effects of elevated co2 (face) on the functional ecology of the drought-deciduous mojave desert...
TRANSCRIPT

Environmental and Experimental Botany 48 (2002) 93–106
Effects of elevated CO2 (FACE) on the functional ecologyof the drought-deciduous Mojave Desert shrub, Lycium
andersonii
Erik P. Hamerlynck a,*, Travis E. Huxman b, Therese N. Charlet c,Stanley D. Smith c
a Department of Biological Sciences, Rutgers Uni�ersity, 101 Warren Street, Newark, NJ 07102-1811, USAb Ecology and E�olutionary Biology, Uni�ersity of Arizona, Biosciences West Room c310, Tucson, AZ 85721, USA
c Department of Biological Sciences, Uni�ersity of Ne�ada, Las Vegas, 4505 Maryland Parkway, Las Vegas, NV 89154-4004, USA
Received 4 October 2001; received in revised form 28 January 2002; accepted 30 January 2002
Abstract
Elevated CO2 may improve the productivity of cool-season active (‘drought-deciduous’) shrubs in the deserts ofsouthwestern North America by reducing early-season phenological constraints imposed by low leaf area whenphotosynthetic capacity is high and later-season physiological limitations from declining photosynthesis and middaywater potentials. Altered productivity under elevated CO2 would depend on the specific responses of short-shoots thatonly provide early-season leaf area display, and long-shoots which determine annual growth increment in these plants.We measured plant water relations, photosynthetic gas exchange, and growth in short- and long-shoots of thedrought-deciduous shrub, Lycium andersonii, under Free Air CO2 Enrichment (FACE) in the field in an intact MojaveDesert ecosystem. We were specifically interested in the differential effects CO2 enrichment would have on short-shootsand actively growing long-shoots during canopy development. Net photosynthesis (Anet) was similar in elevated comparedwith ambient CO2, but stomatal conductance (gs) was reduced by 27% in both shoot types. L. andersonii growing inelevated CO2 had larger leaves on short-shoots, and more leaves per shoot length on long-shoots. Enhanced leaf growthdid not counter lower gs, and midday plant water potential was similar between treatments. In both short- and long-shoots,down-regulation of light-saturated photosynthetic electron transport rate (Jmax) occurred under elevated CO2. However,the balance between rubisco efficiency (estimated by the maximum carboxylation rate of rubisco, Vcmax), and electrontransport capacity (Vcmax/Jmax) remained constant in short-shoots, but increased in elevated CO2 grown long-shoots.Apparent quantum requirement was similar, while light-saturated photosynthetic rates (Amax) decreased by approximately30% under elevated CO2 in both shoot-types. These results suggest that elevated CO2 lowered investment to photosyntheticelectron transport capacity and whole-plant water use, even when leaf growth was stimulated. Such canopy dynamicsare likely to enhance the ability of this drought-deciduous species to better cope with the highly variable inter- andintra-annual climate regimes characteristic of North American deserts. © 2002 Elsevier Science B.V. All rights reserved.
Keywords: Elevated CO2; Growth; Lycium andersonii ; Mojave Desert; Photosynthesis; Water relations
www.elsevier.com/locate/envexpbot
* Corresponding author. Tel.: +1-973-353-1365; fax: +1-973-353-5518.E-mail address: [email protected] (E.P. Hamerlynck).
S0098-8472/02/$ - see front matter © 2002 Elsevier Science B.V. All rights reserved.
PII: S0098 -8472 (02 )00012 -6

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–10694
1. Introduction
Cool-season active (drought-deciduous) C3
shrubs are important contributors to the pro-ductivity and species diversity of North Ameri-can warm deserts (Smith et al., 1997).Drought-deciduous shrubs coordinate canopygrowth and photosynthetic activity with pre-dictable cool-season rains, and are characterizedas having relatively high photosynthetic capac-ities and stomatal conductances (Smith and No-bel, 1986; Ehleringer, 1994). During highprecipitation years, productivity in deserts canbe several orders of magnitude higher than lev-els achieved during low rainfall years, due togreater growth in ephemeral annual species(Huxman and Smith, 2001), and drought-decidu-ous shrubs (Hunter, 1994; Smith et al., 1997).While the effects of elevated CO2 have beenevaluated in terms of the ecologically and evolu-tionarily important characteristics of annual andevergreen life-forms (Huxman et al., 1998;Hamerlynck et al., 2000a; Huxman and Smith,2001), there has been a lack of focus on long-lived shrubs with different life history patterns,despite their overall importance to desert ecosys-tem function.
A critical feature in the annual productivity ofdrought-deciduous shrubs is short-shoot/long-shoot architecture. Short shoots are producedfrom compressed meristems along the mainbranch axes. Short shoots support rapid, earlyseason leaf area expansion, but do not con-tribute significantly to biomass accumulation,which is determined by long shoot growth,which may not occur in low rainfall years. Thisstrategy maximizes carbon gain in favorableyears, and increases survivorship by reducingbiomass mortality in unfavorable years (Smith etal., 1997). Canopy development lags behindphotosynthetic development in this life-form(Comstock and Ehleringer, 1986). Therefore,seasonal productivity in deciduous shrubs ini-tially operates under a phenological constraint(high photosynthetic capacity with low leaf area)followed by a period of physiological constraint(low photosynthetic capacity with high leaf area)as photosynthetic capacity declines in response
to falling water potential well before canopygrowth finishes.
