dipsogenic stimulation in ibotenic drn-lesioned rats induces concomitant sodium appetite
TRANSCRIPT

Neuroscience Letters 374 (2005) 5–10
Dipsogenic stimulation in ibotenic DRN-lesioned rats inducesconcomitant sodium appetite
Haerishton Rubim Cavalcante-Limaa, Hawlinston Rubim Cavalcante Limaa,Ricardo Henrique Costa-e-Sousaa, Emerson Lopes Olivaresa, Pedro Leonardo Cedraz-Merceza,
Rafael Oliveira Reisa, Daniel Badaue-Passos Jr.b, Waldecy De-Lucca Jr.b,Magda Alves de Medeirosa, Wellington da Silva Cortesa, Luıs Carlos Reisa,∗
a Department of Physiological Sciences, Institute of Biology, Federal Rural University of Rio de Janeiro, BR465, Km 7, 23890-000 Serop´edica, RJ, Brazilb Department of Physiology, Faculty of Medicine of Ribeir˜ao Preto, University of S˜ao Paulo, 14049-900 Ribeir˜ao Preto, S˜ao Paulo, Brazil
Received 14 July 2004; received in revised form 31 August 2004; accepted 5 October 2004
Abstract
tenic acidl estion of0 of similard asa 20,000,2 RN alsoe ion of DRNp ich increasec componenti e.©
K
SiFvcis
aua
N)VLT
on-lumeheseNGof
sendsandtores
ncesoup
0d
The main purpose of this study was to investigate whether dipsogenic stimuli influences the sodium appetite of rats with iboesion of the dorsal raphe nucleus (IBO-DRN). Compared to control, rats microinjected with phosphate buffer (PB-DRN), the ing.3 M NaCl was enhanced in IBO-DRN at 21 and 35 days after DRN lesion under a protocol of fluids and food deprivation. Despiteipsogenic response observed both in IBO-DRN and PB-DRN treated with isoproterenol (ISO, 300�g/kg, sc), the 0.3 M NaCl intake wgain significantly enhanced in IBO-DRN at 21 and 35 days post-lesion. Finally, treatment with polyethylene glycol (PEG, MW =0%, w/v, 16.7 ml/kg, sc) induced higher dipsogenic response in IBO-DRN than PB-DRN at 21 day after lesion. In addition, IBO-Dxpressed higher sodium appetite than PB-DRN, concomitantly with a drinking response. These results suggest that ibotenic lesromote an increase of the brain angiotensinergic response, possibly settled within the subfornical organ, through paradigms whirculating ANG II levels. The current paper supports the hypothesis that the ibotenic lesion of DRN suppresses a serotonergicmplicated on the modulation of the sodium appetite and, therefore, furthering homeostatic restoration of extracellular fluid volum
2004 Elsevier Ireland Ltd. All rights reserved.
eywords:Sodium appetite; Angiotensin II; Dipsogenic stimuli; Dorsal raphe nucleus; Serotonergic system
odium appetite constitutes homeostatic behavior involvedn the regulation of extracellular fluid (ECF) volume[10].orebrain structures as subfornical organ (SFO) andorganumasculosum laminae terminalis(OVLT) were recognized asomponents of this behavior. Thus, ECF volume depletionnduces to both sodium appetite and thirst mediated by thesetructures[10,17,27].
Nuclei of brainstem which receive both viscerosensorynd forebrain signals integrate inputs about changes in vol-me and tonicity and thus coordinate serotonergic mech-nisms implicated in the modulation of sodium appetite
∗ Corresponding author. Tel.: +55 21 26821210; fax: +55 21 26821763.E-mail address:[email protected] (L.C. Reis).
[11,20,23,25]. For instance, dorsal raphe nucleus (DRsends serotonergic projections toward SFO and O[2,6,32].
In this context, renal afferents transmit information ccerning decrease in sodium tubular load and/or ECF voand blood pressure induced by furosemide injection. Tinputs are sent toward forebrain where angiotensin I (AI) is converted into angiotensin II (ANG II) and activationangiotensinergic receptors also participates[8].
Probably, a subset of the sensory ascending fibersto the DRN through nucleus tractus solitarius (NTS)thereafter modulates natriorexigenic response which resECF volume in response to sodium depletion. Evidein this reasoning were recently postulated by Vivas’ gr
304-3940/$ – see front matter © 2004 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.neulet.2004.10.017

6 H.R. Cavalcante-Lima et al. / Neuroscience Letters 374 (2005) 5–10
which demonstrated c-Fos expression on the serotonergicneurons of DRN during spontaneous and induced saline in-gestion of sodium-depleted rats following peritoneal dialysis[11].
Data from our laboratory obtained using electrolytic or ex-citotoxic lesions models confirmed that surmising[23]. Earlyevidence provided by Reis and colleagues has shown thatelectrolytic lesion of the DRN induced acute decrease in se-cretory response of atrial natriuretic peptide (ANP), in basalcondition or after blood volume expansion[25]. In this line,the implication of the ANP it would be pertinent taken intoaccount its modulatory role on the natriorexigenic responseinduced by volume depletion[1,33].
Several evidences ascribe to ANP a role on the counterac-tion of the ANG II influence both in central nervous systemand peripheral tissues[1,10,14]. Distribution of specific ANPand ANG II receptors in the SFO constitutes the substantia-tion for a physiological antagonism between those systems inthe blood pressure control and hydroelectrolyte homeostasis[21].
