effect of nonionic surfactants on rhizopus homothallicus lipase activity
TRANSCRIPT

RESEARCH
Molecular Biotechnology © 2007 Humana Press Inc. All rights of any nature whatsoever reserved. ISSN: 1073–6085/Online ISSN: 1559–0305/2007/35:3/205–214/$30.00
MOLECULAR BIOTECHNOLOGY 205 Volume 35, 2007
1. IntroductionLipases (triacylglycerol ester hydrolases EC
3.1.1.3) are ubiquitous enzymes that catalyzethe hydrolysis of long-chain triacylglycerols(TAG). These enzymes play diverse roles in thedegradation of food and fat, and they provide poten-tial means of treating digestive disorders and pancre-atic disease (for a review, see ref. 1). They alsohave many applications in the field of biotechnol-ogy where they are used as detergent additivesand catalysts for the manufacture of specialty
(oleo)chemicals and in the synthesis of organic sub-stances (1). Lipases have been found to be quitestable in organic solvents and therefore, can cat-alyze both the hydrolysis and the synthesis of awide range of natural and synthetic carboxylicesters. One important aspect of lipases is the uniquephysicochemical reaction process they catalyze atlipid/water interfaces, involving interfacialadsorption and subsequent catalysis stricto sensu.
Several lipases from Rhizopus sp. have beenproduced, purified, crystallized, and biochemically
Effect of Nonionic Surfactants on Rhizopus homothallicus Lipase Activity
A Comparative Kinetic Study
J. C. Mateos Diaz,1 J. Cordova,2 J. Baratti,1 F. Carriere,3and A. Abousalham3,4,*
*Author to whom all correspondence and reprint requests should be addressed: 1Groupe de Biocatalyse et Chimie Fine, Universitéde la Méditerranée, Marseille, France; 2Laboratorio de Bioprocesos, Universidad de Guadalajara, Guadalajara, México;3Laboratoire d’Enzymologie Interfaciale et de Physiologie de la Lipolyse, UPR 9025, CNRS, Marseille, France; 4GERME S.A.,Marseille, France. E-mail: [email protected].
Abstract
Based on amino-terminal sequencing and mass spectrometry data on the Rhizopus homothallicus lipaseextracted using solid (SSF) and submerged state fermentation (SmF) methods, we previously establishedthat the two enzymes were identical. Differences were observed, however, in terms of the specific activity ofthese lipases and their inhibition by diethyl p-nitrophenyl phosphate (E600). The specific activity of the SSFlipase (10,700 µmol/min/mg) was found to be 1.2-fold that of SmF lipase (8600 µmol/min/mg). These dif-ferences might be the result of residual Triton X-100 molecules interacting with the SSF lipase. To checkthis hypothesis, the SmF lipase was incubated with submicellar concentrations of Triton X-100. The specificactivity of the lipase increased after this treatment, reaching similar values to those measured with the SSFlipase. Preincubating SSF and SmF lipases with E600 at a molar excess of 100 for 1 h resulted in 80% and60% enzyme inhibition levels, respectively. When the SmF lipase was preincubated with Triton X-100 for 1 h at a concentration 100 times lower than the Triton X-100 critical micellar concentration, the inhibition ofthe lipase by E600 increased from 60% to 80%. These results suggest that residual detergent monomersinteracting with the enzyme may affect the kinetic properties of the Rh. homothallicus lipase.
Index Entries: Nonionic surfactants; lipase; Triton X-100; Rhizopus homothallicus; solid state fermentation;submerged fermentation.

characterized in many previous studies (2–4).Rh. delemar produces a lipase that cleaves andsynthesizes primary esters with a 1,3-regioselec-tivity toward glycerides (2). Lipase fromRh. niveus shows a higher 1,3-regioselectivitythan other known lipases (5). Kinetic studies onRh. oryzae lipase, using the monomolecular filmtechnique and various isomers of dicaprin, haveshown the existence in this lipase of stereoselectivityfor the hydrolysis of the sn-3 position in the 2,3enantiomer of dicaprin (6). All the known aminoacid sequences of lipases from Rhizopus sp. areidentical, except for Rh. oryzae lipase, which containstwo mutations (His134Asn and Ile234Leu) (4).Therefore, it seems likely that the various propertiesof lipases from Rhizopus sp. described in theliterature, such as their temperature and pHprofiles and their thermostability, may not bebecause of the existence of different genes but ofvarious post-translational modifications and possiblyinteractions with small molecules (4).
As in most three-dimensional (3D) lipasestructures, access to the catalytic site of Rhizopussp. lipase is controlled by a so-called lid, whichcovers the catalytic site in the crystal structuresof the Rh. delemar (7) and Rh. niveus (5) lipases.In these studies, the enzyme was found to be inan inactive, closed conformation. Structural stud-ies on related lipase–inhibitor complexes (8,9)have shown that a conformational change canoccur in the lid, resulting in the exposure of thecatalytic site to solvent. Most of the so-calledopen 3D structure of lipases were obtained in thepresence of detergent, and some detergentmonomers have often been observed in the crys-tal structure. Crystals of Rh. delemar lipase con-tain lipase molecules in which the lid was foundto have two different conformations: in the firstone, the lid was in the closed conformation; in thesecond one, the lid was found to be in an inter-mediate position between the open and closedconformations (7). This intermediate lid confor-mation was the result of nonspecific binding of anonionic detergent used in the crystallizationmedium (7). Egloff et al. (10) observed somedetergent molecules (b-octyl glucoside) linked tothe pancreatic lipase–colipase complex inhibited
by alkyl phosphonates. Some detergent moleculesbound to the hydrophobic part of the lid werelocated at the entrance to the active site.Furthermore, Hermoso et al. (11) observed theexistence of monomers of the nonionic detergenttetraethylene glycol monooctyl ether (TGME)bound to the porcine pancreatic lipase–colipasecomplex, where one TGME molecule was pres-ent in the active site, stabilizing the open confor-mation of the lid domain.
