determination of sample sizes for the estimation of onchocerca volvulus (filarioidea: onchocercidae)...
TRANSCRIPT

Vol. 35, No. 5 September 1998
RAPID COMMUNICATIONS
Establishment of the Tortoise Tick Amblyomma marmoreum (Acari: Ixodidae) on a Reptile-BreedingFacility in Florida.
SANDRA A. ALLAN, LEIGH-ANNE SIMMONS, AND MICHAEL J. BfRRIDGE
Experimental Infection of Domestic Cats with Banonella henselae by Inoculation of Ctenocepha]idesfelis (Siphonaptera: Pulicidae) Feces.LANE FOIL, EARL ANDRESS, REBECCA L. FREE-LAND, ALMA F. Roy, ROXANNE RLTLKUIE^PATRICIA C. TRICHE, AND KATHV L. 0’REILLV............................^..2</^...................^..625/
FORUM
Reported Distribution of Ixodes scapularis and I.wdes pacificus (Acari: Ixodidae) in the United States.DAVID T. DENNIS, TRLDI S. NEKOMOTO, JOHN C. VICTOR, WILLIAM S. PALL, AND JoSEPtPlESMAN........................................................................................../:^^................ ^
ARTICLES
Effect of Body Size on Host Seeking and Blood Meal Utilization in Anopheles gambiae sensu stricto(Diptera: Culicidae): the Disadvantage of Being Small. /. ^-/W- Tik-KFN. M- T- KinwnFN- d M f / / ’/ }W. TAKKEN, M. J. KLOWDEN, AND G. M. CHAMBERS. :z.^...
New Procedures to Estimate Water Temperatures and Water Depths for Application in Climate-DengueModeling. /; ’’/’’.’/’ ,^^\SHOLQLAN CHENG, LALRENCE S. KALKSTEIN, DANA A. FOCKS, AND ALSTINE NNAJI ........./646
Minimum Infection Rate of Anibylomma americaniini (Acari: Ixodidae) by Ehrlichia chaffeensis(Rickettsiales: Ehrlichieae in Southern Indiana.CHRISTOPHER T. Bl RKET, CAROLYN N. VANN, ROBERT R. PINGER, CLARE L. CHATOT, AND FME. STEINER............................................................................................^^..^2.^.
(Continued on inside front pages)
Published by

1 -7 6 0- 0 ^Determination of Sample Sizes for the Estimation of Onchocercavolvulus (Filarioidea: Onchocercidae) Infection Rates in Biting
Populations of Simulium ochraceum s.l. (Diptera: Simuliidae) and ItsApplication to Ivermectin Control Programs
MARIA-GLORIA BASAXEZ,’ 2 MARIO ALBERTO RODRTGUEZ-PEREZ,34
FILIBERTO REYES-VILLANUEVA,5 RICHARD C. COLLINS,"AND MARIO HENRY RODRIGUEZ4
El Colegio de la Frontera Sur, Carretera Panamericana y Periferico Sur, San Cristobal de las Casas, Chiapas 29290
Meriw Centro de Investigacion del Paludismo, Institute) Nacional de Salud Publica, 4a. Ave. Norte 19 Ave. Pomente,
Apdo. Postal 53- Tapachula Chiapas 30700 Mexico, and Wellcome Trust Centre for Epidemiology of Infectious Disease,
’Department of Zoology, University of Oxford, South Parks Road, Oxford 0X1 .3PS, UK
J. Mod. Entomol. 35(5): -45-75- (1998)
ABSTRACT Monthly samples ofbiting SilHiJiiim ochraceiim s.l. Walker were collected before and
after ivermectin treatment in southern Mexico and analyzed for Onchocerca cohliliis Leuckart
infection rates infection intensity, and the characteristics of larval distribution among parous flies.
The variance over mean ratio (VMfi) indicated that in all cases this distribution departed from
Poisson and was strongly aggregated (VMR > 1). The negative binomial found to be an adequate
model with a small value of the aggregation parameter k. but the degree of larval overdispersion
increased as the mean larval load decreased, invalidating the use of common k,. value. A linear
relationship between k and the mean (^i) was established, fc(/i) k,^. which permitted exploration
of the relationship between the observed proportion of infected flies, p, and the estimated mean
larval burden per flv, in (all larval stages in parous Hies). This would allow mean numbers of larvae
per parous flv to be predicted from presence-absence data (e.g., from infection rates provided by
polymerase chain reaction methods applied to pools of flies), assuming that k, is known parameter.