On the other hand, raphe neurons interact reciprocally withstructures of the lamina terminalis implicated with hydroelec-trolyte homeostasis[10,18]. Among those structures early ev-idences have shown that the SFO establish bidirectional con-nections with the DRN[10,18,31]. Furthermore, HT2A/2Cand 5-HT1A serotonergic receptors were pharmacologicallyi tot
e hy-p ssi-b ts ofp sub-s RNw ddi-t onta-n dianp e tom
mal nse[ y to( os-m tiono e de-p t ANGI
ANGI pri-v af-t re-l tioni edi-a d byA
poth-e rain
angiotensinergic response on the forebrain structures impli-cated in sodium appetite mediation.
Thus, in the current study we employed experimental ma-neuvers which provoke directly or indirectly increase in cir-culating and/or brain ANG II challenged by (i) food and fluiddeprivation, (ii)�-adrenergic agonist (isoproterenol) or (iii)coloidosmotic polysaccharide (polyethylene glycol) admin-istration.
Considering the integrative role of raphe circuits on themodulation of salt intake mediated at ANG II-sensitive fore-brain structures we aimed to investigate the outcomes of thedipsogenic stimuli on the sodium appetite in rats with DRNibotenic acid lesion.
Male Wistar rats weighting 300–350 g were maintainedin room with lights on from 07:00 h to 19:00 h, controlledtemperature at 25◦C and free access to Purina chow andwater. Before brain stereotaxic surgery rats were housed inmetabolic cages during 5 days, at least. Excitotoxic lesionsof DRN were produced by local ibotenic acid microinjec-tion in rats anesthetized with tribromoethanol (2.5%, i.p.)and fixed in a Harvard stereotaxic frame. Ibotenic acid andtribromoethanol were purchased from Sigma Chemical Co.,St. Louis, MO, USA and Aldrich Chemical Company Inc.,Milwaukee, WI, USA, respectively. The experimental proto-cols and animal procedures were carried out in accordancewith current Brazilian legislation.
xicc r,7 nte-r en-t nso get RN.I -i hout1 sur-g r.A er-a ,C
forh ndlya botd ith0 . Ino icale stem( t ofd ours
mi-c ted,m ereu stionw olicc ve in-
dentified in neurons of the SFO in suchlike proportionhose ANG II-sensitive ones[31].
Regarding this statement, Tanaka’s group raised thothesis that SFO monitors cardiocirculatory and poly hydroelectrolytic oscillations, through measuremenlasma ANG II[38]. These authors demonstrated that aet of SFO neurons responsive to ANG II project to Dhere establish synapse with serotonergic neurons. A
ionally, they have evidenced that SFO neurons of the speously hypertensive rats (SHR) which send fibers to mereoptic nucleus (MnPOn) are significantly more sensitivicroiontophoretic administration of ANG II[39].Dipsogenic stimuli elicited by increase in ANG II plas
evels require mediation of SFO for the ingestive respo9,10,19]. Genesis of thirst and water intake secondari) hypovolemia induced by the administration of coloidotic substance, (ii) arterial hypotension and, (iii) activaf �-adrenergic receptors on the juxtaglomerular cells arendent on the increase of renin release and subsequen
I generation[9,10,13,14,16,26,28–30,35].On the other hand, there are evidences that brain
I plays a role in the osmotic thirst induced by water deation [5,10,30,40]. Late sodium appetite has occurreder water intake and secondarily to (i) increase of reninease, (ii) hypovolemia and hypotension or (iii) dehydranduced by water deprivation. These phenomena are mted by structures of the lamina terminalis and signaleNG II [3,4,7,10,36,41].Based on retrospective observations we raised the hy
sis that excitotoxic lesion of DRN would increase the b
Excitotoxic lesions of DRN were guided by stereotaoordinates of Paxinos and Watson[24]: anterioposterio.6–7.8 mm, posterior to the bregma (1.2–1.4 mm aior to the interaural line), lateral 0.0 mm and dorsovral, 6.4–6.6 mm, below the skull calvaria. Microinjectiof ibotenic acid were made with a Hamilton microsyrin
hough a guide cannula stereotaxically implanted into Dbotenic acid microinjections (1�g/0.2�l dissolved in sterle 0.2 M pH 7.4 phosphate buffer) were made througmin. Sham-lesioned rats were submitted to the sameical stress but microinjected with 0.2�l phosphate buffeprophylactic dose of penicilin was administered in op
ted rats (30,000 IU, i.m.) (Fort Dodge Saude Animal Ltdaampinas, SP, Brazil).At the end of 21 or 35 days the brains were removed
istological procedures. Therefore, the rats were profounesthetized with sodium thionembutal (100 mg/kg) (Abo Brazil, Sao Paulo, Brazil) for transcardiac perfusion w.15 M NaCl followed by buffered 10% paraformaldehyderder to confirm DRN–lesion sites we performed histologxamination of serial coronal sections through the brain30-�m tickness) stained by Nissl. Rats with lesions ouorsoventromedial region of DRN were discarded fromtudy (n= 5).
Rats with DRN–excitotoxic lesion produced by localroinjections of ibotenic acid (IBO-DRN) or sham-operaicroinjected with the phosphate buffer (PB-DRN) wsed. Measurements of water and 0.3 M NaCl ingeere fulfilled in burettes simultaneously placed in metabages. For each experimental condition a representati
H.R. Cavalcante-Lima et al. / Neuroscience Letters 374 (2005) 5–10 7
dex has calculated as follows: [0.3 M NaCl ingested]/[wateringested + 0.3 M NaCl ingested][12]. The levels of water andsaline intake as well as the health status of rats before surgeryprocedure have not shown any differences between groups.