Nonionic surfactants, such as Triton X-100, arewidely used for lipase extraction (12,13), lipasepurification (14,15), lipase desorption from a matrix(16), and the isolation of lipases associated withmembranes (17). Gonzalez-Navarro et al. (18) haveestablished that n-octyl-β-glucopyranoside, at con-centrations below the critical micellar concentra-tion (CMC), induce fungal lipase activation inaqueous solutions. These results are consistent withthe idea that lipase activation involves several equi-librium states (closed, intermediate, and open),which depend on the composition of the micro-environment (i.e., the state of lipases can change inresponse to detergent, water–miscible organic solvents,or other amphiphilic molecules) (18). Bano et al.(19) have reported the occurrence of a noticeableincrease in the hydrolytic activity of two fungallipases assayed in aqueous solution in the absenceof any interface, when submicellar concentra-tions of the amphipathic molecule palmitoyl-CoAwere added. These investigators suggested that othernonaggregated amphipathic constituents may act asregulators of lipase activity (19). In addition, it hasbeen suggested on the basis of fluorescence spec-troscopy data that premicellar detergent assembliesmay stabilize an open lipase conformation (20).
Iwai et al. (21) and Shimada et al. (22) havereported that Rh. delemar lipase activity on lipopro-tein and water-soluble ester substrates increasedafter the lipase had been treated with phos-phatidylcholine (PC), although no change wasobserved on an olive oil substrate. In anotherstudy, Shimada et al. (23) have reported that theeffects of PC on Rh. delemar lipase activity arereversible. In addition, experiments performedwith radiolabeled [14C]dioleoyl PC have shownthat adding about 5 moles of PC per mole of
206 Mateos Diaz et al.
MOLECULAR BIOTECHNOLOGY Volume 35, 2007

lipase resulted in high levels of activity on lipopro-tein substrates (23). These results suggested that alocal structural change in the lipase probablyoccurred with the binding of PC to the lipase mole-cule resulting in an increase in the hydrolyticactivity on water-soluble esters (22). In a laterstudy, Sugihara et al. (24) showed that Rh. delemarlipase activity increased after treatment withphospholipids because of the enhancement of its penetration into organized lipid structures(monomolecular films). These data suggested thatthe hydrophobicity of the lipid-binding domainincreased in the lipase as a result of interactionswith PC molecules. Based on the structural datanow available, it seems likely that PC molecules,like some detergent molecules, may favor theopen conformation of the lipase lid domain,thus exposing a larger hydrophobic surface andpromoting interactions with the lipid substrate.
Although it is well known that amphiphilicmolecules can interact with lipolytic enzymes,only a few studies have focused so far on theeffects of surfactants on lipases’ kinetic properties.Here we present the results of a comparativekinetic study on the effects of the nonionic detergentTriton X-100 on the purified Rh. homothallicuslipases obtained using submerged (SmF) andsolid state fermentation (SSF) methods.
2. Materials and Methods2.1. Reagents and Media
Vinyl acetate (VC2), vinyl propionate (VC3),vinyl butyrate (VC4), vinyl laurate (VC12), TritonX-100, diethyl p-nitrophenyl phosphate (E600),and sodium taurodeoxycholate (NaTDC) werepurchased from Sigma-Aldrich-Fluka Chimie(St-Quentin-Fallavier, France). Triacetylglycerol(TC2), tripropanoylglycerol (TC3), tributyroylglycerol(TC4), tricaproylglycerol (TC6), and trioctanoyl-glycerol (TC8) were from Acros Organics (Noisy-Le-Grand, France). All other chemicals and solventswere of reagent or better quality grade and wereobtained from local suppliers.
2.2. Lipase PurificationThe lipase was extracted using SSF and SmF
methods, and purified using two chromatographic
steps, as described previously (25). Briefly,Rh. homothallicus was grown in solid and liquidculture medium at 40°C for 12 h and 22 h, respec-tively. Broth extract containing lipase activity wasfiltered from submerged cultures or extracted fromsolid cultures with a 0.5% Triton X-100 solution.The extract was then loaded onto a hydrophobicButyl-Sepharose column after adding 1 M ofammonium sulfate. Fractions with lipase activityeluted from the Butyl-Sepharose column werepooled and further subjected to gel filtrationchromatography on a Superdex G-200 columnequilibrated in 10 mM Tris-HCl (pH 8) containing150 mM NaCl, at a flow rate of 1 mL/min.Fractions (1 mL) showing lipase activity on tri-octanoin were collected and pooled for further studies.Only the lipase corresponding to peak 2 (see Fig. 1)was used for the kinetic studies.
2.3. Lipase Activity MeasurementsLipase activity was assayed by measuring the
free fatty acids released from mechanically stirredtriglyceride or vinyl ester emulsions, using a pH-stat(TTT80 radiometer, Copenhagen) adjusted with 0.1 NNaOH at a constant pH end point value. Each assaywas performed in a thermostated (37°C) vesselcontaining 2.5 mM Tris-HCl buffer and 150 mMNaCl. When olive oil or other long-chaintriglycerides were used as the substrates, theseglycerides were pre-emulsified with 10% gumarabic, and CaCl2 (2 mM, final concentration) wasadded in the assay performed at pH 8. The rate ofspontaneous substrate hydrolysis was recorded for2 min before lipase injection. The specific lipolyticactivities are expressed in international units (IU)per milligram of enzyme. One unit corresponds to1 µmol of fatty acid released per minute.
2.4. Protein Determination and GelElectrophoresis
Protein concentration was routinely deter-mined using the Bradford (26) microassayprocedure with Bio-Rad Dye Reagent. Bovineserum albumin (BSA) was used as the referenceprotein. Sodium dodecyl sulfate–polyacrylamidegel electrophoresis (SDS-PAGE) was carried outusing a Bio-Rad Mini Protein II dual vertical slab
Effect of Nonionic Surfactants 207
MOLECULAR BIOTECHNOLOGY Volume 35, 2007
gel electrophoresis cell (Bio-Rad Laboratories)and gels containing 12% of acrylamide asdescribed by Laemmli (27). Low molecular massmarkers (Amersham) were used for molecularmass determination. The proteins separated onthe gel were stained with Coomasie brilliant blueR-250.
