat underlying factors or predispositions are responsiblefor this pathological condition. The inciting organ in thesyndrome is the placenta, and the disease can arise evenwithout the presence of a fetus, as in molar pregnancies(Soto–Wright et al. 1995; Redman 1991). The placenta,which regulates the transfer of materials between themother and the developing fetus, is an organ of fetal originand thus has both maternal and paternal antigens. Althoughthe established common denominator in the genesis of thedisease is abnormal placentation, a maternal response toforeign paternal antigens has been speculated as being animmunological cause of preeclampsia (Robillard et al.1994).
The development of the human placenta depends on thedifferentiation of cytotrophoblast (CTB) cells, the epithe-lial cells of the placenta. This process takes two routes. Inone, the CTB cells remain in the fetal compartment andfuse to form a multinucleate syncytiotrophoblast thatcovers the chorionic villi. The latter are in contact withthe maternal blood in the intervillous space and perform theexchange of nutrients and gas for the fetus. In the otherroute, CTB cells aggregate into cell columns that attach tothe uterine wall, and from there invade the uterine
This work was supported by grants 45691-Q from the MexicanCouncil for Science and Technology (CONACYT) and 2005/1/I/012from the Research Promotion Fund of the Mexican Institute ofSocial Security (IMSS/FOFOI).
S. LiévanoGynecology and Obstetrics Hospital, La Raza Medical Center,Mexican Institute of Social Security (IMSS),México D.F., Mexico
L. Alarcón . L. González–Mariscal (*)Department of Physiology, Biophysics and Neuroscience,Center for Research and Advanced Studies (CINVESTAV),Ave. Politécnico Nacional 2508,México D.F., 07360, Mexicoe-mail: [email protected].: +52-55-50613966Fax: +52-55-50613754
B. Chávez–MunguíaDepartment of Experimental Pathology, CINVESTAV,México D.F., Mexico

interstitium and vasculature, anchoring the fetus to themother and establishing the blood flow to the placenta.CTB colonizing the uterine spiral arterioles replace thematernal endothelium and adopt a vascular phenotype inthe process (Zhou et al. 1997b). In preeclampsia, CTB cellsappear to be arrested in their differentiation program andhence are defective in endovascular invasion and coloni-zation (Zhou et al. 1993, 1997a). This leads to inadequatedilation of the spiral arterioles and to improper perfusion ofthe placenta and fetus (Brosens et al. 1972; Khong et al.1986).
Endothelial cell dysfunction in preeclamptic women is acrucial characteristic of the disease, resulting in vascularreactivity, activation of the coagulation cascade, and loss ofvascular integrity (Pridjian and Puschett 2002). In new-borns of preeclamptic women, the umbilical cord arteriesexhibit endothelial cell disruption, not present in similarvessels from newborns of normal parturients (Roberts1989). However, the reason for this endothelial dysfunctionremains unclear. Here, we have determined whether themolecular composition of the tight junction (TJ) in theendothelia of the placentae is altered in full-termpregnancies complicated by preeclampsia.
The TJ is a seal that regulates the passage of ions andmolecules through the paracellular space (Anderson andCereijido 2001). This structure also works as a “fence” thatblocks the movement of membrane molecules between theapical and basolateral domains of the plasma membrane(Cereijido et al. 1989), which in endothelia respectivelyface the blood and the perivascular spaces (Bazzoni andDejana 2004).
TJs in endothelia are different from those in epithelia andin brain vessels that contribute to the blood-brain barrier(Bazzoni and Dejana 2004). First, their precise location isnot so clear cut. Hence, they can be found at the uppermostportion of the lateral membrane and interspersed withadherent junctions along a broad segment of the lateralmembrane (Leach and Firth 1992; Leach et al. 2000).Second, the TJs of endothelial cells are predominantlyassociated with the E-face and not the P-face in freeze-fracture replicas (Muhleisen et al. 1989; Simionescu et al.1976). Ample variability in TJ complexity has been foundamong different portions of the vascular tree. Thus, in largevessels, TJs are well organized, whereas in post-capillaryvenules, in which extravasation of plasma proteins andleukocytes takes place, loosely organized TJs have beendetected (Bazzoni and Dejana 2004).
TJs are constituted by adaptor and integral proteins (for areview, see Gonzalez–Mariscal et al. 2003). The former areperipheral molecules that usually contain multiple protein/protein-binding domains (e.g. PDZ, SH3, or WW) thatallow them work as scaffolds linked to several TJ integraland adaptor proteins, actin, and/or actin-binding proteins,transcription factors, protein kinases, and phosphatases.Several adaptor proteins, such as ZO–1 and ZO–2, havebeen found to shuttle between the nucleus and the TJ in amanner dependent on the state of confluency of theepithelia (Gottardi et al. 1996; Islas et al. 2002), and recent
data suggest their participation in the regulation of cellproliferation and differentiation (Balda et al. 2003).
Integral proteins occludin and claudins constitute thebackbone of TJ filaments observed by freeze-fracture.Occludin over-expression increases the paracellular resis-tance of epithelial monolayers, and mutated occludinaugments epithelia leakiness (Balda et al. 1996; McCarthyet al. 1996). However, the role of occludin at TJs is difficultto understand, since occludin knockout mice exhibitnormal TJs (Saitou et al. 2000). Claudins constitute afamily of more than 20 different proteins that display atissue-specific distribution (Tsukita and Furuse 1999). Atthe TJ, claudins function as paracellular channels (Tsukitaand Furuse 2000). The ionic selectivity of each claudindepends on the amount and type of charged amino acidspresent on its first extracellular loop. Therefore, the set ofclaudins present in a given epithelium determines itsparticular paracellular ionic selectivity. For example,claudin 16, which functions as a Mg2+ channel, is largelyrestricted to the thick ascending loop of Henle (TALH;Simon et al. 1999), from which 60% of the filtered Mg2+ isreabsorbed (Quamme and de Rouffignac 2000).
The molecular composition of TJs in placental vesselshas begun to be uncovered. Full-term human placentaedisplay occludin and claudin 1 immunoreactivity in largevessels but not in microvessels. Instead, ZO–1 is foundalong the whole vascular tree (Leach et al. 2000, 2002).Here, we have analyzed the expression of ZO–1, occludin,and claudins 1, 3, 4, 5, 7, 15, and 16 in placentae derivedfrom normal and preeclamptic women at term pregnancies.
Materials and methods
Tissue selection
This study was performed on placentae from patients whohad preeclampsia and who delivered singleton live births atthe 38th week of gestation. As controls, we employedplacentae from full-term uncomplicated pregnancies. Allthe placentae were obtained after Cesarean section andwere collected from the La Raza Medical Center, MexicanInstitute of Social Security (IMSS), Mexico City. Placentaewere obtained with informed patient consent and approvalby the local ethical committee.
Preeclampsia was diagnosed as the presence of bloodpressure above 140/90 mm Hg or increases of 30 mm Hgsystolic or 15 mm Hg diastolic over baseline values, withproteinuria being rated with more than one “+” on urineanalysis, and edema (see Table 1).
Tissue processing
Placentae were separated from the decidual tissue im-mediately after delivery, rinsed in ice-cold phosphate-buffered saline (PBS), maintained in ice-cold buffer, andthen transferred to the laboratory where biopsies (1 cm3)were taken. For electron microscopy, the tissue samples
434

were fixed with 2.5% glutaraldehyde in 0.1 M cacodylatebuffer, pH 7.2, for 60 min at room temperature. Thesamples were post-fixed with 1% osmium tetroxide in thesame buffer, containing 5 μg/ml ruthenium red. Sampleswere dehydrated, embedded in Epon, and sectionedfollowing standard procedures. Thin sections were viewedon a Philips Morgagni 268D transmission electronmicroscope.
Placental samples destined for immunofluorescencewere immersed for 2 min in 2-methylbutane, which hadpreviously been cooled in liquid nitrogen. The cubes werethen transferred for 10 min to liquid nitrogen. Sections(6 μm) were cut with a Leica cryostat (Leica CM 1900–3–1). Some sections were processed for hematoxylin-eosinstaining following standard procedures, whereas the restwere processed for immunofluorescence.
