developmental effects of coumarin and the anticoagulant coumarin derivative warfarin on zebrafish...
TRANSCRIPT

Dw
Sa
b
a
ARRAA
KZTCWA
1
pipspmezt(aozgf[ta
2
(h
0d
Reproductive Toxicology 33 (2012) 133– 141
Contents lists available at ScienceDirect
Reproductive Toxicology
jo u r n al hom epa ge: ww w.elsev ier .com/ locate / reprotox
evelopmental effects of coumarin and the anticoagulant coumarin derivativearfarin on zebrafish (Danio rerio) embryos
tefan Weigta,b,∗, Nicole Hueblera, Ruben Streckerb, Thomas Braunbeckb, Thomas H. Broscharda
Institute of Toxicology, Merck KGaA, 64293 Darmstadt, GermanyAquatic Ecology and Toxicology Group, Department of Zoology, University of Heidelberg, 69120 Heidelberg, Germany
r t i c l e i n f o
rticle history:eceived 18 February 2011eceived in revised form 17 June 2011ccepted 2 July 2011vailable online 21 July 2011
a b s t r a c t
Coumarin and warfarin, two substances which are intensively metabolized in animals and humans, weretested for teratogenicity and embryo lethality in a 3-day in vitro assay using zebrafish embryos. Warfarinis a coumarin derivative, but in contrast to the mother substance warfarin has anticoagulant properties.Both substances produced teratogenic and lethal effects in zebrafish embryos. The LC50 and EC50 valuesfor coumarin are 855 �M and 314 �M, respectively; the corresponding values for warfarin are 988 �M
eywords:ebrafisheratogenicoumarinarfarin
and 194 �M. For coumarin, three main or fingerprint endpoints (malformation of head, tail and growthretardation) were identified, whereas malformation of tail was the only fingerprint endpoint of warfarin.The analysis of the ratios between the zebrafish embryo effect concentrations of both substances andhuman therapeutic plasma concentrations confirmed the teratogenic potential of warfarin, as well as theequivocal status of coumarin.
nticoagulants
. Introduction
Over the last years, zebrafish embryos have been discussed as aromising alternative model for teratogenicity testing [1–11]. Orig-
nally it was assumed that zebrafish embryos are not able to activateroteratogenic substances by themselves [12,13]. After havinghown that zebrafish embryos are also able to metabolize/activateroteratogenic substances without addition of an exogenousetabolic activation system [14], we have now investigated the
ffects of coumarin and the coumarin derivative warfarin in aebrafish embryo teratogenicity test. Several aspects make thesewo substances interesting test compounds in zebrafish embryos:i) either substance is intensively metabolized in both humans andnimals [15–20]; (ii) bioavailability of warfarin strongly dependsn CYP metabolism [21–23]; (iii) parallels between human andebrafish CYP metabolism have been described and it was sug-ested that the zebrafish, in some cases, may be a better modelor human toxicity than some commonly used mammalian models
24]; (iv) coumarin derivatives with anticoagulant properties areeratogenic in humans, but do not show teratogenic effects in anynimal developmental toxicity study [25–28]; (v) currently, there is∗ Corresponding author at: Institute of Toxicology, Building U9, Frankfurter Straße50, 64293 Darmstadt, Germany. Tel.: +49 0 6151 72 3704; fax: +49 0 6151 72 7673.
E-mail addresses: [email protected] (S. Weigt), [email protected]. Huebler), [email protected] (R. Strecker), [email protected] (T. Braunbeck), [email protected] (T.H. Broschard).
890-6238/$ – see front matter © 2011 Elsevier Inc. All rights reserved.oi:10.1016/j.reprotox.2011.07.001
© 2011 Elsevier Inc. All rights reserved.
no clear information available concerning the teratogenic potentialof coumarin in humans [29,30].
In contrast to many of its derivatives, coumarin itself has noanticoagulant properties [31,32]. But it is used in other medicalindications, such as treatment of edemas, cancer and chronic infec-tions [33–37]. In humans, the CYP isoform found to be clearlydominant in coumarin metabolism is CYP2A6 [38–40].
Warfarin, also known as coumadin, is an oral anticoagulant drugvery common in the prophylaxis and treatment of thromboem-bolic diseases, especially in the UK and the USA [41]. Warfarinacts as a vitamin K antagonist, and vitamin K is needed as a cofac-tor for the carboxylation of glutamate residues of several clottingfactors [21,42,43]. For oral medication, a racemic mixture of enan-tiomers is used, but the (S) enantiomer exhibits approximately2–5 times more anticoagulant activity than the (R) enantiomer[23,44–46]. The two enantiomers are also metabolized by differ-ent CYP isoforms in humans. The (R) enantiomer is hydroxylatedby CYPs 1A1, 1A2, 2C8, 2C9, 2C18, 2C19, 3A4 and 3A5 (question-able) with CYP1A2 and CYP3A4 being most important. (S)-warfarinhydroxylation is mainly catalyzed by CYP2C9, but CYPs 2A6 (ques-tionable), 2C8, 2C18 and 2C19 also contribute to the metabolism ofthis enantiomer [15,17,22,23,39,47–51]. The CYP enzyme which ismost important for the therapeutic use of warfarin is CYP2C9. How-ever, since warfarin interacts with vitamin K, the enzyme vitamin K
epoxide reductase (VKORC1) also plays a fundamental role [52–57].The terms “coumarin embryopathy” and “coumarin syndrome”used in human medicine today are a little confusing, as theseterms are only related to coumarin derivatives with anticoagulant

1 e Toxi
puataaonctitpf[rtcme
2
2
KmSp(Mta
2
2
zBtcwtsfiswwwdmc
maswut
2
tcctwsTgtt[i
34 S. Weigt et al. / Reproductiv
roperties and not to coumarin itself. Yet, in order to avoid mis-nderstandings, the old-fashioned terms “warfarin embryopathy”nd “warfarin syndrome” are still used in this publication. In 1966,he first case of warfarin embryopathy was described by DiSaia [58],nd Kerber et al. [59] were the first to suggest warfarin to be a ter-togenic agent. Today the embryotoxic potential of warfarin andther coumarin anticoagulants is well-accepted [29,30,60,61], buto epidemiological studies of children born to women treated withoumarin during pregnancy have been reported [29,30]. The lack oferatogenic effects of coumarin in developmental toxicity studiesn animals is not sufficient to conclude a lack of teratogenic poten-ial in humans, because coumarin derivatives with anticoagulantroperties (and human teratogenicity) also have not produced mal-ormations in any experimental animal species exposed prenatally25–28,62]. Pauli et al. suggested that the warfarin embryopathyesulted from the pharmacological effect of warfarin, i.e., inactiva-ion of vitamin K [63]. If this is also the case for zebrafish embryos,oumarin, which lacks anticoagulant properties on the basis of vita-in K antagonism [31,32], would not be able to produce warfarin
mbryopathy in this test system.
. Materials and methods
.1. Materials
Tris (Tris(hydroxymethyl)-aminomethan) and HCl were obtained from MerckGaA (Darmstadt, Germany). Coumarin, warfarin, 3-aminobenzoic acid ethyl esterethanesulfonate (MS-222), and dimethyl sulfoxide (DMSO) were purchased from
igma (Deisenhofen, Germany). 12.1 g Tris was dissolved in fish medium to pre-are a 100 mM buffer solution and 2 M HCl was used to adjust the pH value to 7.4at 26 ◦C). The fish medium (reconstitued water consisting of 2 mM CaCl2, 0.5 mM
gSO4, 0.7 mM NaHCO3, and 0.07 mM KCl) was prepared in the facility accordingo OECD Fish Embryo Test Draft Guideline [64]. All solutions were freshly prepared,nd chemicals were dissolved 2 h before incubation.
