early ordovician conodonts from tarutao island, southern peninsular thailand
TRANSCRIPT

EARLY ORDOVICIAN CONODONTS FROM TARUTAO
ISLAND, SOUTHERN PENINSULAR THAILAND
by SACHIKO AGEMATSU* , KATSUO SASHIDA* , SIROT SALYAPONGSE� and
APSORN SARDSUD�*Graduate School of Life and Environmental Sciences, University of Tsukuba, Ibaraki 305-8572, Japan; e-mail: [email protected]
�Geological Survey Division, Department of Mineral Resources, Rama VI, Bangkok 10400, Thailand
Typescript received 5 March 2007; accepted in revised form 24 July 2008
Abstract: Early Ordovician conodont faunas of the Thung
Song Formation on Tarutao Island, southern peninsular
Thailand, consist of 14 known species belonging to 17 gen-
era, and eight undescribed species. Utahconus tarutaoensis
and Filodontus tenuis are new species. Three conodont zones:
the Rossodus manitouensis Zone, the Utahconus tarutaoensis
Zone and the Filodontus tenuis Zone, in ascending order, are
defined in the study sections. These are coeval with the inter-
val from the Rossodus manitouensis Zone to the Acodus delta-
tus-Oneotodus costatus Zone of the standard zonation in the
North American Midcontinent. Based on the conodonts
studied here and fossils previously reported from Tarutao
Island, the Thung Song Formation is early Tremadocian to
middle Arenig (Ibexian) in age. This formation is lithostrati-
graphically subdivided into the S1 to S5 members, and our
study sections consist of the S1 to S3 members. These strata
accumulated on a shelf in the Early Ordovician. The deposi-
tional environments of the limestones making up the S1 and
S3 members were in deeper-shelf conditions. Limestone and
shale of the S2 member formed in a shallow-water, high-
energy environment.
Key words: biostratigraphy, conodonts, Early Ordovician,
Tarutao Island, Thailand, Thung Song Formation.
The Sibumasu Block is one of the smaller continental
blocks that existed in northern Gondwana during the
early and middle Palaeozoic. It extends today from Myan-
mar and southwestern China in the north, to Sumatora
Island in the south (Metcalfe 1999). Lower Palaeozoic
sedimentary rocks are widely distributed on this block
and contain macro- and microfossils (Kobayashi 1964;
Kobayashi and Hamada 1964a, b; Igo and Koike 1967,
1968; Hamada et al. 1975; Hahn and Siebenhuner 1982;
Stait and Burrett 1984; Boucot et al. 1999; Fortey and
Cocks 1998; Agematsu et al. 2006a–c, 2007). Southern
peninsular Thailand is one of the principal areas for
palaeontological and stratigraphic studies of the Lower
Palaeozoic of the Sibumasu Block (Wongwanich et al.
1990). In particular, Cambrian and Lower Ordovician
rocks are well exposed on Tarutao Island, which is near
the west coast of peninsular Thailand (Cocks et al. 2005).
Several faunas with trilobites, nautiloids, brachiopods and
conodonts have been reported from Tarutao Island (Tera-
oka et al. 1982; Fortey and Cocks 1998; Cocks et al.
2005). These fossil collections added materially to the
detailed biostratigraphy of Lower Palaeozoic sequences in
Southeast Asia. We investigated Ordovician rocks and
collected limestone samples on Tarutao Island in 2004,
and recovered abundant Early Ordovician conodonts. The
present study describes and correlates these conodont fau-
nas and discusses the stratigraphy of the Ordovician
sequence on Tarutao Island.
GEOLOGICAL SETTING
It is the general consensus that the geology of Thailand
comprises two principal continental blocks, the western
Sibumasu Block and the eastern Indochina Block (Buno-
pas 1981; Metcalfe 1999). According to Bunopas (1981),
these two blocks have a suture zone between them that is
bordered by the western Sukhothai and eastern Loei-
Petchabun Fold belts. The area of the suture zone is
referred to as Nan-Uttradit in the north and Sra Kaeo-
Chanthaburi in the south (Text-fig. 1A). The Cambrian
and Ordovician rocks in Thailand have been referred to
the Tarutao and Thung Song groups, respectively (Buno-
pas 1981). The Tarutao Group, which comprises clastic
rocks, is typically exposed on Tarutao Island and is also
present in the Kanchanaburi and Chiang Mai areas of
western and northwestern Thailand respectively (Bunopas
1992). The Thung Song Group is mainly composed of
carbonate rocks and widely distributed in northwestern,
western and southern Thailand (Bunopas 1992). The type
[Palaeontology, Vol. 51, Part 6, 2008, pp. 1435–1453]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2008.00810.x 1435

locality of this group is in Nakhon Sri Thammarat prov-
ince, southern peninsular Thailand.
Tarutao Island, which extends 12 km north to south
and 5 km east to west, lies about 25 km west of Satun in
southern peninsular Thailand (Text-fig. 1B). Cocks et al.
(2005) correlated the Lower Palaeozoic sequences of the
Sibumasu Block and redefined the Tarutao and Thung
Song Groups on Tarutao Island as the Tarutao Formation
and the Thung Song Formation (Thung Song Limestone),
respectively. According to these workers, the Tarutao For-
mation is subdivided into lower, middle and upper mem-
bers. The lower member mainly comprises thickly bedded
mudstones, and the middle member consists of sandstone
strata. The upper member is composed of sandstone beds
with limestone lenses. Although Cocks et al. (2005) did
not subdivide the Thung Song Formation, Teraoka et al.
(1982) classified limestones of this formation into five
members, which are, in ascending order, the S1, S2, S3,
S4 and S5 members. According to Teraoka et al. (1982),
the S1 member is composed of thinly bedded limestones
interlaminated with shales, and the S2 member is charac-
terized by alternations of limestones and red, grey and
green shales. The S3 member is divided into a lower part,
which is similar to the S1 member, and an upper part of
thickly bedded limestone. The lithology of the S4 member
is almost the same as that of the S2 member. The S5
member is composed of thinly and thickly bedded lime-
stones. In this study, we follow the nomenclature of Tera-
oka et al. (1982) and Cocks et al. (2005) and use the
Tarutao and Thung Song Formations and the S1 to S5
members.
Cocks et al. (2005) reviewed occurrences of fossils,
including trilobites, nautiloids, brachiopods and con-
odonts, that have been reported from Tarutao Island.
They concluded that the Tarutao and Thung Song forma-
tions on Tarutao Island range approximately from Cam-
brian to Middle Ordovician (Arenig) in age, and that the
Cambrian–Ordovician boundary probably lies within the
middle member of the Tarutao Formation.
LITHOLOGY AND DEPOSITIONALENVIRONMENTS
Conodonts were recovered from sections A and B on
Tarutao Island (Text-fig. 1B–C). Section A lies along the
southeastern coast of the island, and Section B is situated
along a path to a hilltop in at the northwestern part of
the island. Section A mainly comprises limestones that
are partially intercalated with shale beds. The rocks strike
N 10� to 20�E and dip 30� to 50� east (Text-fig. 1C). Ter-
aoka et al. (1982) showed the distribution of the Tarutao
and Thung Song formations on this island and presumed
that the boundary of these formations could be found on
the southeastern coast between the sample localities TAR-
23 and TAR-24 of the present study. However, the
boundary of the Tarutao and Thung Song formations on
the southeastern coast is actually to the west of sample
locality TAR-16 (Text-fig. 1C). The sequence measured in
this section is about 1000 m thick, and is classified into
three members as follows, in ascending order: the S1
member of limestones with thin shale beds (about
A B C
TEXT -F IG . 1 . A, index map showing the study area. B, map of Tarutao Island and a distribution of the Thung Song Formation. C,
geological map of southern coast of the island and sampling points in Section A.
1436 P A L A E O N T O L O G Y , V O L U M E 5 1

600 m); the S2 member of alternating limestone and shale
(about 200 m); the lower part of the S3 member of grey,
bedded limestones (about 200 m). These three members
are conformable (Text-fig. 2).
Limestones in the S1 member are subdivided into a
lowermost part (about 30 m thick) and a main part
(about 570 m thick) (Text-fig. 2). Rocks in the lowest
part of the S1 member consists of dark grey, argillaceous
TEXT -F IG . 2 . Lihological column with stratigraphic distributions of conodont species, conodont zones and sea-level change in
Section A.
A G E M A T S U E T A L . : E A R L Y O R D O V I C I A N C O N O D O N T S F R O M T A R U T A O I S L A N D , T H A I L A N D 1437