Because human placentae contain large amounts offibrous material, we designed a protocol that increased theefficiency of protein extraction for Western blot analysis,viz., by reducing the tissue to powder. This protocol wasbased on methods previously described by others for thelysis of tissue samples (Sambrook and Russell 2001) andby us for the Western blot determination of claudins inrenal tubules (Reyes et al. 2002). The placental biopsieswere washed six times with ice-cold PBS containing 1 mMphenylmethane sulfonyl-fluoride (PMSF), immersed inliquid nitrogen, and ground with a pre-chilled mortar andpestle. The powdered tissue was next treated with the lysisbuffer (RIPA: 40 mM TRIS-HCl pH 7.6, 150 mM NaCl,2 mM EDTA, 10% glycerol, 1% Triton X–100, 0.5%sodium deoxycholate, 0.2% SDS, and 1 mM PMSF)containing the protease inhibitor cocktail Complete. Thelysate was then sonicated (3×1 min each) in a high intensityultrasonic processor (Vibra cell, Sonics and Materials,Danbury, Conn.) and spun for 15 min at 4°C in a SorvallBiofuge Fresco centrifuge at 13,000 rpm. The resultingsupernatant was employed for Western blotting. For thegeneration of the Triton X–100 soluble and insolublefractions, the powdered tissue of the placentae was treatedwith a gentler buffer lacking SDS (20 mM TRIS-HCl pH7.6, 150 mM NaCl, 2 mM EDTA, 1% Triton X–100, 5 mMsodium fluoride, 1 mM Na orthovanadate, and 2 mMPMSF) containing the protease inhibitor cocktail Com-plete. After incubation for 30 min at 4°C under continuousagitation, the sample was sonicated and centrifuged asdescribed above. The resulting supernatant was designated
as the Triton X–100 soluble fraction. The pellet wasresuspended in RIPA buffer, sonicated three times, andcentrifuged as mentioned above. This supernatant wasdesignated as the Triton X–100 insoluble fraction, and thepellet was discarded.
Immunofluorescence
For immunofluorescence, sections were processed aspreviously described (Gonzalez–Mariscal et al. 2000),with the modification of being fixed for 5 min with ice-cold acetone followed by a 5-min wash with ice-cold PBSand a 10-min incubation with 0.25% Triton X–100. Thesectionswere treatedwith rabbit polyclonal antibodies againstZO–1 (Zymed 61–7300, San Francisco, Calif., USA; dilution1:100), claudin 1 (Zymed 51–9000; dilution 1:25), claudin 3(Zymed 34–1700; dilution 1:13), claudin 5 (Zymed 34–1600;dilution 1:125), claudin 7 (Zymed 34–9100, dilution 1:32),claudin 16 (Zymed 34–5400; dilution 1:85), occludin (Zymed71–1500; dilution 1:100), and the COOH-terminal peptideKETYY (specific for all Na+K+ATPase isoforms; Bayer1990; dilution 1:50; kindly provided to Dr. Liora Shoshani,CINVESTAV, México, by Dr. Nikolai Modyanov, MedicalCollege of Ohio). We also employed mouse monoclonalantibodies against claudin 4 (Zymed 32–9400; dilution 1:167),claudin 15 (Zymed 32–9800: dilution 1:25), glycoproteinGP135 (a generous gift of Dr. George Ojakian, New YorkState University), PECAM–1 (R&D Systems BBA7; dilution1:1,000), and the β subunit of Na+K+ATPase (a generous giftof Dr. Marcelino Cereijido, CINVESTAV, Mexico). Assecondary antibodies we employed fluorescein-isothiocya-nate-conjugated goat anti-rabbit IgG (Zymed 62–6111; dilu-tion 1:100) and goat anti-mouse IgG (Zymed 62–6511;dilution 1:100), a tetramethylrhodamine-isothiocyanate-con-jugated goat anti-mouse IgG (Zymed 62–6514; dilution 1:50),and a CY5-conjugated goat anti-mouse IgG (Zymed 81–6516;dilution 1:50).
Table 1 Patient characteristics
Characteristic Normal pregnancyn=10
Preeclampsian=10
Maternal age (years) 27.3±5.85 29±7.73Gestational age (weeks) 38.1±1 37.5±0.7Newborn weight (grams) 2,970±320 2,440±370Newborn height (cm) 49±2.36 46.3±2.63Systolic pressure (mm Hg) 110±8.8 149±9.95*Diastolic pressure (mm Hg) 69±7.4 106±5.1*Proteinuria Negative ++ to +++
*P<0.001
*
*
*
*
*
s
s
s
s
m
m
m
m
is
is
Fig. 1 Histomorphological view of chorionic villi in a term placentaderived form a normal woman. The placental tissue was stained withhematoxylin and eosin (abbreviations in this and all other figures,other than Fig. 12: asterisks blood vessels, s syncytiotrophoblastlayer, m mesenchyme, is intervillous space)
435

Before mounting, some sections were incubated for 15min with the DNA dye, propidium iodide (0.7 μg/ml; cat.no. 4170, Sigma) and subsequently washed three timeswith PBS. Fluorescence was examined by using a confocalmicroscope (Leica SP2) with argon and helium-neon lasersand by employing Leica confocal software. All images arerepresentative micrographs of four independent experi-ments with placentae derived from different patients.
Western blotting
Proteins were quantified in the total placental extracts andin the Triton X–100 soluble and insoluble fractions, and the
samples were diluted (1:1) in a standard sample buffer, runin 12% polyacrylamide gels, and transferred to polyvinyl-idene fluoride membranes. Blots were obtained aspreviously described (Jaramillo et al. 2004) by using theantibodies against the TJ proteins described above, amonoclonal against actin (a generous gift of Dr. JoséManuel Hernández, CINVESTAV, México), peroxidase-conjugated goat antibodies against rabbit (Zymed 81–6120, dilution 1:2,000) or mouse (Zymed 62–6520,dilution 1:2,000) and a chemiluminiscence detectionsystem (ECL+Plus, Amersham Pharmacia Biotechnology).
ZO 1NORMAL PREECLAMPSIA
B
A
25 µm
* *
*
m
25 µm
*
*
m
N P
ZO 1
ACTIN
220
45
0
1
2
O.
D.
