effects of aripiprazole and terguride on dopamine synthesis in the dorsal striatum and medial...
TRANSCRIPT

J Neural Transm (2008) 115: 97–106
DOI 10.1007/s00702-007-0820-7
Printed in The Netherlands
Effects of aripiprazole and terguride on dopamine synthesis in the dorsalstriatum and medial prefrontal cortex of preweanling rats
S. D. Iniguez, A. M. Cortez, C. A. Crawford, S. A. McDougall
Department of Psychology, California State University, San Bernardino, CA, USA
Received 19 June 2007; Accepted 1 September 2007; Published online 12 November 2007
# Springer-Verlag 2007
Summary. The purpose of this study was to determine whether aripipra-
zole, a D2-like partial agonist increasingly prescribed to children, alters DAsynthesis via actions at autoreceptors in the dorsal striatum and medial
prefrontal cortex (mPFC) of preweanling rats. The ability of dopaminergic
agents to alter DOPA accumulation in the striatum and mPFC was measuredafter NSD-1015 on postnatal day (PD) 20. Dopaminergic tone was manipu-
lated by administering reserpine, g-butyrolactone (GBL), or through am-
phetamine withdrawal. Results showed that the partial agonists aripiprazole
and terguride increased striatal DOPA accumulation under normosensitiveconditions, but decreased DOPA accumulation in states of low dopaminer-
gic tone. A different pattern of results was observed in the mPFC, because
terguride and haloperidol, but not aripiprazole, increased DOPA accumula-
tion under normosensitive conditions. In conclusion, the present data showthat aripiprazole affects striatal synthesis modulating autoreceptors in an
adult-typical manner during the late preweanling period. Unlike in adult
rats, however, the mPFC of preweanling rats appears to contain transitory
synthesis modulating autoreceptors that are sensitive to drug manipulation.
Keywords: Aripiprazole; terguride; DOPA accumulation; autoreceptors;ontogeny; dorsal striatum; prefrontal cortex
Introduction
Dopamine (DA) partial agonist drugs have interesting clin-
ical profiles. Initially employed as anti-parkinsonian agents
(Corsini et al. 1985; Brucke et al. 1987; Critchley and
Parkes 1987) D2-like partial agonists, such as aripiprazole,
are commonly prescribed to adults for schizophrenia and
bipolar disorder (Keck et al. 2003; Davies et al. 2004) and
are a potential pharmacotherapy for psychostimulant abuse
(Pulvirenti and Koob 1994; Platt et al. 2003; Feltenstein
et al. 2007). In terms of schizophrenia and drug abuse, the
therapeutic benefits of aripiprazole presumably result from
stabilizing DA functioning at both pre- and postsynaptic
receptors (Burris et al. 2002; Li et al. 2004; Feltenstein
et al. 2007). An even more recent trend is to prescribe
D2-like partial agonists to pediatric populations. In the
United States, aripiprazole is used to treat various childhood
and adolescent disorders, including Tourette syndrome, per-
vasive developmental disorder, conduct disorder, and pedi-
atric bipolar disorder (Findling et al. 2003; Barzman et al.
2004; Stigler et al. 2004; Biederman et al. 2005; Murphy
et al. 2005; Stachnik and Nunn-Thompson 2007). This in-
creased usage is occurring even though D2-like partial
agonists have seldom been examined using developmental
animal models and results from a minority of human clini-
cal studies suggest caution in prescribing aripiprazole to
children (Rugino and Janvier 2005).
Studies employing adult animal models have established
that D2-like partial agonists exhibit low intrinsic activity
(i.e., function as antagonists) at normosensitive DA presyn-
aptic receptors. In this circumstance, terguride and pre-
clamol increase dorsal striatal DA synthesis and reverse
the quinpirole-induced inhibition of DOPA accumulation
(Hjorth et al. 1983; Kehr 1984; Clark et al. 1991; Svensson
et al. 1991, 1993). In addition to their antagonistic proper-
ties, D2-like partial agonists are capable of showing high
intrinsic activity at synthesis modulating autoreceptors of
adult rats, but only after manipulations that reduce dopa-
minergic tone. For example, D2-like partial agonists inhib-
it, rather than enhance, dorsal striatal dopamine synthesis
in reserpinized rats (Hjorth et al. 1983, 1988; Svensson
Correspondence: Sanders A. McDougall, Department of Psychology,
California State University, 5500 University Parkway, San Bernardino,CA 92407, USA
e-mail: [email protected]

et al. 1991; Kikuchi et al. 1995). Pretreatment with the
nerve impulse inhibitor g-butyrolactone (GBL) also pro-
duces a state of low dopaminergic tone in which aripipra-
zole and terguride inhibit DA synthesis (Hjorth et al. 1983;
Clark et al. 1985a; Svensson et al. 1991; Kikuchi et al.
1995; Oshiro et al. 1998). In adult rats, therefore, biochem-
ical evidence indicates that D2-like partial agonists exhibit
low intrinsic activity at normosensitive presynaptic recep-
tors, while showing high intrinsic activity at the same re-
ceptors under conditions of reduced dopaminergic tone.
Few studies have examined the effects of D2-like partial
agonists in developing animals even though aripiprazole is
commonly administered to pediatric populations. Available
evidence suggests that terguride, like haloperidol, increases
dorsal striatal DA synthesis of preweanling rats under nor-
mosensitive conditions (Yoshida et al. 2006). Conversely,
terguride significantly reduces dorsal striatal DA synthesis
after either one or five days of reserpine treatment (Yoshida
et al. 2006). These agonistic effects at synthesis modulating
autoreceptors are short-lived, because terguride only in-
creases DA synthesis when measured 5 h and 24 h after
conclusion of a 5-day reserpine regimen (Farley et al.
2006). Terguride also decreases striatal DA synthesis after
GBL treatment, an effect that is antagonized by haloperidol
(Yoshida et al. 2006). In conclusion, results suggest that
terguride affects dorsal striatal D2-like receptors of pre-
weanling and adult rats in a generally similar manner,
although other brain regions and more therapeutically rele-
vant partial agonist compounds (e.g., aripiprazole) have not
been examined.
The purpose of the present study was to further examine
the effects of D2-like partial agonists on the synthesis mod-
ulating autoreceptors of preweanling rats during states of
high and low dopaminergic tone. Low dopaminergic tone
was induced by reserpine, GBL, or by administering an
escalating amphetamine regimen that is purported to cause
a withdrawal state after drug discontinuation (Barr and
Phillips 1999; Orsini et al. 2001). Because low, but not
high, doses of aripiprazole have been reported to increase
DA efflux in the medial prefrontal cortex (mPFC) of adult
rats, we also examined the effects of various doses of ari-
piprazole and terguride on DA synthesis in the dorsal stria-
tum and mPFC of rats on postnatal day (PD) 20. The mPFC
was of special interest, because (a) increasing DA neuro-
transmission in the mPFC is of presumed therapeutic
benefit (Jordan et al. 2004; Zocchi et al. 2005) and (b)
D2-like synthesis modulating autoreceptors in the mPFC
exist in functional quantities during the preweanling period
(Andersen et al. 1997), but are either absent or occur in
greatly reduced numbers during adulthood (Bannon et al.
1981; Galloway et al. 1986; but see Fadda et al. 1984). In
all experiments, rats were tested on PD 20 because this age
corresponds with ‘‘childhood’’ in humans (Andersen 2003).
Materials and methods
Subjects
Preweanling rats (n! 221) of Sprague-Dawley descent (Charles River,
Hollister, CA, USA) were obtained from the breeding colony at California
State University, San Bernardino (CSUSB). Litters were culled to ten pupsat postnatal day (PD) 3 (day of parturition is PD 0). Rat pups were kept with
the dam except when undergoing experimental manipulation. The colony
room was maintained at 21–23"C and kept under a 12 h dark=light cycle.Subjects were treated according to the ‘‘Principles of laboratory animalcare’’ (NIH publication no. 86-23, revised 1985) under a research protocol
approved by the Institutional Animal Care and Use Committee of CSUSB.
