donor t-cell alloreactivity against host thymic epithelium limits t-cell development after bone...
TRANSCRIPT

doi:10.1182/blood-2006-07-034157Prepublished online January 9, 2007;2007 109: 4080-4088
Annick Peter, Georg A. Holländer and Werner KrengerMathias M. Hauri-Hohl, Marcel P. Keller, Jason Gill, Katrin Hafen, Esther Pachlatko, Thomas Boulay, development after bone marrow transplantationDonor T-cell alloreactivity against host thymic epithelium limits T-cell
http://bloodjournal.hematologylibrary.org/content/109/9/4080.full.htmlUpdated information and services can be found at:
(1882 articles)Transplantation � (5022 articles)Immunobiology �
Articles on similar topics can be found in the following Blood collections
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#repub_requestsInformation about reproducing this article in parts or in its entirety may be found online at:
http://bloodjournal.hematologylibrary.org/site/misc/rights.xhtml#reprintsInformation about ordering reprints may be found online at:
http://bloodjournal.hematologylibrary.org/site/subscriptions/index.xhtmlInformation about subscriptions and ASH membership may be found online at:
Copyright 2011 by The American Society of Hematology; all rights reserved.Washington DC 20036.by the American Society of Hematology, 2021 L St, NW, Suite 900, Blood (print ISSN 0006-4971, online ISSN 1528-0020), is published weekly
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom

TRANSPLANTATION
Donor T-cell alloreactivity against host thymic epithelium limits T-celldevelopment after bone marrow transplantationMathias M. Hauri-Hohl,1 Marcel P. Keller,1 Jason Gill,1 Katrin Hafen,1 Esther Pachlatko,1 Thomas Boulay,1
Annick Peter,1 Georg A. Hollander,1 and Werner Krenger
1Department of Clinical-Biological Sciences, Laboratory of Pediatric Immunology, University of Basel,Basel University Children’s Hospital (UKBB), Switzerland
Acute graft-versus-host disease (aGVHD)impairs thymus-dependent T-cell regen-eration in recipients of allogeneic bonemarrow transplants through yet to bedefined mechanisms. Here, we demon-strate in mice that MHC-mismatched do-nor T cells home into the thymus ofunconditioned recipients. There, acti-vated donor T cells secrete IFN-�, which
in turn stimulates the programmed celldeath of thymic epithelial cells (TECs).Because TECs themselves are competentand sufficient to prime naive allospecificT cells and to elicit their effector function,the elimination of host-type professionalantigen-presenting cells (APCs) does notprevent donor T-cell activation and TECapoptosis, thus precluding normal thymo-
poiesis in transplant recipients. Hence,strategies that protect TECs may be nec-essary to improve immune reconstitutionfollowing allogeneic bone marrow trans-plantation. (Blood. 2007;109:4080-4088)
© 2007 by The American Society of Hematology
Introduction
Allogeneic bone marrow transplantation (alloBMT) is the preferredtherapeutic option for a number of life-threatening disorders, includingthose of the lymphohematopoietic system.1,2 A favorable outcome ofalloBMT depends, in turn, on the comprehensive reconstitution of thehost’s immune system. Whereas recovery of peripheral T cells occurs intransplant recipients via thymus-dependent and -independent mecha-nisms,3,4 the regeneration of phenotypically naive T cells with a broadTCR repertoire relies on the de novo generation of T cells in thethymus.3,5-11 However, alloBMT is associated with toxicities, includinggraft-versus-host disease (GVHD), which limit thymus-dependent T-cell generation. Acute GVHD (aGVHD) is induced by donor T cellsinteracting with specific cells of the recipient.12,13 In murine models andin human BMT, professional (p) antigen-presenting cells (pAPCs) of thehost are believed to be critical in initiating acute graft-versus-hostresponses.14-19 Activated donor T cells trigger an inflammatory cascadethat includes multiple cellular and cytokine effectors that are responsiblefor cell injury in a restricted set of target tissues.12,20,21 In this respect,aGVHD also affects thymic structure and function.22-27 The resultantdeficiency in the peripheral T-cell pool contributes to a patient’s risk forlife-threatening infections.10,27-29
Thymic T-cell development relies on signals provided by afunctionally competent 3-dimensional network of stromal cells.30
This compartment consists of different cell types that collectivelyenable the attraction, survival, expansion, migration, and differen-tiation of T-cell precursors. The thymic epithelial cells (TECs)constitute the most abundant cell type of the stroma and can bedifferentiated into phenotypically and functionally separate subpopu-lations.30-34 The absence of a regularly structured and composedthymic microenvironment precludes regular thymocyte develop-ment.35 We have previously established in a radiation-independenttransplantation model that aGVHD perturbs the architectural
organization and composition of the thymic microenvironment.25
These alterations were characterized by a decrease and occasionalloss of distinct TEC subpopulations. However, the cellular andmolecular mechanisms by which aGVHD causes these deficitshave remained unknown. Loss of integrity of the thymic microenvi-ronment following alloBMT was firmly associated with an inva-sion of donor T cells into the recipient’s thymus, a reduction inthymopoietic capacity and thymic output, and diminished periph-eral T-cell reconstitution.22,23,25,26
Here we report that TECs undergo programmed cell deathfollowing transfer of T cells into unconditioned, MHC-disparatemurine recipients. In vitro data indicated a direct and causative linkbetween donor T-cell activation to host alloantigens, IFN-� secre-tion, signal transducer and activator of transcription 1 (STAT-1)activation in TECs, and caspase-dependent TEC apoptosis. Impor-tantly, donor T-cell activation in vivo by host pAPCs was notrequired for induction of thymic epithelial injury because TECsthemselves displayed a cell autonomous capacity to prime naiveallogeneic T cells and to elicit IFN-� secretion. From these data weinfer that the depletion of host-derived hematopoietic pAPCs aspart of pretransplant conditioning in clinical BMT will not preventthe activation of alloreactive donor T cells. Hence, strategiesdirected at a protection of TECs from transplantation-relatedinjuries may be necessary to improve T-cell reconstitution.
Materials and methods
Mice
Female C57BL/6 (B6, H-2b, CD45.2�), Balb/c (H-2d), (C57BL/6 xDBA/2)F1 (BDF1, H-2bd, CD45.2�), B6.SJL-PtprcaPep3b/BoyJ (B6.CD45.1;
Submitted July 7, 2006; accepted December 22, 2006. Prepublished online asBlood First Edition Paper, January 9, 2007; DOI 10.1182/blood-2006-07-034157.
The online version of this article contains a data supplement.
An Inside Blood analysis of this article appears at the front of this issue.
The publication costs of this article were defrayed in part by page chargepayment. Therefore, and solely to indicate this fact, this article is herebymarked ‘‘advertisement’’ in accordance with 18 USC section 1734.
© 2007 by The American Society of Hematology
4080 BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom

H-2b, CD45.1�), B6.Cg/Ntac-Foxn1nuN9 (B6.nu/nu), CanN.Cg-Foxn1nu/Crl (Balb/c.nu/nu), 129S6/SvEvTac-Stattm1Rds (129.STAT1�/�; H-2b), and129S6/SvEvTac (129Sv; H-2b) were obtained from Biological ResearchLaboratories (Fullinsdorf, Switzerland), Jackson Laboratories (Bar Harbor,ME), Charles River (Lyon, France), and Taconic Europe (Ry, Denmark),respectively. IFN-��/� mice (GKO) that had been bred on a Balb/cbackground were kindly provided by Dr U. Eriksson (University Hospital,Basel, Switzerland). Animals between 6 and 10 weeks of age were keptunder specific pathogen-free conditions and in accordance with federalregulations.
Flow cytometry
For flow cytometry, monoclonal antibodies (mAbs) against CD3 (cloneKT3), CD4 (RM4-5), CD8 (53-6.7), TCR� (H57-592), CD44 (IM7), CD25(PC61), CD45.1 (Ly5.1; A20), B220 (RA3-6B2), CD11b (M1/70), CD11c(HL3), CD62L (MEL-14), CD69 (H1.2F3), H-2Kb (AF6-88.5), H-2Kd
(SF1-1.1), CD45 (30-F11), and I-Ab (AF6-120.1) were obtained from BDBiosciences PharMingen (San Diego, CA) or eBioscience (San Diego, CA).The anti–IFN-� mAb clone XMG1.2 (eBioscience) was used to detectintracellular IFN-� production by T cells, some of which had been culturedfor 4 hours with phorbol myristate acetate (PMA; 5 ng/mL), ionomycin(500 ng/mL), and brefeldin A (500 ng/mL). To detect apoptosis, cells werelabeled with annexin V-allophycocyanin (BD Biosciences, Allschwil,Switzerland) and propidium iodide (PI). Labeled cells were analyzed on aFACScalibur dual laser (Becton Dickinson, Mountain View, CA). Foranalysis of procaspase-12, cells were fixed, permeabilized, and incubatedwith polyclonal antibody ab8118 (Abcam, Cambridge, United Kingdom).