PB-DRN and IBO-DRN were tested in following exper-imental conditions. (1) Water, sodium and food deprivationfor 16 h in IBO-DRN (n= 9) and PB-DRN (n= 10), 21 and35 days after surgery. At the day of experiments burettes withdistilled water and 0.3 M NaCl were returned to the metaboliccage. (2) Isoproterenol administration (ISO, 300�g/kg, sc) inPB-DRN (n= 9) and IBO-DRN at 7 (n= 9), 21 and 35 daysafter ibotenic acid surgery. Burettes with water and 0.3 MNaCl were immediately returned to the metabolic cages.(3) Polyethylene glycol administration in hyperoncotic dose(PEG, MW = 20,000, 20%, w/v, 16.7 ml/kg, sc) 21 days af-ter surgery. Following PEG injection, IBO-DRN (n= 10) andPB-DRN (n= 7) were returned to metabolic cages suppliedwith water and 0.3 M NaCl. All measurements of fluid inges-tion were accomplished throughout 300 min (60 min interval)and at the end of 24 h.
The data were analyzed by two-way analysis of variancewith repeated measures, and the significance of differencesbetween means were determined by Newman–Keuls test. Thesignificance level was set at 5%.
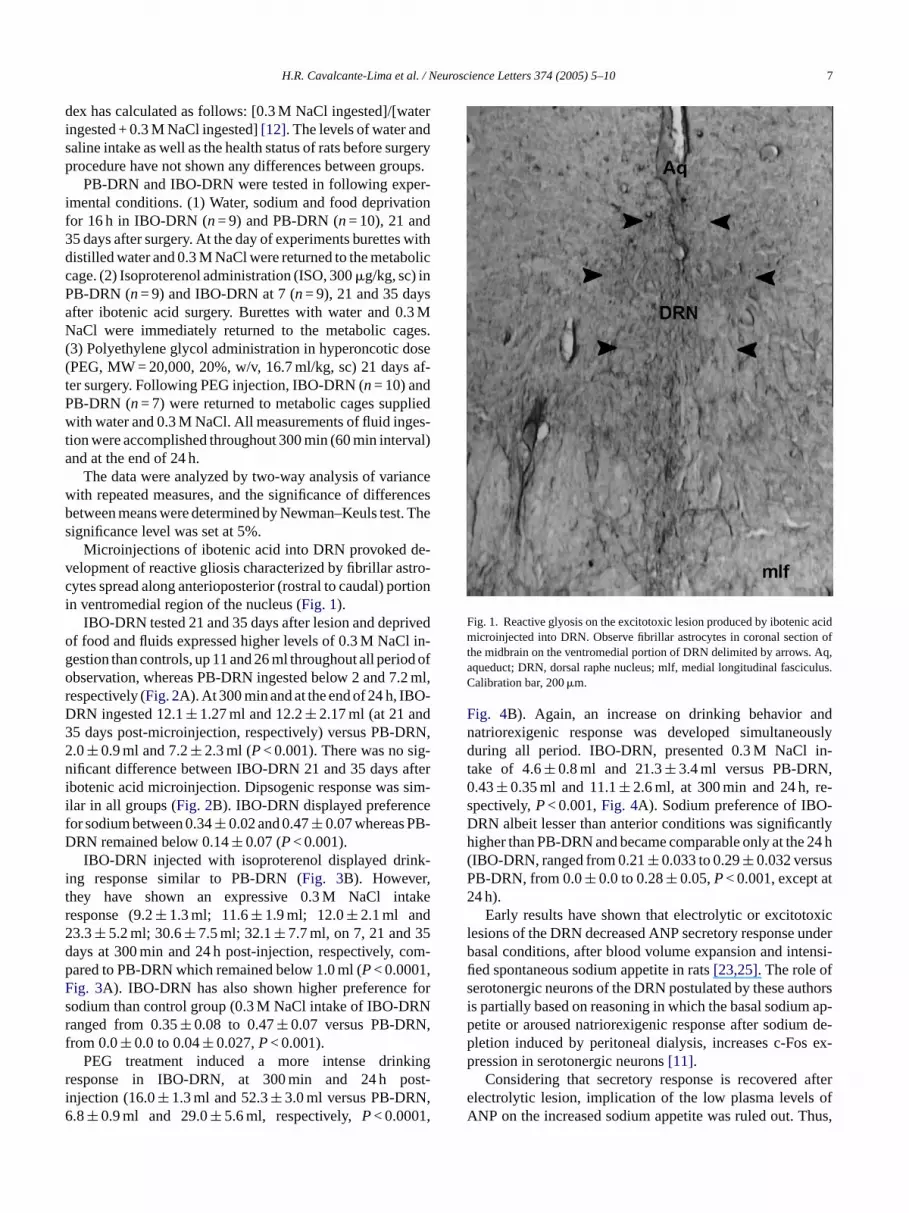
Microinjections of ibotenic acid into DRN provoked de-velopment of reactive gliosis characterized by fibrillar astro-c rtioni
ivedo l in-g d ofo 2 ml,r -D d3 RN,2 g-n fteri sim-i ef -D
k-i ,t aker2 5d om-p ,F fors Nr ,f
ingr st-i ,6 ,
Fig. 1. Reactive glyosis on the excitotoxic lesion produced by ibotenic acidmicroinjected into DRN. Observe fibrillar astrocytes in coronal section ofthe midbrain on the ventromedial portion of DRN delimited by arrows. Aq,aqueduct; DRN, dorsal raphe nucleus; mlf, medial longitudinal fasciculus.Calibration bar, 200�m.