Drugs
R(#)-Terguride, haloperidol, and reserpine were dissolved in a minimal
amount of glacial acetic acid and diluted with saline; whereas, D-amphet-
amine sulfate salt, quinpirole, and 3-hydroxybenzylhydrazine hydrochloride
(NSD-1015) were dissolved in saline. Aripiprazole was dissolved in (2-hydropropyl)-b-cyclodextrin solution (HBC, 45% (w=v) solution in water).
g-Butyrolactone (GBL) was at an initial density of 1.12 g=ml. All drugs
were injected intraperitoneally (i.p.) at a volume of 5ml=kg. With the
exception of aripiprazole (Toronto Research Chemicals, Toronto, Canada),all drugs were purchased from Sigma (St. Louis, MO, USA).
Experimental protocols
Experiment 1: Effects of D2-like partial agonists on DOPA
accumulation after saline, amphetamine, or reserpine pretreatment
Rats (n! 103) were randomly assigned to one of three pretreatment condi-
tions: amphetamine withdrawal, reserpine, or saline controls. To induce a
state of amphetamine withdrawal, rats were injected with an escalating
regimen of amphetamine (starting at 1mg=kg and increasing by 1 unitintervals until 10mg=kg was administered) beginning on PD 16 (see also
Barr and Phillips 1999; Orsini et al. 2001). More specifically, on PD 16-PD
18 rats were injected three times per day (at 800, 1600, and 2400 h) with
amphetamine. The initial dose of amphetamine was 1mg=kg, with eachsubsequent dose of amphetamine increasing by 1mg=kg. On PD 19, rats
received their tenth and last injection of vehicle or amphetamine (in this
case, 10mg=kg) at 800h. Rats in the reserpine condition, received a singleinjection of reserpine at 800h on PD 19. Half of the rats in the saline
condition received a single injection of saline at 800 h on PD 19 (i.e., at
the same time as rats in the reserpine condition), while the other half of the
control rats were given 10 injections of saline on PD 16-PD 20 (i.e., at thesame time as rats in the amphetamine withdrawal condition).
After 24 h (i.e., on PD 20), rats from each pretreatment condition were
further subdivided (n! 5–7 per group) and injected with saline vehicle,
HBC vehicle, quinpirole (0.5mg=kg, i.p.), haloperidol (0.5mg=kg, i.p.),terguride (1.5mg=kg, i.p.), or aripiprazole (3 or 10mg=kg, i.p.) fol-
lowed, 30min later, by an injection of the DOPA decarboxylase inhibitor
NSD-1015 (100mg=kg, i.p.). Rats were killed by rapid decapitation
30min after NSD-1015 treatment and their dorsal striata and prefrontalcortices were removed on an ice-cold dissection plate and stored at
$80"C until assay.
To determine whether amphetamine withdrawal altered DA levels, anadditional 48 rats were injected with an escalating regimen of amphetamine
98 S. D. Iniguez et al.

or saline in the manner described previously. Rats were killed by rapiddecapitation 12, 24, 48, or 72 h after the final amphetamine (n! 8 per
group) or saline (n! 4 per group) injection. Dorsal striata were removed
and stored at $80"C until assay.
Experiment 2: Low-dose effects of aripiprazole on DOPA
accumulation after saline or reserpine pretreatment
On PD 19, rats (n! 47) received a single injection of reserpine or saline at800 h. After 24 h (i.e., on PD 20), rats from each pretreatment condition
were further subdivided (n! 7–8 per group) and injected with saline vehi-
cle, HBC vehicle, terguride (1.5mg=kg, i.p.), or aripiprazole (0.1, 0.3, 3 or10mg=kg, i.p.) followed, 30min later, by an injection of the DOPA decar-
boxylase inhibitor NSD-1015 (100mg=kg, i.p.). The doses of aripiprazole
were based on previous studies (Li et al. 2004; Zocchi et al. 2005). Rats
were killed by rapid decapitation 30min after NSD-1015 treatment and theirdorsal striata and prefrontal cortices were removed on an ice-cold dissection
plate and stored at $80"C until assay.
Experiment 3: Effects of D2-like partial agonists on DOPA
accumulation after GBL treatment
On PD 20, rats (n! 71) were injected with saline vehicle, HBC vehicle,quinpirole (0.5mg=kg, i.p.), haloperidol (0.5mg=kg, i.p.), terguride (0.005,0.015, or 1.5mg=kg, i.p.), or aripiprazole (0.1, 0.3, or 10mg=kg, i.p.)
(n! 6–8 per group) followed, 25min later, by an injection of the nerve
impulse inhibitor GBL (625mg=kg, i.p.). After 5min, rats were injectedwith NSD-1015 (100mg=kg, i.p.). Rats were killed by rapid decapitation
30min after NSD-1015 treatment and their dorsal striata and prefrontal
cortices were removed and stored at $80"C until assay.
HPLC conditions
DOPA accumulation and DA content were determined using high-perfor-
mance liquid chromatography (HPLC). Frozen tissue samples were sonicat-ed in 0.1N HClO4 (dorsal striatum, 10 volumes; mPFC, 5 volumes) and
centrifuged at 20,000g for 30min at 4"C. The supernatant was then filtered
through a 0.22mm centrifugation unit at 2000g for 5min at 4"C. Twenty
microliters of the resulting extract was assayed for DOPA or DA using aBreezeTM HPLC system (Waters, Milford, MA, USA), with a Coulochem II
electrochemical detector (ESA, Chelmsford, MA, USA). For dorsal striatal
samples, the mobile phase consisted of 75mM NaH2PO4, 1.4mM 1-octane
sulfonic acid, 10mM ethylenediamine tetraacetic acid (EDTA), and 10%acetonitrile (pH 3.1). The mobile phase for mPFC samples was adjusted to
75mMNaH2PO4, 0.75mM 1-octane sulfonic acid, 10mMEDTA, and 1.5%
acetonitrile (pH 3.1) to increase peak latency. Both mobile phases were
pumped at a rate of 0.5ml=min.
Statistical evaluation
Assignment of subjects was random, with an equal number of male and
female rats being placed in each group. Individual groups included no morethan one subject from a particular litter and, when possible (i.e., in experi-
ments with ten or fewer groups), each litter provided one subject per group.
In the initial experiments, DOPA accumulation data (ng=mg wet weight
tissue) were analyzed using two-way analyses of variance (ANOVA)(Pretreatment Condition % Drug Group) and further analyzed using sepa-
rate one-way ANOVAs for each pretreatment condition. In the GBL experi-
ment, DOPA accumulation data were analyzed using a one-way randomized
block ANOVA, with litter serving as the blocking factor (Hughes 1979). Forpresentation purposes, DOPA accumulation data were converted to percent
of vehicle controls (see also Baldessarini et al. 1994; Clark et al. 1991).
Saline and HBC vehicle had similar effects on DOPA accumulation, there-
fore data from the two vehicle groups were combined for each experiment.Preliminary statistical analyses showed that DOPA accumulation data did
not vary according to sex of the animals, so the sex variable was excluded
from later analyses. Post hoc analysis of neurochemical data was made
using Newman-Keuls tests (p<0.05).