The functional attributes of drought-deciduousshrubs suggest that they are likely to increasebiomass production with increasing levels of at-mospheric CO2, especially under conditions thatfavor long-shoot growth. In C3 plants, elevatedCO2 reduces diffusional limitations to photosyn-thesis, increases carboxylation, and reduces oxy-genase activity at ribulose-1,5-bisphophatecarboxylase/oxygenase (Rubisco) (Sage, 1994).Reductions in stomatal conductance and stom-atal density can also lead to decreases in wholeplant water use, slowing the rate of soil waterdepletion and improving plant water status andproductivity (Knapp et al., 1996; Huxman et al.,1998; Hamerlynck et al., 2000b). These factorscould reduce the impact of seasonal water limi-tations, an important feature in the functionalecology of desert plants (Smith et al., 1997;Hamerlynck et al., 2000a), and could be espe-cially important in desert plants capable ofrapid growth (Smith et al., 2000; Huxman andSmith, 2001) such as deciduous shrubs.
Prolonged exposure to elevated CO2 also of-ten results in photosynthetic down-regulation,usually expressed as lowered photosynthetic ca-pacity and mesophyll carboxylation efficiency.Photosynthetic down-regulation may be ecologi-cally important, since allocation away from pho-tosynthetic capacity could be used in other plantprocesses (Sage, 1994). The long-term effects ofdown-regulation in long-lived woody specieswould depend on the duration and timing ofdown-regulatory behavior, which is known tovary with water status (Oechel et al., 1995; Hux-man et al., 1998; Hamerlynck et al., 2000a) andtemperature (Lewis et al., 1996; Stirling et al.,1997; Hamerlynck et al., 2000b). Also, the rela-tive proportions of growing and non-growingstructures determine the stimulatory effect ofCO2 on plant growth; actively growing tissuehas been found to show little evidence of down-regulation, while non-growing photosynthetic tis-sue often displays strong down-regulation underelevated CO2 (Wait et al., 1999). The interactionof these factors could be important in determin-ing the ecological role of drought-deciduous

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106 95
shrubs in aridland systems under elevated CO2
conditions.Here, we present findings of a field study on
the water relations, photosynthetic gas exchange,and growth of a drought-deciduous MojaveDesert shrub, Lycium andersonii Gray, duringseasonally declining moisture and increasingtemperature using Free Air CO2 Enrichment(FACE) technology. FACE is ideal for studiesof arid systems with low plant densities, becauseit allows for the fumigation of extensive areasand for the study of intact ecosystems (Jordanet al., 1999). L. andersonii is an excellent repre-sentative of a large range of microphyllousdrought-deciduous shrubs common to NorthAmerican warm deserts (Smith et al., 1997). Ad-ditionally, the Mojave Desert experienced excep-tionally high, El Nino driven precipitation theyear of study (Huxman and Smith, 2001), allow-ing us to evaluate physiological performance un-der conditions amendable to extensivelong-shoot activity and biomass accumulation.We tested four specific hypotheses:1. Seasonal declines in water potential would be
less pronounced under elevated CO2 concen-trations, and would accompany lower stom-atal conductance, prolonging seasonalactivity (Knapp et al., 1996).
2. Responses of short shoots and long shootsto elevated CO2 would vary with their func-tional role. Short shoots supply energy forlong shoot growth, and could therefore showphotosynthetic down-regulation early ingrowth, when long shoots are not acting asstrong carbon sinks, a factor known to medi-ate CO2-related regulatory responses(Thomas and Strain, 1991; Stirling et al.,1997; Wait et al., 1999).
3. Actively growing long shoots would showlimited down-regulation of photosynthetic ca-pacity at elevated CO2 (Wait et al., 1999).
4. Down-regulation under elevated CO2 woulddecrease with declining plant water potential,as noted in other aridland shrub species(Oechel et al., 1995; Huxman et al., 1998;Hamerlynck et al., 2000a,b).
2. Materials and methods
2.1. FACE site description
Exposure of intact Mojave Desert vegetationto elevated CO2 commenced 28 April 1997 onthe DOE Nevada Test Site (NTS, 36°49�N,115°55�W, 970 m) using FACE technology (Jor-dan et al., 1999). Nine 23-m diameter circularplots were established, three at elevated CO2 (setpoint of 550 �mol CO2 mol−1), three ambientCO2 with system blower fans but no CO2 injec-tion system (‘ambient CO2’), and three ambientCO2 non-blower controls. The plots were simi-lar, in overall species composition, but highlyvariable in density and cover of the representa-tive species. Fumigation occurs continuously, ex-cept when wind speeds exceed 6 m s−1 over a 5min period (ca. �5% of the time). A computercontrol algorithm opens and closes valves on 321.5-m high PVC risers connected to an above-ground plenum. Vaporized CO2 from a 60 met-ric ton storage vessel is injected into an ambientair stream created by a large circulating fan at-tached to the plenum. Under calm conditions(�0.1 m s−1) every other valve is opened,while at greater wind speeds, upwind valves areopened and downwind valves are closed. Thisprotocol maintains CO2 concentrations within�5% of the targeted set-point over a 5 minaveraging period. Plants were accessed from asingle within-plot walk, or from a suspended,moveable platform attached to a pivoting 24-mlong rotating aluminum catwalk supported 2.2m above ground level. In order to avoid micro-climate effects created by air exiting the risers,only plants at least 1 m away from the riserswere sampled. A more comprehensive descrip-tion of this facility, its operations, and site char-acteristics can be found in Jordan et al. (1999).
Air temperature was measured at 2 m with ashaded, naturally aspirated Cu–Co (Type T)thermocouple and a Campbell CR21X datalog-ger (Logan, UT). Air temperatures were loggedevery minute, and daily maximum, minimum,and mean temperatures were calculated from 15min averages over the diel cycle. A tippingbucket recording rain guage was used to mea-

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–10696
sure date and size of individual precipitationevents. Volumetric soil moisture (�) was mea-sured monthly with a Time Domain Reflec-trometry (TDR) system (Dynamax Inc, HoustonTX) consisting of a cable tester (Textronix1502C, Textronix, Inc, Beaverton, OR) underlaptop computer control that can be connectedto 16-channel multiplexers at each plot. Sixteensoil probes of three stainless steel rods (0.32 cmdiameter) are connected to the multiplexer viacoaxial cable. � was measured at 20 and 50 cmdepths beneath four Larrea tridentata DC [Cov]canopies and four interspace areas one meterfrom any shrub canopy. Unfortunately, equip-ment failure did not allow us to measure � after13 April, when much of the study occurred. Airtemperature, precipitation data, and descriptionof the TDR system are available at http://www.unlv.edu/Colleges/Sciences/Biology/Climate–Cha-nge–Research/, � data are available on requestfrom the NDFF ecologist at this website.