in a
rbit
rary
un
its
Normal Preeclampsia
0.8
0 +
0.0
8
0.6
5 +
0.2
2
Fig. 2 ZO–1 is conspicuouslyfound in the endothelial cells ofthe placenta from both normaland preeclamptic patients.a Immunofluorescence images.Placentae were incubated withspecific antibodies against ZO–1. b Western blot analysis. LeftRepresentative gel with sam-ples from various normal (N)and preeclamptic (P) patients.Right Graph displaying densi-tometric analysis of all thesamples. Each point on thegraph corresponds to an indi-vidual patient (means±SE)
Table 2 Expression of TJ proteins in term placentae derived from normal (N) and preeclamptic (P) women (S Triton X-100 soluble fraction,I Triton X-100 insoluble fraction, SN Triton X-100 soluble fraction from normal placentae, SP Triton X-100 soluble fraction frompreeclamptic placentae, IN Triton X-100 insoluble fraction from normal placentae, IP Triton X-100 insoluble fraction from preeclampticplacentae)
Protein Immunofluorescence Western blot
Vessels Syncytiotrophoblast membrane Total Triton X-100 fraction
Large Small Lateral Basal N vs P N P SN vs SP IN vs IP
N P N P S vs I S vs I
ZO-1 + + + + =Occludin + + – – > > = =Claudin 1 + + – – > > = >Claudin 3 + + – – > > > =Claudin 4 – – – – – + > > = =Claudin 5 + + + + > > = >Claudin 16 – – – – + + > > = =
436

O. D
. in
arb
itra
ry u
nit
s
N P N P N P N P
S65 S83 I 65 I 83
0
1
2
0.9
5 +
0.1
3
1.2
2 +
0.2
2
0.9
0 +
0.2
1
--
-
0.5
6 +
0.2
1
0.2
8 +
0.1
5
1.0
9 +
0.1
5
0.3
6 +
0.2
2
0.3
6 +
0.2
2
-
-
-
-
-
B
A
PR
EE
CL
AM
PS
IAN
OR
MA
L
*
OCCLUDIN PECAM-1 MERGE
25 µm
25 µm
s
m
is
**
is
is
s
m* s
*
OCCL
ACTIN
65
45
83
N P
S I S I
25 µm
25 µm25 µm
25 µm
S I S I
Fig. 3 Placentae derived fromnormal and preeclamptic womendisplay a similar pattern ofoccludin immunoreactivity.a Immunofluorescence images.Placentae were treated withspecific antibodies against bothoccludin and PECAM–1 (ar-rows small diameter vesselspositive for PECAM–1 but oc-cludin-negative). b Western blotanalysis. Left Representative gelwith samples from various nor-mal (N) and preeclamptic (P)patients (S Triton X–100 solublefraction, I Triton X–100 insolu-ble fraction). Two occludinbands of 65 and 83 kDa weredetected in the S and I fractions.Right Graph displaying densito-metric analysis of all the sam-ples. Each point on the graphcorresponds to an individualpatient (means±SE)
Normal Preeclampsia
O. D
. in
arb
itra
ry u
nit
s
0.7
4 +
0.1
4
0.3
6 +
0.0
9
1.2
3 +
0.1
2
0.8
4 +
0.2
1
S I S I
0
1
2
3*
*
B
S I S I
N P
CL 5
ACTIN
22
45
NO
RM
AL
CLAUDIN 5 PECAM-1 MERGE
PR
EE
CL
AM
PS
IA
A
25 µm
25 µm
25 µm
25 µm25 µm
25 µm
*
*
*
s
m
is
sm
is
s
is
S I S I
Fig. 4 Claudin 5 is present inthe endothelia of normal andpreeclamptic placentae, but inthe latter, the amount of thisclaudin in the Triton X–100insoluble fraction is diminished.a Immunofluorescence imagesdisplay a conspicuous palisadepattern of immunoreactivity forclaudin 5 at the lumen of thevessels of the placenta fromnormal and preeclampticwomen. However, in both con-ditions, several vessels identifiedwith PECAM–1 remain non-reactive to the claudin 5 anti-body (arrows). b Western blotanalysis. Left Representative gelwith samples from various nor-mal (N) and preeclamptic (P)patients. Note the smalleramount of claudin 5 in theTriton X–100 insoluble fraction(I) than in the soluble (S) frac-tion, and the significant decreasein the amount of claudin 5 in theinsoluble fraction of preeclamp-tic placentae. Right Graph dis-playing densitometric analysis ofall the samples. Each point onthe graph corresponds to anindividual patient (means±SE).*P≤0.05; Student′s t-test
437

Results
The human placenta is constituted by mesenchyme and twomajor tissue layers, viz., the syncytiotrophoblast, whosemicrovillous surface is bathed by maternal blood foundthroughout the intervillous space, and the fetal endotheli-um. At term, the endothelia of the terminal villi,responsible for nutrient exchange, are closely applied tothe basal surface of the syncytiotrophoblast, thus formingareas of presumed least resistance for transplacentaldiffusion.
In order to facilitate the interpretation of the subsequentimmunofluorescent images, we present a histomorpholog-ical view of chorionic villi in a term placenta derived froma normal woman (Fig. 1). Numerous blood vessels and thesyncytiotrophoblast layer surrounded by the intervillousspace can be seen in the chorionic villi.
ZO–1
In placentae from both normal and preeclamptic women,conspicuous ZO–1 immunoreactivity was detected invessels of all widths (Fig. 2). A similar amount of ZO–1was detected by Western blot in normal placentae and inthose derived from patients with preeclampsia. Theseresults and those of the following figures are summarizedin Table 2.
Occludin
In normal women, in agreement with a previous report(Leach et al. 2000), anti-occludin gives conspicuousimmunostaining at the endothelial cells of the chorionicvilli, whereas the small diameter vessels are immunonega-
A
B
NO
RM
AL
CLAUDIN 3 PECAM-1 MERGE
PR
EE
CL
AM
PS
IA
25 µm 25 µm 25 µm
25 µm 25 µm 25 µm
NORMAL PREECLAMPSIA
*
*
*
*
25 µm 25 µm
*
is
is
is
*
*
m
m
m
s
s
ssC
LA
UD
IN 3
*m
is s
*
m
is
s
*
m
*
m
*
*
C
Fig. 5 Claudin 3 lines the largeplacental vessels of normal andpreeclamptic women, althoughthe amount of claudin 3 presentin the Triton–X 100 solublefraction is reduced in preeclamp-tic patients. a, b By immunofluo-rescence, claudin 3 displays apalisade pattern lining the lumenof large placental vessels,whereas in terminal capillariesidentified by PECAM–1 immu-noreactivity, it is barely visiblein both normal and preeclampticplacentae. c Western blot analy-sis shows a slight decrease in theamount of soluble claudin 3 inpreeclamptic placentae. Left Rep-resentative gel with samplesfrom various normal (N) andpreeclamptic (P) patients. RightGraph displaying densitometricanalysis of all the samples. Eachpoint on the graph corre-sponds to an individual patient(means±SE). *P≤0.05;Student′s t-test)
438

tive. The latter can be identified by their positive PECAM–1 immunoreactivity (Fig. 3a, second column, arrows), sincethis cell-cell adhesion molecule is known to be found in allendothelial cells of the vascular tree (Bazzoni and Dejana2004; Leach et al. 2000; Newman 1997). In thepreeclamptic placentae, the wide vessels are immunoreac-tive to occludin (Fig. 3a).
Western blot analysis revealed two clear occludin bandsof 65 and 83 kDa both in Triton X–100 soluble andinsoluble fractions. The densitometric analysis showedthat, in normal and preeclamptic placentae, a larger amountof occludin was present in the soluble fraction. Nodifference in the amount of occludin was detected betweennormal and preeclamptic placentae (Fig. 3b).
Claudin 5
Next, we studied the amount and distribution pattern of thedifferent claudins. We started by exploring claudin 5, sincethis protein, although present in certain epithelia (Rahner etal. 2001), has been found to be specially concentrated inendothelial cells (Morita et al. 1999). In normal placentae,the expression of claudin 5 is strong in several vessels,whereas it is absent from others even of a similar width(Fig. 4 arrow). An analogous situation is encountered inplacentae from preeclamptic women (Fig. 4a).
By Western blot, we observed the presence of claudin 5in both Triton X–100 soluble and insoluble fractions,although the amount in the latter fraction was significantlylower (Fig. 4b). Furthermore, the amount of claudin 5
Fig. 6 In placentae derived fromboth normal and preeclampticwomen, claudin 1 has a strongimmunoreactivity in large ves-sels, but a barely detectable signalis found in terminal villous cap-illaries. aClaudin 1 lines the largeblood vessels in the chorionicvilli of both normal and pre-eclamptic placentae. b In smalldiameter vessels detected by theirpositive PECAM–1 immunore-activity, claudin 1 gives a lowsignal. c Western blot analysisshows a smaller amount of clau-din 1 in the insoluble fractionsthan in the soluble fractions, and adecreased amount of claudin 1 inthe insoluble fraction of pre-eclamptic placentae. Left Repre-sentative gel with samples fromvarious normal (N) and pre-eclamptic (P) patients. RightGraph displaying densitometricanalysis of all the samples. Eachpoint on the graph corre-sponds to an individual patient(means±SE). **P≤0.01,***P≤0.001; Student′s t-test
439
present in the insoluble fraction was significantly higher inplacentae from normal patients than in those derived frompreeclamptic women.
Claudin 3
We also analyzed the distribution of claudins 3 and 15,which previously had respectively been located at theblood-brain barrier (Wolburg et al. 2003) and in kidneyvessels (Kiuchi–Saishin et al. 2002). Claudin 15 wasundetectable in small and in large placental vessels in bothnormal and preeclamptic women (data not shown). How-ever, claudin 3 was clearly immunolocalized at the largevessels present in the placentae of both normal andpreeclamptic women (Fig. 5a). In contrast, the smallerplacental vessels displayed a barely detectable signal forclaudin 3 (Fig. 5b). The Western blot assay revealed agreater amount of claudin 3 in the soluble fraction than inthe insoluble fraction in both normal and preeclampticwomen, and a smaller amount of claudin 3 in the Triton X–100 soluble fraction of preeclamptic placentae comparedwith that detected in normal women (Fig. 5c).