Results
Experiment 1: Effects of D2-like partial agonists
on DOPA accumulation after saline, amphetamine,
or reserpine pretreatment
Overall, the dopaminergic compounds differentially af-
fected dorsal striatal DOPA accumulation depending on
pretreatment condition (see Fig. 1) [F(10,85)! 13.08,
p<0.001]. In the saline pretreatment condition, quinpirole
(0.5mg=kg) caused a significant reduction in striatal DOPAaccumulation relative to vehicle controls, whereas haloper-
idol (0.5mg=kg) increased DOPA accumulation (see upper
graph, Fig. 1) [F(5,32)! 160.42, p<0.001]. Both D2-like
partial agonists increased striatal DOPA accumulation, but
the magnitude of the increase was less for terguride (217%
increase relative to vehicle controls) and aripiprazole
(3mg=kg: 260%; 10mg=kg: 208%) than for the full an-
tagonist haloperidol (427%). Rats in the amphetamine
withdrawal condition responded essentially the same as rats
in the saline pretreatment condition, because terguride and
aripiprazole caused a significant increase in DOPA ac-
cumulation that was approximately half as robust as the
haloperidol-induced increase (see middle graph, Fig. 1)
[F(5,26)! 74.56, p<0.001]. The quinpirole-induced re-
duction in striatal DOPA was only marginally significant
[p<0.07], but quinpirole did cause significantly less stria-
tal DOPA accumulation than the partial agonists. Unlike
amphetamine withdrawal, reserpine produced a state in
which aripiprazole and terguride caused agonist-like de-
creases in striatal DOPA accumulation (see lower graph,
Fig. 1). Specifically, terguride and the higher dose of ari-
piprazole (10mg=kg) reduced striatal DOPA accumulation
relative to vehicle controls [F(5,27)! 81.30, p<0.001].
Reserpine did not cause a qualitatively different pattern
of effects in rats given direct acting compounds, because
haloperidol increased, while quinpirole decreased, DOPA
accumulation in reserpinized rats. Therefore, in saline- and
amphetamine-pretreated rats aripiprazole and terguride
caused antagonist-like increases in striatal DOPA accumula-
tion, but in reserpinized rats the partial agonists functioned
in the same manner as the D2-like agonist quinpirole.
In the mPFC, the DA agonists and antagonists produced
different patterns of effects according to pretreatment con-
dition (see Fig. 2) [F(5,84)! 2.67, p<0.01]. In the saline
Effects of D2-like partial agonists on DA synthesis 99

pretreatment condition, both haloperidol and terguride in-
creased DOPA accumulation in the mPFC (see upper
graph, Fig. 2) [F(1,28)! 4.94, p<0.01]. Interestingly, at
neither of the doses tested did aripiprazole (3 or 10mg=kg)enhance DOPA accumulation in the mPFC, while quinpi-
role caused only a nonsignificant decline in DOPA values
(60% of controls) [p< 0.1]. A different pattern of effects
was evident during amphetamine withdrawal, because ter-
guride-treated rats exhibited greater DOPA accumulation
than haloperidol- and vehicle-treated rats (see middle
graph, Fig. 2) [F(1,27)! 3.95, p<0.01]. No differences
in DOPA accumulation were observed in the reserpine con-
dition, since the various DA compounds did not alter mPFC
DOPA accumulation relative to vehicle control values (see
lower graph, Fig. 2).
In order to determine whether DA levels were reduced
during amphetamine withdrawal, striatal DA content of
saline- and amphetamine-pretreated rats was measured
50
00.5 1.5 3 10 0.5
Fig. 2. Mean mPFC DOPA accumulation of PD 20 rats injected with
vehicle, haloperidol (Hal), terguride (Terg), aripiprazole (Aripip), or quin-pirole (Quin) 24 h after conclusion of saline, amphetamine, or reserpine
pretreatment. Data are expressed as percent of vehicle controls (&SEM)
and doses are in mg=kg. 'p<0.05 vs vehicle controls (indicated by the
dashed line); yp<0.05 vs haloperidol
Fig. 1. Mean striatal DOPA accumulation of PD 20 rats injected with
vehicle, haloperidol (Hal), terguride (Terg), aripiprazole (Aripip), or quin-pirole (Quin) 24 h after conclusion of saline, amphetamine, or reserpine
pretreatment. Data are expressed as percent of vehicle controls (&SEM)
and doses are in mg=kg. 'p<0.05 vs vehicle controls (indicated by the
dashed line); yp<0.05 vs haloperidol; zp<0.05 vs quinpirole
100 S. D. Iniguez et al.
12, 24, 48, or 72 h after final injection. As can be seen in
Table 1, amphetamine withdrawal did not alter striatal DA
levels at any time point.
Experiment 2: low-dose effects of aripiprazole on DOPA
accumulation after saline or reserpine pretreatment
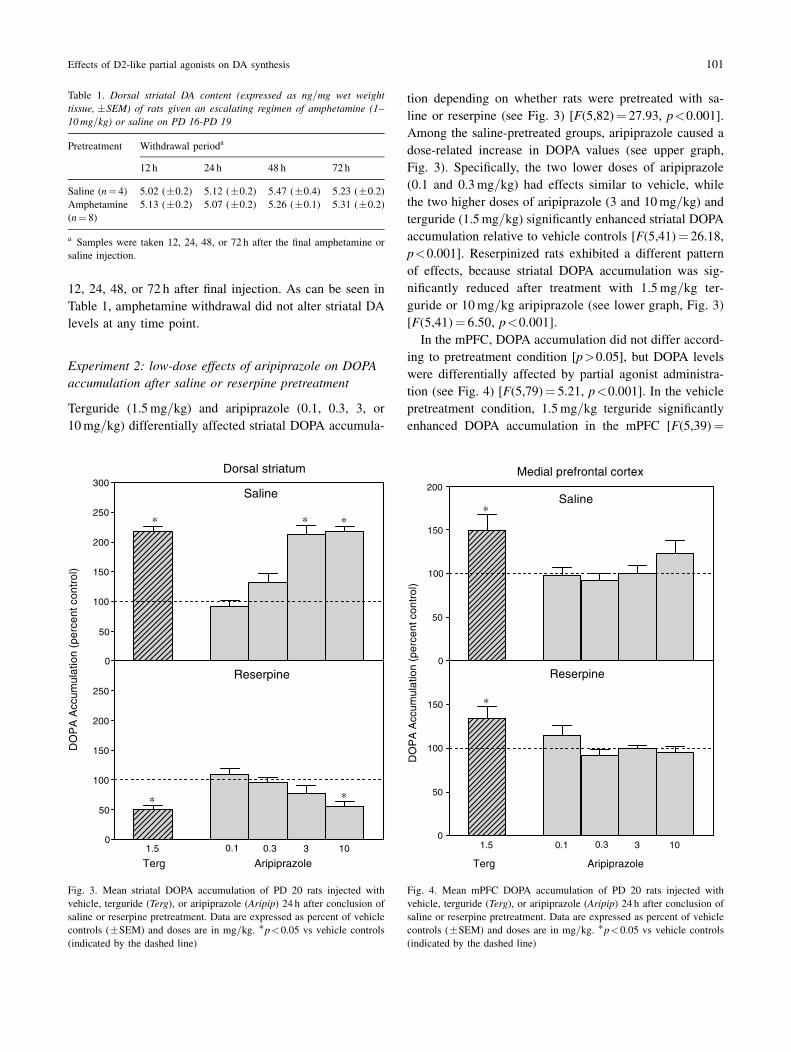
Terguride (1.5mg=kg) and aripiprazole (0.1, 0.3, 3, or
10mg=kg) differentially affected striatal DOPA accumula-
tion depending on whether rats were pretreated with sa-
line or reserpine (see Fig. 3) [F(5,82)! 27.93, p<0.001].
