die analyse stabiler isotope (δ18o, δ13c, δ15n) an schmelz und dentin von hirschzähnen aus der...
TRANSCRIPT

Integrative Prähistorische und Naturwissenschaftliche Archäologie
(IPNA)
Universität Basel
Die Analyse stabiler Isotope (δ18O,
δ13C, δ15N) an Schmelz und Dentin
von Hirschzähnen aus der
horgenzeitlichen Seeufersiedlung
Zug-Riedmatt
Unterschiedliche Jagdgebiete im Laufe der
Besiedlung aufgrund wechselnder klimatischer
Bedingungen?
Masterarbeit
im Studiengang MSc PNA
von Elisa Schumpf
unter der Leitung von Dr. Prof. J. Schibler
2012/2013

1
Inhalt
1. Einleitung ....................................................................................................................................... 3
2. Archäologische und naturwissenschaftliche Grundlagen ........................................................ 4
2.1 Das Neolithikum und die Horgener Kultur in der Zentralschweiz ........................................... 4
2.1.1 Chronologie ..................................................................................................................... 4
2.1.2 Umwelt und Wirtschaft ................................................................................................... 5
2.2 Die Fundstelle Zug-Riedmatt ................................................................................................... 7
2.2.1 Lage und Zustand der Fundstelle .................................................................................... 7
2.2.2 Die Stratigraphie .............................................................................................................. 8
2.3 Der Hirsch .............................................................................................................................. 10
2.3.1 Biologie, Lebensraum und Nahrung .............................................................................. 10
2.3.2 Der Hirsch im Neolithikum ............................................................................................ 11
2.3.3 Zahnaufbau und –entwicklung des Rothirsches ............................................................ 11
2.4 Theoretische Grundlagen zur Isotopenanalyse ..................................................................... 14
2.4.1 Forschungsgeschichte ................................................................................................... 14
2.4.2 Allgemeine Grundlagen ................................................................................................. 15
2.4.3 Temperatur- und Klimarekonstruktion anhand von Sauerstoffisotopen (δ18O) ........... 16
2.4.4 Rekonstruktion von Ernährung und Lebensweise anhand von Kohlenstoff- und
Stickstoffisotopen (δ13C/δ15N) ....................................................................................... 18
2.4.4.1 Anwendungen von Kohlenstoffisotopen (δ13
C) zur Habitat- und
Ernährungsrekonstruktion ......................................................................................... 18
2.4.4.2 δ13
C in Karbonat (aus Apatit) und Kollagen .............................................................. 20
2.4.4.3 Anwendung von Stickstoffisotopen (δ15
N) zur Nahrungs- und Umweltrekonstruktion .
................................................................................................................................... 20
2.5 Die sequenzielle Beprobung von Zahnschmelz und Dentin .................................................. 22
2.5.1 Zahnschmelz .................................................................................................................. 22
2.5.2 Dentin ............................................................................................................................ 24
3. Material ......................................................................................................................................... 25
3.1 Auswahl des Probenmaterials ............................................................................................... 25
3.2 Wasserproben ....................................................................................................................... 26
4. Methoden ...................................................................................................................................... 27
4.1 Einführung in Mainz .............................................................................................................. 27
4.2 Die Analyse von Sauerstoff- und Kohlenstoffisotopen (δ18O/δ13C) aus dem Zahnschmelz .. 28
4.2.1 Die Vorbereitung der Zähne für die Beprobung ............................................................ 28

2
4.2.2 Sequenzierte Entnahme der Zahnschmelzproben ........................................................ 29
4.2.3 Aufreinigung des Zahnschmelzes .................................................................................. 30
4.2.4 Massenspektrometrie ................................................................................................... 31
4.3 Die Analyse von Kohlenstoff- und Stickstoffisotopen (δ13C/δ15N) aus dem Collagen im
Dentin .................................................................................................................................... 32
4.3.1 Vorbereitung der Zähne für die Beprobung .................................................................. 32
4.3.2 Sequenzierte Beprobung des Dentins ........................................................................... 32
4.3.3 Aufreinigung des Dentins und Extraktion des Kollagens ............................................... 33
4.3.4 Massenspektrometrie ................................................................................................... 34
4.4 Wasserproben ....................................................................................................................... 35
5. Resultate ....................................................................................................................................... 35
5.1 Die δ18O/δ13C – Werte aus dem Zahnschmelz....................................................................... 35
5.1.1 Die Messergebnisse aus Mainz und Basel im Vergleich ................................................ 35
5.1.2 Die Messergebnisse aus Mainz...................................................................................... 37
5.1.3 Messergebnisse aus Basel ............................................................................................. 39
5.2 Die δ13C/δ15N-Werte aus dem Collagen des Zahndentins ..................................................... 44
5.3 Die δ13C-Werte aus dem Karbonat und dem Kollagen im Vergleich ..................................... 48
5.4 Resultate Wasserproben ....................................................................................................... 49
6. Diskussion .................................................................................................................................... 50
6.1 Grosse Unterschiede im Verlauf der Isotopenkurven (δ18O/δ13C) aus dem Zahnschmelz –
Hinweis auf verschiedene Populationen oder normale Variation zwischen Individuen? ..... 50
6.2 Der Lebensraum – bewaldete Täler oder luftige Höhen? ..................................................... 52
6.3 Klimatische Bedingungen ...................................................................................................... 54
6.4 Unterschiede zwischen den Siedlungsphasen – ist eine Entwicklung zu erkennen? ............ 55
6.5 Synthese ................................................................................................................................ 56
7. Zusammenfassung ...................................................................................................................... 58
8. Tabellen ........................................................................................................................................ 59
9. Literaturverzeichnis .................................................................................................................... 64
10. Danksagung ................................................................................................................................. 72

3
1. Einleitung
Das Neolithikum ist die erste Epoche, in denen der Mensch nicht mehr nur von seinem Jagdglück
abhängig war, sondern durch Ackerbau und Viehzucht eine gewisse Sicherheit der
Nahrungsversorgung garantieren konnte. Dies setzte ein gewisses Mass an Sesshaftigkeit voraus, da
für den Anbau von Getreide Flächen gerodet werden mussten. Weil das Landschaftsbild im
Neolithikum hauptsächlich von Wald geprägt war und es kaum natürliche Freiflächen gab, war dies
eine aufwändige Arbeit. Nur durch eine ortsgebundene Lebensweise in Dörfern lohnte sich dieser
Aufwand, da durch die Rodung nicht nur Ackerland, sondern auch potentielles Weideland für die
Haustiere entstand. So konnte eine Grundmenge an Nahrungsmitteln produziert werden, welche die
Bevölkerung eines Dorfes sowohl im Sommer als auch im Winter zu ernähren vermochte.
Dennoch wurde im Neolithikum die Jagd nie ganz aufgegeben. Betrachtet man die neolithischen
Siedlungen in der Schweiz so fällt auf, dass die Anteile der Wildtiere im Nahrungsspektrum sehr stark
schwanken (Schibler 2006). Die Gründe dafür können vielfältig sein. Es gibt aber Hinweise für einen
möglichen Zusammenhang zwischen vermehrter Jagdtätigkeit und kurzzeitigen Klimaeinbrüchen.
Sehr nasse, kalte Sommer konnten beträchtliche Einbussen in den Ernteerträgen zur Folge gehabt
haben – was zu Nahrungsknappheit führen konnte. War es nicht möglich, diese Verluste durch
bestehende Vorräte oder durch die Schlachtung von Haustieren auszugleichen, musste die
Bevölkerung auf die wilden Ressourcen zurückgreifen. Dabei spielte vor allem der Rothirsch eine
wichtige Rolle, der im ganzen Neolithikum das wichtigste Jagdwild darstellte.
Hielt eine Klimadepression über längere Zeit an, musste die Jagd auf den Rothirsch intensiviert
werden, was zu erhebliche Schäden an lokalen Hirschpopulationen führen konnte. Für die Jäger
bedeutete dies, für eine erfolgreiche Jagd auf andere Gebiete mit intakten Populationen ausweichen
zu müssen. Bei der Betrachtung des archäozoologischen Materials aus Zug-Riedmatt fielen die hohen
Anteile an Hirschknochen auf - vor allem in den älteren Schichten, wo ein regelrechter
„Knochenhaufen“ von Hirschknochen zu finden war (Anteil Hirschknochen nach Gewicht 93%, mündl.
Mitteil. Sandra Billerbeck).
Ziel der vorliegenden Untersuchung ist es, mit Hilfe von Isotopenanalysen mögliche Unterschiede von
Lebensraum, Ernährung und Herkunft der Hirsche aus den archäologischen Schichten von Zug-
Riedmatt aufzudecken und so Hinweise auf das Jagdverhalten der Bevölkerung der horgenzeitlichen
Seeufersiedlung zu erlangen. Die Analyse von Sauerstoff-, Kohlenstoff- und Stickstoffisotopen an den
Hirschzähnen erlaubt Rückschlüsse auf verschiedene Faktoren der Lebensumstände der Tiere. Es
können Unterschiede in der Saisonalität, der Umwelt, der Nahrung und der klimatischen Bedingungen
sichtbar gemacht werden, was wiederum Hinweise auf verschiedene Hirschpopulationen und folglich
auf unterschiedliche Jagdgründe liefern könnte. Da die Hirschzähne aus verschiedenen
Siedlungsschichten aus einem Zeitraum von ca. 150 Jahren stammen wird auch versucht, allfällige
chronologische Entwicklungen oder Unterschiede zu erkennen und dadurch Hinweise auf eine
Klimaentwicklung und deren Einfluss auf die Jagd zu bekommen.

4
Abb. 1
Tabelle der Chronologie des Neolithikums in
der West-, Zentral- und Ostschweiz (Stöckli
1995, 21).
Die Isotopenanalysen wurden am IPNA (Integrative Prähistorische und Naturwissenschaftliche
Archäologie) Basel in Zusammenarbeit mit dem Institut für Umweltgeowissenschaften der Universität
Basel sowie am Departement für Anthropologie/Institut für Geowissenschaften an der Johannes-
Gutenberg-Universität Mainz durchgeführt. Da in Basel zuvor noch keine Erfahrungen mit
Isotopenanalysen an Zähnen und Knochen aus archäologischen Zusammenhängen gemacht wurden,
ist die erfolgreiche Durchführung dieses Pilotprojektes vor allem der Unterstützung des erfahrenen
Teams der Universität Mainz zu verdanken.
2. Archäologische und naturwissenschaftliche Grundlagen
2.1 Das Neolithikum und die Horgener Kultur in der Zentralschweiz
2.1.1 Chronologie
Die Horgener Kultur datiert zwischen 3500 und 2750 v.
Chr. und fällt in die Zeit des Spätneolithikums. In der
Zentralschweiz folgt sie auf die Cortaillod- bzw. Pfyn-
Kultur. Zu Beginn des 4. Jahrtausends. v. Chr. werden
die Fundstellen in der Zentralschweiz zum klassischen
zentralschweizerischen Cortaillod (Siedlung Egolzwil 5)
zugerechnet. Dieses weist aber schon Einflüsse aus
östlichen Kulturen auf. Um 3800 v. Chr. ist in den
Regionen des Zürichs- und Zugersees ein Wechsel zur
Pfyner Kultur (Zürich-Mozartstrasse Schicht 4, Risch-
Oberrisch) zu beobachten. Ab dieser Zeit scheint der
kulturelle Einfluss in der Zentralschweiz aus Osten bzw.
Mitteleuropa zu kommen. Das darauffolgende Horgen ist
anhand der Keramik sowie den Steinbeilklingen mit
rechteckigem Querschnitt dem östlichen Horgen
zuzuordnen (Abb. 1; Stöckli 1995, 35-36; Hafner/Suter
2003, 35).
Aus der Zeit des frühen Horgens um 3400 v. Chr. sind in
der Zentralschweiz einige Dendrodaten vorhanden,
jedoch ohne dazugehörige Fundkomplexe. Erst ab dem
älteren Horgen sind grössere Fundkomplexe aus mehreren Siedlungen vorhanden. So z.B. vom
Zürichseebecken Zürich Mozartstrasse Schicht 3, Zürich-Kleiner Hafner Schicht 3A/3B, Kansan
Seefeld Schicht 2-4, AKAD/Pressehaus Schicht E/G (ausführlich mit absoluten Datierungen Stöckli
2007, 54/64; Hafner/Suter 2003, 35), sowie Pfäffikon-Burg (Eberli 2010). Vom Zugersee, also der
unmittelbaren Umgebung der Fundstelle Zug-Riedmatt, sind mehrere Fundstellen aus der Zeit um

5
3200 v. Chr. bis 3000 v. Chr. bekannt, so z.B. Hünenberg-Dersbach/-Strandbad, Cham St. Andreas
und Zug-Schützenmatt (Abb. 2). Diese könnten teilweise gleichzeitig mit der Siedlung von Zug-
Riedmatt bestanden haben (Huber/Schaeren 2009, 113; Stöckli 2007, 90). Leider liegen für Zug-
Riedmatt noch keine absoluten Daten aus der Dendrochronologie vor.
Aufgrund der räumlichen Nähe von Zuger- und Zürichsee sind für allfällige Vergleiche zwischen den
Fundstellen die oben genannten Fundstellen zu bevorzugen. Obwohl STÖCKLI (Stöckli 2007, 90) bei
der Keramik von Zug-Schützenmatt und KanSan3 erhebliche Unterschiede feststellt, dürfte die
kulturelle Verbundenheit der beiden Regionen grösser gewesen sein als zur Westschweiz (Bieler- und
Neuenburgersee) sowie zur Bodenseeregion. Diese aus der Keramik ersichtlichen, kulturelle
Dreiteilung – in Westschweiz, Zürichseebecken/Zentralschweiz und Bodenseegebiet/Ostschweiz –
findet erst in der auf die Horgener Kultur folgende Schnurkeramik um ca. 2750 v. Chr. ein Ende
(Stöckli 2007, 196).
2.1.2 Umwelt und Wirtschaft
Ohne den Einfluss des Menschen durch die Landwirtschaft wäre Mitteleuropa im Neolithikum
vollständig von Wald bedeckt gewesen. So zeigen Pollenprofile aus dem Mesolithikum einen Anteil
von weit über 90% an windblütigen Gehölzen (Rösch 2010, 166).
Ab dem Frühneolithikum zeigen pollenanalytische Untersuchungen in der Nordschweiz
Veränderungen in der Vegetation, welche möglicherweise auf moderaten Ackerbau und Viehzucht
zurückzuführen sind. Funde aus der 2. Hälfte des 6. Jahrtausends belegen u.A. den Anbau von
Emmer, Einkorn und Lein (Schibler/Brombacher 1995, 87). Ausgeprägte Waldlandschaften zeigen
aber, dass keine grösseren Flächen entwaldet wurden. Gerodet wurde ausschliesslich für den
Siedlungsbau und dem Anlegen vom Ackerland mittels Brandrodung, worauf feine Holzkohlehorizonte
in Pollendiagrammen hinweisen (Schibler/Brombacher 1995, 90; Rösch 2010, 166). Schon in dieser
frühen Phase des Schweizer Neolithikums wurde der Fleischbedarf fast ausschliesslich über die
Haustierarten Schaf, Ziege und Rind gedeckt, wie Funde aus dem Wallis belegen (Schibler/Chaix
1995, 98-99).
Ab 4300 v. Chr. sind an den Seeufern des Schweizer Mittellandes die ersten Seeufersiedlungen zu
finden. Durch die Feuchtbodenerhaltung und der damit verbundenen guten Erhaltung von organischen
Materialien, vergrössert sich die Datengrundlage zur Interpretation von Wirtschaft und Umwelt
markant (Schibler/Chaix 1995, 102; Schibler 2010, 173). Die angebauten Kulturpflanzen sind jetzt vor
allem Nacktweizen, während Emmer und Einkorn an Bedeutung verlieren. Die Bodenbearbeitung auf
den Feldern war wenig intensiv, was ein längeres Brachestadium der Böden nach mehrjährigem
Anbau nötig machte. Es entstand eine Art „Flickenteppich“ von aktuellen Anbauflächen und Zonen mit
unterschiedlich fortgeschrittener Wiederbewaldung (Schibler/Brombacher 1995, 90; Rösch 2010, 166).
Die Anteile der Haustiere im Knochenspektrum liegen für diese Zeit bei etwa 50%, schwanken jedoch
in Relation zu den Wildtieren bis ca. 3500 v. Chr. immer wieder stark. Es konnte aufgezeigt werden,
dass die hohen Wildtieranteile direkt mit kurzfristigen Klimaschwankungen in Verbindung gebracht
werden können. In Phasen von nassen und kalten klimatischen Bedingungen wurde also vermehrt
gejagt. Da aber gleichzeitig die Fundkonzentrationen der Haustiere mehr oder weniger unverändert

6
blieben ist davon auszugehen, dass die intensivierte Jagd verminderte Ernteerträge von Getreide und
anderen Kulturpflanzen ausgleichen sollte. Aus den Funden der Schichten 4 u, m und o der Siedlung
Zürich „Mozartstrasse“ geht hervor, dass vor allem die Jagd auf den Rothirsch sowie das Sammeln
von kohlenhydrat- und fetthaltigen Samen intensiviert wurde (Schibler/Chaix 1995, 104; Schibler 2010,
173-178).
Ab der Horgener Zeit, also ab ca. 3400 v. Chr., sind schliesslich deutliche Veränderungen in der
Wirtschaftsweise feststellbar. Der Anbau von Emmer nimmt wieder zu und derjenige von Lein und
Mohn erreicht seinen Höhepunkt. Dies scheint die Folge intensiverer Bodennutzung zu sein, welche
zu einer Bodenverschlechterung führte und den Anbau von genügsameren Pflanzen erforderlich
machte. Die Brachezeiten wurden verkürzt und die um die Dörfer liegenden Felder längerfristig
bewirtschaftet. Gleichzeitig deutet ein Rückgang der sommerannuellen Unkräuter auf einen dichteren
Stand der Kulturpflanzen hin, woraus ein höherer Ertrag pro genutzte Fläche resultierte. Diese
Veränderungen in der Landwirtschaft deuten auf eine Zunahme der Bevölkerungsdichte hin, wodurch
der Druck auf die umliegende Natur stieg, da mehr offene Landschaftsfläche geschaffen werden
musste (Brombacher 1995, 87-92).
Die Bedeutung der Jagd (vor allem auf den Rothirsch) ist in der Horgener Kultur nicht einheitlich. In
vielen Fundstellen liegen die Anteile der Haustiere im Knochenspektrum mit ca. 60% höher als die
der Wildtiere (Schibler/Chaix 1995, 104-105). Neuere Untersuchungen zeigen jedoch, dass es auch in
der Zeit zwischen 3200 und 3000 v. Chr. grössere Schwankungen in den Anteilen von Wildtieren im
Knochenspektrum gibt, mit z.T. sehr hohen Werten von bis über 70% (Schibler 2006, Abb. 4, 55). Da
diese Schwankungen nicht von kulturellen Grenzen beeinflusst zu sein scheinen, wird auch hier eine
intensivierte Jagd bei gleichbleibender Dichte von Haustierknochen mit kurzzeitigen
Klimaschwankungen in Verbindung gebracht (Schibler 2006, 54-55).
Bei den Haustieren lässt sich in der Horgener Zeit eine deutliche Tendenz feststellen. War vor der
wirtschaftlichen Krise des 36. Jh. v. Chr. das Rind mit 50-60% mit Abstand das häufigste Haustier, so
übernahm jetzt das Schwein diese Rolle. In den horgenzeitlichen Siedlungen Arbon-Bleiche 3 und
jenen am Zürichsee liegen die Rinderanteile durchschnittlich bei 20% (s. Abb. 5 in Schibler 2006, 56;
Hüster-Plogmann 2002, 103). Der Stellenwert des Rindes scheint sich in dieser Zeit des
landwirtschaftlichen Aufschwungs verändert zu haben. Archäozoologische Untersuchungen haben
gezeigt, dass die Tiere erst in höherem Alter geschlachtet wurden und das Geschlechterverhältnis
ausgeglichen war, also auch männliche Tiere ein höheres Alter erreichten. Dies ist ein Hinweis auf
eine intensivierte Nutzung der Rinder als Zug- und Arbeitstiere, was als Begleiterscheinung der
wachsenden Landwirtschaft bzw. des Ackerbaus interpretiert werden kann (Hüster-Plogmann 2002,
105-106). Ebenfalls aus der Horgener Zeit bzw. aus der Siedlung Arbon-Bleiche 3 stammen
Nachweise von der Nutzung von Milch und Milchprodukten, die aus der Analyse von Fetten aus
angekohlten Speiseresten in Kochtöpfen hervorgehen (Schibler 2010, 177).

7
Abb. 2
Die Lage der fünf horgenzeitlichen Fundstellen Zug-Riedmatt (1), Zug-Schützenmatt (2), Zug-
Schutzengel/Bärenbächli (3), Hünenberg-Strandbad/-Derbach (4), Cham-St.Andreas, Strandbad (5) sowie der
bronzezeitlichen Fundstelle Zug-Sumpf (6). Angaben über die Lage der Fundstellen aus Huber/Schären
(2009). Karte: search.ch, swisstopo.
2.2 Die Fundstelle Zug-Riedmatt
2.2.1 Lage und Zustand der Fundstelle
Die Fundstelle Zug-Riedmatt liegt am nördlichen Zugerseeufer, in der sogenannten „Steinhauser
Bucht“ (Abb. 2, Nr.1). Sie befindet sich somit in der Nähe der bekannten Fundstellen Zug-Sumpf und
Zug-Schützenmatt (Abb. 2, Nr.6/Nr.2).
Da der Seespiegel und folglich auch der Grundwasserspiegel vor über 400 Jahren künstlich
abgesenkt wurden, waren die Schichten von Feuchtbodenfundstellen rund um den Zugersee von
partieller oder vollständiger Austrocknung betroffen. Dies wirkte sich negativ auf die Erhaltung von
organischem Material aus, da dieses sich im äroben Milieu rasch zersetzt (Huber/Schaeren 2009,
128).
Bei der Entdeckung der Siedlung Zug-Riedmatt im Jahre 2006 wurde aber aufgrund des guten
Zustandes der Pfähle in über 5 m Tiefe rasch klar, dass dieser Bereich immer noch auf oder unter
dem Grundwasserniveau liegen musste. Anschliessende Bohrungen ergaben eine Ausdehnung der
erhaltenen Siedlungsschichten von ca. 2500 m2. Da der Untergrund in diesem Gebiet sehr weich ist,
waren für das geplante Bauprojekt auf einer Fläche von etwa 230 m2 Fundationsträger geplant, welche
bis in die Tiefe der Siedlungsschichten gereicht und diese somit zerstört hätten. Da nur vier Monate für
die archäologische Ausgrabung zur Verfügung standen, war eine ausgedehnte Grabung nicht
realisierbar. In Absprache mit der Bauherrschaft konnten die Anzahl Pfähle und somit die Fläche der
Rettungsgrabung auf 80 m2 reduziert werden (Huber/Schaeren 2009, 128).
Die Kulturschichtpakete waren von 5 m Lorze- und Seesedimenten bedeckt und neigten sich gegen
den See hin um etwa 1,5 m. Welchen Einfluss nachträgliche Setzungsvorgänge oder
1
4
5
3
2
6

8
Abb. 3
Schichtpakete von Zug-Riedmatt mit
Kulturschichten und Schwemmhorizonten.
Zwischen dem
Überschwemmungshorizont C2 und der
Siedlungsphase D1/D2 ist ein grösserer
zeitlicher Hiatus zu vermuten (nach E.
Gross 2012, unpubl. Protokoll).
Hervorgehoben sind die
Siedlungsschichten, aus denen
Hirschzähne für die Isotopenanalyse zur
Verfügung standen.
Schichtpressungen hatten, ist beim momentanen Stand der Forschung noch nicht klar. Die Schieflage
und teilweise Zerstörung der Pfähle im Südwesten (Seeseite) der Grabung belegen aber, dass solche
Einflüsse eine Rolle spielten. Insgesamt konnten im ausgegrabenen Bereich 972 Pfähle identifiziert
werden, die z.T. fast einen Meter über die Kulturschicht hinaus ragten (Huber/Schaeren 2009, 129).
Die Funde bewegen sich in einem für die Horgener Zeit zu erwartendem Formenspektrum. Es wurden
grosse Mengen an Keramik geborgen, deren Erhaltungszustand aber aufgrund ihrer tiefen
Brenntemperatur sehr uneinheitlich war. Allgemein herrschte in den Schichten eine sehr grosse
Funddichte. Neben der Keramik gibt es über 400 Knochen- und Geweihartefakte, Steinbeile inkl.
Produktionsbestandteile und zahlreiche Silexwerkzeuge. Durch die guten Erhaltungsbedingungen
kommen auch spezielle Funde aus organischen Materialien dazu, wie etwa Schnüre, Zunderstücke,
Beilholme, ein sog. Horgener Messer und verschiedenste Artefakte aus Holz. Als aussergewöhnlich
darf wohl ein kleines Kupferbeil mit Randleisten bezeichnet werden (Huber/Schaeren 2009, 130).
2.2.2 Die Stratigraphie
Über die Stratigraphie der Fundstelle Zug-Riedmatt liegen noch keine publizierten Angaben vor. Die
folgenden Beschreibungen stammen aus Protokollen von Mai 2012, bereitgestellt von Eda Gross
(Kantonsarchäologie Zug). In der Stratigraphie konnten vier Schichtkomplexe ausgemacht werden, die
je eine Siedlungsphase darstellen. Getrennt werden diese durch organische
Überschwemmungshorizonte, wobei die letzte Siedlungsphase durch einen Erosionshorizont
abgeschlossen wird (Abb. 3). Eine grobe Datierung war vorerst nur durch die Keramik- und
Geweihartefakttypologie möglich. Demnach datieren die Kulturschichten in die Zeit zwischen 3200 v.
Chr. und 3000 v. Chr.
G Lorze-Sand und Torfe
F Limnische Ablagerungen nach Erosion
E3 Erosionshorizont nach Schichtkomplex E
E2 Organische Überschwemmungsschicht
E1 Schichtkomplex, Siedlungsphase E
D3 Organische Überschwemmungsschicht
D1/D2 Schichtkomplex, Siedlungsphase D
_____________________________________
C2 Organische Überschwemmungsschicht
C1 Schichtkomplex, Siedlungsphase C
B3 Organische Überschwemmungsschicht
B2 Schichtkomplex, Lehmschicht
B1 Schichtkomplex, Knochenhaufen
B Schichtkomplex B allgemein
A3 Organische Überschwemmungsschicht
A2 Seekreide, Installationshorizont
A1 Seekreide
A Seekreide Strandplatte vor Besiedlung
A L
T
J
U N
G

9
Die Ausrichtung verschiedener Strukturen und die Lage der Lehmlinsen lassen eine zeitliche Nähe der
Entstehung der Phasen B und C sowie D und E vermuten. Die Überschwemmungsschicht C2 scheint
also markanter zu sein als die anderen Überschwemmungsereignisse. Diese Beobachtung bestätigt
sich in der typochronologischen Gliederung der Zwischenfutter. Es bestehen Unterschiede zwischen
den Siedlungsschichten, vor allem aber zwischen den Phasen C und D. Auch bei der groben
Durchsicht der Tierknochen ist dort eine Veränderung zu beobachten. Während in den Schichten B
und C sehr viele Wildtierknochen (vor allem Hirsch) vorhanden sind, steigt in den jüngeren Phasen der
Anteil an Haustierknochen (v.a. Schwein) und Wildtiere – insbesondere Hirsch – nehmen deutlich ab
(mündl. Mitteil. Sandra Billerbeck).
Die Besiedlung scheint sich im Laufe der Zeit von Nordosten nach Südwesten verschoben zu haben.
Das heisst, die Strukturen der ältesten Besiedlungsphasen liegen am nordöstlichen Ende der
Grabungsfläche und beinhalten u.a. den Knochenhaufen. Nachweise von Hüttenböden oder
Feuerstellen fehlen in dieser Phase, diese könnten weiter landeinwärts und somit ausserhalb der
Grabungsfläche gelegen haben oder die Häuser dieser Phase waren architektonisch anders
konstruiert. Der Knochenhaufen wird am Rand von einer dünnen organischen Schicht überdeckt,
darüber folgt eine kompakte, nicht geschichtete Lehmphase. In diesem Bereich ist die
Überschwemmungsschicht B3 nicht vorhanden, was die eindeutige Trennung zwischen den Phasen B
und C erschwert.
Die Siedlungsphase C ist durch Aschelinsen und markante Keramikkonzentrationen mit recht gut
erhaltenen Gefässen geprägt. Hüttenböden und Herdstellen fehlen aber weiterhin. Ob es sich um die
Überreste eines Brandereignisses oder um Abraum von gereinigten Feuerstellen handelt, ist nicht
geklärt. Die Überschwemmungsschicht C2 ist im seewärtigen Bereich deutlich ausgeprägt, auf den
erhöhten Lagen landeinwärts nur partiell. Dies führt wiederum zu Schwierigkeiten in der
durchgehenden Trennung der Phasen C und D.
Siedlungsphase D liegt deutlich weiter seewärts, sie ist charakterisiert durch eine durchgehende,
dicke, organische Schichtabfolge. Sie wird praktisch auf der ganzen Fläche unterteilt durch eine
dünne (Bau-?)Lehmlage. Die Schicht ist von Keramikkonzentrationen und liegenden Hölzern
durchzogen. Die darüber liegende Überschwemmungsschicht ist z.T. gekappt oder erodiert.
Die jüngste Siedlungsphase E ist stark durch Erosion beeinflusst, sie ist vor allem durch grosse
Lehmlinsen repräsentiert die aufgrund ihres Gewichts in den weichen Untergrund einsanken und so
erhalten blieben. Diese Lehmlinsen liegen direkt über denjenigen von Schicht D und können
möglicherweise mit bestimmten Pfostenreihen (v.a. Eichenspältlinge und Eiben) in Verbindung
gebracht werden. Da aufgrund des intensiven Schneitelungseinflusses noch keine
dendrochronologische Datierung der Pfähle möglich war, sind bisher keine eindeutigen
Hausgrundrisse erkennbar. Im Umkreis der Lehmlinsen sind grosse Steinplatten,
Keramikkonzentrationen und kreuzweise geschichtete Hölzer zu beobachten.