TEC lines
The cortical thymic epithelial cell lines TEC1-2 and TEC1C9 and themedullary lines TEC2-3 and TEC3-10 (Figure S1; available on the Bloodwebsite; see the Supplemental Materials link at the top of the online article)were obtained from B6 mice (kindly provided by Dr M. Kasai, Tokyo,Japan).36,37 Cells were plated at 105 cells/well and recombinant mouse(rm)IFN-� (R&D Systems, Abingdon, United Kingdom) was added. Thepan-caspase inhibitor quinolyl-valyl-O-methylaspartyl-[5,6-difluorophe-noxy]-methyl ketone (Q-VD-OPh; R&D Systems) was included in somecultures (5 �M). Floating and adherent cells were collected for analysisafter 72 hours of culture. For mixed epithelial-lymphocyte reactions(MELRs), 2 � 106 T cells from naive or primed B6 (H-2b) and Balb/c(H-2d) mice, respectively, were added to untreated TEC1-2 (H-2b) platedthe day before. Priming of responder cells prior to MELRs had beenachieved during a 6-day primary mixed lymphocyte reaction (MLR) withallogenic splenic stimulators. After 72 hours of MELRs, supernatants werecollected for IFN-� determination by sandwich enzyme-linked immunosor-bent assay (ELISA) using mAbs from PharMingen, and cells were preparedfor flow cytometric analysis. Lymphocytes were excluded based on theirexpression of CD45. The anti–IFN-� mAb clone XMG1.2 was used toblock IFN-� activity in culture.
Adult TEC cultures
Pure CD45�I-Ab int�high thymic epithelial cells (� 99.9%) were isolatedfrom adult thymic stromal cell preparations of naive B6 mice using aFACSAria cell sorter (Becton Dickinson; Figure S1). Reaggregate cultureswere prepared according to a published protocol.38 Here, intact thymicaggregates formed from cell mixtures within 12 to 18 hours. IFN-� wasadded to the final concentrations indicated. Apoptosis was measured byflow cytometry after 72 hours of culture as described (see “Flow cytom-etry”). To test the T-cell priming potential of adult TECs, purified lymphnode T cells (� 97% purity) were labeled with carboxyfluorescein diacetatesuccinimidyl ester (CFSE; Molecular Probes, Eugene, OR) at a concentra-tion of 2.5 �M. Subsequently, T cells were reaggregated with theCD45
�I-Ab int�high primary adult TECs at a 4:1 ratio and the cell mixture
was placed on Nucleopore filters. T cells were harvested after 96 hours,cultured for 4 hours in the presence of PMA, ionomycin, and brefeldin A(see “Flow cytometry”), and then analyzed by flow cytometry.
Reaggregation cultures and grafting of fetal TECs
Thymic lobes from fetal mice were cultured with 2-deoxyguanosine for6 days, as described,38 and were then depleted from CD45� cells. Theresultant preparations consisted of 80% TECs and were devoid of hemato-poietic cells as determined by flow cytometry (� 99.9% CD45�; FigureS1). Reaggregate cultures were subsequently prepared38 and after 24 hoursin culture, the solidified reaggregate was grafted under the kidney capsuleof athymic recipient mice. Two different models were studied: the graftingof BDF1 fetal TECs (E13) into B6.nu/nu mice (H-2b), and the grafting ofstrain 129 STAT-1�/� or STAT-1�/� fetal TECs (E14, H-2b) into Balb/c.nu/nu recipients (H-2d). Engrafted mice received 4 to 5 weeks later byintravenous injection in the first model 15 � 106 B6.CD45.1 T cells or8 � 106 Balb/c T cells in the second. Heterotopic thymi were analyzed afteranother 3 weeks.
GVHD induction
aGVHD was induced in nonirradiated BDF1 (H-2bd) mice, as described.22
Briefly, recipients were infused intravenously with 15 � 106 B6.CD45.1(H-2b) splenic T cells and were designated as b3bd. As controls, 15 � 106
syngeneic BDF1 splenic T-cells were infused into cohorts (bd3bd).
BM chimeras
To produce [bfbd] BM chimeras, naive BDF1 mice were depleted ofnatural killer (NK) cells using PK136 (400 �g/mouse intraperitoneally; day�1). On day 0, mice were lethally irradiated with 11 Gy (137Cs source at0.91 Gy/min) and subsequently given transplants with T cell-depleted BMcells (1 � 107) from naive B6 donors. At 3 to 4 months after transplanta-tion, host and donor-type pAPCs in [bfbd] chimeras were identified inprimary and secondary lymphoid organs and in the skin by expression ofCD11b and CD11c, and H-2Kb and H-2Kd, respectively (Figure S2). Micewith donor dendritic cell (DC) engraftment of more than 98% in lymphoidtissues were subsequently infused intravenously with 15 � 106 splenicT cells from B6.CD45.1 mice and were designated 45.1�3[bfbd]. Toeliminate residual radioresistant host DCs in the skin,19,39 some [bfbd]chimeras were exposed to UV light 4 weeks prior to infusion of T cells(Figure S2).
Immunoblot analysis
For detection of STAT-1 phosphorylation and caspase-12 cleavage, celllysates were immunoblotted using rabbit anti–phospho-STAT1 (Tyr701)polyclonal IgG (92 kDa; no. 07-307; Millipore Upstate, Lucerne,Switzerland) and ab1881 (Abcam), respectively.
Quantitative PCR and DNA microarray analysis
Caspase-12 mRNA expression was measured by quantitative real-timepolymerase chain reaction (PCR) in CD45�I-Ab int�high TECs, as describedin the Supplemental Materials. To determine gene expression profiles inTECs, biotin-labeled and fragmented cRNA from sorted CD45�
I-Ab�MTS24�TECs was hybridized to DNA microarrays (AffymetrixMouse Expression Set MOE430A) as detailed in Table S1.
Immunofluorescence confocal microscopy
For analysis of medullary and cortical populations of TECs,31-33 frozenthymic sections (6 �m) were fixed with 4% paraformaldehyde/PBS andstained with mAb to cytokeratin (K)18 (Ks18.04, Progene, Heidelberg,Germany), polyclonal antibody to K5 (Covance, Princeton, NJ), biotinyl-ated UEA1 lectin (Vector Laboratories, Burlingame, CA), and mAb againstMTS-10 (BD Biosciences). Antibodies and lectin were applied in differentcombinations as detailed in a previous report25 and also in the SupplementalMaterials. In additional experiments, orthotopic or ectopic thymi wereanalyzed using mAbs against CD31 (clone MTS12, provided by RichardBoyd, Melbourne, Australia), ERTR7 (provided by Willem van Ewijk,Utrecht, The Netherlands), and ab8118. Alexa-, Cy3-, and Cy5-conjugatedsecondary reagents were obtained from Molecular Probes (Invitrogen,Eugene, OR). The level of in situ apoptosis in mouse thymi was evaluatedby localizing internucleosomal DNA degradation on stained tissue sections,
ALLOREACTIVITY TOWARD THYMIC EPITHELIAL CELLS 4081BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom
using the In Situ Cell Death Detection Kit (Roche Diagnostics, Zug,Switzerland), which is a modification of the terminal deoxynucleotidetransferase-mediated dUTP nick end labeling (TUNEL) assay. All imageswere captured on a Zeiss LSM 510 Meta Laser Scanning ConfocalMicroscope system (Carl-Zeiss, Feldbach, Switzerland). Isotype controlswere used in all experiments. Overlays of blue (Cy5), red (Alexa555), andgreen (TUNEL or Alexa488) stainings were colored by computer-assistedmanagement of confocal microscopy data generated with Zeiss LSM 510software version 3.2.
Statistical analysis
Values are mean SD. For 2-group comparisons the nonparametricunpaired U test was used, whereas for multiple group comparisons, analysisof variance was used (StatView, SAS Institute, Cary, NC). The overallstatistical significance level was set to 5%.