2.5. Effects of Surfactants on LipaseActivity
The SmF lipase purified form (0.14 mg/mL inTris-HCl buffer at pH 7.5) was incubated with
micellar concentrations of various surfactants(Tween 80, Triton X-100, and NaTDC) for 1 h at37°C. At the end of the incubation period, lipaseactivity was determined using a trioctanoin sub-strate under standard assay conditions (discussedpreviously). Control experiments were performedwithout surfactants.
Alternatively, the effects of submicellar con-centrations of Triton X-100 (0.0002 mM, finalconcentration) on lipase substrate specificity weretested with various triglycerides and vinyl esterscontaining fatty acids with chain lengths ranging
208 Mateos Diaz et al.
MOLECULAR BIOTECHNOLOGY Volume 35, 2007
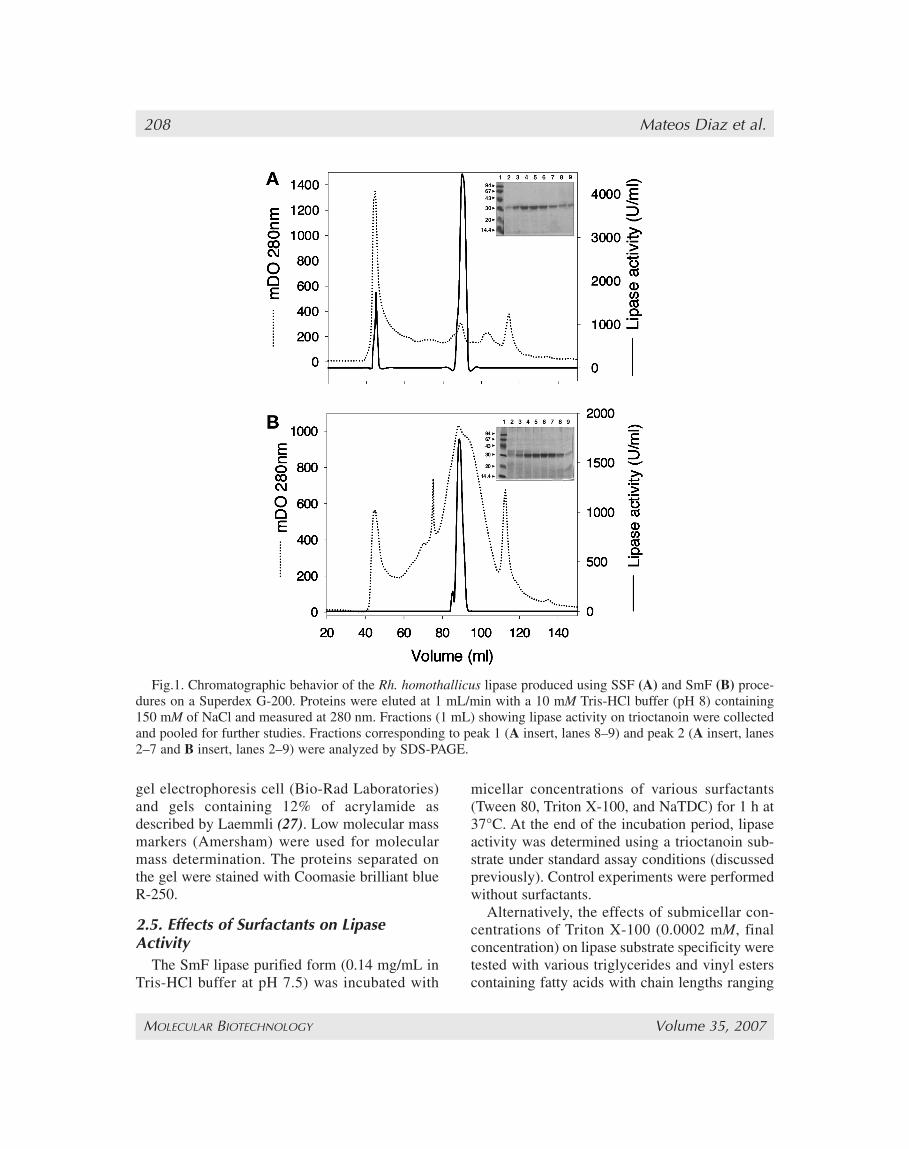
Fig.1. Chromatographic behavior of the Rh. homothallicus lipase produced using SSF (A) and SmF (B) proce-dures on a Superdex G-200. Proteins were eluted at 1 mL/min with a 10 mM Tris-HCl buffer (pH 8) containing150 mM of NaCl and measured at 280 nm. Fractions (1 mL) showing lipase activity on trioctanoin were collectedand pooled for further studies. Fractions corresponding to peak 1 (A insert, lanes 8–9) and peak 2 (A insert, lanes2–7 and B insert, lanes 2–9) were analyzed by SDS-PAGE.

from C2 to C18, using standard assay conditions.On the other hand, the effects of increasing TritonX-100 concentrations on lipase activity werefurther assayed using a trioctanoin substrate.
2.6. Effects of Triton X-100 on the LipaseInhibition by E600
The effects of Triton X-100 on the lipase inhibi-tion exerted by E600 was studied by prein-cubating the SmF lipase (0.14 mg/mL in Tris-HClbuffer at pH 7.5), with Triton X-100 (0.002 mM,final concentration in Tris-HCl buffer at pH 7.5)and E600 (0.5 mM, final concentration) at anenzyme-to-inhibitor molar ratio of 1:100. A stocksolution (10 mM) of E600 was initially preparedin ethanol. Control experiments were performedwithout any inhibitor or Triton X-100. The residuallipase activity was measured at variouspreincubation times, using a trioctanoin substrate(discussed previously).
3. Results and Discussion3.1. Chromatographic Behavior of Rh. homothallicus Lipase on a GelFiltration Column
The two lipases produced in either SSF or SmFwere purified to homogeneity using a two-stepprocedure involving hydrophobic and gel filtrationchromatography (25). Gel filtration chromato-grahy on Superdex G-200 and the SDS-PAGE of theSSF and SmF fractions containing lipase activityare shown in Fig. 1 A,B. When the SSF lipase wassubjected to gel filtration chromatography, twodifferent peaks containing lipase activity wereobserved (Fig. 1A). The fractions corresponding tothe first peak (peak 1) contained around 27% of thelipase activity and were eluted in the void volume(43 mL) of the column; the second peak (peak 2)was eluted at the expected volume (88–92 mL),corresponding to proteins with a molecularmass of 29.5 kDa. When the SmF lipase wassubjected to gel filtration chromatography, a singlepeak (peak 2) corresponding to a 29.5-kDa proteinwas observed (Fig. 1B).