Given that both p and are naturally low in S. ochraceiim, their relationship practical y linear
within the range of observed values. Predictions were tested with the Mexican data from winch the
clumping parameter was estimated well for Guatemalan data for which this information
not available Results showed a highly satisfactory degree of agreement between predictions and ob-
servations The sample sizes required to estimate mean larval loads from prevalence data for fixed levels
of precision (defined as the ratio between SE[m] and m)were calculated for realistic S. oc?irocptii
infection rates (those found in published pre- and postcontrol field surveys well as in this work). For
the special case in which the relationship between k and the is linear and goes through the origin,
k(li) fci/i. the number of flies to be examined for 0 mk-tillis infections does not explicitly depend
the aggregation parameter, but rather on the unknown proportion of infected flies. Practical recom-
mendations for the calculation of sample sizes are discussed. For infection levels <0.2%, minimum
"umber between 6,000 and 13,000 parous flies would have to be examined to estimate the larval
load with precision between 0.20 and 0.30. The linearity between onchocercal infection rate and
infection intensity in the flv population indicates that relationships between the former and onchocer-ciasis patterns in the human population should be further explored for the purposes of monitoring the
impact of ivermectin control programs through entomological evaluations.
KEY WORDS Onchocerca roh’iihis, Simulitim ochraceum s.l
intensity, sample size
ivermectin, infection rate, infection
’’FO^T" "^roceum s.l. Walker is the primary vector^erca wkuhis Leuckart in southern Mexico
^"Ii.in"’’TnJ’’* ^"tre for the Epidemiology of Infectious Dis-
./"’"ro"" "’’Zwiogy, University ofOxford, Oxford 0X1 3PS, UK.’’""lie., "’""""’o para Investigacion y Control de Enfennedades
’.1- Pinm ":’rt ^acucho, Amazonas -101, Venezuela.:^T ^"’ of Biological Sciences, University ofSalford, Salford
"’’^io1’ ’’"’’"’"e-’rione’; sobre Enfennedades Infecciosas. In-
" "f Salud Puhlica, Cuemavaca, Morelos 62508 Mexico.
and Guatemala. With the advent of ivermectin, con-
trol programs based on mass distribution of this mi-
crofilaricide have been started in Latin America underthe auspices of the Onchocerciasis Elimination Pro-gram for the Americas. Even before the administration
Universidad Autonoma de Nuevo Ledn, Facnit.id de Ciencias
Biol6gicas. San Nicolas de lo.s Garza, N.L. 664.50, Mexico.
Departments of Entomology and Veterinary Science. College ofAgriculture. University of Arizona, Tucson, AZ 85-21.
0022-2585/98/0745-OT57S02.00/0 ’(C) 1998 Entomological Society of America

JOURNAL OF MEDICAL ENTOMOLOGY746
of ivermectin, infection rates ofS. ochraceiim s.l. with
filarial larvae indistinguishable from 0. volvulus werenaturally low. This probably is caused by the relatively
low vector competence of this simuliid (it possesses a
well-developed cibarial armature that destroys a sig-
nificant proportion of the ingested microfilariae [mf][Omar and Garms 1975, Basanez et al. 1995]), and to
the relatively long extrinsic incubation period of the
nematode in comparison to the life expectancy of the
vector (Collins et al. 1992, Basanez et al. 1996). Re-
petitive ivermectin treatment has been shown to de-
crease even further infection rates in S. ochraceum
populations (Cupp 1992; Cupp et al. 1989, 1992;
Rodriguez-Perez et al. 1995).Entomological indices of onchocerciasis transmis-
sion intensity include measurements of the annual
biting rate, annual infective biting rate, and annualtransmission potential (Duke 1968, Walsh et al. 1978).Control programs based on chemotherapy alonewould affect only the latter 2 parameters. The annualinfective biting rate requires the estimation of the
proportion of infective flies in the vector sample,whereas the annual transmission potential requires
the estimation of the mean number of infective larvae
(L.,) per fly. Because mass delivery of ivermectin af-
fects the microfilarial load in the skin of the human
host population and therefore parasite uptake by the
simuliid vector (Cupp et al. 1989, Basanez et al. 1994),it has been suggested that entomological indicators of
the impact of ivermectin on transmission be based onthe total number of 0, colvidiis larvae (all stages)rather than on detection of Lg only. In this way,
changes in infection rates (rather than in infectivity
rates alone) would reflect levels of ivermectin cover-
age (Cupp et al. 1992).Detection ofnatural infection levels in simuliid pop-
ulations consists of the dissection of thousands of bit-
ing flies, first to detect parity and then infection. Dis-
section methods to determine infection are extremelytime-consuming and labor-intensive. In some situa-
tions specific identification of the parasite becomesdifficult; simuliids may act as vectors of >1 filarial
species in the same geographical region (Garms 1985,
1987). Novel methods include the species-specific de-tection of 0. volvulus larvae in pools of flies by means
of polymerase chain reaction (PCR)-based DNA
probes (Meredith et al. 1991, Toe et al. 1994, Katholi
et al. 1995, Oskam et al. 1996). The drawback of this
technique is that although it is specific, it only permits
calculation of the proportion of flies infected and not
of the mean larval load per fly. Because in onchocer-ciasis, as well as in many other macroparasitic infec-
tions, parasite load correlates better with morbidity in
host populations than infection rates (Anderson and
May 1985), an estimation of larval burden per fly
would be desirable when monitoring chemotherapy
control programs by entomological methods.Whether dissections of individual flies or DNA-
probes on pools of flies are to be adopted by thenational control programs of Mexico and Guatemalafor the entomological evaluation of the effect of iver-
mectin on transmission, it will be necessary to know
^rf ...Vol.