Results
TECs undergo apoptosis during aGVHD
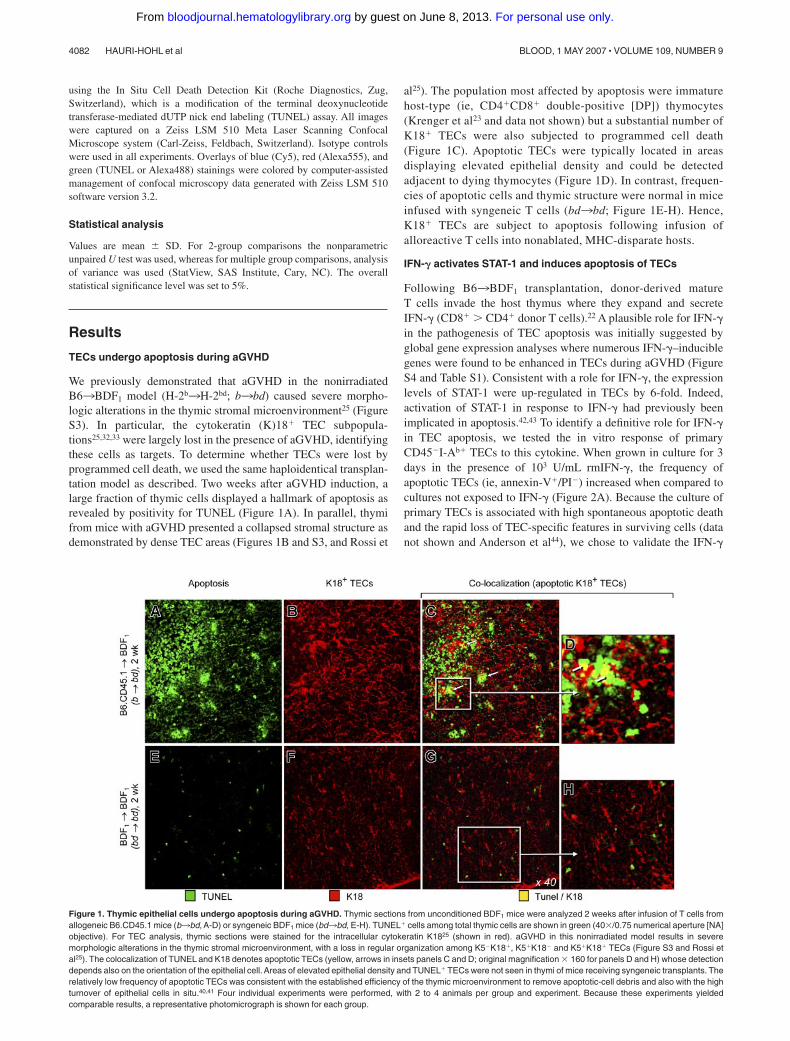
We previously demonstrated that aGVHD in the nonirradiatedB63BDF1 model (H-2b3H-2bd; b3bd) caused severe morpho-logic alterations in the thymic stromal microenvironment25 (FigureS3). In particular, the cytokeratin (K)18� TEC subpopula-tions25,32,33 were largely lost in the presence of aGVHD, identifyingthese cells as targets. To determine whether TECs were lost byprogrammed cell death, we used the same haploidentical transplan-tation model as described. Two weeks after aGVHD induction, alarge fraction of thymic cells displayed a hallmark of apoptosis asrevealed by positivity for TUNEL (Figure 1A). In parallel, thymifrom mice with aGVHD presented a collapsed stromal structure asdemonstrated by dense TEC areas (Figures 1B and S3, and Rossi et
al25). The population most affected by apoptosis were immaturehost-type (ie, CD4�CD8� double-positive [DP]) thymocytes(Krenger et al23 and data not shown) but a substantial number ofK18� TECs were also subjected to programmed cell death(Figure 1C). Apoptotic TECs were typically located in areasdisplaying elevated epithelial density and could be detectedadjacent to dying thymocytes (Figure 1D). In contrast, frequen-cies of apoptotic cells and thymic structure were normal in miceinfused with syngeneic T cells (bd3bd; Figure 1E-H). Hence,K18� TECs are subject to apoptosis following infusion ofalloreactive T cells into nonablated, MHC-disparate hosts.
IFN-� activates STAT-1 and induces apoptosis of TECs
Following B63BDF1 transplantation, donor-derived matureT cells invade the host thymus where they expand and secreteIFN-� (CD8� � CD4� donor T cells).22 A plausible role for IFN-�in the pathogenesis of TEC apoptosis was initially suggested byglobal gene expression analyses where numerous IFN-�–induciblegenes were found to be enhanced in TECs during aGVHD (FigureS4 and Table S1). Consistent with a role for IFN-�, the expressionlevels of STAT-1 were up-regulated in TECs by 6-fold. Indeed,activation of STAT-1 in response to IFN-� had previously beenimplicated in apoptosis.42,43 To identify a definitive role for IFN-�in TEC apoptosis, we tested the in vitro response of primaryCD45�I-Ab� TECs to this cytokine. When grown in culture for 3days in the presence of 103 U/mL rmIFN-�, the frequency ofapoptotic TECs (ie, annexin-V�/PI�) increased when compared tocultures not exposed to IFN-� (Figure 2A). Because the culture ofprimary TECs is associated with high spontaneous apoptotic deathand the rapid loss of TEC-specific features in surviving cells (datanot shown and Anderson et al44), we chose to validate the IFN-�
Figure 1. Thymic epithelial cells undergo apoptosis during aGVHD. Thymic sections from unconditioned BDF1 mice were analyzed 2 weeks after infusion of T cells fromallogeneic B6.CD45.1 mice (b3bd, A-D) or syngeneic BDF1 mice (bd3bd, E-H). TUNEL� cells among total thymic cells are shown in green (40�/0.75 numerical aperture [NA]objective). For TEC analysis, thymic sections were stained for the intracellular cytokeratin K1825 (shown in red). aGVHD in this nonirradiated model results in severemorphologic alterations in the thymic stromal microenvironment, with a loss in regular organization among K5�K18�, K5�K18� and K5�K18� TECs (Figure S3 and Rossi etal25). The colocalization of TUNEL and K18 denotes apoptotic TECs (yellow, arrows in insets panels C and D; original magnification � 160 for panels D and H) whose detectiondepends also on the orientation of the epithelial cell. Areas of elevated epithelial density and TUNEL� TECs were not seen in thymi of mice receiving syngeneic transplants. Therelatively low frequency of apoptotic TECs was consistent with the established efficiency of the thymic microenvironment to remove apoptotic-cell debris and also with the highturnover of epithelial cells in situ.40,41 Four individual experiments were performed, with 2 to 4 animals per group and experiment. Because these experiments yieldedcomparable results, a representative photomicrograph is shown for each group.
4082 HAURI-HOHL et al BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom
effect using different TEC lines.36,37 Exposure of cortical TEC1-2cells to rmIFN-� led to phosphorylation of STAT-1 at the Tyr701residue, demonstrating activation in response to IFN-� (Figure 2B).In parallel, IFN-� treatment reduced the number of TEC1-2 cells byhalf within 3 days (Figure 2C), whereas the frequency of apoptoticcells increased when compared to control cultures (Figure 2D).Identical results were found for cortical TEC1C9 cells and themedullary lines TEC2-3 and TEC3-10 (Figure S1). This IFN-�–induced TEC-cell death was effected via the activation of pro-caspases since the addition of the pan-caspase inhibitor Q-VD-OPhreduced apoptosis to background levels (Figure 2D). Consistently,preliminary data indicated the activation of caspase-12 in TEC1-2in response to IFN-�, and also its enhanced expression in TECsduring aGVHD, suggesting a putative role for this caspase in TECapoptosis (Figure S4).