Among the saline-pretreated groups, aripiprazole caused a
dose-related increase in DOPA values (see upper graph,
Fig. 3). Specifically, the two lower doses of aripiprazole
(0.1 and 0.3mg=kg) had effects similar to vehicle, while
the two higher doses of aripiprazole (3 and 10mg=kg) andterguride (1.5mg=kg) significantly enhanced striatal DOPA
accumulation relative to vehicle controls [F(5,41)! 26.18,
p<0.001]. Reserpinized rats exhibited a different pattern
of effects, because striatal DOPA accumulation was sig-
nificantly reduced after treatment with 1.5mg=kg ter-
guride or 10mg=kg aripiprazole (see lower graph, Fig. 3)
[F(5,41)! 6.50, p<0.001].
In the mPFC, DOPA accumulation did not differ accord-
ing to pretreatment condition [p>0.05], but DOPA levels
were differentially affected by partial agonist administra-
tion (see Fig. 4) [F(5,79)! 5.21, p<0.001]. In the vehicle
pretreatment condition, 1.5mg=kg terguride significantly
enhanced DOPA accumulation in the mPFC [F(5,39)!
Table 1. Dorsal striatal DA content (expressed as ng=mg wet weighttissue, &SEM) of rats given an escalating regimen of amphetamine (1–
10mg=kg) or saline on PD 16-PD 19
Pretreatment Withdrawal perioda
12 h 24 h 48 h 72 h
Saline (n! 4) 5.02 (&0.2) 5.12 (&0.2) 5.47 (&0.4) 5.23 (&0.2)
Amphetamine(n! 8)
5.13 (&0.2) 5.07 (&0.2) 5.26 (&0.1) 5.31 (&0.2)
a Samples were taken 12, 24, 48, or 72 h after the final amphetamine orsaline injection.
Fig. 3. Mean striatal DOPA accumulation of PD 20 rats injected with
vehicle, terguride (Terg), or aripiprazole (Aripip) 24 h after conclusion ofsaline or reserpine pretreatment. Data are expressed as percent of vehicle
controls (&SEM) and doses are in mg=kg. 'p<0.05 vs vehicle controls
(indicated by the dashed line)
Fig. 4. Mean mPFC DOPA accumulation of PD 20 rats injected with
vehicle, terguride (Terg), or aripiprazole (Aripip) 24 h after conclusion ofsaline or reserpine pretreatment. Data are expressed as percent of vehicle
controls (&SEM) and doses are in mg=kg. 'p<0.05 vs vehicle controls
(indicated by the dashed line)
Effects of D2-like partial agonists on DA synthesis 101
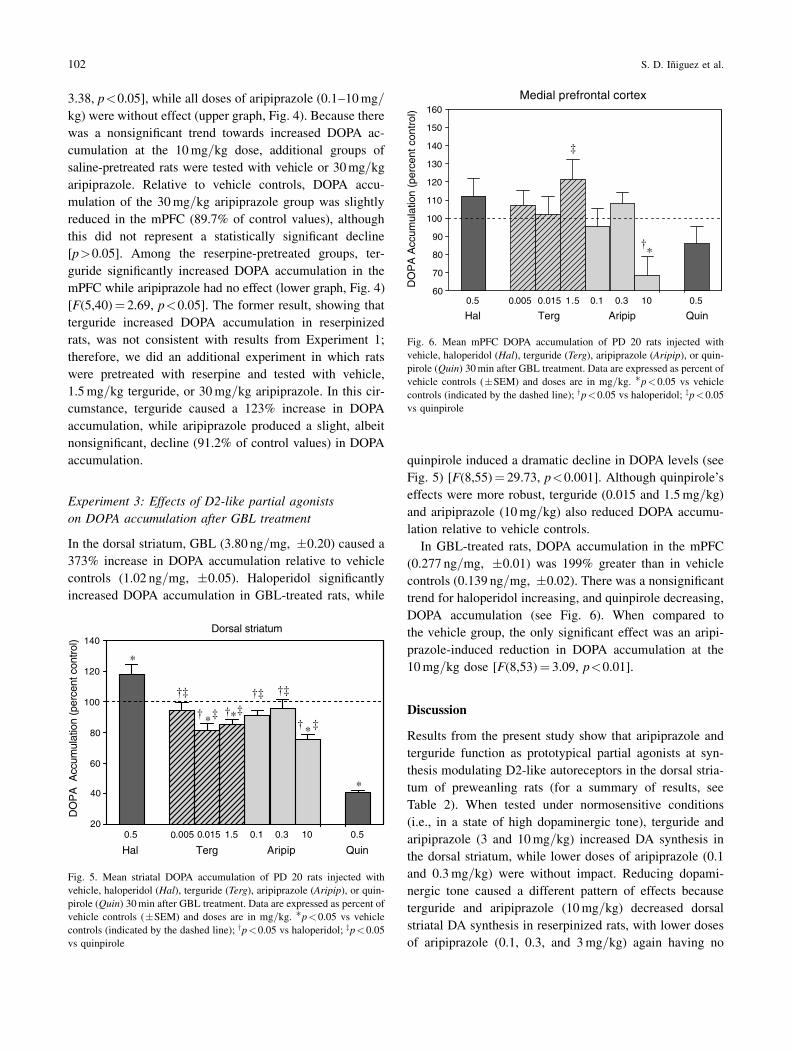
3.38, p<0.05], while all doses of aripiprazole (0.1–10mg=kg) were without effect (upper graph, Fig. 4). Because there
was a nonsignificant trend towards increased DOPA ac-
cumulation at the 10mg=kg dose, additional groups of
saline-pretreated rats were tested with vehicle or 30mg=kgaripiprazole. Relative to vehicle controls, DOPA accu-
mulation of the 30mg=kg aripiprazole group was slightly
reduced in the mPFC (89.7% of control values), although
this did not represent a statistically significant decline
[p>0.05]. Among the reserpine-pretreated groups, ter-
guride significantly increased DOPA accumulation in the
mPFC while aripiprazole had no effect (lower graph, Fig. 4)
[F(5,40)! 2.69, p<0.05]. The former result, showing that
terguride increased DOPA accumulation in reserpinized
rats, was not consistent with results from Experiment 1;
therefore, we did an additional experiment in which rats
were pretreated with reserpine and tested with vehicle,
1.5mg=kg terguride, or 30mg=kg aripiprazole. In this cir-
cumstance, terguride caused a 123% increase in DOPA
accumulation, while aripiprazole produced a slight, albeit
nonsignificant, decline (91.2% of control values) in DOPA
accumulation.