how many flies must be collected and anaLdetect changing infection levels in the vectg,^lation. Methods for estimating sample sizes ^’,mological surveys may depend on the underly, ^tribution of the number of individuals per 5^’.unit (Gerrard and Chiang 1970, Elliot 197 -.H1986). In the case of filarial infections in b) ’^(Simuliidae), the sampling unit is the fly ^,^.assumptions will have to be made regarding A*tribution of the numbers of larvae per fly, -n^Hbution generally is aggregated strongly (n^harbor none or few larvae, whereas a few Bies L"a heavy larval load) and therefore may be des^"by the negative binomial (Cheke et al. 1982,and Cheke 1985, Basanez et al. 1995). For the n^-binomial, there are mathematical expressions\Qscribe the relationship between the proportion }dividuals infected (prevalence or infection rate).1the mean number of parasites per host (inf^intensity) (Pielou 1960, Anderson 1987), This,’tionship could, in principle, permit estimation o(,mean number of larvae per fly, if the fraction o(fected flies in the sample is determined and the’gregation parameter has been estimated from asfcof preliminary samples considered to be represstive of those for which predictions will be later n^The idea of estimating mean population values f^,presence-absence data is not new, and the Istrep^date back to the late 1910s when these methods^applied to bacterial growth (McCrady 1915) andpi^ecology (Gleason 1920). It was assumed that the^tribution of such organisms was mainly random nhence adequately described by the Poisson modelThe objectives ofour study were to apply preseiio
absence sampling procedures to the detection ofnfection levels with 0. volvulus larvae in S. ochracy,s.l. populations, to explore the feasibility of estimatemean larval load from the proportion of flies (larvae, and to determine sample sizes (number of Bitto be collected) for varying infection and precision
levels under ivermectin control programs. These pn,cedures could be adapted to other Onc/iocerca-Sin;lium combinations and other filarial infections.
Materials and Methods
Study Site. The study was conducted in Las Goloi.drinas, a village of325 inhabitants located at 92 39’1)E
W, 15 25’ 59" N and at an elevation of 890 m infkState ofChiapas, southern Mexico, with an initial a,
chocerciasis prevalence of 78% and a communitya
crofilarial load of 12 mf/mg (Rodriguez-Perez eti
1995).Entomological and Parasitological Procedures,fc
control blackfly collections were made from Septenber 1990 through September 1991 at sampling site
within the village and a nearby coffee plantation. I
each sampling site 2 volunteers from the comniunih
working as bait and the other as collector, captureall S. ochraceum females that landed during the firstil
min of each hour from 0700 to 1730 hours. These flia
were dissected immediately, and all parous female
^A",,>-;N;^1"-<\|91’*,i’)l
K’>’\^t’’.....’""’"’,.t .^
V.r
f1’-’"lo.’1’,1’111.11^’iolc’,l.ln’’VKC1-1i.M-f!n>o"’l.ii-v.’lio"0
R.iUTl’1’II>OIIbit’srati-in 1
tionstratifniCaieac’con;lSS(
t\v<
;icr’
listfoilde--mitalCO
pl>re1te’teHsc

were examined for developmental stages (L[, L.^, andL, larvae) indistinguishable from 0. volvulus accord-ing to Porter and Collins (1984).An onchocerciasis control program based on mass
semiannual administration of ivermectin started in
November 1991 (Rodriguez-Perez et al. 1995). Post-control collections were carried out from December1991 to April 1992 and from January to March 1993,1994, and 1995 because the dry season has been shownto constitute the main transmission season in Guate-mala (Ochoa 1982, Porter and Collins 1988). Entomo-iogical measurements of transmission intensity forevaluations of the control program therefore were
concentrated during these months (Rodriguez-Perez^,1995).Variables of Interest. The variables of interest were
Pirity rate, proportion of infected flies, mean larval!ri, distribution of the number of larvae per fly, andlumber of flies to be collected and analyzed to detect^"’en infection rate with a fixed level of precision (a^lerable error about the estimate of the 0. volvulus’ana! population mean). Data from successive months*ere treated as independent observations with theOption of the postcontrol years 3 and 4 for which"onthly data were pooled to increase the number of^al categories. This was necessary for the applica-llon 01 goodness-of-fit tests.Comparison of Sampling Sites According to Parity
"ale, Infection Rate, and Larval Distribution per Fly.Mantel-Haenszel method jointly analyzed
.^wly pre- and postcontrol 2 by 2 contingency ta-’es ’or an association between parity rate or infection
^e "id sampling site (village and coffee plantation)’"1
"r^t" ^le ’^P’""^ months to obtain a single, un-
fm {a^e, might have provided incorrect resultsof the year were a confounder (Pagano and
^reau 1993) The odds ratios were calculated for
’’tseparate ^ble and a test of homogeneity was
>^o .ucte^ to ^^’""’"ne whether the strength of the
*’-Pn1 ’"r" ^etween parity and sampling site, or be-
lfry ""Sttion rate and sampling site, was uniform’i^, he Pre- and postcontrol months (the test sta-
’"’’lloit ""^geneity across a series of 2 by 2 tables
See5 ’’^’’"’"""ately a chi-square distribution with
linn01 freedom equal to the number of tables
<h|^ " ^e odds ratios were not constant across’’nt,^5111^ are reported for each month. If, on the
iti L6 ^^ were homogeneous among sam-
’’’^tivg i"’1’ a "ngle summary value for the overall,’"’<P(t ^
w^ computed and the null hypothesis
h’^1 an !10 ’"^’’^on between the variable of in-
^ns;g] ’’""pling site by means of the Mantel-’l’l^t(> ^..i^’^11’ (approximately distributed as a chi-
^ ^iT""0" of the distribution of 0. volvulus-like
’.^^hot"!8 parous flies between village and coffeellv iTip,.