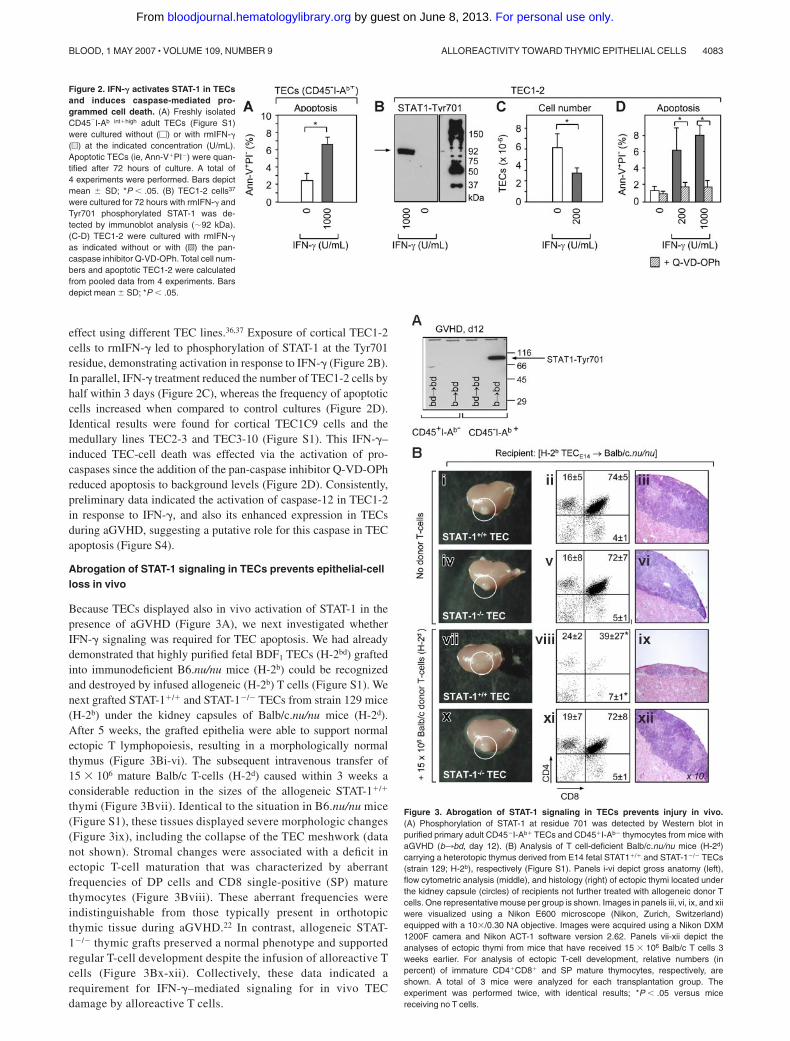
Abrogation of STAT-1 signaling in TECs prevents epithelial-cellloss in vivo
Because TECs displayed also in vivo activation of STAT-1 in thepresence of aGVHD (Figure 3A), we next investigated whetherIFN-� signaling was required for TEC apoptosis. We had alreadydemonstrated that highly purified fetal BDF1 TECs (H-2bd) graftedinto immunodeficient B6.nu/nu mice (H-2b) could be recognizedand destroyed by infused allogeneic (H-2b) T cells (Figure S1). Wenext grafted STAT-1�/� and STAT-1�/� TECs from strain 129 mice(H-2b) under the kidney capsules of Balb/c.nu/nu mice (H-2d).After 5 weeks, the grafted epithelia were able to support normalectopic T lymphopoiesis, resulting in a morphologically normalthymus (Figure 3Bi-vi). The subsequent intravenous transfer of15 � 106 mature Balb/c T-cells (H-2d) caused within 3 weeks aconsiderable reduction in the sizes of the allogeneic STAT-1�/�
thymi (Figure 3Bvii). Identical to the situation in B6.nu/nu mice(Figure S1), these tissues displayed severe morphologic changes(Figure 3ix), including the collapse of the TEC meshwork (datanot shown). Stromal changes were associated with a deficit inectopic T-cell maturation that was characterized by aberrantfrequencies of DP cells and CD8 single-positive (SP) maturethymocytes (Figure 3Bviii). These aberrant frequencies wereindistinguishable from those typically present in orthotopicthymic tissue during aGVHD.22 In contrast, allogeneic STAT-1�/� thymic grafts preserved a normal phenotype and supportedregular T-cell development despite the infusion of alloreactive Tcells (Figure 3Bx-xii). Collectively, these data indicated arequirement for IFN-�–mediated signaling for in vivo TECdamage by alloreactive T cells.
Figure 2. IFN-� activates STAT-1 in TECsand induces caspase-mediated pro-grammed cell death. (A) Freshly isolatedCD45�I-Ab int�high adult TECs (Figure S1)were cultured without (�) or with rmIFN-�(u) at the indicated concentration (U/mL).Apoptotic TECs (ie, Ann-V�PI�) were quan-tified after 72 hours of culture. A total of4 experiments were performed. Bars depictmean SD; *P � .05. (B) TEC1-2 cells37
were cultured for 72 hours with rmIFN-� andTyr701 phosphorylated STAT-1 was de-tected by immunoblot analysis (92 kDa).(C-D) TEC1-2 were cultured with rmIFN-�as indicated without or with (o) the pan-caspase inhibitor Q-VD-OPh. Total cell num-bers and apoptotic TEC1-2 were calculatedfrom pooled data from 4 experiments. Barsdepict mean SD; *P � .05.
Figure 3. Abrogation of STAT-1 signaling in TECs prevents injury in vivo.(A) Phosphorylation of STAT-1 at residue 701 was detected by Western blot inpurified primary adult CD45�I-Ab� TECs and CD45�I-Ab� thymocytes from mice withaGVHD (b3bd, day 12). (B) Analysis of T cell-deficient Balb/c.nu/nu mice (H-2d)carrying a heterotopic thymus derived from E14 fetal STAT1�/� and STAT-1�/� TECs(strain 129; H-2b), respectively (Figure S1). Panels i-vi depict gross anatomy (left),flow cytometric analysis (middle), and histology (right) of ectopic thymi located underthe kidney capsule (circles) of recipients not further treated with allogeneic donor Tcells. One representative mouse per group is shown. Images in panels iii, vi, ix, and xiiwere visualized using a Nikon E600 microscope (Nikon, Zurich, Switzerland)equipped with a 10�/0.30 NA objective. Images were acquired using a Nikon DXM1200F camera and Nikon ACT-1 software version 2.62. Panels vii-xii depict theanalyses of ectopic thymi from mice that have received 15 � 106 Balb/c T cells 3weeks earlier. For analysis of ectopic T-cell development, relative numbers (inpercent) of immature CD4�CD8� and SP mature thymocytes, respectively, areshown. A total of 3 mice were analyzed for each transplantation group. Theexperiment was performed twice, with identical results; *P � .05 versus micereceiving no T cells.
ALLOREACTIVITY TOWARD THYMIC EPITHELIAL CELLS 4083BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom
TECs elicit IFN-� secretion by primed allospecific T cellsand as a result undergo apoptosis
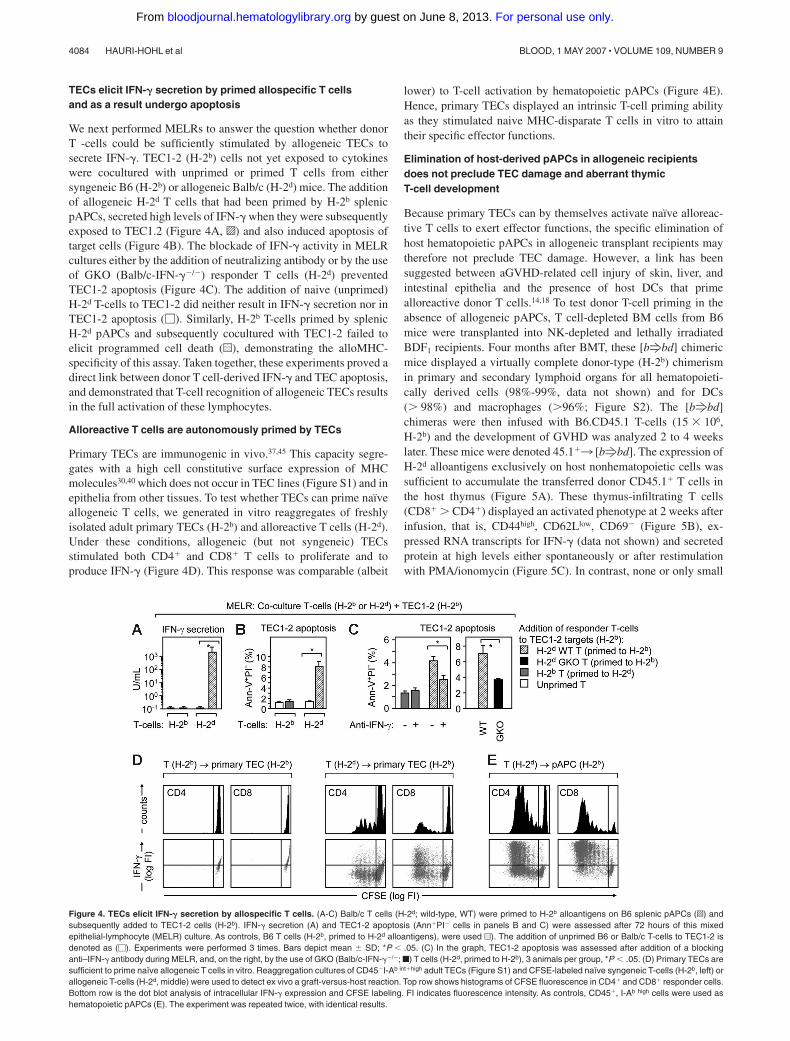
We next performed MELRs to answer the question whether donorT -cells could be sufficiently stimulated by allogeneic TECs tosecrete IFN-�. TEC1-2 (H-2b) cells not yet exposed to cytokineswere cocultured with unprimed or primed T cells from eithersyngeneic B6 (H-2b) or allogeneic Balb/c (H-2d) mice. The additionof allogeneic H-2d T cells that had been primed by H-2b splenicpAPCs, secreted high levels of IFN-� when they were subsequentlyexposed to TEC1.2 (Figure 4A, o) and also induced apoptosis oftarget cells (Figure 4B). The blockade of IFN-� activity in MELRcultures either by the addition of neutralizing antibody or by the useof GKO (Balb/c-IFN-��/�) responder T cells (H-2d) preventedTEC1-2 apoptosis (Figure 4C). The addition of naive (unprimed)H-2d T-cells to TEC1-2 did neither result in IFN-� secretion nor inTEC1-2 apoptosis (�). Similarly, H-2b T-cells primed by splenicH-2d pAPCs and subsequently cocultured with TEC1-2 failed toelicit programmed cell death (u), demonstrating the alloMHC-specificity of this assay. Taken together, these experiments proved adirect link between donor T cell-derived IFN-� and TEC apoptosis,and demonstrated that T-cell recognition of allogeneic TECs resultsin the full activation of these lymphocytes.