2.2. Experimental species
Lycium andersonii (Solanaceae) is a cool-sea-son active, drought-deciduous, perennial shrubcommon to middle to low elevations across theMojave Desert. L. andersonii co-ordinates leafgrowth and flowering with the heaviest seasonalrains, which usually occur on the NTS fromMarch through May, and sheds leaves at theonset of summer drought. Summer dormancycan be broken following exceptionally heavy fallrains, though this is infrequent (Hunter, 1994;Hamerlynck, personal observation). Leaves aresmall at maturity (2.0–11.0 mm), cylindrical tospatulate, and fleshy, often described as hemi-succulent (Hunter, 1994).
2.3. Ecophysiological measurements
From 19 March to 30 May 1998 biweeklyA/Ci (assimilation rate versus internal CO2 con-centration) were made on short-shoots and cur-rent year long-shoots of L. andersonii using aportable open-flow gas exchange system (Li6400, LiCOR, Inc, Lincoln, NE). Temperatureof a Peltier-cooled block was held at 25 °C, and
cuvette relative humidity was maintained at 15–25%, resulting in leaf temperatures of 18 to26 °C, and leaf-to-air vapor pressure deficits of1.7–2.8 kPa. Measurements were made undersaturating photosynthetic photon flux densities(1500 �mol m−2 s−1 PPFD) from an LED redlight source. Measurements were made from08:00 to 13:00 h to minimize diurnal changes instomatal conductance. Cuvette CO2 concentra-tions (Ca) were set by mixing vaporized CO2
from an internal CO2 source with ambient air.Shoots were exposed for 5 min to 200 �molCO2 mol−1 to stimulate stomatal opening. Ca
was reduced to 75 �mol mol−1, then raised in-crementally to 150, 250, 350, 550, 700, 800, 900,1200, and 2000 �mol mol−1. After a minimum7 min equilibration, Ci at each Ca was calcu-lated using the equations of Von Caemmererand Farquhar (1981). Mechanistic analyses ofA/Ci derived parameters of maximum electrontransport rate (Jmax), carboxylation velocity ofRubisco (Vcmax) and triose-phosphate utilization(TPU) were estimated using the recorded cuvetteconditions, with the formulas utilized in PHOTO-
SYNTHESIS ASSISTANT V. 2.1 software (DundeeScientific, Dundee, UK).
Midday (1000–1300) plant water potential (�)was estimated using a Scholander type pressurechamber (Soil Moisture Stress Inc, Santa Bar-bara, CA). Two shoots from different L. ander-sonii in each ring were harvested, with the �
values pooled to give a plot mean value. L. an-dersonii � sampling was concurrent with mea-surements of net CO2 assimilation (Anet) andstomatal conductance to water vapor (gs) madeusing a LiCOR 6400 open-flow portable photo-synthesis system. These measurements alternatedwith the biweekly A/Ci measurements. Humidityand air temperature conditions were maintainedat ambient levels obtained after shading theopen cuvette for 5 min prior to measurement.Cuvette CO2 was set to that of the treatmentplot being sampled (550 �mol mol−1 for ele-vated CO2, 360 �mol mol−1 for ambient) bymixing ambient air with CO2 from an internalCO2 injection system.

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106 97
When peak photosynthetic capacity wasreached, photosynthetic light saturation responseswere made on short- and long-shoots of the sameplant sampled for A/Ci measurements. CuvetteCO2 concentrations were set to the appropriatetreatment means (360 �mol mol−1 for ambient,550 �mol mol−1 for elevated CO2), with thetemperature control block set to 20 C, giving leaftemperatures ranging from 18 to 23 °C, andVPD’s of 1.3 to 1.7 kPa. PPFD in the cuvette wasinitially set to darkness, then raised incrementallyto 2000 �mol m−2 s−1. At each step, the enclosedleaves were allowed to equilibrate for a minimumof 2 min or a maximum of 5 min prior torecording.
2.4. Growth of canopy constituents
On 13 March 1998, two plants per plot wereselected for measurement of leaf area, long shootgrowth, and leaf phenology. Plants ranged inheight from 29 to 37 cm, and were similar inhemispherical canopy volume (Hamerlynck et al.,2002) between treatments (0.016–0.074 m3; one-way ANOVA of log-transformed data F=0.03;P=0.88; 1, 4 df). Four branch segments of 5 to30 cm length were labeled with aluminum tags oneach plant. Projected leaf area (LA) of 60 individ-ual leaves per plant from short-shoots were esti-mated weekly, using an allometric relationship[LA=0.0164*length (R2=0.97; F=2260.6; P�0.00001, 1, 74 df)] determined by relating pro-jected leaf area from leaves harvested fromoff-plot plants with a Delta-T leaf area meter(Delta-T Devices, Cambridge, UK) and leaflength measured with a digital caliper. Digitalcalipers were used to measure length and diameterof four to eight randomly selected long shoots oneach branch. Leaf number and individual leafarea on these long shoots were measured as well.Short-shoot and long-shoot leaf phenology wastracked by recording the number of leaves permm stem length on each of the four markedbranches. Also, for any the marked branches pro-ducing 1 and 2° long shoots, we counted andmeasured length, diameter, total number ofleaves, and individual leaf areas on each newshoot. For statistical analyses, all obtained values
were pooled for all plants per plot to give a meanplot value.