10
2.3 Der Hirsch
2.3.1 Biologie, Lebensraum und Nahrung
Der Rothirsch (Cervus elaphus) gehört zur Ordnung der Paarhufer (Artiodactyla) und zur
Unterordnung der Wiederkäuer (Ruminantia). Die Gattung der Edelhirsche (Cervus) umfasst rund 15
Arten, wobei die ersten „Ur-Rothirsche“ vor ca. einer Million Jahre auftraten (Righetti 1995, 434-435).
Heutzutage ist der Rothirsch in der Schweiz mit Schulterhöhen bis zu 150 cm und einem Gewicht bis
260 kg das grösste einheimische Säugetier (Righetti 1995, 434). Die durchschnittliche Grösse der
mitteleuropäischen Hirsche (männlich, ausgewachsen) liegt aber wohl eher bei etwa 115 cm bis max.
124 cm (Wagenknecht 1996, 38). Dies ist deutlich kleiner als die errechneten Grössen für
prähistorische Zeiträume. In Arbon-Bleiche 3 zum Beispiel schwanken die Wiederristhöhen der
männlichen Tiere zwischen 125 und knapp 132 cm (Deschler-Erb/Marti-Grädel 2004, 196).
Hirsche leben in Familien- sowie Junggesellen-Rudeln. Diese verlassen sie lediglich in der Brunftzeit
im September, wenn die Männchen möglichst viele Weibchen um sich scharen und ihr Harem gegen
Rivalen verteidigen. Nach der Brunft im Oktober kehren Männchen sowie Weibchen wieder in ihre
Rudel zurück, bis die Hirschkühe im Mai und Juni nach ca. 34 Wochen Tragezeit abseits der Herde
meist ein Kalb gebären (Righetti 1995, 435; Bützler 1986, 128). Normalerweise erfolgt die
Entwöhnung des Kalbes im Alter von 5-7 Monaten. Wird die Mutter im Herbst aber nicht erneut
trächtig, kann das Kalb bis weit in den nächsten Sommer hinein gesäugt werden (Bützler 1986, 131-
132).
Der bevorzugte Lebensraum des Rothirsches sind grundsätzlich lichte Auenwälder im Tal, wo er auch
im Winter genügend Nahrung findet. In der warmen Jahreszeit wandert er in höhere Lagen der
Voralpen und Alpen, im Hochsommer sogar über die Baumgrenze hinaus. Die Besiedlung der Täler
durch den Menschen sowie die Rodung der Auenwälder und Nutzung der Alpen führte aber immer
mehr dazu, dass der Hirsch sich in grössere, dichte Waldgebiete zurückzog (Bützler 1986, 125,
Righetti 1995, 436). Durch diesen menschlichen Einfluss kann der heutige Lebensraum und somit
auch die Lebensweise des Rothirsches z.T. nicht mehr als natürlich betrachtet werden. Es zeigt aber
die aussergewöhnliche Anpassungsfähigkeit dieser Art.
Der Rothirsch wird in Bezug auf seine Nahrung als Intermediärtyp bezeichnet. D.h. er verwertet
sowohl Rauhfutter wie auch reichhaltigere Nahrung wie Gräser und Früchte. Das Nahrungsspektrum
umfasst darüber hinaus auch Knospen, Zweige und Rinde von Bäumen und Sträuchern sowie
Flechten, Moose und Pilze (Bützler 1986, 125-127). Untersuchungen haben gezeigt, dass der Hirsch
über 90% der verfügbaren Pflanzen als Nahrung nutzt und einige Arten wie z.B. Esche, Feldahorn und
Sahlweide dadurch stark dezimiert werden können (Bützler 1986, 126). Täglich benötigt der Rothirsch
3 bis 4 kg Futter und deckt darüber auch den Grossteil seines Flüssigkeitsbedarfs (Righetti 1995,
436).

11
2.3.2 Der Hirsch im Neolithikum
Während des gesamten Schweizer Neolithikums war der Rothirsch die wichtigste Jagdbeute
(Schibler/Hüster-Plogmann 1995, 77; Schibler 1997a, 60; Schibler et al. 1997b, 89; Schibler/Steppan
1999, 95; Schibler 2006, 60). In den zahlreichen neolithischen Seeufersiedlungen erreicht der Hirsch
in gewissen Zeitabschnitten Anteile in den Wildtierüberesten von bis zu 90% (Schibler/Steppan 1999,
95). Besonders in der Zeit von 3950 bis ca. 3600 v. Chr. ist eine sehr intensive Hirschjagd
festzustellen, die zu einer messbaren negativen Beeinflussung der Rothirschpopulationen geführt hat.
Dazu ist eine anhaltende intensive Bejagung von 80-120 Jahren notwendig, was sogar zu lokaler
Ausrottung der Hirsche führen kann (Schibler/Hüster-Plogmann 1995, 77; Schibler et al. 1997b, 93;
Schibler/Steppan 1999, 95; Schibler 2006, 60). Anhand der Hirschknochen aus den Schichten 4-6 der
Fundstelle Mozartstrasse am Zürichsee konnte gezeigt werden, dass eine hohe Bejagung zu einer
geringeren Körpergrösse der Hirsche führte (Schibler et al. 1997b, 96; Schibler/Steppan 1999, 95-96).
Gleichzeitig ist anzunehmen, dass die Dichte der Hirschpopulationen im Umkreis der
Seeufersiedlungen abnahm, was zu einem grösseren Einzugsgebiet des Jagdrevieres geführt haben
könnte.
In den ersten Horgener Siedlungen am Zürichsee im 33. Jh. v. Chr. ist der Anteil der Hirschknochen
mit weniger als 10% im Knochenspektrum relativ gering, stieg aber bis zum 28. Jh. v. Chr. erneut
kontinuierlich an. Neben der Haltung von Schweinen nahm die Bedeutung der Jagd also wieder zu
(Schibler et al. 1997b, 97). Die Schwankungen lassen sich primär mit dem unterschiedlichen Anteil
von Fleisch in der Gesamternährung erklären. Bei Ernteausfällen aufgrund schlechter klimatischer
Bedingungen oder kurzfristigen negativen Einflüssen, liess sich das Defizit an produzierter Nahrung
nicht mit vermehrtem Schlachten von Haustieren ausgleichen, sondern führte zu einem höheren
Konsum von Wild und Sammelfrüchten wie Nüssen und Früchten (Schibler/Hüster-Plogmann 1995,
116; Schibler/Steppan 1999, 93). Dieses Phänomen ist vor allem für die Zeit der Cortaillod- und Pfyn-
Kultur zwischen 4300 und 3400 v. Chr. belegt (Schibler/Hüster-Plogmann 1995, 116), kann aber auch
für steigenden Wildtier-Anteilen im Horgen verantwortlich sein. In der Fundstelle von Zug-Riedmatt
erscheinen die Anteile der Rothirsche im Vergleich zu den Haustieren relativ hoch, was auf eine
negative Veränderung des Klimas in dieser Zeit schliessen lässt.
2.3.3 Zahnaufbau und –entwicklung des Rothirsches
Die Zahnformel des Hirsches ist 0 1 3 3
3 1 3 3. Die Zähne bestehen wie bei allen Säugetieren aus einer
Krone (Corona dentis) und einer oder mehreren Wurzeln (Radix dentis), der Übergang von Krone zur
Wurzel wird als Zahnhals (Collum dentis) bezeichnet. Sowohl die Krone als auch die Wurzel sind aus
Dentin oder Zahnbein aufgebaut, das eine knochenähnliche Zusammensetzung hat. Das Dentin der
Zahnkrone wird von Zahnschmelz (Enamelum) umschlossen, die Zahnwurzeln sind mit
knochenähnlichem Zahnzement (Cementum) überzogen (Brown/Chapman 1991, 371; Vohberger
2011, 22 nach Drenckhahn/Fleischhauer 1994; Knipper 2011, 130 nach Schumacher et al. 1990)

12
Abb. 4
Bildung von Zahnschmelz an einem
menschlichen Eckzahn. (Knipper
2011, 131, Abb. 8.2 nach Vorlage
von Hillson & Antoine 2003, Fig. 6.l)
Zahnschmelz ist das härteste vom Körper gebildete Gewebe. Es hat mit 95 Gewichts-% einen viel
höheren Anteil an mineralischen Komponenten als Dentin (ca. 70 Gew-%) oder Knochen (ca. 45
Gew.-%). Die Mineralphase besteht hauptsächlich aus karbonathaltigem Hydroxylapatit
(mineralogische Bezeichnung: Dahllit; Summenformel Ca10(PO4)6OH2), das neben den
Hauptelementen Kalzium und Phosphor unter anderem auch Karbonat enthält, was für die Messung
von Sauerstoff- und Kohlenstoffisotopen entscheidend ist. Die Apatitkristalle sind sehr gross und
bilden regelmässige Schmelzprismen, die sehr dicht angeordnet sind. Dadurch resultiert eine viel
geringere Porosität als bei Dentin oder Knochen, was den Zahnschmelz sehr hart und
widerstandsfähig macht. Es ermöglicht eine gute Erhaltung über Jahrtausende, ohne dass die
Element- oder Isotopenzusammensetzung diagenetisch verändert würde. Höchstens an der
Oberfläche kann es zum Austausch von in Wasser gelösten Ionen kommen. Entscheidend ist, dass
der Zahnschmelz nach seiner Bildung keinem aktiven Stoffwechsel unterliegt, d.h. er enthält keine
lebenden Zellen. Das Verhältnis und die Konzentration von
Elementen und Isotopen im Zahnschmelz verändert sich also
nach seiner Entstehung nicht mehr (u.a. Tütken 2010, 35-36;
zusammenfassend Knipper 2011, 130 und Knipper 2004, 14-
15).
Die Bildung und Mineralisation des Zahnschmelzes beginnt an
der Schmelz-Dentin-Grenze an der Zahnspitze und wächst
schrittweise von oben nach unten respektive nach aussen. So
entstehen verschieden Schichten, die sich schuppenförmig
überlappen. Trotz dieser Überschneidung bildet sich eine
chronologische Abfolge des gebildeten Zahnschmelzes von
oben nach unten bis zur Schmelzgrenze am Zahnhals (Abb.
4). Dies ermöglicht eine zeitliche Auflösung durch
sequenziertes Beproben des Zahnschmelzes (s. Kapitel 2.4.4)
(Suga 1982; Brown/Chapman 1991, 88; zusammenfassend
Knipper 2011, 131).
Das Dentin macht den grössten Teil des Zahnes aus, es bildet den Kern der Zahnkrone sowie die
Wurzel. Es ist viel weicher als der Zahnschmelz, jedoch härter als Knochen und besteht zu 70% aus
Hydroxylapatit. Die restlichen organischen Anteile sind Wasser (10%) und Kollagen (20%). Es können
drei Arten von Dentin unterschieden werden: Primär-, Sekundär- und Tertiärdentin. Das Primärdentin
bildet die Zahnkrone und –wurzel und wird während des Wachstums des Zahns gebildet. Danach folgt
am Rande der Pulpahöhle das Sekundärdentin, das eine ähnliche Struktur aufweist wie das
Primärdentin. Davon unterscheidet sich das Tertiärdentin, das punktuell bei Beschädigung des Zahns
durch Abrasion oder Karies entstehen kann (Knipper 2004, 15 u.a. nach Frank/Nalbandian 1989 und
Schumacher et al. 1990).
Wie auch beim Zahnschmelz beginnt das Wachstum des Dentins in der Kronspitze des Zahns und
setzt sich bis zur Wurzel fort. Dies kann wesentlich länger dauern als die Bildung des Zahnschmelzes.
Da das Dentin keinen festen Umbauprozessen unterliegt wie Knochen, entsteht auch hier eine

13
chronologische Abfolge der geochemischen Zusammensetzung des Dentins ähnlich wie beim
Zahnschmelz. Dieser Effekt betrifft aber nur das Primärdentin: das später gebildete Sekundär- oder
Tertiärdentin kann durchaus eine andere chemische Zusammensetzung mit abweichenden
Isotopenverhältnissen aufweisen als das Primärdentin. Dies kann bei einer sequenzierten Beprobung
des Dentins zu einer Abschwächung oder Verfälschung der sich chronologisch verändernden
Isotopenwerte kommen, da es nicht möglich ist, ausschliesslich das Primärdentin zu beproben. Dazu
kommt, dass Dentin viel poröser ist als Zahnschmelz, was eine höhere Kontaminationsgefahr bei
längerer Bodenlagerung zur Folge hat (Knipper 2004, 15-20 u.a. nach Buddecke 1981,
Frank/Nalbandian 1989 und Schumacher et al. 1990).
Über die Zahnentwicklung beim Hirsch liegen aus älteren und jüngeren Quellen sehr unterschiedliche
Daten vor, aus denen hervorgeht, dass der Zeitpunkt der Zahnmineralisation und –durchbruch je nach
Individuum stark variieren kann. Im Folgenden werden nur der 2. und 3. Molar näher betrachtet, da
diese aufgrund ihrer relativ späten Entwicklung für die Isotopenanalysen in dieser und anderen
Arbeiten verwendet werden.
BROWN (1991) untersuchte die Zahnentwicklung von 113 rezenten Rothirschen aus halbwilden
Verhältnissen aus dem Richmond Park (Surrey, UK), wobei nur 82 Individuen für die
Altersklassenbildung verwendet werden konnten, da bei den restlichen Hirschen die Zahnentwicklung
nach dem 40. Lebensmonat schon abgeschlossen war. Es wurden Röntgenbilder gemacht, um den
Stand des Wachstums, der Mineralisation und der Wurzelentwicklung eruieren zu können. So konnten
sechs Stufen (a-g) der Zahnentwicklung bestimmt werden, die zur Altersbestimmung bis zum 50.
Lebensmonat herangezogen werden können (s. Brown 1991, 88-89). Gemäss diesen
Untersuchungen beginnt die Mineralisation des M2 schon vor dem 3. Lebensmonat und ist spätestens
mit 9 Monaten abgeschlossen. Die Entwicklung dieses Zahns scheint für alle Individuen recht
einheitlich zu sein, es wurden keine grösseren Unterschiede festgestellt. Anders beim M3, wo die
erste Mineralisation zwischen dem 9. und 13. Lebensmonat beginnen kann und zwischen dem 18.
und 26. Lebensmonat abgeschlossen ist (Brown 1991, 91). Es scheinen also beim Hirschen
Entwicklungsunterschiede bei der Zahnschmelzbildung des M3 von über einem halben Jahr möglich
zu sein. Dies könnte sich auch in den Isotopenwerten der vorliegenden Untersuchung zeigen, da die
Werte des Zahnschmelzes von der Jahreszeit, in der er gebildet wurde, beeinflusst werden (s. Kapitel
2.4.3).
Wie neuere Untersuchungen von AZORIT ET AL. (2002) zeigen, gibt es nicht nur grosse Unterschiede
beim Mineralisationszeitraum der Zähne, sondern auch beim Zeitpunkt ihres vollständigen
Durchbruchs. Es wurden Unterkiefer von 158 rezenten spanischen Hirschen aus Gefangenschaft im
Alter zwischen 4 und 44 Monaten untersucht. Von jedem Hirsch war der Zeitpunkt seiner Geburt
bekannt, was ein exaktes Bestimmen seines Alters ermöglichte (Azorit et al. 2002, 108). Die
beobachteten Zahndurchbruchsalter wurden mit älteren Forschungsergebnissen von verschiedenen
Autoren aus Europa verglichen. Dabei variiert der Zeitpunkt des vollständigen Wachstums des M2
zwischen 12 Monaten (Wagenknecht 1980; Muller-Using 1971; Mitchell 1967; Godawa 1989) und 14-
16 Monaten (O.N.C. 1984; Mitchell 1967; Azorit et al. 2002). Beim M3 schwanken die Werte noch
stärker, nämlich zwischen 21 Monaten (Wagenknecht 1980; Muller-Using 1971) und 31-32 Monaten

14
(Mariezkurrena 1983; Mitchell 1967; Azorit et al. 2002), wobei die hinterste Säule des M3 sogar erst
mit 37-44 Monaten gänzlich hochgewachsen ist (Azorit et al 2002, 112). Das bedeutet, dass bei
Hirschen bis zum 3. Lebensjahr kaum mit Abrasion an der Kaufläche des M3 zu rechnen ist. Dies ist
ein Vorteil für die Beprobung des Zahns für Isotopenanalysen, da in diesem Fall keine Reduktion der
im Zahnschmelz repräsentierten Zeitspanne auftritt. Bei älteren Individuen verringert sich die Höhe der
Zahnkrone mit jedem Lebensjahr – im vorliegenden Material war die Krone eines M3 von einem 10-
jährigen Individuum noch knapp 11mm hoch, während die Höhe eines M3 ohne Abrasion bei jüngeren
Hirschen bis zu 25mm betrug.
2.4 Theoretische Grundlagen zur Isotopenanalyse
2.4.1 Forschungsgeschichte
Die Analyse stabiler Isotope aus Knochen oder Zähnen in archäologischen Zusammenhang wird
schon seit den 60er- Jahren angewendet, um Informationen über (paläo-)klimatische Bedingungen
und später auch über Klimaentwicklung, Umwelt, Lebensräume, Migration und Ernährung von
Menschen oder Tieren zu gewinnen. Die ersten Forschungen beschäftigten sich mit der
Klimageschichte anhand der Sauerstoffisotopen-Verhältnissen in Molluskenschalen, Knochen und
Zähnen aus maritimem Umfeld (z.B. Longinelli 1966). Später wurden sie auf terrestrische Systeme
und verschiedenste archäologische Fundorte ausgeweitet (Stephan 2008, Übersicht in Longinelli
1995).
Es gibt mittlerweile unzählige Arbeiten, die anhand von Sauerstoff-, Kohlenstoff- und
Stickstoffisotopen Erkenntnisse zu Klima (z.B. Fricke/O‘Neil 1996; Fricke et al.1998; Stephan 1999;
Zazzo 2002; Bernard et al. 2009), Umwelt (z.B. Cerling/Herris 1999; Balasse 2002; Hoppe et al.
2004), Ernährung (z.B. Luz/Kolodny 1985; Ambrose/Norr 1993; Richards et al. 2002;
Bocherens/Drucker 2003; Bösl et al. 2006; Frémondeau et al. 2012) und Lebensbedingungen/Mobilität
(z.B. Balasse et. al 2002; Bentley/Knipper 2005; Balasse/Ambrose 2005; Balasse/Tresset 2007;
Balasse et al. 2012a; Knipper 2011) von Menschen und Tieren aus allen Epochen gewinnen konnten.
Mit der Entwicklung des Verfahrens der sequentiellen Beprobung von Zahnschmelz entlang der
vertikalen Achse von hochkronigen Tierzähnen in den 90er- Jahren erweiterten sich die Möglichkeiten
zur Untersuchung von Saisonalität, Nahrungsumstellungen, Lebensraumwechsel und kurzzeitigen
Klimaschwankungen erheblich (siehe Kapitel *Sequenzierung* und Zusammenstellung von Balasse
2003, 3). Mittlerweile wird diese Art der Beprobung standardmässig zur Untersuchung von
hochkronigen Zähnen angewendet. Die meisten Resultate gibt es für Rinderzähne (z.B. Balasse et al.
2001; Balasse/Tresset 2002; Zazzo et al. 2005; Hoppe 2006; Blasse/Tresset 2007; Knipper 2011),
Schafszähne (z.B. Balasse et al. 2002; Balasse et al. 2003; Balasse et al. 2006; Balasse et al. 2012a;
Balasse et al. 2012b), Schweinezähne (z.B. Richards et al. 2002; Frémondeau 2012) und
Pferdezähne (z.B. Hoppe 2004; Stevens/Hedges 2004).
Der Hirsch eignet sich als weit verbreitetes und sehr anpassungsfähiges Säugetier gut für
Isotopenanalysen aller Art (Saisonalität, Klimarekonstruktion und –entwicklung,

15
Landschaftsrekonstruktion etc.). DRUCKER ET AL. (2003) führten Messungen von Kohlenstoff- und
Stickstoffisotopen im Knochenkollagen von holozänen Hirschen aus dem nördlichen Jura
(Rochedane) durch (Drucker et al. 2003). Die Ergebnisse wurden mit Werten einer früheren
Auswertung aus Frankreich verglichen, in der ebenfalls Hirschknochen aus verschiedenen
archäologischen Schichten (Präboreal bis Bölling-Alleröd) beprobt wurden (Drucker 2001;
Drucker/Célérier 2001). In einer weiteren Untersuchung der δ18
O-Werte des Knochenphosphates
derselben Individuen aus Rochedane konnten DRUCKER ET AL. aufzeigen, dass eine positive
Korrelation zwischen den δ15
N- und den δ18
O-Werten besteht (Drucker et al. 2009). Eine neuere
Studie zeigt anhand von Kohlenstoff-, Stickstoff-, Sauerstoff- und Sulfurisotopen die unterschiedliche
Entwicklung von Hirschpopulationen im Französischen Jura und den westlichen Alpen von der
Späteiszeit bis zum frühen Holozän (Drucker et al. 2011). Überreste von Hirschen aus
archäologischen Schichten sind jedoch weitgehend nur in Einzelfällen beprobt worden, zusammen mit
der restlichen Fauna aus der jeweiligen Epoche bei Betrachtung eines ganzen Ökosystems (z.B.
Jedrzejewski et al. 2000; Bösl et al. 2006).
Zwei sehr umfangreiche Studien wurden an rezenten Hirschen aus Nordeuropa durchgeführt.
STEVENS ET AL. (2006) beprobten Knochenkollagen von 154 Hirschen aus fünf Populationen aus
Grossbritannien und Polen. Es konnten so wichtige Erkenntnisse über die Interpretierbarkeit von
Kohlenstoff- und Stickstoffisotopenwerte gewonnen werden (Stevens et al. 2006). In einer weiteren
Studie wurde erstmals der Zahnschmelz von Hirschen sequentiell beprobt. Dabei handelte es sich um
neun rezente, ausgewachsene Hirsche von der Isle of Rum, Schottland. Es konnte u.a. gezeigt
werden, dass die δ18
O-Werte aus den 2. und 3. Molaren der Hirsche eine positive Korrelation mit den
Sauerstoffisotopen-Kurven des Niederschlags im jahreszeitlichen Verlauf aufweisen (Stevens et al.
2011).
2.4.2 Allgemeine Grundlagen
Die Isotope eines Elements unterscheiden sich lediglich in ihrer Anzahl Neutronen im Atomkern und
weisen alle prinzipiell dieselben chemischen Eigenschaften auf. Sauerstoff hat drei stabile Isotope
(16
O, 17
O, 18
O), Kohlenstoff und Stickstoff deren zwei (12
C, 13
C; 14
N, 15
N). Grundlegend für die Isotopie
ist die Tatsache, dass die Isotope aufgrund ihrer unterschiedlichen Masse (± 1-2 Neutronen) in
physikalischen, chemischen und biologischen Prozessen unterschiedliche
Reaktionsgeschwindigkeiten aufweisen. Dadurch kommt es im Verlaufe dieser Prozesse zu einer
Veränderung der Isotopenverhältnisse, was als Isotopenfraktionierung bezeichnet wird. Innerhalb
einer Probe wird immer das leichtere (häufigere) Isotop zu dem schwereren (selteneren) Isotop ins
Verhältnis gesetzt. Dieses wird gegen einen internationalen Standard verglichen und die Abweichung
davon wird als δ-Wert in Promille (‰) angegeben. Folgende Gleichung wird dafür verwendet (Sharp
2005; Hoefs 2009):
δ (‰) = [(RProbe – RStandard)/(RStandard)] * 1000
Wobei R = 18
O/16
O; 13
C/12
C; 15
N/14
N

16
Abb. 5
Vereinfachte Darstellung
der Fraktionierung von
Sauerstoffisotopen
während dem
atmosphärischen
Wasserzyklus (Stephan
2008, nach Hoefs 1980).
Resultiert aus dieser Gleichung ein positiver δ-Wert, so ist der Anteil des schwereren Isotopes in der
Probe höher als im Standard (Anreicherung), bei einem negativen δ-Wert ist der Anteil geringer als im
Standard (Abreicherung). Diese Standards werden üblicherweise verwendet (Sharp 2005; Stephan
2008; Hoefs 2009):
18O/
16O (aus Phosphatgruppe): VSMOW = Vienna Standard Mean Ozean Water (Ozeanwasser)
18O/
16O (aus strukturellem Karbonat): PDB = Pee Dee Belemnit (aus kreidezeitlichem Fossil,
mittlerweile durch andere Standards ersetzt, die auf PDB kalibriert sind)
13C/
12C: PDB = Pee Dee Belemnit (aus kreidezeitlichem Fossil, mittlerweile durch andere Standards
ersetzt, die auf PDB kalibriert sind)
15N/
14N: AIR = Atmosphärischer Stickstoff N2
2.4.3 Temperatur- und Klimarekonstruktion anhand von Sauerstoffisotopen (δ18O)
Sauerstoffisotope im Wasser durchlaufen zwei Fraktionierungsprozesse zwischen ihrem Ursprung im
Ozean und dem Verhältnis, wie sie in biogenem Hartgewebe (Knochen oder Zahn) von Tieren oder
Menschen eingelagert werden. Die erste Fraktionierung findet im Verlauf des atmosphärischen
Wasserzyklus statt. Sie beginnt mit dem Verdampfen von Wasser aus dem Ozean. Dabei verdunsten
die leichteren H216
O-Moleküle eher als die schwereren H218
O-Moleküle, es kommt also zu einer
Abreicherung von 18
O im Wasserdampf. Daraus folgt, dass der Niederschlag, der aus diesem zu
Wolken kondensierten Wasserdampf auf die Erde fällt („meteorisches Wasser“), in jedem Fall negative
δ18
O-Werte aufweist.
Durch das bevorzugte Abregnen von H218
O-Molekülen wird dieser Effekt verstärkt, je weiter die
Regenwolken schon gezogen sind. Der fallende Niederschlag wird mit 18
O angereichert, im
übriggebliebenen Wasserdampf hingegen sinkt die Konzentration, d.h. es resultieren niedrigere δ18
O-
Werte (Abb. 5). Folgende vier Effekte sind dabei entscheidend (Stephan 2008; Knipper 2011 u.a.
nach Dansgaard 1964, Sharp 2005 und Stephan 1999):

17
Kontinentaleffekt: Je weiter im Inland bzw. je weiter vom Ozean entfernt, umso negativer
werden die δ18
O-Werte des Niederschlags im Vergleich zum Ozeanwasser.
Höheneffekt: Durch die verstärkte Abreicherung von 18
O an Gebirgen resultieren in höheren
Lagen besonders niedrige δ18
O-Werte.
Temperatureffekt (besonders wichtig): Bei hohen Temperaturen ist die Konzentration von
18O im Niederschlag grösser als bei niedriger Temperatur. Dies gilt sowohl für längerfristige
(klimatische) Bedingungen einer Region, als auch für die jahreszeitlichen
Temperaturschwankungen. D.h. in warmen Regionen (oder im Sommer) sind die δ18
O-Werte
höher bzw. weniger negativ als in kälteren Regionen (oder im Winter). In der Schweiz
betragen die Schwankungen der δ18
O-Werte im Niederschlag zwischen kalter und warmer
Jahreszeit ca. 4-6‰ (Schotterer et al. 2000).
„amount-Effekt“, Niederschlagsmenge: Bei sehr starken saisonalen Schwankungen in der
Niederschlagsmenge lassen sich in Trockenzeiten höhere δ18
O-Werte feststellen als in der
Niederschlagsreichen Zeit (Regenzeit).
Die zweite Fraktionierung der Sauerstoffisotope findet innerhalb der Säugetiere während der
Aufnahme und Metabolismus bis zum Einbau des Sauerstoffs in das biogene Gewebe statt. Dabei
besteht eine positive Korrelation zwischen den δ18
O-Werten des Trinkwassers und dem δ18
O-Wert im
Phosphat des Knochen- oder Zahnapatits. Dieser Zusammenhang kann mittels einer
Regressionsgleichung beschrieben werden, die aufgrund der unterschiedlichen
Nahrungszusammensetzung und Stoffwechsel je Tierart variiert (Stephan 1999; Zusammenstellung
siehe Stephan 2008, 52-53). Da schon sehr früh eine direkte Korrelation zwischen Temperatur und O-
Isotopenverhältnis im Niederschlag nachgewiesen wurde (Dansgaard 1964), besteht so im besten Fall
die Möglichkeit, mithilfe der δ18
O-Werten aus Knochenphosphat von Faunenresten den 18
O-Anteil im
Niederschlag und somit die Durchschnittstemperatur eines archäologisch relevanten Zeitraums zu
bestimmen (Stephan 1999; Stephan 2008).
Für den Hirsch wurde bisher keine empirische Regressionsgleichung für den Zusammenhang von
δ18
O-Werten im Trinkwasser und Knochenphosphat erstellt. An den rezenten Hirschen von Schottland
gelang es STEVENS ET AL. (2011) aber, einen linearen Zusammenhang zwischen den δ18
O-Werten
aus dem Karbonat des Zahnschmelzes und den Sauerstoffisotopenverhältnissen im Niederschlag
bzw. der Temperatur zu ermitteln. Die Umrechnung der Maxima bzw. Minima der Isotopenwerte im
Zahnschmelz in Durchschnittstemperaturen von Sommer und Winter der entsprechenden Lebensjahre
der Hirsche erwies sich beim Vergleich mit den aufgezeichneten Temperaturmessungen als zutreffend
(Stevens et al. 2011, 68). Dies könnte die Grundlage für zukünftige Versuche, mithilfe von δ18
O-
Werten aus Hirschzähnen Hinweise auf paläoklimatische Verhältnisse zu gewinnen, bilden. Da aber
die Auswirkung von unterschiedlichen Lebensräumen von Hirschpopulationen auf die Fraktionierung
von Sauerstoffisotopen innerhalb der Tiere (noch) nicht abgeschätzt werden kann, sind
Rekonstruktionen von klimatischen Bedingungen aufgrund solcher Modelle mit Zurückhaltung zu

18
handhaben. Sie könnten allenfalls als Ergänzung zu anderen klimaanalytischen Methoden wie z.B.
Pollenanalyse, Seeablagerungen oder Eisbohrkernen dienen.
Weiter ist zu bedenken, dass das Trinkwasser von Wildtieren (z.B. Hirsche) nicht direkt von kürzlich
gefallenem Niederschlag stammen muss. Möglich ist auch ein Wasserbezug aus Flüssen, Seen und
Tümpeln, in denen sehr unterschiedliche Verweilzeiten des Wassers vorliegen und sich Niederschläge
aus längeren Zeiträumen vermischen. Dies kann zu einer Dämpfung des Isotopensignals im
Zahnschmelz führen und somit saisonale Unterschiede nahezu unsichtbar machen (vertiefend z.B.
Fricke et al. 1996; Hoppe et al. 2004; Stevens et al. 2011).
2.4.4 Rekonstruktion von Ernährung und Lebensweise anhand von Kohlenstoff- und
Stickstoffisotopen (δ13C/δ15N)
Die Kohlenstoff- und Stickstoffisotopenwerte im Gewebe eines Säugetiers stehen in direktem
Zusammenhang zu denjenigen seiner aufgenommenen Nahrung (z.B. DeNiro/Eppstein 1981;
Ambrose 1993; Ambrose/Norr 1993; Ambrose 2000). Die isotopische Zusammensetzung in der
Nahrung sowie die Fraktionierung zwischen Nahrung und Konsument hängen von verschiedensten
Umweltfaktoren ab, deren Einfluss und Wirkungsweise z.T. noch ungeklärt oder umstritten sind. Dies
hat zur Folge, dass das Ausmass der möglichen Variabilität von Isotopenwerten auch innerhalb einer
Population im gleichen Habitat nicht genau eingeschätzt werden kann. Solange gewisse
Zusammenhänge und ihr Ablauf nicht an einer genügend grossen Anzahl von Proben belegt sind und
rekonstruiert werden können, sollte bei der Interpretation von absoluten Aussagen abgesehen werden
(Bocherens/Drucker 2003; Stevens et al. 2006).
2.4.4.1 Anwendungen von Kohlenstoffisotopen (δ13
C) zur Habitat- und
Ernährungsrekonstruktion
Der Kohlenstoff in den Pflanzen stammt aus dem atmosphärischen Kohlendioxid in der Luft und wird
im Laufe der Photosynthese in die Zellen eingebaut. Beim Verzehr der Pflanzen durch Landsäugetiere
werden die Kohlenstoffisotope aufgenommen, wobei weitere Fraktionierungsprozesse ablaufen.
Unabhängig von den δ13
C-Werten der Pflanzen beträgt die 13
C-Anreicherung im Kollagen von
Herbivoren gegenüber der Nahrung ca. 5‰ und im Apatit ca. 9-14‰ (z.B. DeNiro/Eppstein 1978;
Ambrose/Norr 1993; Cerling/Harris 1999).
Im Folgenden werden die wichtigsten Fraktionierungsprozesse von Kohlenstoffisotopen kurz
vorgestellt. Eingegangen wird vor allem auf diejenigen, die für die Interpretation der vorliegenden
Daten der Hirsche aus Zug-Riedmatt relevant sind.
Canopy-Effekt (Baldachin-Effekt): Pflanzen, die unter einem geschlossenen Blätterdach
wachsen haben bis zu 5‰ tiefere δ13
C-Werte als die Vegetation im Offenland oder in den
Baumkronen (z.B. Vogel 1978; Tieszen 1991). Die Wissenschaft macht mehrere Gründe für
diesen Umstand verantwortlich (siehe Drucker 2003, 379-380), u.a. die geringere
Luftzirkulation am Waldboden, veränderte CO2-Konzentration (Durchmischung von biogenem
und atmosphärischem CO2 vermindert), geringerer Lichteinfall sowie Wasserverfügbarkeit.