Other claudins
We subsequently studied the immunoreactivity of placentaltissue to claudins 1, 4, 7, and 16. Whereas claudin 7 wasbarely detectable, the other three claudins showed aconspicuous presence in placental tissue.
Claudin 1
In full term human placentae, claudin 1 has been previouslylocalized in large vessels corresponding to arteries andveins of stem villi, whereas terminal villous capillarieswere immunonegative (Leach et al. 2002). Our immuno-fluorescence results with placentae from normal andpreeclamptic women are in agreement with these observa-tions (Fig. 6a,b). Western blot analysis indicates a smalleramount of claudin 1 in the TX–100 insoluble fraction thanin the soluble fraction. In the insoluble samples derivedfrom preeclamptic patients, the amount of claudin 1 issignificantly reduced (Fig. 6c).
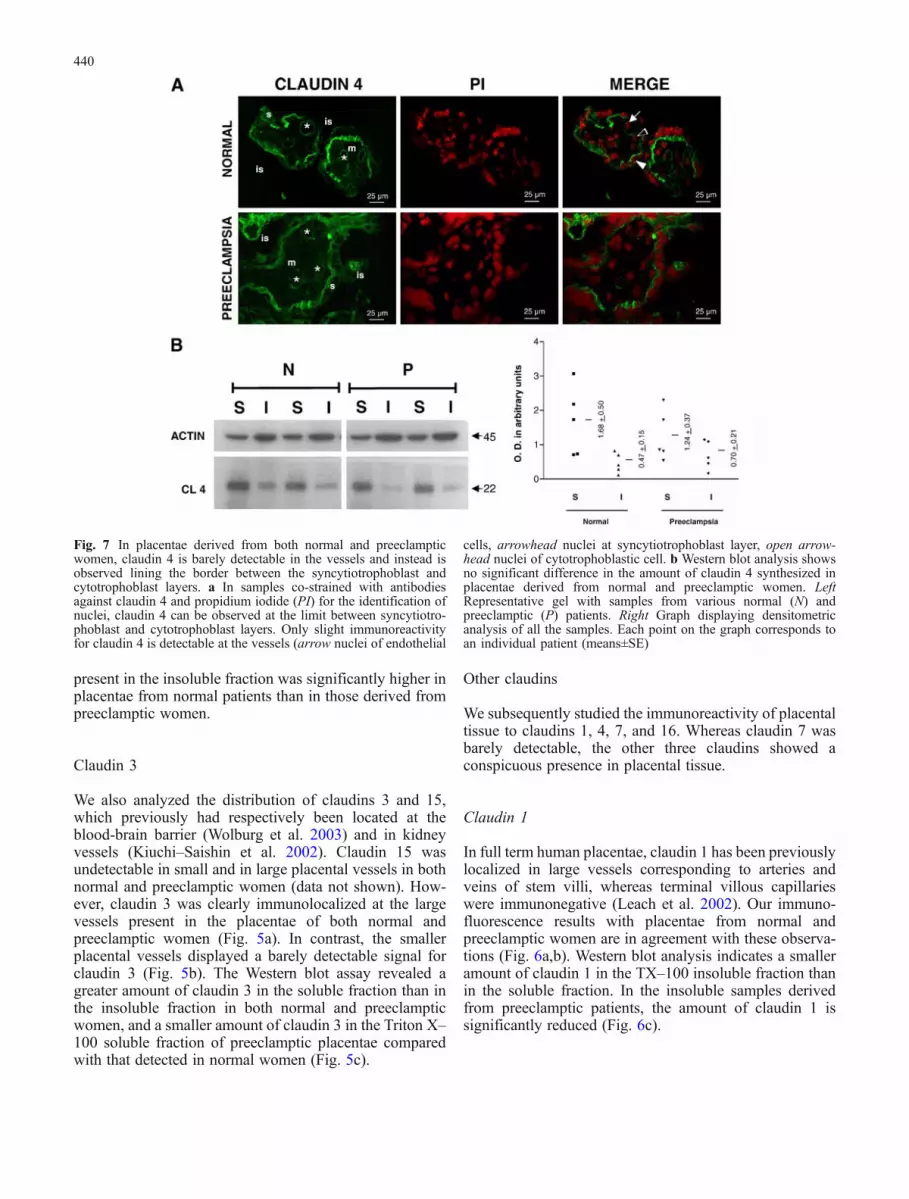
Fig. 7 In placentae derived from both normal and preeclampticwomen, claudin 4 is barely detectable in the vessels and instead isobserved lining the border between the syncytiotrophoblast andcytotrophoblast layers. a In samples co-strained with antibodiesagainst claudin 4 and propidium iodide (PI) for the identification ofnuclei, claudin 4 can be observed at the limit between syncytiotro-phoblast and cytotrophoblast layers. Only slight immunoreactivityfor claudin 4 is detectable at the vessels (arrow nuclei of endothelial
cells, arrowhead nuclei at syncytiotrophoblast layer, open arrow-head nuclei of cytotrophoblastic cell. b Western blot analysis showsno significant difference in the amount of claudin 4 synthesized inplacentae derived from normal and preeclamptic women. LeftRepresentative gel with samples from various normal (N) andpreeclamptic (P) patients. Right Graph displaying densitometricanalysis of all the samples. Each point on the graph corresponds toan individual patient (means±SE)
440

Claudin 4
Claudin 4 displayed a barely detectable immunoreaction atthe placental endothelia. However, in both normal andpreeclamptic placentae, claudin 4 gave a strong signal atthe basal membrane of the syncytiotrophoblast layer thatfaced the fetal endothelia, the stroma, and some remnantcytotrophoblastic cells still present at full-term. We treatedsections with both the anti-claudin 4 antibody andpropidium iodide (Fig. 7), which distinguished the nucleiof the endothelial cells (arrow), the syncytiotrophoblastlayer (full arrowhead), and the cytotrophoblastic or stromacells (empty arrowhead). Thus, claudin 4 appeared to be aprotein present at the basal membrane of the syncytiotro-phoblast layer and not a protein of the placental vessels.Western blot analysis revealed a higher amount of claudin 4in the Triton X–100 soluble fraction than in the insolublefraction in both normal and preeclamptic women. Theamount of claudin 4 found in each fraction did not varywith preeclampsia.
Claudin 16
In the human placental tissue, claudin 16 is absent from theendothelia, whereas it is conspicuously found in thesyncytiotrophoblast layer (Fig. 8a). In the placentae ofnormal women, instead of generating a claudin-4-like basalpattern, claudin 16 immunolocalizes at the plasma mem-brane of the syncytiotrophoblast layer. In the placentae
obtained from preeclamptic women, a less-well-definedpattern of claudin 16 immunolocalization is detected. ByWestern blot analysis, a similar amount of claudin 16 wasfound in both groups in the Triton-X soluble and insolublefractions (Fig. 8b).