.2. Methods
.2.1. Animal care and egg productionA breeding stock of unexposed and healthy mature wild type Tuebingen strain
ebrafish (Danio rerio) (original supplier: Max Planck Institute for Developmentaliology Tübingen, Germany; breeder: Institute of Toxicology, Merck KGaA) olderhan 6 months was used for egg production. Spawners were maintained in a recir-ulating zebtec housing system (Tecniplast, Hohenpeißenberg, Germany) at 26 ◦Cith a loading capacity of a minimum of 1 L water per gram fish. The housing sys-
em is equipped with mechanical, biological, UV light and activated carbon filterystems; pH value is automatically kept at 7.8 and the conductivity at 800 �S. Thesh medium in the housing system was prepared by the system itself from stockolutions according to OECD Fish Embryo Test Draft Guideline [64]. The automaticater exchange was adjusted to 10% of the system total volume per day. Lightingas controlled by a timer to provide a 12/12 h light/dark cycle. Females and malesere continuously kept together in a ratio of 1:2 or 1:1. Dry flake food was fed twiceaily and frozen food (Cyclops and Artemia) was fed once a day, occasionally supple-ented with Artemia nauplii [64,65]. The concentration of nitrate and nitrite were
hecked once a week, but were consistently <100 mg/L and 1.0 mg/L, respectively.Mating and spawning took place within 30 min after turning on the lights in the
orning. To prevent adult zebrafish from egg predation, egg trays were covered with 2 mm plastic mesh. Plastic plant imitations fixed to the mesh served as spawningubstrate. About 30 min after the onset of light, egg trays were removed and eggsere collected [64]. Under the culture conditions described above, fertilized eggsndergo the first cleavage after approximately 15 min. Based on their transparency,he 4- to 32-cell stage eggs can clearly be identified as fertilized.
.2.2. Embryo exposureCoumarin and warfarin were dissolved in DMSO; with a final solvent concentra-
ion of 0.5% in the test media. For both substances, range-finding experiments wereonducted with a constant spacing factor of 2. The highest tested concentrations foroumarin and warfarin were 5 mM and 1.5 mM, respectively. Concentration selec-ions for the main study were based on the number of affected embryos (embryosith teratogenic and/or lethal effects) in the range-finding experiments (data not
hown). Coumarin was tested in 5 concentrations and warfarin in 7 concentrations.ris-buffer with 0.5% DMSO was used as control. Twenty embryos were used per
roup and each experiment was repeated 3 times. Tris-buffer was chosen as vehicleo keep the pH value at a physiological level. On the one hand, a controlled pH inhe test media is relevant since zebrafish embryos are very sensitive to pH changes66]; on the other hand, the test substance is made available at a pH similar to thatn human blood.cology 33 (2012) 133– 141
Eggs were first rinsed twice in glass Petri dishes with fish medium. Within 2hours post-fertilization (hpf), fertilized eggs (from 4- to 64-cell blastomeres) wereselected under a CKX41 stereomicroscope (Zeiss, Göttingen, Germany) into a plasticPetri dish containing Tris-buffer.
At latest 2.5 hpf, the incubation was started by addition of the fertilized fish eggsto the test solution. The embryos were exposed individually in 24-well plates (Nunc,Wiesbaden, Germany) containing a final volume of 2 mL per well at 26 ◦C with a12:12-h light/dark cycle in a precision incubator (Memmert, Schwabach, Germany).The well plates were sealed with self-adhesive foil (MicroAmp® optical adhesivefilm, Applied Biosystems, Darmstadt, Germany) to prevent evaporation. Before theembryos were added to the test solutions, the well plates were pre-warmed to 26 ◦C.
2.2.3. Evaluation (scoring) of lethality and teratogenic effectsAt 8 hpf as well as at 1, 2 and 3 dpf (days post-fertilization), the embryos were
evaluated and scored for lethal or teratogenic effects using a Zeiss CKX41 invertedmicroscope equipped with phase contrast optics, a mounted time-lapse recorderand the analySIS software (Olympus, Hamburg, Germany). The 8 hpf time pointserved as a control step to identify unfertilized eggs, which had entered the testaccidentally. Scoring for effects at 1 and 2 dpf was performed to track the fate of thetreated embryos and to give information about the time point when the differentendpoints were detectable. The final scoring at 3 dpf was performed on embryosanesthetized by addition of MS-222 (ethyl 3-aminobenzoate methanesulfonate, tri-caine, Sigma–Aldrich) solution (concentration about 0.1%). After final scoring theanesthetized embryos were killed by freezing at −20 ◦C for ≥24 h.
All embryos were staged as described by Kimmel et al. [67], and lethal or ter-atogenic effects were recorded according to Bachmann and Nagel [1,7] (Table 1).Teratogenic effects were considered as fingerprint endpoints, if the following cri-teria were fulfilled: (i) concentration–response relationship is present and (ii) theendpoint must be observed in ≥50% of all embryos showing teratogenic effects inall test groups of a test substance.
2.2.4. Calculation of LC50, EC20, EC50 and teratogenicity index (TI)For the calculation of EC and LC values, the program ToxRatPro (ToxRat® ,
Software for the Statistical Analysis of Biotests, ToxRat Solutions GmbH, Alsdorf,Germany, Version 2.10), using probit analysis with linear maximum likelihoodregression, was used. In order to characterize the teratogenic potential of a test sub-stance, the teratogenicity index (TI), which is traditionally defined as the quotient ofLC50 and EC50, was calculated. If the TI of a given substance is >1, the substance is con-sidered to be teratogenic; if the TI is ≤1, the substance produces mainly embryolethaleffects. The EC20 is used for the comparison between zebrafish embryo and humanexposure data.
2.2.5. Validity criteria and statisticsEgg batches were only used, if fertilization rates were ≥80%. An assay was con-
sidered valid, if the controls did not show >10% teratogenic plus lethal effects at 3dpf.
A Student’s one-tailed t-test was performed to identify statistically significantdifferences between treatment and control groups. Statistics were done on the basisof affected embryos (embryos with lethal and/or teratogenic effects).
3. Results
Lethal and teratogenic effects recorded at 3 dpf are summarizedin Tables 2 and 3. All controls (Fig. 1A) fulfilled the acceptancecriteria of ≤10% affected embryos at 3 dpf (Tables 2 and 3).Except for yolk deformities, all malformations listed in Table 1were also observed in embryos treated with coumarin and war-farin. In the low-dose treatment groups of coumarin and warfarin,the chorda malformations appeared as isolated lesions; in thehigh-dose groups, the chorda structure often displayed completedisintegration.