19
Der Canopy-Effekt wurde anfangs vor allem in tropischen bzw. äquatorialen Gebieten
beobachtet (z.B. Tieszen/Button 1989; Broadmeadow et al. 1992). Mittlerweile wird er aber
auch in Studien aus Mitteleuropa zur Rekonstruktion von Lebensräumen miteinbezogen (z.B.
Bocherens et al. 1999; für Hirsche Drucker et al. 2003; Stevens et al. 2006; Drucker et al.
2009; Drucker et al. 2011).
DRUCKER ET AL. (2003) haben mit Hilfe von δ13
C-Werten aus modernem Kollagen von
Rehen aus Frankreich (aus Rodière et al. 1996) einen maximalen δ13
C-Wert von -22.5‰
errechnet, den Tiere bzw. Hirsche im Kollagen aufweisen sollten, wenn sie ausschliesslich
Nahrung vom Waldboden unter geschlossenem Blätterdach konsumieren (Drucker et al.
2003, 381). Dieser Wert ist aber lediglich eine Konstruktion und konnte in einem späteren
Artikel (Drucker et al. 2008) nicht absolut bestätigt werden. Er kann aber als Richtlinie dienen,
um bei grösseren Probenmengen mit hoher Variabilität einen allfälligen Unterschied der
Lebensräume in Betracht ziehen zu können.
Unterscheidung C3/C4-Pflanzen: Während der Photosynthese von C3-Pflanzen (Calvin-
oder C3-Zyklus) wird stärker gegen das schwere Isotop 13C diskriminiert als bei der
Photosynthese von C4-Pflanzen (Hatch-Slack- oder C4-Zyklus). Dies hat zur Folge, dass C3-
Pflanzen mit durchschnittlich ca. -26.5‰ deutlich tiefere δ13
C-Werte aufweisen als C4-
Pflanzen mit ca. -12.5‰. C4-Pflanzen (z.B. Mais, Hirse, Zuckerrohr, Savannengräser)
kommen vor allem in warmen, trockenen Regionen vor und machen nur ca. 10-20% der
Vegetation aus (z.B. Amrose/DeNiro 1986; Ambrose 1993; Hoefs 2009, 178; Stephan 2008,
58-60). Da die früheste C4-Pflanze in Mitteleuropa - die Hirse - erst ab der Bronzezeit
angebaut wurde (Schibler et al. 1997, 234), ist diese Möglichkeit der
Ernährungsrekonstruktion für neolithische Hirsche aber nicht von Bedeutung.
Feuchtigkeit: Die Fraktionierung von Kohlenstoff innerhalb der Pflanzen hängt stark mit der
Aktivität der Stomata zusammen. Bei zunehmender Trockenheit schliessen sich die Stomata,
es wird weniger CO2 aufgenommen. Dies führt zu einer Anreicherung von 13
C im Blatt, da der
leichte Kohlenstoff (12
C) bevorzugt in der Photosynthese verarbeitet wird. Daraus resultieren
bei trockenen Verhältnissen höhere δ13
C-Werte als bei grosser Wasserverfügbarkeit (v.a.
Luftfeuchtigkeit!). Es besteht also eine negative Korrelation zwischen den δ13
C-Werten und
der Feuchtigkeit (Übersicht Heaton 1999; Drucker et al. 2011, 269.).
Höheneffekt: Die Leitfähigkeit der Stomata, die Effizienz der Wassernutzung sowie die
Intensität der Photosynthese können durch verschiedene Faktoren wie Lichtintensität,
Temperatur, Salzgehalt des Bodens sowie CO2-Druck beeinflusst werden (Drucker et al. 2011
nach Tieszen 1991, Heaton 1999). Diese Faktoren können durch die Höhenlage beeinflusst
werden. So steigen die δ13
C-Werte in der Höhe mit grösserer Wassernutzungseffizienz bei
sinkenden Temperaturen und CO2-Druck (Drucker et al. 2011 z.B. nach Körner et al. 1991).

20
Grosse Variabilität in aquatischen Systemen: Da aquatische Lebewesen auch Kohlenstoff
aus dem Kohlendioxid und dem Bicarbonat des Wassers sowie Carbonate aus Gesteinen und
Böden aufnehmen, ist die Variabilität der δ13
C-Werte viel höher als bei terrestrischen
Organismen. Aufgrund der Anreicherung von 13
C im Bicarbonat des Meerwassers um ca. 7‰,
weisen marine Organismen höhere δ13
C-Werte auf als terrestrische Lebewesen.
Für die Interpretation der δ13
C-Werte der neolithischen Hirsche aus Zug-Riedmatt sind vor allem der
Canopy-Effekt sowie die Faktoren Feuchtigkeit und Höhe wichtig. Es wäre zu erwarten, dass Tiere
aus tieferen/bewaldeten Lagen niedrigere Kohlenstoffwerte aufweisen als solche aus höher
gelegenen, offenen Landschaften (Drucker et al 2011, 269).
2.4.4.2 δ13
C in Karbonat (aus Apatit) und Kollagen
Die Kohlenstoffisotopenverhältnisse werden üblicherweise zusammen mit dem Sauerstoff im Karbonat
sowie mit dem Stickstoff im Kollagen gemessen. Dabei fällt auf, dass beim gleichen Individuum
unterschiedliche δ13
C-Werte aus diesen beiden Quellen resultieren. Dies liegt an der z. T.
unterschiedlichen Herkunft des Kohlenstoffs, der in das jeweilige biogene Gewebe eingebaut wird.
Während die δ13
C-Werte im Karbonat des Apatits (im vorliegenden Fall Zahnschmelz) die
Kohlenstoffwerte der gesamten Nahrung reflektieren, stammen die δ13
C-Werte im Kollagen vor allem
von den Proteinen der zuletzt aufgenommenen Nahrung. Zusammen mit der unterschiedlich starken
Fraktionierung während des Einbaus des Kohlenstoffs entstehen Abweichungen zwischen den Werten
aus Kollagen und Apatit von durchschnittlich 4-7‰. Diese sind von der Nahrungszusammensetzung
(Trophiestufe), der Verstoffwechselung sowie der Knochen- und Zahnbildung einer Tierart abhängig
(Lee-Thorp et al. 1989; Ambrose/Norr 1993; Hedges 2003). Das heisst, die beobachteten
Unterschiede zwischen den δ13
C-Werte aus Kollagen und Apatit variieren zwischen verschiedenen
Tierarten (siehe Zusammenstellung Stephan 2008, Abb. 1.24) und ermöglichen eine Unterscheidung
von herbivorer, omnivorer und carnivorer Ernährung (Lee-Thorp 1989; Hedges 2003).
Bei Hirschen, für die eine rein herbivore Ernährung angenommen werden kann, sind δ13
C-Differenzen
aus Kollagen und Apatit von 7±1‰ zu erwarten (Lee-Thorp 1989). Da die Fraktionierung im Apatit des
Zahnschmelzes mit bis zu 14‰ stärker ist als im Knochenapatit (ca. 9.4‰), könnte diese Differenz
aber auch etwas höher ausfallen (z.B. Ambrose/Norr 1993; Cerling/Harris 1999).
2.4.4.3 Anwendung von Stickstoffisotopen (δ15
N) zur Nahrungs- und Umweltrekonstruktion
Stickstoff gelangt bei der Fixierung durch Mikroorganismen im Boden in den terrestrischen Kreislauf.
Die Pflanzen nehmen den im Boden verfügbaren Stickstoff auf und spiegeln die grosse Variation der
δ15
N-Verhältnisse wieder, wobei es immer zu einer Anreicherung mit dem schwereren Isotop kommt.
Diese Verhältnisse werden wiederum mit einer durchschnittlichen Anreicherung von 3-5‰ an ihre
Konsumenten (Herbivoren) weitergegeben. Daraus folgt eine stetige Anreicherung von 15
N mit
aufsteigender Trophie-Stufe (Schöninger/DeNiro 1984; Ambrose 1993; Ambrose 2000).
Da bei Hirschen von einer rein herbivoren Ernährung ausgegangen werden kann, müssen für allfällige
Variationen der δ15
N-Werte ausschliesslich die Unterschiede zwischen den gefressenen Pflanzen

21
verantwortlich sein. In verschiedensten Untersuchungen konnten folgende Faktoren für die Variabilität
der δ15
N-Werte in Pflanzen festgestellt werden:
Trockenheit: In vielen Arbeiten wurde Trockenheit als Faktor für erhöhte δ15
N-Werte
angegeben (z.B. Ambrose/DeNiro 1986; Bocherens et al. 2000). DRUCKER ET AL. (2003)
geben aber zu bedenken, dass sehr grosse Variationen in den Stickstoffisotopenwerten nicht
alleine auf die unterschiedlichen Niederschlagsmengen zurückzuführen sind (Drucker et al.
2003, 382). Für ein gemässigtes Klima wie beispielsweise im nördlichen Jura ist sogar der
umgekehrte Trend beobachtet worden – dass trockene und kalte Gebiete wie z.B.
Bergkuppen durch geringen Stickstoffkreislauf niedrigere δ15
N-Werte produzieren (Drucker et
al. 2003, 383). In der Studie von STEVENS ET AL. (2006) an rezenten Hirschen ist der
Zusammenhang zwischen Niederschlag und δ15
N-Werten nicht beobachtet worden, was damit
begründet wird, dass der Effekt nur in Regionen mit unter 400 mm Niederschlag/Jahr auftritt –
z.B. bei Savannen- oder Wüstenböden (Stevens et al. 2006, 17; Stephan 2008, 60).
Stickstoffkreislauf: Bei erhöhtem Stickstoffkreislauf steigen die δ15
N-Werte in den Pflanzen
an, da mehr vom leichten 14
N von den Pflanzen und Mikroorganismen absorbiert wird oder
verdunstet. Diese Faktoren können zu einem erhöhten Stickstoffkreislauf führen:
- Düngung/Feuer: Die Bearbeitung von Boden mittels Düngung bzw. Feuerlegung führt zu
einem gesteigerten Stickstoffkreislauf und somit zu erhöhten δ15
N-Werten
(zusammenfassend: Drucker et al. 2003, 382-383),
- Temperatur: Die Resultate verschiedenster Untersuchungen lassen vermuten, dass die
Konzentration von schweren Stickstoffisotopen mit der durchschnittlichen
Jahrestemperatur zusammenhängt (Drucker et al. 2003; Stevens et al. 2006; Drucker et
al. 2009; Drucker et al. 2011). Höhere Temperaturen begünstigen den Abbau von
organischem Material, was zu einer natürlichen Düngung des Bodens führt (Drucker et al.
2011, 273) und die δ15
N-Werte ansteigen lässt.
- Höhenunterschied: Ähnliche Gründe wie bei der Temperaturabhängigkeit sind für die
Variation von δ15
N-Werten im Boden und somit in Pflanzen/Herbivoren in verschiedenen
Höhenlagen verantwortlich. So wurden z.B. bei Hirschen aus dem frühen Holozän in den
Alpen deutlich niedrigere Stickstoffwerte gemessen als bei den Hirschen aus dem Jura
zur gleichen Zeit (Drucker et al. 2011). Bei den rezenten Hirschen aus den
Untersuchungen von STEVENS ET AL. (2006) konnte dieser Zusammenhang aber nicht
bestätigt werden (Stevens et al. 2006, 17).
Zusammenfassend kann angenommen werden, dass bei optimalen Wachstumsbedingungen
für die Vegetation höhere δ15
N-Werte resultieren. Limitierende Faktoren wie niedrige
Temperaturen, Trockenheit oder Nährstoffknappheit bremsen den Stickstoffkreislauf und
führen zu niedrigen δ15
N-Werten.
Durch die Anreicherung von 15
N von einer Trophie-Stufe zur nächsten, weisen auch Jungtiere
während ihrer Stillphase durch die Aufnahme von Muttermilch höhere δ15
N-Werte auf als erwachsene

22
Abb. 6
Beispiel von drei Beprobungsstellen (Sequenzen),
die verschiedene Zahnschmelzinkremente
anschneiden (Knipper 2011, 135, Abb. 8.5)
Abb. 7
Hirschzahn aus Zug-Riedmatt mit sieben
Beprobungsstellen (Sequenzen).
Individuen. Dieses Still-Signal ist vor allem im Dentin des 1. Molaren nachzuweisen bzw. in dem
Gewebe, das während der Säuglingsphase gebildet wurde (z.B. Balasse 2001; Balasse/Tresset
2002). Aus diesem Grund sollten Proben zur Nahrungs-, Lebensraum- und Klimarekonstruktion mittels
Stickstoffisotopenverhältnissen immer aus den 2. oder 3. Molaren stammen, um einen Einfluss des
Stillsignals ausschliessen zu können.
2.5 Die sequenzielle Beprobung von Zahnschmelz und Dentin
2.5.1 Zahnschmelz
Die Veränderung der Sauerstoff-Isotopenverhältnisse im Zahnschmelz von hochkronigen Tierzähnen
wurde erstmals 1996 von FRICKE und O’NEIL (Fricke/O’Neil 1996) mit Hilfe der sequenziellen oder
seriellen Beprobungstechnik untersucht und seither in zahlreichen verschiedenen Forschungsarbeiten
angewandt und weiterentwickelt (z.B. Wiedemann et al. 1999; Balasse 2002; Balasse 2003;
Higgins/MacFadden 2004; Zazzo et al. 2005; Knipper et al. 2006; Stevens et al. 2011; Balasse et al.
2012; Frémondeau et al. 2012; Knipper 2011; Weitere Literatur aufgelistet von Balasse 2003, 155-156
sowie Knipper 2011, 135). Die Grundlage dieser Methode ist die in Kapitel 2.3.3 beschriebene Art der
Zahnschmelzbildung. Der Zahnschmelz bildet sich von der Zahnspitze bis zum Zahnhals in einer
bestimmten Zeit aus, was zu verschiedenen Schmelzinkrementen führt, die sich schuppenartig
überlappen. Die isotopische Zusammensetzung des Zahnschmelzes an einer bestimmten Stelle
spiegelt die Zeit wieder, zu welcher die Mineralisation stattgefunden hat. Durch die sequentielle
Beprobung entlang der Zahnkrone können so Veränderungen der Isotopenwerte festgestellt werden,
die sich während der Zeitspanne der Zahnschmelzmineralisation ergeben haben. Diese können
aufgrund von veränderter Nahrung oder Aufenthaltsort sowie Saisonalität entstehen (s. Kapitel 2.4).
Es ist jedoch zu beachten, dass bei einer Probe aus einer bestimmten Stelle bzw. Höhe des
Zahnschmelzes immer ein Mischsignal aus einem gewissen Zeitraum gemessen wird. Einerseits
findet die Mineralisation einer Schmelzschicht in mehreren Schritten statt, das heisst der äussere

23
Tab. 1
Verringerung der Kronhöhen mit
zunehmendem Alter, ersichtlich an
Hirschzähnen aus Zug-Riedmatt.
Zahnschmelz und derjenige an der Schmelz-Dentin-Grenze werden nicht zur gleichen Zeit gebildet.
Man unterscheidet zwischen der schwach mineralisierten Matrix zu Beginn der Reifung und der
darauffolgenden sogenannten sekundären Mineralisation, in der sich 80-90% der anorganischen
Komponenten des Zahnschmelzes ausbilden. Dieser Schmelzreifungsprozess breitet sich
kontinuierlich in verschiedene Richtungen über der ursprünglichen Matrix aus (Balasse 2002; Balasse
2003; Zazzo et al. 2005; Knipper 2011). Andererseits bilden sich die Wachstumsinkremente über dem
Dentin nicht horizontal, sondern in einem Winkel von ca. 45 Grad aus. Das heisst, dass bei einer
horizontalen Beprobung immer mehrere Inkremente durchschnitten werden, was wiederum zu einer
Mischung von Isotopenverhältnissen aus verschiedenen Bildungszeiträumen führt (Abb. 6 und Abb.
7). Eine Untersuchung von BALASSE (2003) zeigt aber, dass die Beprobungsrichtung am
Zahnschmelz keinen Einfluss auf die resultierenden Isotopenwerte hat (Balasse 2003).
Trotz dieser Einschränkungen zeigen serielle Beprobungen aus den verschiedensten Untersuchungen
einen graduellen Verlauf der Isotopenwerte entlang der Zahnkrone, welche die Veränderung der
Isotopenverhältnisse in der aufgenommenen Nahrung und im Trinkwasser wiederspiegeln. Die
Isotopenwerte aus einem Zahnschmelzquerschnitt von ca. 1mm sind aber als Mittelwerte aus einem
gewissen Zeitraum zu verstehen, in der die verschiedenen Schmelzschichten mineralisiert wurden.
Dies kann eventuell zu einer Dämpfung der Isotopenkurve führen, deren Ausmass wesentlich durch
die Dauer der Zahnschmelzmineralisation sowie der Probengrösse beeinflusst wird (z.B. Balasse
2002; Balasse 2003; Higgins/MacFadden 2004; Zazzo et al. 2005; Knipper 2011). Der effektive
Zeitraum, der in einer durchschnittlichen Probe von ca. 1-2mm Länge, 2-5mm Breite und 1mm Tiefe
(durch die gesamte Dicke des Zahnschmelzes) repräsentiert ist, wird unterschiedlich geschätzt. Für
Pferde werden 3-4 Monate (Hoppe et al. 2004) bzw. 1-1,5 Monate (Balasse 2002; Kohn et al. 2002)
angegeben und für Rinder bzw. Bisons ca. 2 Monate (Higgins/MacFadden 2004). Für Hirsche liegen
keine solche Schätzungen vor, STEVENS ET AL. (2011) sprechen von einem unbestimmten
Zeitraum, den eine Sequenz an einem Hirschzahn abdeckt (Stevens et al. 2011, 65).
Ohne Zweifel ist es sinnvoll, möglichst kleine bzw. viele Proben von einem Zahn zu entnehmen, um
die Auflösung des Ergebnisses zu erhöhen. An hochkronigen Zähnen wie bei denen vom Rind oder
Schaf sind bei aufwendiger Probenentnahme 20 bis über 30 Sequenzen an einer einzigen Zahnkrone
möglich (Balasse et al. 2012a; Balasse et al. 2012b), wobei entscheidend ist, wie viel Material an der
Kaufläche durch natürliche Abrasion schon verlorengegangen ist (Stevens et al. 2011). Das heisst, je
jünger ein Individuum und je höher die Zahnkrone, umso mehr Sequenzen pro Zahn sind möglich.
Hirsche weisen eine wesentlich kürzere Zahnkrone auf als Rinder, Pferde oder Schafe. Nach dem
Durchbruch sind die Zähne einer starken Abrasion ausgesetzt, ohne dass sie sich weiter aus dem
Kiefer schieben könnten. Das heisst, die Höhe der Zahnkrone verringert sich mit jedem Lebensjahr um
ca. 1mm, wie aus dem vorliegenden Material aus Zug-Riedmatt sichtbar wurde:
Nr. Alter Kronhöhe M3 [mm]
1067.1.3 Ca. 1 Jahr 24.2
1348.2.3 4-5 Jahre 18.6
1535.1.3 Ca. 10 Jahre 13.9

24
Bei älteren Tieren muss also beachtet werden, dass durch die Abrasion bereits bis zu 10mm
Zahnschmelz nicht mehr vorhanden sind und somit auch die Anfangszeit der Schmelzbildung nicht
mehr in den Isotopenverhältnissen repräsentiert wird. Die Einschätzung des Kurvenverlaufs ist auch
mit nur vier bis fünf Datenpunkten möglich, bei der Interpretation muss aber die verkürzte
repräsentierte Zeitspanne berücksichtigt werden
2.5.2 Dentin
Dentin aus Tier- und Menschenzähnen diente neben Knochen schon in einigen Untersuchungen als
Quelle für Ernährungs- und Umweltrekonstruktionen (z.B. Bocherens et al. 1994; Koch et al. 1995;
Balasse et al. 1999; Wright/Schwarcz 1999). Die sequenzierte Beprobung von Dentin ist aber erst in
wenigen Studien angewandt worden. Als Vorstufe davon könnte man die Studien von KOCH ET AL.
(1989) und HOBSON/SEASE (1998) betrachten, die Wachstums-Inkremente von Stosszähnen bzw.
Zähnen von Meeressäuger untersuchten und Veränderungen in Umwelt- und Ernährungs-
bedingungen der Tiere aufzeigen konnten. Diese Methode ist aber bei den meisten Landsäugetieren
nicht anwendbar, da deren Zähne keine so klaren Wachstumsstrukturen des Dentins aufweisen oder
schlichtweg zu klein sind (Balasse et al. 2001, 236). BALASSE ET AL. (2001) testeten eine andere
Form der Sequenzierung, welche auf die meisten Zähne angewendet werden kann. Anstatt in der
Dicke wird das Dentin der Längsachse des Zahns nach beprobt (Balasse et al. 2001, 236). Sie
konnten an den Zähnen von rezenten Rindern nachweisen, dass sich sowohl der Zeitpunkt der
Entwöhnung als auch die Ernährungsumstellung von C3 auf C4-Pflanzen in den Stickstoff- und
Kohlenstoffisotopen-verhältnissen des Dentins erkennen lassen. In einer darauffolgenden Studie
konnten diese Erkenntnisse auf das archäologische Material von Bercy (F) angewendet werden
(Balasse/Tresset 2002).
Wie in Kapitel 2.3.3 beschrieben, unterliegt das Dentin - anders als Knochen - nach der Bildung
keinen Umbau- und Erneuerungsprozessen und speichert somit wie der Zahnschmelz die
Isotopenverhältnisse vom Zeitpunkt der Entstehung. Dies gilt aber lediglich für das Primärdentin.
Sekundärdentin wird erst im darauffolgenden Zeitraum gebildet und enthält somit auch abweichende
Isotopensignale. Weiter muss mit einer stärkeren diagenetischen Überprägung gerechnet werden als
beim Zahnschmelz, da die Struktur von Dentin wesentlich poröser ist. Trotzdem kann der Vergleich
der Isotopenverhältnissen aus Dentin und Knochen v.a. bei älteren Individuen einen Hinweis über
veränderte Lebensumstände zwischen Jugend- und Erwachsenenalter liefern. Denn während das
Dentin die mehr oder weniger deutliche Signale aus der Zeit der Bildung des Gewebes enthält, wird
das Kollagen des Knochens ständig erneuert und die isotopische Zusammensetzung so an die
aktuellen Lebensumstände angepasst (Balasse et al. 2001).
Bei Isotopenanalysen zur Rekonstruktion von Lebensraum und Ernährung sollte Dentin aus den 3.
Molaren beprobt werden. Die 1. und 2. Molaren werden in jenem Zeitraum gebildet, in dem die
Jungtiere noch gesäugt werden. Das hat zur Folge, dass die Stickstoffisotopenzusammensetzung
noch vom sogenannten „Stilleffekt“ (s. Kapitel 2.4.2.3) beeinflusst wird.

25
Das Wachstum des Dentins dauert länger als das des Zahnschmelzes, da es nicht nur die Zahnkrone,
sondern auch die Zahnwurzel bildet. Die Zeitspanne, die mit der Beprobung des Dentins abgedeckt
wird, beinhaltet die Anfangsphase der Zahnbildung bis zur Vervollständigung der Wurzel. Die dauert
beim M3 des Rothirsches ca. vom 9. bis zum 40. Lebensmonat, also über 2 Jahre (Brown 1991, 91-
94).
3. Material
3.1 Auswahl des Probenmaterials
Mittels der Isotopenanalysen an den Hirschzähnen könnte es möglich sein, Unterschiede zwischen
den Siedlungsphasen in Bezug auf Jagdtätigkeit und Herkunft der Hirsche zu beobachten. Da es in
den Phasen D und E aber weniger Hirschknochen und kaum vollständige Zähne gibt, wurde der
Grossteil der Zähne aus den Phasen B (n=3), B1 (n=8), B2 (n=7) und C1 (n=4) entnommen. Das
heisst die meisten Zähne stammen aus dem Knochenhaufen und den darüber liegenden Lehmlagen.
Aus der Schicht D (n=1) konnten lediglich ein 1. Molar aus dem Oberkiefer eines jungen Hirsches
beprobt werden, da keine Unterkiefer von Hirschen vorhanden waren. Insgesamt standen also 23
Zähne von 22 Individuen für die Isotopenanalysen zur Verfügung (Tab. 2).
Bei der Auswahl des zu beprobenden Materials musste auf verschieden Faktoren geachtet werden.
Für das Auswahlverfahren wurden nur die mehr oder weniger vollständig erhalten Unterkiefer
ausgelegt. Da die Altersbestimmung schon gemacht war und die Kieferhälften nach rechts und links
getrennt wurden, konnte die doppelte Beprobung eines Individuums ausgeschlossen werden.
Die meisten Unterkiefer stammten aus der Siedlungsphase B, vor allem aus dem Knochenhaufen (B1)
und der Lehmschicht (B2). Diejenigen Stücke, die keinem der beiden Strukturen zugeordnet werden
konnten, wurden zur allgemeinen Schicht B gezählt. Auch in der Schicht C waren genug Unterkiefer
vorhanden, so dass eine Auswahl getroffen werden konnte.
Da vor allem die 3. Molaren beprobt werden sollten, war deren Vorhandensein im Kiefer das
wichtigste Kriterium. Bei den Probeläufen in Mainz wurde darüber hinaus festgestellt, wie wichtig die
gute Erhaltung der Zähne für eine genaue und vollständige Beprobung ist. Risse und Absplitterungen
im Zahnschmelz sind ein Anzeichen dafür, dass der Zahn während der Probenentnahme weiter
zerfallen und ein sauberes Arbeiten dadurch erschweren würde.
Weiter wurde darauf geachtet, in jedem Schichtkomplex Hirsche aus verschiedenen Altersklassen zu
beproben. Die Altersbestimmung wurde von Sandra Billerbeck im Rahmen der archäozoologischen
Auswertung nach WAGENKNECHT (2000) und RAESFELD (1988) durchgeführt. Aus den Schichten
B und C konnten Individuen im Alter zwischen 19-27 Monaten und 9-10 Jahre ausgelesen werden. So
können Altersgruppen gebildet werden und die Isotopenwerte untereinander auf Unterschiede hin
verglichen werden. Zudem variiert die Kronhöhe der M3 durch die unterschiedliche Abrasion.