Experiment 3: Effects of D2-like partial agonists
on DOPA accumulation after GBL treatment
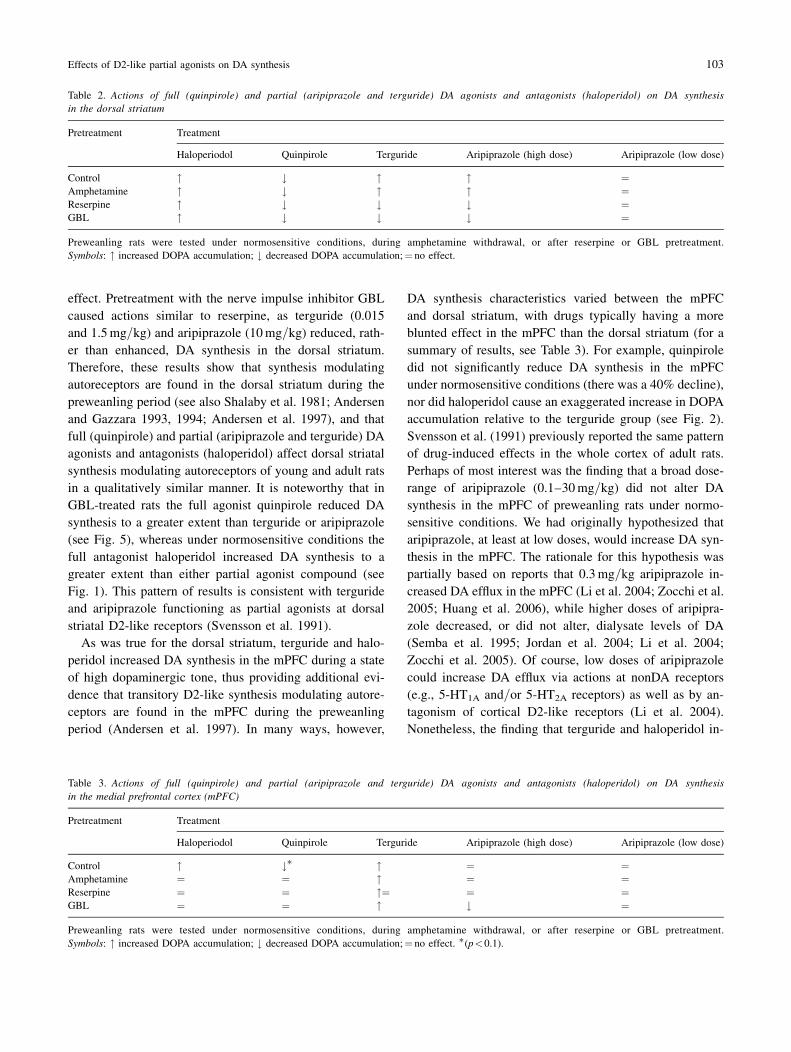
In the dorsal striatum, GBL (3.80 ng=mg, &0.20) caused a
373% increase in DOPA accumulation relative to vehicle
controls (1.02 ng=mg, &0.05). Haloperidol significantly
increased DOPA accumulation in GBL-treated rats, while
quinpirole induced a dramatic decline in DOPA levels (see
Fig. 5) [F(8,55)! 29.73, p<0.001]. Although quinpirole’s
effects were more robust, terguride (0.015 and 1.5mg=kg)and aripiprazole (10mg=kg) also reduced DOPA accumu-
lation relative to vehicle controls.
In GBL-treated rats, DOPA accumulation in the mPFC
(0.277 ng=mg, &0.01) was 199% greater than in vehicle
controls (0.139 ng=mg, &0.02). There was a nonsignificant
trend for haloperidol increasing, and quinpirole decreasing,
DOPA accumulation (see Fig. 6). When compared to
the vehicle group, the only significant effect was an aripi-
prazole-induced reduction in DOPA accumulation at the
10mg=kg dose [F(8,53)! 3.09, p<0.01].
Discussion
Results from the present study show that aripiprazole and
terguride function as prototypical partial agonists at syn-
thesis modulating D2-like autoreceptors in the dorsal stria-
tum of preweanling rats (for a summary of results, see
Table 2). When tested under normosensitive conditions
(i.e., in a state of high dopaminergic tone), terguride and
aripiprazole (3 and 10mg=kg) increased DA synthesis in
the dorsal striatum, while lower doses of aripiprazole (0.1
and 0.3mg=kg) were without impact. Reducing dopami-
nergic tone caused a different pattern of effects because
terguride and aripiprazole (10mg=kg) decreased dorsal
striatal DA synthesis in reserpinized rats, with lower doses
of aripiprazole (0.1, 0.3, and 3mg=kg) again having no
Fig. 5. Mean striatal DOPA accumulation of PD 20 rats injected with
vehicle, haloperidol (Hal), terguride (Terg), aripiprazole (Aripip), or quin-
pirole (Quin) 30min after GBL treatment. Data are expressed as percent ofvehicle controls (&SEM) and doses are in mg=kg. 'p<0.05 vs vehicle
controls (indicated by the dashed line); yp<0.05 vs haloperidol; zp<0.05
vs quinpirole
Fig. 6. Mean mPFC DOPA accumulation of PD 20 rats injected with
vehicle, haloperidol (Hal), terguride (Terg), aripiprazole (Aripip), or quin-pirole (Quin) 30min after GBL treatment. Data are expressed as percent of
vehicle controls (&SEM) and doses are in mg=kg. 'p<0.05 vs vehicle
controls (indicated by the dashed line); yp<0.05 vs haloperidol; zp<0.05
vs quinpirole
102 S. D. Iniguez et al.
effect. Pretreatment with the nerve impulse inhibitor GBL
caused actions similar to reserpine, as terguride (0.015
and 1.5mg=kg) and aripiprazole (10mg=kg) reduced, rath-er than enhanced, DA synthesis in the dorsal striatum.
Therefore, these results show that synthesis modulating
autoreceptors are found in the dorsal striatum during the
preweanling period (see also Shalaby et al. 1981; Andersen
and Gazzara 1993, 1994; Andersen et al. 1997), and that
full (quinpirole) and partial (aripiprazole and terguride) DA
agonists and antagonists (haloperidol) affect dorsal striatal
synthesis modulating autoreceptors of young and adult rats
in a qualitatively similar manner. It is noteworthy that in
GBL-treated rats the full agonist quinpirole reduced DA
synthesis to a greater extent than terguride or aripiprazole
(see Fig. 5), whereas under normosensitive conditions the
full antagonist haloperidol increased DA synthesis to a
greater extent than either partial agonist compound (see
Fig. 1). This pattern of results is consistent with terguride
and aripiprazole functioning as partial agonists at dorsal
striatal D2-like receptors (Svensson et al. 1991).
As was true for the dorsal striatum, terguride and halo-
peridol increased DA synthesis in the mPFC during a state
of high dopaminergic tone, thus providing additional evi-
dence that transitory D2-like synthesis modulating autore-
ceptors are found in the mPFC during the preweanling
period (Andersen et al. 1997). In many ways, however,
DA synthesis characteristics varied between the mPFC
and dorsal striatum, with drugs typically having a more
blunted effect in the mPFC than the dorsal striatum (for a
summary of results, see Table 3). For example, quinpirole
did not significantly reduce DA synthesis in the mPFC
under normosensitive conditions (there was a 40% decline),
nor did haloperidol cause an exaggerated increase in DOPA
accumulation relative to the terguride group (see Fig. 2).
Svensson et al. (1991) previously reported the same pattern
of drug-induced effects in the whole cortex of adult rats.
Perhaps of most interest was the finding that a broad dose-
range of aripiprazole (0.1–30mg=kg) did not alter DA
synthesis in the mPFC of preweanling rats under normo-
sensitive conditions. We had originally hypothesized that
aripiprazole, at least at low doses, would increase DA syn-
thesis in the mPFC. The rationale for this hypothesis was
partially based on reports that 0.3mg=kg aripiprazole in-
creased DA efflux in the mPFC (Li et al. 2004; Zocchi et al.
2005; Huang et al. 2006), while higher doses of aripipra-
zole decreased, or did not alter, dialysate levels of DA
(Semba et al. 1995; Jordan et al. 2004; Li et al. 2004;
Zocchi et al. 2005). Of course, low doses of aripiprazole
could increase DA efflux via actions at nonDA receptors
(e.g., 5-HT1A and=or 5-HT2A receptors) as well as by an-
tagonism of cortical D2-like receptors (Li et al. 2004).
Nonetheless, the finding that terguride and haloperidol in-
Table 2. Actions of full (quinpirole) and partial (aripiprazole and terguride) DA agonists and antagonists (haloperidol) on DA synthesisin the dorsal striatum
Pretreatment Treatment
Haloperiodol Quinpirole Terguride Aripiprazole (high dose) Aripiprazole (low dose)
Control " # " " !Amphetamine " # " " !Reserpine " # # # !GBL " # # # !