3.1. Exposure of zebrafish embryos to coumarin
The percentage of embryos with teratogenic and lethaleffects clearly increased with increasing coumarin concentrations(Table 2): teratogenic effects in zebrafish embryos exposed to 200and 400 �M coumarin were almost exclusively observed at 3 dpf;higher concentrations of coumarin (600, 800 and 1000 �M), how-ever, resulted in teratogenic effects already at 1 dpf (Table 4). At
the two highest concentrations, many of these early malforma-tions were very prominent and resulted in embryo lethality withinthe next 2 days, reaching 85% lethality at 1000 �M (Table 2). Themajor lethal endpoint in the experiments with coumarin was lack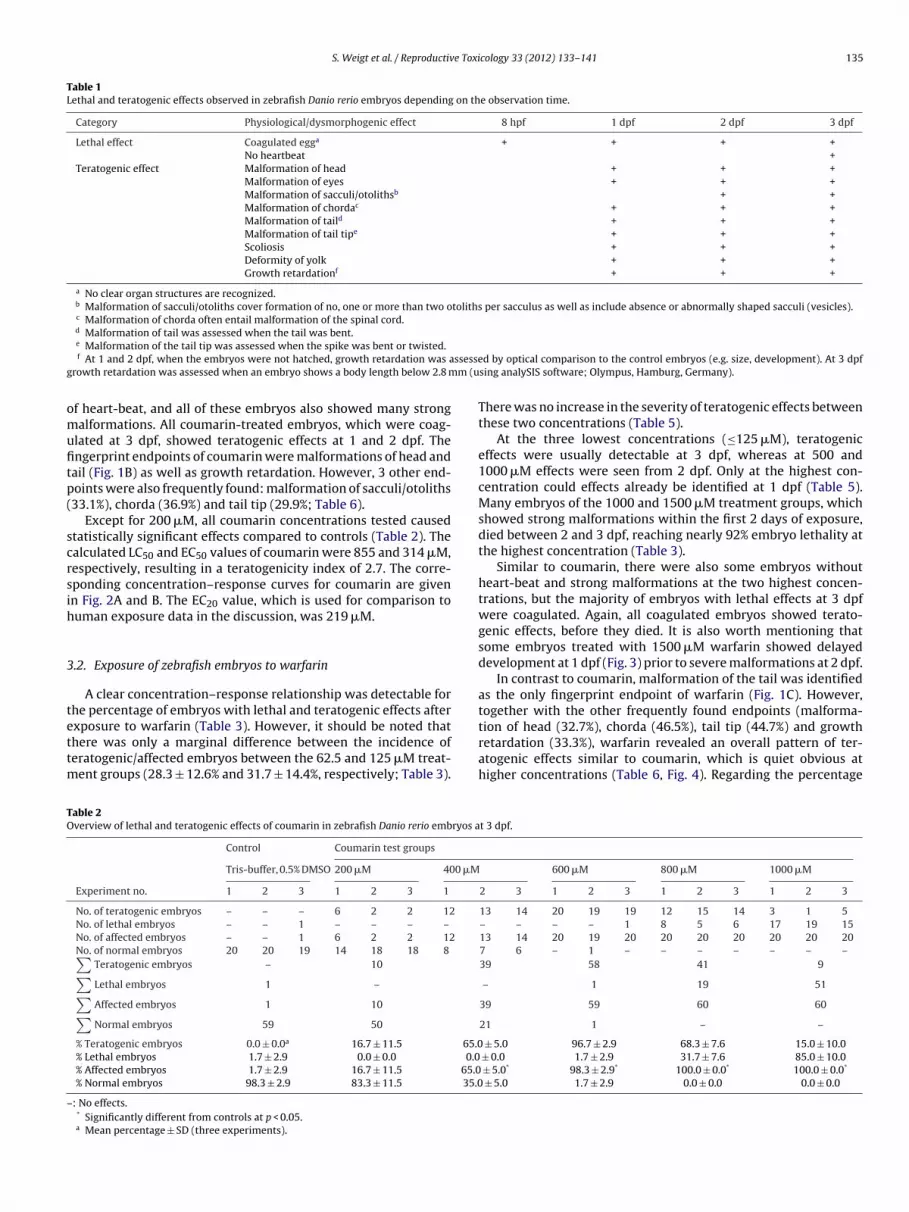
S. Weigt et al. / Reproductive Toxicology 33 (2012) 133– 141 135
Table 1Lethal and teratogenic effects observed in zebrafish Danio rerio embryos depending on the observation time.
Category Physiological/dysmorphogenic effect 8 hpf 1 dpf 2 dpf 3 dpf
Lethal effect Coagulated egga + + + +No heartbeat +
Teratogenic effect Malformation of head + + +Malformation of eyes + + +Malformation of sacculi/otolithsb + +Malformation of chordac + + +Malformation of taild + + +Malformation of tail tipe + + +Scoliosis + + +Deformity of yolk + + +Growth retardationf + + +
a No clear organ structures are recognized.b Malformation of sacculi/otoliths cover formation of no, one or more than two otoliths per sacculus as well as include absence or abnormally shaped sacculi (vesicles).c Malformation of chorda often entail malformation of the spinal cord.d Malformation of tail was assessed when the tail was bent.
ssesseg m (u
omufitp(
scrsih
3
tettm
TO
–
e Malformation of the tail tip was assessed when the spike was bent or twisted.f At 1 and 2 dpf, when the embryos were not hatched, growth retardation was a
rowth retardation was assessed when an embryo shows a body length below 2.8 m
f heart-beat, and all of these embryos also showed many strongalformations. All coumarin-treated embryos, which were coag-
lated at 3 dpf, showed teratogenic effects at 1 and 2 dpf. Thengerprint endpoints of coumarin were malformations of head andail (Fig. 1B) as well as growth retardation. However, 3 other end-oints were also frequently found: malformation of sacculi/otoliths33.1%), chorda (36.9%) and tail tip (29.9%; Table 6).
Except for 200 �M, all coumarin concentrations tested causedtatistically significant effects compared to controls (Table 2). Thealculated LC50 and EC50 values of coumarin were 855 and 314 �M,espectively, resulting in a teratogenicity index of 2.7. The corre-ponding concentration–response curves for coumarin are givenn Fig. 2A and B. The EC20 value, which is used for comparison touman exposure data in the discussion, was 219 �M.
.2. Exposure of zebrafish embryos to warfarin
A clear concentration–response relationship was detectable forhe percentage of embryos with lethal and teratogenic effects after
xposure to warfarin (Table 3). However, it should be noted thathere was only a marginal difference between the incidence oferatogenic/affected embryos between the 62.5 and 125 �M treat-ent groups (28.3 ± 12.6% and 31.7 ± 14.4%, respectively; Table 3).
able 2verview of lethal and teratogenic effects of coumarin in zebrafish Danio rerio embryos a
Control Coumarin test groups
Tris-buffer, 0.5% DMSO 200 �M 400 �M
Experiment no. 1 2 3 1 2 3 1
No. of teratogenic embryos – – – 6 2 2 12
No. of lethal embryos – – 1 – – – –
No. of affected embryos – – 1 6 2 2 12
No. of normal embryos 20 20 19 14 18 18 8 ∑Teratogenic embryos – 10
∑Lethal embryos 1 –
∑Affected embryos 1 10
∑Normal embryos 59 50
% Teratogenic embryos 0.0 ± 0.0a 16.7 ± 11.5 65.0% Lethal embryos 1.7 ± 2.9 0.0 ± 0.0 0.0% Affected embryos 1.7 ± 2.9 16.7 ± 11.5 65.0% Normal embryos 98.3 ± 2.9 83.3 ± 11.5 35.0
: No effects.* Significantly different from controls at p < 0.05.a Mean percentage ± SD (three experiments).
d by optical comparison to the control embryos (e.g. size, development). At 3 dpfsing analySIS software; Olympus, Hamburg, Germany).
There was no increase in the severity of teratogenic effects betweenthese two concentrations (Table 5).
At the three lowest concentrations (≤125 �M), teratogeniceffects were usually detectable at 3 dpf, whereas at 500 and1000 �M effects were seen from 2 dpf. Only at the highest con-centration could effects already be identified at 1 dpf (Table 5).Many embryos of the 1000 and 1500 �M treatment groups, whichshowed strong malformations within the first 2 days of exposure,died between 2 and 3 dpf, reaching nearly 92% embryo lethality atthe highest concentration (Table 3).
Similar to coumarin, there were also some embryos withoutheart-beat and strong malformations at the two highest concen-trations, but the majority of embryos with lethal effects at 3 dpfwere coagulated. Again, all coagulated embryos showed terato-genic effects, before they died. It is also worth mentioning thatsome embryos treated with 1500 �M warfarin showed delayeddevelopment at 1 dpf (Fig. 3) prior to severe malformations at 2 dpf.