26
Tab. 2
Auflistung der 23 beprobten Zähne aus Zug-Riedmatt. Der Abkauungsgrad steht in direktem Zusammenhang
mit der Höhe des Zahns sowie der Anzahl der entnommenen Sequenzen aus Zahnschmelz und Dentin. Die
Höhe des Dentins [mm] beinhaltet die Wurzellänge und die Krondentinhöhe.
Siedlungs- phase
Nr. Alter Zahn Abrasion Höhe Krone [mm]
Anz. Sequenzen
Zahnschmelz
Höhe Dentin [mm]
Anz. Sequenzen
Dentin
B 799.1 10 J M2 sehr stark 10.8 4 27.8 5
B 1890.1 19-27 M M3 sehr leicht 22.8 8 27.5 4
B 1901.1 2 J M3 leicht 23.5 8 22.0 5
B1 1763.1 19-27 M M3 keine; kaum durchgebrochen 25.2 6 17.1 3
B1 1763.2 8-9 J M3 sehr stark 11.2 4 29.6 5
B1 1776.1 4-5 J M3 deutlich 19.3 6 28.1 5
B1 1846.1 2-3 J M2 leicht 22.0 6 30.2 6
2-3 J M3 sehr leicht 24.0 9 29.2 6
B1 1846.2 2-3 J M3 leicht 23.0 8 23.2 5
B1 1846.3 2 J M3 leicht-deutlich 23.6 7 25.8 5
B1 1846.4 3 J M3 stark 18.7 6 29.0 5
B2 1342.1 4-5 J M3 leicht 18.0 6 26.9 6
B2 1342.2 3 J M3 stark 20.0 6 32.6 5
B2 1348.1 4-5 J M3 deutlich 19.3 6 28.8 6
B2 1348.2 4-5 J M3 deutlich 18.6 6 26.4 5
B2 1348.3 4-5 J M3 stark 16.1 6 30.1 5
B2 1464.1 19-27 M M3 nicht- sehr schwach 23.5 7 23.1 4
B2 1535.1 bis 10 J M3 sehr stark 13.9 5 25.6 5
C1 952.1 19-27 M M3 nicht-sehr schwach 23.9 7 24.4 4
C1 1067.1 22-25 M M3 leicht 24.2 5 31.2 6
C1 1238.1 6-7 J M3 sehr stark 17.9 6 29.5 5
C1 1244.1 4-5 J M3 leicht 17.0 5 32.7 5
D1/D2 710.1 11-12 M M1 OK leicht 18.5 6 18.1 3
3.2 Wasserproben
Im Rahmen der nationalen Grundwasserbeobachtung wurde 1992 ein Modul zur Beobachtung der
Isotope im Wasserkreislauf gegründet (ISOT). Dieses umfasst 23 Messstationen in den verschiedenen
Klimaregionen der Schweiz, wo die Anteile an 18
O, Deuterium und Tritium im Wasser gemessen
werden. Die Messdaten dieser Stationen werden an die International Atomic Energy Agency (IAEA)
weitergeleitet und sind dort für die Öffentlichkeit zugänglich bzw. können für die Forschung genutzt
werden (Schotterer et al. 2000, Schotterer 2010; http://www.univie.ac.at/cartography/project/wiser/).
Leider liegt keine dieser Messstationen im Gebiet um Zug, was bedeutet, dass keine genauen
Isotopendaten für die Gewässer im möglichen Einzugsgebiet der neolithischen Hirschpopulation
vorhanden sind. Aus diesem Grund wurden für diese Untersuchungen eigene Wasserproben
genommen, um einen ungefähren Eindruck zu bekommen, in welcher Grössenordnung sich die δ18
O-
Daten bewegen. Zudem kann der Temperatur- und Höheneffekt an konkreten Daten nachvollzogen
werden.
Am Tag der Beprobung (Anfang Oktober 2012) regnete es stark und ausdauernd, was einen Einfluss
auf die Werte aus den Seen haben kann. Aufgrund des Höheneffektes kann deshalb mit
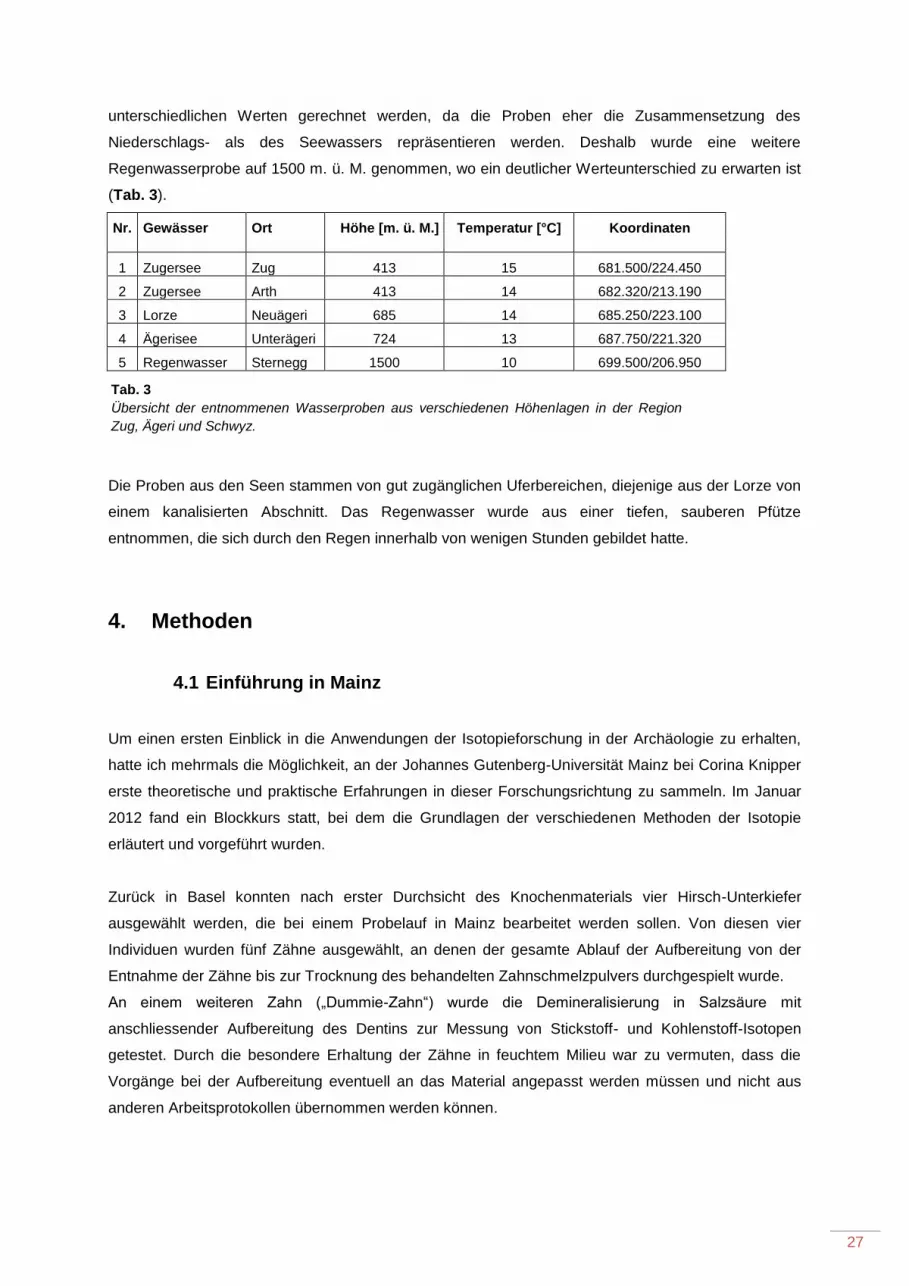
27
Tab. 3
Übersicht der entnommenen Wasserproben aus verschiedenen Höhenlagen in der Region
Zug, Ägeri und Schwyz.
unterschiedlichen Werten gerechnet werden, da die Proben eher die Zusammensetzung des
Niederschlags- als des Seewassers repräsentieren werden. Deshalb wurde eine weitere
Regenwasserprobe auf 1500 m. ü. M. genommen, wo ein deutlicher Werteunterschied zu erwarten ist
(Tab. 3).
Nr. Gewässer Ort Höhe [m. ü. M.] Temperatur [°C] Koordinaten
1 Zugersee Zug 413 15 681.500/224.450
2 Zugersee Arth 413 14 682.320/213.190
3 Lorze Neuägeri 685 14 685.250/223.100
4 Ägerisee Unterägeri 724 13 687.750/221.320
5 Regenwasser Sternegg 1500 10 699.500/206.950
Die Proben aus den Seen stammen von gut zugänglichen Uferbereichen, diejenige aus der Lorze von
einem kanalisierten Abschnitt. Das Regenwasser wurde aus einer tiefen, sauberen Pfütze
entnommen, die sich durch den Regen innerhalb von wenigen Stunden gebildet hatte.
4. Methoden
4.1 Einführung in Mainz
Um einen ersten Einblick in die Anwendungen der Isotopieforschung in der Archäologie zu erhalten,
hatte ich mehrmals die Möglichkeit, an der Johannes Gutenberg-Universität Mainz bei Corina Knipper
erste theoretische und praktische Erfahrungen in dieser Forschungsrichtung zu sammeln. Im Januar
2012 fand ein Blockkurs statt, bei dem die Grundlagen der verschiedenen Methoden der Isotopie
erläutert und vorgeführt wurden.
Zurück in Basel konnten nach erster Durchsicht des Knochenmaterials vier Hirsch-Unterkiefer
ausgewählt werden, die bei einem Probelauf in Mainz bearbeitet werden sollen. Von diesen vier
Individuen wurden fünf Zähne ausgewählt, an denen der gesamte Ablauf der Aufbereitung von der
Entnahme der Zähne bis zur Trocknung des behandelten Zahnschmelzpulvers durchgespielt wurde.
An einem weiteren Zahn („Dummie-Zahn“) wurde die Demineralisierung in Salzsäure mit
anschliessender Aufbereitung des Dentins zur Messung von Stickstoff- und Kohlenstoff-Isotopen
getestet. Durch die besondere Erhaltung der Zähne in feuchtem Milieu war zu vermuten, dass die
Vorgänge bei der Aufbereitung eventuell an das Material angepasst werden müssen und nicht aus
anderen Arbeitsprotokollen übernommen werden können.
28
Tab. 4
In Mainz
analysierte
Hirschzähne aus
Zug-Riedmatt.
Die Bearbeitung dieser Zähne wurde in Mainz durchgeführt:
Probe Nr. Zahn Alter Abrasion Siedlungsphase
RIZG1846.1.2 1846.1 M2 2-3 J leicht B1
RIZG1846.1.3
M3 2-3 J sehr leicht B1
RIZG1846.2.3 1846.2 M3 2-3 J leicht B1
RIZG1342.1.3 1342.1 M3 4-5 J leicht B2
RIZG1244.1.3 1244.1 M3 4-5 J leicht C1
Die Benennung der Proben wurde folgendermassen festgelegt:
RIZG 1846. 1. 3
Fundstelle (Riedmatt Zug) Fundkomplex Zahn aus Fundkomplex M3 oder M2
Diese Identifikation soll einheitlich für alle Proben aus dem Zahnschmelz gelten. Da jeder Zahn
sequenziell beprobt wird, folgt hinter der Probennummer noch die Nummerierung der Sequenz. Für
die Zahnschmelzproben ergibt sich also folgende definitive Bezeichnung einer einzelnen gemessen
Probe: RIZG 1846.1.3.1-9. Bei den Sequenzen aus dem Dentin hat die Probennummer denselben
Aufbau, zur Unterscheidung von den Zahnschmelzproben wird aber noch ein „d“ hinten angefügt.
Während der Bearbeitung dieser fünf Zähne in Mainz wurde ein Arbeits- und Laborprotokoll erstellt,
welches bei der weiteren Beprobung in Basel genau eingehalten wurde. Abweichungen betreffen
höchstens einzelne Geräte oder Laborutensilien, da diese am IPNA teilweise extra für diese
Forschungsarbeit neu angeschafft werden mussten. Falls diese Unterschiede einen Einfluss auf den
Ablauf der Aufbereitung der Proben hatten, wird dies in der folgenden Beschreibung der Methodik
erwähnt.
Für die Aufnahme und Verarbeitung aller Daten wurde von Corina Knipper eine Access-Datenbank zur
Verfügung gestellt, die für dieses Projekt beliebig verändert und ausgebaut werden konnte.
4.2 Die Analyse von Sauerstoff- und Kohlenstoffisotopen (δ18O/δ13C)
aus dem Zahnschmelz
4.2.1 Die Vorbereitung der Zähne für die Beprobung
Von jedem Individuum wurden der 2. und der 3. Molar aus dem Kieferknochen entnommen, sofern
beide vorhanden waren. Bei einem Individuum (RIZG799.1) war lediglich der M2 vorhanden. Da in
dieser Arbeit aber primär die möglichst hohe Anzahl an beprobten Hirschen im Vordergrund steht,
konnte pro Tier nur ein Zahn bearbeitet werden. Die übrigen Zähne, vorwiegend also M2, könnten
aber für zukünftige Analysen verwendet werden. Weiter musste auch damit gerechnet werden, dass
während der Beprobung oder Aufbereitung Material verloren gehen oder zerstört werden könnte. So
hätte man mit den restlichen Zähnen weiterarbeiten können und nicht auf Messergebnisse von
einzelnen Tieren verzichten müssen.

29
Nachdem die Kiefer mitsamt Zähnen fotografiert waren, konnten die einzelnen Zähne mit Hilfe einer
Trennscheibe (Bohrer EV250-230 Emax EVOlution) aus dem Knochen entnommen werden. Durch die
gute Erhaltung des Materials war es grösstenteils nicht möglich, die Zähne ohne teilweise Zerstörung
des Kieferknochens zu entfernen. Die Knochenteile, die durch die Entnahme der Zähne vom Kiefer
gelöst werden mussten, wurden beschriftet und aufgehoben. In weiteren Untersuchungen könnte so
noch das Isotopenverhältnis von Stickstoff und Kohlenstoff aus dem Knochenkollagen des
zugehörigen Unterkiefers gemessen werden.
Die entnommenen Zähne mussten gründlich gereinigt werden, da sich in offenen Wurzeln und
zwischen den Schmelzfalten viel Sediment und organisches Material abgelagert hatte. Dies würde
sich bei der folgenden Entnahme der Zahnschmelzproben lösen und die Proben verunreinigen.
Zur Reinigung wurden die Zähne ins Ultraschallbad gelegt und wo nötig anschliessend manuell vom
restlichem Sediment befreit. Nach der Trocknung im Trockenschrank wurde von jedem Zahn die Säule
ausgewählt, die am besten erhalten war bzw. keine oder möglichst wenig Haarrisse aufwies. Da sich
zwischen den einzelnen Säulen immer noch kleinste Mengen an störendem Sediment befinden
konnten, wurde die ausgewählte Säule mit der Trennscheibe abgetrennt und nochmals gereinigt.
Anschliessend wurde die Schmelzoberfläche der Zahnsäule mit dem Dremel sehr gründlich gereinigt,
bis keine dunklen Verfärbungen mehr sichtbar waren. Dabei muss sehr vorsichtig vorgegangen
werden, denn durch die Vibrationen des Dremels können weitere Haarrisse im Zahnschmelz
entstehen und zu dessen Absplitterung führen. Bei Zähnen die schon Risse aufweisen, kann die
Oberflächenreinigung auch mit dem Schleifkopf 1,8mm erfolgen. Dies dauert länger, es entstehen
aber weniger starke Vibrationen als mit dem Dremel.
4.2.2 Sequenzierte Entnahme der Zahnschmelzproben
Die Sequenzen wurden in möglichst regelmässigen Abständen von ca. 3-4mm entnommen, was aber
aufgrund der Erhaltung und unterschiedlicher Schmelzdicke nicht immer eingehalten werden konnte.
So ergaben sich bei den Zähnen, bei denen die Schmelzsäule abbrach und die Proben mit der
Trennscheibe gesägt werden mussten, tendenziell weniger Sequenzen, da der Materialverlust relativ
gross war. Die Anzahl der Sequenzen bzw. deren Abstand musste so gewählt werden, dass sich pro
Probe 15-20mg Zahnschmelzpulver ergab. Dies führte zu einer sehr unterschiedlichen Anzahl von
Sequenzen pro Zahn – es wurden zwischen vier und neun Proben entnommen. Durch eine feinere
Beprobung mit kleineren Probenmengen wäre es möglich gewesen, eine höhere Auflösung der
Ergebnisse zu erzielen. Dies hätte aber zu einer wesentlich grösseren Gesamtzahl der Proben geführt
und somit zu einem grösseren zeitlichen und finanziellen Aufwand, was im Rahmen dieser Arbeit nicht
sinnvoll gewesen wäre. Im vorliegenden Zusammenhang war es wichtiger, die Isotopenverhältnisse
möglichst vieler Zähne bzw. Individuen messen zu können.
Das Verfahren der Sequenzierung hing davon ab, ob der Zahnschmelz keine Risse aufwies und so
direkt vom Dentin abgefräst werden konnte - oder ob er sich der Länge nach vom Zahn ablöste.

30
Abb. 8
Sequenziell
beprobter
Hirschzahn (M3)
von Zug-Riedmatt.
Zahnschmelz bricht nicht
Mit dem Schleifkopf (1,8mm) wurden die Sequenzen aus dem Zahnschmelz
gebohrt. Diese verlaufen quer und sind ca. 2mm breit. Zwischen jeder
Sequenz blieb ein Steg von ca. 1-2mm stehen. Dies ergab pro Zahn zwischen
vier und acht Proben, je nach Höhe und Erhaltung der Zahnkrone. Um pro
Sequenz genügend Probenmaterial zu erhalten (15-20mg), wurde bis kurz vor
die Zahnschmelz-Dentin-Grenze gebohrt. Bei der untersten Probe an der
Schmelzgrenze war es trotzdem notwendig, eine etwas breitere Sequenz zu
bohren, da der Schmelz dort oft schon sehr dünn ist (Abb. 8).
Bei einigen Zähnen brach der Zahnschmelz während der Beprobung trotz aller
Vorsicht weg, was ein Bohren an dieser Stelle unmöglich machte. In diesem
Fall konnte auf eine andere Säule des Zahns ausgewichen werden, wobei der
Abstand von der Zahnschmelzgrenze natürlich genau abgemessen werden
musste.
Die Abstände der einzelnen Probesequenzen wurde notiert und das Pulver in
beschriftete 2ml-Eppendorftubes abgefüllt.
Zahnschmelz ist abgebrochen
Um abgesplitterten Zahnschmelz zur Probengewinnung zu verwenden, ist eine durchgehender Splitter
der Zahnsäule von 4-5mm Breite nötig. Dieser kann dann mit Hilfe der Trennscheibe in einzelne
Sequenzen von 2-4mm Höhe zerlegt werden.
Von den 23 Hirschzähnen wurde dieses Verfahren an insgesamt sieben Zähnen angewendet. Da die
Unterseite der Zahnschmelzsplitter noch deutlich braun verfärbt waren, musste diese mit dem
Schleifkopf gründlich gereinigt werden, bis nur noch weisser Zahnschmelz übrig war. In einigen Fällen
war dies extrem schwierig, weil die einzelnen Proben weiter zersplitterten und kaum mehr zu
handhaben waren. Bei Schwierigkeiten in der Reinigung wurde dies im Probenheft vermerkt und kann
allenfalls bei der Auswertung berücksichtigt werden.
Die Zahnschmelzstückchen wurden anschliessend einzeln in einem Achat-Mörser zu Pulver zerrieben,
was für die Aufreinigung des Karbonates nötig ist. Auch hier sollte das Gewicht jeder Probe zwischen
15 und 20mg liegen. Nach der Pulverisierung im Mörser konnte mit diesen Proben gleich verfahren
werden wie mit denjenigen, die gefräst wurden.
4.2.3 Aufreinigung des Zahnschmelzes
Die Aufbereitung des Karbonats erfolgte mit leichten Abweichungen der von BALASSE (2002)
entwickelten und von VOHBERGER (2011) modifizierten Methode. Weitere Änderungen für die
vorliegenden Untersuchungen beruhen auf mündlichen Mitteilungen von Corina Knipper.
Vor der Aufreinigung wurde von jeder Probe 10-11mg Material in ein frisches, beschriftetes 2ml-
Eppendorftube eingewogen. Soviel Pulver ist nötig, um nach der Reinigung noch genügend Material
für mehrere Messungen im Massenspektrometer übrig zu haben. Das restliche Pulver wird
unaufbereitet für allfällige weitere Messungen aufbewahrt.

31
Als erstes wurde jede Probe mit 1,8ml Natriumhypochlorid (NaOCl 2%) versetzt und über Nacht bei
Raumtemperatur gemischt. Dies führt zur Oxidation der organischen Fraktion im Zahnschmelz, welche
sich unter Gasbildung herauslöst.
Anschliessend wurden die Proben dreimal mit deionisiertem Wasser gespült, wobei das Material bei
jedem Durchgang auf dem Vortexer aufgeschüttelt und anschliessend wieder zentrifugiert wurde, um
das Wasser abpippetieren zu können. Im nächsten Schritt folgte eine Behandlung mit 0.1 molarer
Essigsäure, die mit einem Litiumacetate-Puffer versehen war. Nach dem Zufügen von 1,8ml
Essigsäure wurden die Proben 10 Minuten ins Ultraschallbad gestellt. Das Ziel der Zugabe leichter
Säure ist, sekundär absorbiertes Karbonat zu entfernen und nur das strukturelle Karbonat im Apatit zu
erhalten.
Es folgten erneut drei Spüldurchgänge mit Wasser und anschliessend konnten die Proben bei
offenem Deckel im Trockenschrank getrocknet und daraufhin ausgewogen werden. Die Ausbeute lag
je nach Einwaage und Sequenz zwischen sechs und neun Milligramm, was genügend Material für bis
zu fünf Messdurchgänge entspricht.
4.2.4 Massenspektrometrie
Die in Mainz aufbereiteten Proben der ersten fünf Zähne wurden schon im Mai 2012 am Institut für
Geowissenschaften der Universität Mainz gemessen. Diese Resultate dienten zur ersten Orientierung
und erlaubten einen Einblick in die Interpretationsmöglichkeiten.
Da genügend Probenmaterial vorhanden war, wurden dieselben Proben zusammen mit den
restlichen 18 Zähnen in Basel am Institut für Umweltgeowissenschaften vom Labor für stabile Isotope
(Leitung: Moritz Lehmann) erneut gemessen, um eine Kontrolle über die Vergleichbarkeit der beiden
Labors zu erhalten.
Basel
Am Institut für Umweltgeowissenschaften in Basel wurden die Karbonat-Messungen mit einem
Thermo Finnigan Gasbench II und einem Thermo Finnigan Delta V Advantage durchgeführt. Dafür
wurden zwischen 1.5 und 2.0 mg aufgereinigtes Zahnschmelzpulver auf einer Mettler Toledo
Präzisionswaage abgewogen und in Langhals-Rundbodengläschen (Exetainer) gegeben.
Als Standards wurden NBS 19 und UBC 1 verwendet, wovon je 4 kleine (40-60 µg) und vier grosse
(90-110 µg) Proben eingewogen wurden. Beide Standards sind auf PDB kalibriert. Die Messungen
erfolgten nach einem leicht modifizierten Protokoll nach VENNEMANN ET AL. (2002). Für einige
Proben erfolgten routinemässige Doppelmessungen, wobei davon der Durchschnittswert errechnet
wurde. Die Abweichungen der Kohlenstoffwerte waren dabei meist gering, diejenigen der
Sauerstoffwerte aber bis zu 0.95‰.

32
Mainz
Im Institut für Geowissenschaften in Mainz wurden die Proben mit einem Thermo Finnigan MAT 253
gekoppelt mit einer Gas Bench II gemessen. Dafür wurden 800 ± 50 µg Probenmaterial auf einer
Präzisionswaage eingewogen. Von dem Standard IVA Carrara wurden drei Proben an je 40, 65, 90
und 120 µg eingewogen und vom internationalen Standard NBS 19 zweimal 80-100 µg.
4.3 Die Analyse von Kohlenstoff- und Stickstoffisotopen (δ13C/δ15N) aus
dem Collagen im Dentin
4.3.1 Vorbereitung der Zähne für die Beprobung
Da die Zähne für die Beprobung bzw. Sequenzierung des Dentins demineralisiert werden müssen,
steht der Zahnschmelz danach nicht mehr für weitere Beprobungen zur Verfügung. Deshalb wurden
nur die Teile des Zahns für diese Analyse ausgewählt, die sich zuvor für die Sequenzierung des
Zahnschmelzes als ungeeignet erwiesen hatten. Das heisst meistens konnten zwei Säulen des M3
verwendet werden, in einzelnen Fällen (M2 oder sehr schlechte Erhaltung) nur eine Säule. Da die
Schmelzgrenze nach der Demineralisation eventuell nicht mehr klar erkennbar sein würde, wurden für
alle Zähne die Höhe der Krone und die Länge der Wurzel notiert.
Die Zahnteile wurden gereinigt, aber ansonsten unbehandelt in Bechergläser mit 0.5 molarer
Salzsäure (HCl) gegeben. Je nach Grösse und Erhaltung des Zahns dauerte es 4-6 Wochen, bis sich
alle Zahnschmelzreste aufgelöst hatten und das Dentin vollständig demineralisiert war. Die Salzsäure
wurde einmal pro Woche ausgetauscht und die Zähne immer wieder stundenweise bei
Raumtemperatur stehen gelassen, um die Reaktion zu beschleunigen. Ansonsten lagerten die Proben
im Kühlraum bei 4 Grad Celsius.
Nach vollständiger Demineralisation wurden die Zähne 5-6 Mal mit deionisiertem Wasser gründlich
gespült, bis ein ph-Wert von 7 erreicht war. Einige Zähne hatten sich trotz dem wochenlangen
Säurebad vollständig erhalten, die meisten waren aber stark zerfallen und beim Spülen musste darauf
geachtet werden, nicht zu viele Dentin-Stückchen zu verlieren. Die kleinsten Teile wurden entsorgt, da
sie bei der sequenzierten Probenentnahme nicht mehr ihrer ursprünglichen Lage am Zahn zugeordnet
werden konnten.
4.3.2 Sequenzierte Beprobung des Dentins
Die Erhaltung des Dentins war nach der Demineralisierung sehr unterschiedlich und führte zu einer
verschiedenen Anzahl Probesequenzen, die pro Zahn genommen werden konnte. Es waren zwischen
drei und sechs Sequenzen möglich, wobei zwei bis drei aus dem Bereich der Zahnkrone stammen
und ein bis drei aus der Wurzel. In zwei Fällen konnten nur drei Proben gewonnen werden, da die
Erhaltung des Dentins sehr schlecht war (RIZG1763.1.3 (Abb. 9) und RIZG710.1.1).

33
Abb. 11
Dentinsequenzen der Hirschzähne aus Zug-Riedmatt,
eingelegt in Natronlauge.
Abb. 10
Hirschzahn aus Zug-Riedmatt (RIZG1763.2.3), vor und
nach der Sequenzierung des Dentins.
Abb. 9
Hirschzahn aus Zug-Riedmatt (RIZG1763.1.3) demineralisiert, vor
Sequenzierung des Dentins.
RIZG1763.1.3.1d Bereich Zahnkrone
RIZG1763.1.3.2d
RIZG1763.1.3.3d Bereich Zahnwurzel
Die Zerlegung des Zahns erfolgte mit einem Skalpell, wobei bei der Krone begonnen wurde und nach
jedem Schnitt die Entfernung zur Schmelzgrenze - die in den meisten Fällen noch als Abdruck auf
dem Dentin sichtbar war - gemessen wurde (Abb. 10).
Zerteilung mit Skalpell in fünf Sequenzen
4.3.3 Aufreinigung des Dentins und Extraktion des Kollagens
Die einzelnen Dentinsequenzen wurden in ein 15ml-
Tube gegeben (passend zum EZEE-Filter), beschriftet
und anschliessend mit 5ml Natronlauge (NaOH 0.5N)
versetzt. Der Deckel wurde mit Luftlöchern versehen
und die Proben 2-3 Tage stehen gelassen. Je nach

34
Erhaltung und Grad der Verfärbung des Dentins konnte so eine Aufhellung des Dentins erreicht
werden. Bei manchen Proben musste das NaOH nach zwei Tagen ausgetauscht werden. Abbildung
11 zeigt deutlich die unterschiedlichen Verfärbungen der Proben durch Huminstoffe. Diese Sequenzen
stammen von verschiedenen Zähnen aus unterschiedlichen Positionen. Generell waren die
Sequenzen aus der Krone weniger stark verfärbt als jene aus der Wurzel und konnten durch die
Natronlauge dementsprechend besser aufgehellt werden. Dies muss eventuell bei der
Qualitätskontrolle der Messungen von δ15
N/δ13
C berücksichtigt werden. Es ist jedoch nicht bekannt,
welchen Einfluss die starke Verfärbung auf die Messergebnisse haben kann.
Sobald bei allen Proben eine zufriedenstellende Aufhellung erreicht war, wurden die Proben erneut 6-
7 Mal mit deionisiertem Wasser gespült, bis der ph-Wert wieder 7 betrug. Anschliessend wurden 4ml
Wasser und ca. 100µl Salzsäure (HCl 0.5N) hinzugefügt. Der ph-Wert sollte bei 2 liegen, was bei
manchen Proben einige Tropfen zusätzliches HCl erforderte.
Die angesäuerten Proben wurden anschliessend 2 Tage bei 70 Grad im Ofen geschmolzen, bis keine
Kollagen-Klümpchen mehr sichtbar waren. Anschliessend wurden mit den EZEE-Filtern alle
Verunreinigungen rausgefiltert, sodass nur reines gelöstes Kollagen übrig blieb. Diese Lösung wurde
in beschriftete 5ml-Tubes umgefüllt, welche daraufhin geöffnet und mit Parafilm umwickelt in den
Gefrierschrank gestellt wurden.
Für die folgende Gefriertrocknung wurde das Gerät vom Botanischen Institut der Uni Basel zur
Verfügung gestellt. Die tiefgefrorenen Proben wurden in einer isolierten Kiste transportiert, um ein
Auftauen zu verhindern. Nach der Instruktion zur Handhabung des Gefriertrockners durch den
Techniker Giacomo Busco, konnten alle Proben auf einmal während zwei Tagen getrocknet werden.
Leider stelle sich heraus, dass die Temperatur der Proben beim Einfüllen in den Trockner doch schon
zu hoch war, was zu einem mangelhaften Trocknungsvorgang führte und die Kollagenlösung z.T.
überschäumen liess. Nach zwei Tagen waren neun Proben noch nicht trocken und mussten erneut
eingefroren werden. Auf Hinweis von Herrn Thomas Boller (Botanisches Institut Universität Basel)
wurden diese Proben vor erneutem Trocknen noch für eine Stunde in den -80 Grad Gefrierschrank
gestellt. Dies stellte sich als eine wirkungsvolle Massnahme heraus, denn die Trocknung verlief
problemlos und vollständig.
Übrig blieb ein Kollagenfadennetz, das vor der Einwaage für die Massenspektrometrie noch mit einem
Spatel durchmischt wurde.
4.3.4 Massenspektrometrie
Für die Messungen der Kohlenstoff- und Stickstoffwerte aus dem Collagen wurde ca. 0.5 - 1.0 mg
Probenmenge eingewogen. Als Standards wurde EDTA und AO-1 verwendet, wovon je 0.3 – 1.0 mg
zur Messung eingewogen wurde. Die Messungen erfolgten mit einem Thermo Finnigan FlashEA 1112
in Verbindung mit dem Thermo Finnigan Delta V Advantage.
Es erfolgten routinemässige Doppelmessungen von ca. jeder 10. Probe, wobei sich die Abweichungen
für die Kohlenstoff- sowie Stickstoffwerte als gering herausstellten.

35
4.4 Wasserproben
Das Wasser wurde bei der Probenentnahme in 5ml-Tubes gefüllt und möglichst ohne Lufteinschluss
verschlossen. Die Tubes wurden beschriftet und im Labor für stabile Isotope des Institutes für
Umweltgeowissenschaften der Universität Basel abgegeben. Die Sauerstoffisotopenverhältnisse im
Wasser wurden dort mit einem LGR liquid water isotopes analyser gemessen.
5. Resultate
5.1 Die δ18O/δ13C – Werte aus dem Zahnschmelz
5.1.1 Die Messergebnisse aus Mainz und Basel im Vergleich
Die ersten Messungen in Mainz wurden mit Proben aus fünf verschiedenen Zähnen von vier
Individuen durchgeführt. Es handelt sich dabei um vier 3. Molare und einen 2. Molaren von
ausgewachsenen Hirschen. In Basel wurden anschliessend die Isotopenverhältnisse im Zahnschmelz
von allen zur Verfügung stehenden 23 Hirschzähnen gemessen (21 3. Molare, zwei 2. Molare und ein
1. Molar). Daraus resultieren die Doppelmessungen der ersten fünf Zähne. Es ergab sich somit die
Möglichkeit, die Messungen von beiden Labors (Mainz, Basel) zu vergleichen und allenfalls
Rückschlüsse auf die Qualität und Genauigkeit der Resultate zu ziehen.
Die ersten Ergebnisse der δ18
O-Werte aus Mainz (Abb. 12, grün) zeigen klare Kurvenverläufe, was
eine genaue und zuverlässige Datengenauigkeit vermuten lässt. Von den Messungen in Basel wurde
somit erwartet, dass sie diesen Werten möglichst nahe kommen, sich im Idealfall sogar decken
würden. Aus dem ersten Messdurchgang im Basler Labor resultierten allerdings Werte (Abb. 12,
blau), die z.T. grob von denjenigen aus Mainz abwichen (z.B. RIZG1846.1.2.2, RIZG1846.1.2.4,
RIZG1846.1.3.1). Zudem zeigen die Kurven keinen klaren Verlauf, sondern weisen durch einige
Ausreisser eher einen Zick-Zack-Kurs auf. Da dies die Interpretation erheblich erschwert, wurde eine
zweite Messung durchgeführt.

36
Abb. 12
δ18
O-Werte der fünf Hirschzähne aus Zug-Riedmatt; Messung der Universität Mainz sowie der 1. Und 2.
Messung der Universität Basel im Vergleich. Zur Erklärung der Probenbenennung s. Kapitel 4.1.
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
-8.00
-7.00
-6.00
12
44
.1.3
.1
12
44
.1.3
.3
12
44
.1.3
.5
13
42
.1.3
.1
13
42
.1.3
.3
13
42
.1.3
.5
18
46
.1.2
.2
18
46
.1.2
.4
18
46
.1.2
.6
18
46
.1.3
.1
18
46
.1.3
.3
18
46
.1.3
.5
18
46
.1.3
.7
18
46
.1.3
.9
18
46
.2.3
.1
18
46
.2.3
.3
18
46
.2.3
.5
18
46
.2.3
.7
δ18
O (
‰)
δ18O-Werte aus Mainz und Basel im Vergleich
1.δ18O_BS
2.δ18O_BS
δ18O_MZ
Die Resultate dieser zweiten Messung in Basel (Abb. 12, rot) ergaben klarere Kurvenverläufe in der
Grafik. Einzelne Ausreisser sind zwar auch hier vorhanden (z.B. Probe RIZG1846.1.3.4 mit
Standardfehler von 1.3‰ in der Doppelmessung), doch allgemein passen sich die Verläufe der Kurven
gut an diejenigen von Mainz an (schön zu sehen bei Zahn RIZG1846.2.3). Auffallend ist aber die klare
Verschiebung der Werte um ca. 1.3‰. Es bleibt abzuklären, wodurch diese deutliche Verschiebung
ausgelöst wird. Denkbar wären methodische Unterschiede während den Messabläufen im Labor. Bis
der Grund für diese beobachteten Unterschiede gefunden worden ist und somit die Glaubwürdigkeit
der Werte gesichert ist, wird auf eine Interpretation der absoluten δ18
O-Werte verzichtet. Die
Kurvenverläufe sowie die relativen Unterschiede zwischen den einzelnen Individuen können aber
trotzdem in die Auswertung miteinbezogen werden.
Trotz dieser Unklarheit wird im Folgenden mit den Resultaten aus der 2. Messung gearbeitet, da sie
deutlich weniger Ausreisser zeigen und somit klarere Kurvenverläufe (Tendenzen) sichtbar machen.
Die Kohlenstoffwerte (Abb. 13) zeigen sich im Gegensatz zu den δ18
O-Werten grösstenteils einheitlich
und reproduzierbar. Sowohl im Vergleich zu den δ13
C-Werten aus Mainz als auch zwischen den
beiden Messungen aus Basel bewegen sich die Daten in einem ähnlichen Rahmen und mit
übereinstimmenden Tendenzen. Einzige Ausnahme bildet die 2. Basler Messung von Zahn
RIZG1846.1.2. Da aber für die Sauerstoffisotope die 2. Messung genauer zu sein scheint, werden im
Folgenden auch nur die δ13
C-Werte aus derselben Messung berücksichtigt.