Preweanling rats were tested under normosensitive conditions, during amphetamine withdrawal, or after reserpine or GBL pretreatment.Symbols: " increased DOPA accumulation; # decreased DOPA accumulation;! no effect.
Table 3. Actions of full (quinpirole) and partial (aripiprazole and terguride) DA agonists and antagonists (haloperidol) on DA synthesis
in the medial prefrontal cortex (mPFC)
Pretreatment Treatment
Haloperiodol Quinpirole Terguride Aripiprazole (high dose) Aripiprazole (low dose)
Control " #' " ! !Amphetamine ! ! " ! !Reserpine ! ! "! ! !GBL ! ! " # !
Preweanling rats were tested under normosensitive conditions, during amphetamine withdrawal, or after reserpine or GBL pretreatment.
Symbols: " increased DOPA accumulation; # decreased DOPA accumulation;! no effect. '(p<0.1).
Effects of D2-like partial agonists on DA synthesis 103

creased DA synthesis in the mPFC, while aripiprazole
(0.1–30mg=kg) did not, is surprising and suggests that
nonDA receptors may mediate some of these effects.
When tested under conditions of low dopaminergic tone
the full agonist and antagonist compounds had only minor
effects on DA synthesis in the mPFC. More specifically,
neither quinpirole nor haloperidol significantly altered DA
synthesis in the mPFC of reserpine- or GBL-treated pre-
weanling rats, although nonsignificant trends were some-
times apparent. These results are more consistent with the
adult rat literature, because full D2-like agonists and an-
tagonists either do not alter DA synthesis in the mPFC after
reserpine or GBL treatment (Bannon et al. 1981; Galloway
et al. 1986; Svensson et al. 1991) or have reduced effects
when compared to actions in the dorsal striatum (Fadda
et al. 1984). In partial contrast to our results, Andersen
et al. (1997) reported that the full D2-like agonists (&)-
7-OH-DPAT and quinpirole dramatically reduced DA
synthesis in the mPFC of GBL-treated 15-day-old rats;
however, the dose of quinpirole (5mg=kg) needed to fully
reverse GBL-induced DOPA accumulation was substantial-
ly larger than the dose (0.5mg=kg) employed in the present
study.
In terms of the partial agonist compounds, terguride
and aripiprazole had opposing effects on DA synthesis
in the mPFC under conditions of low dopaminergic tone.
Specifically, 1.5mg=kg terguride enhanced DOPA accumu-
lation in the mPFC of GBL-treated preweanling rats, while
10mg=kg aripiprazole significantly reversed the GBL-
induced increase in DOPA accumulation. In reserpinized
rats, aripiprazole had minimal effects on DA synthesis in
the mPFC, while 1.5mg=kg terguride increased DA syn-
thesis (see Fig. 4). The latter effect was not observed in
Experiment 1 (see Fig. 2), so we did an additional ex-
periment employing the same procedures and found that
1.5mg=kg terguride increased DOPA accumulation in the
mPFC by 123% relative to reserpinized controls. In the
mPFC, therefore, terguride was probably responsible for
marginal increases in DA synthesis under conditions of
low dopaminergic tone, while aripiprazole decreased DA
synthesis in GBL-treated rats. The reason why these two
partial agonists had divergent effects is uncertain, but it
may have been due to the nondopaminergic actions of these
compounds (e.g., see Stark et al. 2007).
In adult rats, amphetamine withdrawal has been reported
to reduce dopaminergic tone to such an extent that partial
agonists will stimulate, rather than antagonize, D2-like re-
ceptors. Specifically, aripiprazole and terguride were able
to reverse the decline in saccharin-reinforced bar press
responding that was evident during amphetamine with-
drawal (Orsini et al. 2001; Schwabe and Kock 2007; see
also Hoefer et al. 2006). In the present study, the identical
amphetamine withdrawal paradigm did not induce a state
in which D2-like partial agonists were able to stimulate the
synthesis modulating autoreceptors of preweanling rats. In
the dorsal striatum, for example, terguride and aripiprazole
(3 and 10mg=kg) increased DOPA accumulation in am-
phetamine- and saline-pretreated rats, whereas the same
partial agonists decreased DOPA accumulation in reserpi-
nized rats (see Fig. 1). The inability of D2-like partial
agonists to stimulate presynaptic receptors during amphet-
amine withdrawal was probably not surprising, because
endogenous DA levels in the dorsal striatum of preweanl-
ing rats were not altered during amphetamine withdrawal
(Table 1). In comparison, reserpine (1mg=kg) causes a
substantial reduction in dorsal striatal DA content (to about
10% of control values) when measured 24 h later on PD 20
(Wacan et al. 2006). The relative impact of these pretreat-
ments (i.e., amphetamine withdrawal vs reserpine) on DA
content is meaningful, because D2-like partial agonists typ-
ically exhibit enhanced intrinsic activity only under condi-
tions of past low receptor occupancy (e.g., after reserpine
treatment or 6-OHDA lesions) (Carlsson 1983; Clark et al.
1985b). Meller et al. (1987) have also proposed that manip-
ulations (e.g., reserpine) which increase the receptor re-
serve are needed before partial agonists will exhibit high
intrinsic activity at synthesis modulating autoreceptors.
Regardless of explanation, amphetamine withdrawal was
unable to induce a state in which terguride or aripiprazole
stimulated, rather than antagonized, the synthesis modulat-
ing autoreceptors of preweanling rats.
Although there is substantial evidence that terguride and
aripiprazole act as partial agonists at D2-like receptors,
both drugs exhibit high affinity for other receptor types.
In addition to being partial agonists at D2, D3, and D4
receptors (Newman-Tancredi et al. 2002a; Shapiro et al.
2003), terguride and aripiprazole function as antagonists
at the 5-HT2B receptor and as partial agonists at the
5-HT1A receptor (Jordan et al. 2002; Newman-Tancredi
et al. 2002b; Shapiro et al. 2003). Actions at the 5-HT1A
receptor may be relevant because R(#)-8-OH-DPAT (a
5-HT1A receptor agonist with some D2 agonist properties)
has been reported to decrease DOPA accumulation in the
mPFC (Kuroki et al. 2000; see also Hillegaart et al. 2000).
Both partial agonists also have significant affinity for
a1A-adrenergic and histamine receptors (Millan et al. 2002;
Newman-Tancredi et al. 2002a; Shapiro et al. 2003).
Perhaps of most interest is the finding that aripiprazole is
an antagonist at 5-HT2A receptors (Burris et al. 2002; Byars
et al. 2002), whereas Newman-Tancredi et al. (2002b) re-
104 S. D. Iniguez et al.

port that terguride is a potent agonist at human 5-HT2A
receptors. Whether differential actions at the 5-HT2A re-
ceptor are responsible for some of the disparate effects of
terguride and aripiprazole is uncertain. Interestingly, Urban
et al. (2007) have proposed that aripiprazole has a unique
role as a ‘‘functionally selective D2 ligand’’ rather than as a
‘‘simple partial agonist’’ (see also Lawler et al. 1999).
According to this hypothesis, aripiprazole is capable of ex-
hibiting different intrinsic activities at a given receptor type
depending on the effector system being measured. Because
terguride has the characteristics of a ‘‘simple partial ago-
nist’’, it is possible that some of the differential effects of
terguride and aripiprazole at synthesis modulating auto-
receptors, especially in the mPFC, may be due to aripipra-
zole’s actions as a functionally selective D2-like ligand.
In conclusion, partial agonist compounds affected the
synthesis modulating autoreceptors of the dorsal striatum
in an adult-typical manner during the preweanling period.