Alloreactive T cells are autonomously primed by TECs
Primary TECs are immunogenic in vivo.37,45 This capacity segre-gates with a high cell constitutive surface expression of MHCmolecules30,40 which does not occur in TEC lines (Figure S1) and inepithelia from other tissues. To test whether TECs can prime naıveallogeneic T cells, we generated in vitro reaggregates of freshlyisolated adult primary TECs (H-2b) and alloreactive T cells (H-2d).Under these conditions, allogeneic (but not syngeneic) TECsstimulated both CD4� and CD8� T cells to proliferate and toproduce IFN-� (Figure 4D). This response was comparable (albeit
lower) to T-cell activation by hematopoietic pAPCs (Figure 4E).Hence, primary TECs displayed an intrinsic T-cell priming abilityas they stimulated naive MHC-disparate T cells in vitro to attaintheir specific effector functions.
Elimination of host-derived pAPCs in allogeneic recipientsdoes not preclude TEC damage and aberrant thymicT-cell development
Because primary TECs can by themselves activate naıve alloreac-tive T cells to exert effector functions, the specific elimination ofhost hematopoietic pAPCs in allogeneic transplant recipients maytherefore not preclude TEC damage. However, a link has beensuggested between aGVHD-related cell injury of skin, liver, andintestinal epithelia and the presence of host DCs that primealloreactive donor T cells.14,18 To test donor T-cell priming in theabsence of allogeneic pAPCs, T cell-depleted BM cells from B6mice were transplanted into NK-depleted and lethally irradiatedBDF1 recipients. Four months after BMT, these [bfbd] chimericmice displayed a virtually complete donor-type (H-2b) chimerismin primary and secondary lymphoid organs for all hematopoieti-cally derived cells (98%-99%, data not shown) and for DCs(� 98%) and macrophages (�96%; Figure S2). The [bfbd]chimeras were then infused with B6.CD45.1 T-cells (15 � 106,H-2b) and the development of GVHD was analyzed 2 to 4 weekslater. These mice were denoted 45.1�3 [bfbd]. The expression ofH-2d alloantigens exclusively on host nonhematopoietic cells wassufficient to accumulate the transferred donor CD45.1� T cells inthe host thymus (Figure 5A). These thymus-infiltrating T cells(CD8� � CD4�) displayed an activated phenotype at 2 weeks afterinfusion, that is, CD44high, CD62Llow, CD69� (Figure 5B), ex-pressed RNA transcripts for IFN-� (data not shown) and secretedprotein at high levels either spontaneously or after restimulationwith PMA/ionomycin (Figure 5C). In contrast, none or only small
Figure 4. TECs elicit IFN-� secretion by allospecific T cells. (A-C) Balb/c T cells (H-2d; wild-type, WT) were primed to H-2b alloantigens on B6 splenic pAPCs (o) andsubsequently added to TEC1-2 cells (H-2b). IFN-� secretion (A) and TEC1-2 apoptosis (Ann�PI� cells in panels B and C) were assessed after 72 hours of this mixedepithelial-lymphocyte (MELR) culture. As controls, B6 T cells (H-2b, primed to H-2d alloantigens), were used u). The addition of unprimed B6 or Balb/c T-cells to TEC1-2 isdenoted as (�). Experiments were performed 3 times. Bars depict mean SD; *P � .05. (C) In the graph, TEC1-2 apoptosis was assessed after addition of a blockinganti–IFN-� antibody during MELR, and, on the right, by the use of GKO (Balb/c-IFN-��/�; f) T cells (H-2d, primed to H-2b), 3 animals per group, *P � .05. (D) Primary TECs aresufficient to prime naıve allogeneic T cells in vitro. Reaggregation cultures of CD45�I-Ab int�high adult TECs (Figure S1) and CFSE-labeled naıve syngeneic T-cells (H-2b, left) orallogeneic T-cells (H-2d, middle) were used to detect ex vivo a graft-versus-host reaction. Top row shows histograms of CFSE fluorescence in CD4� and CD8� responder cells.Bottom row is the dot blot analysis of intracellular IFN-� expression and CFSE labeling. FI indicates fluorescence intensity. As controls, CD45�, I-Ab high cells were used ashematopoietic pAPCs (E). The experiment was repeated twice, with identical results.
4084 HAURI-HOHL et al BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom

percentages of the host populations secreted IFN-� (Figure 5F).Analogous to nonchimeric mice with aGVHD,22 the recipient type(ie, CD45.1�) thymocytes were reduced in total cellularity (Figure5D) and the relative frequencies of immature DP and mature CD4and CD8 SP cells were abnormal (Figure 5E).
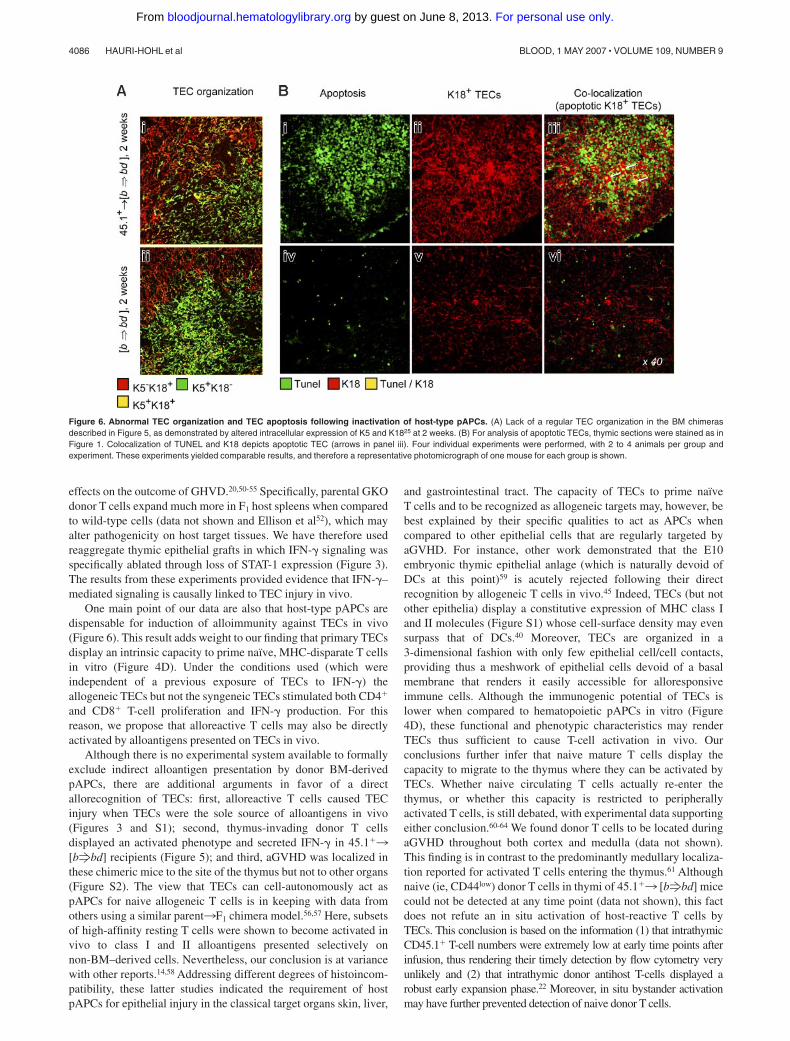
Extensive morphologic changes occurred in thymi of 45.1�3[bfbd] mice that were characterized by altered architecturalorganization of TECs (Figure 6A). Specifically, K18� TECs werefrequently grouped in dense areas (Figure 6Bii) and thus displayedan abnormal pattern indistinguishable from that of nonchimericmice with aGVHD.25 Moreover, a fraction of the K18� TECs wasalso apoptotic (Figure 6Biii). Taken together, 45.1�3 [bfbd]mice developed all the hallmarks of thymic aGVHD despite theabsence of allogeneic host pAPC. Signs of aGVHD were notdetected in the classical target organs, liver and skin, of 45.1�3[bfbd] mice (Figure S2), demonstrating that aGVHD specificallytargeted the thymus in this model. Skin disease did not developeven though residual radioresistant host-type pAPCs were stillpresent at substantial frequencies (7% of total pAPCs) inepidermal tissue from [bfbd] chimeras (Figure S2) and Merad etal.19,39 Moreover, the exposure of chimeras to UV light furtherreduced host DCs and macrophages but did not prevent thymicGVHD (Figure S2). Hence, peripheral activation of donor T cellsby residual host DCs in the skin, and subsequent migration of Tcells to the thymus, was unlikely to be a cause for TEC injury inthis model. Taken together, our results suggested that TECs maysuffice to prime alloreactive donor T cells not only in vitro but alsoin vivo and they clearly demonstrated that changes in the composi-tion, organization, and viability of TECs as a consequence ofGVHD is sufficient to cause irregular thymocyte maturation.