37
Abb. 14
δ18
O-Werte der fünf Hirschzähne aus Zug-Riedmatt; Messungen der Universität Mainz. Zur Erklärung der
Probenbenennung s. Kapitel 4.1.
Abb. 13
δ13
C-Werte der fünf Hirschzähne aus Zug-Riedmatt; Messung der Universität Mainz (grün) sowie der 1. (blau)
und 2. Messung (rot) der Universität Basel im Vergleich. Zur Erklärung der Probenbenennung s. Kapitel 4.1.
-17.50
-17.00
-16.50
-16.00
-15.50
-15.00
-14.50
-14.00
12
44
.1.3
.1
12
44
.1.3
.3
12
44
.1.3
.5
13
42
.1.3
.1
13
42
.1.3
.3
13
42
.1.3
.5
18
46
.1.2
.2
18
46
.1.2
.4
18
46
.1.2
.6
18
46
.1.3
.1
18
46
.1.3
.3
18
46
.1.3
.5
18
46
.1.3
.7
18
46
.1.3
.9
18
46
.2.3
.1
18
46
.2.3
.3
18
46
.2.3
.5
18
46
.2.3
.7
δ1
3 C (
‰)
δ13C-Werte aus Mainz und Basel im Vergleich
1.δ13C_BS
2.δ13C_BS
δ13C_MZ
5.1.2 Die Messergebnisse aus Mainz
Die δ18
O-Werte
-13.00
-12.50
-12.00
-11.50
-11.00
-10.50
-10.00
-9.50
-9.00
-8.50
-8.00
12
44
.1.3
.2
12
44
.1.3
.4
13
42
.1.3
.2
13
42
.1.3
.4
13
42
.1.3
.6
18
46
.1.2
.1
18
46
.1.2
.3
18
46
.1.2
.5
18
46
.1.3
.2
18
46
.1.3
.4
18
46
.1.3
.6
18
46
.1.3
.8
18
46
.2.3
.2
18
46
.2.3
.4
18
46
.2.3
.6
18
46
.2.3
.8
δ18
O (
‰)
δ18O-Werte aus Mainz
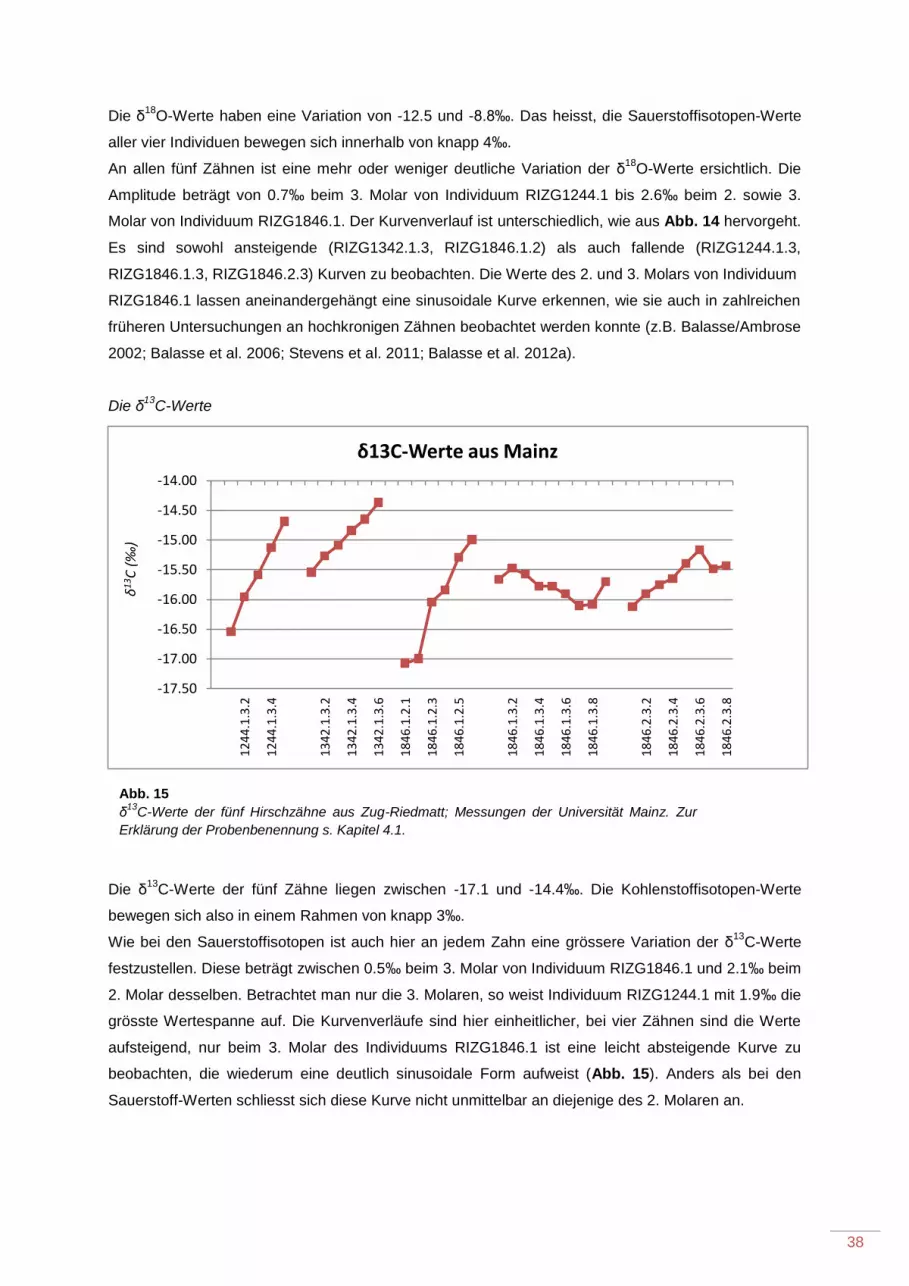
38
Abb. 15
δ13
C-Werte der fünf Hirschzähne aus Zug-Riedmatt; Messungen der Universität Mainz. Zur
Erklärung der Probenbenennung s. Kapitel 4.1.
-17.50
-17.00
-16.50
-16.00
-15.50
-15.00
-14.50
-14.00
12
44
.1.3
.2
12
44
.1.3
.4
13
42
.1.3
.2
13
42
.1.3
.4
13
42
.1.3
.6
18
46
.1.2
.1
18
46
.1.2
.3
18
46
.1.2
.5
18
46
.1.3
.2
18
46
.1.3
.4
18
46
.1.3
.6
18
46
.1.3
.8
18
46
.2.3
.2
18
46
.2.3
.4
18
46
.2.3
.6
18
46
.2.3
.8
δ1
3 C (
‰)
δ13C-Werte aus Mainz
Die δ18
O-Werte haben eine Variation von -12.5 und -8.8‰. Das heisst, die Sauerstoffisotopen-Werte
aller vier Individuen bewegen sich innerhalb von knapp 4‰.
An allen fünf Zähnen ist eine mehr oder weniger deutliche Variation der δ18
O-Werte ersichtlich. Die
Amplitude beträgt von 0.7‰ beim 3. Molar von Individuum RIZG1244.1 bis 2.6‰ beim 2. sowie 3.
Molar von Individuum RIZG1846.1. Der Kurvenverlauf ist unterschiedlich, wie aus Abb. 14 hervorgeht.
Es sind sowohl ansteigende (RIZG1342.1.3, RIZG1846.1.2) als auch fallende (RIZG1244.1.3,
RIZG1846.1.3, RIZG1846.2.3) Kurven zu beobachten. Die Werte des 2. und 3. Molars von Individuum
RIZG1846.1 lassen aneinandergehängt eine sinusoidale Kurve erkennen, wie sie auch in zahlreichen
früheren Untersuchungen an hochkronigen Zähnen beobachtet werden konnte (z.B. Balasse/Ambrose
2002; Balasse et al. 2006; Stevens et al. 2011; Balasse et al. 2012a).
Die δ13
C-Werte
Die δ13
C-Werte der fünf Zähne liegen zwischen -17.1 und -14.4‰. Die Kohlenstoffisotopen-Werte
bewegen sich also in einem Rahmen von knapp 3‰.
Wie bei den Sauerstoffisotopen ist auch hier an jedem Zahn eine grössere Variation der δ13
C-Werte
festzustellen. Diese beträgt zwischen 0.5‰ beim 3. Molar von Individuum RIZG1846.1 und 2.1‰ beim
2. Molar desselben. Betrachtet man nur die 3. Molaren, so weist Individuum RIZG1244.1 mit 1.9‰ die
grösste Wertespanne auf. Die Kurvenverläufe sind hier einheitlicher, bei vier Zähnen sind die Werte
aufsteigend, nur beim 3. Molar des Individuums RIZG1846.1 ist eine leicht absteigende Kurve zu
beobachten, die wiederum eine deutlich sinusoidale Form aufweist (Abb. 15). Anders als bei den
Sauerstoff-Werten schliesst sich diese Kurve nicht unmittelbar an diejenige des 2. Molaren an.

39
Abb. 16
δ18
O- und δ13
C-Werte der fünf Hirschzähne aus Zug-Riedmatt im Vergleich; Messungen der Universität Mainz.
Zur Erklärung der Probenbenennung s. Kapitel 4.1.
Die δ18
O-Werte und die δ13
C-Werte im Vergleich
Legt man die Kurven der Sauerstoff- und Kohlenstoffisotopenwerte übereinander so fällt auf, dass sie
bei zwei Zähnen (RIZG1342.1.3, RIZG1846.1.2) beinahe deckungsgleich sind und auch bei
RIZG1846.1.3 eine ähnliche Tendenz aufweisen. Die Kurven der Zähne RIZG1244.1.3 sowie
RIZG1846.2.3 weichen jedoch deutlich voneinander ab, bewegen sich sogar in gegensätzliche
Richtungen. Für die Beschreibung der Lage der Isotopenkurve zueinander werden im Folgenden die
Begriffe gleichläufig bzw. gegenläufig verwendet. Bei der Interpretation ist vor allem die Tendenz (also
auf- oder absteigend) wichtig und nicht die genaue Deckungsgleichheit der δ18
O- und δ13
C-Kurven. So
werden auch die Kurven von Zahn RIZG1846.1.3 als gleichläufig angesprochen.
Auffallend bei Zahn RIZG1244.1.3 ist, dass sich die Spanne der Werte stark unterscheidet. So ist die
Amplitude der δ13
C-Kurve sehr gross (1.9‰), diejenige der δ18
O-Kurve aber auffallend klein (0.8‰).
Bei den beiden ebenfalls gegenläufigen Kurven von Zahn RIZG1846.2.3 liegen die Amplituden (-1.0
bzw. -1.9‰) im Vergleich mit den anderen Zähnen im mittleren Bereich.
5.1.3 Messergebnisse aus Basel
Die δ18
O-Werte
Die δ18
O-Werte der 23 Hirschzähne bewegen sich zwischen -12.4 und -5.7‰, also innerhalb von fast
7‰. Wie schon bei den fünf Zähnen die in Mainz gemessen wurden, sind auch hier die Werte
innerhalb eines Zahnes sehr unterschiedlich. Die Amplitude reicht von 0.8‰ bei Zahn RIZG1244.1.3
bis zu 4.5‰ bei Zahn RIZG1890.1.3 (Abb. 19-21).
Die Kurvenverläufe sind sehr unterschiedlich, es ist kein einheitliches Bild zu erkennen. Vereinzelt
zeigen sich angedeutete sinusoidale Kurven wie bei den Zähnen RIZG1763.1.3 und RIZG952.1.3,
ansonsten sind die Isotopenwerte der einzelnen Proben auf- oder absteigend. Da auch grössere
Messungenauigkeiten nicht gänzlich ausgeschlossen werden können, sind die „peaks“ in den
-17.50
-17.00
-16.50
-16.00
-15.50
-15.00
-14.50
-14.00
-13.00
-12.50
-12.00
-11.50
-11.00
-10.50
-10.00
-9.50
-9.00
-8.50
-8.00
12
44
.1.3
.2
12
44
.1.3
.4
13
42
.1.3
.2
13
42
.1.3
.4
13
42
.1.3
.6
18
46
.1.2
.1
18
46
.1.2
.3
18
46
.1.2
.5
18
46
.1.3
.2
18
46
.1.3
.4
18
46
.1.3
.6
18
46
.1.3
.8
18
46
.2.3
.2
18
46
.2.3
.4
18
46
.2.3
.6
18
46
.2.3
.8
δ1
3 C (‰
)
δ1
8 O (
‰)
δ18O- und δ13C-Werte aus Mainz
δ18O
δ13C

40
-13
-12
-11
-10
-9
-8
-7
-6
-5
δ1
8 O (
‰)
δ18O-Wertespannen nach Schicht
Abb. 17
δ18
O-Wertespannen aus dem Zahnschmelz aller 23
Hirschzähne aus Zug-Riedmatt. Nach Siedlungsschichten
getrennt und innerhalb derer aufsteigend nach Minimalwert
geordnet. Erläuterungen zur Stratigraphie s. Kapitel 2.2.2.
einzelnen Kurven nur bedingt zu interpretieren (wie z.B. bei den Zähnen RIZG1890.1.3, RIZG1846.3.3
oder RIZG1464.1.3). Es können lediglich die Trends sowie die Unterschiede zwischen den Individuen
bewertet werden.
Die δ18
O-Wertespannen getrennt nach Schicht
Um allfällige Unterschiede in den δ18
O-Werten zwischen den einzelnen Siedlungsschichten sichtbar
zu machen, werden die Wertespannen nach Schicht getrennt und nach aufsteigendem Minimalwert
dargestellt (Abb. 17).
Es zeigt sich, dass zwischen den
Schichten keine deutlichen Unterschiede
in der Werteverteilung vorhanden sind,
die Spannweite der Werte liegt in allen
Siedlungsphasen (ausser D1/D2, da nur
ein Individuum) zwischen 4.7‰ (Schicht
C1) und 5.5‰ (Schicht B2). D.h. es fällt
keine Schicht mit einer Anhäufung von
besonders tiefen oder hohen Werten auf.
Auf eine genauere statistische Analyse
der Verteilung wird verzichtet, da bei der
Messgenauigkeit der Sauerstoffisotopen-
Werte noch Verbesserungen
vorgenommen werden müssen.
In allen vier Siedlungsphasen mit
mehreren beprobten Zähnen befinden
sich Individuen, deren δ18
O-Werte sich
nicht oder nur ganz leicht mit denjenigen
von anderen Hirschen aus der gleichen
Schicht überschneiden.
Die δ13
C-Werte
Die δ13
C-Werte der 23 Hirschzähne bewegen sich zwischen -16.8‰ und -13.0‰, also innerhalb von
fast 4‰. Die Variation innerhalb der Zähne schwankt sehr stark zwischen 0.4‰ (RIZG1901.1.3) und
1.9‰ (RIZG1244.1.3) (Abb. 19-21).
Die Kurvenverläufe sind etwas klarer als diejenige der Sauerstoffwerte - Ausreisser die nicht in den
Verlauf passen sind nur selten zu beobachten (z.B. RIZG1348.3.3.4, RIZG1342.2.3.4). Mehrheitlich
verlaufen die Kurven aufsteigend, entweder gerade (z.B. RIZG1342.1.3, RIZG799.1.2, RIZG1348.1.3)
oder mehr oder weniger deutlich sinusoidal (z.B. RIZG1890.1.3, RIZG1763.1.3, RIZG1464.1.3,
RIZG952.1.3, RIZG710.1.1).
D1/
D2
B B1
1
B2 C1

41
-17.5
-16.5
-15.5
-14.5
-13.5
-12.5
δ1
3 C (
‰)
δ13C- Wertespannen nach Schicht
B B1 B2 C1 D1/D2
Abb. 18
δ13
C-Wertespannen aus dem
Zahnschmelz aller 23 Hirschzähne aus
Zug-Riedmatt. Nach Siedlungsschichten
getrennt und innerhalb derer aufsteigend
nach Minimalwert geordnet. Erläuterungen
zur Stratigraphie s. Kapitel 2.2.2.
Die δ13
C- Wertespannen getrennt nach Schicht
Wie bei den δ18
O-Werten sind keine offensichtlichen Unterschiede der Wertespannen zwischen den
Siedlungsschichten erkennbar. Die Werte liegen alle ca. zwischen -14.3 und -16.7‰ (Abb. 18,
markierter Bereich). Nur die Werte der beiden Individuen RIZG799.1.2 (Schicht B) und RIZG1763.1.3
(Schicht B1, Knochenhaufen) liegen deutlich höher, bei einer überdurchschnittlich hohen Spannweite
der Werte innerhalb des markierten Bereichs überschneiden sich die Werte der einzelnen Individuen
einer Schicht grösstenteils deutlich bzw. teilweise.
Die δ18
O- und die δ13
C-Werte im Vergleich
Die Beobachtungen aus den Resultaten der Mainzer Messungen bestätigen sich an den Sauerstoff-
und Kohlenstoffisotopen-Werten aus Basel. Die Verläufe der Sauerstoff- und Kohlenstoff-Kurven
scheinen zumindest teilweise einen Zusammenhang zu haben. Es können Gruppen von gleichläufigen
(Abb. 19) und gegenläufigen (Abb. 20) Kurvenpaaren gebildet werden, sowie von solchen mit nicht
genau zu definierendem Verhältnis zueinander (Abb. 21). Um als gleich- oder gegenläufig
beschrieben zu werden, müssen die Kurven mindestens zu 50% die definierte Tendenz aufweisen.
Bei Kurven mit unklarem Verlauf oder stark schwankenden Werten (Ausreisser) muss die Möglichkeit
in Betracht gezogen werden, dass die Ursache dafür in Messfehlern mit bisher unbekannter Ursache
und Ausmass liegen könnte.

42
-17.50
-16.50
-15.50
-14.50
-13.50
-12.50
-13.00
-12.00
-11.00
-10.00
-9.00
-8.00
-7.00
-6.00
-5.00
13
42
.1.3
.21
34
2.1
.3.4
13
42
.1.3
.67
99
.1.2
.17
99
.1.2
.3
18
90
.1.3
.21
89
0.1
.3.4
18
90
.1.3
.61
89
0.1
.3.8
17
63
.1.3
.11
76
3.1
.3.3
17
63
.1.3
.5
17
63
.2.3
.21
76
3.2
.3.4
17
76
.1.3
.11
77
6.1
.3.3
17
76
.1.3
.5
13
48
.1.3
.21
34
8.1
.3.4
13
48
.1.3
.61
53
5.1
.3.1
15
35
.1.3
.31
53
5.1
.3.5
95
2.1
.3.1
95
2.1
.3.3
95
2.1
.3.5
95
2.1
.3.7
10
67
.1.3
.11
06
7.1
.3.3
10
67
.1.3
.5
δ13
C (
‰)
δ18
O (
‰)
δ18O- und δ13C-Kurve gleichläufig
δ18O
δ13C
Abb. 19
Die δ18
O- und δ13
C-Werte aus dem Zahnschmelz der 10 Hirsche aus Zug-Riedmatt, deren Kurvenverlauf
tendenziell gleichläufig ist. Messungen der Universität Basel. Zur Erklärung der Probenbenennung s. Kapitel
4.1.
Abb. 20
Die δ18
O- und δ13
C-Werte aus dem Zahnschmelz der 4 Hirsche aus Zug-Riedmatt, deren Kurvenverlauf
tendenziell gegenläufig ist. Messungen der Universität Basel. Zur Erklärung der Probenbenennung s.
Kapitel 4.1.
-17.50
-16.50
-15.50
-14.50
-13.50
-12.50
-13.00
-12.00
-11.00
-10.00
-9.00
-8.00
-7.00
-6.00
-5.00
12
44
.1.3
.11
24
4.1
.3.2
12
44
.1.3
.31
24
4.1
.3.4
12
44
.1.3
.5
18
46
.2.3
.11
84
6.2
.3.2
18
46
.2.3
.31
84
6.2
.3.4
18
46
.2.3
.51
84
6.2
.3.6
18
46
.2.3
.71
84
6.2
.3.8
18
46
.3.3
.11
84
6.3
.3.2
18
46
.3.3
.31
84
6.3
.3.4
18
46
.3.3
.51
84
6.3
.3.6
18
46
.3.3
.7
13
48
.2.3
.11
34
8.2
.3.2
13
48
.2.3
.31
34
8.2
.3.4
13
48
.2.3
.51
34
8.2
.3.6
δ13
C (
‰)
δ18
O (
‰)
δ18O- und δ13C-Kurve gegenläufig
δ18O
δ13C

43
Abb. 21
Die δ18
O- und δ13
C-Werte aus dem Zahnschmelz der 9 Hirsche aus Zug-Riedmatt, deren Kurvenverlauf
zueinander keine klare Tendenz aufweist. Messungen der Universität Basel. Zur Erklärung der
Probenbenennung s. Kapitel 4.1.
-17.50
-16.50
-15.50
-14.50
-13.50
-12.50
-13.00
-12.00
-11.00
-10.00
-9.00
-8.00
-7.00
-6.00
-5.00
18
46
.1.2
.3
18
46
.1.2
.6
18
46
.1.3
.2
18
46
.1.3
.5
18
46
.1.3
.8
19
01
.1.3
.1
19
01
.1.3
.4
19
01
.1.3
.7
18
46
.4.3
.1
18
46
.4.3
.4
13
42
.2.3
.3
13
42
.2.3
.6
13
48
.3.3
.2
13
48
.3.3
.5
14
64
.1.3
.1
14
64
.1.3
.4
14
64
.1.3
.7
12
38
.1.3
.2
12
38
.1.3
.5
71
0.1
.1.1
71
0.1
.1.4
δ13
C (
‰)
δ18
O (
‰)
δ18O- und δ13C-Kurven eher unklar
δ18O
δ13C

44
5.2 Die δ13C/δ15N-Werte aus dem Collagen des Zahndentins
Überprüfung der Kollagen-Qualität
Aufgrund der Tatsache, dass die Kollagen-Proben erst nach der Demineralisierung sequenziert
wurden, bestand keine Möglichkeit das Trockengewicht der einzelnen Proben zu nehmen und somit
die Kollagen-Ausbeute genau zu bestimmen. Alle Proben wiesen jedoch eine grosse Menge an
Kollagen auf (durchschnittlich etwa 40-100mg), was auf eine gute Erhaltung schliessen lässt. Dies war
bei der Feuchtbodenerhaltung auch zu erwarten.
Die Anteile von Kohlenstoff (%C) und Stickstoff (%N) sowie die das C/N-Verhältnis im Kollagen
weisen sehr einheitliche Werte auf, in der Grössenordnung wie sie auch bei modernem Kollagen aus
Dentin zu beobachten sind. Es gibt nur eine Probe - RIZG1848.2.3.4 -, deren Anteile von Kohlenstoff
und Stickstoff tiefer liegen (%C=34.8, %N=12.3) als bei den restlichen Proben. Das C/N-Verhältnis
liegt mit 3.29 jedoch im normalen Bereich.
Bei allen anderen Proben liegen die Anteile von Kohlenstoff (%C) zwischen 40.6 und 45.6% und
diejenigen von Stickstoff (%N) zwischen 14.4 und 16.3%. Die C/N-Verhältnisse im Kollagen bewegen
sich zwischen 3.24 und 3.34. Diese Werte bestätigen den optischen Eindruck, dass das Material (also
die Zähne inkl. Dentin) sehr gut erhalten ist und somit seine ursprünglich isotopische
Zusammensetzung erhalten geblieben ist. Dies ist eine wichtige Voraussetzung zur Interpretation der
gemessenen δ13
C- und δ15
N-Werte.
Die δ13
C-Werte
Die δ13
C-Werte aus dem Kollagen des Zahndentins bewegen sich in einer Grössenordnung zwischen
-26.0 und -22.2‰. Die Wertespannen innerhalb der Zähne sind sehr unterschiedlich, sie liegen
zwischen 0.2‰ (RIZG1348.3.3) und 1.8‰ (RIZG1846.1.2; M2!). Die grösste Amplitude unter den M3
hat RIZG1846.1.3 mit knapp 1.5‰, also das gleiche Individuum von dem auch der M2 mit der
höchsten Amplitude stammt (Abb. 24-25).
Die Kurvenverläufe sind sehr unterschiedlich, wobei aber bei einigen Individuen (n=13; z.B.
RIZG1846.3.3, RIZG1342.2.3, RIZG1348.1.3, RIZG1067.1.3) die Kurve als eindeutig aufsteigend
beschrieben werden kann und bei anderen (n=6; z.B. RIZG1763.1.3, RIZG952.1.3, RIZG1244.1.3)
eher als abfallend. Bei den restlichen (n=4; z.B. RIZG1776.1.3, RIZG1348.3.3) ist der Verlauf ohne
klare bzw. mit schwacher Tendenz. Die meisten Kurven weisen einen Umbruch in der Richtung auf,
also einen Knick. Diese könnten wohl als sinusoidale Formen angesprochen werden, falls die
Sequenzierung eine höhere Auflösung hätte, d.h. eine dichtere Beprobung erfolgt wäre.

45
-26.5
-26
-25.5
-25
-24.5
-24
-23.5
-23
-22.5
-22
-21.5
δ13
C (
‰)
δ13C- Wertespannen nach Schicht
M2!
M2
B B1 B2 C1 D1/
D2
Abb. 22
δ13
C-Wertespannen (Kollagen) aller 23 Hirschzähne aus
Zug-Riedmatt. Nach Siedlungsschichten getrennt und
innerhalb derer aufsteigend nach Minimalwert geordnet.
Erläuterungen zur Stratigraphie s. Kapitel 2.2.2.
Die δ13
C- Wertespannen getrennt nach Schicht
Die aussergewöhnlich tiefen Werte in
Schicht B1 stammen vom Zahn
RIZG1846.1.2, also einem 2. Molar. Auch
die hohen Werte über -23‰ aus Schicht
B stammen von einem M2. In den
Schichten B1 und B2 fallen je zwei
Individuen (RIZG1763.1.3, RIZG1763.2.3
bzw. RIZG1348.1.3, RIZG1342.1.3) mit
erhöhten Werten auf, die sich mit den
restlichen Werten aus den Schichten
kaum (B2) oder gar nicht (B1)
überschneiden (Abb. 22).
Dies sind die einzigen sichtbaren
Ausreisser, die restlichen Wertespannen
liegen in allen Siedlungsschichten auf
einem ähnlichen Niveau zwischen -25.2
und 23.0‰. In Schicht C1 sind keine
höheren Werte vorhanden wie in Schicht
B-B2, was aber aufgrund der kleinen
Probenmenge von vier Zähnen nicht als
signifikant betrachtet werden kann.
Die δ15
N-Werte
Die δ15
N-Werte aus dem Kollagen reichen von 4.1 bis 7.8‰, weisen also eine Variation von 3.7‰ auf.
Die Amplituden sind wiederum sehr unterschiedlich und liegen zwischen 0.15‰ (RIZG1846.3.3) und
2.35‰ (RIZG1846.1.2; M2) bzw. 1.62‰ (RIZG1763.1.3; M3). Die Verläufe der Kurven sind vielfältig.
Ca. 12 Kurven können als ansteigend beschrieben werden (z.B. RIZG1901.1.3, RIZG1846.2.3,
RIZG1348.1.3, RIZG1244.1.3), nur eine als deutlich absteigend (RIZG1846.1.3). Einige Proben haben
zu wenige Sequenzen, um eine Tendenz festzulegen bzw. haben eine sehr kleine Amplitude mit einer
angedeuteten sinusoidalen Kurve (z.B. RIZG1348.3.3, RIZG1464.1.3, RIZG1535.1.3) (Abb. 24-25).
Die δ15
N-Wertespannen nach Schicht
Die δ15
N-Werte zeigen eine ziemlich regelmässige Streuung innerhalb der Schichten in der
Grössenordnung zwischen 4.1 und 6.8‰. Lediglich der M2 (RIZG1846.1.2) und der M1 (710.1.1)
weisen Werte über 7‰ auf.
Obwohl die Anzahl der Zähne in Schicht B (n=3) und C1 (n=4) sehr gering sind, kann festgehalten
werden, dass die δ15
N-Werte in der ältesten Siedlungsschicht B tiefer sind (4.1 - 5.7‰) als in der
jüngeren Schicht C1 (5.4 – 6.6‰). In den Schichten B1 und B2 mit acht bzw. sieben beprobten
Zähnen spannen sich die Werte fast über den ganzen Wertebereich der 3. Molare (Abb. 23).