pre- ^d postcontrol periods was carried"-. ^’e^el i^0 t^e Kolmogorov-Smirnov 2-sample
^’^bin and by testing whether or not the"’ial distribution was an adequate model
-& Golondrinas. The simple sum of the observa-
a(Ludwig and Reynolds 1988). The fit also providedestimates for the parameters ^. (mean) and k (degreeof aggregation). The outcome of these tests indicatedthat the variables of interest should refer only to par-ous flies and not to total flies in the remainder of thestudy, and that the data for village and coffee area
could be pooled and analyzed together (see Results).Indices of Aggregation. The variance over mean
ratio (VWR) of the larval burden per parous fly wascalculated for each separate sample. Departures oftheVMR from Poisson were tested by the d statistic, whichis appropriate when sample sizes are large (n > 30).Agreement with a Poisson series (VMR 1) would beaccepted with P > 0.05 ifthe absolute value of d were<1.96 (Elliot 1977, Ludwig and Reynolds 1988). In allcases the VMR was significantly >1.0, indicating a
strong degree of larval aggregation within the parousfly population. Consequently, the negative binomialdistribution was fitted to the monthly data using theprogram NEGBINOM.BAS (Ludwig and Reynolds1988), and its parameters estimated by maximum like-lihood (Grenfell et al. 1990). Goodness of fit betweenobserved and expected frequencies was tested by chi-square.The relationship between the parameters of the
negative binomial (i.e., between the degree of larvalaggregation, k, and the arithmetic mean larval load perparous fly, p.) was investigated. Only data sets thatpermitted testing for departures from the negativebinomial distribution were included (some samplesdid not have enough larval categories for the appli-cation ofthe chi-square test). Because a positive lineartrend was observed between the estimates of these 2parameters, a common k^. value could not be calcu-lated (Bliss and Owen 1958, Elliot 1977). Therefore,the relationship between k and m was explored bylinear regression methods, with least squares estima-
tion of the parameters weighted by the number ofparous flies in the sample. The most parsimoniousmodel was found to be A-(^) k^ (see Results).
Relationship Between the Average Larval Burdenper Parous Fly and the Fraction of Infected Flies.During entomological evaluations of ivermectin con-
trol programs, it may be more practical to assess theproportion of positive parous flies (e.g., with PCRmethods) than to calculate the mean number oflarvaeper fly. For an underlying negative binomial distribu-tion in which k depends linearly on /A as describedabove, the mean intensity of larval infection, m, may;be estimated from the observed proportion, p, of par-;ous flies with larvae as follows,
ln(l p)m
fci lnl +
Expressions for the variance, V(m); the bias, B(m),and the confidence limits (CL) of the mean are pre-sented in Appendix 1.Sample Size for a Specified Degree ofPrecision. The
ratio ofthe standard error ofthe mean [SE(fn) to thearithmetic mean, m, is an index of precision (D) (i.e.;

JOURNAL OF MEDICAL ENTOMOLOGY748 Vol. ^.
Table 1. PropMexico) during pr
Period
Pre-control
Tan. 1991
Post-control
Post-control 2
Post-control 3
Post-control
ortion ofand pc
Mo
Sept.Oct.Nov.Dec.
Feb.Mar.Apr.MayJuneAug.Sept.Dec.
Jan.Feb.Mar.Apr.Jan.Feb.Mar.Jan.Feb.Mar.Jan,Feb.Mar.
parons fliist-iverme
nth
1990199019901990
199119911991199119911991199119911992199219921992199319931993199419941994199,5
19951995
in biting 5.etin sampling
Vill.