limestone alternated with black shale beds, and are
1–30 cm in thickness. The main part of the S1 member
comprises thickly bedded, grey limestones. These lime-
stones are micrite or pelmicrite. The S2 member consists
of interbeds of grey limestones, 0.1–1 m thick, and red
shales (Text-fig. 2). Seen under a microscope, the lime-
stone is micrite or biomicrite with abundant quartz
grains, which frequently form cross-laminations. The
shale beds, mostly red in colour, are composed of silt-
size quartz and calcite grains and contain some fine
sand-size grains. Grey, bedded limestone in the lower
part of the S3 member has beds 1–30 cm thick (Text-
fig. 2) and are micrites or pelmicrites similar to those of
the S1 member.
Section B is made up of 40 m of limestone beds, which
strike N 40�W to N 70�W and dip 30� to the northeast.
This sequence comprises grey, massive limestones inter-
bedded with laminated limestones. One interval of mas-
sive limestone is more than 50 cm thick. The limestones
of this section correspond to member S1.
Depositional environments
The depositional environments of Section A are inferred
from field observations and thin section analyses. The
reconstructed paleoenvironments and sea-level changes
within Section A are shown in Text-fig. 2. The lime-
stone facies in the lowermost part of the S1 member
implies that its depositional environment was a shelf
with high-energy conditions with a continuous influx of
terrigenous grains. The environment became a deeper-
water shelf owing to transgression, and persisted during
a limestone deposition of the S1 member. Interbeds of
limestone and shale of the S2 member represent the
existence of a low-sea-level period. This interval was
characterized by the inflow of large amounts of terrige-
nous material and strong currents, which formed cross
laminations. The alternation of limestone and shale
implies cyclical changes of environmental factors, such
as depth, salinity, temperature and turbidity. These fac-
tors may have been more or less influenced by trans-
gressions and regressions. The lower part of the S3
member is thought to have been deposited under deeper
water conditions, similar to the environment of the S1
member.
CONODONT BIOSTRATIGRAPHY
Most samples from each stratigraphic section yielded con-
odonts. Seventeen species belonging to 14 genera, and
eight unidentified species, are identified (Table 1). The
following three conodont zones are established in Section
A, in ascending order: the Lower Ordovician Rossodus
manitouensis Zone, the Utahconus tarutaoensis Zone and
the Filodontus tenuis Zone (Text-fig. 2).
Rossodus manitouensis Range Zone
This zone covers the S1 member in Section A (Text-
fig. 2). The lower and upper boundaries of this zone are
marked by the first and last occurrences of Rossodus man-
itouensis Repetski and Ethington, 1983. This zone yields
other species, including Variabiloconus bassleri (Furnish,
1938), Teridontus obesus Ji and Barnes, 1994, Utahconus
longipinnatus Ji and Barnes, 1994 and Acanthodus uncina-
tus Furnish, 1938. Acanthodus lineatus Furnish, 1938 and
Scolopodus sp. are present in the lower part of the zone.
Coniform indet. A, B and C are included in the upper
part. Drepanoistodus forceps (Lindstrom, 1955) appears in
the uppermost part of this zone and ranges into younger
strata of the section (Text-fig. 2).
Utahconus tarutaoensis Range Zone
This zone ranges through the upper 100 m of the S2
member in Section A (Text-fig. 2), and coincides with the
stratigraphical range of Utahconus tarutaoensis sp. nov.
Drepanoistodus pervetus Nowlan, 1985, Paroistodus num-
arcuatus (Lindstrom 1955) and Striatodontus prolificus Ji
and Barnes, 1994 are another species characteristic of this
zone (Text-fig. 2). This zone also includes U. longipinna-
tus, D. forceps, Protoprioniodus yapu Cooper, 1981 and
Coniform indet. H.
Filodontus tenuis Range Zone
This is the uppermost zone of Section A. The base and
top of the zone, about 100 m and 30 m below the top
of the section, respectively, are defined by the appearance
and disappearance of Filodontus tenuis sp. nov. Other
representative species of this zone are D. forceps and
Parapanderodus striatus (Graves and Ellison, 1941) (Text-
fig. 2). S. prolificus, Scolopodus sp., P. yapu, Acodus sp.,
Coniform indet. D and Coniform indet. F also occur in
this zone.
Comparison between sections A and B on Tarutao Island
Section B includes five species belonging to five genera
and two unidentified species (Table 1). Teridontus obe-
sus, Variabiloconus bassleri and Acanthodus uncinatus
occur through the section. The lower and middle parts
1438 P A L A E O N T O L O G Y , V O L U M E 5 1

of this section yields Coniform indet. E and Drepanodus
arcuatus Pander, 1856, respectively. Utahconus longipinn-
atus and Coniform indet. A are present in the upper
part of this section. Section B does not includes Rosso-
dus manitouensis. However, this section is inferred to be
comparable to the R. manitouensis Zone of Section A,
because Section B shares all species, except for Coni-
form indet. E, with the zone of Section A. This fact
supports the above-mentioned correlation based on
lithostratigraphy.
CORRELATION
Correlations of the study sections with other Ordovician
sequences worldwide are possible. First, we will compare
Section A to Lower Ordovician successions in adjacent
areas. Next, the zonation established in Section A will be
correlated to the standard zonation of the North Ameri-
can Midcontinent (Ross et al. 1997; Webby et al. 2004),
to zones of the Midcontinent realm in British Columbia
(Pyle and Barnes 2002) and to shallow-water zones in
Newfoundland (Ji and Barnes 1994). These correlations
are shown in Text-fig. 3.
TABLE 1 . List of conodont species from sections A and B. Samples 3–14 and 16–47 are collected from Section B and Section A,
respectively. About 1.5 kg of limestone was processed for each sample.
TEXT -F IG . 3 . Correlation of the Ordovician conodont zones
of the study section with zonations of the North American
Midcontinent, North Atlantic, British Columbia and
Newfoundland.
A G E M A T S U E T A L . : E A R L Y O R D O V I C I A N C O N O D O N T S F R O M T A R U T A O I S L A N D , T H A I L A N D 1439

Comparison with conodont faunas in Thailand and
Malaysia
Metcalfe (1980) and Agematsu et al. (2006c) studied Early
Ordovician conodonts from Malaysia and Thailand,
respectively. Metcalfe (1980) recovered Ordovician con-
odonts from the Setul Limestone in the Peris area of pen-
insular Malaysia. He presented a list of the Early and
Middle Ordovician conodont faunas, which occurred at
localities 1, 2 and 3 in (Metcalfe 1980). According to
him, the ages of the faunas from localities 1 and 3 are
Early Ordovician and the fauna from locality 2 is Middle
Ordovician. The fauna from locality 1 is Tremadocian in
age, because it includes Loxodus bransoni Furnish, 1938,
which has been described from strata corresponding to
the Rossodus manitouensis Zone of the standard zonation
of the North American Midcontinent (Ethington and
Clark 1982). However, the fauna he studied does not con-
tain species in common with faunas of the present study.
On the basis of the occurrence of Serratognathus bilobatus
Lee, 1970, the fauna from locality 3 Arenig, and therefore
younger than the faunas from Tarutao Island.
Teraoka et al. (1982) first reported Ordovician cono-
dont faunas from Tarutao Island. They recovered Drepan-
odus arcuatus, Drepanoistodus basiovalis (Sergeeva, 1963),
Drepanoistodus forceps, Paroistodus parallelus (Pander,
1856), Paroistodus proteus (Lindstrom, 1955) and Proto-
panderodus rectus (Lindstrom, 1955) from limestones of
the S1 member and the lower part of the S3 member.
However, P. parallelus and P. proteus clearly indicate a
younger age than the faunas in our study. Teraoka et al.
(1982) photographically illustrated only D. arcuatus,
D. basiobalis and D. forceps without description, so it is
difficult to compare the details of their specimens with
conodonts in this study. Taking into account the poor
preservation of specimens in Teraoka et al. (1982), their
identification may be questionable.
Comparison with the zonation of the North American
Midcontinent
The standard zonation for Early Ordovician conodonts in
the North American Midcontinent, summarized by Ross
et al. (1997) and Webby et al. (2004), is shown in Text-
fig. 3. Rossodus manitouensis, Variabiloconus bassleri,
Acanthodus lineatus and Acanthodus uncinatus, which are
representative species of the R. manitouensis Zone of Sec-
tion A in this study, are common to the R. manitouensis
Zone in the Midcontinent area (Ethington and Clark
1981; Repetski and Ethington 1983; Landing et al. 1996;
Sweet and Tolbert 1997). D. forceps and Parapanderodus
striatus occur from the upper part of Section A in this
study and are relatively long-ranging species. The oldest
occurrence of P. striatus is in the Low-diversity interval of
the Midcontinent (Ethington and Clark 1981). Paroistodus
numarcuatus has not been described from the Midconti-
nent, but it is a representative species of the North Atlantic
region and has been reported from the upper Paltodus
deltifer Zone to the lower P. proteus Zone (Lofgren 1997).
Therefore, the R. manitouensis Zone in this study is coeval
with the R. manitouensis Zone in the North American Mid-
continent. Based on the stratigraphic range of Pa. striatus
and P. numarcuatus, the Utahconus tarutaoensis and Fil-
odontus tenuis zones of the study section are comparable to
the Low-diversity interval to the Acodus deltatus-Oneotodus
costatus Zone in the Midcontinent (Text-fig. 3).
Comparison with zonation in British Columbia
Pyle and Barnes (2002) used the Lower Ordovician zona-
tion of the Midcontinent realm for the Kechika Forma-
tion in British Columbia (Text-fig. 3). Because the
R. manitouensis Zone in the present study shares several
species, including R. manitouensis, V. bassleri, Utahconus
longipinnatus, A. lineatus and A. uncinatus, with the
R. manitouensis Zone of the Kechika Formation, these
zones are correlated with each other. T. obesus has been
described from the Iapetognathus Zone and D. pervetus
and S. prolificus from the R. manitouensis Zone of the
Kechika Formation (Pyle and Barnes 2002). The oldest
known Pa. striatus in British Columbia has been reported
from the Scolopodus subrex Zone. As mentioned above,
the upper limit of the occurrence of P. numarcuatus
is the P. proteus Zone in the North Atlantic region.
These lines of biostratigraphic evidence indicate that the
U. tarutaoensis and F. tenuis zones of the study section
are comparable to the S. subrex Zone and to the Acodus
kechikaensis Zone in British Columbia.
Comparison with zonation in Newfoundland
The Lower Ordovician shallow-water zones of the
St. George Group in Newfoundland, proposed by Ji and
Barnes (1994), are shown in Text-fig. 3. Several
representative Tarutao Island species are known from
Newfoundland. T. obesus, V. bassleri, R. manitouensis,
A. lineatus, A. uncinatus, U. longipinnatus and D. pervetus
occur from the R. manitouensis-Plycostatus sulcatus Zone
to the Colaptoconus floweri-Colaptoconus bolites Zone
of the St. George Group. S. prolificus ranges from the C.
floweri-C. bolites to S. prolificus-Striatodontus lanceolatus
zones in Newfoundland. Pa. striatus is one of the youngest
species in the St. George Group and occurs in the Para-
panderodus carlae-Paraserratognathus ovatus Zone and
overlying zones. Based on the stratigraphical range of
1440 P A L A E O N T O L O G Y , V O L U M E 5 1