2.5. Statistical analyses
Three-way split-plot, two-way repeated mea-sures GLM estimate model (SYSTAT V8.0, SPSSSoftware, Chicago, IL) was used to compareshort-shoot and long-shoot growth (individualleaf area and leaves per unit stem length) andseasonal performance (Anet, gs, Vcmax, Jmax, TPUand Vcmax/Jmax) of L. andersonii growing underelevated and ambient CO2 conditions. CO2 treat-ment was the whole-plot factor, with a nestedCO2 treatment by replicate plot (n=3 for eachtreatment) interaction as the whole-plot errorterm. Date and shoot-type and all subsequentinteraction effects were the sub-plot factors, usingthe treatment-date-by-shoot-by-nested replicateinteraction as the subplot error term. Of specificinterest in these analyses were the CO2-by-date,CO2-by-shoot and CO2-by-date-by-shoot interac-tions, which would indicate treatment-specific dif-ferences of short and long shoots in growth andperformance through time.
Two-way ANOVA (SYSTAT V8.0) was used totest differences between CO2 treatment and short-shoot and long-shoot photosynthetic light satura-tion parameters, and to compare CO2 treatmenteffects on the growth of long-shoots (length anddiameter). For all statistical analyses, post-hoc, �adjusted general linear contrasts (Scheffe’s F)were used to test the significance of individualcontrasts that may have contributed to any sig-nificant main or interaction effects.
3. Results
3.1. En�ironmental conditions
The spring of 1998 was unusually wet and cool(Fig. 1) due to the influence of a strong SouthernOscillation El-Nino pattern in the SouthernPacific. Total precipitation from 1 October 1997(the end of the previous growing season) to 21May 1998 was 213 mm, 64.7 mm of which fellduring the study period (after 13 March Fig. 1);

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–10698
nearly twice the average annual precipitation atthis location on the NTS over the last decade (123mm; Hunter, 1994). Early morning air tempera-tures were consistently near or below 0 °Cthroughout the study period, with the coldesttemperature occurring the morning of 16 April(−7.3 °C), and the warmest maximum daytimetemperature in the afternoon of 20 April(27.5 °C). The average high temperature over thestudy period was 20.8 °C, with an average mini-mum of 0.2 °C. Volumetric soil moisture washigh at the start of the study (�=6.7�0.5 S.E. at0–20 cm and 7.90�0.56 S.E. at 0–50 cm), andwere still high in mid-June, two weeks after thelast sampling (�=8.2�0.59 S.E. at 0–20 cm and8.0�0.66 S.E. at 0–50 cm), and did not varysignificantly between CO2 treatments.
3.2. Ecophysiological responses
Midday water potentials (�mid) of L. andersoniiwere not significantly higher under elevated CO2
compared with ambient grown plants, though�mid levels were higher under elevated CO2 later inthe growing season, when water potentials showedsignificant declines (F=88.24; P�0.05; 6, 60 df;Fig. 2). Mid-day net photosynthetic rates in short-shoots and long-shoots did not vary between CO2
treatments, or display any CO2-by-time interac-
Fig. 2. Midday water potentials of Lycium andersonii at theNevada Desert FACE Facility. Each point is the mean ofthree measurements, bars indicate �1 S.E.M.
tion, but there was a significant CO2-by-shootinteraction (MS=49.31; F=3.54; P=0.068; 1,36 df). This was due to higher Anet in long-shootsunder elevated CO2 being lower than rates inambient-CO2 grown plants (Scheffe’s F=2.87;P=0.098), as well as to overall higher Anet inlong-shoots compared with short-shoots in ambi-ent CO2 plants (Scheffe’s F=3.19; P=0.082; Fig.3). Anet roughly followed a bell-shaped curve, withlow rates during early sampling (ca. 8.0 �molm−2 s−1 on 31 March), reaching seasonal highrates by 21 April (23 �mol m−2 s−1), then declin-ing to 12.6 �mol m−2 s−1 by the end of the studyin May. Pooled short-shoot and long-shoot stom-atal conductances to water vapor (gs) under ele-vated CO2 were 27% lower compared withambient-grown plants in both shoot types (F=5.99; P=0.07; 1, 4 df), with no higher orderinteractions (Fig. 3). Ci was consistently higher by51% in elevated CO2 plants (F=81.83; P=0.0008; 1, 4 df) compared with ambient CO2
counterparts (Fig. 3). Depressions in Ci on 21April accounted for the significant date effect(F=24.63; P�0.0001; 4, 36 df) due to relativelylow stomatal conductances when Anet was at aseasonal high (Fig. 3).
Mechanistic analyses of A/Ci curves showedthat L. andersonii altered the control of photosyn-
Fig. 1. Maximum and minimum temperatures and precipita-tion at the Nevada Desert FACE facility from 1 January to 15June 1998. Sampling of Lycium andersonii was done between19 March and 19 May 1998.

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106 99
thetic demand through the growing season (Table1). Pooled across the season and shoot types,light-saturated Rubisco carboxylation velocity(Vcmax) was 19% lower under elevated CO2 (41.6
�mol m−2 s−1) compared with Vcmax in ambientCO2 grown plants (51.2 �mol m−2 s−1). Corre-sponding electron transport rates (Jmax) werenearly 40% lower (118.1 mmol m−2 s−1 vs. 165.1
Fig. 3. Midday net assimilation rates (Anet), stomatal conductances to water vapor (gs) and internal CO2 concentrations (Ci) ofLycium andersonii growing under elevated (550 �mol mol−1) and ambient (360 �mol mol−1) CO2. Each point is the mean of threemeasurements; bars indicate �1 S.E.M.