As previously described (25), Triton X-100 wasused in the SSF lipase purification procedure toelute the lipase from the solid fermentation
medium. In addition, because of the initial presenceof olive oil in the solid culture medium (olive oilwas not present in the case of SmF), lipolysis prod-ucts (i.e., free fatty acids, mono- and di-glycerides)were also extracted with the enzyme during elution.The gel filtration chromatography pattern observedin Fig. 1A suggests that a partial aggregation oflipase with lipolysis products may have occurred,thus generating a “fast” lipase eluted with the voidvolume (peak 1). To check whether the two lipaseactivities observed in peaks 1 and 2 were the resultof the same enzyme, various fractions correspon-ding to these two peaks were subjected to SDS-PAGE analysis (Fig. 1A, insert). In both cases, asingle protein band corresponding to a molecularmass of around 29.5 kDa was observed (Fig. 1A).
As established previously (25), SSF and SmFlipases are identical proteins. However, the lipasescorresponding to peak 2 were found to have differ-ent physicochemical and kinetic properties. It wassuggested that some nonproteic compounds fromthe initial culture medium might be noncovalentlyassociated with the lipase produced using the SSFprocedure. Because the apparent molecular massof the SSF lipase was not found to be significantlydifferent from that of the SmF lipase (as estimatedfrom the elution of peak 2 in both cases), onepossible explanation for the differences observedwas that small detergent monomers (Triton X-100)might have bound tightly to the lipase.
3.2. Effects of Surfactants on LipaseActivity
The SmF and SSF Rh. homothallicus lipaseswere purified to homogeneity (25) and foundto have specific activities of 8600 ± 680 U/mgand 10,700 ± 390 U/mg, respectively, usingtrioctanoin as substrate. To test the hypothesisthat the kinetic properties of Rh. homothallicuslipase might be affected by the association withthe SSF lipase with detergent molecules, theSmF lipase was preincubated with variousamphiphilic molecules at concentrations abovetheir CMC. Tween 80, Triton X-100, and NaTDCwere found to significantly increase (by 25% to92%) the lipase activity (Table 1). It is worthnoting that once the lipase sample was diluted in
Effect of Nonionic Surfactants 209
MOLECULAR BIOTECHNOLOGY Volume 35, 2007
the pH-stat vessel, the final detergent concen-tration was found to be below the CMC ineach case.
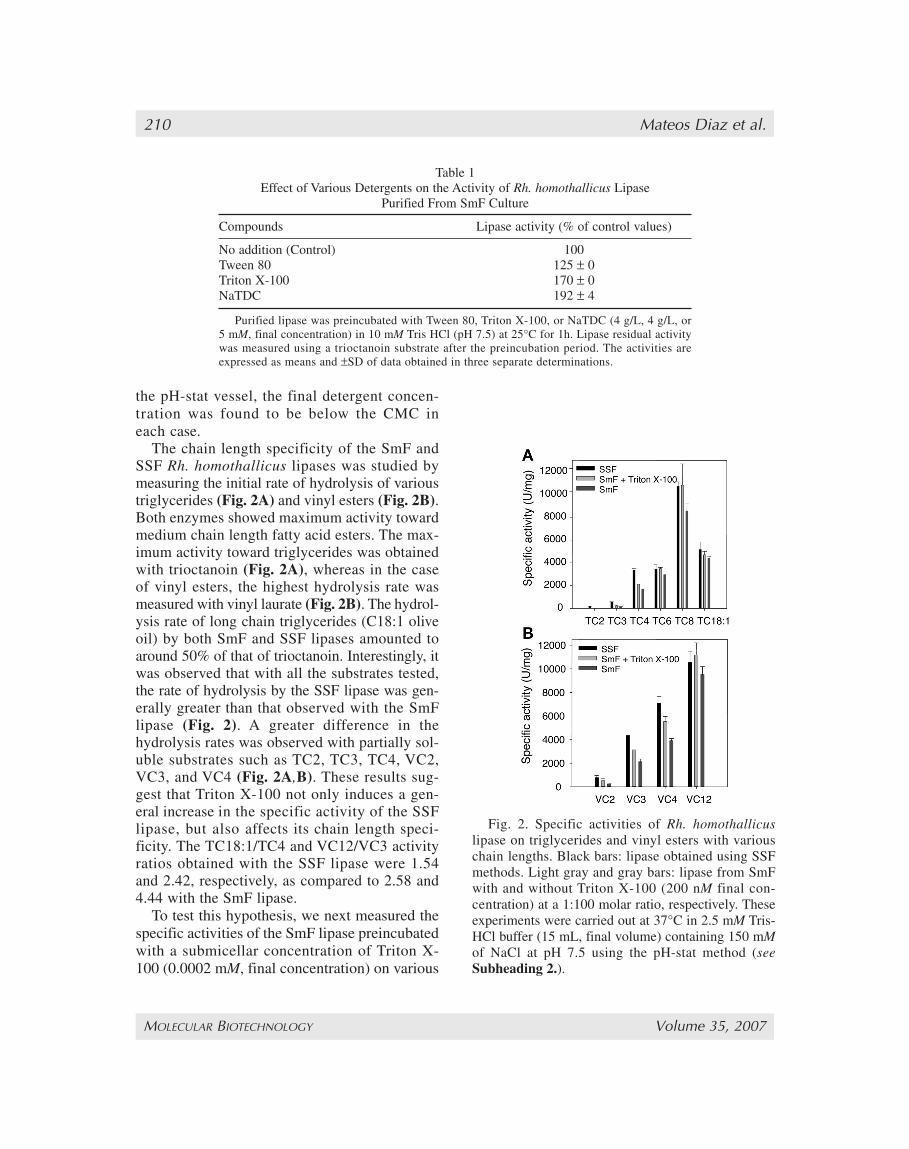
The chain length specificity of the SmF andSSF Rh. homothallicus lipases was studied bymeasuring the initial rate of hydrolysis of varioustriglycerides (Fig. 2A) and vinyl esters (Fig. 2B).Both enzymes showed maximum activity towardmedium chain length fatty acid esters. The max-imum activity toward triglycerides was obtainedwith trioctanoin (Fig. 2A), whereas in the caseof vinyl esters, the highest hydrolysis rate wasmeasured with vinyl laurate (Fig. 2B). The hydrol-ysis rate of long chain triglycerides (C18:1 oliveoil) by both SmF and SSF lipases amounted toaround 50% of that of trioctanoin. Interestingly, itwas observed that with all the substrates tested,the rate of hydrolysis by the SSF lipase was gen-erally greater than that observed with the SmFlipase (Fig. 2). A greater difference in thehydrolysis rates was observed with partially sol-uble substrates such as TC2, TC3, TC4, VC2,VC3, and VC4 (Fig. 2A,B). These results sug-gest that Triton X-100 not only induces a gen-eral increase in the specific activity of the SSFlipase, but also affects its chain length speci-ficity. The TC18:1/TC4 and VC12/VC3 activityratios obtained with the SSF lipase were 1.54and 2.42, respectively, as compared to 2.58 and4.44 with the SmF lipase.