these species and above-mentioned P. numarcuatus, zonal
correlation between the study section and the St. George
Group is as follows: the R. manitouensis Zone in the study
section is coeval with the R. manitouensis-P. sulcatus and
C. floweri-C. bolites zones; the U. tarutaoensis and F. tenuis
zones in this study are compared to the C. floweri-
C. bolites to S. prolificus-S. lanceolatus zones and the
P. carlae-S. ovatus Zone, respectively. The stratigraphic
ranges of S. prolificus and D. pervetus on Tarutao Island
are thought to extend into younger strata than do those
in other areas.
AGE OF THE THUNG SONGFORMATION
Text-figure 4 shows the stratigraphic sequence of the
Tarutao and Thung Song Formations and their ages,
based on conodonts in this study and previously reported
fossils. Section A in this study correlates lithostratigraphi-
cally with the interval from the S1 member to the lower
part of the S3 member in the Thung Song Formation. As
mentioned above, Teraoka et al. (1982) reported Ordo-
vician conodont faunas from the S1 and S3 members on
Tarutao Island, but several species were probably mis-
identified. Teraoka et al. (1982) also illustrated conodonts
from the uppermost part of the Tarutao Formation and
the S4 and S5 members of the Thung Song Formation.
These include abundant well-preserved specimens, in con-
trast with their faunas from the S1 and S3 members. The
presence of Chosonodina herfurthi Muller, 1964 indicates
that the uppermost strata of the Tarutao Formation also
correlate with the R. manitouensis Zone in the North
American Midcontinent. The faunas from the S4 and S5
members contain Acodus deltatus Lindstrom, 1955, and
Oepikodus evae (Lindstrom, 1955), which are early and
middle Arenig species. The ages of these faunas accords
with those in the present study.
Several macrofossils faunas have been reported from
the Thung Song Formation of Tarutao Island (Text-
fig. 4). Tremadocian brachiopods have been illustrated by
Fortey and Cocks (1998) from limestone in the S1 mem-
ber, which is exposed along the northwestern coast of the
island. Although Cocks et al. (2005) revised the age of
this fauna to Arenig, conodonts in the present study sug-
gest that the S1 member is Tremadocian in age. Stait and
Burrett (1984) also described nautiloids from the S1
member, which crops out along the northwestern coast of
the island. The sum of evidence is that the S1 member is
a Tremadocian sequence. Limestone in the S4 member,
which is distributed on Lae Tong Island, southeastern
Tarutao Island, contains Arenig nautiloids (Stait and Bur-
rett, 1984). This age coincides with that of conodonts
reported by Teraoka et al. (1982) from the S4 member.
To sum up the faunal evidence given above, the ages of
the S1, S2, S3 and S4 to S5 members of the Thung Song
Formation are early Tremadocian, middle Tremadocian,
late Tremadocian to early Arenig and middle Arenig,
respectively (Text-fig. 4).
CONCLUSION
The Thung Song Formation of Tarutao Island, southern
peninsular Thailand, was investigated bio- and litho-
stratigraphically. Early Ordovician conodonts occur from
study sections A and B and three conodont zones are
established in Section A. These zones are the Rossodus
manitouensis Range Zone, the Utahconus tarutaoensis
Range Zone and the Filodontus tenuis Range Zone, in
ascending order. Section B is equivalent to the R. manito-
uensis Zone of Section A. The zones of Section A are
comparable to the interval from the Rossodus manitouen-
sis Zone to the Acodus deltatus-Oneotodus costatus Zone
of the standard zonation in the North American Mid-
continent. The zones in the present study also correlate
with the intervals from the R. manitouensis Zone to the
Acodus kechikaensis Zone in British Columbia and the
R. manitouensis-Polycostatus sulcatus Zone to the Para-
panderodus carlae-Paraserratognathus ovatus Zone in
Newfoundland.
The Thung Song Formation is subdivided into S1–S5
members, and Section A of this study consists of the
S1–S3 members. Our data of conodonts, and fossils pre-
viously reported from Tarutao Island, imply that the
ages of the S1, S2, S3 and S4 to S5 members of the
TEXT -F IG . 4 . Column showing the stratigraphic sequences
from the Late Cambrian to the Early Ordovician on Tarutao
Island and relative positions of some of faunas reported from
the island.
A G E M A T S U E T A L . : E A R L Y O R D O V I C I A N C O N O D O N T S F R O M T A R U T A O I S L A N D , T H A I L A N D 1441

Thung Song Formation are early Tremadocian, middle
Tremadocian, late Tremadocian to early Arenig, and
middle Arenig, respectively. The depositional environ-
ments of the S1–S3 members are inferred to be as fol-
lows. The lowermost limestone of the S1 member was
deposited on a shallow-water shelf in the early Trem-
adocian. The environment changed to a deeper-water
shelf owing to transgression, and this condition persisted
during deposition of the limestones of the S1 member.
The middle Tremadocian was a low-sea-level interval.
The limestone and shale of the S2 member were depos-
ited in a shallow-water setting, which experienced
deeper water due to a transgression before the late
Tremadocian. Limestone in the S3 member accumulated
on a deeper shelf.
SYSTEMATIC PALAEONTOLOGY
Conodonts from the study section are shown in Plates
1–4. The illustrated specimens are deposited in the Insti-
tute of Geoscience, University of Tsukuba, with the prefix
IGUT. Our collection includes several taxa represented by
quite a few specimens. Therefore, 11 taxa, including
stratigraphically important species and new species, are
described here. Conodonts are only classified to genus
and species, because a suprageneric classification is still
uncertain. Element terminology essentially follows that of
Barnes et al. (1979).
Genus ACANTHODUS Furnish, 1938
Type species. Acanthodus uncinatus Furnish, 1938.
Remarks. Landing et al. (1996) revised the diagnosis of
Acanthodus as consisting of three elemental morphologies,
drepanodiform, scandodiform and suberectiform. The
apparatus of A. uncinatus in the Tarutao material is
composed of a (drepanodiform), c (suberectiform) and
e (scandodiform) elements, which is in accordance with
the definition of Landing et al. (1996).
Acanthodus uncinatus Furnish, 1938
Plate 2, figures 1–4
* 1938 Acanthodus uncinatus Furnish, p. 337–338, pl. 42,
fig. 30, text-fig. 2B.
1996 Acanthodus uncinatus Furnish; Ji and Barnes,
figs. 12.7–12.11.
. 1996 Acanthodus uncinatus Furnish; Landing et al.,
p. 669, 671, figs. 8.1–8.9, 9.13, 9.5–17.
2002 Acanthodus uncinatus Furnish; Pyle and Barnes,
p. 59, pl. 1, figs. 1–2.
2003 Acanthodus uncinatus Furnish; Landing et al.
fig. 10.7.
Material. Fifty-one specimens; 36 a, 4 c and 11 e elements
(IGUT-ag2166–2167, 2898–2946).
EXPLANATION OF PLATE 1
Figs 1–4. Rossodus manitouensis Repetski and Ethington, 1983. R. manitouensis Zone, Thung Song Formation. 1a–b, IGUT-ag3216,
sample TAR-21; posterior and aboral views of c element; ·150. 2a–b, IGUT-ag3076, sample TAR-33; lateral and aboral views of e
element; ·100. 3a–b, IGUT-ag3213, sample TAR-17; lateral and aboral views of a element ·100. 4a–b, IGUT-ag1978, sample
TAR-16; lateral and aboral views of a element; ·100.
Figs 5–7. Teridontus obesus Ji and Barnes, 1994. R. manitouensis Zone, Thung Song Formation. 5a–b, IGUT-ag3308, sample TAR-20;
lateral and aboral views of a ⁄ b element; ·150. 6a–b, IGUT-ag1941, sample TAR-6; lateral and aboral views of e element; ·100.
7a–b, IGUT-ag3283, sample TAR-8; posterior and aboral views of c element; ·150.
Figs 8–10. Utahconus longipinnatus Ji and Barnes, 1994. R. manitouensis and U. tarutaoensis zones, Thung Song Formation. 8a–ab,
IGUT-ag2951, sample TAR-17; posterior and aboral views of c element; ·100. 9a–b, IGUT-ag2959, sample TAR-32; lateral and
aboral views of e element; ·80. 10a–b, IGUT-ag3577, sample TAR-36; lateral and aboral views of a element; ·100.
Figs 11–12, 15–16. Variabiloconus bassleri (Furnish, 1938). R. manitouensis Zone, Thung Song Formation. 11a–b, IGUT-ag3388,
sample TAR-6; lateral and aboral views of e element; ·100. 12a, IGUT-ag3434, sample TAR-14; posterior view of c element; ·100.
15a–b, IGUT-ag1935, sample TAR-8, lateral and aboral views of a element; ·100. 16a–b, IGUT-ag1936, sample TAR-4; lateral and
aboral views of b element; ·100.
Figs 13–14. Paroistodus numarcuatus (Lindstrom, 1955). U. tarutaoensis Zone to Filodontus tenuis Zone, Thung Song Formation. 13a–
b, IGUT-ag3018, sample TAR-38; lateral and aboral views of r element; ·100. 14a–b, IGUT-ag3013, sample TAR-38; lateral and
aboral views of q element; ·100.
Figs 17–18. Drepanodus arcuatus Pander, 1856. R. manitouensis Zone, Thung Song Formation. 17a–b, IGUT-ag1951, sample TAR-6;
lateral and aboral views of q element; ·60. 18a–b, IGUT-ag3008, sample TAR-36; lateral and aboral views of q element; ·80.
Scale bars indicate 100 lm.
1442 P A L A E O N T O L O G Y , V O L U M E 5 1
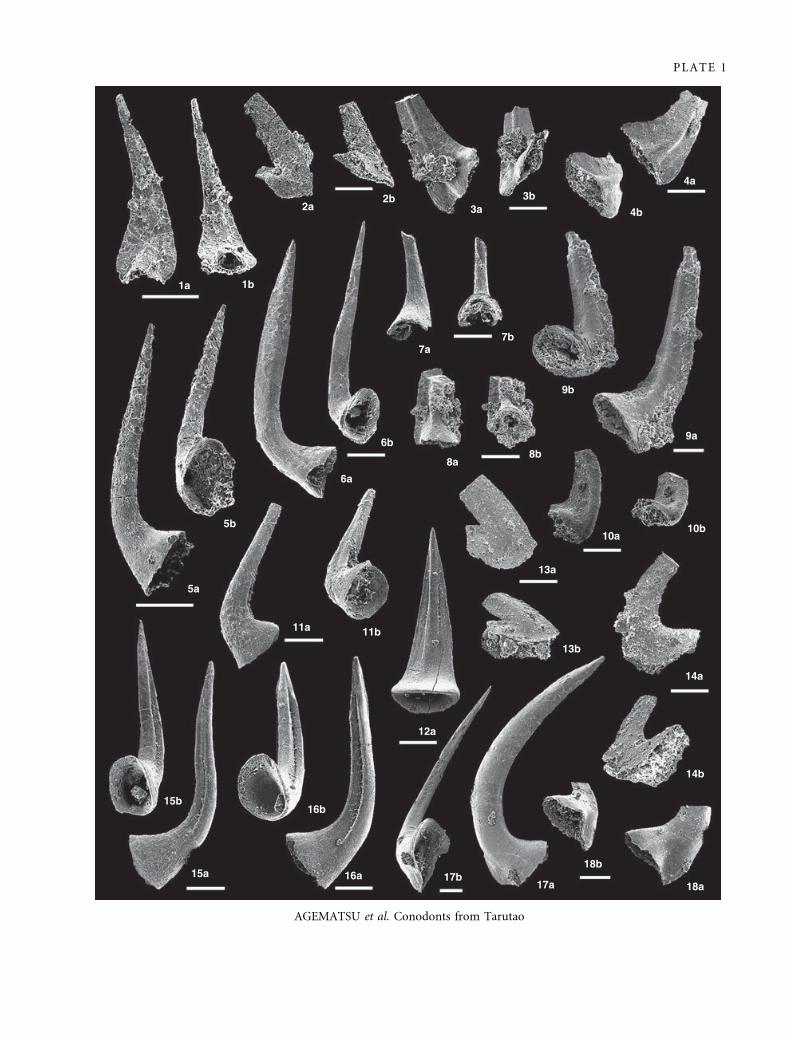
PLATE 1
AGEMATSU et al. Conodonts from Tarutao
1a
2a 3a
7a 7b
9b
10b 10a
13a
14a
14b
18b 17b 16a 15a
18a 17a
8a
6a
6b
5b
5a
11a
12a
11b
13b
16b 15b
8b
9a
4a 3b
4b 2b
1b