Parityrate
0.2480.3190.4080.4080.3440.3990.4750.3510.3640.5360.3450.3580.4610.4010.4100.4790.4480.4210.4220.4240.5530.6630.6650.6640.6950.590
trhrdceum B.I.months
>ge
Total
6741,117579579
2,0251.9151,3615895388,38
1,7341,2251,4802,8723,4831,4081,0847,0453,7612,6806,1871,701600
3,7531,641787
collected in til
Cod
Parityrate
0.2840.4340.4070.4070.5400.5820.4990.4970.5850.5670.5280.3790.3690.6020.4890.5930.6050.4660.4530.4480.5630,615
0.6600.7970.7230.533
housing
Total
440715194194
1,0451,1911.069654176321654507515951
1,652513405
2.8222,3271,5424,9271,213968
1,6801,1572,414
coffee of
OB
1.205NS1.636*0.998NS0.998NS2.234*2.097*1.097NS1.823*2.462*1.134NS2.116*1.091NS0.684*2.257*1.376*1.584*1.884-1.197*1.135*1.099NS1.039NS0.814*0.978NS1.992*1.141NS0.795*
Las Golonnrio,
95%Q
Lower
0.9191.3470.71-0.7171.9181.8100.9,35
1.4511.7390.8751.7620.8810.5571.9431.2231.2911.4931.0961.0230.9690.9630.6980.7891.7370.9670.675
"C
^^^,
ll1<U(tsI*4!.!,liii,
I’i.I;1:14l*itdi,1<l4Ot.
The estimated odds ratios (OR) the strength of the association between parity and coffee plantation. OR > 1, the proponeparous flies is higher in the coffee plantation; OR < 1, the (higher parity is associated with the housing area). *, Signij^different from unity (the 95% CL does not include 1); NS, not significant. The odds ratios sampling months not constant^summary overall value not calculated (for the precontrol months the chi-square test of homogeneity among odds ratios y2 y,with 11 df freedom, P < 0.001; for the postcontrol months this figure ^ 246.1 with 13 df, P < 0.001; text).
a measure of how large a fractional error will occur in
the estimate ofthe population average). Because SE(m)is simply VV(m), it is possible to relate the equations forD, m, and V(m) to obtain an expression for Up, thenumber of parous flies that must be analyzed given a
prevalence of infection in the vector population anda specified degree of precision,
[2]ID D^l Trn-lnO Tr)]
Equation 2 permits calculation, under the assumption
of a negative binomial larval distribution with lineardependency between k and the mean, of the numberofparous flies, n, to be examined for 0. volvultis larvaefor any given true proportion ofinfected flies, TT, whenthe mean larval load is to be estimated with an ac-
cepted level of error, D. Only under the particularassumption k(p-) fci(ui, does n^ not depend explicitlyon the aggregation parameter, but on the (unknown)fraction of infected flies in the parous population.Details of the derivation of equations and 2 are
outlined in Appendix LThis rationale assumes that fc,is a known constant (i.e., derived with relatively highprecision from prior data). Appreciable errors in es-
timating fc; would add to the imprecision of m.
Results
Comparison of Sampling Sites According to PanRate, Infection Rate, and Larval Distribution perp,Parity Rate. For both pre- and postcontrol periods,!!,monthly parity rate appeared often to be higher in tcoffee plantation than in the village. The strength;the association between parity and coffee samplesite in Las Colondrinas was measured for each sss.pling month by the corresponding odds ratio. Becanodds ratios were not uniform across months, a sui
mary value could not be calculated. Table reportsll,fraction of parous flies and the odds ratios (witht95% CL) for an association between coffee plantafio-and parity for each one of the pre- and postcontnsampling months. These results indicated that eiiti
mological indices of transmission should be consic
ered in relation only to the parous fly population, iftkdata from both sampling sites were to be analyze;together.
Infection Rate. The odds ratios for the associate
between the proportion of infected parous flies an
sample site in Las Golondrinas were homogene<nacross pre- and postcontrol sampling months andovnall relative odds values therefore were obtained, tha^95% CL calculated, and the null hypothesis ofiassociation tested with the Mantel-Haenszel sta

3 ^|5 r^
1 T
’ rj60 Trecootrol 0.014 0020 2014 4.110
.,- 199M1) W93) (3,530) f00 0.6B1 1.862 0.319
,90 Ptcontro 1-4 0.003 ^0003 1.135-N.’
:^ ,, N9.158) (12..9.) ----------------------------,02
129;S5
291185t-0540352^41.621348.9443"’30".26024’120
.94S1.2142.2841.34S0.936
i0" ofc;"’111.>..’’10’
p^-.-I";Js.t11.int1".,?t" ^^i""^<.^;-
^^c0^>t>’"^"l^1^:%:
^
r33-c.~^ ^iRmBCTiggS-------------BASANEzrn^SAHFLE^TZESWDrnN^.W^aK-Wt".