To test this hypothesis, we next measured thespecific activities of the SmF lipase preincubatedwith a submicellar concentration of Triton X-100 (0.0002 mM, final concentration) on various
210 Mateos Diaz et al.
MOLECULAR BIOTECHNOLOGY Volume 35, 2007
Fig. 2. Specific activities of Rh. homothallicuslipase on triglycerides and vinyl esters with variouschain lengths. Black bars: lipase obtained using SSFmethods. Light gray and gray bars: lipase from SmFwith and without Triton X-100 (200 nM final con-centration) at a 1:100 molar ratio, respectively. Theseexperiments were carried out at 37°C in 2.5 mM Tris-HCl buffer (15 mL, final volume) containing 150 mMof NaCl at pH 7.5 using the pH-stat method (seeSubheading 2.).
Table 1Effect of Various Detergents on the Activity of Rh. homothallicus Lipase
Purified From SmF Culture
Compounds Lipase activity (% of control values)
No addition (Control) 100Tween 80 125 ± 0Triton X-100 170 ± 0NaTDC 192 ± 4
Purified lipase was preincubated with Tween 80, Triton X-100, or NaTDC (4 g/L, 4 g/L, or5 mM, final concentration) in 10 mM Tris HCl (pH 7.5) at 25°C for 1h. Lipase residual activitywas measured using a trioctanoin substrate after the preincubation period. The activities areexpressed as means and ±SD of data obtained in three separate determinations.
triglyceride and vinyl ester substrates. As shownin Fig. 2, the specific activities of the Triton X-100-treated SmF lipase were found to be inter-mediate between the specific activities of SSFand SmF lipases on various triglycerides (Fig. 2A)and vinyl esters (Fig. 2B).
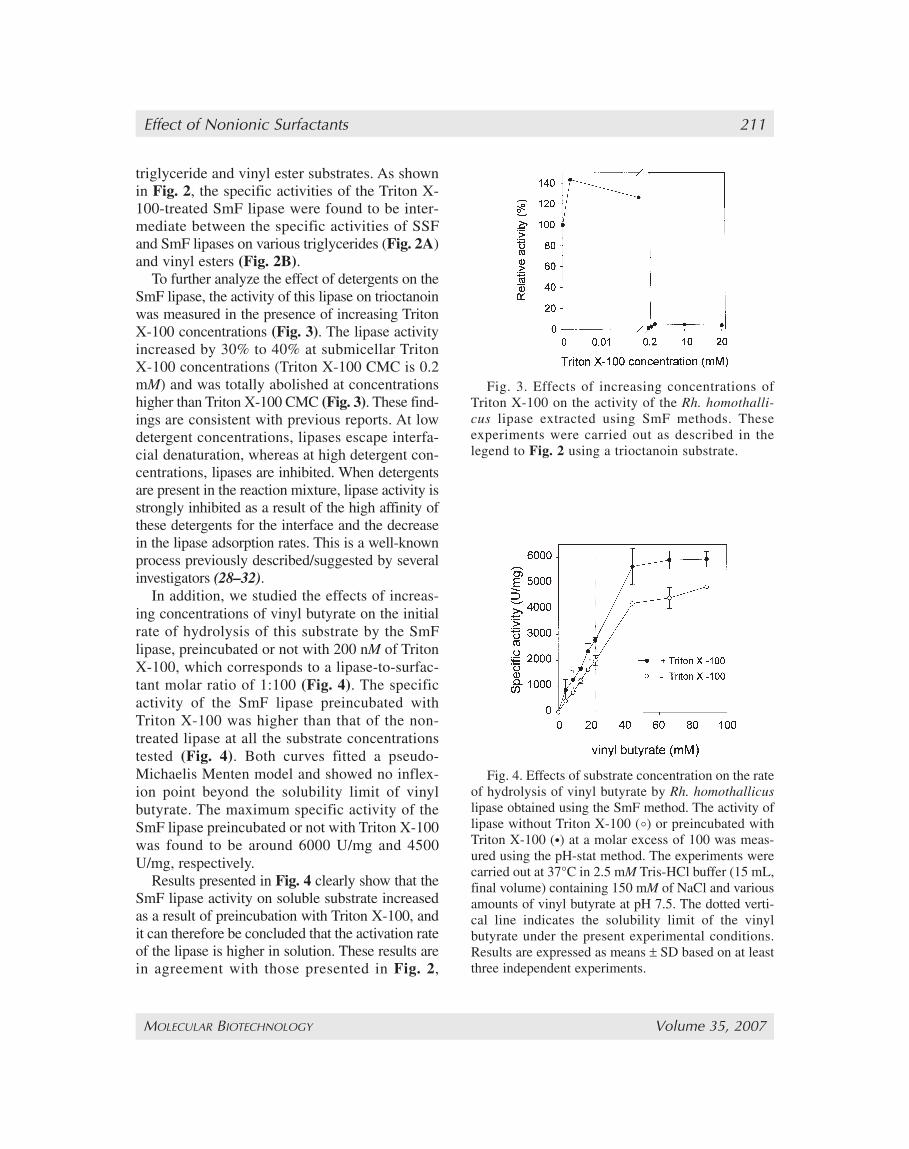
To further analyze the effect of detergents on theSmF lipase, the activity of this lipase on trioctanoinwas measured in the presence of increasing TritonX-100 concentrations (Fig. 3). The lipase activityincreased by 30% to 40% at submicellar TritonX-100 concentrations (Triton X-100 CMC is 0.2mM) and was totally abolished at concentrationshigher than Triton X-100 CMC (Fig. 3). These find-ings are consistent with previous reports. At lowdetergent concentrations, lipases escape interfa-cial denaturation, whereas at high detergent con-centrations, lipases are inhibited. When detergentsare present in the reaction mixture, lipase activity isstrongly inhibited as a result of the high affinity ofthese detergents for the interface and the decreasein the lipase adsorption rates. This is a well-knownprocess previously described/suggested by severalinvestigators (28–32).