Remarks. Specimens in the Tarutao collection are identi-
fied with these forms. The a element of this study is a
synonym of ‘A. uncinatus’ and ‘D. suberectus’ of Furnish
(1938). The form of the c element described here equals
with ‘O. erectus’ of Furnish (1938). This is similar to the c
element of species of Drepanoistodus, but it is distin-
guished by having a straight and robust cusp and short
base. The e element of A. uncinatus in this study is a syn-
onym of ‘D. tortus’ of Furnish (1938). Mature elements of
this species are known to have serrations along the pos-
terior margin. Although specimens of this study are
poorly preserved and lack the distal part of cusp, some
elements show some serrations (Pl. 2, fig. 4). Landing
et al. (1996) also described a drepanodiform element,
which bears an antero-lateral costa, but the Tarutao speci-
mens do not contain this element. A larger collection is
necessary to more precise discussion.
Occurrence. A. uncinatus is widespread in the Rossodus
manitouensis Zone in North America and its equivalents
in North China, Siberia, Australia, Korea and Iran (Land-
ing et al. 1996). In this study, the species is from the
R. manitouensis Zone of the Thung Song Formation.
Genus DREPANOISTODUS Lindstrom, 1971
Type species. Oistodus forceps Lindstrom, 1955.
Drepanoistodus pervetus Nowlan, 1985
Plate 2, figure 8
* 1985 Drepanoistodus? pervetus Nowlan, p. 112–113, text-
figs 5.53–5.55, 6.1–6.3.
1981 Drepanoistodus? n. sp.; Landing and Barnes,
p. 1615–1616, pl. 2, figs. 1–2, 5, 7–8.
1994 Drepanoistodus pervetus Nowlan; Ji and
Barnes, p. 35–36, pl. 7, figs. 21–27, text-fig.
24B.
1996 Drepanoistodus pervetus Nowlan; Ji and Barnes, fig.
11.12–11.19.
2002 Drepanoistodus pervetus Nowlan; Pyle and Barnes,
p. 64, pl. 6, figs. 16–18.
Material. Four specimens (IGUT-ag2169, 2965, 3002–3003).
Remarks. Laterally compressed elements of this study
have a cusp with keeled anterior and posterior margins
and an antero-posteriorly expanded base. These speci-
mens are comparable with nongeniculate elements
described by Nowlan (1985).
Occurrence. This species has been described from the
uppermost Cambrian to Lower Ordovician strata in Can-
ada (Landing and Barnes 1981; Nowlan 1985; Ji and
Barnes 1994). The Utahconus tarutaoensis Zone of the
Thung Song Formation also yields D. pervetus on Tarutao
Island.
EXPLANATION OF PLATE 2
Figs 1–4. Acanthodus uncinatus Furnish, 1938. Rossodus manitouensis Zone, Thung Song Formaiton. 1a–b, IGUT-ag2946, sample TAR-
31; lateral and aboral views of c element; ·50. 2a–b, IGUT-ag2898, sample TAR-3; lateral and aboral views of a element; ·60. 3a–
b, IGUT-ag2167, sample TAR-13; lateral and aboral views of e element; ·50. 4a–b, IGUT-ag2166, sample TAR-13; lateral and
aboral views of e element; ·50.
Figs 5–7. Drepanoistodus forceps (Lindstrom, 1955). R. manitouensis Zone to Filodontus tenuis Zone, Thung Song Formation. 5a–b,
IGUT-ag2066, sample TAR-46; lateral and aboral views of a element; ·100. 6a–b, IGUT-ag3115, sample TAR-46; lateral and
aboral views of e element; ·100. 7a–b, IGUT-ag2005, sample TAR-46; lateral and aboral views of c element; ·100.
Fig 8. Drepanoistodus pervetus Nowlan, 1985. Utahconus tarutaoensis Zone, Thung Song Formation. 8a–b, IGUT-ag2169, sample TAR-
38; lateral and aboral views; ·100.
Fig 9. Acanthodus lineatus Furnish, 1938. R. manitouensis Zone, Thung Song Formation. 9a–b, IGUT-ag2001, sample TAR-17; lateral
and aboral views; ·100.
Figs 10–12. Striatodontus prolificus Ji and Barnes, 1994. U. tarutaoensis Zone to F. tenuis Zone, Thung Song Formation. 10a–b, IGUT-
ag2170, sample TAR-41; lateral and aboral views of e element; ·100. 11a–b, IGUT-ag3257, sample TAR-46; lateral and aboral
views of b element; ·100. 12a–b, IGUT-ag3256, sample TAR-46; lateral and aboral views of b element; ·150.
Fig 13. Protoprioniodus yapu Cooper, 1981. U. tarutaoensis Zone to F. tenuis Zone, Thung Song Formation. 13a–b, IGUT-ag3202,
sample TAR-42; lateral and aboral views of b element; ·80.
Figs 14–15. Acodus sp. F. tenuis Zone, Thung Song Formation. 14a–b, IGUT-ag2000, sample TAR-46; lateral and aboral views of b
element; ·50. 15a–b, IGUT-ag2949, sample TAR-46; lateral and aboral views of g element; ·100.
Figs 16–17. Scolopodus sp. R. manitouensis Zone to F. tenuis Zone, Thung Song Formation. 16a–b, IGUT-ag1966, sample TAR-18;
lateral and aboral views; ·80. 17a–b, IGUT-ag1968, sample TAR-46; lateral and aboral views; ·100.
Scale bars indicate 100 lm.
1444 P A L A E O N T O L O G Y , V O L U M E 5 1
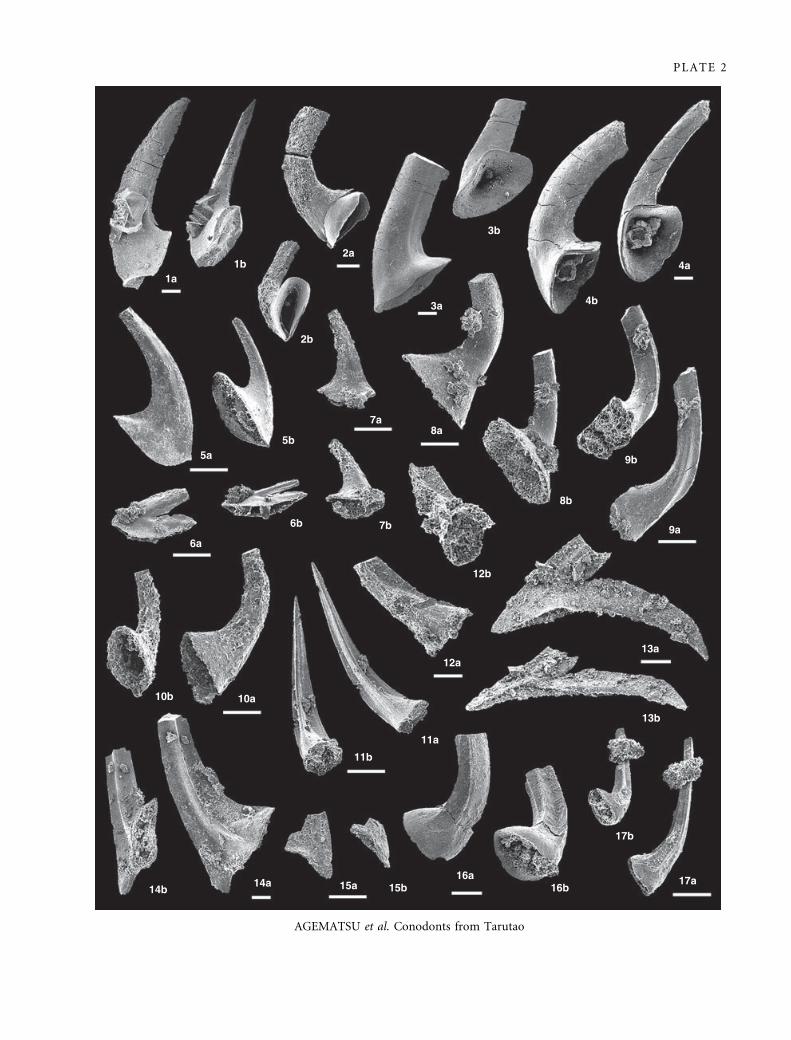
PLATE 2
AGEMATSU et al. Conodonts from Tarutao
1a 1b
2b
2a
3a 4b
9b
9a
8a 7a
7b 6b
5b 5a
6a
10b
11b
11a
12a 13a
13b
17b
17a 16b
16a 15b 14b 15a 14a
10a
8b
12b
4a
3b