... Tl,ep.opo^of^^p^H,e.,n^^^^^-.>..-"ee.ed,n.HeHo^^.o.ee.^.or^OoW.^W^, Mexico) during pre- and po.t-ivermeclin .ampling month,
95% CL (OR)Intection rate Si.mimn’ V2.,!,
Period Village Coffee OR Lower L’Pper
(Parnusn) (Parousn)
^e---v odds ratios (OR)~ the stre^^e^^^^^^^^^S^d-:^rS:^^^association is significant, the 95% CL do not include 1. The odds ratios acrosss^1"1^0 ^ n y^ 11 df, P > 0.10; for the
Plated (for the precontrol .nonthsthe ch-square test ^^^"^^.H^^i^rstati^ follows approxin,ately chi-square
S.^df^e^^e^^T^^^^^^^^^TOulation and sampling site for the post-ivennectin months (see text).
,v’ ^/^f-l^^ rplaHnnsiiin between the niaxiinum likelihood esti-
^^^ Thistest.stat;st’cl ^’^a( OOS) mite of fcTrom the negative binomial and the arith-slight y above the critical value (3.841, a U.U>; maie ui
oarous fly for theforthe precontrol period, but it was well below this metc,mean,numb^lo^^^t^pe^ testing ofyr the postcontrol months. Consequently, the mon^^^^^ wert incWed.KSL-^^^^^^^^^ .2-sample Kolmo.- ^ re^ssion line^^^^^^-S.imov test revealed no differences between ^q^^^^ sapling sites regarding larvae distribuhon ^^^^^^^^yee plantation i’n the remaining analvse^ t^^^^^d ^dL these data""lices of Aggreeation and Relationship Between line is not a regression
oredicted’^^tion oflnfeTted Flies and the Average Larval points, but instead them^^^^^^^^ Fly. Table 4 summarizes the monthly data using equation and the fc, value calculated trom g
Sy6’and P^^0"11-01 sampling periods, the 1.^5 tested using an independent
^^S^^^S^pS: da^^mSe^adheor^ldatafromCupp
^- Th. di^WUon of .11^^ of 0. ^H, ..on, p..ou. S.^c^. nie. .o,le.,ed in .he hou.n, and coiTee
’>^ Golondrinas (Chiapa,, Mexico) durins pr<- and post-ivenneelm -amphng months
Fostcontroll.in,,p Prprontrol -------------------------~~------------------------ff------ Village (Jottee
--^ Ohse.JillaeeExpected OWr^^^-----4.21------^21------3:^59------3,459 19,10^ 19,105 1^3 ^
32 33 36 6 ^ ^ , 217 " 1, 6 4 5 56
5 ^ 3 2 ^. 2 1 9 3 0 ^ ^^^ \^ 9 3,530 19,15^ ^-> ^ w^ ^ ^s’.,, 0.230 0.191 0^4 ^’^"- 5.78 ^Oe
,,.40 203.58%( 140.21 f.33 l35A9
0.00190.0082 0.013.5 ^3.38 3.5’
4 4
V^-- ’030 >(}AO >0’05 >al0-----
"^^’^^Pond’to negative binomial fit with Jean^dTesH.ates shown. Agreement between observed and expected
""d by the chi-square test.

750 JOURNAL OF MEDICAL ENTOMOLOGY VolM
Table 4. Pro- and post-in din levels of 0. volvulus larval infection and aggregation in S. ochraceum s.l. parous flipand coffee plantation in Las Golondrinas, Chiapas, Mexico ^^
Period Month N(7a^ous Infected flies Larvaey^R , ^^Precontrol
Sept. 1991Postcontrol
Postcontrol 2
Postcontrol 3+
Sept. 1990Oct.Nov.Dec,
Jan.Feb.Mar.Apr.MayJuneAug.
Dec.Jan.Feb.Mar.Apr.Jan.Feb.Mar.Jan.