In addition, we studied the effects of increas-ing concentrations of vinyl butyrate on the initialrate of hydrolysis of this substrate by the SmFlipase, preincubated or not with 200 nM of TritonX-100, which corresponds to a lipase-to-surfac-tant molar ratio of 1:100 (Fig. 4). The specificactivity of the SmF lipase preincubated withTriton X-100 was higher than that of the non-treated lipase at all the substrate concentrationstested (Fig. 4). Both curves fitted a pseudo-Michaelis Menten model and showed no inflex-ion point beyond the solubility limit of vinylbutyrate. The maximum specific activity of theSmF lipase preincubated or not with Triton X-100was found to be around 6000 U/mg and 4500U/mg, respectively.
Results presented in Fig. 4 clearly show that theSmF lipase activity on soluble substrate increasedas a result of preincubation with Triton X-100, andit can therefore be concluded that the activation rateof the lipase is higher in solution. These results arein agreement with those presented in Fig. 2,
Effect of Nonionic Surfactants 211
MOLECULAR BIOTECHNOLOGY Volume 35, 2007
Fig. 3. Effects of increasing concentrations ofTriton X-100 on the activity of the Rh. homothalli-cus lipase extracted using SmF methods. Theseexperiments were carried out as described in thelegend to Fig. 2 using a trioctanoin substrate.
Fig. 4. Effects of substrate concentration on the rateof hydrolysis of vinyl butyrate by Rh. homothallicuslipase obtained using the SmF method. The activity oflipase without Triton X-100 (°) or preincubated withTriton X-100 (•) at a molar excess of 100 was meas-ured using the pH-stat method. The experiments werecarried out at 37°C in 2.5 mM Tris-HCl buffer (15 mL,final volume) containing 150 mM of NaCl and variousamounts of vinyl butyrate at pH 7.5. The dotted verti-cal line indicates the solubility limit of the vinylbutyrate under the present experimental conditions.Results are expressed as means ± SD based on at leastthree independent experiments.

opening; and (2) Triton X-100 may interact with thesubstrate and either prevent interfacial lipase denat-uration by lowering the surface tension (increasedactivity levels occur at low detergent concentra-tions) or increase the substrate concentration insolution in the presence of partly soluble esters andtherefore, increase the activation of the lipase insolution. It is difficult to choose between these twopossible modes of action on the basis of the resultsobtained here. The fact that Rh. homothallicus lipaseis inhibited by E600 at a higher rate in the presenceof Triton X-100 suggests, however, that the openconformation of the lid is favored by Triton X-100molecules. These results are reminiscent of the pre-viously described process of “interfacial activation-based molecular (bio)-imprinting” of lipases bydetergent (34). Lipases, including the fungal lipasefrom Rhizopus miehei, which is homologous toRh. homothallicus, were found in the latter studyto display a much higher level of activity in water-restricted media/environments after being preincu-bated with detergent such as Triton X-100 andfreeze drying. In line with the present conclusions,we also hypothesized that the presence of detergentmolecules associated with the lipase may havefavored its open “activated” conformation.
AcknowledgmentsWe acknowledge the financial support from
CONACyT. We wish also to thank Dr. R. Vergerand Dr. H. Chahinian (CNRS Marseille) for theirhelpful advice and discussions during the prepa-ration of the manuscript, and Dr. Jessica Blancfor revising the English manuscript.
References1. Schmid, R. D. and Verger, R. (1998) Lipases: interfa-
cial enzymes with attractive applications. Angew.Chem. Int. Ed. 37, 1608–1633.
2. Haas, M. J., Bailey, D. G., Baker, W., et al. (1999)Biochemical and molecular biological characterizationof a lipase produced by the fungus Rhizopus delemar.Fett-Lipid 101, 364–370.
3. Kohno, M., Kugimiya, W., Hashimoto,Y., and Morita,Y.(1994) Purification, characterization, and crystalliza-tion of two types of lipase from Rhizopus niveus.Biosci. Biotechnol. Biochem. 58, 1007–1012.
4. Beer, H. D., McCarthy, J. E., Bornscheuer, U. T., andSchmid, R. D. (1998) Cloning, expression, character-ization and role of the leader sequence of a lipase from
Rhizopus oryzae. Biochim. Biophys. Acta 1399,173–180.
5. Kohno, M., Funatsu, J., Mikami, B., Kugimiya,W.,Matsuo, T., and Morita, Y. (1996) The crystal structureof lipase II from Rhizopus niveus at 2.2 A resolution.J. Biochem. (Tokyo) 120, 505–510.
6. Ben Salah, A., Sayari, A., Verger, R., and Gargouri, Y.(2001) Kinetic studies of Rhizopus oryzae lipaseusing monomolecular film technique. Biochimie 83,463–469.
7. Derewenda, Z. S. (1994) Structure and function oflipases. Adv. Protein Chem. 45, 1–52.
8. Brzozowski, A. M., Derewenda, U., Derewenda, Z. S.,et al. (1991) A model for interfacial activation in lipasesfrom the structure of a fungal lipase–inhibitor complex.Nature 351, 491–494.