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106100
Table 1Split-plot three-way repeated-measures ANOVA mean-square values of the light-saturated Rubisco carboxylation (Vcmax), electrontransport rate (Jmax), triose-phosphate utilization (TPU) and the ratio of Vcmax/Jmax
Vcmax Jmax TPUEffect Vc/Jdf
1091.6*** 26 461.0***Treat (T) 85.2*1 0.034**Error a 4 50.6 918.8 17.2 0.003
52.3ns 2998.8*Shoot (S) 101.4**1 0.006ns
3447.0*** 12 022.3***3 47.5**Date (D) 0.049**1T×S 625.7** 140.8ns 1.2ns 0.023*
472.1** 7347.2***T×D 37.9*3 0.008ns
38.1ns 218.9ns3 1.6nsS×D 0.003ns
3T×S×D 255.1* 2024.6* 12.8ns 0.004ns
Error b 28 106.8 839.6 14.6 0.007
Vcmax/Jmax mean-square values are for log-transformed data. Effect significant at *P�0.1, **P�0.05, ***P�0.01.
�mol m−2 s−1, for elevated and ambient CO2
grown plants, respectively). Triosphosphate uti-lization rates (TPU) were moderately lower underelevated CO2 (9.98 �mol m−2 s−1) comparedwith TPU in ambient CO2-grown plants (12.65�mol m−2 s−1).
CO2 treatment effects on Vcmax, Jmax and TPUshowed a significant interaction with date, and aninteraction with shoot type in the case of Vcmax
and the ratio of Vcmax to Jmax (Table 1). TheCO2-by-date interactions are due to greater pro-portional increases in ambient CO2 plants com-pared with elevated CO2 plants later in theseason, and are most clearly apparent in shortshoots (Fig. 4). The treatment-by-shoot interac-tion in Vcmax was due to significant CO2 treatmenteffects within short shoots, where Vcmax was 32%lower in elevated CO2 plants (Scheffe’s F=16.89;P�0.05), with Vcmax similar between treatmentsin long-shoots (Fig. 4). Additionally, under ele-vated CO2, long-shoots Vcmax were 14% higherthan in short shoots (Scheffe’s F=5.21; P�0.05),while Vcmax did not significantly differ betweenshort and long shoots in ambient CO2 grownplants (Fig. 4). The CO2-by-shoot interaction inVcmax/Jmax was due to significant CO2 effects inlong shoots (F=8.42; P�0.05), where late-sea-son increases in elevated CO2 plants were appar-ent (Fig. 5). The mildly significant three-wayinteractions in Vcmax and Jmax (Table 1) were dueto depressed rates in these parameters in longshoots under elevated CO2 that were apparent on
the 11 May sampling date, and to the strongerCO2 treatment differences in short shoots on thelast sampling date (Fig. 4).
Photosynthetic light response curves (Fig. 6)showed significant differences only in light-satu-rated photosynthetic rate (Amax) between CO2
treatments (MS=263.2; F=10.55; P�0.05; 1, 4df) and shoot type (MS=184.08; F=9.22; P�0.05; 1, 4 df), with no shoot-by-treatment interac-tion. L. andersonii growing under elevated CO2
had Amax 33% lower than in ambient CO2 grownplants (18.4 �mol m−2 s−1 vs. 27.8 �mol m−2
s−1), while Amax in long shoots (27.0 �mol m−2
s−1) was 41% higher than in short shoots (19.1�mol m−2 s−1; Fig. 6). Apparent quantum re-quirement (Qreq) was lower in ambient CO2 plants(13.0 mol mol−1�1.18 S.E.) compared withplants grown under elevated CO2 (18.9 molmol−1�2.79 S.E.), but this, as well as smalldifferences in dark respiration rate, was not sig-nificant at the P�0.1 level.
3.3. Plant growth
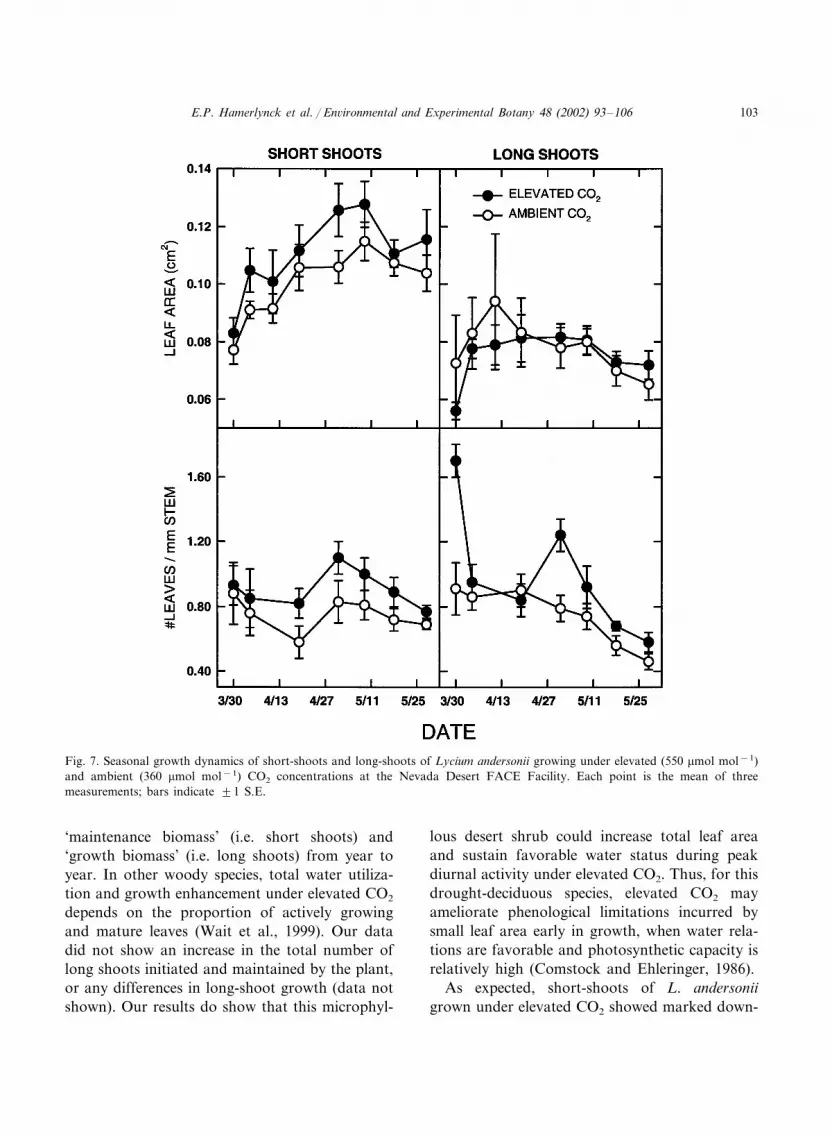
There was a significant CO2-by-shoot interac-tion in leaf area (F=4.19; P=0.045; 7, 60 df).Leaf areas of leaves on short-shoots were signifi-cantly higher in elevated CO2 plants (Scheffe’sF=4.17; P=0.0453), while leaf areas from long-shoot leaves, which were significantly smaller thanthose on short-shoots (F=85.5; P�0.0001), didnot differ between CO2 treatments (Fig. 7). There

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106 101
was also a mildly significant CO2-by-shoot inter-action (F=2.94; P=0.092; 1, 52 df) and CO2-by-date interaction (F=2.01; P=0.081; 6, 52 df) inleaf number per mm stem length. The CO2-by-
shoot interaction was due to significantly greaterleaves per stem length in long-shoots in elevatedCO2 grown plants (Scheffe’s F=10.92; P=0.0017) that was not apparent in ambient CO2