Fig. 4B). Again, an increase on drinking behavior andnatriorexigenic response was developed simultaneouslyduring all period. IBO-DRN, presented 0.3 M NaCl in-take of 4.6± 0.8 ml and 21.3± 3.4 ml versus PB-DRN,0.43± 0.35 ml and 11.1± 2.6 ml, at 300 min and 24 h, re-spectively,P< 0.001,Fig. 4A). Sodium preference of IBO-DRN albeit lesser than anterior conditions was significantlyhigher than PB-DRN and became comparable only at the 24 h(IBO-DRN, ranged from 0.21± 0.033 to 0.29± 0.032 versusPB-DRN, from 0.0± 0.0 to 0.28± 0.05,P< 0.001, except at24 h).
Early results have shown that electrolytic or excitotoxiclesions of the DRN decreased ANP secretory response underbasal conditions, after blood volume expansion and intensi-fied spontaneous sodium appetite in rats[23,25]. The role ofserotonergic neurons of the DRN postulated by these authorsis partially based on reasoning in which the basal sodium ap-petite or aroused natriorexigenic response after sodium de-pletion induced by peritoneal dialysis, increases c-Fos ex-pression in serotonergic neurons[11].
Considering that secretory response is recovered afterelectrolytic lesion, implication of the low plasma levels ofANP on the increased sodium appetite was ruled out. Thus,
ytes spread along anterioposterior (rostral to caudal) pon ventromedial region of the nucleus (Fig. 1).
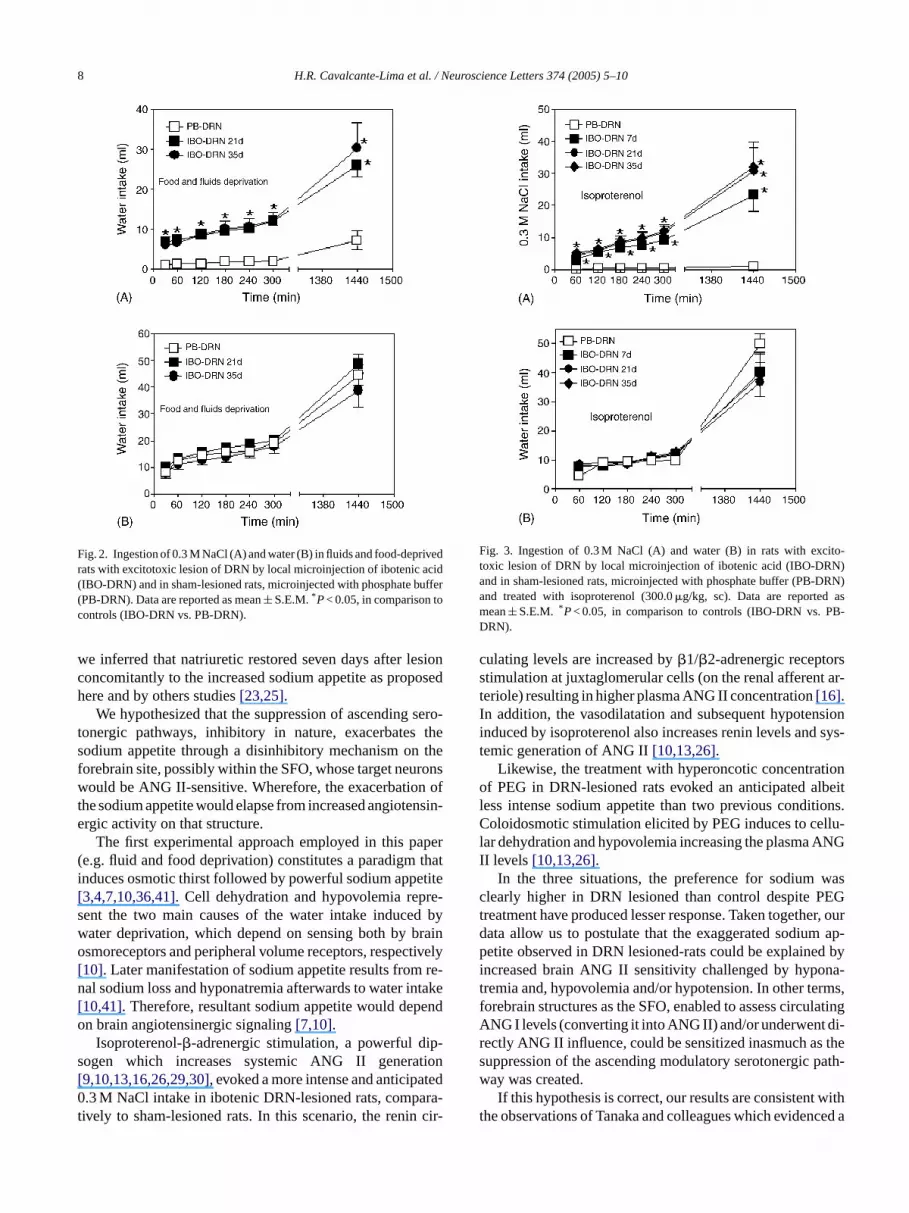
IBO-DRN tested 21 and 35 days after lesion and deprf food and fluids expressed higher levels of 0.3 M NaCestion than controls, up 11 and 26 ml throughout all periobservation, whereas PB-DRN ingested below 2 and 7.espectively (Fig. 2A). At 300 min and at the end of 24 h, IBORN ingested 12.1± 1.27 ml and 12.2± 2.17 ml (at 21 an5 days post-microinjection, respectively) versus PB-D.0± 0.9 ml and 7.2± 2.3 ml (P< 0.001). There was no siificant difference between IBO-DRN 21 and 35 days a