9. Derewenda, U., Brzozowski, A. M., Lawson, D. M.,and Derewenda, Z. S. (1992) Catalysis at the interface:the anatomy of a conformational change in a trigly-ceride lipase. Biochemistry 31, 1532–1541.
10. Egloff, M.-P., Marguet, F., Buono, G., Verger, R.,Cambillau, C., and van Tilbeurgh, H. (1995) The 2.46Å resolution structure of the pancretic lipase-colipasecomplex inhibited by a C11 alkyl phosphonate.Biochemistry 34, 2751–2762.
11. Hermoso, J., Pignol, D., Kerfelec, B., Crenon, I.,Chapus, C., and Fontecilla-Camps, J. C. (1996)Lipase activation by nonionic detergents. The crystalstructure of the porcine lipase-colipase-tetraethyleneglycol monooctyl ether complex. J Biol Chem 271,18007–18016.
12. Ferrer, M., Plou, F. J., Nuero, O. M., Reyes, F., andBallesteros, A. (2000) Purification and properties of alipase from Penicillium chrysogenum isolated fromindustrial wastes. J. Chem. Technol. Biotechnol. 75, 1–8.
13. Rodriguez, J. A., Mateos, J. C., Nungaray, J., et al. (2006)Improving lipase production by nutrient source modifi-cation using Rhizopus homothallicus cultured in solidstate fermentation. Process Biochem. 41, 2264–2269.
14. Lee, M. W. and Severson, D. L. (1994) Partial purifi-cation of a diacylglycerol lipase from bovine aorta.Biochem. J. 298, 213–219.
15. Snellman, E. A., Sullivan, E. R., and Colwell, R. R.(2002) Purification and properties of the extracellu-lar lipase, LipA, of Acinetobacter sp. RAG-1. Eur.J. Biochem. 269, 5771–5779.
16. Gupta, N., Rathi, P., Singh, R., Goswami, V. K., andGupta, R. (2005) Single-step purification of lipasefrom Burkholderia multivorans using polypropylenematrix. Appl. Microbiol. Biotechnol. 67, 648–653.
17. Misset, O., Gerritse, G., Jaeger, K. E., et al. (1994) Thestructure–function relationship of the lipases fromPseudomonas aeruginosa and Bacillus subtilis. ProteinEng. 7, 523–529.
18. Gonzalez-Navarro, H., Bano, M. C., and Abad, C.(2001) The closed/open model for lipase activation.Addressing intermediate active forms of fungal
Effect of Nonionic Surfactants 213
MOLECULAR BIOTECHNOLOGY Volume 35, 2007

opening; and (2) Triton X-100 may interact with thesubstrate and either prevent interfacial lipase denat-uration by lowering the surface tension (increasedactivity levels occur at low detergent concentra-tions) or increase the substrate concentration insolution in the presence of partly soluble esters andtherefore, increase the activation of the lipase insolution. It is difficult to choose between these twopossible modes of action on the basis of the resultsobtained here. The fact that Rh. homothallicus lipaseis inhibited by E600 at a higher rate in the presenceof Triton X-100 suggests, however, that the openconformation of the lid is favored by Triton X-100molecules. These results are reminiscent of the pre-viously described process of “interfacial activation-based molecular (bio)-imprinting” of lipases bydetergent (34). Lipases, including the fungal lipasefrom Rhizopus miehei, which is homologous toRh. homothallicus, were found in the latter studyto display a much higher level of activity in water-restricted media/environments after being preincu-bated with detergent such as Triton X-100 andfreeze drying. In line with the present conclusions,we also hypothesized that the presence of detergentmolecules associated with the lipase may havefavored its open “activated” conformation.
AcknowledgmentsWe acknowledge the financial support from
CONACyT. We wish also to thank Dr. R. Vergerand Dr. H. Chahinian (CNRS Marseille) for theirhelpful advice and discussions during the prepa-ration of the manuscript, and Dr. Jessica Blancfor revising the English manuscript.
References1. Schmid, R. D. and Verger, R. (1998) Lipases: interfa-
cial enzymes with attractive applications. Angew.Chem. Int. Ed. 37, 1608–1633.
2. Haas, M. J., Bailey, D. G., Baker, W., et al. (1999)Biochemical and molecular biological characterizationof a lipase produced by the fungus Rhizopus delemar.Fett-Lipid 101, 364–370.
3. Kohno, M., Kugimiya, W., Hashimoto,Y., and Morita,Y.(1994) Purification, characterization, and crystalliza-tion of two types of lipase from Rhizopus niveus.Biosci. Biotechnol. Biochem. 58, 1007–1012.
4. Beer, H. D., McCarthy, J. E., Bornscheuer, U. T., andSchmid, R. D. (1998) Cloning, expression, character-ization and role of the leader sequence of a lipase from
Rhizopus oryzae. Biochim. Biophys. Acta 1399,173–180.
5. Kohno, M., Funatsu, J., Mikami, B., Kugimiya,W.,Matsuo, T., and Morita, Y. (1996) The crystal structureof lipase II from Rhizopus niveus at 2.2 A resolution.J. Biochem. (Tokyo) 120, 505–510.
6. Ben Salah, A., Sayari, A., Verger, R., and Gargouri, Y.(2001) Kinetic studies of Rhizopus oryzae lipaseusing monomolecular film technique. Biochimie 83,463–469.
7. Derewenda, Z. S. (1994) Structure and function oflipases. Adv. Protein Chem. 45, 1–52.
8. Brzozowski, A. M., Derewenda, U., Derewenda, Z. S.,et al. (1991) A model for interfacial activation in lipasesfrom the structure of a fungal lipase–inhibitor complex.Nature 351, 491–494.
9. Derewenda, U., Brzozowski, A. M., Lawson, D. M.,and Derewenda, Z. S. (1992) Catalysis at the interface:the anatomy of a conformational change in a trigly-ceride lipase. Biochemistry 31, 1532–1541.