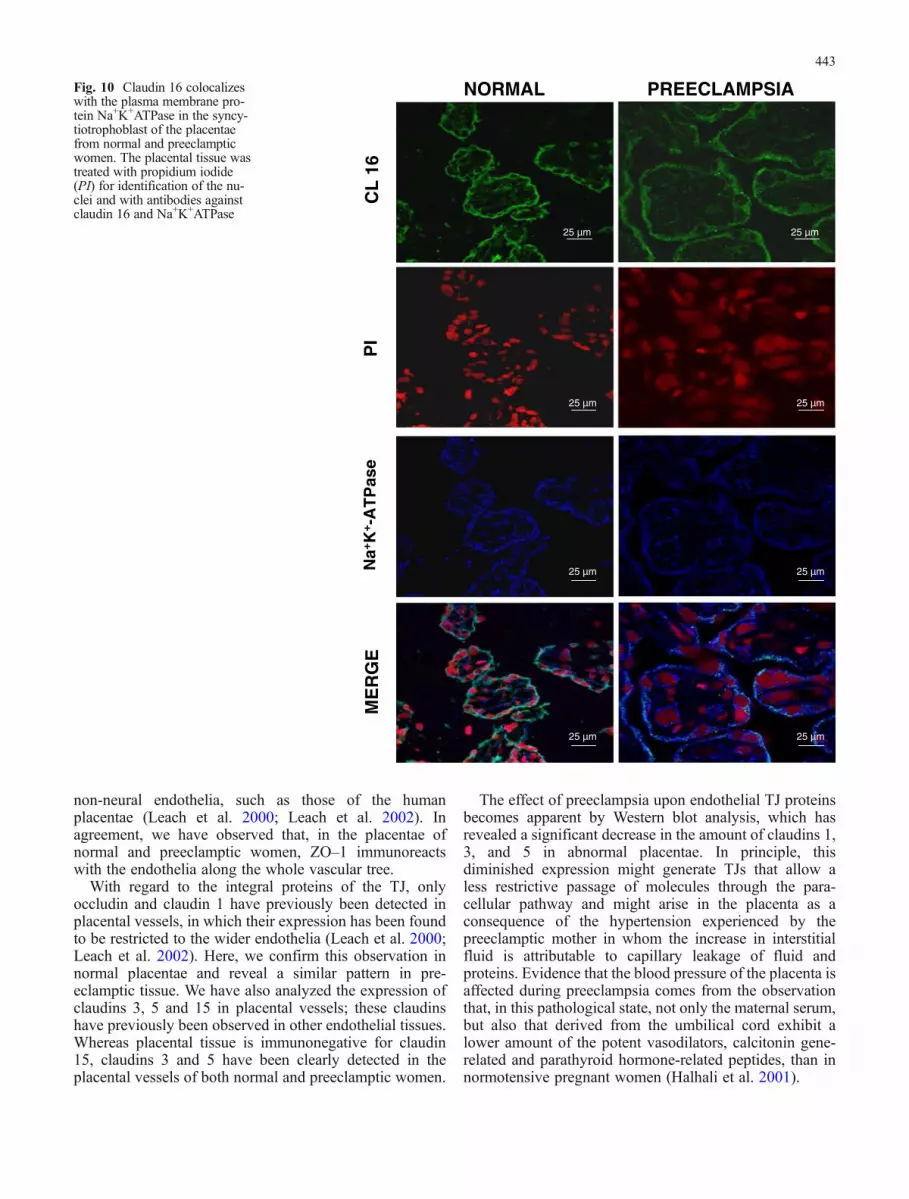
To confirm the presence of claudin 16 at the plasmamembrane, we incubated placental tissue with antibodiesagainst claudin 16 and the apical membrane glycoproteinGP135 or the basolateral membrane protein Na+K+-ATPase and, in both cases, stained the cell nuclei withpropidium iodide. Claudin 16 immunoreactivity colocal-ized with that of GP135 (Fig. 9) and Na+K+-ATPase(Fig. 10) at the plasma membrane. Whereas in epithelialcells, the distribution of these proteins was markedlypolarized to either the apical or the basolateral plasmamembrane surfaces, they appeared to display a less-well-defined pattern of polarization in the syncytiotrophoblastlayer. This was further confirmed when the placental tissuewas co-stained with antibodies against GP135 and Na+K+-ATPase (Fig. 11a,b). The confocal xz sections of normaltissue (Fig. 11b) showed in more detail that, although theNa+K+-ATPase was concentrated at the lateral membranesof the syncytiotrophoblast layer, significant immunoreac-tivity could also be detected at the apical membrane. In asimilar fashion, GP135 was clearly detectable at the apicalsurface (arrowhead) but still displayed some positiveimmunostaining at the lateral surface (arrow). The xzsections of the preeclamptic placentae displayed a morediffuse pattern of staining (data not shown).
Fig. 8 Claudin 16 is absentfrom the placental vessels,whereas it strongly immuno-reacts with the plasma mem-brane of the syncytiotrophoblastlayer. a No claudin 16 is de-tected in the placental endotheliaidentified by its positivePECAM–1 immunoreactivity.However, the plasma membraneof the syncytiotrophoblast layerof the placenta from a normalwoman is positive to claudin 16.Inset: Higher magnification ofthe syncytiotrophoblast. In pre-eclamptic placentae a less-well-defined pattern of claudin 16 isobserved at the syncytiotropho-blast layer. b Western blot anal-ysis shows no difference in theamount of claudin 16 in theplacentae of normal and pre-eclamptic women. Left Repre-sentative gel with samples fromvarious normal (N) and pre-eclamptic (P) patients. RightGraph displaying densitometricanalysis of all the samples. Eachpoint on the graph corre-sponds to an individual pa-tient (means±SE)
441

Electron-microscopic observations
Electron microscopy revealed that, in both normal andpreeclamptic placentae, the extracellular marker rutheniumred stained the microvilli of the apical surface of thesyncytiotrophoblast layer (Fig. 12, arrows). The penetra-tion of the marker was prevented at the uppermost portionof the lateral membrane, the location of the TJs (arrow-heads). The paracellular pathway traversed the syncytio-trophoblast layer connecting the maternal intervillousspace (Fig. 12, asterisk) to the fetal extracellular compart-ment found below the syncytiotrophoblast layer (Fig. 12,double asterisks). In this tissue, the paracellular pathwayexhibited both a winding (Fig. 12a) and a linear-like(Fig. 12b) pattern; however, the former was morefrequently observed in both the normal and the preeclamp-tic condition.
Discussion
Preeclampsia is a hypertensive disorder of pregnancy thatcan be life-threatening for the fetus and the mother. Centralto the pathogenesis of preeclampsia is an initial defectiveplacentation that leads to maternal endothelial dysfunction.We have explored the molecular composition of the TJ atthe terminal villi of normal and preeclamptic human termplacentae and have found that, in the pathological condi-tion, the expression of claudins 1, 3, and 5 diminishes,whereas the amount of ZO–1 and occludin remainsunchanged.
We have also analyzed the expression of ZO–1, sincethis submembranous protein is a ubiquitous marker of TJsin both epithelia and endothelia, independent of the degreeof sealing of the tissue. Thus, ZO–1 has been previouslyreported in high and low permeable epithelia, such as theproximal and collecting tubules of the kidney (Gonzalez–Mariscal et al. 2000), and in neural (Nitta et al. 2003) and
NORMAL
CL
16
PREECLAMPSIA
PI
GP
135
ME
RG
E
25 µm25 µm
25 µm25 µm
25 µm25 µm
25 µm25 µm
Fig. 9 Claudin 16 colocalizeswith the plasma membrane pro-tein GP135 in the syncytiotro-phoblast of the placentae fromnormal and preeclampticwomen. The placental tissuewas treated with propidium io-dide (PI) for identification of thenuclei and with antibodiesagainst claudin 16 and GP135
442
non-neural endothelia, such as those of the humanplacentae (Leach et al. 2000; Leach et al. 2002). Inagreement, we have observed that, in the placentae ofnormal and preeclamptic women, ZO–1 immunoreactswith the endothelia along the whole vascular tree.
With regard to the integral proteins of the TJ, onlyoccludin and claudin 1 have previously been detected inplacental vessels, in which their expression has been foundto be restricted to the wider endothelia (Leach et al. 2000;Leach et al. 2002). Here, we confirm this observation innormal placentae and reveal a similar pattern in pre-eclamptic tissue. We have also analyzed the expression ofclaudins 3, 5 and 15 in placental vessels; these claudinshave previously been observed in other endothelial tissues.Whereas placental tissue is immunonegative for claudin15, claudins 3 and 5 have been clearly detected in theplacental vessels of both normal and preeclamptic women.
The effect of preeclampsia upon endothelial TJ proteinsbecomes apparent by Western blot analysis, which hasrevealed a significant decrease in the amount of claudins 1,3, and 5 in abnormal placentae. In principle, thisdiminished expression might generate TJs that allow aless restrictive passage of molecules through the para-cellular pathway and might arise in the placenta as aconsequence of the hypertension experienced by thepreeclamptic mother in whom the increase in interstitialfluid is attributable to capillary leakage of fluid andproteins. Evidence that the blood pressure of the placenta isaffected during preeclampsia comes from the observationthat, in this pathological state, not only the maternal serum,but also that derived from the umbilical cord exhibit alower amount of the potent vasodilators, calcitonin gene-related and parathyroid hormone-related peptides, than innormotensive pregnant women (Halhali et al. 2001).