In the mPFC aripiprazole was without significant action,
while terguride and haloperidol increased DA synthesis
under normosensitive conditions. The latter results are con-
sistent with studies showing that synthesis modulating D2-
like autoreceptors are functional in the mPFC by PD 10
and are able to mediate agonist-induced decreases in DA
synthesis until at least PD 30 (Andersen et al. 1997). That
aripiprazole did not alter DA synthesis characteristics in the
mPFC under normosensitive conditions may have relevance
to pediatric clinical populations, because these results sug-
gest that aripiprazole has similar actions at the synthesis
modulating autoreceptors of young and adult animals.
Acknowledgments
We thank Faiza Furqan for technical contributions. This work was partially
supported by a grant (GM073842) from the National Institutes of Health.
References
Andersen SL (2003) Trajectories of brain development: point of vulnera-
bility or window of opportunity. Neurosci Biobehav Rev 27: 3–18
Andersen SL, Gazzara RA (1993) The ontogeny of apomorphine-induced
alterations of neostriatal dopamine release: effects on spontaneousrelease. J Neurochem 61: 2247–2255
Andersen SL, Gazzara RA (1994) The development of D2 autoreceptor-
mediated modulation of K(#)-evoked dopamine release in the neo-
striatum. Dev Brain Res 78: 123–130Andersen SL, Dumont NL, Teicher MH (1997) Developmental differences
in dopamine synthesis inhibition by (&)-7-OH-DPAT. Naunyn
Schmiedebergs Arch Pharmacol 356: 173–181Baldessarini RJ, Marsh ER, Huston-Lyons D (1994) Effects of dopamine
partial-agonist aminoergolines on dopamine metabolism in limbic
and extrapyramidal regions of rat brain. Biochem Pharmacol 47:
1917–1919
Bannon MJ, Michaud RL, Roth RH (1981) Mesocortical dopamine neu-rons: lack of autoreceptors modulating dopamine synthesis. Mol
Pharmacol 19: 270–275
Barr AM, Phillips AG (1999) Withdrawal following repeated exposure to
D-amphetamine decreases responding for a sucrose solution as mea-sured by a progressive ratio schedule of reinforcement. Psychophar-
macology 141: 99–106
Barzman DH, DelBello MP, Kowatch RA, Gernert B, Fleck DE, Pathak S,
Rappaport K, Delgado SV, Campbell P, Strakowski SM (2004) Theeffectiveness and tolerability of aripiprazole for pediatric bipolar
disorders: a retrospective chart review. J Child Adolesc Psychophar-
macol 14: 593–600Biederman J,McDonnellMA,Wozniak J, Spencer T, AleardiM, Falzone R,
Mick E (2005) Aripiprazole in the treatment of pediatric bipolar
disorder: a systematic chart review. CNS Spectr 10: 141–148
Brucke T, DanielczykW, Simanyi M, Sofic E, Riederer P (1987) Terguride:partial dopamine agonist in the treatment of Parkinson’s disease. Adv
Neurol 45: 573–576
Burris KD, Molski TF, Xu C, Ryan E, Tottori K, Kikuchi T, Yocca FD,
Molinoff PB (2002) Aripiprazole, a novel antipsychotic, is a high-affinity partial agonist at human dopamine D2 receptors. J Pharmacol
Exp Ther 302: 381–389
Byars A, Burris K, Jordan S, Tottori K, Kikuchi T, McQuade R (2002)Aripiprazole: a dopamine-serotonin system stabilizer. Eur Neuro-
psychopharmacol 12(Suppl 3): S290–S291
Carlsson A (1983) Dopamine receptor agonists: intrinsic activity vs. state of
the receptor. J Neural Transm 57: 309–315Clark D, Hjorth S, Carlsson A (1985a) (#)- and ($)-3-PPP exhibit different
intrinsic activity at striatal dopamine autoreceptors controlling dopa-
mine synthesis. Eur J Pharmacol 106: 185–189
Clark D, Hjorth S, Carlsson A (1985b) Dopamine receptor agonists:mechanisms underlying autoreceptor selectivity. II. Theoretical con-
siderations. J Neural Transm 62: 171–207
Clark D, Salah RS, Galloway MP (1991) Differential agonist profile of theenantiomers of 3-PPP at striatal dopamine autoreceptors: dependence
on extracellular dopamine. Synapse 8: 169–176
Corsini GU, Bonuccelli U, Rainer E, Del Zompo M (1985) Therapeutic
efficacy of a partial dopamine agonist in drug-free parkinsonianpatients. J Neural Transm 64: 105–111
Critchley P, Parkes D (1987) Transdihydrolisuride in parkinsonism. Clin
Neuropharmacol 10: 57–64
Davies MA, Sheffler DJ, Roth BL (2004) Aripiprazole: a novel atypicalantipsychotic drug with a uniquely robust pharmacology. CNS Drug
Rev 10: 317–336
Fadda F, Gessa GL, Marcou M, Mosca E, Rossetti Z (1984) Evidence for
dopamine autoreceptors in mesocortical dopamine neurons. Brain Res293: 67–72
Farley CM, Baella SA,Wacan JJ, Crawford CA,McDougall SA (2006) Pre-
and postsynaptic actions of a partial D2 receptor agonist in reser-pinized young rats: longevity of agonistic effects. Brain Res 1124:
37–44
Feltenstein MW, Altar CA, See RE (2007) Aripiprazole blocks reinstate-
ment of cocaine seeking in an animal model of relapse. Biol Psychiatry61: 582–590
Findling RL, Blumer J, Kauffman R, Batterson J, Gilbert D, Bramer S,
Marcus R (2003) Aripiprazole in pediatric conduct disorder: a pilot
study. Eur Neuropsychopharm 13(Suppl 4): S335GallowayMP,WolfME, Roth RH (1986) Regulation of dopamine synthesis
in the medial prefrontal cortex is mediated by release modulating
autoreceptors: studies in vivo. J Pharmacol Exp Ther 236: 689–698Hillegaart V, Magnusson O, Ahlenius S (2000) A9 and A10 dopamine
nuclei as a site of action for effects of 8-OH-DPATon locomotion in the
rat. Pharmacol Biochem Behav 67: 55–63
Hjorth S, Carlsson A, Clark D, Svensson K, Wikstrom H, Sanchez D,Lindberg P, Hacksell U, Arvidsson LE, Johansson A, Nilsson JLG
Effects of D2-like partial agonists on DA synthesis 105

(1983) Central dopamine receptor agonist and antagonist actions of theenantiomers of 3-PPP. Psychopharmacology 81: 89–99
Hjorth S, Clark D, Carlsson A (1988) Dopamine (DA) autoreceptor efficacy
of 3-PPP enantiomers after short-term synaptic DA deprivation. Eur J
Pharmacol 152: 207–215Hoefer ME, Voskanian SJ, Koob GF, Pulvirenti L (2006) Effects of
terguride, ropinirole, and acetyl-L-carnitine on methamphetamine
withdrawal in the rat. Pharmacol Biochem Behav 83: 403–409
Huang M, Li Z, Ichikawa J, Dai J, Meltzer HY (2006) Effects of divalproexand atypical antipsychotic drugs on dopamine and acetylcholine efflux
in rat hippocampus and prefrontal cortex. Brain Res 1099: 44–55
Hughes CW (1979) Outcome of early experience studies as affected bybetween-litters variance. J Nutr 109: 642–645
Jordan S, Koprivica V, Chen R, Tottori K, Kikuchi T, Altar CA (2002) The
antipsychotic aripiprazole is a potent, partial agonist at the human
5-HT1A receptor. Eur J Pharmacol 441: 137–140Jordan S, Koprivica V, Dunn R, Tottori K, Kikuchi T, Altar CA (2004) In
vivo effects of aripiprazole on cortical and striatal dopaminergic and
serotonergic function. Eur J Pharmacol 483: 45–53
Keck PE Jr, Marcus R, Tourkodimitris S, Ali M, Liebeskind A, Saha A,Ingenito G (2003) A placebo-controlled, double-blind study of the
efficacy and safety of aripiprazole in patients with acute bipolar mania.