In contrast to coumarin, malformation of the tail was identifiedas the only fingerprint endpoint of warfarin (Fig. 1C). However,together with the other frequently found endpoints (malforma-
tion of head (32.7%), chorda (46.5%), tail tip (44.7%) and growthretardation (33.3%), warfarin revealed an overall pattern of ter-atogenic effects similar to coumarin, which is quiet obvious athigher concentrations (Table 6, Fig. 4). Regarding the percentaget 3 dpf.
600 �M 800 �M 1000 �M
2 3 1 2 3 1 2 3 1 2 3
13 14 20 19 19 12 15 14 3 1 5– – – – 1 8 5 6 17 19 1513 14 20 19 20 20 20 20 20 20 207 6 – 1 – – – – – – –39 58 41 9
– 1 19 51
39 59 60 60
21 1 – –
± 5.0 96.7 ± 2.9 68.3 ± 7.6 15.0 ± 10.0 ± 0.0 1.7 ± 2.9 31.7 ± 7.6 85.0 ± 10.0
± 5.0* 98.3 ± 2.9* 100.0 ± 0.0* 100.0 ± 0.0*
± 5.0 1.7 ± 2.9 0.0 ± 0.0 0.0 ± 0.0

136 S. Weigt et al. / Reproductive Toxi
Tab
le
3O
verv
iew
of
leth
al
and
tera
toge
nic
effe
cts
of
war
fari
n
in
zebr
afish
Dan
io
reri
o
embr
yos
at
3
dp
f.
Con
trol
War
fari
n
test
grou
ps
Tris
-bu
ffer
, 0.5
%D
MSO
31.2
5
�M
62.5
�M
125
�M
250
�M
500
�M
1000
�M
1500
�M
Exp
erim
ent
No.
12
31
23
12
31
2
3
1
2
3
1
2
3
1
2
3
1 2
3N
o.
of
tera
toge
nic
embr
yos
1
2
–
2
3
2
8
6
3
8
8
3
13
9
12
17
16
12
15
9
8
5 –
–N
o.
of
leth
al
embr
yos
––
––
––
–
–
–
–
–
–
–
1
1
–
1
1
5
11
12
15
20
20N
o.
of
affe
cted
embr
yos
1
2
–
2
3
3
8
6
3
8
8
3
13
10
13
17
17
13
20
20
20
20
20
20N
o.
of
nor
mal
embr
yos
1918
20
18
17
18
12
14
17
12
12
17
7
10
7
3
3
7
–
–
– –
–
–∑
Tera
toge
nic
embr
yos
3
7
17
19
34
45
32
5∑
Leth
al
embr
yos
––
––
22
2855
∑A
ffec
ted
embr
yos
37
1719
36
47
60
60∑
Nor
mal
embr
yos
57
53
43
41
24
13
–
–
%
Tera
toge
nic
embr
yos
5.0
±
5.0a
11.7
±
2.9
28.3
±
12.6
31.7
±
14.4
56.7
±
10.4
75.0
±
13.2
53.3
±
18.9
8.3
±
14.4
%
Leth
al
embr
yos
0.0
±
0.0
0.0
±
0.0
0.0
±
0.0
0.0
±
0.0
3.3
±
2.9
3.3
±
2.9
46.7
±
18.9
91.7
±
14.4
%
Aff
ecte
d
embr
yos
5.0
±
5.0
11.7
±
2.9
28.3
±
12.6
*31
.7
±
14.4
*60
.0
±
8.7*
78.3
± 11
.5*
100.
0
±
0.0*
100.
0
±
0.0*
%
Nor
mal
embr
yos
95.0
±
5.0
88.3
±
2.9
71.7
±
12.6
68.3
±
14.4
40.0
±
8.7
21.7
± 11
.5
0.0
±
0.0
0.0
±
0.0
–:
No
effe
cts.
*Si
gnifi
can
tly
dif
fere
nt
from
con
trol
s
at
p
<
0.05
.a
Mea
n
per
cen
tage
±
SD
(th
ree
exp
erim
ents
).
cology 33 (2012) 133– 141
of affected embryos in the experiments with warfarin, differencesfrom controls were statistically significant from 62.5 �M (Table 3).Calculation of LC50 and EC50 led to values of 988 and 194 �M,respectively, with a corresponding teratogenicity index of 5.1,which is about twice as high as that one for coumarin. The cor-responding concentration–response curves for warfarin are givenin Fig. 2C and D. The EC20 value, which is used for comparison tohuman exposure data in the discussion, was 52.4 �M.
4. Discussion
In contrast to all other well-accepted human teratogens, anti-coagulant coumarin derivatives have not produced malformationsin any experimental animal species exposed prenatally [25–28].Howe and Webster described a model in the rat, which managed todemonstrate most of the features of human warfarin teratogenic-ity, but in these experiments the rats were exposed to warfarinpostnatally [68]. In the experiments with prenatal exposure of war-farin in conjunction with vitamin K supplementation Howe andWebster only detected haemorrhages and some associated changesmainly in the brain, but no other external malformations [69]. Eventhalidomide, which is famous for its strong species-specific terato-genicity, produced teratogenic effects at least in some experimentalanimal species, such as rabbits and primates [70–72]. In this con-text, the classification of warfarin as clearly teratogenic in zebrafishembryos is of outstanding importance. The teratogenic potentialof coumarin in humans is unknown/unclear [29,30], but in thezebrafish embryo assay coumarin also was shown to be clearly ter-atogenic. Both warfarin- and coumarin-exposed embryos showedsimilar effects, even if extent and frequency of the different end-points varied (Table 6).
In humans, the first trimester, especially the 6th to 9th weekof gestation, is supposed to be the critical period of exposurefor warfarin embryopathy, while the critical period of exposurefor warfarin syndrome seems to be mainly during the 2nd and3rd trimester of pregnancy [73–77]. Warfarin embryopathy refersto bone defects, and the warfarin syndrome to central nervoussystem anomalies [76]. The exposure period in the zebrafishembryo assay mainly covers the critical period of exposure forwarfarin embryopathy, but to some extent also for warfarinsyndrome. The following features of human warfarin embryopa-thy/syndrome [25,60,76–78] could also be identified in zebrafishembryos exposed to warfarin (Tables 5 and 6): developmentaldelay, growth retardation, eye defects, scoliosis and ear defects.Nasal hypoplasia, which is also characteristic of human foetusesexposed to warfarin, has no clear analogy in the zebrafish embryo,but malformation of the head can perhaps be considered as a corre-sponding effect. The chorda/spinal cord malformations observed inzebrafish embryos partly correspond to the central nervous systemanomalies known from human foetuses with warfarin syndrome.The only endpoint frequently found in humans after warfarin expo-sure, which is completely missing in zebrafish, is hypoplasia of theextremities.
The EC50 is traditionally used for calculation of the teratogenicityindex, which is just a first estimation for the teratogenic poten-tial of a substance. For the comparison to human exposure datathe EC20 was used, because experience has shown that statisti-cal significance is generally reached around the 20% effect level inthe zebrafish embryo assay (provided that the variation betweenthe replicates is not too high). Furthermore it is known that manypopular teratogens have a teratogenic risk between 15 and 30% inhumans [79].
Human therapeutic plasma concentrations of warfarin are usu-ally reported to be between 1.2 and 17.8 �M [80–82]. The EC20 ofwarfarin in the zebrafish embryo assay was 52.4 �M, which is in asimilar range of concentrations found under therapeutic conditions

S. Weigt et al. / Reproductive Toxicology 33 (2012) 133– 141 137
Fig. 1. Inverted microscope images of zebrafish embryos at 3 dpf. (A) Embryo of the control group. (B) Embryo exposed to 200 �M coumarin with malformation of head andtail. (C) Embryo exposed to 125 �M warfarin with malformation of head, eye and tail; H: head; E: eye; S/O: sacculi/otoliths; C: chorda; T: tail; TT: tail tip.