botenic acid microinjection. Dipsogenic response waslar in all groups (Fig. 2B). IBO-DRN displayed preferencor sodium between 0.34± 0.02 and 0.47± 0.07 whereas PBRN remained below 0.14± 0.07 (P< 0.001).IBO-DRN injected with isoproterenol displayed drin
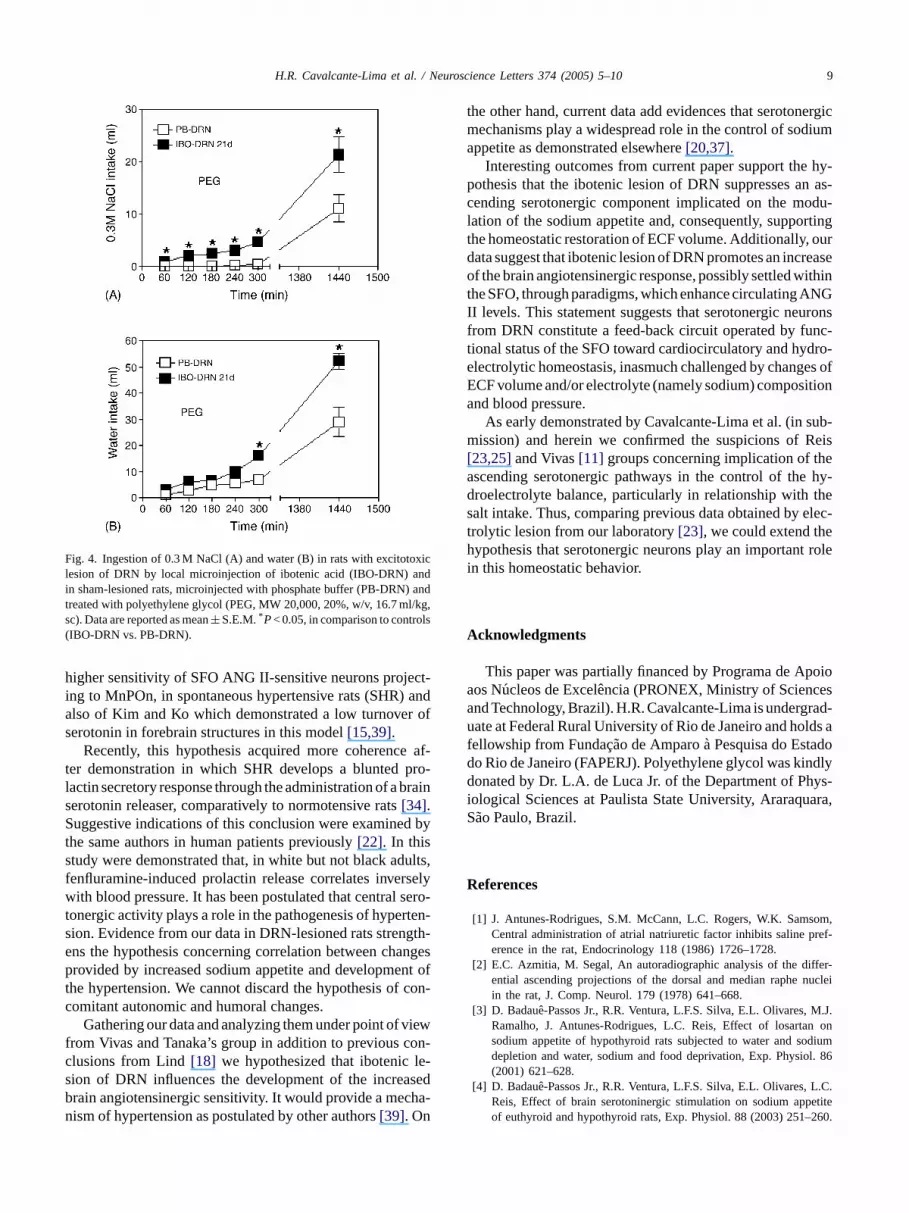
ng response similar to PB-DRN (Fig. 3B). Howeverhey have shown an expressive 0.3 M NaCl intesponse (9.2± 1.3 ml; 11.6± 1.9 ml; 12.0± 2.1 ml and3.3± 5.2 ml; 30.6± 7.5 ml; 32.1± 7.7 ml, on 7, 21 and 3ays at 300 min and 24 h post-injection, respectively, cared to PB-DRN which remained below 1.0 ml (P< 0.0001ig. 3A). IBO-DRN has also shown higher preferenceodium than control group (0.3 M NaCl intake of IBO-DRanged from 0.35± 0.08 to 0.47± 0.07 versus PB-DRNrom 0.0± 0.0 to 0.04± 0.027,P< 0.001).
PEG treatment induced a more intense drinkesponse in IBO-DRN, at 300 min and 24 h ponjection (16.0± 1.3 ml and 52.3± 3.0 ml versus PB-DRN.8± 0.9 ml and 29.0± 5.6 ml, respectively,P< 0.0001
8 H.R. Cavalcante-Lima et al. / Neuroscience Letters 374 (2005) 5–10
Fig. 2. Ingestion of 0.3 M NaCl (A) and water (B) in fluids and food-deprivedrats with excitotoxic lesion of DRN by local microinjection of ibotenic acid(IBO-DRN) and in sham-lesioned rats, microinjected with phosphate buffer(PB-DRN). Data are reported as mean± S.E.M.*P< 0.05, in comparison tocontrols (IBO-DRN vs. PB-DRN).
we inferred that natriuretic restored seven days after lesionconcomitantly to the increased sodium appetite as proposedhere and by others studies[23,25].