Discussion
The present study was designed to understand the pathways bywhich aGVHD impairs T-cell regeneration after alloBMT. Ourwork was based on previous observations that the contribution ofthe thymus to the patient’s T-cell recovery following alloBMT isinfluenced by the presence of GVHD.6,10,46 Indeed, thymic stromalintegrity is affected during both clinical and experimental dis-ease,28,47-49 with TECs as a disease-relevant GVHD target.25
Because regular development of T cells is dependent on TECs,30,35
any injury to the latter following alloBMT caused by GVHDshould have severe consequences on normal T-cell maturation,repertoire selection, and export. This prediction led us to investi-gate the cellular and molecular mechanism by which aGVHDcauses TEC injury. The use of a nonablative parent3F1 transplan-tation model for these studies was guided by the necessity todistinguish pathogenic influences on TECs by graft-versus-hostreactions from those caused by chemo/radiotherapy.
Here we describe that TECs undergo apoptosis on recognitionby MHC-disparate donor T cells and we identify molecularmechanisms by which this occurs. Using an in vitro system, a directand causative link was established between donor T-cell activationto TEC alloantigens, IFN-� secretion, STAT-1 activation in TECs,and caspase-mediated TEC apoptosis (Figures 2 and 4). Directlytesting the role of IFN-� and its association to thymic GVHD in anin vivo situation was, however, not straightforward since asystemic abrogation of IFN-� signaling in transplant recipients orthe use of GKO donor T cells exert variable and unpredictable
Figure 5. Thymic donor T-cell invasion and abnormal T-cell development following inactivation of host-type pAPC. [B63BDF1] BM chimeras (Figures S2-S3)remained untreated [bfbd], or received B6.CD45.1 T-cells, denoted 45.1�3 [bfbd]. Analysis of intrathymic CD45.1� donor T cells (A-C) and CD45.1� host-type thymocytes(D-F). (A) Frequencies and total numbers of donor CD4� (u) and CD8� (o) T cells at 2 to 4 weeks after T-cell infusion. For comparison, donor T-cell infiltration during aGVHD innonchimeric mice is given (F). (B) Activation markers of thymic CD45.1� T cells (shaded) were compared to splenic T cells from naive B6 mice (not filled). The x-axes show logFI of flow cytometric analyses. Numbers depict relative frequencies (%) of cells expressing the indicated surface marker. (C) Intracytoplasmic (i.c.) IFN-� expression bydonor-derived intrathymic T cells was detected in 8 mice per group. (D) Absolute number of host-type thymocytes in 45.1�3 [bfbd] (u) was compared to [bfbd] mice (�).(E) Frequencies of CD45.1� thymocyte populations. (F) Intracytoplasmic IFN-� in CD45.1� thymocytes in the same mice shown in panel C. For panels A, D, and E, a total of 16(controls) and 26 (aGVHD) mice were analyzed. Data were compiled from results of 6 experiments, with 2 to 4 animals per group. For panels B, C, and F, 8 mice (GVHD) and 4mice (controls), respectively, were studied per group. *P � .05 versus controls.
ALLOREACTIVITY TOWARD THYMIC EPITHELIAL CELLS 4085BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom
effects on the outcome of GHVD.20,50-55 Specifically, parental GKOdonor T cells expand much more in F1 host spleens when comparedto wild-type cells (data not shown and Ellison et al52), which mayalter pathogenicity on host target tissues. We have therefore usedreaggregate thymic epithelial grafts in which IFN-� signaling wasspecifically ablated through loss of STAT-1 expression (Figure 3).The results from these experiments provided evidence that IFN-�–mediated signaling is causally linked to TEC injury in vivo.
One main point of our data are also that host-type pAPCs aredispensable for induction of alloimmunity against TECs in vivo(Figure 6). This result adds weight to our finding that primary TECsdisplay an intrinsic capacity to prime naıve, MHC-disparate T cellsin vitro (Figure 4D). Under the conditions used (which wereindependent of a previous exposure of TECs to IFN-�) theallogeneic TECs but not the syngeneic TECs stimulated both CD4�
and CD8� T-cell proliferation and IFN-� production. For thisreason, we propose that alloreactive T cells may also be directlyactivated by alloantigens presented on TECs in vivo.
Although there is no experimental system available to formallyexclude indirect alloantigen presentation by donor BM-derivedpAPCs, there are additional arguments in favor of a directallorecognition of TECs: first, alloreactive T cells caused TECinjury when TECs were the sole source of alloantigens in vivo(Figures 3 and S1); second, thymus-invading donor T cellsdisplayed an activated phenotype and secreted IFN-� in 45.1�3[bfbd] recipients (Figure 5); and third, aGVHD was localized inthese chimeric mice to the site of the thymus but not to other organs(Figure S2). The view that TECs can cell-autonomously act aspAPCs for naive allogeneic T cells is in keeping with data fromothers using a similar parent3F1 chimera model.56,57 Here, subsetsof high-affinity resting T cells were shown to become activated invivo to class I and II alloantigens presented selectively onnon-BM–derived cells. Nevertheless, our conclusion is at variancewith other reports.14,58 Addressing different degrees of histoincom-patibility, these latter studies indicated the requirement of hostpAPCs for epithelial injury in the classical target organs skin, liver,
and gastrointestinal tract. The capacity of TECs to prime naıveT cells and to be recognized as allogeneic targets may, however, bebest explained by their specific qualities to act as APCs whencompared to other epithelial cells that are regularly targeted byaGVHD. For instance, other work demonstrated that the E10embryonic thymic epithelial anlage (which is naturally devoid ofDCs at this point)59 is acutely rejected following their directrecognition by allogeneic T cells in vivo.45 Indeed, TECs (but notother epithelia) display a constitutive expression of MHC class Iand II molecules (Figure S1) whose cell-surface density may evensurpass that of DCs.40 Moreover, TECs are organized in a3-dimensional fashion with only few epithelial cell/cell contacts,providing thus a meshwork of epithelial cells devoid of a basalmembrane that renders it easily accessible for alloresponsiveimmune cells. Although the immunogenic potential of TECs islower when compared to hematopoietic pAPCs in vitro (Figure4D), these functional and phenotypic characteristics may renderTECs thus sufficient to cause T-cell activation in vivo. Ourconclusions further infer that naive mature T cells display thecapacity to migrate to the thymus where they can be activated byTECs. Whether naive circulating T cells actually re-enter thethymus, or whether this capacity is restricted to peripherallyactivated T cells, is still debated, with experimental data supportingeither conclusion.60-64 We found donor T cells to be located duringaGVHD throughout both cortex and medulla (data not shown).This finding is in contrast to the predominantly medullary localiza-tion reported for activated T cells entering the thymus.61 Althoughnaive (ie, CD44low) donor T cells in thymi of 45.1�3 [bfbd] micecould not be detected at any time point (data not shown), this factdoes not refute an in situ activation of host-reactive T cells byTECs. This conclusion is based on the information (1) that intrathymicCD45.1� T-cell numbers were extremely low at early time points afterinfusion, thus rendering their timely detection by flow cytometry veryunlikely and (2) that intrathymic donor antihost T-cells displayed arobust early expansion phase.22 Moreover, in situ bystander activationmay have further prevented detection of naive donor T cells.
Figure 6. Abnormal TEC organization and TEC apoptosis following inactivation of host-type pAPCs. (A) Lack of a regular TEC organization in the BM chimerasdescribed in Figure 5, as demonstrated by altered intracellular expression of K5 and K1825 at 2 weeks. (B) For analysis of apoptotic TECs, thymic sections were stained as inFigure 1. Colocalization of TUNEL and K18 depicts apoptotic TEC (arrows in panel iii). Four individual experiments were performed, with 2 to 4 animals per group andexperiment. These experiments yielded comparable results, and therefore a representative photomicrograph of one mouse for each group is shown.