Fig. 2. Concentration–response curves used for the calculations of the EC and LC values (ToxRatPro®; probit analysis with linear maximum likelihood regression). (A) ECcurve for coumarin. (B) LC curve for coumarin. (C) EC curve for warfarin. (D) LC curve for warfarin; 95%-CV: 95%-confidence limits.

138 S. Weigt et al. / Reproductive Toxicology 33 (2012) 133– 141
F controH
(zt5afholgtIfbscieec
FtE
ig. 3. Inverted microscope images of zebrafish embryos at 1 dpf. (A) Embryo of the
: head; E: eye; S/O: sacculi/otoliths; YS: yolk sac; C: chorda; T: tail; TT: tail tip.
3 to 44-fold higher), showing a very good concordance betweenebrafish and human data. In the case of coumarin, the humanherapeutic plasma concentrations range between 3.4 nM and.1 �M [83–85]. The corresponding EC20 in the zebrafish embryossay was 219 �M, which is 43 to 64,400-fold the concentrationsound under therapeutic conditions. The ratio of the EC20 andigh-dose coumarin therapy concentrations is very close to thatf warfarin low-dose therapy. However, the ratio of the EC20 andow-dose coumarin therapy concentrations is extremely high, sug-esting that, there is a teratogenic potential but under normalherapeutic conditions, teratogenicity is not an issue for coumarin.n summary, these data confirm the teratogenic potential of war-arin, but also the questionable status of coumarin. Finally it shoulde mentioned that extrapolation of aquatic concentrations of testubstances in the zebrafish assay to human therapeutic plasmaoncentration is a bit difficult, because not only the pH value maynfluence the uptake of a test substance, but also other elements as
.g. the protein level [86]. For this reason the concentration in thembryonic tissue and the nominal concentration in the mediumould be different and measurement of the test substance in theig. 4. Inverted microscope images of zebrafish embryos at 3 dpf. (A) Embryo exposed to
ip and growth retardation. (B) Embryo exposed to 500 �M warfarin with malformation
: eye; S/O: sacculi/otoliths; C: chorda; T: tail; TT: tail tip.
l group. (B) Embryo with developmental delay after exposure to 1500 �M warfarin;
embryo would be more accurate and will be considered in futurework.
Similar to the data presented here, coumarin was also reportedto be teratogenic in the frog embryo teratogenesis assay usingXenopus (FETAX), with an EC50 value of 547 �M. Addition ofAroclor-induced rat liver microsomes decreased the FETAX EC50value 2.7-fold to 205 �M and further experiments with differentxenobiotic metabolizing enzyme inhibitors suggested CYP1A1/2-mediated activation of coumarin to a reactive epoxide metabolitewas critical [87]. The metabolite, o-hydroxyphenylacetaldehyde,which arises after CYP1A- and CYP2E-mediated epoxidation ofcoumarin and subsequent spontaneous breakdown is involved inliver carcinogenicity in the rat [16,88]. However, since rat offspringdid not show malformations after maternal exposure to coumarin[30], o-hydroxyphenylacetaldehyde does not seem to reach the ratembryos, perhaps because of its very short half-life [89]. In contrastto the hepatotoxicity and carcinogenicity, the teratogenic effects of
coumarin have not been investigated in sufficient detail.It has been assumed that teratogenicity induced by manycoumarin derivatives is due to the anticoagulant properties of this
800 �M coumarin with malformation of head, eye, sacculi/otoliths, chorda, tail, tailof head, eye, sacculi/otoliths, chorda, tail, tail tip and growth retardation; H: head;

S. Weigt et al. / Reproductive Toxicology 33 (2012) 133– 141 139
Table 4Summary of teratogenic effects of coumarin in zebrafish Danio rerio embryos.
Malformation Time Control Coumarin test groups∑
t
Tris-buffer, 0.5% DMSO 200 �M 400 �M 600 �M 800 �M 1000 �M
Head 1 dpf – 0 2 20 56 552 dpf – 3 6 39 53 413 dpf – 6 (10.0)a 19 (31.7) 47 (78.3) 41 (68.3) 9 (15.0) 122
Eyes 1 dpf – 0 0 0 17 332 dpf – 0 0 0 22 303 dpf – 0 (0.0) 1 (1.7) 1 (1.7) 15 (25.0) 4 (6.6) 21
Sacculi/otoliths 2 dpf – 1 3 11 39 353 dpf – 0 (0.0) 3 (5.0) 12 (20.0) 30 (50.0) 7 (11.7) 52
Chorda 1 dpf – 0 0 1 18 382 dpf – 0 2 11 41 363 dpf – 3 (5.0) 6 (10.0) 13 (21.7) 29 (48.3) 7 (11.7) 58
Tail 1 dpf – 0 2 27 59 552 dpf – 0 2 6 47 363 dpf – 3 (5.0) 28 (46.7) 57 (95.0) 39 (65.0) 9 (15.0) 136
Tail tip 1 dpf – 0 0 1 21 392 dpf – 0 2 4 27 303 dpf – 0 (0.0) 5 (8.3) 9 (15.0) 18 (30.0) 6 (10.0) 38
Scoliosis 1 dpf – 0 0 0 5 152 dpf – 0 0 0 9 233 dpf – 0 (0.0) 0 (0.0) 0 (0.0) 6 (10.0) 4 (6.7) 10
Yolk deformity 1 dpf – 0 0 0 0 02 dpf – 0 0 0 0 03 dpf – 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0
Growth retardation 1 dpf – 0 0 6 51 542 dpf – 0 1 4 22 313 dpf – 1 (1.7) 16 (26.7) 59 (98.3) 40 (66.7) 9 (15.0) 125
Sum teratogenic embryosb 3 dpf – 10 39 58 41 9 157
–: No effects;∑
t: sum of teratogenic embryos per endpoint out of all teratogenic embryos in the three test groups at 72 hpf; one embryo can display multiple teratogenicmalformations.
a Number of embryos displaying this endpoint (%).b Sum of the teratogenic embryos per group at 3 dpf of three experiments.
Table 5Summary of teratogenic effects of warfarin in zebrafish Danio rerio embryos.
Malformation Time Control Warfarin test groups∑
t
Tris-buffer, 0.5% DMSO 31.25 �M 62.5 �M 125 �M 250 �M 500 �M 1000 �M 1500 �M
Head 1 dpf – 0 1 1 2 2 5 192 dpf – 0 3 1 7 6 37 423 dpf – 2 (3.3)a 2 (3.3) 3 (5.0) 10 (16.7) 8 (13.3) 23 (38.3) 4 (6.7) 52
Eyes 1 dpf – 0 1 0 2 1 4 42 dpf – 0 1 0 1 0 2 73 dpf – 0 (0.0) 2 (3.3) 0 (0.0) 1 (1.7) 0 (0.0) 2 (3.3) 1 (1.7) 6
Sacculi/otoliths 2 dpf – 0 3 0 2 2 28 433 dpf – 0 (0.0) 2 (3.3) 1 (1.7) 3 (5.0) 2 (3.3) 18 (30.0) 3 (5.0) 29
Chorda 1 dpf – 0 1 0 2 3 8 142 dpf – 0 1 0 3 0 25 423 dpf – 2 (3.3) 6 (10.0) 3 (5.0) 15 (25.0) 14 (23.3) 29 (48.3) 5 (8.3) 74
Tail 1 dpf – 0 1 0 2 2 5 142 dpf – 0 1 0 6 4 31 433 dpf 3 (5.0) 6 (10.0) 8 (13.3) 18 (30.0) 28 (46.7) 37 (61.7) 36 (60.0) 5 (8.3) 138
Tail tip 1 dpf – 0 1 0 2 2 4 32 dpf – 0 1 0 4 3 15 233 dpf 1 (1.7) 0 (0.0) 5 (8.3) 3 (5.0) 16 (26.7) 15 (25.0) 27 (45.0) 5 (8.3) 71
Scoliosis 1 dpf – 0 1 0 1 1 1 02 dpf – 0 1 0 0 0 1 03 dpf – 0 (0.0) 2 (3.3) 0 (0.0) 2 (3.3) 0 (0.0) 0 (0.0) 0 (0.0) 4
Yolk deformity 1 dpf – 0 0 0 0 0 0 02 dpf – 0 0 0 0 0 0 03 dpf – 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0
Growth retardation 1 dpf – 0 1 0 2 1 4 52 dpf – 0 1 0 2 0 13 363 dpf – 1 (1.7) 2 (3.3) 1 (1.7) 11 (18.3) 9 (15.0) 24 (40.0) 5 (8.3) 53
Sum teratogenic embryosb 3 dpf 3 7 17 19 34 45 32 5 159
–: No effects;∑
t: sum of teratogenic embryos per endpoint out of all teratogenic embryos in the three test groups at 72 hpf; one embryo can display multiple teratogenicmalformations.
a Number of embryos displaying this endpoint (%).b Sum of the teratogenic embryos per group at 3 dpf of three experiments.