Genus FILODONTUS Pyle, Barnes and Ji, 2003
Type species. Scolopodus filosus Ethington and Clark, 1964.
Remarks. Pyle et al. (2003) erected the genus Filodontus
and assigned a single species, Filodontus filosus, to this
genus. They considered Filodontus as a bi-membrate
apparatus. Elements of this genus are characterized by
bearing many extremely fine longitudinal costae from the
tip of the cusp to above the basal margin.
Filodontus tenuis sp. nov.
Plate 3, figures 5–8.
Derivation of name. From the Latin, slim.
Holotype. a element IGUT-ag1987, Thung Song Formation,
Tarutao Island, Thailand; Early Ordovician; Plate 3, figure 6.
Material. Twenty-two specimens; 17 a and 5 e elements (IGUT-
g1987, 2017–2018, 3133–3151).
Diagnosis. Apparatus comprises coniform a and e ele-
ments, with a proclined, slender cusp and an unexpanded,
cylindrical base. The cusp and the upper part of the base
bear many extremely fine longitudinal costae. The cross
section of the a element is circular through the unit. The
posterior margin of the cusp of the e element is laterally
compressed and sharply edged.
Description. a elements are simple and coniform with a circu-
lar cross-section throughout their length. The unit slightly
curves posteriorly at one-third to one-half length from the
basal margin. The cusp gradually tapers toward the tip. The
base is unexpanded and cylindrical. All sides of the surface are
ornamented by fine longitudinal costae that extend from the
tip of the cusp to the upper part of the base. The costae of a
distal part are thicker than those of a basal part of elements
and are 24 lm in maximum width. The surface is smooth at
the lower part of the base. The height of the basal cavity is
1.5 times of the diameter of the basal margin. The lower part
of the base of some specimens twists anteriorly. The basal
margin is straight in lateral view and circular in aboral view.
e elements are similar to the a elements in the curvature
of the unit and ornamentation of the surface. However, the e
EXPLANATION OF PLATE 3
Figs 1–4. Parapanderodus striatus (Graves and Ellison, 1941). Filodontus tenuis Zone, Thung Song Formation. 1a–b, IGUT-ag2010,
sample TAR-46; lateral and aboral views of a element; ·100. 2a–b, IGUT-ag2012, sample TAR-46; lateral and aboral views of b
element; ·200. 3a–b, IGUT-ag3167, sample TAR-46; posterior and aboral views of c element; ·100. 4a–b, IGUT-ag3182, sample
TAR-46; lateral and aboral views of e element; ·200.
Figs 5–8. Filodontus tenuis n. sp. F. tenuis Zone, Thung Song Formation. 5a–b, paratype; IGUT-ag3135, sample TAR-46; lateral and
aboral views of a element; ·150. 6a–b, holotype; IGUT-ag1987, sample TAR-46; lateral and aboral views of a element; ·200.
7a–b, paratype; IGUT-ag2018, sample TAR-46; lateral and aboral views of e element; ·150. 8a–b, paratype; IGUT-ag2017, sample
TAR-46; lateral and aboral views of e element; ·150.
Fig 9. Coniform indet. E. R. manitouensis Zone, Section B, Thung Song Formation. 9a–b, IGUT-ag3463, sample TAR-4; posterior and
aboral views; ·100.
Fig 10. Coniform indet. H. Utahconus tarutaoensis Zone to F. tenuis Zone, Thung Song Formation. 10a–b, IGUT-ag1964, sample
TAR-46; lateral and aboral views; ·100.
Figs 11–14. Utahconus tarutaoensis sp. nov. U. tarutaoensis Zone, Thung Song Formation. 11a–b, paratype; IGUT-ag3357, sample
TAR-42; lateral and aboral views of e element; ·100. 12a–b, paratype; IGUT-ag1991, sample TAR-42; lateral and aboral views of b
element; ·80. 13a–b, holotype; IGUT-ag1988, sample TAR-39; lateral and aboral views of a element; ·100. 14a–b, paratype;
IGUT-ag3326, sample TAR-39; lateral and aboral views of a element; ·100.
Fig 15. Coniform indet. A. R. manitouensis Zone, Thung Song Formation. 15a–b, IGUT-ag3449, sample TAR-30; lateral and aboral
views; ·80.
Fig 16. Coniform indet. B. R. manitouensis Zone, Thung Song Formation. 16a–b, IGUT-ag1982, sample TAR-32; lateral and aboral
views; ·60.
Figs 17–18. Coniform indet. C. R. manitouensis Zone, Thung Song Formation. 17a–b, IGUT-ag1975, sample TAR-27; lateral and
aboral views; ·100. 18a–b, GUT-ag1974, sample TAR-27; lateral and aboral views; ·100.
Fig 19. Coniform indet. D. F. tenuis Zone, Thung Song Formation. 19a–b, IGUT-ag3463, sample TAR-46; lateral and aboral views;
·200.
Fig 20. Coniform indet. F. F. tenuis Zone, Thung Song Formation. 20a–b, IGUT-ag1969, sample TAR-46; lateral and aboral views;
·60.
Fig 21. Coniform indet. G. Thung Song Formation. 21a–b, IGUT-ag1985, sample TAR-36; lateral and aboral views; ·100.
Scale bars indicate 100 lm.
1446 P A L A E O N T O L O G Y , V O L U M E 5 1
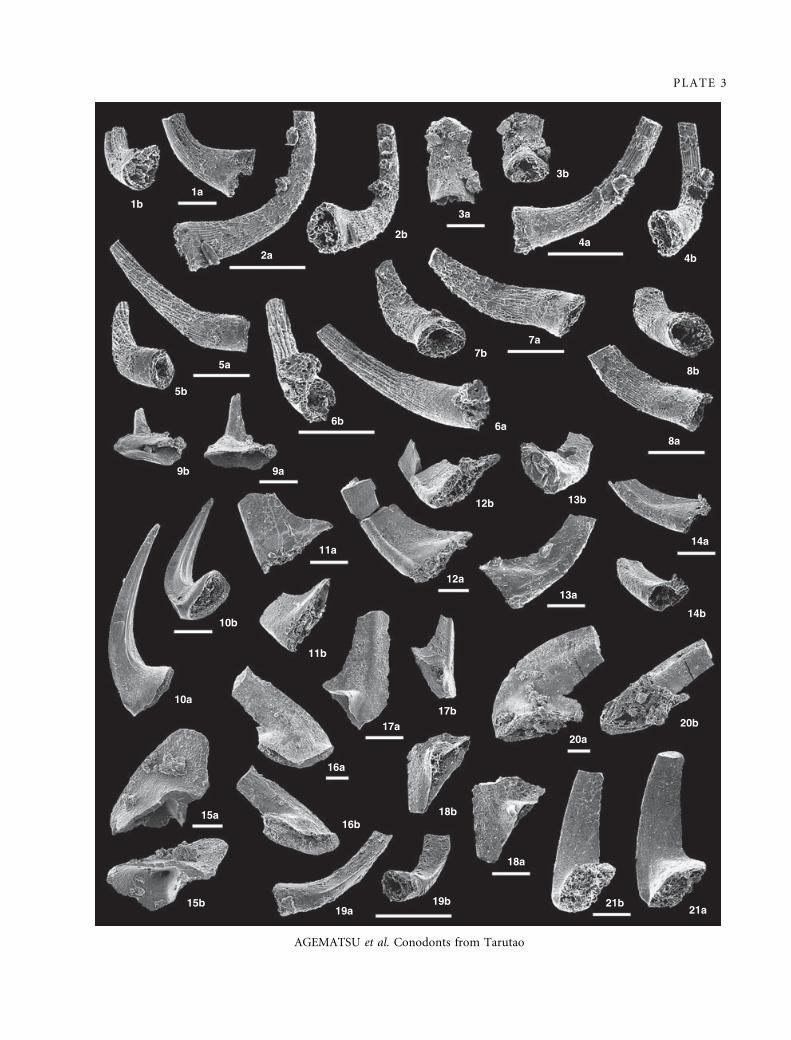
PLATE 3
AGEMATSU et al. Conodonts from Tarutao
1b 1a
2a
5a
5b
9b
10b
10a
16a
17a 17b
18b 16b
15a
15b 19a
18a
19b 21b 21a
20a
20b
11b
9a
11a
2b
3a
3b
4a
4b
8b
8a
7a 7b
6a
12b 13b
13a
12a
14a
14b
6b

element differs from the a element in that the posterior margin
is keeled at the cusp and that the height of the basal cavity is
twice of the diameter of the basal margin. The lower part of the
base twists anteriorly.
Remarks. Pyle et al. (2003) described two element mor-
photypes of F. filosus. One is a slender form, and the
other is stout and squat. Both types have a reclined cusp
and a more or less expanded base. All specimens of F. ten-
uis in this study are more slender, and have a proclined
cusp and a shorter base than elements of F. filosus of Pyle
et al. (2003). Repetski (1982) reported proclined coniform
elements as S. filosus, which are similar to the a element
of F. tenuis, but his specimens bear thinner and weaker
costae on the surface than those of F. tenuis.
Occurrence. This species occurs in the F. tenuis Zone of
the Thung Song Formation on Tarutao Island.
Genus PARAPANDERODUS Stouge, 1984
Type species. Parapanderodus arcuatus Stouge, 1984.
Parapanderodus striatus (Graves and Ellison, 1941)
Plate 3, figures 1–4
* 1941 Drepanodus striatus n. sp. Graves and Ellison, p. 11,
pl. 1, figs. 3, 12.
. 2000 Parapanderodus striatus (Graves and Ellison);
Johnston and Barnes, p. 30, pl. 12, figs. 11, 13–15,
17–19.
2003 Parapanderodus striatus (Graves and Ellison); Pyle
and Barnes, figs. 12.18–12.19.
2003 Parapanderodus striatus (Graves and Ellison); Pyle
et al., figs.8.9–8.10.
Material. Forty-eight specimens; 15 a, 30 b, 1 c and 2 e elements
(IGUT-ag2010, 2012, 3152–3197).
Remarks. Four types of coniform elements, a, b, c and e,
are included in the apparatus. All specimens have a long
cusp and a slender base, and are finely striated. Ji and
Barnes (1994) defined a, b, c and e elements as an appara-
tus of this species. Specimens of this study are in accor-
dance with the description of these elements.
Occurrence. P. striatus has been described from the Lower
and Middle Ordovician strata in North America and
Argentina (Johnston and Barnes 2000). The oldest occur-
rence of this species is known from sequences in Utah
(Ethington and Clark 1981) and Argentina (Albanesi
et al. 1998), corresponding to the Low diversity interval
of the standard zonation in North American Midconti-
nent. The Filodontus tenuis Zone of the Thung Song
Formation in this study also yields this species.
Genus PAROISTODUS Lindstrom, 1971
Type species. Oistodus parallelus Pander, 1856.
Paroistodus numarcuatus (Lindstrom, 1955)
Plate 1, figures 13–14
* 1955 Drepanodus numarcuatus Lindstrom, pp. 564–565,
pl. 2, figs 48–49, text-fig. 3I.
. 1997 Paroistodus numarcuatus (Lindstrom); Lofgren,
pp. 921–922, text-figs 2O–U, 3A–G, 4A–K.
Material. Sixteen specimens; 15 q and 1 r elements (IGUT-
ag3010–3025).
Remarks. Elements of this species are distinguished by
Lofgren (1997) into seven types, a, b, c, d, e, f and g.
Johnston and Barnes (2000) redescribed a Paroistodus
apparatus consisting of two element forms, r (oistodi-
form) and q (drepanodiform) elements. We follow the
concept of Johnston and Barnes (2000).
Occurrence. This species is widely known from the
uppermost Tremadocian strata in the North Atlantic areas
(Lofgren 1997). In this study, the species is present in the
Utahconus tarutaoensis and Filodontus tenuis zones of the
Thung Song Formation on Tarutao Island.
Genus ROSSODUS Repetski and Ethington, 1983
Type species. Rossodus manitouensis Repetski and Ethington,
1983.
Rossodus manitouensis Repetski and Ethington, 1983
Plate 1, figures 1–4
1982 New genus A, n. sp. A, Repetski, p. 56–57, pl. 28,
figs. 1–4.
* 1983 Rossodus manitouensis Repetski and Ethington,
p. 293–300, figs. 1A–V, 2A–T, 3A–R, 4A–D.
1993 Rossodus manitouensis Repetski and Ethington;
Wang, p. 199, pl. 7, figs. 15–17, 25.
. 1994 Rossodus manitouensis Repetski and Ethington; Ji
and Barnes, p. 56, pl. 17, figs. 1–9.
1994 Rossodus manitouensis Repetski and Ethington; Seo
et al., figs. 7.23–7.29.
1995 Rossodus manitouensis Repetski and Ethington;
Chen et al., pl. 1, fig. 2.
1448 P A L A E O N T O L O G Y , V O L U M E 5 1