NORMAL PREECLAMPSIA
CL
16
PI
Na+ K
+ -A
TP
a se
ME
RG
E
25 µm 25 µm
25 µm 25 µm
25 µm 25 µm
25 µm 25 µm
Fig. 10 Claudin 16 colocalizeswith the plasma membrane pro-tein Na+K+ATPase in the syncy-tiotrophoblast of the placentaefrom normal and preeclampticwomen. The placental tissue wastreated with propidium iodide(PI) for identification of the nu-clei and with antibodies againstclaudin 16 and Na+K+ATPase
443

In this study, we have unexpectedly found that claudins4 and 16 are present at the plasma membrane of thesyncytiotrophoblast layer. To our knowledge, this is thefirst time that claudins have been reported in a mammalian
scyncytium. The finding that these claudins are notconcentrated at the limit between the apical and the lateralmembranes is not surprising, since several claudins havebeen found to display a non-TJ restricted distribution. Forexample, in the rabbit bladder epithelium, claudin 4 isfound along the basolateral margins of the umbrella celllayer (Acharya et al. 2004), and in the rat uterus, claudins 1,3, and 5 change their expression from the uppermost regionof the lateral plasma membrane to the lateral and basaldomains during the estrous cycle, whereas claudin 7 isalways detected at the basolateral membrane of the ratuterus (Mendoza–Rodriguez et al. 2005).
In epithelial cells, claudin 4 diminishes the paracellularconductance through a selective decrease in Na+ perme-ability (Van Itallie et al. 2001). This effect can be reversedby substituting a negative for a positive charge in the firstextracellular loop of claudin 4 (Colegio et al. 2003).However, in the human placentae that we have studied, wehave not observed claudin 4 at the lateral membrane whereit could function as a paracellular regulator of Na+
permeability. The reason for the presence of claudin 4 atthe basal membrane of the syncytiotrophoblast layerremains unclear. However, from a speculative point ofview, its presence could be attributable to another role ofclaudins related to adhesion to molecules of the extracel-lular matrix, a point whose validity should be clarified byfuture research.
Claudin 16 has been found to be largely restricted to theTALH (Simon et al. 1999). The intraluminal positiveelectrical potential that is only present in this part of thenephron drives cations back to the blood through the TJs(Monnens et al. 2000). Indeed, in every species studied todate, 60% of the filtered Mg2+ is reabsorbed in the loop ofHenle, and this process occurs within the cortical TALH bypassive diffusion through the paracellular pathway(Quamme and de Rouffignac 2000). The discovery thatmutations in the gene for claudin 16 are responsible for afamilial Mg2+ wasting disease named hypomagnesemiawith hypercalciuria and nephrocalcinosis (Simon et al.1999) has given rise to the concept that claudins areselective ionic channels of the paracellular pathway (Wongand Goodenough 1999). In agreement, the first extracel-lular loop of claudin 16 contains a large number of negativeresidues that are compatible with the formation of a cationselective channel. Recently, another human disease, viz.idiopathic hypercalciuria, which is characterized by abnor-mal renal loss, has been shown to result from mutations inclaudin 16 that block its localization at the TJ and insteadlead to its accumulation at the lysosome (Muller et al.2003). The reason for the human syncytiotrophoblast layerexpressing claudin 16 at the plasma membrane remains
PI
ME
RG
EG
P 1
35NORMAL PREECLAMPSIA
25 µm 25 µm
25 µm 25 µm
25 µm 25 µm
25 µm 25 µm
Na+ K
+ -A
TP
a se
PI
ME
RG
EG
P 1
3 5
8 µm
8 µm
8 µm
8 µm
Na+
K+ -
AT
Pas
eA
B
3Fig. 11 Na+K+ATPase and GP135 display a mildly polarized patternin the plasma membrane of the syncytiotrophoblast layer. Theplacental tissue was treated with propidium iodide (PI) for identifi-cation of the nuclei and with antibodies against Na+K+ATPase andGP135. a Confocal xy sections. b Confocal xz sections. GP135 wasreadily detected at the apical surface (arrowhead) and also exhibitedpositive immunostaining at the lateral surface (arrow)
444

unknown. However, since Mg2+ supplementation reducesblood pressure (Jee et al. 2002), it has been employed inhuman pregnancy. Indeed, hypomagnesemia has beenfound to be more pronounced in women with preeclampsiathan in women with normal pregnancies (Standley et al.1997; Kisters et al. 1998), suggesting that this situationmay contribute to the development of hypertension in thesepatients (Kisters et al. 2000; Adam et al. 2001). However,whereas some studies report that Mg2+ supplementationduring pregnancy reduces the frequency of fetal growthretardation and preeclampsia (Conradt et al. 1984) or ofmaternal and fetal morbidity (Spatling and Spatling 1988),others have observed that magnesium supplementationdoes not improve pregnancy outcome (Sibai et al. 1989;Morris et al. 2001). This is, of course, unrelated to theeffectiveness of parenteral MgSO4 as an anticonvulsantdrugs given to women with preeclampsia to reduce the riskof seizures (Belfort et al. 2003; Altman et al. 2002;Eclampsia Trial Collaborative Group 1995). The way thatMgSO4 controls eclamptic convulsions is unclear. Its effectmay be in part through the role of magnesium as a blockerof excitatory amino acid receptors, such as NMDA,
kainate, and AMPA, in the brain (Hallak et al. 2000;Cotton et al. 1993) and/or attributable to a vasodilationeffect. Mg2+ may have a generalized effect on all smoothmuscle, including the peripheral vasculature and the uterus,and hence have antihypertensive and toxolytic effects.
The presence of claudin 16 at the lateral plasmamembrane of the syncytiotrophoblast layer is the firstevidence to suggest the occurrence of paracellular transportof Mg2+ between the maternal blood and the intercellularspace of the human placenta. Of note, fetal blood hashigher levels of ionized Mg2+ in comparison with maternalblood (Standley and Standley 2002). The absence ofclaudin 16 immunolocalization at the placental vesselsfurther suggests that once Mg2+ enters the lumen ofplacental vessels, by an as yet undefined mechanism, thecation might not be driven out through the TJ. With regardsto Mg2+ transport in the human placenta, only the presenceof a functional Na+/Mg2+ exchanger that maintains lowintracellular Mg2+ in choriocarcinoma JEG–3 cells inculture has been reported (Standley and Standley 2002).
From a speculative perspective, the conspicuous expres-sion of claudin 16 at the lateral plasma membrane of the
Fig. 12 Paracellular pathway ofthe syncytiotrophoblast layerconnects the fetal interstitiumwith the maternal blood spaceand blocks the passage of ruthe-nium red at the TJ region (as-terisks maternal blood space,double asterisks fetal extracellu-lar compartment located underthe syncytiotrophoblast layer).Placentae from both normal(a) and preeclamptic (b) womenwere treated with ruthenium redand processed for electron mi-croscopy. The electron-densemarker is detected at the micro-villi of the syncytiotrophoblastlayer (arrows), but its passagethrough the paracellular route isblocked at the TJ region (ar-rowheads)
445

syncytiotrophoblast layer might give rise to a passiveparacellular flux of Mg2+ to the placental tissue that may, inturn, exert a vasodilation effect on the placental vessels.Accordingly in preeclampsia, the less-well-defined patternof claudin 16 expression in the syncytiotrophoblast layermight generate a lower influx of Mg2+ to the placentaltissue with a consequent minimization of the vasodilationeffect.
The presence of a paracellular network in the syncytiallayer of the hemochorial placentae was postulated severalyears ago (Stulc et al. 1969; Stulc 1989a). Transportexperiments demonstrated that the transplacental flux ofsmall hydrophilic molecules was proportional to theirdiffusion coefficients in water (Stulc 1989b; Thornburgand Faber 1977; Hedley and Bradbury 1980; Stulc andStulcova 1986; Robinson et al. 1988), thus suggestingtransport across the placenta via wide aqueous channels.