Am J Psychiatry 160: 1651–1658Kehr W (1984) Transdihydrolisuride, a partial dopamine receptor an-
tagonist: Effects on monoamine metabolism. Eur J Pharmacol 97:
111–119
Kikuchi T, Tottori K, Uwahodo Y, Hirose T, Miwa T, Oshiro Y, Morita S(1995) 7-(4-[4-(2,3-Dichlorophenyl)-1-piperazinyl]butyloxy)-3,4-
dihydro-2(1H)-quinolinone (OPC-14597), a new putative antipsychot-
ic drug with both presynaptic dopamine autoreceptor agonistic activity
and postsynaptic D2 receptor antagonistic activity. J Pharmacol ExpTher 274: 329–336
Kuroki T, Dai J, Meltzer HY, Ichikawa J (2000) R(#)-8-OH-DPAT, a
selective 5-HT1A receptor agonist, attenuated amphetamine-induceddopamine synthesis in rat striatum, but not nucleus accumbens or
medial prefrontal cortex. Brain Res 872: 204–207
Lawler CP, Prioleau C, Lewis MM, Mak C, Jiang D, Schetz JA, Gonzalez
AM, Sibley DR (1999) Interactions of the novel antipsychotic aripi-prazole (OPC-14597) with dopamine and serotonin receptor subtypes.
Neuropsychopharmacology 20: 612–627
Li Z, Ichikawa J, Dai J, Meltzer HY (2004) Aripiprazole, a novel antipsy-
chotic drug, preferentially increases dopamine release in the prefrontalcortex and hippocampus in rat brain. Eur J Pharmacol 493: 75–83
Meller E, Bohmaker K, Namba Y, Friedhoff AJ, Goldstein M (1987)
Relationship between receptor occupancy and response at striatal au-
toreceptors. Mol Pharmacol 31: 592–598Millan MJ, Maiofiss L, Cussac D, Audinot V, Boutin J-A, Newman-
Tancredi A (2002) Differential actions of antiparkinson agents at
multiple classes of monoaminergic receptor. I. A multivariate analysisof the binding profiles of 14 drugs at 21 native and cloned human
receptor subtypes. J Pharmacol Exp Ther 303: 791–804
Murphy TK, BengtsonMA, Soto O, Edge PJ, SajidMW, Shapira N, YangM
(2005) Case series on the use of aripiprazole for Tourette syndrome. IntJ Neuropsychopharmacol 8: 489–490
Newman-Tancredi A, Cussac D, Audinot V, Nicolas JP, De Ceuninck F,
Boutin J-A, Millan MJ (2002a) Differential actions of antiparkinson
agents at multiple classes of monoaminergic receptor. II. Agonist andantagonist properties at subtypes of dopamine D2-like receptor and
a1=a2-adrenoceptor. J Pharmacol Exp Ther 303: 805–814
Newman-Tancredi A, Cussac D, Quentric Y, Touzard M, Verri!eele L,Carpentier N, Millan MJ (2002b) Differential actions of antiparkinson
agents at multiple classes of monoaminergic receptor. III. Agonist and
antagonist properties at serotonin, 5-HT1 and 5-HT2, receptor sub-types. J Pharmacol Exp Ther 303: 815–822
Orsini C, Koob GF, Pulvirenti L (2001) Dopamine partial agonist re-
verses amphetamine withdrawal in rats. Neuropsychopharmacology
25: 789–792Oshiro Y, Sato S, Kurahashi N, Tanaka T, Kikuchi T, Tottori K, Uwahodo Y,
Nishi T (1998) Novel antipsychotic agents with dopamine autoreceptor
agonist properties: synthesis and pharmacology of 7-[4-(4-phenyl-
1-piperazinyl)butoxy]-3,4-dihydro-2(1H)-quinolinone derivatives. JMed Chem 41: 658–667
Platt DM, Rodefer JS, Rowlett JK, Spealman RD (2003) Suppression of
cocaine- and food-maintained behavior by the D2-like receptor par-tial agonist terguride in squirrel monkeys. Psychopharmacology 166:
298–305
Pulvirenti L, Koob GF (1994) Dopamine receptor agonists, partial agonists
and psychostimulant addiction. Trends Pharmacol Sci 15: 374–379Rugino TA, Janvier YM (2005) Aripiprazole in children and adolescents:
clinical experience. J Child Neurol 20: 603–610
Schwabe K, Koch M (2007) Effects of aripiprazole on operant responding
for a natural reward after psychostimulant withdrawal in rats. Psycho-pharmacology 191: 759–765
Semba J, Watanabe A, Kito S, Toru M (1995) Behavioural and neurochem-
ical effects of OPC-14597, a novel antipsychotic drug, on dopaminer-gic mechanisms in rat brain. Neuropharmacology 34: 785–791
Shalaby IA, Dendel PS, Spear LP (1981) Differential functional ontogeny
of dopamine presynaptic receptor regulation. Brain Res 227: 434–439
Shapiro DA, Renock S, Arrington E, Chiodo LA, Liu LX, Sibley DR, RothBL, Mailman R (2003) Aripiprazole, a novel atypical antipsychotic
drug with a unique and robust pharmacology. Neuropsychopharma-
cology 28: 1400–1411
Stachnik JM, Nunn-Thompson C (2007) Use of atypical antipsychotics inthe treatment of autistic disorder. Ann Pharmacother 41: 626–634
Stark AD, Jordan S, Allers KA, Bertekap RL, Chen R, Kannan TM, Molski
TF, Yocca FD, Sharp T, Kikuchi T, Burris KD (2007) Interaction of thenovel antipsychotic aripiprazole with 5-HT1A and 5-HT2A receptors:
functional receptor-binding and in vivo electrophysiological studies.
Psychopharmacology 190: 373–382
Stigler KA, Posey DJ, McDougle CJ (2004) Aripiprazole for maladaptivebehavior in pervasive developmental disorders. J Child Adolesc
Psychopharmacol 14: 455–463
Svensson K, Ekman A, Piercey MF, Hoffmann WE, Lum JT, Carlsson A
(1991) Effects of the partial dopamine receptor agonists SDZ 208-911,SDZ 208-912 and terguride on central monoamine receptors. A
behavioral, biochemical and electrophysiological study. Naunyn-
Schmiedebergs Arch Pharmacol 344: 263–274
Svensson K, Eriksson E, Carlsson A (1993) Partial dopamine receptoragonists reverse behavioral, biochemical and neuroendocrine effects
of neuroleptics in the rat: potential treatment of extrapyramidal side
effects. Neuropharmacology 32: 1037–1045Urban JD, Vargas GA, von Zastrow M, Mailman RB (2007) Aripiprazole
has functionally selective actions at dopamine D2 receptor-mediated
signaling pathways. Neuropsychopharmacology 32: 67–77
Wacan JJ, Reichel CM, Farley CM, McDougall SA (2006) The partialdopamine D2-like receptor agonist terguride functions as an agonist in
preweanling rats after a 5-day reserpine regimen. Psychopharmacology
185: 104–111
Yoshida ST, Baella SA, Stuebner NM, Crawford CA, McDougall SA (2006)Effects of a partial D2-like receptor agonist on striatal dopamine autore-
ceptor functioning in preweanling rats. Brain Res 1073=1074: 269–275Zocchi A, Fabbri D, Heidbreder CA (2005) Aripiprazole increases dopa-
mine but not noradrenaline and serotonin levels in the mouse prefrontal
cortex. Neurosci Lett 387: 157–161
106 S. D. Iniguez et al.: Effects of D2-like partial agonists on DA synthesis