46
3.50
4.00
4.50
5.00
5.50
6.00
6.50
7.00
7.50
8.00
δ15
N(‰
)
δ15N-Wertespannen nach Schicht Abb. 23
δ15
N-Wertespannen (Kollagen) aller 23
Hirschzähne aus Zug-Riedmatt. Nach
Siedlungsschichten getrennt und innerhalb
derer aufsteigend nach Minimalwert
geordnet. Erläuterungen zur Stratigraphie
s. Kapitel 2.2.2.
B B1 B2 C1 D1/
D2
Die δ13
C- und die δ15
N-Werte aus dem Kollagen des Dentins im Vergleich
Im Gegensatz zu den δ18
O- und δ13
C-Werten aus dem Karbonat konnten hier alle Kurven in
gleichläufig (Abb. 24) oder gegenläufig (Abb. 25) eingeteilt werden. Dies liegt vermutlich an der
geringeren Anzahl Sequenzen pro Zahn aus dem Dentin, was eine geringere Auflösung des
Kurvenverlaufs zur Folge hat. Trotzdem sind deutliche Unterschiede zu erkennen.
Der Verlauf der Kurven – also die Gleich- oder Gegenläufigkeit – ist unabhängig vom Wertebereich
(hohe oder niedrige Werte) und von der Wertespanne (grosse oder kleine Amplitude). Auch zeigen
sich keine Häufungen einer Tendenz in bestimmten Siedlungsschichten. Auch das Alter bzw. der
Abkauungsgrad der Zähne hat keinen Einfluss auf das Verhältnis der δ13
C- und δ15
N-Kurven
zueinander.
Auffallend ist, dass bei den gleichläufig verlaufenden Kurven die δ15
N-Werte schon einen
„Richtungswechsel“ vollziehen, während die δ13
C-Werte noch in eine Richtung weiterlaufen (z.B.
RIZG1342.1.3, RIZG1342.2.3, RIZG1348.1.3, RIZG1067.1.2). Eine Veränderung in den Umwelt- oder
Lebensbedingungen könnte sich also womöglich schneller in den Stickstoff- als in den
Kohlenstoffisotopen niedergeschlagen haben.
M2 M1

47
Abb. 24
Die δ13
C- und die δ15
N-Werte aus dem Collagen der 13 Hirsche aus Zug-Riedmatt, deren Kurvenverlauf
tendenziell gleichläufig ist. Messungen der Universität Basel. Zur Erklärung der Probenbenennung s. Kapitel
4.1.
3.50
4.00
4.50
5.00
5.50
6.00
6.50
7.00
7.50
8.00
-26.50
-26.00
-25.50
-25.00
-24.50
-24.00
-23.50
-23.00
-22.50
-22.00
-21.50
18
90
.1.3
.3d
19
01
.1.3
.1d
19
01
.1.3
.4d
17
63
.2.3
.1d
17
63
.2.3
.4d
18
46
.1.2
.1d
18
46
.1.2
.4d
18
46
.2.3
.3d
18
46
.4.3
.3d
13
42
.1.3
.3d
13
42
.1.3
.6d
13
42
.2.3
.2d
13
42
.2.3
.5d
13
48
.1.3
.2d
13
48
.1.3
.5d
13
48
.2.3
.1d
13
48
.2.3
.4d
13
48
.3.3
.1d
13
48
.3.3
.4d
15
35
.1.3
.1d
15
35
.1.3
.4d
10
67
.1.3
.1d
10
67
.1.3
.4d
δ15
N(‰
)
δ13
C (
‰)
δ13C- und δ15N-Kurve gleichläufig
δ13C
δ15N
Abb. 25
Die δ13
C- und die δ15
N-Werte aus dem Collagen der 10 Hirsche aus Zug-Riedmatt, deren
Kurvenverlauf tendenziell gegenläufig ist. Messungen der Universität Basel. Zur Erklärung der
Probenbenennung s. Kapitel 4.1.
3.50
4.00
4.50
5.00
5.50
6.00
6.50
7.00
7.50
8.00
-26.50
-26.00
-25.50
-25.00
-24.50
-24.00
-23.50
-23.00
-22.50
-22.00
-21.50
79
9.1
.2.2
d7
99
.1.2
.4d
17
63
.1.3
.2d
17
76
.1.3
.2d
17
76
.1.3
.4d
18
46
.1.3
.2d
18
46
.1.3
.4d
18
46
.1.3
.6d
18
46
.3.3
.1d
18
46
.3.3
.3d
18
46
.3.3
.5d
14
64
.1.3
.1d
14
64
.1.3
.3d
95
2.1
.3.2
d9
52
.1.3
.4d
12
44
.1.3
.1d
12
44
.1.3
.3d
12
44
.1.3
.5d
12
38
.1.3
.1d
12
38
.1.3
.3d
12
38
.1.3
.5d
71
0.1
.1.1
d7
10
.1.1
.3d
δ15
N(‰
)
δ13
C (
‰)
δ13C- und δ15N-Kurve gegenläufig
δ13C
δ15N
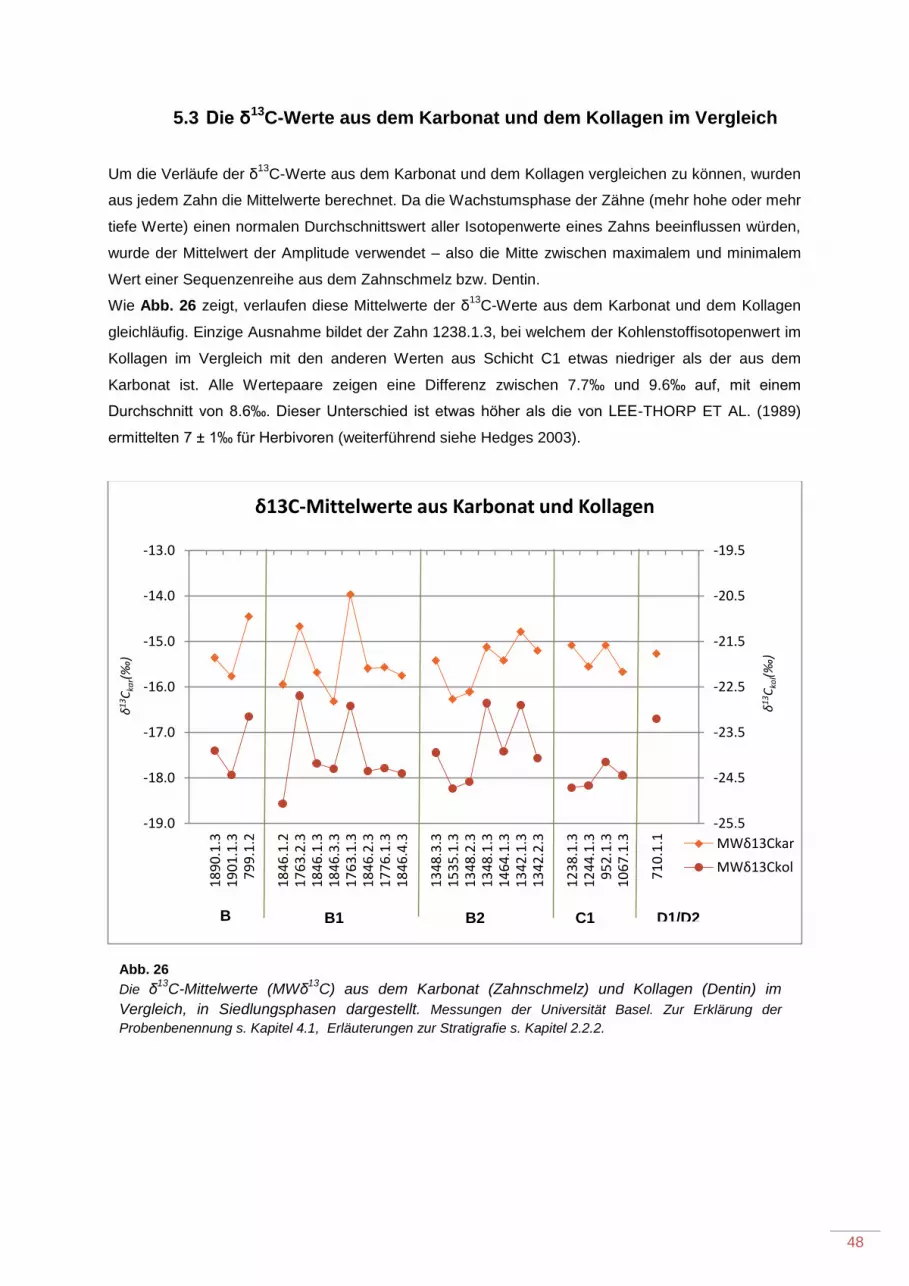
48
Abb. 26
Die δ13
C-Mittelwerte (MWδ13
C) aus dem Karbonat (Zahnschmelz) und Kollagen (Dentin) im
Vergleich, in Siedlungsphasen dargestellt. Messungen der Universität Basel. Zur Erklärung der
Probenbenennung s. Kapitel 4.1, Erläuterungen zur Stratigrafie s. Kapitel 2.2.2.
5.3 Die δ13C-Werte aus dem Karbonat und dem Kollagen im Vergleich
Um die Verläufe der δ13
C-Werte aus dem Karbonat und dem Kollagen vergleichen zu können, wurden
aus jedem Zahn die Mittelwerte berechnet. Da die Wachstumsphase der Zähne (mehr hohe oder mehr
tiefe Werte) einen normalen Durchschnittswert aller Isotopenwerte eines Zahns beeinflussen würden,
wurde der Mittelwert der Amplitude verwendet – also die Mitte zwischen maximalem und minimalem
Wert einer Sequenzenreihe aus dem Zahnschmelz bzw. Dentin.
Wie Abb. 26 zeigt, verlaufen diese Mittelwerte der δ13
C-Werte aus dem Karbonat und dem Kollagen
gleichläufig. Einzige Ausnahme bildet der Zahn 1238.1.3, bei welchem der Kohlenstoffisotopenwert im
Kollagen im Vergleich mit den anderen Werten aus Schicht C1 etwas niedriger als der aus dem
Karbonat ist. Alle Wertepaare zeigen eine Differenz zwischen 7.7‰ und 9.6‰ auf, mit einem
Durchschnitt von 8.6‰. Dieser Unterschied ist etwas höher als die von LEE-THORP ET AL. (1989)
ermittelten 7 ± 1‰ für Herbivoren (weiterführend siehe Hedges 2003).
-25.5
-24.5
-23.5
-22.5
-21.5
-20.5
-19.5
-19.0
-18.0
-17.0
-16.0
-15.0
-14.0
-13.0
18
90
.1.3
19
01
.1.3
79
9.1
.2
18
46
.1.2
17
63
.2.3
18
46
.1.3
18
46
.3.3
17
63
.1.3
18
46
.2.3
17
76
.1.3
18
46
.4.3
13
48
.3.3
15
35
.1.3
13
48
.2.3
13
48
.1.3
14
64
.1.3
13
42
.1.3
13
42
.2.3
12
38
.1.3
12
44
.1.3
95
2.1
.31
06
7.1
.3
71
0.1
.1
δ13
Cko
l(‰)
δ13
Cka
r(‰
)
δ13C-Mittelwerte aus Karbonat und Kollagen
MWδ13Ckar
MWδ13Ckol
B
B
B1
B
B
B2
B
B
C1
B
B
D1/D2
B
B

49
Abb. 27
Korrelation der δ18
O-Werte
der beprobten Gewässer
rund um Zug und Umgebung
mit der Höhenlage. Trendlinie
in Rot.
-10.0
-9.5
-9.0
-8.5
-8.0
-7.5
-7.0
0 500 1000 1500
δ18
O (
‰)
Höhe [M.ü.M.]
d18O Werte des Wassers
5.4 Resultate Wasserproben
Die Resultate der Messungen aus verschiedenen Gewässern (Zugersee, Lorze, Ägerisee und
Regenwasser) zeigen eine deutliche Korrelation zwischen Höhenlage und δ18
O-Wert im Wasser.
Der höchste Wert mit -7.5‰ stammt aus dem Zugersee auf 413 M.ü.M. und der niedrigste Wert mit
-9.2‰ vom Regenwasser aus 1500 M.ü.M. Diese Differenz von 1.7‰ entspricht einer Abnahme von
knapp 0.16‰ auf 100 Höhenmeter. Dies ist etwas geringer als die oft zitierten 0.2‰ auf 100 Meter,
laut SCHOTTERER (2010) sind Schwankungen aber die Regel und fallen über alle Messstationen der
Schweiz oft beträchtlich aus (Schotterer 2010, 1077).
Da es während der Sammlung der Proben im Oktober 2012 stark regnete, handelt es sich bei den
vorliegenden Werten vermutlich um die Isotopenzusammensetzung des Regenwassers von jenem
Tag. Aus diesem Grund können die Daten mit denjenigen aus der internationalen Datenbank des
IAEA (International Atomic Energy Agency) verglichen werden, die öffentlich zugänglich ist
(http://www.univie.ac.at/cartography/project/wiser/). Diese Datensammlung enthält Isotopenwerte von
Messstationen aus der ganzen Welt. Idealerweise werden Werte aber mit solchen aus örtlich nahen
Stationen verglichen. In diesem Fall sind die nächsten Stationen Bern (511 M.ü.M.), Meiringen (632
M.ü.M.) und Guttannen (1055 M.ü.M.). Der Niederschlag im Oktober an diesen Orten weist deutlich
tiefere δ18
O-Werte auf als in dieser Studie in der Region Zug-Schwyz gemessen wurden (Bern: -
10.3‰, Meiringen: -11.7‰, Guttannen: -12.7‰). Ob diese Differenzen von ca. 4‰ auf regionale
Unterschiede zurückzuführen sind oder auf einen Messfehler an den vorliegenden Proben kann an
dieser Stelle nicht abschliessend bewertet werden. Die publizierten Daten von SCHOTTERER (2010,
1077) bestätigen aber die Vermutung, dass die δ18
O-Werte dieser Studie zu hoch ausfallen.
B B1 B2
Zugersee (Zug/Arth)
Lorze/Ägerisee
Pfütze Sternegg

50
Abb. 28
δ18
O-Werte aus den sequenziell
beprobten Zähnen des Individuums
RIZG1846.1. Messung aus Mainz,
da der Kurvenverlauf hier deutlicher
ist als bei der Messung in Basel von
den gleichen Zähnen. Die Kurve
reflektiert die saisonal
unterschiedlichen,
temperaturabhängigen
Sauerstoffwerte aus dem
Trinkwasser der Hirsche
(sinusoidale Kurve in Rot,
angepasst). Zur Erklärung der
Probenbenennung s. Kapitel 4.1.
6. Diskussion
6.1 Grosse Unterschiede im Verlauf der Isotopenkurven (δ18O/δ13C) aus
dem Zahnschmelz – Hinweis auf verschiedene Populationen oder
normale Variation zwischen Individuen?
Bei einer Hirschpopulation, in welcher alle Jungtiere zwischen Mai und Juni geboren werden und die
gleiche physische Entwicklung durchlaufen wäre, zu erwarten, dass auch die Zahnentwicklung mehr
oder weniger synchron verläuft. Wie in Kapitel 2.3.3 beschrieben, gibt es aber vor allem bei der
Mineralisation des M3 grosse Unterschiede zwischen den einzelnen Individuen – sowohl beim
Zeitpunkt des Mineralisationsbeginns als auch bei der Mineralisationsdauer.
Die Kurvenverläufe der δ18
O-Werten aus den Sequenzen des Zahnschmelzes entstehen durch die
unterschiedliche Isotopenkonzentration im Regenwasser im Verlauf eines Jahres. Durch die
Aufnahme des meteorischen Wassers durch den Hirsch werden die Isotopensignale in dem sich
bildenden Zahnschmelz festgehalten, ohne sich nachträglich wieder zu verändern (s. Kapitel 2.4.3 und
2.5.1). Bei den Zähnen von jüngeren Individuen – wo noch kein Materialverlust am Zahn durch
Abkauung entstanden ist – sollte also die gesamte Zeitspanne der Zahnmineralisation aus den
Isotopenwerten sichtbar sein.
Dies ist bei Individuum RIZG1846.1 der Fall, von dem sowohl der M2 als auch der M3 beprobt wurde.
Durch das geringe Alter von 2-3 Jahren wies der M2 nur schwache und der M3 kaum
Abnutzungsspuren auf. Da der M3 etwa zu dem Zeitpunkt zu mineralisieren begann, als die Krone des
M2 vollständig war, ergibt sich eine sinusoidale Kurve (Abb. 28, in rot stilisierte Sinuskurve) die den
Zeitraum von über einem Jahr abdeckt (ca. Januar bis Februar des nächsten Jahres; Vgl. Stevens et
al. 2011, 67).
-13.00
-12.50
-12.00
-11.50
-11.00
-10.50
-10.00
-9.50
-9.00
RIZ
G 1
84
6.1
.2.1
RIZ
G 1
84
6.1
.2.2
RIZ
G 1
84
6.1
.2.3
RIZ
G 1
84
6.1
.2.4
RIZ
G 1
84
6.1
.2.5
RIZ
G 1
84
6.1
.2.6
RIZ
G 1
84
6.1
.3.1
RIZ
G 1
84
6.1
.3.2
RIZ
G 1
84
6.1
.3.3
RIZ
G 1
84
6.1
.3.4
RIZ
G 1
84
6.1
.3.5
RIZ
G 1
84
6.1
.3.6
RIZ
G 1
84
6.1
.3.7
RIZ
G 1
84
6.1
.3.8
RIZ
G 1
84
6.1
.3.9
δ18
O (
‰)
Jahreszyklus in den δ18O-Werten von M2 und M3
Jahreszyklus 12 Monate Jan-Jan
M2 M3

51
-13.00
-12.00
-11.00
-10.00
-9.00
-8.00
-7.00
-6.00
-5.00
18
90
.1.3
.1
18
90
.1.3
.4
18
90
.1.3
.7
17
63
.1.3
.1
17
63
.1.3
.4
14
64
.1.3
.3
14
64
.1.3
.6
95
2.1
.3.1
95
2.1
.3.4
95
2.1
.3.7
10
67
.1.3
.2
10
67
.1.3
.5
δ18
O (
‰)
δ18O-Werte aus dem M3 von jungen Hirschen (ohne Abkauung)
Abb. 29
δ18
O-Werte von den 3. Molaren fünf junger Hirsche, die
praktisch keine Abkauungsspuren aufwiesen. Die
ähnlichen Kurvenverläufe spiegeln eine saisonal
vergleichbare Zahnentwicklung wieder. Zur Erklärung
der Probenbenennung s. Kapitel 4.1.
Davon ausgehend, dass schon etwas Material vom M2 abgekaut war und somit nicht im
Wertespektrum vertreten ist, dauerte die Mineralisationsphase vom M2 von ca. Dezember bis Juli.
Laut BROWN (1991) beginnt die Mineralisation des M2 ziemlich einheitlich bei allen Hirschen im 3.
Lebensmonat und dauert bis zum 9. Lebensmonat. Das würde im vorliegenden Fall bedeuten, dass
das Hirschkalb erst im September/Oktober geboren wurde und nicht wie üblich im Frühling zwischen
Mai und Juni. Diese Annahme bestätigt sich in den Werten des M3. Dessen Mineralisation begann ca.
im Juli, was etwa dem 9. Lebensmonats des Kalbs entsprechen würde und dauerte ca. bis im Februar.
Diese Phase ist eher zu kurz, da von einer Mineralisationsdauer von ca. 9-13 Monaten ausgegangen
werden kann (Brown 1991). Auch ist eine so stark verzögerte Geburt eher unwahrscheinlich, denn
Hirsche haben eine feste Brunftzeit im Herbst (s. Kapitel 2.3.1) die sich allenfalls bis in den Dezember
verlängern kann (mündl. Mitteil. Marguerita Schäfer, IPNA). Bei einer durchschnittlichen Tragezeit von
34 Wochen sollten die spätesten Geburten im August erfolgen. Allenfalls ist eine etwas verzögerte
Zahnentwicklung in Betracht zu ziehen, die zusammen mit einer späteren Geburt zu einer
aussergewöhnlich späten Mineralisation des M2 geführt hat. Ähnliche Beobachtungen wurden an der
rezenten Hirschpopulation von der Insel Rum (Schottland) gemacht (Stevens et al. 2011, 70-71). Die
Mineralisation des M2 von mehreren Hirschen dauerte ebenfalls von Januar bis in den Hochsommer,
also etwa vom 6. bis 12. Lebensmonat. Für die Insel Rum seien solche Verzögerungen in Wachstum
und Reproduktion aufgrund der kargen Umwelt- und Ernährungsbedingungen nicht ungewöhnlich
(Stevens et al. 2001, 71).
Beim zweiten beprobten M2 (RIZG799.1.2) scheint die Mineralisation ebenfalls im Sommer
abgeschlossen gewesen zu sein. Da es sich aber um ein sehr altes Tier handelt (ca. 10 jährig), sind
der Beginn und die Dauer der Mineralisation nicht mehr nachvollziehbar.
Bei der Betrachtung aller Sauerstoffwerte der M3 fällt auf, dass es eine sehr grosse Variabilität in den
Kurvenverläufen gibt. Da die Mineralisationsphase von 3. Molaren zwischen Individuen (und
Populationen) in ihrem Beginn und der Dauer
stark variieren kann, ist dies zu erwarten. Dazu
kommt, dass durch die verschiedenen Alter
der Hirsche unterschiedliche Abkauungsgrade
an den Zähnen vorhanden sind und somit die
repräsentierte Zeit der Isotopenkurven nicht
einheitlich ist. Auffallend ist aber, dass bei fünf
jungen Hirschen mit nahezu vollständigem
Zahnschmelz (ohne Abnutzung) ähnliche
Kurvenverläufe zu erkennen sind (Abb. 29;
1464.1.3.3 ist als Ausreisser zu betrachten).
Bei allen fünf Zähnen scheint die Mineralisation
im Frühling begonnen zu haben und bis im
darauffolgenden Winter/Frühling gedauert zu
haben. Bei einer Geburt im Mai/Juni hätte die
Mineralisation also ca. vom 12. bis 20./22.

52
Tab. 5
Mögliche Rückschlüsse auf den Lebensraum aufgrund der
Sauerstoff- und Kohlenstoffisotopenwerte (Höhen- und
Canopy-Effekt).
Lebensmonat gedauert. Dies würde mit den Beobachtungen von BROWN (1991) übereinstimmen und
auf eine „normale“ Zahnentwicklung hindeuten. Im Gegensatz zum Individuum 1846.1 decken sich
diese Beobachtungen gar nicht mit jenen von STEVENS ET AL. (2011) an den Schottischen Hirschen,
wo die Mineralisation der M3 bei allen Hirschen im Sommer begann und bis zum nächsten
Frühling/Frühsommer dauerte. Dies ist mit grosser Wahrscheinlichkeit auf die speziellen
Gegebenheiten auf der schottischen Insel zurückzuführen (s. oben). Das würde bedeuten, dass das
Klima am Zugersee zur horgener Zeit keine Verzögerung in der physischen Entwicklung der Hirsche
bewirkt hat. Eine Ausnahme könnte tatsächlich der Hirsch 1846.1 darstellen.
Bei den älteren Individuen ist aufgrund des Materialverlusts an der Krone durch die Abnutzung der
Zeitpunkt des Mineralisationsbeginns nicht mehr zu bestimmen. Die δ18
O-Werte deuten aber darauf
hin, dass das Ende der Mineralisationsphase sowohl im Winter als auch im Sommer liegen kann. Dies
ist vermutlich auf die natürliche Variation der Zahnentwicklung zurückzuführen.
Mit systematischer Beprobung von Zahnreihen – also M1, M2 und M3 vom gleichen Individuum –
könnten starke Verzögerungen in der Entwicklung wie bei Hirsch 1846.1 besser nachgewiesen
werden und eventuell Hinweise auf klimatische Einflüsse oder populationsspezifische Unterschiede
gefunden werden. Die vorliegenden Daten ermöglichen aber die Feststellung deutlicher Variationen
was den Zeitpunkt und die Dauer der Mineralisation der 3. Molaren von den horgenzeitlichen Hirschen
aus Zug-Riedmatt betrifft. Klare Gruppierungen sind nicht zu erkennen, was einen Rückschluss auf
populationsabhängige Unterschiede verunmöglicht.
6.2 Der Lebensraum – bewaldete Täler oder luftige Höhen?
Unterschiedliche Lebensräume können sich in den verschiedensten Isotopenmerkmalen bemerkbar
machen. Zwei Zusammenhänge zwischen Lebensraum und Isotopenzusammensetzung in
Knochen/Zähnen, die sich schon in sehr vielen Untersuchungen als konsistent erwiesen haben, sind
der Canopy-Effekt (δ13
C) und der Höhen/Temperatur-Effekt (δ13
C /δ18
O). Diese können sowohl einzeln
als auch in Kombination auftreten (Tab.
5). Die Bezeichungen „hoch“ und „niedrig“
müssen relativ betrachtet werden, da vor
allem die Sauerstoffwerte stark von
Region und Klima abhängig sind (siehe
Kapitel Grundlagen). Auch dürfen diese
Interpretationsmöglichkeiten nicht als
absolut angesehen werden sondern
können lediglich eine Tendenz darstellen,
da noch viele weitere Faktoren in die Isotopenzusammensetzungen einfliessen können.
Der Einfluss von dichter Bewaldung auf die Isotopenzusammensetzung von Flora und Fauna der
gemässigten Klimazone wurde mittlerweile in einigen Arbeiten sowohl an rezentem als auch
archäologischem Material bestätigt (z.B. Drucker et al. 2003; Bocherens/Drucker 2003; Drucker et al.
2008; Drucker et al. 2011; Balasse et al. 2012). Ausgehend von diesen Daten ist anzunehmen, dass
δ18
O hoch δ18
O niedrig
δ13
C niedrig Im Flachland
gelegene Waldgebiete
Höher gelegene
Waldgebiete
δ13
C hoch
Offene Landschaft im
Flachland, Lichtungen
oder Felder/Äcker
Offenland in höheren
Lagen, ev. über der
Baumgrenze

53
Abb. 30
Mögliche δ13
C-Werte der aufgenommenen Nahrung der Hirsche aus Zug-Riedmatt. Berechnet aus den
maximalen Kohlenstoffisotopenwerten aus dem Zahnschmelz und Dentin von allen Hirschen.
die Nahrung von Herbivoren unter einem geschlossenen Blätterdach (dichter Wald) einen maximalen
δ13
C-Wert von -26.5 – 28.4‰ aufweisen sollte (Drucker et al. 2008; Drucker et al. 2011; Balasse et al.
2012).
Da für die Hirsche von Zug-Riedmatt Kohlenstoffisotopen-Daten sowohl aus dem Kollagen (δ13
Ckol) als
auch aus dem Karbonat (δ13
Ckar) vorhanden sind, kann aus beiden Quellen der ungefähre
Isotopenwert der Nahrung berechnet werden. Für das Kollagen wird eine Anreicherung von 5.1‰
(berechneter Durchschnittswert von Drucker et al. 2008, 72) und für das Karbonat im Zahnschmelz
eine Anreicherung von 14.1‰ (Cerling/Harris 1999) angenommen. Werden diese Werte von den
maximalen δ13
C-Werten aus Kollagen und Karbonat der Hirsche aus Zug-Riedmatt abgezogen,
resultieren ausschliesslich Kohlenstoffwerte unter -27‰ (Abb. 30). Dies deutet darauf hin, dass sich
die Hirsche vor allem oder ausschliesslich in bewaldetem Gebiet aufgehalten haben und vermutlich
auch in selbigem gejagt wurden.
Die Anreicherung von 5.1 bzw. 14.1‰ darf aber nicht als sicher angesehen werden - es gibt auch
Quellen, die von niedrigeren Werten ausgehen (z.B. Ambrose/Norr 1993, Rodière et al. 1996, Passey
et al. 2005). Einige Individuen aus Zug-Riedmatt würden sich dann wohl eher in einem Mischbereich
zwischen Wald und Offenland bewegen. Besonders die beiden Hirsche 1348.1 und 1342.1 (Abb. 30,
rot eingekreist) würden in diesen kritischen Bereich fallen – ihre maximalen δ13
Ckol-Werte liegen mit -
22.3 und -22.2‰ knapp über dem von DRUCKER ET AL. (2003) definierten Bereich für waldlebende
Individuen (Drucker et al. 2003, 381). Ebenfalls hoch aber nicht über -22.5‰ liegen die δ13
Ckol-Werte
der beiden Hirsche 1763.1 und 1763.2 (Abb 30, blau eingekreist).
Betrachtet man nun die δ18
O-Werte dieser vier Individuen, ist keine gemeinsame Auffälligkeit in deren
Höhe festzustellen. Würde man von einem eher offenen Lebensraum dieser vier Hirsche aufgrund
einer erhöhten Lage (über der Waldgrenze) ausgehen, so müssten die Sauerstoffisotopenwerte
deutlich niedriger sein als der Durchschnitt. Dies trifft am ehesten auf Individuum 1763.2 zu, dessen
δ18
O-Werte mit -11.9 (Minimum) und -10.1 (Maximum) niedrig ausfallen (Abb. 19). Auch die tiefen
δ15
N-Werte desselben Tieres (Abb. 23) könnten als Hinweis für einen latidudinal erhöhten
Lebensraum mit tieferen Temperaturen und geringerer Vegetationsaktivität angesehen werden.
-32
-31
-30
-29
-28
-27
-26
-25
-24
δ13
C (‰
) N
ah
run
g
δ13CkolMax - 5.1‰
δ13CkarMax - 14.1‰
O f f e n l a n d
d i c h t e r W a l d

54
Insgesamt ist anzunehmen, dass die meisten Hirsche in stark bewaldetem Gebiet gelebt haben.
Natürlich ist nicht auszuschliessen, dass auch Nahrungsquellen in offeneren Landschaften genutzt
wurden (Gras, Getreide auf Feldern, Büsche und Sträucher am Waldrand), was einzelne erhöhte
δ13
C-Werte hervorrufen kann. Den Grossteil ihrer Nahrung haben die Hirsche aber im Wald gesucht.
Da kein klarer Zusammenhang zwischen dem Verlauf der Sauerstoff- und Kohlenstoffisotopenkurven
zu erkennen ist (Abb. 19-21), ist auch eine saisonal bedingte Änderung von Ernährung/Aufenthaltsort
nicht nachzuweisen.
Da die Sauerstoffisotopenzusammensetzung im Zahnschmelz hauptsächlich durch die
Isotopenverhältnisse im Trinkwasser beeinflusst wird, lassen sich Rückschlüsse auf die
Trinkwasserquelle der Tiere ziehen. In grossen Gewässern wie Seen oder Flüssen und im
Grundwasser vermischen sich diese Signale und ergeben einen Durchschnittswert aus
unterschiedlichen Zeiträumen. Würde ein Tier sein Trinkwasser nun hauptsächlich aus diesen Quellen
beziehen, würde das saisonale Signal stark abgeschwächt oder verfälscht (Fricke et al. 1996, 98). Bei
einer starken Ausprägung der saisonalen Sauerstoffisotopenschwankungen wie im vorliegenden Fall
bei den Hirschen von Zug-Riedmatt ist also davon auszugehen, dass sie vor allem
Niederschlagswasser aus Pfützen oder Bächen bzw. aus Pflanzen aufgenommen haben.
Aufgrund der unterschiedlichen Amplituden (0.8‰ – 4.5‰) der δ18
O-Werte von einzelnen Individuen
ist eine abschwächende Wirkung von Seewasser auf die saisonalen Schwankungen nicht
auszuschliessen. Vor allem der Hirsch 1244.1 fällt durch sehr einheitliche Werte (Amplitude 0.8‰) auf.
6.3 Klimatische Bedingungen
Da sowohl die absoluten δ18
O-Werte als auch die Daten des rezenten Niederschlagswassers nicht
verlässlich sind (Shift von 1.3‰ zwischen Mainzer und Basler Messung) bzw. überprüft werden
müssten, ist von einer Rekonstruktion der Isotopenverhältnisse im Trinkwasser der Hirsche oder gar
einer Jahresmitteltemperaturrekonstruktion abzusehen (Bsp. Sevens et al. 2011, 68). Die
Sauerstoffisotopendaten können lediglich relativ miteinander verglichen werden und die saisonalen
Verläufe mit den Kohlenstoffdaten kombiniert werden. Dies ermöglicht Rückschlüsse auf den
Zusammenhang von Temperatur/Jahreszeit und Trockenheit/Niederschlag, welche durch die
zusätzliche Betrachtung der δ15
N-Werte unterstützt werden könnten. Idealerweise würden dafür die
Isotopenwerte aus dem Zahnschmelz und dem Dentin auf einer Skala aufgetragen (z.B. Entfernung
von der Schmelzgrenze in mm), sodass man die Werte aus dem Zahnmaterial, das ungefähr im
gleichen Zeitraum gebildet wurde, miteinander verknüpfen könnte. Problematisch bliebe aber die
Frage, welche Zeitspanne zwischen der Bildung des Dentins und des darüber liegenden
Zahnschmelzes liegt. Eine korrekte Korrelation der Isotopenwerte aus Dentin und Zahnschmelz dürfte
also schwierig bleiben.
Wie weiter oben schon erwähnt, ist die grosse Variabilität der δ18
O-Kurvenverläufe und der
Verhältnisse der Sauerstoff- und Kohlenstoffisotopenkurven zueinander auffallend. Bei 10 Hirschen
verlaufen die Kurven mehr oder weniger gleichläufig (Abb. 19). Geht man nun davon aus, dass diese
Hirsche in einem ähnlichen Lebensraum (vorwiegend Wälder in nicht allzu grosser Höhe) gelebt