4086 HAURI-HOHL et al BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom

Whereas the reaggregate thymic epithelial grafts demonstratedthat thymic epithelium is indeed sufficient to induce in vivo T-cellresponses leading to TEC apoptosis (Figures 3 and S1), this may ormay not be the usual course of events in aGVHD. The present viewholds that host-derived hematopoietic pAPCs are required totrigger alloimmune responses in vivo and to induce host tissueinjury during aGVHD.14,15,17-19,58 Thus, it seems likely that underregular conditions (ie, the presence of host pAPC), alloreactive Tcells are activated by pAPCs in the periphery, acquire the capacityto produce IFN-�, enter the thymus as activated T cells, secreteIFN-�, and induce TEC apoptosis. In additional experiments using“reverse” BM chimeras (ie, 45.1�3 [Ebdfbd]), we formallydemonstrated such a role for pAPCs in TEC injury (Figure S3).These chimeras expressed the H-2d alloantigen on their pAPCs butnot on TECs. On infusion of B6.CD45.1� T cells, TEC injury anddeficits in thymic T-cell development (data not shown) were clearlyevident in 45.1�3 (bdfb) mice and the extent of aGVHDpathology was comparable to that described for nonchimericmice.22,23,25 Thus, the expression of alloantigens on host thymicepithelium was not a prerequisite for the initiation of TEC injury,confirming previous data for other types of host epithelia.15
Our data infer that the depletion of host-derived hematopoieticpAPCs as part of pretransplant conditioning in clinical BMT willlikely not prevent the activation of alloreactive donor T cells andthe ensuing injury of TECs, and the generation of an aberrantperipheral T-cell compartment. Here, direct recognition of MHC-disparate TECs by donor T cells may only be preventable by donorT-cell depletion, which, in turn, has its own shortcomings.65
Furthermore, our conclusion remains to have clinical significanceshould TECs be destroyed in aGVHD via mechanisms involvingindirect presentation (ie, possibly by residual radioresistant hostpAPC or by donor BM-derived elements). Thus, our findingsprovide a clear rationale to establish therapeutic approaches thatpreserve recipient TEC organization, differentiation, and functionfollowing alloBMT. We have initiated studies addressing TECcytoprotection using fibroblast growth factor-7 (FGF7, also calledkeratinocyte growth factor [KGF]). which promotes epithelial-cellproliferation and differentiation in a variety of tissues (includingTECs, Werner66 and Min et al67 and Rossi et al68. Signaling via its
receptor FGFR2IIIb, whose expression in the murine thymus isrestricted to TECs,25,69 FGF7 preserved the structure of the thymicmicroenvironment during aGVHD in mice.25 This TEC cytoprotec-tion was achieved despite the presence of alloreactive donor T cellswithin the confines of the thymic microenvironment. Importantly,the integrity of the TEC network allowed for regular thymopoiesisto occur. Thus, future approaches to limit immune deficiencyfollowing alloBMT should include alloreactive donor T cells fortheir advantageous antileukemic effects and novel interventions toprotect TEC integrity and function.
Acknowledgments
This work was supported by the Swiss National Science Founda-tion grants 3100-61782.00 (W.K.) and 3100-68310.02 (G.A.H.), bya grant from the European Community 6th Framework ProgramEuro-Thymaide Integrated Project (G.A.H.), by National Institutesof Health grant ROI-A1057477-01 (G.A.H), a Roche ResearchFoundation grant (M.M.H.), and by the Basel Cancer League(W.K., M.P.K.).
The authors thank Drs Bruce B. Blazar, Dale I. Godfrey andJonathan Sprent for helpful discussions.
Authorship
Contribution: M.M.H. and M.P.K. contributed equally to this workand share first authorship and both designed and performed work;W.K. and G.A.H. share senior authorship, and both designed workand wrote the paper; J.G. performed microscopy and imageacquisition; K.H., E.P., T.B., and A.P. performed work.
Conflict-of-interest disclosure: The authors declare no compet-ing financial interests.
Correspondence: Werner Krenger or Georg A. Hollander, Labora-tory of Pediatric Immunology, Department of Clinical-Biological Sci-ences, Center for Biomedicine, University of Basel, Mattenstrasse 28,4058 Basel, Switzerland; e-mail: [email protected]; [email protected].
References
1. Loberiza F. Report on the state of the art in bloodand marrow transplantation. IBMTR/ABMTRNewsletter. 2003;10:7-9.
2. Gratwohl A, Baldomero H, Passweg J. Hemato-poietic stem cell transplantation for hematologicalmalignancies in Europe. Leukemia. 2003;17:941-959.
3. Hakim FT, Gress RE. Reconstitution of thymicfunction after stem cell transplantation in humans.Curr Opin Hematol. 2002;9:490-496.
4. Van den Brink MR, Alpdogan O, Boyd RL. Strate-gies to enhance T-cell reconstitution in immuno-compromised patients. Nat Rev Immunol. 2004;4:856-867.
5. Roux E, Dumont-Girard F, Starobinski M, et al.Recovery of immune reactivity after T-cell-de-pleted bone marrow transplantation depends onthymic activity. Blood. 2000;96:2299-2303.
6. Douek DC, Vescio RA, Betts MR, et al. Assess-ment of thymic output in adults after hematopoi-etic stem cell transplantation and prediction ofT-cell reconstitution. Lancet. 2000;355:1875-1881.
7. Haynes BF, Markert ML, Sempowski GD, PatelDD, Hale LP. The role of the thymus in immunereconstitution in aging, bone marrow transplanta-tion, and HIV-1 infection. Annu Rev Immunol.2000;18:529-560.
8. Heitger A, Greinix H, Mannhalter C, et al. Re-quirement of residual thymus to restore normalT-cell subsets after human allogeneic bone mar-row transplantation. Transplantation. 2000;69:2366-2373.
9. Patel DD, Gooding ME, Parrott RE, Curtis KM,Haynes BF, Buckley RH. Thymic function afterhematopoietic stem-cell transplantation for thetreatment of severe combined immunodeficiency.N Engl J Med. 2000;342:1325-1332.
10. Weinberg K, Blazar BR, Wagner JE, et al. Factorsaffecting thymic function after allogeneic hemato-poietic stem cell transplantation. Blood. 2001;97:1458-1466.
11. Muraro PA, Douek DC, Packer A, et al. Thymicoutput generates a new and diverse TCR reper-toire after autologous stem cell transplantation inmultiple sclerosis patients. J Exp Med. 2005;201:805-816.
12. Reddy P, Ferrara JL. Immunobiology of acutegraft-versus-host disease. Blood Rev. 2003;17:187-194.
13. Blazar BR, Murphy WJ. Bone marrow transplan-tation and approaches to avoid graft-versus-hostdisease (GVHD). Philos Trans R Soc Lond B BiolSci. 2005;360:1747-1767.
14. Shlomchik WD, Couzens MS, Tang CB, et al.Prevention of graft-versus-host disease by inacti-
vation of host antigen-presenting cells. Science.1999;285:412-415.
15. Teshima T, Ordemann R, Reddy P, et al. Acutegraft-versus-host disease does not require al-loantigen expression on host epithelium. NatMed. 2002;8:575-581.
16. Ruggeri L, Capanni M, Urbani E, et al. Effective-ness of donor natural killer cell alloreactivity inmismatched hematopoietic transplants. Science.2002;295:2097-2100.
17. Zhang Y, Louboutin JP, Zhu J, Rivera AJ, Emer-son SG. Preterminal host dendritic cells in irradi-ated mice prime CD8� T cell-mediated acutegraft-versus-host disease. J Clin Invest. 2002;109:1335-1344.
18. Shlomchik WD. Antigen presentation in graft-vs-host disease. Exp Hematol. 2003;31:1187-1197.
19. Merad M, Hoffmann P, Ranheim E, et al. Deple-tion of host Langerhans cells before transplanta-tion of donor alloreactive T cells prevents skingraft-versus-host disease. Nat Med. 2004;5:510-517.
20. Krenger W, Hill GR, Ferrara JLM. Cytokine cas-cades in acute graft-versus-host disease. Trans-plantation. 1997;64:553-558.
21. Ferrara JLM, Yanik G. Acute graft versus host
ALLOREACTIVITY TOWARD THYMIC EPITHELIAL CELLS 4087BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom

disease: pathophysiology, risk factors, and pre-vention strategies. Clin Adv Hematol Oncol. 2005;428:415-419.
22. Krenger W, Rossi S, Piali L, Hollander GA. Thy-mic atrophy in murine acute graft-versus-host dis-ease is effected by impaired cell cycle progres-sion of host pro- and pre-T cells. Blood. 2000;96:347-354.
23. Krenger W, Rossi S, Hollander GA. Apoptosis ofthymocytes during acute graft-versus-host dis-ease is independent of glucocorticoids. Trans-plantation. 2000;69:2190-2193.
24. Van den Brink MR, Moore E, Ferrara JL, BurakoffSJ. Graft-versus-host-disease-associated thymicdamage results in the appearance of T cell cloneswith anti-host reactivity. Transplantation. 2000;69:446-449.
25. Rossi S, Blazar BR, Farrell CL, et al. Keratinocytegrowth factor preserves normal thymopoiesis andthymic microenvironment during experimentalgraft-versus-host disease. Blood. 2002;100:682-691.
26. Krenger W, Schmidlin H, Cavadini G, HollanderGA. On the relevance of T-cell receptor rear-rangement circles (TRECs) as molecular markersfor thymic output during experimental graft-ver-sus-host disease. J Immunol. 2004;172:7359-7367.
27. Fry TJ, Mackall CL. Immune reconstitution follow-ing hematopoietic progenitor cell transplantation:challenges for the future. Bone Marrow Trans-plant. 2005;35(suppl 1):S53–57.