1990199019901991199119911991199119911991
199119921992199219921993199319931994-Mar. 1995
292666315315
1,2611,4571,180532299631944631872
1,7232,234978731
4,2822,6431,827
16,665
56841925337y8156857591299
21
P
0.0170.0090.0250.0130.0150.0180.0280.0130.0230.0130,016
0.0100.0090.0030.0030.0050.0120.0030.0030.0050.001
111020265566731318213319121018201129141423
0.0380.0150.0640.0830.0440.0450.0620.0240.0600.0330.0350.0300.0140.0060.0080.0200.0150.0070.0050.0080.001
3.16*1.79*4.35*
13.04*6.93*4.41*4.20*2.83*3.28*5.83*3.27*5.93*1.99*2.40*3.99*
11.59*1.53*3.34*1.85*1.85*1.26*
18.7512.3127.2365.4481.9259.4050.9922.2219.8550.2535.1350.9317.1332.1566.73
106.319.10
76.5526.2521.7822.33
k
0.01^"0.00970.01610.00420.00830.01030.02020.01180.01450.00730.01150.00480.01220.00230.00190.00220.03270.00180.00410.00600.0071
^",’t’it,iinttil
>tiHiHiI*t
^’d,tIt:Oji
Natural infection levels expressed the proportion of infected parous flies (with all larval stages) and the oflarvae perfly. The degree of larval aggregation is measured by the VMS and the estimate of the negative binomial parameter k. *, Significantlythan unity (rf statistic > 1.96). srtl
et al. [1992]), consisting of pre- and postcontrolmonthly samples (1988-1990) from which the pro-portion of parous S. ochraceum infected with 0. vol-vulus and the mean larval burden per fly could becalculated. The underlying larval distribution per par-ous fly was assumed to follow that observed in theMexican data (i.e., fe; was a known parameter). Theobserved and predicted mean larval burdens agreedwell in relation to the line of unity slope (Fig. 3).
Sample Size for a Specified Degree of Precision’calculate sample sizes within a realistic range of,.ural infection levels for S. ochraceum s.l., publishedaunpublished information dating from 1974 wasmarized in Table 5. The pre-ivermectin infectionwith all larval stages ranged from 0.7 to 17.0%-’;li,introduction ofthis control measure lowered inferti,levels below 0.5% in those villages where it has be..possible to conduct repetitive interventions within,coverage and compliance [Los Andes in Guatenii
Fig. 1. Maximum likelihood estimate of the degree of0volvulus larva! aggregation (parameter k of the negative bi-nomial) plotted against mean larval load per fly, m, for eachseparate precontrol (open squares) and postcontrol (closedsquares) parous S. ochraceum s.l. monthly sample from LasGolondrinas, Chiapas, Mexico. Control refers to thesemiannual administration ofivermectin. Only data sets thatpermit testing for departure from negative binomial are in-
cluded. The regression line (fc(m) k^m} is weighted by thenumber of parous flies in the sample (r 0.87, fej 0.273).Error bars are SE (k) as in Table 4.
f
Proportion Larvu,
Fig. 2. Mean larval burden per parous fly plotted againthe proportion ofparous S. ochraceum flies naturally infectewith 0 volvulus in precontrol (open squares) and post(trol (closed squares) samples from Las Golondrinas. Thelitis not a regression line but the predicted infection intenAin the flies according to equation with parameter k as inr(1. Bars the 95% CL about the predicted mean witli(number of flies) as in the corresponding sample (seeffpendix 1). Al! but of the observed means lie within tha^confidence intervals.