We hypothesized that the suppression of ascending sero-tonergic pathways, inhibitory in nature, exacerbates thesodium appetite through a disinhibitory mechanism on theforebrain site, possibly within the SFO, whose target neuronswould be ANG II-sensitive. Wherefore, the exacerbation ofthe sodium appetite would elapse from increased angiotensin-ergic activity on that structure.
The first experimental approach employed in this paper(e.g. fluid and food deprivation) constitutes a paradigm thatinduces osmotic thirst followed by powerful sodium appetite[3,4,7,10,36,41]. Cell dehydration and hypovolemia repre-sent the two main causes of the water intake induced bywater deprivation, which depend on sensing both by brainosmoreceptors and peripheral volume receptors, respectively[10]. Later manifestation of sodium appetite results from re-nal sodium loss and hyponatremia afterwards to water intake[10,41]. Therefore, resultant sodium appetite would dependon brain angiotensinergic signaling[7,10].
Isoproterenol-�-adrenergic stimulation, a powerful dip-sogen which increases systemic ANG II generation[9,10,13,16,26,29,30], evoked a more intense and anticipated0.3 M NaCl intake in ibotenic DRN-lesioned rats, compara-tively to sham-lesioned rats. In this scenario, the renin cir-
Fig. 3. Ingestion of 0.3 M NaCl (A) and water (B) in rats with excito-toxic lesion of DRN by local microinjection of ibotenic acid (IBO-DRN)and in sham-lesioned rats, microinjected with phosphate buffer (PB-DRN)and treated with isoproterenol (300.0�g/kg, sc). Data are reported asmean± S.E.M. *P< 0.05, in comparison to controls (IBO-DRN vs. PB-DRN).
culating levels are increased by�1/�2-adrenergic receptorsstimulation at juxtaglomerular cells (on the renal afferent ar-teriole) resulting in higher plasma ANG II concentration[16].In addition, the vasodilatation and subsequent hypotensioninduced by isoproterenol also increases renin levels and sys-temic generation of ANG II[10,13,26].
Likewise, the treatment with hyperoncotic concentrationof PEG in DRN-lesioned rats evoked an anticipated albeitless intense sodium appetite than two previous conditions.Coloidosmotic stimulation elicited by PEG induces to cellu-lar dehydration and hypovolemia increasing the plasma ANGII levels [10,13,26].
In the three situations, the preference for sodium wasclearly higher in DRN lesioned than control despite PEGtreatment have produced lesser response. Taken together, ourdata allow us to postulate that the exaggerated sodium ap-petite observed in DRN lesioned-rats could be explained byincreased brain ANG II sensitivity challenged by hypona-tremia and, hypovolemia and/or hypotension. In other terms,forebrain structures as the SFO, enabled to assess circulatingANG I levels (converting it into ANG II) and/or underwent di-rectly ANG II influence, could be sensitized inasmuch as thesuppression of the ascending modulatory serotonergic path-way was created.
If this hypothesis is correct, our results are consistent withthe observations of Tanaka and colleagues which evidenced a
H.R. Cavalcante-Lima et al. / Neuroscience Letters 374 (2005) 5–10 9
Fig. 4. Ingestion of 0.3 M NaCl (A) and water (B) in rats with excitotoxiclesion of DRN by local microinjection of ibotenic acid (IBO-DRN) andin sham-lesioned rats, microinjected with phosphate buffer (PB-DRN) andtreated with polyethylene glycol (PEG, MW 20,000, 20%, w/v, 16.7 ml/kg,sc). Data are reported as mean± S.E.M.*P< 0.05, in comparison to controls(IBO-DRN vs. PB-DRN).
higher sensitivity of SFO ANG II-sensitive neurons project-ing to MnPOn, in spontaneous hypertensive rats (SHR) andalso of Kim and Ko which demonstrated a low turnover ofserotonin in forebrain structures in this model[15,39].
Recently, this hypothesis acquired more coherence af-ter demonstration in which SHR develops a blunted pro-lactin secretory response through the administration of a brainserotonin releaser, comparatively to normotensive rats[34].Suggestive indications of this conclusion were examined bythe same authors in human patients previously[22]. In thisstudy were demonstrated that, in white but not black adults,fenfluramine-induced prolactin release correlates inverselywith blood pressure. It has been postulated that central sero-tonergic activity plays a role in the pathogenesis of hyperten-sion. Evidence from our data in DRN-lesioned rats strength-ens the hypothesis concerning correlation between changesprovided by increased sodium appetite and development ofthe hypertension. We cannot discard the hypothesis of con-comitant autonomic and humoral changes.
Gathering our data and analyzing them under point of viewfrom Vivas and Tanaka’s group in addition to previous con-clusions from Lind[18] we hypothesized that ibotenic le-sion of DRN influences the development of the increasedbrain angiotensinergic sensitivity. It would provide a mecha-nism of hypertension as postulated by other authors[39]. On
the other hand, current data add evidences that serotonergicmechanisms play a widespread role in the control of sodiumappetite as demonstrated elsewhere[20,37].
Interesting outcomes from current paper support the hy-pothesis that the ibotenic lesion of DRN suppresses an as-cending serotonergic component implicated on the modu-lation of the sodium appetite and, consequently, supportingthe homeostatic restoration of ECF volume. Additionally, ourdata suggest that ibotenic lesion of DRN promotes an increaseof the brain angiotensinergic response, possibly settled withinthe SFO, through paradigms, which enhance circulating ANGII levels. This statement suggests that serotonergic neuronsfrom DRN constitute a feed-back circuit operated by func-tional status of the SFO toward cardiocirculatory and hydro-electrolytic homeostasis, inasmuch challenged by changes ofECF volume and/or electrolyte (namely sodium) compositionand blood pressure.