140 S. Weigt et al. / Reproductive Toxi
Table 6Frequency of endpoints of coumarin and warfarin at 3 dpf.
Malformation Coumarin Warfarin∑
t [%]∑
t [%]
Head 122a 77.7b 52 32.7Eyes 21 13.4 6 3.8Sacculi/otoliths 52 33.1 29 18.2Chorda 58 36.9 74 46.5Tail 136 86.6 138 86.8Tail tip 38 29.9 71 44.7Scoliosis 10 6.4 4 2.5Yolk deformity 0 0.0 0 0.0Growth retardation 125 79.6 53 33.3
Major endpoints given in bold.a Sum of embryos with teratogenic effects displaying this endpoint (in all exper-
iments).b
es
saictgptshcacwo
oedim
C
A
eW(s
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[DEC on filaritic lymphoedema and elephantiasis in India. Preliminary results.
Sum of embryos with teratogenic effects displaying this endpoint/sum ofmbryos with teratogenic effects detected in all concentrations of the given testubstance.
ubstance class [63]. In contrast to warfarin and other coumarinnticoagulants, standard and low molecular weight heparin, whichs used as an anticoagulant during pregnancy, cannot pass the pla-ental barrier and do not reach the embryo [90–94]. Assuminghat anticoagulant properties are directly connected with terato-enicity, coumarin or coumarin derivates without anticoagulantroperties should not to be teratogenic or should produce othereratogenic effects than those known from anticoagulant expo-ure. The zebrafish embryo data presented seem to contradict thisypothesis and might have multiple possible explanations. In thisontext two main questions have to be considered: (i) Are the ter-togenic effects of warfarin and coumarin in zebrafish embryosaused by the same mechanism? (ii) Are the teratogenic effects ofarfarin in zebrafish embryos and humans caused by one common
r two different mechanisms?However, clarification of these aspects goes beyond the scope
f this communication and should be considered in separatexperiments using adequate molecular techniques. As coumarinerivatives are used for different therapeutic indications, it is
mportant to clarify if the teratogenic activity is due to the coumarinoiety or due to the anticoagulant properties alone.
onflict of interest
None.
cknowledgments
The authors wish to thank Julia Pohl and Andreas Gado forxcellent technical assistance. We also want to thank Dr. Brigittealderich from the Max Planck Institute of Developmental Biology
Tübingen, Germany) for the breeding stock of wild type tuebingentrain zebrafish.
eferences
[1] Bachmann J. Entwicklung und Erprobung eines Teratogenitäts-ScreeningTestes mit Embryonen des Zebrabärblings Danio rerio. Ph.D. Thesis. TechnicalUniversity Dresden, Germany; 2002.
[2] Brannen KC, Panzica-Kelly JM, Danberry TL, Augustine-Rauch KA. Developmentof a zebrafish embryo teratogenicity assay and quantitative prediction model.Birth Defects Res B Dev Reprod Toxicol 2010;89(1):66–77.
[3] Chakraborty C, Hsu CH, Wen ZH, Lin CS. Agoramoorthy G. Zebrafish: a completeanimal model for in vivo drug discovery and development. Curr Drug Metab
2009;10(2):116–24.[4] Hill AJ, Teraoka H, Heideman W, Peterson RE. Zebrafish as a model vertebratefor investigating chemical toxicity. Toxicol Sci 2005;86(1):6–19.
[5] Loucks E, Carvan 3rd MJ. Strain-dependent effects of developmental ethanolexposure in zebrafish. Neurotoxicol Teratol 2004;26(6):745–55.
[
cology 33 (2012) 133– 141
[6] McGrath P, Li CQ. Zebrafish: a predictive model for assessing drug-inducedtoxicity. Drug Discov Today 2008;13(9–10):394–401.
[7] Nagel R. DarT: The embryo test with the zebrafish Danio rerio—a general modelin ecotoxicology and toxicology. ALTEX 2002;19(Suppl. 1):38–48.
[8] Selderslaghs IW, Hooyberghs J, De Coen W, Witters HE. Locomotor activityin zebrafish embryos: a new method to assess developmental neurotoxicity.Neurotoxicol Teratol 2010;32(4):460–71.
[9] Selderslaghs IW, Van Rompay AR, De Coen W, Witters HE. Development of ascreening assay to identify teratogenic and embryotoxic chemicals using thezebrafish embryo. Reprod Toxicol 2009;28(3):308–20.
10] Ton C, Lin Y, Willett C. Zebrafish as a model for developmental neurotoxicitytesting. Birth Defects Res A Clin Mol Teratol 2006;76(7):553–67.
11] Yang L, Ho NY, Alshut R, Legradi J, Weiss C, Reischl M, et al. Zebrafish embryos asmodels for embryotoxic and teratological effects of chemicals. Reprod Toxicol2009;28(2):245–53.
12] Busquet F, Nagel R, von Landenberg F, Mueller SO, Huebler N, Broschard TH.Development of a new screening assay to identify proteratogenic substancesusing zebrafish Danio rerio embryo combined with an exogenous mammalianmetabolic activation system (mDarT). Toxicol Sci 2008;104(1):177–88.
13] Weigt S, Huebler N, Braunbeck T, von Landenberg F, Broschard TH. Zebrafishteratogenicity test with metabolic activation (mDarT): effects of phase Iactivation of acetaminophen on zebrafish Danio rerio embryos. Toxicology2010;275(1–3):36–49.
14] Weigt S, Huebler N, Strecker R, Braunbeck T, Broschard TH. Zebrafish(Danio rerio) embryos as a model for testing proteratogens. Toxicology2011;281(1–3):25–36.
15] Kaminsky LS, Zhang ZY. Human P450 metabolism of warfarin. Pharmacol Ther1997;73(1):67–74.
16] Lake BG. Coumarin metabolism, toxicity and carcinogenicity: relevance forhuman risk assessment. Food Chem Toxicol 1999;37(4):423–53.
17] Miller GP, Jones DR, Sullivan SZ, Mazur A, Owen SN, Mitchell NC, et al. Assessingcytochrome P450 and UDP-glucuronosyltransferase contributions to warfarinmetabolism in humans. Chem Res Toxicol 2009;22(7):1239–45.
18] Ishizuka M, Okajima F, Tanikawa T, Min H, Tanaka KD, Sakamoto KQ, et al.Elevated warfarin metabolism in warfarin-resistant roof rats (Rattus rattus) inTokyo. Drug Metab Dispos 2007;35(1):62–6.
19] Guengerich FP, Dannan GA, Wright ST, Martin MV, Kaminsky LS. Purificationand characterization of liver microsomal cytochromes p-450: electrophoretic,spectral, catalytic, and immunochemical properties and inducibility ofeight isozymes isolated from rats treated with phenobarbital or beta-naphthoflavone. Biochemistry 1982;21(23):6019–30.