However, the morphological evidence for this pathwayhas been a matter of debate because of the syncytial natureof the syncytiotrophoblast layer. The existence of mem-brane-lined channels that cross the syncytiotrophoblast hasbeen proposed after observations of the perfusion ofelectron-dense markers and their increased movementacross the rat and human placenta resulting from elevatedfetal hydrostatic pressures (Kertschanska et al. 2000;Kertschanska et al. 1997). This so-called transtrophoblasticchannel system, which apparently originates from therichly folded basal plasmalemma invaginations, providesan open paracellular route that connects the fetal intersti-tium with the maternal blood space. Our transmissionelectron micrographs illustrate the way that the paracellularpathway of the syncytiotrophoblast layer connects theintervillous space to the fetal extracellular compartmentfound below the syncytiotrophoblast layer. The immuno-localization of claudin 16, Na+K+-ATPase, and (to a lesserextent) GP135 allows a further characterization of thisparacellular network of the human placentae.
Acknowledgement We thank Dr. Oscar Arturo Martinez Rodri-guez, Director of the Gynecological Hospital of La Raza MedicalCenter-IMSS in Mexico City, for his help in obtaining the placentae.This work was submitted in partial fulfilment of the requirements forthe PhD degree awarded to Samuel Liévano (Doctorate in Biomed-ical Science of the National Autonomous University of Mexico;UNAM).
References
Acharya P, Beckel J, Ruiz WG, Wang E, Rojas R, Birder L,Apodaca G (2004) Distribution of the tight junction proteinsZO–1, occludin, and claudin–4, –8, and –12 in bladderepithelium. Am J Physiol Renal Physiol 287:F305–F318
Adam B, Malatyalioglu E, Alvur M, Talu C (2001) Magnesium, zincand iron levels in pre-eclampsia. JMatern Fetal Med 10:246–250
Altman D, Carroli G, Duley L, Farrell B, Moodley J, Neilson J,Smith D (2002) Do women with pre-eclampsia, and theirbabies, benefit from magnesium sulphate? The Magpie Trial: arandomised placebo-controlled trial. Lancet 359:1877–1890
Anderson JM, Cereijido M (2001) Evolution of ideas on the tightjunction. In: Cereijido M, Anderson JM (eds) Tight junctions.CRC Press, Boca Raton, pp 1–18
Balda MS, Garrett MD, Matter K (2003) The ZO–1–associated Y–box factor ZONAB regulates epithelial cell proliferation andcell density. J Cell Biol 160:423–432
Balda MS, Whitney JA, Flores C, Gonzalez S, Cereijido M, MatterK (1996) Functional dissociation of paracellular permeabilityand transepithelial electrical resistance and disruption of theapical-basolateral intramembrane diffusion barrier by expres-sion of a mutant tight junction membrane protein. J Cell Biol134:1031–1049
Bayer R (1990) Topological disposition of the sequences -QRKIVE-and -KETYY in native (Na+ + K+)-ATPase.Biochemistry29:2251–2256
Bazzoni G, Dejana E (2004) Endothelial cell-to-cell junctions:molecular organization and role in vascular homeostasis.Physiol Rev 84:869–901
Belfort MA, Anthony J, Saade GR, Allen JC Jr (2003) Acomparison of magnesium sulfate and nimodipine for theprevention of eclampsia. N Engl J Med 348:304–311
Brosens IA, Robertson WB, Dixon HG (1972) The role of the spiralarteries in the pathogenesis of preeclampsia. Obstet GynecolAnnu 1:177–191
Cereijido M, Contreras RG, Gonzalez–Mariscal L (1989) Develop-ment and alteration of polarity. Annu Rev Physiol 51:785–795
Colegio OR, Van Itallie C, Rahner C, Anderson JM (2003) Claudinextracellular domains determine paracellular charge selectivityand resistance but not tight junction fibril architecture. Am JPhysiol Cell Physiol 284:C1346–C1354
Conradt A, Weidinger H, Algayer H (1984) On the role ofmagnesium in fetal hypotrophy, pregnancy induced hyperten-sion and preeclampsia. Magnesium Bull 6:68–76
Cotton DB, Hallak M, Janusz C, Irtenkauf SM, Berman RF (1993)Central anticonvulsant effects of magnesium sulfate on N-methyl-D-aspartate-induced seizures. Am J Obstet Gynecol168:974–978
Eclampsia Trial Collaborative Group (1995) Which anticonvulsantfor women with eclampsia? Evidence from the CollaborativeEclampsia Trial. Lancet 345:1455–1463
Gonzalez–Mariscal L, Namorado MC, Martin D, Luna J, Alarcon L,Islas S, Valencia L, Muriel P, Ponce L, Reyes JL (2000) Tightjunction proteins ZO–1, ZO–2, and occludin along isolatedrenal tubules. Kidney Int 57:2386–2402
Gonzalez–Mariscal L, Betanzos A, Nava P, Jaramillo BE (2003)Tight junction proteins. Prog Biophys Mol Biol 81:1–44
Gottardi CJ, Arpin M, Fanning AS, Louvard D (1996) The junction-associated protein, zonula occludens–1, localizes to the nucleusbefore the maturation and during the remodeling of cell-cellcontacts. Proc Natl Acad Sci USA 93:10779–10784
Halhali A, Wimalawansa SJ, Berentsen V, Avila E, Thota CS, LarreaF (2001) Calcitonin gene- and parathyroid hormone-relatedpeptides in preeclampsia: effects of magnesium sulfate. ObstetGynecol 97:893–897
Hallak M, Hotra JW, Custodio D, Kruger ML (2000) Magnesiumprevents seizure-induced reduction in excitatory amino acidreceptor (kainate and alpha-amino-3-hydroxy-5-methyliso-xazole-4-propionic acid) binding in pregnant rat brain. Am JObstet Gynecol 183:793–798
Hedley R, Bradbury MW (1980) Transport of polar non-electrolytesacross the intact and perfused guinea-pig placenta. Placenta1:277–285
Islas S, Vega J, Ponce L, Gonzalez–Mariscal L (2002) Nuclearlocalization of the tight junction protein ZO–2 in epithelialcells. Exp Cell Res 274:138–148
Jaramillo BE, Ponce A, Moreno J, Betanzos A, Huerta M, Lopez–Bayghen E, Gonzalez–Mariscal L (2004) Characterization ofthe tight junction protein ZO–2 localized at the nucleus ofepithelial cells. Exp Cell Res 297:247–258
Jee SH, Miller ER, III, Guallar E, Singh VK, Appel LJ, Klag MJ(2002) The effect of magnesium supplementation on bloodpressure: a meta-analysis of randomized clinical trials. Am JHypertens 15:691–696
446

Kertschanska S, Kosanke G, Kaufmann P (1997) Pressure depen-dence of so-called transtrophoblastic channels during fetalperfusion of human placental villi. Microsc Res Tech 38:52–62
Kertschanska S, Stulcova B, Kaufmann P, Stulc J (2000) Distensibletranstrophoblastic channels in the rat placenta. Placenta21:670–677
Khong TY, De Wolf F, Robertson WB, Brosens I (1986) Inadequatematernal vascular response to placentation in pregnanciescomplicated by pre-eclampsia and by small-for-gestational ageinfants. Br J Obstet Gynaecol 93:1049–1059
Kisters K, Korner J, Louwen F, Witteler R, Jackisch C, Zidek W, OttS, Westermann G, Barenbrock M, Rahn KH (1998) Plasma andmembrane Ca2+ and Mg2+ concentrations in normal pregnancyand in preeclampsia. Gynecol Obstet Invest 46:158–163
Kisters K, Barenbrock M, Louwen F, Hausberg M, Rahn KH, KoschM (2000) Membrane, intracellular, and plasma magnesium andcalcium concentrations in preeclampsia. Am J Hypertens13:765–769
Kiuchi–Saishin Y, Gotoh S, Furuse M, Takasuga A, Tano Y, TsukitaS (2002) Differential expression patterns of claudins, tightjunction membrane proteins, in mouse nephron segments. J AmSoc Nephrol 13:875–886