As early demonstrated by Cavalcante-Lima et al. (in sub-mission) and herein we confirmed the suspicions of Reis[23,25]and Vivas[11] groups concerning implication of theascending serotonergic pathways in the control of the hy-droelectrolyte balance, particularly in relationship with thesalt intake. Thus, comparing previous data obtained by elec-trolytic lesion from our laboratory[23], we could extend thehypothesis that serotonergic neurons play an important rolein this homeostatic behavior.
A
poioa esa rad-u lds af dod ndlyd ys-i ara,S
R
som,ref-
ffer-uclei
M.J.n ondiuml. 86
L.C.etite260.
cknowledgments
This paper was partially financed by Programa de Aos Nucleos de Excelencia (PRONEX, Ministry of Sciencnd Technology, Brazil). H.R. Cavalcante-Lima is undergate at Federal Rural University of Rio de Janeiro and ho
ellowship from Fundac¸ao de Amparoa Pesquisa do Estao Rio de Janeiro (FAPERJ). Polyethylene glycol was kionated by Dr. L.A. de Luca Jr. of the Department of Ph
ological Sciences at Paulista State University, Araraquao Paulo, Brazil.
eferences
[1] J. Antunes-Rodrigues, S.M. McCann, L.C. Rogers, W.K. SamCentral administration of atrial natriuretic factor inhibits saline perence in the rat, Endocrinology 118 (1986) 1726–1728.
[2] E.C. Azmitia, M. Segal, An autoradiographic analysis of the diential ascending projections of the dorsal and median raphe nin the rat, J. Comp. Neurol. 179 (1978) 641–668.
[3] D. Badaue-Passos Jr., R.R. Ventura, L.F.S. Silva, E.L. Olivares,Ramalho, J. Antunes-Rodrigues, L.C. Reis, Effect of losartasodium appetite of hypothyroid rats subjected to water and sodepletion and water, sodium and food deprivation, Exp. Physio(2001) 621–628.
[4] D. Badaue-Passos Jr., R.R. Ventura, L.F.S. Silva, E.L. Olivares,Reis, Effect of brain serotoninergic stimulation on sodium appof euthyroid and hypothyroid rats, Exp. Physiol. 88 (2003) 251–

10 H.R. Cavalcante-Lima et al. / Neuroscience Letters 374 (2005) 5–10
[5] J.R. Blair-West, K.D. Carey, D.A. Denton, R.S. Weisinger, R.E.Sade, Evidence that brain angiotensin II is involved in both thirstand sodium appetite in baboons, Am. J. Physiol. 275 (1998)R1639–R1646.
[6] O. Bosler, L. Descarries, Monoamines innervation of the organumvasculosum laminae terminalis (OVLT): a high resolution radioauto-graphic study in the rat, J. Comp. Neurol. 272 (1988) 545–561.
[7] L.A. De Luca Jr., Z. Xu, G.H. Schoorlemmer, R.L. Thunhorst, T.G.Beltz, J.V. Menani, A.K. Johnson, Water deprivation-induced sodiumappetite: humoral and cardiovascular mediators and immediate earlygenes, Am. J. Physiol. 282 (2002) R552–R559.
[8] G.K. Fitch, M.L. Weiss, Activation of renal afferent pathways follow-ing furosemide treatment. II. Effect of angiotensin blockade, BrainRes. 861 (2000) 377–389.
[9] D.A. Fitts, Angiotensin II receptors in SFO but not in OVLT mediateisoproterenol-induced thirst, Am. J. Physiol. 267 (1994) R7–R15.
[10] J.T. Fitzsimons, Angiotensin, thirst, and sodium appetite, Physiol.Rev. 78 (1998) 583–686.
[11] L.F. Franchini, A.K. Johnson, J. de Olmos, L. Vivas, Sodium ap-petite and Fos activation in serotonergic neurons, Am. J. Physiol.282 (2002) R235–R243.
[12] S.P. Frankmann, P.M. Dorsa, R.R. Sakai, J.B. Simpson, A singleexperience with hyperoncotic colloid dialysis persistently alters waterand sodium intake, in: G. de Caro, A.N. Epstein, M. Massi (Eds.),The Physiology of Thirst and Sodium Appetite, Plenum Press, NewYork, 1986, pp. 115–121.
[13] A.K. Johnson, T.L. Thunshorst, Sensory mechanisms in the behav-ioral control of body balance: thirst and salt appetite, Prog. Psy-chobiol. Physiol. Psychol. 16 (1995) 145–176.
[14] A.K. Johnson, R.L. Thunhorst, The neuroendocrinology of thirst andsalt appetite: visceral sensory signals and mechanisms of central
[ uslyl. 20
[ oleinkin,
[ petitemina
[ ionsBrain
[ gan. 32
[ chialpetite
[ or,an-of
987)
[ ws,ucedon 32
[ .R.C.sion
of dorsal raphe nucleus on water intake and sodium appetite, Braz.J. Med. Biol. Res. 36 (2003) 1709–1716.
[24] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates,second ed., Academic Press, New York, 1986.
[25] L.C. Reis, M.J. Ramalho, A.L.V. Favaretto, J. Gutkowska, S.M. Mc-Cann, J. Antunes-Rodrigues, Participation of the ascending seroton-ergic system in the stimulation of atrial natriuretic peptide release,Proc. Natl. Acad. Sci. U.S.A. 91 (1994) 12022–12026.
[26] R. Rettig, D. Ganten, A.K. Johnson, Isoproterenol-induced thirst:renal and extrarenal mechanisms, Am. J. Physiol. 241 (1981)R152–R157.