1996 Rossodus manitouensis Repetski and Ethington;
Ji and Barnes, figs. 13.13–13.20.
1996 Rossodus manitouensis Repetski and Ethington;
Landing et al. 1996, figs. 5.18, 5.20, 5.22–5.24.
1999 Rossodus manitouensis Repetski and Ethington;
Lofgren et al., pl. 3, figs. 13, 15, 21.
2002 Rossodus manitouensis Repetski and Ethington;
Pyle and Barnes, p. 100, pl. 13, figs. 1–6.
2005 Rossodus manitouensis Repetski and Ethington;
Wu et al., pl. 1, fig. 5.
Material. Thirteen specimen; 4 a, 7 c and 2 e elements (IGUT-
ag1978, 1980, 2966, 3076, 3209–3217).
Remarks. The apparatus of this genus consists of follow-
ing four types of element; acontiodiform a element, drep-
anodiform b element, suberect and symmetrical c element
and oistodiform e element (Ji and Barnes 1994). Speci-
mens of this study are classified into asymmetrical a,
symmetrical c and oistdontiform e elements.
Occurrence. This is a representative species of the R. manitouensis
zone and its equivalents in North American Midcontinent, New-
foundland, New York, British Columbia, Quebec and South
China (Landing et al. 1986). Lofgren et al. (1999) reported this
species from the middle Tremadocian strata in the Baltica area.
On Tarutao Island, this species range through the R. manitouensis
Zone of the Thung Song Formation.
Genus STRIATODONTUS Ji and Barnes, 1994
Type species. Striatodontus prolificus Ji and Barnes, 1994.
Striatodontus prolificus Ji and Barnes, 1994
Plate 2, figures 10–12
* 1994 Striatodontus prolificus Ji and Barnes, p. 61,
pl. 20, figs. 1–26, text-fig. 36A.
1998 Striatodontus prolificus Ji and Barnes; Lehnert
et al., p. 57, pl. 1, figs. 1–4, 7, 8.
non 2002 Striatodontus prolificus Ji and Barnes; Pyle and
Barnes, p. 83, pl. 15, figs. 1–4.
2003 Striatodontus prolificus Ji and Barnes; Landing
et al., figs. 11.11–11.15.
Materials. Nine specimens; 7 b and 2 e elements (IGUT-ag2170,
3254–3258, 3260–3262).
Description. Specimens in the Tarutao collection are slender
coniform and distinguished into b and e elements. b elements
are transitional triangulariform and have a proclined cusp and
a short base. Two types of the b element are recognized in
this study. One is a symmetrical form having smooth lateral
faces and a wide longitudinal depression on the posterior face.
The other is asymmetrical and characterized by deep antero-
and postero-lateral grooves on the inner face. These grooves
are bounded by sharply developed anterior, inner lateral and
posterior costae. The grooves and costae are carried from the
tip of the cusp to the mid base. The outer face is rounded.
The base slightly expands posteriorly and is a circle in aboral
view.
e elements are symmetrical with an erect cusp. The anterior
margin is narrowly rounded. A median groove on the posterior
face ranges from the tip of the cusp to above the basal margin.
The base slightly flares antero-posteriorly. The basal margin is
elliptical in aboral view.
Remarks. Ji and Barnes (1994) included a, b, c and e ele-
ments in the apparatus of this species. The material of
this study does not contain the a and c elements. The b
elements show two forms, as mentioned above, without a
transitional form. It may be necessary to classify the b ele-
ments in more detail. Figured specimens of Pyle and
Barnes (2002) seem to differ from the type specimens of
Ji and Barnes (1994).
Occurrence. S. prolificus has been reported from the
Colaptoconus floweri-Colaptoconus bolites to Protopanderodus in-
constans-Scolopodus subrex zones of the St. George Group in
Newfoundland (Ji and Barnes 1994) and from the lower part of
the Ponon Trehue Formation in Argentina (Lehnert et al.
1998). This species is also included in the Utahconus tarutao-
ensis Zone and the Filodontus tenuis Zone of the Thung Song
Formation on Tarutao Island.
Genus TERIDONTUS Miller, 1980
Type species. Oneotodus nakamurai Nogami, 1967.
Remarks. Ji and Barnes (1994) re-defined the apparatus
of this genus to comprise four types of element: a, b, c
and e. The e elements are partly identified with species of
Monocostodus described by Miller (1980). On the other
hand, Nicoll (1994) recognized six forms, a, b, c, d, f and
g elements, in an apparatus. We follow the definition of Ji
and Barnes (1994), because the Tarutao material contains
laterally compressed e elements.
Teridontus obesus Ji and Barnes, 1994
Plate 1, figures 5–7
* 1994 Teridontus obesus Ji and Barnes, p. 65, 66, pl. 24,
figs. 10–17, text-fig. 37B.
1998 Teridontus obesus Ji and Barnes; Rao and Tortello,
p. 37, 38, pl. 2, figs. 1–7.
2002 Teridontus obesus Ji and Barnes; Pyle and Barnes,
p. 71, pl. 15, figs. 20–22.
A G E M A T S U E T A L . : E A R L Y O R D O V I C I A N C O N O D O N T S F R O M T A R U T A O I S L A N D , T H A I L A N D 1449

Materials. Fifty-six specimens; 38 a ⁄ b, 4 c and 14 e elements
(IGUT-ag1941, 3263–3317).
Remarks. Ji and Barnes (1994) described four types of
element as components of the apparatus of this species.
The Tarutao collection contains a ⁄ b, c and e elements.
a ⁄ b elements are compared with the a and b elements
of Ji and Barnes (1994). The form of a ⁄ b elements grad-
ually varies from symmetrical to subsymmetrical. A few
specimens bear faint groove on one lateral face, recog-
nized by Ji and Barnes (1994). These specimens are dis-
tinguished from elements of Teridontus nakamurai
(Nogami 1967) by the a ⁄ b element with a short,
expanded base and the c element with a straight, slender
cusp and a flared base.
Occurrence. This species has been reported from the Polycostatus
falsioneotensis-Rossodus tenuis Zone to the Rossodus manitouen-
sis-Polycostatus sulcatus Zone of the St. George Group in New-
foundland (Ji and Barnes 1994), the lower Tremadocian
Cardonal Formation in Argentina (Rao and Tortello 1998), the
lowest Tremadocian strata of the Kechika Formation in British
Columbia (Pyle and Barnes 2002) and the R. manitouensis Zone
of the Thung Song Formation in this study.
Genus UTAHCONUS Miller, 1980
Type species. Paltodus utahensis Miller, 1969.
Remarks. According to Pyle and Barnes (2002), an appa-
ratus of Utahconus species contains three or four types of
element characterized by compression along the cusp and
rounded to triangular basal outline.
Utahconus longipinnatus Ji and Barnes, 1994
Plate 1, figures 8–10.
* 1994 Utahconus longipinnatus n. sp. Ji and Barnes,
p. 66, 67, pl. 25, figs. 1–8, text-fig. 38A.
1985 Utahconus aff. U. utahensis (Miller, 1969); Nowlan,
p. 117, figs. 5.45–5:52.
p 1996 Scalpellodus longipinnatus (Ji and Barnes, 1994);
Landing et al., p. 675, 676, figs. 7.9–7.10, 7.12–19,
7.24, 9.22, 9.23 [non figs. 7.8, 7.11 (= indet.)].
2002 Utahconus longipinnatus Ji and Barnes; Pyle and
Barnes, p. 72, pl. 17, figs. 1–3.
Material. Twenty specimens; 3a, 8c and 9e elements (IGUT-
ag1959, 1960, 2951–2957, 2959–2964, 3000, 3575–3578).
Remarks. Ji and Barnes (1994) recognized a, c and e ele-
ments as constituting the apparatus of this species. Speci-
mens in this study are symmetrical to asymmetrical
coniform, which are also distinguished into a, c and e
elements, and characteristics of the element conform to
the description of Ji and Barnes (1994). Cusps of these
elements are longer than U. utahensis specimens.
Occurrence. The Lower Ordovician strata in North America
(Nowlan 1985; Ji and Barnes 1994; Landing et al. 1996; Pyle and
Barnes 2002) and the Rossodus manitouensis and Utahconus taru-
taoensis zones in the Thung Song Formation, in this study,
include U. longipinnatus.
Utahconus tarutaoensis sp. nov.
Plate 3, figures 11–14
1982 Protopanderodus? n. sp. 1 s.f.; Repetski, p. 41, pl. 18,
figs. 4, 6.
Derivation of name. Refers to Tarutao Island where the study
sections crop out.
Holotype. A element IGUT-ag1988; Plate 3, figure 13; Thung
Song Formation, Tarutao Island, southern peninsular Thailand;
Early Ordovician.
Material. Thirty-six specimens; 18 a, 7 b and 11 e elements
(IGUT-ag1988, 1991, 3323–3351, 3353–3357).
Diagnosis. Specimens are simple, coniform and classified
into a, b and e elements. All the elements have a proclined
to erect cusp and an edged posterior margin. a elements
are subsymmetrical and bear a rounded anterior margin
with antero-lateral costae. Subsymmetrical to asymmetrical
b elements are laterally compressed. The anterior margin is
narrowly rounded and bounded by antero-lateral grooves,
developed on the both or inner side of the unit. e elements
are symmetrical and laterally compressed strongly.
Description. Elements are a long, slender unit with a proclined to
erect cusp and a posteriorly expanded base. Subsymmetrical a
elements have a cusp that inclines inwardly. The anterior margin
is broadly convex and rounded costae are carried on antero-lat-
eral faces. The posterior margin is sharply keeled from the tip of
the cusp to the basal margin. The basal cavity points toward the
anterior face and its height is 1–1.5 times of the antero-posterior
width of the basal margin. Outline of the basal margin is
expanded laterally and tapers to the posterior end.
b elements are a subsymmetrical to asymmetrical unit and lat-
erally compressed. The cusp bends inwardly. The anterior mar-
gin is narrowly rounded and bounded by antero-lateral grooves
on the both or inner side of the unit. The basal cavity opens
narrowly and its height is one-third to one half of the antero-
posterior width of the basal margin. The postero-basal corner
twists outwardly.
Laterally compressed e elements are a symmetrical acostate
unit. The anterior margin is edged or narrowly rounded. The
1450 P A L A E O N T O L O G Y , V O L U M E 5 1