55
haben, so wäre die Saisonalität der δ13
C-Werte eventuell mit einer grossen Schwankung der
Niederschlagsmengen bzw. Luftfeuchtigkeit zu erklären. Da bei Trockenheit der Anteil an schwerem
13C steigt, würden die gleichläufigen Kurven auf trockene Sommer und nasse Winter hindeuten.
Umgekehrt bei den vier Individuen mit gegenläufigen Kurven (Abb. 20). Dort korrelieren die tiefen
Kohlenstoffwerte mit hohen Sauerstoffwerten, was auf grosse Feuchtigkeit im Sommer und
Trockenheit im Winter zurückzuführen sein könnte. Bei neun weiteren Hirschen ist die Tendenz unklar
bzw. die Kurven verlaufen teilweise gegenläufig und gleichläufig. Dies schliesst aber eine
Abhängigkeit der δ13
C-Werte von der Niederschlagsmenge/Luftfeuchtigkeit nicht aus. Denkbar wäre
auch ein Aufenthalt der Hirsche in feuchteren oder trockeneren Gebieten, z.B. See-/Flussufern und
Talsenken oder trockeneren Stellen wie Hügelkuppen.
6.4 Unterschiede zwischen den Siedlungsphasen – ist eine Entwicklung
zu erkennen?
Sauerstoff-Werte
Die δ18
O-Wertespannen nach Siedlungsschicht aufgetrennt zeigen eine regelmässige Verteilung
(Abb. 17). Innerhalb jeder Siedlungsphase ist eine grosse Variation an Werten vorhanden, sogar in
Schicht B und C1 mit nur drei bzw. vier beprobten Individuen.
Die gejagten Hirsche stammen möglicherweise aus unterschiedlichen Höhenlagen oder nutzten
verschiedene Wasserquellen. Eine signifikante Veränderung der Jahresdurchschnittstemperaturen im
Verlaufe der Besiedlung von Zug-Riedmatt ist nicht zu erkennen, weil in diesem Fall davon
ausgegangen werden müsste, dass sich die δ18
O-Werte erkennbar nach oben oder nach unten
entwickeln würden. Es ist also anzunehmen, dass die Wertebereichvariation auf lokale und/oder
kurzfristige Unterschiede bzw. Veränderungen zurückzuführen sind, welche in allen Siedlungsphasen
eine ähnliche Variation der Sauerstoffisotopenkonzentrationen in den Zähnen der Hirsche auslöste.
Kohlenstoff-Werte
Sowohl bei den δ13
C-Werten aus dem Karbonat als auch bei jenen aus dem Kollagen sind keine
signifikanten Unterschiede zwischen den Siedlungsschichten zu erkennen. Die meisten Werte
bewegen sich innerhalb von ca. 2.5‰, mit nur wenigen Ausreissern (Abb. 18 und Abb. 22). Dabei ist
die Schicht C1 die einzige Phase, deren δ13
C-Werte aus dem Kollagen -23.8‰ nicht übersteigen und
somit keine Hinweise auf einen Aufenthalt der Hirsche ausserhalb des Waldes liefern. Der
Lebensraum scheint also einheitlicher gewesen zu sein als in den vorhergehenden Phasen B-B2, was
zu einer geringeren Variation der δ13
C-Werte geführt hat.
Stickstoff-Werte
Zwischen der ältesten Schicht B und der jüngeren Schicht C1 gibt es einen deutlichen Unterschied der
Wertebereiche der δ15
N-Werte (Abb. 23). Während sie in Schicht B zwischen 4.1 und 5.7‰ liegen,
bewegen sich diejenigen in Schicht C1 zwischen 5.4 und 6.6‰. Dies könnte als eine Verbesserung
des Klimas im Verlaufe der Besiedlung von Zug-Riedmatt gewertet werden, aufgrund der geringen
Probenzahl ist dies jedoch problematisch. In den dazwischenliegenden Schichten B1 und B2 mit acht

56
bzw. sieben beprobten Individuen bewegen sich die Werte über die gesamte Spanne von 4.2 bis
6.8‰.
Der Verlauf der Kurven
Weder bei den Isotopenwerten aus dem Karbonat noch bei jenen aus dem Kollagen ist ein
Zusammenhang zwischen Kurvenverlauf und Phasenzugehörigkeit auszumachen. Sowohl bei den
gleichläufig verlaufenden Kurven (Abb. 19 und Abb. 24) als auch bei den gegenläufig verlaufenden
Kurven (Abb. 20 und Abb. 25) sind Zähne aus allen Siedlungsschichten vertreten. Innerhalb der
einzelnen Isotopengruppen (O, C, N) fehlt ebenfalls jeglicher Zusammenhang zwischen auf- und
absteigenden Kurven und der Phasenzugehörigkeit.
Insgesamt lassen sich zwischen den Siedlungsphasen nur geringfügige Unterschiede zwischen den
Isotopenwerten und deren Variation erkennen. Klare Hinweise auf eine Klimaveränderung oder ein
verändertes Jagdverhalten der Menschen aus Zug-Riedmatt sind nicht gegeben.
Aufgrund der geringen Probenmengen aus Schicht C1 und D1/D2 ist die Interpretation der
einheitlicheren (tiefen) δ13
C-Werten und den ähnlich hohen δ15
N-Werten aus dem Zahnkollagen dieser
beiden Phasen vorsichtig zu formulieren. Dazu kommt, dass die hohen C/N-Werte des M1 – dem
einzigen zur Verfügung stehenden Hirschzahn aus Schicht D1/D2 – mit hoher Wahrscheinlichkeit auf
ein Stillsignal zurückzuführen sind und somit nicht in die Auswertung miteinbezogen werden dürfen.
6.5 Synthese
Aus den Isotopenanalysen der Hirschzähne aus der Fundstelle Zug-Riedmatt geht eine sehr grosse
Variation an Sauerstoff-, Kohlenstoff- und Stickstoffwerten hervor. Anhand der Wertebereiche, der
Kurvenverläufe sowie aus der Kombination der Messungen können folgende Aussagen gemacht
werden:
Die saisonal geprägten Kurven der Sauerstoffisotopenwerte im Zahnschmelz weisen eine
hohe Variabilität auf. Zumindest für einen Teil der Hirsche kann aber ein normales
Zahnwachstum des 3. Molaren angenommen werden, wie es auch bei rezenten Hirschen in
gemässigten Klimazonen üblich ist. Das heisst, der Geburtszeitpunkt lag etwa im Mai/Juni und
die körperliche Entwicklung der Jungtiere verlief durchschnittlich.
Bei einem Individuum (RIZG1846.1) aus dem Knochenhaufen in Schicht B1 finden sich in den
Isotopensignalen im 2. und 3. Molar Hinweise auf eine möglicherweise verspätete Geburt oder
verzögerte Entwicklung des Zahnwachstums. Dies könnte die Folge von schwierigen
Lebensbedingungen wie Nahrungsknappheit, Stress oder Klimaverschlechterung sein.
Aufgrund der durchgehend tiefen Kohlenstoffwerte kann angenommen werden, dass sich die
Hirsche hauptsächlich in bewaldetem Gebiet aufgehalten haben. Die grosse Variation der
Werte aus Zahnschmelz und Dentin zeigen aber, dass die Zusammensetzung der Nahrung
nicht einheitlich war, was u.a. mit unterschiedlichen Habitaten zusammenhängen kann.

57
Auffällig sind die Werte des Hirsches RIZG1763.2 (Knochenhaufen, B1), bei dem die
Kohlenstoffwerte eher hoch und die Sauerstoff- sowie Stickstoffwerte auffallend gering
ausfallen. Für dieses Individuum kann ein aussergewöhnlicher Lebensraum in eher offenem,
kargem und kühlerem Gebiet wie einer Hügelkuppe (z.B. Zugerberg) postuliert werden.
Die Wertebereiche der Kohlenstoff- und Stickstoffisotope aus dem Kollagen des Dentins
unterscheiden sich in Schicht C1 von denjenigen in Schicht B-B2. Die Kohlenstoffwerte der
vier Individuen sind einheitlich tief, während sich die Stickstoffwerte in einem oberen Bereich
bewegen. Dies könnte auf eine Klimaberuhigung bzw. Temperaturanstieg im Laufe der
Besiedlung von Zug-Riedmatt hindeuten, was ein engräumigeres Einzugsgebiet der
Jagdgründe zur Folge hatte.
Geht man von der Hypothese aus, dass das Jagdverhalten der Bevölkerung von Zug-Riedmatt
zumindest teilweise von klimatischen Veränderungen beeinflusst wurde, so ist in den Isotopendaten
der Hirschzähne zumindest kein Wiederspruch dazu zu finden.
Aus den ältesten Schichten mit sehr hohem Anteil an Wildtier- bzw. Hirschknochen sind die
Isotopenwerte sowie der saisonale Verlauf der Kurven sehr unterschiedlich. Dies kann auf
verschiedene Populationen, unterschiedliche Lebensräume sowie auf klimatische Unterschiede
zurückzuführen sein. Tatsächlich finden sich an zwei Individuen Anzeichen für klimatische Einflüsse –
einerseits eine verzögerte körperliche Entwicklung und andererseits Isotopenwerte, die auf ein Leben
unter relativ wiedrigen Umständen zurückzuführen sein könnten. Durch die Tatsache, dass aus diesen
Schichten die meisten Proben stammen, könnte die auffallend grosse Variabilität aber auch
methodischen Ursprungs sein.
Aus Schicht C1 stammen nur vier beprobte Individuen, da hier der Anteil von Wildtieren im
Knochenspektrum schon wesentlich geringer ist als in den älteren Schichten. Diese Hirsche scheinen
alle aus dem gleichen Lebensraum zu kommen, in der die Vegetation sehr aktiv war – also gute
klimatische Verhältnisse herrschten. Dies könnte gleichzeitig der Grund für eine verminderte
Jagdtätigkeit sein, da die Bevölkerung durch bessere Ernteerträge nicht mehr im gleichen Masse auf
Jagderfolge angewiesen war wie in den vorangegangenen Jahren oder Generationen.
Um die Hypothese einer sukzessiven Klimaverbesserung während der Besiedlung von Zug-Riedmatt
zu erhärten, wären weitere Isotopenanalysen an Hirschknochen aus den Siedlungsphasen D und E
notwendig. Da keine Hirschzähne aus diesen Schichten vorhanden sind, müsste auf die Kohlenstoff-
und Stickstoffisotopenverhältnisse in den Knochen ausgewichen werden. Es ist aber anzunehmen,
dass eine Klimaveränderung an den Werten erkennbar wäre.
Grundlegend für weitere Untersuchungen sind erneute Messungen und die Überprüfung der
Sauerstoffisotopenwerte. Nur so könnte eine Entwicklung der Jahresdurchschnittstemperaturen
sichtbar gemacht werden – ein wichtiger Faktor für die Klima- und Lebensraumrekonstruktion.
Erstrebenswert wären zudem Vergleichswerte von Hirschzähnen aus möglichst zeitnahen
Seeufersiedlungen des Alpenvorlandes.

58
Zur genaueren Bestimmung des Lebensraumes und der saisonalen Bewegungen der Hirsche wäre
die Durchführung von Strontiumisotopenanalysen am Zahnschmelz hilfreich. Damit könnten
Unterschiede und Variabilität im geologischen Untergrund des Habitats der Hirsche erkannt und mit
den vorliegenden Daten kombiniert werden.
7. Zusammenfassung
In der vorliegenden Forschungsarbeit an der Universität Basel wurden Isotopenanalysen (δ18
O, δ13
C,
δ15
N) an 23 Zähnen von 22 Hirschen aus der horgenzeitlichen Seeufersiedlung Zug-Riedmatt
durchgeführt. Diese stammen aus verschiedenen Siedlungsphasen, die einen nicht genau zu
definierenden Zeitraum zwischen 3200 und 3000 v. Chr. abdecken. Da die Schichtpakete eine klar
chronologische Abfolge haben war das Ziel, anhand der Isotopendaten eine Veränderung im Laufe
der Besiedlung von Zug-Riedmatt zu erkennen. Die archäozoologische Auswertung ergab, dass in
den ältesten Schichten die Anteile der Wildtiere (v.a. Hirsch) im Knochenspektrum markant höher
liegen als in den darüber liegenden, jüngeren Phasen. Daraus lässt sich ableiten, dass die Jagd auf
den Rothirsch zu Beginn der Besiedlung sehr intensiv war und in der darauffolgenden Zeit
kontinuierlich an Bedeutung verlor. Mittels der Analyse von Sauerstoff- und Kohlenstoffisotopen am
Zahnschmelz sowie von Kohlenstoff- und Stickstoffisotopen am Dentin der Hirschzähne sollten
Informationen über die Herkunft, den Lebensraum und die Ernährung der Tiere gewonnen und so
Rückschlüsse auf das Jagdgebiet der Bevölkerung von Zug-Riedmatt ermöglicht werden.
Durch die sequenzierte Beprobung der Zähne wurden saisonale Entwicklungen sichtbar, welche die
Isotopenzusammensetzung im Zahn während des Zeitraumes seiner Bildung beeinflusst haben. Dabei
zeigte sich, dass die Variabilität der Isotopenwerte sowohl innerhalb als auch zwischen den Individuen
sehr gross ist und eine Vielzahl an Interpretationsmöglichkeiten zulässt, welche jedoch beim
momentanen Stand der Forschung nur schwer verifiziert werden können. Dennoch konnte die
Hypothese, dass die nachlassende Jagdtätigkeit im Laufe der Besiedlung von Zug-Riedmatt mit einer
Klimaverbesserung einhergeht und zu einem kleineren Einzugsgebiet des Jagdrevieres führte,
zumindest ansatzweise bestätigt werden. Ein Hinweis darauf ist die geringe Spannweite der
Kohlenstoff- und Stickstoffwerte in der jüngeren Schicht C1, was auf eine lokalere Herkunft der
Hirsche hindeuten könnte. Zudem steigen die Stickstoffwerte im Vergleich zu den älteren Phasen
leicht an – dies könnte die Folge einer klimatischen Verbesserung sein.
Mit diesem Pilotprojekt an der Universität Basel konnte aufgezeigt werden, dass die Analyse von
stabilen Isotopen an Knochen- und Zahnmaterial aus neolithischen Seeufersiedlungen grosses
Potential zur Ergänzung der traditionellen archäologischen und archäozoologischen Methoden
darstellen kann. Das Ziel weiterer Forschung sollte eine breite und umfangreiche Sammlung von
Isotopendaten aus möglichst vielen Fundstellen sein, um Vergleiche zwischen Tierarten, Regionen
und Epochen zu ermöglichen und so die Interpretierbarkeit von jeder einzelnen Untersuchung zu
verbessern.

59
Tab. 6
Vollständige Übersicht der δ18
O- und δ13
C-Werte aus dem Karbonat des Zahnschmelzes der Hirsche
aus Zug-Riedmatt. Messungen von Basel und Mainz.
8. Tabellen
Schicht Individuum Zahn Sequenz
Basel Mainz
δ18
O[‰] V-PDB
δ13
C[‰] V-PDB
δ18
O[‰] V-PDB
δ13
C[‰] V-PDB
B
799.1 M2 799.1.2.1 -8.94 -15.45
799.1.2.2 -8.60 -15.08
799.1.2.3 -7.61 -14.08
799.1.2.4 -6.43 -13.46
1890.1 M3 1890.1.3.1 -9.92 -15.21
1890.1.3.2 -7.28 -14.87
1890.1.3.3 -8.02 -14.67
1890.1.3.4 -9.42 -14.48
1890.1.3.5 -8.60 -14.60
1890.1.3.6 -9.44 -15.09
1890.1.3.7 -10.11 -15.55
1890.1.3.8 -11.75 -16.24
1901.1 M3 1901.1.3.1 -8.78 -15.97
1901.1.3.2 -9.66 -15.91
1901.1.3.3 -9.91 -15.94
1901.1.3.4 -11.04 -15.87
1901.1.3.5 -10.99 -15.70
1901.1.3.6 -11.32 -15.70
1901.1.3.7 -10.87 -15.63
1901.1.3.8 -10.59 -15.56
B1
1763.1 M3 1763.1.3.1 -8.40 -13.81
1763.1.3.2 -7.25 -13.04
1763.1.3.3 -6.29 -13.16
1763.1.3.4 -7.19 -14.21
1763.1.3.5 -7.92 -14.49
1763.1.3.6 -10.33 -14.89
1763.2 M3 1763.2.3.1 -10.84 -14.41
1763.2.3.2 -11.09 -14.71
1763.2.3.3 -11.92 -14.93
1763.2.3.4 -10.11 -14.51
1776.1 M3 1776.1.3.1 -7.97 -15.34
1776.1.3.2 -8.38 -15.31
1776.1.3.3 -8.88 -15.40
1776.1.3.4 -8.70 -15.42
1776.1.3.5 -9.41 -15.83
1776.1.3.6 -9.77 -15.66
1846.1 M2 1846.1.2.1 -12.43 -16.03 -12.5 -17.1
1846.1.2.2 -11.52 -15.66 -12.3 -17.0
1846.1.2.3 -9.54 -15.73 -11.1 -16.0
1846.1.2.4 -8.53 -15.98 -11.2 -15.8
1846.1.2.5 -8.22 -15.89 -10.1 -15.3
1846.1.2.6 -8.18 -16.23 -9.7 -15.0
M3 1846.1.3.1 -8.42 -15.95 -9.8 -15.7
1846.1.3.2 -8.46 -15.39 -9.9 -15.5
1846.1.3.3 -9.81 -15.58 -10.6 -15.6
1846.1.3.4 -7.81 -15.96 -10.7 -15.8
1846.1.3.5 -10.20 -15.63 -10.8 -15.8
1846.1.3.6 -10.85 -15.76 -11.6 -15.9
1846.1.3.7 -11.76 -15.94 -12.2 -16.1
1846.1.3.8 -10.38 -15.97 -12.5 -16.1
1846.1.3.9 -11.15 -15.66 -12.2 -15.7
1846.2 M3 1846.2.3.1 -8.22 -16.01 -9.6 -16.1
1846.2.3.2 -8.71 -15.82 -10.1 -15.9
1846.2.3.3 -9.34 -15.59 -10.4 -15.8
1846.2.3.4 -9.51 -15.69 -10.8 -15.7
1846.2.3.5 -9.86 -15.34 -11.2 -15.4
1846.2.3.6 -10.13 -15.09 -11.5 -15.2
1846.2.3.7 -9.85 -15.52 -11.2 -15.5
1846.2.3.8 -9.54 -15.36 -11.0 -15.4

60
Tab. 6 (Fortsetzung)
Vollständige Übersicht der δ18
O- und δ13
C-Werte aus dem Karbonat des Zahnschmelzes der Hirsche
aus Zug-Riedmatt. Messungen aus Basel und Mainz.
Schicht Individuum Zahn Sequenz
Basel Mainz
δ18
O[‰] V-PDB
δ13
C[‰] V-PDB
δ18
O[‰] V-PDB
δ13
C[‰] V-PDB
B1
1846.3 M3 1846.3.3.1 -10.15 -16.78
1846.3.3.2 -8.29 -16.44
1846.3.3.3 -9.48 -16.67
1846.3.3.4 -10.00 -16.50
1846.3.3.5 -10.09 -16.65
1846.3.3.6 -10.62 -16.46
1846.3.3.7 -11.02 -15.87
1846.4 M3 1846.4.3.1 -8.91 -16.48
1846.4.3.2 -8.37 -16.62
1846.4.3.3 -8.27 -16.46
1846.4.3.4 -9.51 -15.34
1846.4.3.5 -8.89 -15.26
1846.4.3.6 -7.79 -14.88
B2
1342.1 M3 1342.1.3.1 -8.96 -15.34 -10.2 -15.5
1342.1.3.2 -8.53 -15.19 -9.8 -15.3
1342.1.3.3 -7.99 -14.89 -9.3 -15.1
1342.1.3.4 -7.46 -14.60 -9.1 -14.8
1342.1.3.5 -7.80 -14.44 -9.0 -14.6
1342.1.3.6 -7.60 -14.24 -8.8 -14.4
1342.2 M3 1342.2.3.1 -7.94 -16.00
1342.2.3.2 -8.54 -15.74
1342.2.3.3 -8.68 -15.32
1342.2.3.4 -7.10 -15.60
1342.2.3.5 -6.67 -15.35
1342.2.3.6 -5.70 -14.41
1348.1 M3 1348.1.3.1 -9.49 -15.58
1348.1.3.2 -8.17 -15.32
1348.1.3.3 -7.90 -15.19
1348.1.3.4 -7.51 -15.06
1348.1.3.5 -7.89 -14.87
1348.1.3.6 -7.64 -14.67
1348.2 M3 1348.2.3.1 -7.41 -16.38
1348.2.3.2 -7.70 -16.35
1348.2.3.3 -7.47 -16.26
1348.2.3.4 -6.57 -16.17
1348.2.3.5 -7.50 -16.13
1348.2.3.6 -9.62 -15.84
1348.3 M3 1348.3.3.1 -7.85 -15.92
1348.3.3.2 -8.57 -15.13
1348.3.3.3 -9.93 -14.94
1348.3.3.4 -10.21 -15.35
1348.3.3.5 -10.18 -14.93
1348.3.3.6 -11.24 -15.30
1464.1 M3 1464.1.3.1 -8.59 -15.28
1464.1.3.2 -8.06 -15.00
1464.1.3.3 -9.29 -14.86
1464.1.3.4 -8.25 -14.87
1464.1.3.5 -9.15 -15.12
1464.1.3.6 -9.34 -15.42
1464.1.3.7 -9.22 -15.98
1535.1 M3 1535.1.3.1 -9.20 -16.47
1535.1.3.2 -9.73 -16.63
1535.1.3.3 -9.37 -16.36
1535.1.3.4 -8.75 -16.23
1535.1.3.5 -8.50 -15.92

61
Tab. 6 (Fortsetzung)
Vollständige Übersicht der δ18
O- und δ13
C-Werte aus dem Karbonat des Zahnschmelzes der Hirsche
aus Zug-Riedmatt. Messungen aus Basel und Mainz.
Schicht Individuum Zahn Sequenz
Basel Mainz
δ18
O[‰] V-PDB
δ13
C[‰] V-PDB
δ18
O[‰] V-PDB
δ13
C[‰] V-PDB
C1
952.1 M3 952.1.3.1 -9.21 -14.91
952.1.3.2 -7.87 -14.42
952.1.3.3 -7.25 -14.22
952.1.3.4 -7.42 -14.55
952.1.3.5 -8.98 -15.02
952.1.3.6 -10.09 -15.29
952.1.3.7 -10.23 -15.33
1067.1 M3 1067.1.3.1 -7.33 -15.44
1067.1.3.2 -6.58 -15.18
1067.1.3.3 -7.50 -15.67
1067.1.3.4 -7.64 -16.15
1067.1.3.5 -7.43 -15.94
1238.1 M3 1238.1.3.1 -10.16 -15.83
1238.1.3.2 -11.32 -15.50
1238.1.3.3 -10.21 -14.96
1238.1.3.4 -10.43 -14.71
1238.1.3.5 -10.98 -14.33
1238.1.3.6 -9.94 -14.50
1244.1 M3 1244.1.3.1 -9.57 -16.50 -10.8 -16.5
1244.1.3.2 -9.86 -16.27 -10.8 -16.0
1244.1.3.3 -10.00 -15.81 -11.5 -15.6
1244.1.3.4 -10.40 -15.02 -11.6 -15.1
1244.1.3.5 -10.33 -14.61 -11.4 -14.7
D1/D3
710.1 M1 710.1.1.1 -5.91 -15.62
710.1.1.2 -6.05 -15.77
710.1.1.3 -6.03 -15.07
710.1.1.4 -6.73 -14.77
710.1.1.5 -7.56 -14.86
710.1.1.6 -9.32 -15.25

62
Tab. 7
Vollständige Übersicht der δ13
C- und δ15N-Werte aus dem Kollagen des Dentins der Hirsche aus
Zug-Riedmatt. Messungen aus Basel. Anteile von Kohlenstoff und Stickstoff im Kollagen in % sowie
ihr Verhältnis (C:N).
Schicht Individuum Zahn Sequenz d13
C d15
N %C %N C:N
B
799.1 M2 799.1.2.1 -23.18 4.80 42.52 15.03 3.30
799.1.2.2 -23.04 4.76 41.79 14.82 3.29
799.1.2.3 -22.78 4.72 45.41 16.09 3.29
799.1.2.4 -22.52 4.74 44.93 15.93 3.29
799.1.2.5 -22.65 4.54 44.19 15.61 3.30
1890.1 M3 1890.1.3.1 -23.33 4.14 44.71 15.99 3.26
1890.1.3.2 -24.00 4.50 45.24 16.15 3.27
1890.1.3.3 -24.48 4.45 43.70 15.54 3.28
1890.1.3.4 -24.07 4.51 44.64 15.89 3.28
1901.1 M3 1901.1.3.1 -24.37 4.72 41.60 14.93 3.25
1901.1.3.2 -24.79 4.76 44.14 15.59 3.30
1901.1.3.3 -24.45 5.10 44.06 15.62 3.29
1901.1.3.4 -24.11 5.65 43.69 15.51 3.29
1901.1.3.5 -24.08 5.74 42.77 15.14 3.30
B1
1763.1 M3 1763.1.3.1 -22.79 5.15 42.41 15.03 3.29
1763.1.3.2 -22.87 5.70 44.12 15.60 3.30
1763.1.3.3 -23.06 6.77 42.02 14.96 3.28
1763.2 M3 1763.2.3.1 -22.91 4.47 42.13 14.87 3.30
1763.2.3.2 -22.88 4.51 44.91 15.91 3.29
1763.2.3.3 -22.72 4.87 45.10 16.08 3.27
1763.2.3.4 -22.49 4.86 44.22 15.78 3.27
1763.2.3.5 -22.48 4.81 44.02 15.36 3.34
1776.1 M3 1776.1.3.1 -24.31 6.28 42.62 15.16 3.28
1776.1.3.2 -24.36 6.53 43.04 15.37 3.26
1776.1.3.3 -24.55 6.59 44.04 15.85 3.24
1776.1.3.4 -24.51 6.72
1776.1.3.5 -24.03 6.59 42.72 15.18 3.28
1846.1 M2 1846.1.2.1 -25.97 6.15 42.93 15.17 3.30
1846.1.2.2 -25.61 6.71 42.85 15.35 3.26
1846.1.2.3 -24.16 7.17
1846.1.2.4 -24.94 7.18 42.98 15.16 3.31
1846.1.2.5 -24.65 7.50 43.37 15.37 3.29
1846.1.2.6 -24.88 6.39 43.60 15.32 3.32
M3 1846.1.3.1 -24.91 6.74 42.00 14.92 3.28
1846.1.3.2 -24.84 6.61 41.89 14.82 3.30
1846.1.3.3 -24.25 6.39 44.76 15.93 3.28
1846.1.3.4 -23.91 6.44 44.78 15.93 3.28
1846.1.3.5 -23.45 6.10 43.77 15.48 3.30
1846.1.3.6 -23.53 5.88 43.16 15.25 3.30
1846.2 M3 1846.2.3.1 -24.55 4.99 44.52 15.89 3.27
1846.2.3.2 -24.53 5.10 43.70 15.47 3.29
1846.2.3.3 -24.43 5.45 41.41 14.84 3.25
1846.2.3.4 -24.38 5.79 34.79 12.32 3.29
1846.2.3.5 -24.15 5.82 40.75 14.39 3.30
1846.3 M3 1846.3.3.1 -24.85 6.43 43.77 15.64 3.26
1846.3.3.2 -24.64 6.39 45.01 15.93 3.29
1846.3.3.3 -24.25 6.35 45.00 16.11 3.26
1846.3.3.4 -23.89 6.39 44.88 16.01 3.27
1846.3.3.5 -23.76 6.28 43.74 15.49 3.29
1846.4 M3 1846.4.3.1 -24.73 4.87 42.37 15.03 3.29
1846.4.3.2 -24.66 4.77 42.82 15.21 3.28
1846.4.3.3 -24.29 5.29 44.18 15.66 3.29
1846.4.3.4 -24.08 5.18 43.89 15.61 3.28
1846.4.3.5 -24.31 4.75 43.90 15.49 3.30

63
Tab. 7 (Fortsetzung)
Vollständige Übersicht der δ13
C- und δ15N-Werte aus dem Kollagen des Dentins der Hirsche aus
Zug-Riedmatt. Messungen aus Basel. Anteile von Kohlenstoff und Stickstoff im Kollagen in % sowie
ihr Verhältnis (C:N).
Schicht Individuum Zahn Sequenz d13
C d15
N %C %N C:N
B2
1342.1 M3 1342.1.3.1 -23.50 5.13 42.81 15.30 3.26
1342.1.3.2 -23.50 5.31 42.65 15.09 3.30
1342.1.3.3 -23.11 5.25 45.57 16.26 3.27
1342.1.3.4 -22.66 5.58 44.86 15.96 3.28
1342.1.3.5 -22.52 5.65 45.24 16.03 3.29
1342.1.3.6 -22.30 5.36 44.86 15.92 3.29
1342.2 M3 1342.2.3.1 -24.67 4.19 41.66 14.85 3.27
1342.2.3.2 -24.42 4.39 45.02 16.07 3.27
1342.2.3.3 -23.79 4.84 43.74 15.62 3.26
1342.2.3.4 -23.57 5.10 43.88 15.59 3.28
1342.2.3.5 -23.46 4.59 43.99 15.60 3.29
1348.1 M3 1348.1.3.1 -23.54 4.86 42.94 15.36 3.26
1348.1.3.2 -23.42 5.24 44.56 15.95 3.26
1348.1.3.3 -22.96 5.29 45.10 16.05 3.28
1348.1.3.4 -22.54 5.59 44.67 15.78 3.30
1348.1.3.5 -22.39 5.79 43.93 15.66 3.27
1348.1.3.6 -22.18 5.61 43.93 15.70 3.26
1348.2 M3 1348.2.3.1 -24.95 5.13 44.00 15.76 3.26
1348.2.3.2 -24.71 5.36 44.06 15.62 3.29
1348.2.3.3 -24.35 5.60 44.96 16.08 3.26
1348.2.3.4 -24.23 5.93 43.55 15.43 3.29
1348.2.3.5 -24.32 5.79 44.99 16.01 3.28
1348.3 M3 1348.3.3.1 -23.96 5.24 44.14 15.74 3.27
1348.3.3.2 -24.01 5.56 44.65 15.68 3.32
1348.3.3.3 -23.94 5.69 45.44 15.86 3.34
1348.3.3.4 -24.04 5.52 43.95 15.74 3.26
1348.3.3.5 -23.85 5.70 44.59 15.84 3.28
1464.1 M3 1464.1.3.1 -23.31 4.21 43.74 15.71 3.25
1464.1.3.2 -23.73 4.62 43.11 15.40 3.26
1464.1.3.3 -24.52 4.64 43.50 15.56 3.26
1464.1.3.4 -24.05 4.48 42.02 15.01 3.27
1535.1 M3 1535.1.3.1 -24.91 5.98 42.99 15.15 3.31
1535.1.3.2 -25.03 5.78 43.39 15.33 3.30
1535.1.3.3 -24.71 5.94 44.72 15.87 3.29
1535.1.3.4 -24.48 6.15 43.50 15.37 3.30
1535.1.3.5 -24.44 5.78 43.56 15.26 3.33
C1
952.1 M3 952.1.3.1 -23.77 5.65 42.79 15.20 3.28
952.1.3.2 -24.03 5.75 40.58 14.37 3.29
952.1.3.3 -24.32 5.99 44.39 15.68 3.30
952.1.3.4 -24.52 6.42 43.19 15.41 3.27
1067.1 M3 1067.1.3.1 -24.83 5.42 44.95 16.06 3.26
1067.1.3.2 -24.90 5.67 43.55 15.54 3.27
1067.1.3.3 -24.76 5.74 44.29 15.85 3.26
1067.1.3.4 -24.52 5.90 44.70 15.86 3.29
1067.1.3.5 -24.20 6.02 43.68 15.50 3.29
1067.1.3.6 -23.99 5.72 43.12 15.24 3.30
1238.1 M3 1238.1.3.1 -24.63 5.55 42.11 15.00 3.27
1238.1.3.2 -24.23 5.93 43.69 15.50 3.29
1238.1.3.3 -24.53 6.52 42.78 15.24 3.27
1238.1.3.4 -24.85 6.63 42.85 15.24 3.28
1238.1.3.5 -25.20 6.22 41.54 14.81 3.27
1244.1 M3 1244.1.3.1 -24.32 5.37 42.02 14.92 3.28
1244.1.3.2 -24.25 5.96 42.87 15.25 3.28
1244.1.3.3 -24.42 6.15 44.07 15.66 3.28
1244.1.3.4 -24.88 6.31 41.63 14.79 3.28
1244.1.3.5 -25.09 6.63 41.67 14.79 3.29
D1/D3
710.1 M1 710.1.1.1 -23.04 6.79 42.39 14.99 3.30
710.1.1.2 -23.36 7.16 43.54 15.31 3.32
710.1.1.3 -23.33 7.78 42.30 14.83 3.33

64
Tab. 8
Die δ18
O-Werte aus den Wasserproben der verschiedenen Entnahmestellen.
Nr. Gewässer Ort Höhe [ M. ü. M.] Temperatur [°C] Koordinaten δ18O[‰]
1 Zugersee Zug 413 15 681.500/224.450 -7.8
2 Zugersee Arth 413 14 682.320/213.190 -7.5
3 Lorze Neuägeri 685 14 685.250/223.100 -8.3
4 Ägerisee Unterägeri 724 13 687.750/221.320 -8.4
5 Regenwasser Sternegg 1500 10 699.500/206.950 -9.2
9. Literaturverzeichnis
Ambrose, S. H., DeNiro, M.J., 1986. The isotopic ecology of East African mammals. Oecologie 69,
395-406.
Ambrose, S. H., 1993. Isotopic analysis of paleodiets: methodological and interpretive considerations.
In : Stanford, M.K. (Ed.). Investigations of Ancient Human Tissue. Chemical Analysis in
Anthropology. Langhorne: Gordon & Breach Science Publishers, 59–130.
Ambrose, S.H., Norr, L., 1993. Experimental evidence for the relationship of the carbon isotope ratios
of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert, B.,
Grupe, G. (Eds.), Prehistoric Human Bone. Archaeology at the Molecular Level. Springer-
Verlag, Berlin, 1-37.
Ambrose, S.H., 2000. Controlled diet and climate experiments on nitrogen isotope ratiosin rats. In :
Ambrose, S.H., Katzenberg, M.A. (Hrsg.). Biochemical Approaches to Paleodietary Analysis.
Kluwer Academic/Plenum Publishers : New York, 243-259.
Azorit, C., Analla, M., Carrasco, R., Calvo, J.A., Muñoz-Cobo, J., 2002. Teeth eruption pattern in red
deer (Cervus elaphus hispanicus) in southern Spain. Ann. Biol. 24, 107–114.
Balasse, M., Bocherens, H., Mariotti, A., 1999. Intra-bone variability of collagen and apatite isotopic
composition as evidence of a change of diet. Journal of Archaeological Science 26, 593–598.
Balasse, M., Bocherens, H., Mariotti, A., Ambrose, S. H., 2001. Detection of dietary changes by intra-
tooth carbon and nitrogen isotopic analysis: An experimental study of dentine collagen of
cattle (Bos taurus). Journal of Archaeological Science 28, 235-245.
Balasse, M., Ambrose, S. H., Smith, A. B., Price, T. D., 2002. The Seasonal Mobility Model for
Prehistoric Herders in the South-western Cape of South Africa Assessed by Isotopic Analysis
of Sheep Tooth Enamel. Journal of Archaeological Science 29, 917–932.
Balasse, M., 2002. Reconstructing dietary and environmental history from enamel isotopic analysis:
time resolution of intra-tooth sequential sampling. International Journal of Osteoarchaeology
12, 155–165.