20] Guo Y, Weller P, Farrell E, Cheung P, Fitch B, Clark D, et al. In silico pharmaco-genetics of warfarin metabolism. Nat Biotechnol 2006;24(5):531–6.
21] Hirsh J, Dalen J, Anderson DR, Poller L, Bussey H, Ansell J, et al. Oral antico-agulants: mechanism of action, clinical effectiveness, and optimal therapeuticrange. Chest 2001;119(1 Suppl):8S–21S.
22] Wadelius M, Pirmohamed M. Pharmacogenetics of warfarin: current status andfuture challenges. Pharmacogenomics J 2007;7(2):99–111.
23] Zielinska A, Lichti CF, Bratton S, Mitchell NC, Gallus-Zawada A, Le VH, et al.Glucuronidation of monohydroxylated warfarin metabolites by human livermicrosomes and human recombinant UDP-glucuronosyltransferases. J Phar-macol Exp Ther 2008;324(1):139–48.
24] Rubinstein AL. Zebrafish assays for drug toxicity screening. Expert Opin DrugMetab Toxicol 2006;2(2):231–40.
25] Brent RL, Beckman DA. Environmental teratogens. Bull N Y Acad Med1990;66(2):123–63.
26] Nau H. Species differences in pharmacokinetics and drug teratogenesis. EnvironHealth Perspect 1986;70:113–29.
27] Schardein JL, Schwetz BA, Kenel MF. Species sensitivities and prediction ofteratogenic potential. Environ Health Perspect 1985;61:55–67.
28] Shepard TH, Fantel AG, Mirkes PE, Greenaway JC, Faustman-Watts E, CampbellM, et al. Teratology testing: I. Development and status of short-term prescreens.II. Biotransformation of teratogens as studied in whole embryo culture. ProgClin Biol Res 1983;135:147–64.
29] Shepard TH, Lemire RJ. Catalog of teratogenic agents. 11th ed. Baltimore: TheJohns Hopkins University Press; 2004.
30] Friedman JM, Polifka JE. Teratogenic effects of drugs, a resource for clinicans(TERIS). 2nd ed. Baltimore and London: The John Hopkins University Press;2000.
31] McKnight JT, Maxwell AJ, Anderson RL. Warfarin necrosis. Arch Fam Med1992;1(1):105–8.
32] Merkel U, Sigusch H, Hoffmann A. Grapefruit juice inhibits 7-hydroxylationof coumarin in healthy volunteers. Eur J Clin Pharmacol 1994;46(2):175–7.
33] Marshall ME, Mohler JL, Edmonds K, Williams B, Butler K, Ryles M, et al. Anupdated review of the clinical development of coumarin (1,2-benzopyrone)and 7-hydroxycoumarin. J Cancer Res Clin Oncol 1994;120(Suppl.):S39–42.
34] Mohler JL, Williams BT, Thompson IM, Marshall ME. Coumarin (1,2-benzopyrone) for the treatment of prostatic carcinoma. J Cancer Res Clin Oncol1994;120(Suppl.):S35–8.
35] Jamal S, Casley-Smith JR. The effects of 5,6 benzo-[a]-pyrone (coumarin) and
Ann Trop Med Parasitol 1989;83(3):287–90.36] Egan D, O’Kennedy R, Moran E, Cox D, Prosser E, Thornes RD. The pharmacol-
ogy, metabolism, analysis, and applications of coumarin and coumarin-relatedcompounds. Drug Metab Rev 1990;22(5):503–29.

e Toxi
[
[
[
[
[
[
[[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
S. Weigt et al. / Reproductiv
37] Abraham K, Wohrlin F, Lindtner O, Heinemeyer G, Lampen A. Toxicologyand risk assessment of coumarin: focus on human data. Mol Nutr Food Res2010;54(2):228–39.
38] Guengerich FP. Characterization of human cytochrome P450 enzymes. FASEB J1992;6(2):745–8.
39] Guengerich FP. Cytochromes P450, drugs, and diseases. Mol Interv2003;3(4):194–204.
40] Lewis DF, Lake BG. Species differences in coumarin metabolism: a molec-ular modelling evaluation of CYP2A interactions. Xenobiotica 2002;32(7):547–61.
41] Rojas JC, Aguilar B, Rodriguez-Maldonado E, Collados MT. Pharmacogenetics oforal anticoagulants. Blood Coagul Fibrinolysis 2005;16(6):389–98.
42] Freude S, Pabinger-Fasching I, Kozel-Lachmann D, Braun F, Pollak A. Warfarinembryopathy in maternal coumarin therapy for protein S deficiency. PadiatrPadol 1991;26(5):239–41.
43] Hirsh J. Oral anticoagulant drugs. N Engl J Med 1991;324(26):1865–75.44] Breckenridge AM. Interindividual differences in the response to oral anticoag-
ulants. Drugs 1977;14(5):367–75.45] Chan E, McLachlan A, O’Reilly R, Rowland M. Stereochemical aspects of warfarin
drug interactions: use of a combined pharmacokinetic–pharmacodynamicmodel. Clin Pharmacol Ther 1994;56(3):286–94.
46] Pitsiu M, Parker EM, Aarons L, Rowland M. A Bayesian method based on clottingfactor activity for the prediction of maintenance warfarin dosage regimens.Ther Drug Monit 2003;25(1):36–40.
47] Rettie AE, Korzekwa KR, Kunze KL, Lawrence RF, Eddy AC, Aoyama T, et al.Hydroxylation of warfarin by human cDNA-expressed cytochrome P-450: arole for P-4502C9 in the etiology of (S)-warfarin-drug interactions. Chem ResToxicol 1992;5(1):54–9.
48] Zhang Z, Fasco MJ, Huang Z, Guengerich FP, Kaminsky LS. Human cytochromesP4501A1 and P4501A2: R-warfarin metabolism as a probe. Drug Metab Dispos1995;23(12):1339–46.
49] Grossman SJ, Herold EG, Drey JM, Alberts DW, Umbenhauer DR, Patrick DH, et al.CYP1A1 specificity of Verlukast epoxidation in mice, rats, rhesus monkeys, andhumans. Drug Metab Dispos 1993;21(6):1029–36.
50] Freeman BD, Zehnbauer BA, McGrath S, Borecki I, Buchman TG. CytochromeP450 polymorphisms are associated with reduced warfarin dose. Surgery2000;128(2):281–5.
51] Huang W, Lin YS, McConn 2nd DJ, Calamia JC, Totah RA, Isoherranen N, et al.Evidence of significant contribution from CYP3A5 to hepatic drug metabolism.Drug Metab Dispos 2004;32(12):1434–45.
52] Bell RG, Matschiner JT. Warfarin and the inhibition of vitamin K activity by anoxide metabolite. Nature 1972;237(5349):32–3.
53] Bell RG, Sadowski JA, Matschiner JT. Mechanism of action of warfarin. Warfarinand metabolism of vitamin K1. Biochemistry 1972;11(10):1959–61.
54] Li T, Chang CY, Jin DY, Lin PJ, Khvorova A, Stafford DW. Identification of the genefor vitamin K epoxide reductase. Nature 2004;427(6974):541–4.
55] Rieder MJ, Reiner AP, Gage BF, Nickerson DA, Eby CS, McLeod HL, et al. Effectof VKORC1 haplotypes on transcriptional regulation and warfarin dose. N EnglJ Med 2005;352(22):2285–93.
56] Rost S, Fregin A, Ivaskevicius V, Conzelmann E, Hortnagel K, Pelz HJ, et al. Muta-tions in VKORC1 cause warfarin resistance and multiple coagulation factordeficiency type 2. Nature 2004;427(6974):537–41.