form of the basal cavity resembles that of the b element, but the
basal opening is straight in aboral view.
Remarks. This species does not bear a symmetrical sub-
erectiform c element. The a element of U. tarutaoensis is
distinguished from that of U. utahensis and Utahconus
longipinnatus Ji and Barnes (1994) by the long cusp and
the slender base. The b element of this species is charac-
terized by the antero-lateral grooves along the anterior
margin and the twisted postero-basal corner of the base.
The e element of U. tarutaoensis differs from that of the
other species in a lateral compression of the element and
the unexpanded base.
Occurrence. This species is included in the lower part of the El
Paso Group in Texas (Repetski 1982) and the U. tarutaoensis
zone of the Thung Song Formation on Tarutao Island.
Genus VARIABILOCONUS Landing, Barnes and Stevens, 1986
Type species. Paltodus bassleri Furnish, 1938.
Variabiloconus bassleri (Furnish, 1938)
Plate 1, figures 11–12, 15–16
* 1938 Paltodus bassleri Furnish, p. 331, pl. 42, fig. 1.
1986 Variabiloconus bassleri (Furnish); Landing et al.,
p. 1946, 1947, pl. 3, figs. 1–7, 9.
1994 Variabiloconus bassleri (Furnish); Ji and Barnes,
p. 67, 68, pl. 25, figs. 14–26, text-fig. 38D.
1994 Variabiloconus bassleri (Furnish); Seo et al.,
figs.10.25–10.26.
1996 Variabiloconus bassleri (Furnish); Ji and Barnes,
figs. 14.19–14.25.
1996 Variabiloconus bassleri (Furnish); Landing et al.,
figs. 7.20–7.23, 7.25–7.33, 9.28–9.33.
2003 Variabiloconus bassleri (Furnish); Landing et al.,
fig. 8.2–8.5.
Material. One hundred and twenty-two specimens; 75 a, 38 b, 6
c and 3 e elements (IGUT-ag1935, 1936, 3218–3253, 3358–3447).
Remarks. Ji and Barnes (1994) re-defined this species as a
conodont bearing a, b, c and e elements. Specimens in
the Tarutao collection are classified into four elements, a,
b, c and e. The characteristics of these specimens accord
with the description of Ji and Barnes (1994).
Occurrence. V. bassleri is one of the representative species of the
Early Ordovician in North America, Australia and North China
(Wang 1984; Landing et al. 1986). The upper limit of its strati-
graphic range is the Rossodus manitouensis zone and the Low
diversity interval of the standard zonation in North American
Midcontinent (Sweet and Tolbert 1997). This species occurs
from the Rossodus muskwaensis to Acanthodus uncinatus zones of
the Thung Song Formation, in this study.
Acknowledgements. We are indebted to the referees and the
editorial board, and to J. Repetski, for reading the manuscript
and offering many useful comments and suggestions. We
would like to thank the Department of Mineral Resources of
Thailand for providing facilities for our research in Tarutao
Island. The fieldwork was funded by a grant from the Mom-
busho International Scinetific Research (Number 11440148 to
K. Sashida).
REFERENCES
A G E M A TS U, S., S A S H I D A , K., S A L Y A PO N G S E , S. and
S A R DS UD , A. 2006a. Ordovician conodonts from the
Thong Pha Phum area, western Thailand. Journal of Asian
Earth Sciences, 26, 49–60.
————————2006b. Ordovician-Silurian boundary grapto-
lites of the Satun area, southern peninsular Thailand. Paleon-
tological Research, 10, 207–214.
—— —— —— —— 2006c. Lower and Middle Ordovician con-
odonts from the Thung Song and Thung Wa areas, southern
peninsular Thailand. Paleontological Research, 10, 215–231.
—— —— —— —— 2007. Ordovician conodonts from the Satun
area, southern peninsular Thailand. Journal of Paleontology,
81, 19–37.
A L B A N E S I , G. L., HU N I C K E N , M. A. and B A R N E S ,
C. R. 1998. Biostratigraphia de conodontes de las secuencias
Ordovicicas del cerro potrerillo, Precordillera central de San
Juan, R. Argentina. 7–72. In H U N I CK E N , M. A. (ed.).
Biostratigrafia, biofacies y taxonomia de conodontes de las secu-
encias Ordovicicas del derro potrerillo, Precordillera central
de San Juan, R. Argentina. Actas de la Academia Nacional de
Ciencias, Cordoba, 12, 253 pp. [In Spanish].
B A R N E S , C. R., K E N N E D Y , D. J., M c C RA CK E N , A. D.,
N OW L A N , G. S. and T A R R A N T , G. A. 1979. The struc-
ture and evolution of Ordovician conodont apparatuses.
Lethaia, 12, 125–151.
B O UC OT , A. J., CO C KS , L. R. M. and R A CH E B OE U F ,
P. R. 1999. Early Devonian brachiopods from Satun Province,
southern Thailand. Journal of Paleontology, 73, 850–859.
B UN OP A S , S. 1981. Paleogeographic history of western Thai-
land and adjacent part of southeast Asia: a plate tectonics
interpretation. Geological Survey Paper, Special Issue, 5, 1–
810.
—— 1992. Regional stratigraphic correlation in Thailand. 189–
208. In P I A N C H A R OE N , C. (ed.). Proceedings of a National
Conference on Geologic Resources of Thailand. Department of
Mineral Resources, Bangkok.
C H E N X I A O Y U A N , P E N G M A O H U A and J I N
CA I S H E N G 1995. Lower Ordovician conodonts from
Tudi’ao, Yanhe County, Guizhou. Acta Microlalaeontologica
Sinica, 12(3), 323–332. [In Chinese].
C OC K S , L. R. M., F OR T E Y , R. A. and L E E , C. P. 2005. A
review of Lower and Middle Palaeozoic biostratigraphy in west
peninsular Malaysia and southern Thailand in its context
A G E M A T S U E T A L . : E A R L Y O R D O V I C I A N C O N O D O N T S F R O M T A R U T A O I S L A N D , T H A I L A N D 1451