[27] N.E. Rowland, M.J. Fregly, L. Han, G. Smith, Expression of Fos inrat brain in relation to sodium appetite: furosemide and cerebroven-tricular rennin, Brain Res. 278 (1996) 90–96.
[28] N.E. Rowland, B.E. Goldstein, K.L. Robertson, Role of angiotensinin body fluid homeostasis of mice: fluid intake, plasma hormones,and brain Fos, Am. J. Physiol. 284 (2003) R1586–R1594.
[29] W.A. Saad, L.A. Camargo, A. Renzi, L.A. de Luca Jr., J. Antunes-Rodrigues, W.A. Saad, Alterations in the water intake caused bycentral inhibition of angiotensin-converting enzyme in the rat, Neu-rosci. Lett. 134 (1992) 212–214.
[30] W.A. Saad, A.C. Luiz, L.A. Camargo, A. Renzi, J.E. Silveira, M.A.Paulino, W.A. Saad, Injection of ramipril into the lateral ventricleinterferes with the drinking response induced by pharmacological andnatural thirst stimuli, Braz. J. Med. Biol. Res. 27 (1994) 2909–2913.
[31] K.E. Scrogin, A.K. Johnson, H.A. Schmid, Multiple receptor sub-types mediate the effects of serotonin on rat subfornical organ neu-rons, Am. J. Physiol. 275 (1998) 2035–2042.
[32] H.W.M. Steinbusch, Distribution of serotonin-immunoreactivity inthe central nervous system of the rat cell bodies and terminals, Neu-roscience 6 (1981) 557–618.
[ te, J.
[ ked003)
[ t inNeu-
[ e in-siol.
[ In-cleuseu-
[ ofhem-
[ Dif-path-s be-nsive
[ of. J.
[ on,. 34
integration, Front. Neuroendocrinol. 18 (1997) 292–353.15] S.J. Kim, K.H. Ko, Abnormal serotonergic activities in spontaneo
hypertensive rats (SHR), Methods Find. Exp. Clin. Pharmaco(1998) 473–478.
16] R.F. Kirby, C.M. Novak, R.L. Thunhorst, A.K. Johnson, The rof beta1 and beta2 adrenoceptors in isoproterenol-induced drBrain Res. 656 (1994) 79–84.
17] J.M. Lane, J. Herbert, J.T. Fitzsimons, Increased sodium apstimulates c-fos expression in the organum vasculosum of the laterminalis, Neuroscience 78 (1997) 1167–1176.
18] R.W. Lind, Bi-directional, chemically specified neural connectbetween the subfornical organ and the midbrain raphe system,Res. 384 (1986) 250–261.
19] R.W. Lind, R.L. Thunhorst, A.K. Johnson, The subfornical orand the integration of multiple factors in thirst, Physiol. Behav(1984) 69–74.
20] J.V. Menani, R.L. Thunhorst, A.K. Johnson, Lateral parabranucleus and serotonergic mechanisms in the control of salt apin rats, Am. J. Physiol. 270 (1996) R162–R168.
21] F.A. Mendelsohn, A.M. Allen, S.Y. Chai, P.M. Sexton, R. FigdOverllaping distributions of receptors for natriuretic peptide andgiotensin II by in vitro autoradiography: morphological basisphysiological antagonism, Can. J. Physiol. Pharmacol. 65 (11517–1521.
22] M.F. Muldoon, A.F. Sved, J.D. Flory, J.M. Perel, K.A. MattheS.B. Manuck, Inverse relationship between fenfluramine-indprolactin release and blood pressure in humans, Hypertensi(1998) 972–975.
23] E.L. Olivares, R.H. Costa e Sousa, R.H. Cavalcante-Lima, HLima, P.L. Cedraz-Mercez, L.C. Reis, Effects of electrolytic le
33] E. Stellar, A.N. Epstein, Neuroendocrine factors in salt appetiPhysiol. Pharmacol. 42 (1991) 345–355.
34] S.D. Stocker, M.F. Muldoon, A.F. Sved, Blunted fenfluramine-evoprolactin secretion in hypertensive rats, Hypertension 42 (2719–724.
35] E.M. Stricker, Thirst and sodium appetite after colloid treatmenrats: role of the renin-angiotensin-aldosterone system, Behav.rosci. 97 (1983) 725–737.
36] E.M. Stricker, K.S. Gannon, J.C. Smith, Thirst and salt appetitduced by hypovolemia in rats: analysis of drinking behavior, PhyBehav. 51 (1992) 27–37.
37] J. Tanaka, Y. Hayashi, K. Yamato, H. Miyakubo, M. Nomura,volvement of serotonergic systems in the lateral parabrachial nuin sodium and water intake: a microdialysis study in the rat, Nrosci. Lett. 357 (2004) 341–344.
38] J. Tanaka, T. Okumura, K. Sakamati, H. Miyakubo, Activationserotonergic pathways raphe system to the subfornical organ byorrhage in the rat, Exp. Neurol. 169 (2001) 156–162.
39] J. Tanaka, Y. Yamamuro, H. Saito, M. Matsuda, M. Nomura,ferences in electrophysiological properties of angiotensinergicways from the subfornical organ to the median preoptic nucleutween normotensive Wistar–Kyoto and spontaneously hyperterats, Exp. Neurol. 134 (1995) 192–198.
40] R.S. Weisinger, J.R. Blair-West, D.A. Denton, E. Tarjan, Rolebrain angiotensin II in thirst and sodium appetite of sheep, AmPhysiol. 273 (1997) R187–R196.
41] R.S. Weisinger, D.A. Denton, M.J. McKinley, J.F. NelsDehydration-induced sodium appetite in rats, Physiol. Behav(1985) 45–50.