Fig. 4. Light-saturated Rubisco carboxylation velocity (Vcmax), electron transport rate (Jmax), and triose-phosphate utilization (TPU)of short- and long-shoots of Lycium andersonii growing under elevated (550 �mol mol−1) and ambient (360 �mol mol−1) CO2
concentrations. Each point is the mean of three measurements, bars indicate �one S.E. * indicate significant general linear contrasts(Scheffe’s F ; P�0.05) contributing to a significant higher order interaction (Table 1).

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106102
Fig. 5. Co-regulatory responses of Vcmax and Jmax in short-and long-shoots of Lycium andersonii grown under elevated(550 �mol mol−1) and ambient (360 �mol mol−1) CO2. Eachpoint is the mean of five measurements, error bars indicate�1 S.E.M.
marked reductions in both short- and long-shootstomatal conductance under elevated CO2 (Fig.3). The lower gs in elevated CO2 grown L. ander-sonii occurred under conditions of unusually highprecipitation and volumetric soil water contents(ca. 7–8%). This agrees with findings in otherMojave Desert plants, which show decreased gs
under elevated CO2 during moister conditions(Hamerlynck et al., 2000b; Pataki et al., 2000;Nowak et al., 2001). Concurrent with lower gs
was an increase in individual leaf area on shortshoots and by an initiation of new leaf cohorts,reflected in peaks in the number of leaves per unitstem length, especially in long shoots (Fig. 7).These suggest that L. andersonii could sustaingreater standing biomass during years favorablefor biomass accumulation. The degree to whichthis is met would depend on the proportions of
Fig. 6. Short-shoot and long-shoot photosynthetic light satura-tion response curves of L. andersonii growing under elevatedand ambient CO2 at the Nevada Desert FACE Facility. Eachbar is the mean of three measurements, bars indicate �1 S.E.
plants (Fig. 7). The CO2-by-date interactionreflected a significant increase in the number ofleaves per stem length in elevated CO2 plants inMay compared with values in March and April(Scheffe’s F=2.8; P=0.019) while ambient CO2
plants showed a continued decline in leaf numberper stem length (Fig. 7). In addition, there was asignificant date-by-shoot interaction (F=4.41;P=0.0011). Unlike short-shoots, long shoots hadsignificantly more leaves on 31 March and 2 Maycompared with the other dates pooled (Scheffe’sF=3.54; P=0.0049), and this was due solely tothe sharp increases in the number of leaves onlong-shoots of elevated CO2 plants (Fig. 7). De-spite the increase in the initiation of leaves onlong-shoots under elevated CO2, there were nosignificant CO2 direct or interaction effects on thenumber, length, or diameter of 1 and 2° long-shoots in L. andersonii (data not shown).
4. Discussion
Lycium andersonii only showed improved mid-day water potentials under elevated CO2 later inthe growing season (Fig. 2), accompanied by
E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106 103
Fig. 7. Seasonal growth dynamics of short-shoots and long-shoots of Lycium andersonii growing under elevated (550 �mol mol−1)and ambient (360 �mol mol−1) CO2 concentrations at the Nevada Desert FACE Facility. Each point is the mean of threemeasurements; bars indicate �1 S.E.
‘maintenance biomass’ (i.e. short shoots) and‘growth biomass’ (i.e. long shoots) from year toyear. In other woody species, total water utiliza-tion and growth enhancement under elevated CO2
depends on the proportion of actively growingand mature leaves (Wait et al., 1999). Our datadid not show an increase in the total number oflong shoots initiated and maintained by the plant,or any differences in long-shoot growth (data notshown). Our results do show that this microphyl-
lous desert shrub could increase total leaf areaand sustain favorable water status during peakdiurnal activity under elevated CO2. Thus, for thisdrought-deciduous species, elevated CO2 mayameliorate phenological limitations incurred bysmall leaf area early in growth, when water rela-tions are favorable and photosynthetic capacity isrelatively high (Comstock and Ehleringer, 1986).