Leach L, Firth JA (1992) Fine structure of the paracellular junctionsof terminal villous capillaries in the perfused human placenta.Cell Tissue Res 268:447–452
Leach L, Lammiman MJ, Babawale MO, Hobson SA, Bromilou B,Lovat S, Simmonds MJ (2000) Molecular organization of tightand adherens junctions in the human placental vascular tree.Placenta 21:547–557
Leach L, Babawale MO, Anderson M, Lammiman M (2002)Vasculogenesis, angiogenesis and the molecular organisation ofendothelial junctions in the early human placenta. J Vasc Res39:246–259
McCarthy KM, Skare IB, Stankewich MC, Furuse M, Tsukita S,Rogers RA, Lynch RD, Schneeberger EE (1996) Occludin is afunctional component of the tight junction. J Cell Sci109:2287–2298
Mendoza–Rodriguez CA, Gonzalez–Mariscal L, Cerbon M (2005)Changes in the distribution of ZO–1, occludin, and claudins inthe rat uterine epithelium during the estrous cycle. Cell TissueRes 319:315–330
Monnens L, Starremans P, Bindels R (2000) Great strides in theunderstanding of renal magnesium and calcium reabsorption.Nephrol Dial Transplant 15:568–571
Morita K, Sasaki H, Furuse M, Tsukita S (1999) Endothelialclaudin: claudin–5/TMVCF constitutes tight junction strands inendothelial cells. J Cell Biol 147:185–194
Morris CD, Jacobson SL, Anand R, Ewell MG, Hauth JC, Curet LB,Catalano PM, Sibai BM, Levine RJ (2001) Nutrient intake andhypertensive disorders of pregnancy: evidence from a largeprospective cohort. Am J Obstet Gynecol 184:643–651
Muhleisen H, Wolburg H, Betz E (1989) Freeze-fracture analysis ofendothelial cell membranes in rabbit carotid arteries subjectedto short-term atherogenic stimuli. Virchows Arch B Cell Pathol56:413–417
Muller D, Kausalya PJ, Claverie–Martin F, Meij IC, Eggert P,Garcia–Nieto V, Hunziker W (2003) A novel claudin 16mutation associated with childhood hypercalciuria abolishesbinding to ZO–1 and results in lysosomal mistargeting. Am JHum Genet 73:1293–1301
Newman PJ (1997) The biology of PECAM–1. J Clin Invest 99:3–8Nitta T, Hata M, Gotoh S, Seo Y, Sasaki H, Hashimoto N, Furuse M,
Tsukita S (2003) Size-selective loosening of the blood-brainbarrier in claudin-5-deficient mice. J Cell Biol 161:653–660
Pridjian G, Puschett JB (2002) Preeclampsia. Part 2: experimentaland genetic considerations. Obstet Gynecol Surv 57:619–640
Quamme GA, de Rouffignac C (2000) Epithelial magnesiumtransport and regulation by the kidney. Front Biosci 5:D694–D711
Rahner C, Mitic LL, Anderson JM (2001) Heterogeneity inexpression and subcellular localization of claudins 2, 3, 4,and 5 in the rat liver, pancreas, and gut. Gastroenterology120:411–422
Redman CW (1991) Current topic: pre-eclampsia and the placenta.Placenta 12:301–308
Reyes JL, Lamas M, Martin D, Carmen NM del, Islas S, Luna J,Tauc M, Gonzalez–Mariscal L (2002) The renal segmentaldistribution of claudins changes with development. Kidney Int62:476–487
Roberts JM (1989) Pregnancy related hypertension. In: Creasy RK,Resnik R (eds) Maternal fetal medicine: principles and practice,2nd edn. Saunders, Philadelphia, pp 783–808
Robillard PY, Hulsey TC, Perianin J, Janky E, Miri EH, Papiernik E(1994) Association of pregnancy-induced hypertension withduration of sexual cohabitation before conception. Lancet344:973–975
Robinson NR, Atkinson DE, Jones CJ, Sibley CP (1988) Perme-ability of the near-term rat placenta to hydrophilic solutes.Placenta 9:361–372
Saitou M, Furuse M, Sasaki H, Schulzke JD, Fromm M, Takano H,Noda T, Tsukita S (2000) Complex phenotype of mice lackingoccludin, a component of tight junction strands. Mol Biol Cell11:4131–4142
Sambrook J, Russell DW (2001) Preparation and analysis ofeukaryotic genomic DNA. In: Sambrook J, Russell DW (eds)Molecular cloning: a laboratory manual, 3rd edn. Cold SpringHarbor Laboratory Press, Cold Spring Harbor, NY, pp 6.1–6.64
Sibai BM, Villar MA, Bray E (1989) Magnesium supplementationduring pregnancy: a double-blind randomized controlled clin-ical trial. Am J Obstet Gynecol 161:115–119
Simionescu M, Simionescu N, Palade GE (1976) Segmentaldifferentiations of cell junctions in the vascular endothelium.Arteries and veins. J Cell Biol 68:705–723
Simon DB, Lu Y, Choate KA, Velazquez H, Al Sabban E, Praga M,Casari G, Bettinelli A, Colussi G, Rodriguez–Soriano J,McCredie D, Milford D, Sanjad S, Lifton RP (1999)Paracellin–1, a renal tight junction protein required forparacellular Mg2+ resorption. Science 285:103–106
Soto–Wright V, Bernstein M, Goldstein DP, Berkowitz RS (1995)The changing clinical presentation of complete molar pregnan-cy. Obstet Gynecol 86:775–779
Spatling L, Spatling G (1988) Magnesium supplementation inpregnancy. A double-blind study. Br J Obstet Gynaecol95:120–125
Standley CA, Whitty JE, Mason BA, Cotton DB (1997) Serumionized magnesium levels in normal and preeclamptic gesta-tion. Obstet Gynecol 89:24–27
Standley PR, Standley CA (2002) Identification of a functionalNa+/Mg2+ exchanger in human trophoblast cells. Am JHypertens 15:565–570
Stulc J (1989a) Extracellular transport pathways in the haemochorialplacenta. Placenta 10:113–119
Stulc J (1989b) Study of the permeability of the intact guinea pigplacenta to hydrophilic molecules. Placenta 10:427–428
Stulc J, Stulcova B (1986) Transport of calcium by the placenta ofthe rat. J Physiol (Lond) 371:1–16
Stulc J, Friedrich R, Jiricka Z (1969) Estimation of the equivalentpore dimensions in the rabbit placenta. Life Sci 8:167–180
Thornburg KL, Faber JJ (1977) Transfer of hydrophilic moleculesby placenta and yolk sac of the guinea pig. Am J Physiol 233:C111–C124
447

Tsukita S, Furuse M (1999) Occludin and claudins in tight-junctionstrands: leading or supporting players? Trends Cell Biol 9:268–273
Tsukita S, Furuse M (2000) Pores in the wall: claudins constitutetight junction strands containing aqueous pores. J Cell Biol149:13–16
Van Itallie C, Rahner C, Anderson JM (2001) Regulated expressionof claudin–4 decreases paracellular conductance through aselective decrease in sodium permeability. J Clin Invest107:1319–1327
Wolburg H, Wolburg–Buchholz K, Kraus J, Rascher–Eggstein G,Liebner S, Hamm S, Duffner F, Grote EH, Risau W, EngelhardtB (2003) Localization of claudin–3 in tight junctions of theblood-brain barrier is selectively lost during experimentalautoimmune encephalomyelitis and human glioblastoma multi-forme. Acta Neuropathol (Berl) 105:586–592
Wong V, Goodenough DA (1999) Paracellular channels! Science285:62
Zhou Y, Damsky CH, Chiu K, Roberts JM, Fisher SJ (1993)Preeclampsia is associated with abnormal expression of adhe-sion molecules by invasive cytotrophoblasts. J Clin Invest91:950–960
Zhou Y, Damsky CH, Fisher SJ (1997a) Preeclampsia is associatedwith failure of human cytotrophoblasts to mimic a vascularadhesion phenotype. One cause of defective endovascularinvasion in this syndrome? J Clin Invest 99:2152–2164
Zhou Y, Fisher SJ, Janatpour M, Genbacev O, Dejana E, WheelockM, Damsky CH (1997b) Human cytotrophoblasts adopt avascular phenotype as they differentiate. A strategy forsuccessful endovascular invasion? J Clin Invest 99:2139–2151
448