10. Egloff, M.-P., Marguet, F., Buono, G., Verger, R.,Cambillau, C., and van Tilbeurgh, H. (1995) The 2.46Å resolution structure of the pancretic lipase-colipasecomplex inhibited by a C11 alkyl phosphonate.Biochemistry 34, 2751–2762.
11. Hermoso, J., Pignol, D., Kerfelec, B., Crenon, I.,Chapus, C., and Fontecilla-Camps, J. C. (1996)Lipase activation by nonionic detergents. The crystalstructure of the porcine lipase-colipase-tetraethyleneglycol monooctyl ether complex. J Biol Chem 271,18007–18016.
12. Ferrer, M., Plou, F. J., Nuero, O. M., Reyes, F., andBallesteros, A. (2000) Purification and properties of alipase from Penicillium chrysogenum isolated fromindustrial wastes. J. Chem. Technol. Biotechnol. 75, 1–8.
13. Rodriguez, J. A., Mateos, J. C., Nungaray, J., et al. (2006)Improving lipase production by nutrient source modifi-cation using Rhizopus homothallicus cultured in solidstate fermentation. Process Biochem. 41, 2264–2269.
14. Lee, M. W. and Severson, D. L. (1994) Partial purifi-cation of a diacylglycerol lipase from bovine aorta.Biochem. J. 298, 213–219.
15. Snellman, E. A., Sullivan, E. R., and Colwell, R. R.(2002) Purification and properties of the extracellu-lar lipase, LipA, of Acinetobacter sp. RAG-1. Eur.J. Biochem. 269, 5771–5779.
16. Gupta, N., Rathi, P., Singh, R., Goswami, V. K., andGupta, R. (2005) Single-step purification of lipasefrom Burkholderia multivorans using polypropylenematrix. Appl. Microbiol. Biotechnol. 67, 648–653.
17. Misset, O., Gerritse, G., Jaeger, K. E., et al. (1994) Thestructure–function relationship of the lipases fromPseudomonas aeruginosa and Bacillus subtilis. ProteinEng. 7, 523–529.
18. Gonzalez-Navarro, H., Bano, M. C., and Abad, C.(2001) The closed/open model for lipase activation.Addressing intermediate active forms of fungal
Effect of Nonionic Surfactants 213
MOLECULAR BIOTECHNOLOGY Volume 35, 2007

enzymes by trapping of conformers in water-restrictedenvironments. Biochemistry 40, 3174–3183.
19. Bano, M. C., Gonzalez-Navarro, H., and Abad, C.(2003) Long-chain fatty acyl-CoA esters induce lipaseactivation in the absence of a water-lipid interface.Biochim. Biophys. Acta 1632, 55–61.
20. Jutila, A., Zhu, K., Patkar, S. A., Vind, J., Svendsen,A., and Kinnunen, P. K. (2000) Detergent-induced con-formational changes of Humicola lanuginosa lipasestudied by fluorescence spectroscopy. Biophys. J. 78,1634–1642.
21. Iwai, M., Shimada, Y., and Tsujisaka, Y. (1980)Modification of Rhizopus delemar lipase by its bindingwith phospholipids. J. Biochem. (Tokyo) 88, 533–538.
22. Shimada, Y., Tominaga, Y., Iwai, M., and Tsujisaka, Y.(1983) Increase in the activity of Rhizopus delemarlipase on water-soluble esters by its binding withphosphatidylcholine. J. Biochem. (Tokyo) 93, 1655–1660.
23. Shimada, Y., Iwai, M., and Tsujisaka, Y. (1981)Reversibility of the modification of Rhizopus delemarlipases by phosphatidylcholine. J. Biochem. (Tokyo)89, 937–942.
24. Sugihara, A., Gargouri, Y., Piéroni, G., Rivière, C.,Sarda, L., and Verger, R. (1986) Activities and inter-facial properties of Rhizopus delemar and porcine pan-creatic lipases after treatment with phospholipids.Biochemistry 25, 3430–3434.
25. Diaz, J. C. M., Rodríguez, J. A., Roussos, S., et al.(2006) Lipase from the thermotolerant fungusRhizopus homothallicus is more thermostable whenproduced using solid state fermentation than liquid fer-mentation procedures. Enzyme Microb. Tech. 39,1042–1050.
26. Bradford, M. M. (1976) A rapid and sensitive methodfor quantitation of microgram quantities of proteinutilizing the principle of protein-deye binding. Anal.Biochem. 72, 248–254.
27. Laemmli, U. K. (1970) Cleavage of structural proteinsduring the assembly of the head of bacteriophage T4.Nature 227, 680–685.
28. Momsen, W. E. and Brockman, H. L. (1976) Inhibitionof pancreatic lipase B activity by taurodeoxycholate andits reversal by colipase. J. Biol. Chem. 251, 384–388.
29. Momsen, W. E. and Brockman, H. L. (1976) Effectsof colipase and taurodeoxycholate on the catalytic andphysical properties of pancreatic lipase B at an oilwater interface. J. Biol. Chem. 251, 378–383.
30. Morgan, R. G., Barrowman, J., and Borgstrom, B.(1969) The effect of sodium taurodesoxycholate andpH on the gel filtration behavior of rat pancreatic pro-tein and lipases. Biochim. Biophys. Acta 175, 65–75.
31. Brockerhoff, H. (1973) A model of pancreatic lipaseand the orientation of enzymes at interfaces. Chem.Phys. Lipids 10, 215–222.
32. Canioni, P., Julien, R., Rathelot, J., and Sarda, L. (1976)Inhibition of sheep pancreatic lipase activity againstemulsified tributyrin by non-ionic detergents. Lipids12, 393–397.
33. Derewenda, U., Swenson, L., Wei, Y., et al. (1994)Conformational lability of lipases observed in theabsence of an oil–water interface: crystallographic stud-ies of enzymes from the fungi Humicola lanuginosa andRhizopus delemar. J. Lipid Res. 35, 524–534.
34. Mingarro, I., Abad, C., and Braco, L. (1995) Interfacialactivation-based molecular bioimprinting of lipolyticenzymes. Proc. Natl. Acad. Sci. USA 92, 3308–3312.
214 Mateos Diaz et al.
MOLECULAR BIOTECHNOLOGY Volume 35, 2007