within the Sibumasu Terrane. Journal of Asian Earth Sciences,
24, 703–717.
C OO P E R , B. J. 1981. Early Ordovician conodonts from the
Horn Valley Siltstone, central Australia. Palaeontology, 24, 147–
183.
E T H I N G T O N , R. L. and C L A R K, D. L. 1964. Conodonts
from the El Paso Formation (Ordovician) of Texas and Arisona.
Journal of Paleontology, 38, 685–704.
—— —— 1981. Lower and Middle Ordovician conodonts from the
Ibex area Western Millard County, Utah. A publication of the
Department of Geology Brigham Young University, Provo,
Utah, 155 pp.
F O R TE Y , R. A. and C OC K S , L. R. M. 1998. Biogeography
and palaeogeography of the Sibumasu terrane in the Ordovi-
cian: a new review. 45–56. In H A L L , R. and H O L L O W A Y ,
J. D. (eds). Biogeography and geological evolution of SE Asia.
Backhuys, Lieden, 417 pp.
F UR N I S H , W. M. 1938. Conodonts from the Prairie du Chien
(Lower Ordovician) beds of the Upper Mississippi Valley.
Journal of Paleontology, 12, 318–340.
G R A V E S , R. W. and E L L I S O N , S. 1941. Ordovician conodonts
of the Marathon basin, Texas. University of Missouri School of
Mining and Metallurgy, Bulletin, Technical Series, 14, 1–26.
H A H N , L. and S I E B E N HU N E R , M. 1982. Explanatory notes
(Paleontology) on the geological maps of northern and western
Thailand 1:250,000. Bundesanstalt fur Geowissenschaften und
Rohstoffe, Hannover, 76 pp.
H A M A D A , T., I G O, H., KO B A Y A S H I , T. and KO I K E , T.
1975. Older and Middle Palaeozoic formations and fossils of
Thailand and Malaysia. Japanese Journal of Geology and Geo-
graphy, 45, 1–39.
I GO , H. and K OI KE , T. 1967. Ordovician and Silurian con-
odonts from the Langkawi Islands, Malaysia, Part I. Geology
and Palaeontology of Southeast Asia, 3, 1–35.
—— —— 1968. Ordovician and Silurian conodonts from the
Langkawi Islands, Malaya, Part II. Geology and Palaeontology
of Southeast Asia, 5, 1–21.
J I Z A I L I A N G and B A R N E S , C. R. 1994. Lower Ordovician
conodonts of the St. George Group, Port au Port Peninsula,
western Newfoundland, Canada. Palaeontographica Canadiana,
11, 1–149.
—— —— 1996. Uppermost Cambrian and Lower Ordovician
conodont biostratigraphy of the Survey Peak Formation
(Ibexian ⁄ Tremadoc) Wilcox Pass, Alberta, Canada. Journal of
Paleontology, 70(5), 871–890.
J OH N S T O N , D. I. and B A R N E S , C. R. 2000. Early and
Middle Ordovician (Arenig) conodonts form St. Pauls Inlet and
Martin Point, Cow Head Group, Western Newfoundland,
Canada 2. Systematic paleontology. Geologica et Palaeontologica,
34, 11–87.
K OB A Y A S HI , T. 1957. Upper Cambrian fossils from peninsu-
lar Thailand. Journal of the Faculty of Science University of
Tokyo, 10, 367–382.
—— 1964. Palaeontology of Thailand (1916–62). Geology and
Palaeontology of Southeast Asia, 1, 17–30.
—— and H A M A D A , T. 1964a. On a new Malayan species of
Dalmanitina. Geology and Palaeontology of Southeast Asia, 1,
221–236.
—— —— 1964b. On the Middle Ordovician fossils from Satun,
the Malaysian frontier of Thailand. Geology and Palaeontology
of Southeast Asia, 1, 209–220.
L A N D I N G , E. and BA R N E S , C. R. 1981. Conodonts from
the Cape Clay Formation (Lower Ordovician), southern
Devon Island, Arctic Archipelage. Canadian Journal of Earth
Sciences, 18, 1609–1628.
—— —— and S T E V E N S , R. K. 1986. Tempo of earliest Ordo-
vician graptolite faunal succession: conodont-based correla-
tions from the Tremadocian of Quebec. Canadian Journal of
Earth Sciences, 23, 1928–1949.
—— W E S T R OP , S. R. and KN O X , L. A. 1996. Conodonts,
stratigraphy and relative sea-level changes of the Tribes Hill
Formations (Lower Ordovician, east-central New York).
Journal of Paleontology, 70, 656–680.
—— —— and H E R N I C K , L. A. 2003. Uppermost Cambrian-
Lower Ordovician faunas and Laurentian platform sequence
stratigraphy, eastern New York and Vermont. Journal of Paleo-
ntology, 77, 78–98.
L E E , H. 1970. Conodonten aus der Chosen-Gruppe (Unteres
Ordovizium) von Korea. Neues Jahrbuch fur Geologie und
Palaontologie, Abhandelungen, 136, 303–344. [In German].
L E H N E R T, O., K E L L E R , M. and B O R DO N A R O, O.
1998. Early Ordovician conodonts from the Southern Cuyania
Terrane (Mendoza Province, Argentina). Palaeontologia
Polonica, 58, 47–65.
L I N D S T R O M , M. 1955. Conodonts from the lowermost
Ordovician strata of south-central Sweden. Geologiska Forenin-
gens i Stockholm Forhandlingar, 76, 517–604.
—— 1971. Lower Ordovician conodonts of Europe. Geological
Society of America Memoir, 127, 21–61.
L O FG R E N , A. 1997. Reinterpretaion of the Lower Ordovician
conodont apparatus Paroistodus. Palaeontology, 40, 913–929.
—— R E P E T S K I , J. E. and E T H I N G T O N , R. L. 1999. Some
tran-Iapetus conodont faunal connections in the Tremadocian.
Bollettino della Societa Paleontologica Italiana, 37, 159–173.
M E TC A L F E , I. 1980. Ordovician conodonts from the Kaki
Bukit area, West Malaysia. Warta Geologi, 6, 63–68.
—— 1999. Gondwana dispersion and Asian accretion: an over-
view. 9–28. In M E T C A L FE , I. (ed.). Gondwana dispersion
and Asian accretion. A. A. Balkema, Rotterdam.
M I L L E R , J. F. 1969. Conodont fauna of the Notch Peak Lime-
stone (Cambro-Ordovician), House Range, Utah. Journal of
Paleontology, 43, 413–439.
—— 1980. Taxonomic revisions of some Upper Cambrian and
Lower Ordovician conodonts with comments on their evolu-
tion. The University Kansas Paleontological Contributions Paper,
99, 1–43.
M U L L E R , K. J. 1964. Conodonten aus dem unteren Ordovizi-
um von Sudkorea. Neues Jahrbuch fur Geologie und Palaontolo-
gie, Abhandlungen, 119, 93–102.
N I CO L L , R. S. 1994. Seximembrate apparatus structure of the
Late Cambrian coniform conodont Teridontus nakamurai
from the Chatsworth Limestone, Georgina Basin, Queensland.
AGSO Journal of Australian Geology and Geophysics, 15(3),
367–379.
N O G A M I , Y. 1967. Kambrische Conodonten von China, Teli 2.
Conodonten aus den hoch oberkambrischen Yencho-Schich-
1452 P A L A E O N T O L O G Y , V O L U M E 5 1

ten. Memoirs of the College of Science, University of Kyoto,
Geology and Mineralogy, Series B, 33, 211–218.
N O W L A N , G. S. 1985. Late Cambrian and Early Ordovician
conodonts from the Franklinian Miodeosyncline, Canadian
Atcric Islands. Journal of Paleontology, 59, 96–122.
P A N D E R , C. H. 1856. Monographie der fossilen Fische der silu-
rischen Systems der russischenbaltischen Gouvernements. Akade-
mie der Wissenschaften, St. Petersburg, 91 pp.
P Y L E , L. J. and BA RN E S , C. R. 2002. Taxonomy, evolution,
and biostratigraphy of conodonts from the Kechika formation,
Skoki formation, and Road River Group (Upper Cambrian to
Lower Silurian), Northeastern British Columbia. National
Research Council of Canada, Ottawa, 169 pp., 29 pls.
—— —— 2003. Conodonts from a platform-to-basin transect,
Lower Ordovician to Lower Silurian, northeastern British
Columbia, Canada. Journal of Paleontology, 77, 146–171.
—— —— and J I Z A I L I A N G 2003. Conodont fauna and
biostratigraphy of the Outram, Skoki, and Owen Creek
Formations (Lower to Middle Ordovician), Wilcox Pass,
Alberta, Canada. Journal of Paleontology, 77, 958–976.
R A O , R. I. and TO R T E L L O, M. F. 1998. Tremadoc con-
odonts and trilobites from the Cardonal Formation, Incamayo
Creek, Salta Province, northwestern Argentina. 31–45. In
S Z A N I A W S KI , H. (ed.). Proceedings of the Sixth European
conodont symposium (ECOS VI). Palaeontologia Polonica, 58,
Warszawa, 287.
R E P E T S K I , J. E. 1982. Conodonts from El Paso Group (Lower
Ordovician) of western Texas and southern New Mexico. New
Mexico Bureau of Mines and Mineral Resources Memoir, 40,
1–119.
—— and E T HI N G T ON , R. L. 1983. Rossodus manitouensis
(Conodonta), a new Early Ordovician index fossil. Journal of
Paleontology, 57, 289–301.
R O S S , R. J. Jr., H I N T Z E , L. F., E T H I N G T ON , R. L.,
M I L L E R , J. F., T A Y L O R, M. E. and R E P E T S KI , J. E. 1997.
The Ibexian, lowermost series in the North American Ordovi-
cian. U.S. Geological Survey Professional Paper, 1579 (A), 51–84.
S E O , K., L E E , H. and E T H I N G T ON , R. L. 1994. Early
Ordovician conodonts from the Dumugol formation in the
Baegunsan Syncline, eastern Yeongweol and Samcheog areas,
Kangweon-Do, Korea. Journal of Paleontology, 68, 599–
616.
S E R G E E V A , S. P. 1963. Conodonts from the Lower Ordovi-
cian of the Leningrad region. Paleontologicheskii Zhurnal, 2,
93–108.
S H E R G O L D, J. H., B UR R E T T , C. F., A KE R M A N , T. and
S T A I T , B. 1988. Late Cambrian trilobites from Tarutao
Island, Thailand. Memoir of New Mexico Bureau of Mine and
Mineral Resources, 44, 303–320.
S T A I T, B. A. and B UR R E T T , C. F. 1984. Ordovician nauti-
loid faunas of central and southern Thailand. Geological Maga-
zine, 121 (2), 115–124.
—— —— and W O N GW A N I C H , T. 1984. Ordovician trilo-
bites from the Tarutao Formation, southern Thailand. Geologi-
cal Magazine, 121 (2), 115–124.
S T O UG E , S. S. 1984. Conodonts of the Middle Ordovician
table head formation, western Newfoundland. Fossils and
Strata, 16, 1–145.
S W E E T , W. C. and TO L B E R T , C. M. 1997. An Ibexian
(Lower Ordovician) reference section in the Southern Egan
Range, Nevada, for a conodont-based chronostratigraphy. U.S.
Geological Survey Professional Paper, 1579 (B), 51–84.
T E R A O KA , Y., S A W A D A , H., Y OS H I D A , T. and P UN -
G R A S S A M I , T. 1982. Lower Paleozoic formations of Taru-
tao Islands, southern Thailand. Prince of Songkhla University
Geological Research Project Publication, 6, 1–54.
W A N G CH E N G Y U A N . 1993. Conodonts of Lower Yangtze
Valley. An indexes to biostratigraphy and organic metamor-
phic maturity. Science Press, Beijing, 326 pp. [In Chinese].
W A N G Z H I H A O 1984. Late Cambrian and Early Ordovician
conodonts from North and Northeast China with comments
on the Cambrian-Ordovician boundary. 195–258. In C H E N ,
J. and W A N G Z H I H A O . (eds). Stratigraphy and palaeontol-
ogy of systemic boundaries in China. Cambrian-Ordovician
Boundary (2). Nanjing Institute of Geology and Palaeontology,
Anhui Science and Technology Publishing House, Nanjing,
326 pp., 60 pls.
W E B B Y , B. D., C O OP E R , R. A., B E R GS TR O M , S. M. and
PA R I S , F. 2004. Stratigraphic framework and time slices. 41–
47. In W E BB Y , B. D., P A R I S , F., DR O S E R , M. L. and
PE R C I V A L , I. G. (eds). The Great Ordovician Biodiversifica-
tion Event. Columbia University Press, New York, 484 pp.
W O N G W A N I CH , T., B UR R E T T , C., T A N S A T H E I N , W.
and C HA O D UM R O N G , P. 1990. Lower to Mid Palaeozoic
stratigraphy of mainland Satun province, southern Thailand.
Journal of Southeast Asian Earth Science, 4, 1–9.
W U G U I C H U N , Y A O J I A N X I N , J I Z H A N S H E N G and
L I U S H U CA I 2005. Latest biostratigraphic advances of
Cambrian-Ordovician boundary in Qingzhou area, Shandong.
Acta Palaeontologica Sinica, 44 (1), 106–116 [In Chinese].
A G E M A T S U E T A L . : E A R L Y O R D O V I C I A N C O N O D O N T S F R O M T A R U T A O I S L A N D , T H A I L A N D 1453