28. Hakim FT, Mackall CL. The immune system: ef-fector and target of graft-versus-host disease. In:Ferrara JLM, Deeg HJ, Burakoff SJ, eds. Graft-vs-Host Disease. New York, NY: Marcel Dekker;1997:257-289.
29. Peggs KS, Mackinnon S. Immune reconstitutionfollowing hematopoietic stem cell transplantation.Br J Haematol. 2004;124:407-420.
30. Gill J, Malin M, Sutherland J, Gray DHD, Hol-lander GA, Boyd RL. Thymic generation and re-generation. Immunol. Rev. 2003;195:28-50.
31. Kampinga J, Berges S, Boyd RL, et al. Thymicepithelial antibodies: immunohistological analysisand introduction of nomenclature. Thymus. 1989;13:165-173.
32. Godfrey DI, Izon DJ, Tucek CL, Wilson TJ, BoydRL. The phenotypic heterogeneity of mouse thy-mic stromal cells. Immunology. 1990;70:66-74.
33. Klug DB, Carter C, Crouch E, Roop D, Conti CJ,Richie ER. Interdependence of cortical thymicepithelial cell differentiation and T-lineage com-mitment. Proc Natl Acad Sci U S A. 1998;95:11822-11827.
34. Gill J, Malin M, Hollander GA, Boyd RL. Genera-tion of a complete thymic microenvironment byMTS24� thymic epithelial cells. Nat Immunol.2002;3:635-642.
35. Hollander GA, Wang B, Nichogiannopoulou A, etal. Developmental control point in induction ofthymic cortex regulated by a subpopulation ofprothymocytes. Nature. 1995;373:350-353.
36. Mizuochi T, Kasai M, Kokuho T, Kakiuchi T, Hiro-kawa K. Medullary but not cortical thymic epithe-lial cells present soluble antigens to helper Tcells. J Exp Med. 1992;175:1601-1605.
37. Kasai M, Hirokawa K, Kajino K, et al. Differencein antigen presentation pathways between corti-
cal and medullary thymic epithelial cells. EurJ Immunol. 1996;26:2101-2107.
38. Anderson G, Jenkinson EJ, Moore NC, Owen JJ.MHC class II-positive epithelium and mesen-chyme cells are both required for T-cell develop-ment in the thymus. Nature. 1993;362:70-73.
39. Merad M, Manz MG, Karsunky H, et al. Langer-hans cells renew in the skin throughout life understeady-state conditions. Nat Immunol. 2002;3:1135-1141.
40. Yang SJ, Ahn S, Park CS, et al. The quantitativeassessment of MHC II on thymic epithelium: im-plications in cortical thymocyte development. IntImmunol. 2006;18:729-739.
41. Gillard GO, Farr AG. Features of medullary thy-mic epithelium implicate postnatal developmentin maintaining epithelial heterogeneity and tissue-restricted antigen expression. J Immunol. 2006;176:5815-5824.
42. Stephanou A, Latchman DS. STAT-1: a novelregulator of apoptosis. Int J Exp Pathol. 2003;84:239-244.
43. Chilakamarti V, Ramana CV, Gil MP, SchreiberRD, Stark GR. STAT-1-dependent and -indepen-dent pathways in IFN-gamma-dependent signal-ing. Trends Immunol. 2002;23:96-101.
44. Anderson KL, Moore NC, McLoughlin DE, Jenkin-son EJ, Owen JJ. Studies on thymic epithelialcells in vitro. Dev Comp Immunol. 1998;22:367-377.
45. Pimenta-Araujo R, Mascarell L, Huesca M, CumanoA, Bandeira A. Embryonic thymic epithelium naturallydevoid of APCs is acutely rejected in the absence ofindirect recognition. J Immunol. 2001;167:5034-5041.
46. Mackall CL, Gress RE. Pathways of T-cell regen-eration in mice and humans: implications for bonemarrow transplantation and immunotherapy. Im-munol Rev. 1997;157:61-72.
47. Beschorner WE, Hutchins GM, Elfenbein GJ,Santos GW. The thymus in patients with alloge-neic bone marrow transplants. Am J Pathol.1978;92:173-181.
48. Seddik M, Seemayer TA, Lapp WS. The graft-versus-host reaction and immune function, II: re-cruitment of pre-T-cells in vivo by graft-versus-host-induced dysplastic thymuses followingirradiation and bone marrow treatment. Trans-plantation. 1984;37:286-290.
49. Fukushi N, Arase H, Wang B, et al. Thymus: adirect target tissue in graft-versus-host reactionafter allogeneic bone marrow transplantation thatresults in abrogation of induction of self-toler-ance. Proc Natl Acad Sci U S A. 1990;87:6301-6305.
50. Mowat A. Antibodies to IFN-gamma prevent im-munological mediated intestinal damage in mu-rine graft-versus-host reactions. Immunology.1989;68:18-24.
51. Brok HPM, Heidt PJ, van der Meide PH, ZurcherC, Vossen JM. Interferon-� prevents graft-versus-host disease after allogeneic bone marrow trans-plantation in mice. J Immunol. 1993;151:6451-6459.
52. Ellison CA, Fischer JM, HayGlass KT, GartnerJG. Murine graft-versus-host disease in an F1-hybrid model using IFN-gamma gene knockoutdonors. J Immunol. 1998;161:631-640.
53. Murphy WJ, Welniak LA, Taub DD, et al. Differen-
tial effects of the absence of interferon-� and IL-4in acute graft-versus-host disease after alloge-neic bone marrow transplantation in mice. J ClinInvest. 1998;102:1742-1748.
54. Yang YG, Dey BR, Sergio JJ, Pearson DA, SykesM. Donor-derived interferon gamma is requiredfor inhibition of acute graft-versus-host diseaseby interleukin 12. J Clin Invest. 1998;102:2126-2135.
55. Nikolic B, Lee S, Bronson RT, Grusby MJ, SykesM. Th1 and Th2 mediate acute graft-versus-hostdisease, each with distinct end-organ targets.J Clin Invest. 2000;105:1289-1298.
56. Gao EK, Kosaka H, Surh CD, Sprent J. T cellcontact with Ia antigens on nonhemopoietic cellsin vivo can lead to immunity rather than tolerance.J Exp Med. 1991;174:435-446.
57. Kosaka H, Surh CD, Sprent J. Stimulation of ma-ture unprimed CD8� T cells by semiprofessionalantigen-presenting cells in vivo. J Exp Med. 1992;176:1291-1302.
58. Zhang Y. APCs in the liver and spleen recruit acti-vated allogeneic CD8� T cells to elicit hepaticgraft-versus-host disease. J Immunol. 2002;169:7111-7118.
59. Dakic A, Shao QX, D’Amico A, et al. Developmentof the dendritic cell system during mouse ontog-eny. J Immunol. 2004;172:1018-1027.
60. Surh CD, Sprent J, Webb SR. Exclusion of circu-lating T cells from the thymus does not apply inthe neonatal period. J Exp Med. 1993;177:379-385.
61. Agus D, Surh C, Sprent J. Reentry of T cells tothe adult thymus is restricted to activated T cells.J Exp Med. 1991;173:1039-1046.
62. Westermann J, Smith T, Peters U, et al. Both acti-vated and nonactivated leukocytes from the pe-riphery continuously enter the thymic medulla ofadult rats: phenotypes, sources and magnitude oftraffic. Eur J Immunol. 1996;26:1866-1874.
63. Michie SA, Kirkpatrick EA, Rouse RV. Rare pe-ripheral T cells migrate to and persist in normalmouse thymus. J Exp Med. 1988;168:1929-1934.
64. Gopinathan R, DePaz HA, Oluwole OO, et al.Role of reentry of in vivo alloMHC peptide-acti-vated T cells into the adult thymus in acquiredsystemic tolerance. Transplantation. 2001;72:1533-1541.
65. Appelbaum FR. Haematopoietic cell transplanta-tion as immunotherapy. Nature. 2001;411:385-389.
66. Werner S. Keratinocyte growth factor: a uniqueplayer in epithelial repair processes. CytokineGrowth Factor Rev. 1998;9:153-165.
67. Min D, Taylor PA, Panoskaltsis-Mortari A, et al.Protection from thymic epithelial cell injury by ker-atinocyte growth factor: a new approach to im-prove thymic and peripheral T-cell reconstitutionafter bone marrow transplantation. Blood. 2002;99:4592-4600.
68. Rossi SW, Jeker LT, Ueno T, et al. Keratinocytegrowth factor (KGF) enhances postnatal T-celldevelopment via enhancements in proliferationand function of thymic epithelial cells. Blood.2007;109:3803-3811.
69. Erickson M, Morkowski S, Lehar S, et al. Regula-tion of thymic epithelium by keratinocyte growthfactor. Blood. 2002;100:3269-3278.
4088 HAURI-HOHL et al BLOOD, 1 MAY 2007 � VOLUME 109, NUMBER 9
For personal use only. by guest on June 8, 2013. bloodjournal.hematologylibrary.orgFrom