65
Balasse, M., Tresset, A., 2002. Early weaning of neolithic domestic cattle (Bercy, France) revealded
by intra-tooth variation in nitrogen isotope ratios. Journal of Archaeological Science 29, 853-
859.
Balasse, M., 2003. Potential biases in sampling design and interpretation of intra-tooth isotope
analysis. International Journal of Osteoarchaeology 13, 3-10.
Balasse, M., Smith, A. B., Ambrose, S. H., Leigh, S. R., 2003. Determining sheep birth seasonality by
analysis of tooth enamel oxygen isotope ratios: The late Stone Age site of Kasteelberg (South
Africa). Journal of Archaeological Science 30, 205-215.
Balasse, M., Ambrose, S.H., 2005. Mobilité altitudinale des pasteurs néolithiques dans la vallée du Rift
(Kenya): premiers indices de l’analyse du d13C de l’émail dentaire du cheptel domestique.
Anthropozoologica 40, 147-166.
Balasse, M., Tresset, A., Ambrose, S.H., 2006. Stable isotope evidence (d13
C, d18
O) for winter
feeding on seaweed by Neolithic sheep of Scotland. Journal of Zoology 270, 170–176.
Balasse, M., Tresset, A., 2007. Environmental constraints on the reproductive activity of domestic
sheep and cattle: what latitude for the herder? Anthropozoologica 42, 71-88.
Balasse, M., Boury, L., Ughetto-Monfrin, J., Tresset, A., 2012a. Stable isotope insights (d18O, d13C)
into cattle and sheep husbandry at Bercy (Paris, France, 4th millennium BC) : birth
seasonalitiy and winter leaf foddering. Environmental Archaeology 17:1, 29-44.
Balasse, M., Obein, G., Ughetto-Monfrin, J., Mainland, I., 2012b. Investigating seasonality and season
of birth in past herds: A reference set of sheep enamel stable oxygen isotope ratios.
Archaeometry 54 :2, 349–368.
Bentley, R. A., C. Knipper, 2005. Geographical patterns in biologically available strontium, carbon and
oxygen isotope signatures in prehistoric SW Germany. Archaeometry 47, 629-644.
Bernard, A., Daux, V., Lécuyer, C., Brugal, J.-P., Genty, D., Wainer, K., Gardien, V., Fourel, F.,
Jaubert, J., 2009. Pleistocene seasonal variations recorded in the d18O of Bison priscus teeth.
Earth and Planetary Science Letters 283, 133e143.
Broadmeadow, M.S.J., Griffiths, H., Maxwell, C., Borland, A.M., 1992. The carbon isotope ratio of
plant organic material reflects temporal and spatial variations in CO2 within tropical forest
formations in Trinidad. Oecologia 89, 435-441.
Bocherens, H., Billiou, D., Mariotti, A., Patou-Mathis, M., Otte, M., Bonjean, D., Toussaint, M., 1999.
Palaeoenvironmental and palaeodietary implications of isotopic biogeochemistry of Last
Interglacial Neanderthal and mammal bones in Scladina Cave (Belgium). Journal of
Archaeological Science 26, 599-607.
Bocherens, H., Fizet, M., Mariotti, A., 1994. Diet, physiology and ecology of fossil mammals as
inferred from stable carbon and nitrogen isotope biogeochemistry: implications for Pleistocene
bears. Paleogeography, Palaeoclimatology, Palaeoecology 107, 213–225.
Bocherens, H., Mashkour, M., Billiou, D., 2000. Palaeoenvironmental and archaeological implications
of isotopic analyses (13C, 15N) from Neolithic to Present in Qazvin Plain (Iran). Env. Archaeol.
5, 1-19.

66
Bocherens, H., Drucker, D., 2003. Trophic Level Isotopic Enrichment of Carbon and Nitrogen in Bone
Collagen: Case Studies from Recent and Ancient Terrestrial Ecosystems. International Journal
of Osteoarchaeology 13, 46–53.
Bösl, C., Gruppe, G., Peters, J., 2006. A late Neolithic vertebrate food web based on stable isotope
analyses. International Journal of Osteoarchaeology 16, 296-315.
Brown, W.A.B., 1991. Age assessment of red deer (Cervus elaphus): from a scoring scheme based on
radiographs of developing permanent molariform teeth. J. Zool. 225, 85–97.
Brown, W. A. B. & Chapman, N. G., 1991. Age assessment of fallow deer (Dama dama): from a
scoring scheme based on radiographs of developing permanent molariform teeth. J. Zool.
224, 367-379.
Buddecke, E., 1981. Biochemische Grundlagen der Zahnmedizin (Berlin, New York: Walter de
Gruyter).
Bützler, W., 1986. Cervus elaphus – Rothirsch. In: Jochen Niethammer und Franz Krapp (Hrsg.),
Handbuch der Säugetiere Europas. Paarhufer: 107–139.
Cerling, T.E., Harris, J.M. 1999. Carbon isotope fractionation between diet and bioapatite in
ungulate mammals and implications for ecological and paleoecological studies. Oecologia
120, 347–363.
Dansgaard, W., 1964. Stable isotopes in precipitation. Tellus 16, 436–468.
DeNiro, M. J., Epstein, S., 1981. Influence of diet on the distribution of nitrogen isotopes in animals.
Geochimica et Cosmochimica Acta 45, 341-351.
Deschler-Erb, S., Marti-Grädel, E., 2004. Viehhaltung und Jagd. Ergebnisse der Untersuchung der
handaufgelesenen Tierknochen. In: Jacomet, S., Leuzinger, U., Schibler, J., 2004. Die
jungsteinzeitliche Seeufersiedlung Arbon-Bleiche 3. Umwelt und Wirtschaft. Archäologie
im Thurgau 12, 2004, 158-251.
Drenckhahn, D., Fleischhauer, K., 1994. Mundhöhle. In: Drenckhahn D., Zenker W. (Hrsg.).
Benninghoff Anatomie. 15. Auflage. München, Wien, Baltimore: Urban & Schwarzenberg.
780-815.
Drucker, D., 2001. Validation méthodologique de l’analyse isotopique d’ossements fossiles et
apports aux reconstitutions paléoécologiques du Paléolithique supérieur du sudouest
de la France. Unpubl. Ph.D. Thesis, Universite. Paris 6, 222 pp.
Drucker, D., Célérier, G., 2001. Teneurs en carbone-13 du collagène de grands mammifères du site
de Pont d’Ambon (Dordogne, France): Implications pour l’environnement et son
exploitation au Tardiglaciaire dans le sud-ouest de la France. Paleo 13, 145-153.
Drucker, D., Bocherens, H., Bridault, A., Billiou, D,. 2003. Carbon and nitrogen isotopic composition
of red deer (Cervus elaphus) collagen as a tool for tracking palaeoenvironmental change
during the Late-Glacial and Early Holocene in the northern Jura (France). Palaeogeography,
Palaeoclimatology, Palaeoecology 195, 375–388.
Drucker, D.G., Bridault, A., Iacumin, P., Bocherens, H., 2009. Bone stable isotopic signatures (15N,
18O) as tracers of temerature variation during the Late-glacial and early Holocene: case study
on red deer Cervus elaphus from Rochedane (Jura, France). Geological Journal 44, 593-604.

67
Drucker, D.G., Bridault, A., Cupillard, C., Hujic, A., Bocherens, H., 2011. Evolution of habitat and
environment of red deer (Cervus elaphus) during theLate-glacial and early Holocene in
eastern France (French Jura and the western Alps) using multi-isotope analysis (δ13
C, δ15
N,
δ18
O, δ34
S) of archaeological remains. Quaternary International 245, 268-278.
Eberli, U., 2010. Die horgenzeitliche Siedlung Pfäffikon-Burg. Monographien der Kantonsarchäologie
Zürich 40.
Frank, R.M., Nalbandian, J., 1989. Structure and ultrastructure of dentine. In: Berkovitz, B. K. B.,
Boyde, A., Frank, R., M., Höhling, H. J., Moxham, B. J., Nalbandian, J., Tonge, C. H., Teeth
(Berlin, Heidelberg, New York, London, Paris, Tokyo) 173-247.
Frémondeau, D., Cucchi, T., Casabianca, F., Ughetto-Monfrin, J., Horard-Herbin, M.-P., Balasse, M.,
2012. Seasonality of birth and diet of pigs from stable isotope analyses of tooth enamel (d18O,
d13C): a modern reference data set from Corsica, France. Journal of Archaeological Science
39, 2023-2035.
Fricke, H. C., O’Neil, J. R., 1996. Inter- and intra-tooth variation in the oxygen isotope composition of
mammalian tooth enamel phosphate: implications for palaeoclimatological and
palaeobiological research. Palaeogeography, Palaeoclimatology, Palaeoecology 126, 91-99.
Fricke, H.C., Clyde, W.C., O’Neil, J.R., Gingerish, P.D., 1998. Evidence for rapid climate change in
North America during the latest Paleocene thermal maximum: oxygen isotope compositions of
biogenic phosphate from the Bighorn Basin (Wyoming). Earth and Planetary Science Letters
160, 193-208.
Godawa, J., 1989. Age determination in the Red deer (Cervus elaphus). Acta Theriologyca 34: 381-
384.
Hafner, A., Suter, P.J., 2003. Das Neolithikum in der Schweiz. www.jungsteinSITE.de
Heaton, T.H.E., 1999. Spatial, species, and temporal variations in the 13C/12C ratios of C3 plants:
implications for palaeodiet studies. Journal of Archaeological Science 26, 637-649.
Hedges, R.E.M., 2003. On Bone Collagen - Apatite-Carbonate Isotopic Relationships. International
Journal of Osteoarchaeology 13, 66–79.
Higgins, P., MacFadden, B. J., 2004. „Amount Effect’’ recorded in oxygen isotopes of Late Glacial
horse (Equus) and bison (Bison) teeth from the Sonoran and Chihuahuan deserts,
southwestern United States. Palaeogeography, Palaeoclimatology, Palaeoecology 206, 337-
353.
Hobson, K. A., Sease, J. L., 1998. Stable isotope analyses of tooth annuli reveal temporal dietary
records: an example using Steller sea lions. Marine Mammal Science 14, 116–129.
Hoefs, J., 2009. Stable Isotope Geochemistry. Berlin : Springer.
Hoppe, K. A., Amundson, R., Vavra, M., McClaran, M.P., Anderson, D. L., 2004. Isotopic analysis of
tooth enamel carbonate from modern North American feral horses: implications for
paleoenvironmental reconstructions. Palaeogeography, Palaeoclimatology, Palaeoecology
203, 299-311.

68
Huber, R., Schaeren, G., 2009. Zum Stand der Pfahlbauforschung im Kanton Zug. Tugium 25, 111-
140.
Hüster-Plogmann, H., 2002. Früheste archöologische Hinweise zur Nutzung von Rindern als Zugtiere
in neolithischen Siedlungen der Schweiz. In: J. Köninger et al. (Hrsg), Schleife, Schlitten, Rad
und Wagen. Zur Frage früher Transportmittel nördlich der Alpen. Hemmenhofener Skripte 3,
103-108.
Jedrzejewski, W., Jedrzejewska, B., Okarma, H., Schmidt, K., Zub, K., Musiani, M., 2000. Prey
selection and predation by wolves in Bialowieza primeval forest, Poland. Journal of
Mammalogy 81, 197–212.
Knipper, C., 2004. Die Strontiumisotopenanalyse: eine naturwissenschaftliche Methode zur Erfassung
von Mobilität in der Ur- und Frühgeschichte. Jahrb. RGZM 51, 2004, 589–685.
Knipper, C., Paulus, S., Uerpmann, H-P, 2006. Sauerstoff- und Strontium-Isotopenanalysen an
Tierknochenfunden aus Udabno (Georgien). Archäologische Mitteilungen aus Iran und Turan
38, 213-217.
Knipper, C., 2011. Die räumliche Organisation der linearbandkeramischen Rinderhaltung:
naturwissenschaftliche und archäologische Untersuchungen. British Archaeological Reports.
International Series 2305, Oxford.
Koch, P. L., Fischer, D. C., Dettman, D., 1989. Oxygen isotope variation in the tusks of extinct
proboscideans: a measure of season of death and seasonality. Geology 17, 515–519.
Koch, P. L., Heisinger, J., Moss, C., Carlson, R. W., Fogel, M. L., Behrensmeyer, A. K., 1995.
Isotopic tracking of change in diet and habitat use in African elephants. Science 267, 1340–
1343.
Kohn, M. J., Miselis, J. L., Fremd, T. J., 2002. Oxygen isotope evidence for progressive uplift of the
Cascade Range, Oregon. Earth and Planetary Science Letters 204, 151-165.
Körner, C., Farquhar, G.D., Wong, S.C., 1991. Carbon isotope discrimination by plants follows
latitudinal and altitudinal trends. Oecologia 88, 30-40.
Lee-Thorp, J.A., Sealy, J.C., van der Merwe, N.J., 1989. Stable carbon isotope ratio differences
between bone collagen and bone apatite, and their relationship to diet. Journal of
Archaeological Science 16, 585–599.
Longinelli, A., 1966. Ratios of oxygen-18 : oxygen-16 in phosphate and carbonate from living and
fossil marine organisms. Nature 211, 923-927.
Longinelli, A., 1995. Stable isotope ratios in phosphate from mammal bone and tooth as climatic
indicators. In : Frenzel, B., Stauffer, B., Weiss, M.M. (Hrsg.). Problems of stable isotopes in
tree-rings, lake sediments and peat bogs as climatic evidence for the Holocene.
Paläoklimaforschung 15, 57-70.
Luz, B., Y. Kolodny, 1985. Oxygen isotope variations in phosphate of biogenic apatites, IV. Mammal
teeth and bones. Earth and Planetary Science Letters 75, 29-36.
Mariezkurrena, K., 1983. Contribución al conocimiento del desarrollo de la dentición y el esqueleto
postcraneal de Cervus elaphus. Munibe 35: 149-202.

69
Mitchell, B., 1967. Growth layers in dental cement for determining the age of red deer (Cervus elaphus
L.). Journal Animal Ecology 36: 279-293.
Müldner, G.,Richards, M. P., 2005. „Fast or feast: reconstructing diet in later medieval England by
stable isotope analysis”, Journal of Archaeological Science, 32, 39-48.
Müller-Using, D., 1981. Rotwildalter-Merkblatt. 7th edition. Bonn: D.J.V.
O.N.C. (Office National de la Chasse), 1984. Le Cerf d´Europe. París.
Passey, B.H., Robinson, T.F., Ayliffe, L.K., Cerling, T.E., Sponheimer, M., Dearing, D., Roeder, B.L.,
Ehleringer, J.R., 2005. Carbon isotope fractionation between diet, breath CO2, and bioapatite in
different mammals. Journal of Archaeological Science 32, 1459–1470.
Raesfeld, F., 1988. Das Rotwild: Naturgeschichte, Hege, Jagdausübung. Hamburg: Paul Parey.
Richards, M., Dobney, K., Albarella, U., Fuller, B., Pearson, J., Müldner, G., Jay, M., Molleson, T.,
Schibler, J., 2002. Stable isotope evidence of Sus diets from European and Near Eastern
archaeological sites. In: Paper Presented at the International Council of Archaeozoology (ICAZ)
9th Conference. University of Durham.
Righetti, A., 1995. Cervus elaphus – Rothirsch. In: Hausser J., Denkschriftenkommission der
Schweizerischen Akademie der Naturwissenschaften (Hrsg.) Säugetiere der Schweiz:
Verbreitung, Biologie, Ökologie, 433-439.
Rodière, É., Bocherens, H., Angibault, J.-M., Mariotti, A.,1996. Particularite.s isotopiques chez le
chevreuil (Capreolus capreolus L.): Implications pour les reconstitutions
paléoenvironnementales. C. R. Acad. Sci. Paris 323, 179-185.
Rösch, M., 2010. Landnutzung, Pflanzenproduktion und Sammelwirtschaft. Badisches
Landesmuseum Karlsruhe (Hrsg.), Jungsteinzeit im Umbruch. Die Michelsberger Kultur und
Mitteleuropa vor 6000 Jahren. Ausstellungskatalog, 164-168.
Schibler, J., Brombacher, C., 1995. Wirtschaft. In: Stöckli, W. E., Niffeler, U. Gross-Klee, E. (Hrsg.),
Die Schweiz vom Paläolithikum bis zum Frühen Mittelalter. SPM II: Neolithikum. Basel: Verlag
Schweizerische Gesellschaft für Ur- und Frühgeschichte, 85-120.
Schibler, J., Chaix, L., 1995. Wirtschaftliche Entwicklung aufgrund archäozoologischer Daten. In:
Stöckli, W. E., Niffeler, U. Gross-Klee, E. (Hrsg.), Die Schweiz vom Paläolithikum bis zum
Frühen Mittelalter. SPM II: Neolithikum. Basel: Verlag Schweizerische Gesellschaft für Ur- und
Frühgeschichte, 97-118.
Schibler, J., Hüster-Plogmann, H., 1995. Die neolithische Wildtierfauna und ihr Aussagegehalt
betreffend Umwelt und Umweltveränderungen. In: Stöckli, W. E., Niffeler, U. Gross-Klee, E.
(Hrsg.), Die Schweiz vom Paläolithikum bis zum Frühen Mittelalter. SPM II: Neolithikum. Basel:
Verlag Schweizerische Gesellschaft für Ur- und Frühgeschichte, 76-83.
Schibler, J., 1997a. Haus- und Wildtiernutzung in der jungsteinzeitlichen Feuchtbodensiedlungen des
Kantons Thurgau. Archäologie der Schweiz 20/2, 57–61.
Schibler, J., Hüster-Plogmann, H., Jacomet, S., Brombacher, C.,Gross-Klee, E. and Rast-Eicher, A.,
1997b. Ökonomie und Ökologie Neolithischer und Bronzezeitlicher Ufersiedlungen am
Zürichsee. Monographien der Kantonsarchäologie Zürich 20.

70
Schibler, J., Steppan, K., 1999. Human impact on the habitat of large herbivores in Eastern
Switzerland and Southwest Germany in the Neolithic. Archaeofauna 8, 87–99.
Schibler, J., 2006. The economy and environment of the 4th and 3rd millennia BC in the northern
Alpine foreland based on studies of animal bones. Environmental Archaeology 2006, Vol. 11,
No. 1, 49-64.
Schibler, J., 2010. Crisis? What crisis? Die Nahrungskrise im 37. Jh. v.Chr. und ihre Bewältigung.
Badisches Landesmuseum Karlsruhe (Hrsg.), Jungsteinzeit im Umbruch. Die Michelsberger
Kultur und Mitteleuropa vor 6000 Jahren. Ausstellungskatalog, 173-178.
Schotterer, U., Stocker, T., Bürki, H., Hunziker, J., Kozel, R., Grasso, D. A., Tripet, J.-P., 2000. Das
Schweizer Isotopen-Messnetz. Trends 1992-1999. gwa 10/00, 733–741.
Schotterer, U., 2010. Wasserisotope in der Schweiz. Neue Ergebnisse und Erfahrungen aus dem
nationalen Messnetz ISOT. gwa 12/10, 1073-1081.
Schöninger, M. J., DeNiro, M. J., 1984. Nitrogen and carbon composition of bone collagen from
marine and terrestrial animals. Geochimica et Cosmochimica Acta 48, 625-639.
Schumacher, G.-H., Schmidt, H., Börnig, H., Richter, W., 1990. Anatomie und Biochemie der Zähne.
4. Auflage (Stuttgart, New York: Gustav Fischer).
Sharp, Z. D., 2005. Principles of stable isotope geochemistry (Upper Saddle River, New Jersey).
Stephan, E., 1999. Sauerstoffisotopenverhältnisse im Knochengewebe großer terrestrischer
Säugetiere. Tübinger Geowiss. Arb. E 6.
Stephan, E., 2008. Stabile Isotope in fossilen Faunenfunden: Erforschung von Klima, Umwelt und
Ernährung prähistorischer Tiere. In: Haupmann, A., Pingel, V. (Hrsg.). Archäometrie. Methoden
und Anwendungsbeispiele, 46-66.
Stevens, R.E., Hedges, R.E.M., 2004. Carbon and nitrogen stable isotope analysis of Northwest
European horse bone and tooth collagen, 40,000 BP–present: palaeoclimatic interpretations.
Quaternary Science Reviews 23:7–8, 977–991.
Stevens, R.E., Lister, A.M., Hedges R.E.M., 2006. Predicting diet, trophic level and palaeoecology
from bone stable isotope analysis: a comparative study of five red deer populations. Oecologia
149, 12-21.
Stevens, R.E., Balasse, M., O’Connell, T.C., 2011. Intra-tooth oxygen isotope variation in a known
population of red deer: Implications for past climate and seasonality reconstructions.
Palaeogeography, Palaeoclimatology, Palaeoecology 301, 64-74.
Stöckli W.E., 1995. Geschichte des Neolithikums in der Schweiz. In: SPM II. Die Schweiz vom
Paläolithikum bis zum frühen Mittelalter. Band II: Neolithikum, 19–52.
Stöckli, W.E., 2009. Chronologie und Regionalität des jüngeren Neolithikums (4300-2400 v. Chr.) im
Schweizer Mittelland, in Süddeutschland und in Ostfrankreich aufgrund der Keramik und der
absoluten Datierungen, ausgehend von den Forschungen in den Feuchtbodensiedlungen der
Schweiz. Antiqua 45.
Suga, S., 1982. Progressive mineralization pattern of developing enamel during the maturation stage.
J. Dent. Res. 61, 1532–1542.

71
Tieszen, L.L., Boutton, T.W., 1989. Stable carbon isotopes in terrestrial ecosystem research. In:
Rundell, P.W., Ehleringer, J.R., Nagy, K.A. (Eds.), Stable Isotopes in Ecological Research, 167-
195.
Tieszen, L.L., 1991. Natural variations in the carbon isotope values of plants: Implications for
archaeology, ecology, and paleoecology. Journal of Archaeological Science 18, 227-248.
Tütken, T., 2010. Die Isotopenanalyse fossiler Skelettreste – Bestimmung der Herkunft und Mobilität
von Menschen und Tieren. Tagungen des Landesmuseums für Vorgeschichte Halle, Band 3,
2010.
Vennemann, T. W., Fricke, H. C., Blake, R. E., O’Neil, J. R., Colman, A., 2002. Oxygen isotope
analyses of phosphates: a comparison of techniques for analysis of Ag3PO4. Chemical Geology
185, 321-336.
Vohberger, M. A., 2011. Lokal oder eingewandert? Interpretationsmöglichkeiten und Grenzen lokaler
Strontium- und Sauerstoffisotopensignaturen am Beispiel einer Altgrabung in Wenigumstadt.
Unpubl. Dissertation an der Ludwig-Maximilians-Universität München.
Vogel, J. C., 1978. Recycling of carbon in a forest environment. Oecologia Plantarum 13, 89-94.
Wagenknecht, E., 1980. Der Rothirsch. Wittenberg Lutherstad: A. Ziemsen Verlag.
Wagenknecht, E., 1996. Der Rothirsch. Heidelberg: Die neue Brehm-Bücherei 129.
Wagenknecht, E., 2000. Rotwild. Suderburg: Nimrod-Verlag.
Wiedemann, F. B., Bocherens, H., Mariotti, A., v. d. Driesch, A., Gruppe, G., 1999. Methodological and
archaeological implications of intra-tooth isotopic variations (δ13C, δ18O) in herbivores from Ain
Ghazal (Jordan, Neolithic). Journal of Archaeological Science 26, 697-704.
Wright, L. E., Schwarcz, H. P., 1999. Correspondence between stable carbon, oxygen and nitrogen
isotopes in human tooth enamel and dentine: infant diets at Kaminaljuyu´. Journal of
Archaeological Science 26, 1159–1170.
Zazzo, A., Mariotti, A., Lécuyer, C., Heintz, E., 2002. Intra-tooth isotopic variations in Late Miocene
bovid enamel from Afghanistan: paleobiological, taphonomical, and climatical implications.
Palaeogeography, Palaeoclimatology, Palaeoecology 186, 145-161.
Zazzo, A., Balasse, M., Patterson, W.P., 2005. High-resolution d13C intratooth profiles in bovine
enamel: implications for mineralization pattern and isotopic attenuation. Geochimica et
Cosmochimica Acta 69, 3631-3642.
Internetadressen
http://www.univie.ac.at/cartography/project/wiser/

72
10. Danksagung
Die erfolgreiche Durchführung dieser Arbeit war nur mit der Hilfe und Unterstützung von zahlreichen
Personen möglich, die mir bei all meinen Fragen und Anliegen viel Zeit, Geduld und Verständnis
entgegenbrachten.
Mein grösster Dank geht an Corina Knipper, für die engagierte Betreuung vor und während meiner
Masterarbeit sowie für die kompetente Einführung in die Isotopenanalytik in der Archäologie. Durch sie
habe ich gelernt, eigene Interpretationswege in diesem komplexen und für mich neuen
Forschungsgebiet zu wagen.
Speziell möchte ich Herrn Prof. Schibler danken, durch dessen Unterstützung dieses Projekt
ermöglicht wurde der mir diese grosse Herausforderung zugetraut hat. Die gemeinsamen Gespräche
haben mir geholfen, die archäologische Fragestellung im Isotopie-Dschungel nicht aus den Augen zu
verlieren.
Ebenfalls zu grossem Dank verpflichtet bin ich der Kantonsarchäologie Zug, die mir trotz des
ungewissen Ausgangs der Untersuchungen vollstes Vertrauen für die Bearbeitung ihres
Fundmaterials entgegenbrachte. Vor allem Eda Gross und Gishan Schären möchte ich danken für ihre
Informationsbereitschaft, wenn es um die Klärung von Fragen bezüglich der Fundstelle und der
Befunde ging.
Ein grosses Dankeschön geht an Moritz Lehmann, Leiter des Labors für Stabile Isotope am Institut für
Umweltgeowissenschaften der Universität Basel sowie an Mark Rollog, den Profi am
Massenspektrometer. Auch wenn mir die Zeit fehlte, mich in die Tiefen der Massenspektrometrie
einzuarbeiten bin ich überzeugt, dass sie hervorragende Arbeit geleistet haben.
Herzlichen Dank an Prof. Dr. Thomas Boller und Giacomo Busco vom Botanischen Institut Basel für
die Unterstützung am Gefriertrockner und im Kampf gegen den Kollagen-Schaum.
Danke an Michael Maus vom Institut für Geowissenschaften der Universität Mainz sowie dem ganzen
Team für die Unterstützung bei den Vorbereitung und Messungen der ersten Proben in Mainz.
Nicht zuletzt geht ein riesen grosses Dankeschön an alle Dozenten, Mitarbeitenden und Studierenden
vom IPNA, die mir bei der Bewältigung und dem Lösen von praktischen sowie theoretischen
Problemen geholfen haben und mir mit Rat und Tat zur Seite standen. An den anregenden
Diskussionen ist diese Arbeit gewachsen.
Allen Verwandten, Bekannten und Freunden ist ein grosses Lob auszusprechen für ihre Bereitschaft,
sich für mich mit so einem komplexen Thema auseinanderzusetzen und Interesse dafür zu zeigen.
Mein herzlichster Dank geht an meine Eltern. Für meine Förderung, für die Unterstützung und ihr
Glaube an mich. Danke.
Und an C.K.G.