As expected, short-shoots of L. andersoniigrown under elevated CO2 showed marked down-

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106104
regulation in photosynthetic demand comparedwith ambient grown plants (Fig. 4). However,these differences in photosynthetic demand be-tween elevated and ambient CO2 grown plantsincreased as water potential declined (Figs. 2 and4). This contrasts findings in other arid and semi-arid shrubs, which show reductions in down-regu-lation of assimilatory capacity with decreasingmidday water potential (Oechel et al., 1995; Hux-man et al., 1998; Hamerlynck et al., 2000a,b).However, even by the last measurement period,after which there was widespread leaf senescenceacross all study plots, midday water potentialswere relatively high (Fig. 3), especially for a desertshrub (Jones, 1984; Comstock and Ehleringer,1986; Hamerlynck et al., 2000a, 2002), as werevolumetric soil water contents. Thus, the declinesin �midday likely reflect higher transpirationallosses accompanying canopy development (Fig. 7)and warmer temperatures (Fig. 1). These findingsalso suggest there may be controls to leaflongevity that initiate leaf shedding even underfavorable, well-watered conditions in L. ander-sonii. Indeed, early-spring cold temperaturesstrongly limited flowering, which was apparent inonly two of our test plants, both of which abortedtheir seeds (data not shown). Possibly, this curtail-ment of reproductive effort facilitated leafshedding.
Contrary to our hypotheses, down-regulationwas apparent in short-shoots while long-shootswere actively growing, as well as in long-shoots,even early in development (Fig. 4). There weremarked differences in how L. andersonii balancedphotosynthetic down-regulation in different shoottypes (Fig. 5). In short-shoots, Vcmax and Jmax
co-varied through the season, resulting in similarseasonal Vcmax/Jmax ratios between CO2 treat-ments (Fig. 5). In long-shoots, elevated CO2
grown plants showed increases in the contributionof Vcmax compared with Jmax, which was relativelyinvariant (Fig. 5). This differs from shade-adaptedwoody species, where Jmax under elevated CO2 isfrequently up-regulated compared with Vcmax
(DeLucia and Thomas, 2000). Thus, the capacityto regenerate RuBP in this high-light adaptedspecies is reduced under elevated CO2 in activelygrowing tissue. This is further supported by pat-
terns of triose-phosphate utilization, which weremarkedly higher in long-shoots, and was signifi-cantly lower under elevated CO2 (Fig. 4). Thesefindings suggest a greater limitation due to regen-eration of substrates for photosynthetic enzymeintermediates, as found in other C3 species(Harley and Sharkey, 1991; Harley et al., 1992).
These findings, coupled with lower levels oflight-saturated net assimilation rates estimatedfrom light-saturation curves (Fig. 6) are stronglysuggestive of reduced allocation to photosyntheticelectron transport capacity in this deciduousshrub. This seems to be most apparent in activelygrowing long-shoots, as evidenced by increasedVcmax/Jmax ratios (Fig. 5), and by greater propor-tional reductions in light saturation responses(Fig. 6). Possibly, the greater variation in elevatedCO2 L. andersonii photosynthetic light responses(Fig. 6) may be due to irregular leaf ages follow-ing initiation of new leaf cohorts (Fig. 7). It seemsthe balance of limiting factors to photosynthesisin growing leaves resulted in increased relativeefficiency of Rubisco. Indeed, Vcmax in elevatedCO2 grown long-shoots was significantly higherthan in ambient CO2 counterparts by the end ofthe study, while Jmax levels were similar (Fig. 4).Greater reliance on regulation of enzymatic effi-ciency and reduction in Jmax resulted in similarAnet at lower gs and higher internal CO2 concen-trations in L. andersonii under elevated CO2 (Fig.3).
In summary, we found that elevated CO2 elic-ited marked down-regulation in photosyntheticcapacity in two functionally different canopycomponents of this drought-deciduous shrub.Photosynthetic down-regulation resulted in simi-lar levels of instantaneous gas exchange withgreater leaf growth, but lower gs maintained simi-lar, favorable midday water status. Together,these findings suggest that drought-deciduousshrubs may require less resources per unit leafarea for processes needed to sustain positive car-bon assimilation rates when moisture conditionsare favorable, which would allow them to bettercope with the highly variable precipitation andtemperature regimes that characterize NorthAmerican warm desert systems (Smith et al.,1997).

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106 105
Acknowledgements
This research was supported by a NSF/TECO(IBN 9524036) grant to SDS, a DOE/TCP grantfor support of the Nevada Desert FACE Facility,and a DOE Traineeship to TEH. Additional oper-ations support to the NDFF was provided by theDOE–Nevada Operations Office and BechtelNevada.
References
Comstock, J.P., Ehleringer, J.R., 1986. Canopy dynamicsand carbon gain in response to soil moisture availabilityin Encelia frutescens Gray, a drought deciduous shrub.Oecologia 68, 271–278.
DeLucia, E.H., Thomas, R.B., 2000. Photosynthetic re-sponses to CO2 enrichment of four hardwood species ina forest understory. Oecologia 122, 11–19.
Ehleringer, J.R., 1994. Variation in gas exchange characteris-tics among desert plants. In: Schulze, E.-D., Caldwell,M.M. (Eds.), Ecophysiology of Photosynthesis. Springer,Berlin, pp. 361–392.
Hamerlynck, E.P., Huxman, T.E., Nowak, R.S., Reader, S.,Loik, M.E., Jordan, D.N., Zitzer, S.F., Coleman, J.S.,Seemann, J.R., Smith, S.D., 2000a. Photosynthetic re-sponses of Larrea tridentata to a step-increase in atmo-spheric CO2 at the Nevada Desert FACE Facility. J.Arid. Environ. 44, 425–436.