57] Geisen C, Watzka M, Sittinger K, Steffens M, Daugela L, Seifried E, et al. VKORC1haplotypes and their impact on the inter-individual and inter-ethnical variabil-ity of oral anticoagulation. Thromb Haemost 2005;94(4):773–9.
58] DiSaia PJ. Pregnancy and delivery of a patient with a Starr-Edwardson mitralvalve prothesis: report of a case. Obstet Gynecol 1966;29:469–72.
59] Kerber IJ, Warr 3rd OS, Richardson C. Pregnancy in a patient with a prostheticmitral valve. Associated with a fetal anomaly attributed to warfarin sodium.JAMA 1968;203(3):223–5.
60] Briggs GG, Freeman RK, Yaffe SJ. A Reference guide to fetal and neonatal risk– drugs in pregnancy and lactation. 8th ed. Hong Kong: Lippincott Williams &Wilkins; 2008.
61] Lewis RJ. Reproductive active chemicals – a reference guide. New York: VanNostrand Reinhold; 1991.
62] Schardein JL, Macina OT. Human developmental toxicants: aspects of toxicol-ogy and chemistry. 1st ed. Boca Raton: CRC Press; 2007.
63] Pauli RM, Lian JB, Mosher DF, Suttie JW. Association of congenital deficiencyof multiple vitamin K-dependent coagulation factors and the phenotype of thewarfarin embryopathy: clues to the mechanism of teratogenicity of coumarinderivatives. Am J Hum Genet 1987;41(4):566–83.
64] OECD. OECD draft proposal for a new guideline, 1st version. Guideline for thetesting of chemicals. Fish Embryo Toxicity, FET Test; 2006.
65] Westerfield M. The zebrafish book: a guide for the laboratory use of zebrafish,
Danio rerio. 4th ed. Eugene: University of Oregon Press; 2000.66] Augustine-Rauch K, Zhang CX, Panzica-Kelly JM. In vitro developmental toxicol-ogy assays: a review of the state of the science of rodent and zebrafish wholeembryo culture and embryonic stem cell assays. Birth Defects Res C EmbryoToday 2010;90(2):87–98.
[
[
cology 33 (2012) 133– 141 141
67] Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embry-onic development of the zebrafish. Dev Dynam 1995;203(3):253–310.
68] Howe AM, Webster WS. The warfarin embryopathy: a rat model show-ing maxillonasal hypoplasia and other skeletal disturbances. Teratology1992;46(4):379–90.
69] Howe AM, Webster WS. Exposure of the pregnant rat to warfarin and vitaminK1: an animal model of intraventricular hemorrhage in the fetus. Teratology1990;42(4):413–20.
70] Schumacher H, Blake DA, Gurian JM, Gillette JR. A comparison of the ter-atogenic activity of thalidomide in rabbits and rats. J Pharmacol Exp Ther1968;160(1):189–200.
71] Szabo KT, Steelman RL. Effects of maternal thalidomide treatment on preg-nancy, fetal development, and mortality of the offspring in random-bred mice.Am J Vet Res 1967;28(127):1823–8.
72] Teo SK, Colburn WA, Tracewell WG, Kook KA, Stirling DI, JaworskyMS, et al. Clinical pharmacokinetics of thalidomide. Clin Pharmacokinet2004;43(5):311–27.
73] Holzgreve W, Carey JC, Hall BD. Warfarin-induced fetal abnormalities. Lancet1976;2(7991):914–5.
74] Hall JG, Pauli RM, Wilson KM. Maternal and fetal sequelae of anticoagulationduring pregnancy. Am J Med 1980;68(1):122–40.
75] Kaplan LC. Congenital Dandy Walker malformation associated with firsttrimester warfarin: a case report and literature review. Teratology1985;32(3):333–7.
76] Van Driel D, Wesseling J, Rosendaal FR, Odink RJ, Van der Veer E, Gerver WJ,et al. Growth until puberty after in utero exposure to coumarins. Am J MedGenet 2000;95(5):438–43.
77] Gärtner BC, Seifert CB, Michalk DV, Roth B. Phenprocoumon therapyduring pregnancy: case report and comparison of the teratogenic riskof different coumarin derivatives. Z Geburtshilfe Perinatol 1993;197(6):262–5.
78] Hetzel PG, Glanzmann R, Hasler PW, Ladewick A, Buhrer C. Coumarinembryopathy in an extremely low birth weight infant associated withneonatal hepatitis and ocular malformations. Eur J Pediatr 2006;165(6):358–60.
79] Banhidy F, Lowry RB, Czeizel AE. Risk and benefit of drug use during pregnancy.Int J Med Sci 2005;2(3):100–6.
80] Zacchigna M, Di Luca G, Cateni F, Maurich V. Improvement of warfarin bio-pharmaceutics by conjugation with poly(ethylene glycol). Eur J Pharm Sci2004;23(4–5):379–84.
81] Vaz-da-Silva M, Almeida L, Falcao A, Soares E, Maia J, Nunes T, et al. Effectof eslicarbazepine acetate on the steady-state pharmacokinetics and pharma-codynamics of warfarin in healthy subjects during a three-stage, open-label,multiple-dose, single-period study. Clin Ther 2010;32(1):179–92.
82] Sun S, Wang M, Su L, Li J, Li H, Gu D. Study on warfarin plasma concentrationand its correlation with international normalized ratio. J Pharm Biomed Anal2006;42(2):218–22.
83] Lamiable D, Vistelle R, Trenque T, Fay R, Millart H, Choisy H. Sensitivehigh-performance liquid chromatographic method for the determination ofcoumarin in plasma. J Chromatogr 1993;620(2):273–7.
84] Ritschel WA, Brady ME, Tan HS, Hoffmann KA, Yiu IM, Grummich KW.Pharmacokinetics of coumarin and its 7-hydroxy-metabolites upon intra-venous and peroral administration of coumarin in man. Eur J Clin Pharmacol1977;12(6):457–61.
85] Ritschel WA. Therapeutic concentration of coumarin and predicted dosage reg-imens. Arzneimittelforschung 1984;34(8):907–10.
86] Kramer NI, van Eijkeren JCH, Hermens JLM. Influence of albumin on sorptionkinetics in solid-phase microextration: consequences for chemical analysesand uptake processes. Anal Chem 2007;79:6941–8.
87] Fort DJ, Stover EL, Propst T, Hull MA, Bantle JA. Evaluation of the developmen-tal toxicities of coumarin, 4-hydroxycoumarin, and 7-hydroxycoumarin usingFETAX. Drug Chem Toxicol 1998;21(1):15–26.
88] Vassallo JD, Hicks SM, Daston GP, Lehman-McKeeman LD. Metabolic detoxi-fication determines species differences in coumarin-induced hepatotoxicity.Toxicol Sci 2004;80(2):249–57.
89] EFSA. Coumarin in flavourings and other food incrediants with flavouring prop-erties. Scientific opinion on the panel on food additives, flavourings, processingaids and material in contact with food (ACF). EFSA J 2008;793:1–15.
90] Harbison RD. Chemical–biological reactions common to teratogenesis andmutagenesis. Environ Health Perspect 1978;24:87–100.
91] Andrew M, Boneu B, Cade J, Cerskus AL, Hirsh J, Jefferies A, et al. Placentaltransport of low molecular weight heparin in the pregnant sheep. Br J Haematol1985;59(1):103–8.
92] Flessa HC, Kapstrom AB, Glueck HI, Will JJ. Placental transport of heparin. Am J
Obstet Gynecol 1965;93(4):570–3.93] Hirsh J, Cade JF, O’Sullivan EF. Clinical experience with anticoagulant therapyduring pregnancy. Br Med J 1970;1(5691):270–3.
94] Pieper PG, Balci A, Van Dijk AP. Pregnancy in women with prosthetic heartvalves. Neth Heart J 2008;16(12):406–11.