Hamerlynck, E.P., Huxman, T.E., Loik, M.E., Smith, S.D.,2000b. Effects of extreme high temperature, drought, andelevated CO2 on photosynthesis of Larrea tridentata, aMojave Desert evergreen. Plant Ecol. 148, 185–195.
Hamerlynck, E.P., McAuliffe, J.R., McDonald, E.V., Smith,S.D., 2002. Ecological responses of two Mojave Desertshrubs to soil horizon development and soil water dy-namics. Ecology 83, 768–779.
Harley, P.C., Sharkey, T.D., 1991. An improved model ofC3 photosynthesis at high CO2: Reversed O2 sensitivityexplained by lack of glycerate re-entry into the chloro-plast. Photosyn. Res. 27, 169–178.
Harley, P.C., Thomas, R.B., Reynolds, J.F., Strain, B.R.,1992. Modeling photosynthesis of cotton grown in ele-vated CO2. Plant Cell Environ. 15, 271–282.
Hunter, R.B., 1994. Status of the Flora and Fauna on theNevada Test Site, 1993. DOE/NV/11432-195 UC–721.US Department of Commerce, Springfield, VA, USA.
Huxman, T.E., Smith, S.D., 2001. Photosynthesis in an inva-sive grass and native forb at elevated CO2 during an ElNino year in the Mojave Desert. Oecologia 128, 193–301.
Huxman, T.E., Hamerlynck, E.P., Moore, B.d., Smith, S.D.,Jordan, D.N., Zitzer, S.F., Nowak, R.S., Coleman, J.S.,Seemann, J.R., 1998. Photosynthetic down-regulation in
Larrea tridentata exposed to elevated atmospheric CO2:Interaction with drought under glasshouse and field(FACE) exposure. Plant Cell Environ. 21, 1153–1161.
Jones, C.S., 1984. The effect of axis splitting on xylem pres-sure potentials and water relations in the desert shrub,Ambrosia dumosa (Gray) Payne (Asteraceae). Bot. Gaz.145, 125–131.
Jordan, D.N., Zitzer, S.F., Hendrey, G.R., Lewin, K.F.,Nagy, J., Nowak, R.S., Smith, S.D., Coleman, J.S., See-mann, J.R., 1999. Biotic, abiotic and performance as-pects of the Nevada Desert Free-Air CO2 Enrichment(FACE) Facility. Global Change Biol. 5, 659–668.
Knapp, A.K., Hamerlynck, E.P., Ham, J.M., Owensby,C.E., 1996. Responses in stomatal conductance to ele-vated CO2 in 12 grassland species that differ in growthform. Vegetatio 125, 31–41.
Lewis, J.D., Tissue, D.T., Strain, B.R., 1996. Seasonal re-sponse of photosynthesis to elevated CO2 in loblolly pine(Pinus tadea L.) over two growing seasons. GlobalChange Biol. 2, 103–114.
Nowak, R.S., DeFalco, L.A., Wilcox, C.S., Jordan, D.N.,Coleman, J.S., Seemann, J.R., Smith, S.D., 2001. Leafconductance decreased under free-air CO2 enrichment(FACE) for three perennials in the Nevada desert. NewPhytol. 150, 449–458.
Oechel, W.C., Hastings, S.J., Vourlitis, G.L., Jenkins, M.A.,Hinkson, C.L., 1995. Direct effects of elevated CO2 inchaparral and Mediterranean-type ecosystems. In:Moreno, J., Oechel, W. (Eds.), Global Change and Med-iterranean-type Ecosystems, Ecological Studies, vol. 117.Springer–Verlag, Berlin, pp. 58–75.
Pataki, D.E., Huxman, T.E., Jordan, D.N., Zitzer, S.F.,Coleman, J.S., Smith, S.D., Nowak, R.S., Seemann, J.R.,2000. Water use of two Mojave Desert shrubs underelevated CO2. Global Change Biol. 6, 889–898.
Sage, R.F., 1994. Acclimation of photosynthesis to increas-ing atmospheric CO2: the gas exchange perspective. Pho-tosyn. Res. 39, 351–368.
Smith, S.D., Nobel, P.S., 1986. Deserts. In: Baker, N.R.,Long, S.P. (Eds.), Photosynthesis in Contrasting Envi-ronments. Topics in Photosynthesis 7. Elsevier, Amster-dam, pp. 13–62.
Smith, S.D., Monson, R.K., Anderson, J.E., 1997. Physio-logical Ecology of North American Desert Plants.Springer, Berlin.
Smith, S.D., Huxman, T.E., Zitzer, S.F., Charlet, T.N.,Housman, D.C., Coleman, J.S., Fenstermaker, L.K., See-mann, J.R., Nowak, R.S., 2000. Elevated CO2 increasesproductivity and invasive species success in an aridecosystem. Nature 408, 79–82.
Stirling, C.M., Davey, P.A., Williams, T.G., Long, S.P.,1997. Acclimation of photosynthesis to elevated CO2 andtemperature in five British native species of contrastingfunctional type. Global Change Biol. 3, 237–246.
Thomas, R.B., Strain, B.R., 1991. Root restriction as a fac-

E.P. Hamerlynck et al. / En�ironmental and Experimental Botany 48 (2002) 93–106106
tor in photosynthetic acclimation of cotton seedlingsgrown in elevated carbon dioxide. Plant Physiol. 96,627–634.
Wait, D.A., Jones, C.G., Wynn, J., Woodward, F.I., 1999.The fraction of expanding to expanded leaves determines
the biomass response of Populus to elevated CO2. Oe-cologia 121, 193–200.
Von Caemmerer, S., Farquhar, G.D., 1981. Some relation-ships between the biochemistry of photosynthesis and thegas exchange rates of leaves. Planta 153, 376–387.