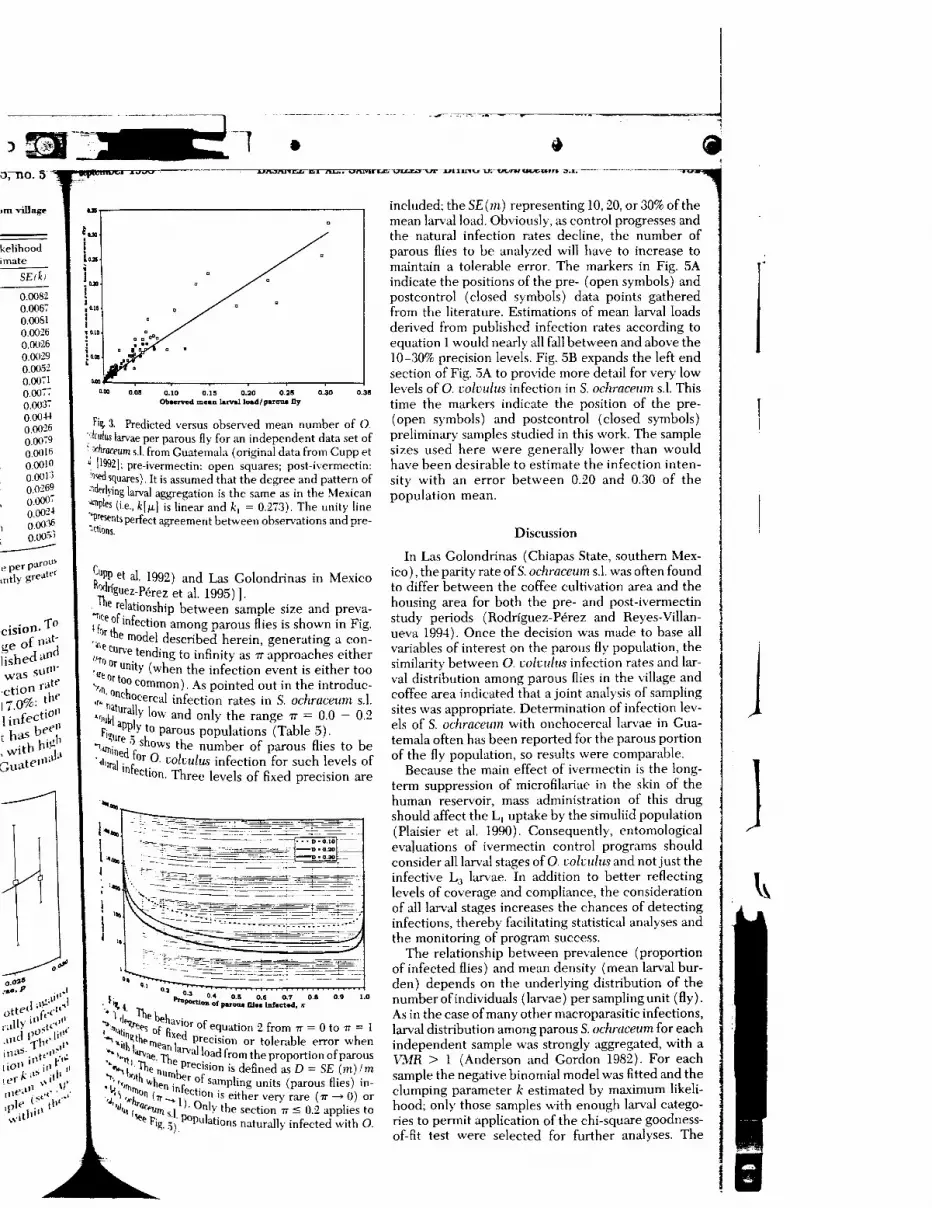
0.00820.006"0.00810.00260.00260.00290.00520.00"!0.00"0.003"0.00-H0.00260.00:90.00160.00100001"’0.02690.000"0.002-10.00’16O.OO^’’
eperp."-0’"Jlygr^
^
y.P’5’"!.u’"1^,’-^int1’ r"11-
,il’
ti?. 3. Predicted versus observed mean number of 0,kiilus larvae per parous fly for independent data set ofvhraceum s.l. from Guatemala (original data from Cupp et11992]; pre-ivermectin: open squares; post-ivermectin:
’"ri squares). It is assumed that the degree and pattern ofwriying larval aggregation is the same in the Mexican^P’K (i.e., k\p.\ is linear and fc, 0.273). The unity lineT’ttents perfect agreement between observations and pre-vious.
"Pp et al. 1992) and Las Golondrinas in Mexico"Wguez-Perez et al. 1995)].
relationship between sample size and preva-’"rection among parous flies is shown in Fig.
~t’ "’"del described herein, generating a con-
,^curvre Ending to infinity as IT approaches either
wor unity ^"en the infection event is either too
\n ^""^’"""on). As pointed out in the introduc-
nat"0 ocerca^ infection rates in S. ochraceum s.l.
^"iMlow a"d ""’y the ""s(R) w o- -2
Fi^^-y to P^ous populations (Table 5)."i^, e, ^hows the number of parous flies to be^ra] .^’ ^wlus infection for such levels of
sction. Three levels of fixed precision are
’*.,
^^Y^’’’01" of equation 2 from 0 to
^xiir^ertieanT P’’ecision or tolerable error when"’’..( "’^ae -m ’lrva^oad from the proportion ofparous
’^’ The numh1"’*’0""0" is ^""ed as D S (m)/m^"^l^M ^’""^""g ""its (parous flies) in-
"’ ,,i10" (TT^ ,\ " is eit"er ve^>’ (-n- 0) or
’’’’11 (^m ""’y the section a 0.2 applies to
’’f^ 5) ""Potions naturally infected with 0.
-i
..t’-’
included; the SE(m) representing 10, 20, or 30% ofthemean larval load. Obviously, as control progresses andthe natural infection rates decline, the number ofparous flies to be analyzed will have to increase tomaintain a tolerable error. The markers in Fig. 5Aindicate the positions of the pre- (open symbols) andpostcontrol (closed symbols) data points gatheredfrom the literature. Estimations of mean larval loadsderived from published infection rates according toequation would nearly all fall between and above the10-30% precision levels. Fig. 5B expands the left endsection of Fig. 5A to provide more detail for very lowlevels of 0. volvulus infection in S. ochraceum s.l. Thistime the markers indicate the position of the pre-(open symbols) and postcontrol (closed symbols)preliminary samples studied in this work. The samplesizes used here were generally lower than wouldhave been desirable to estimate the infection inten-
sity with an error between 0.20 and 0.30 of thepopulation mean.
Discussion
In Las Golondrinas (Chiapas State, southern Mex-ico), the parity rate ofS. ochraceum s.l. was often foundto differ between the coffee cultivation area and thehousing area for both the pre- and post-ivermectinstudy periods (Rodriguez-Perez and Reyes-Villan-ueva 1994). Once the decision was made to base allvariables of interest on the parous fly population, thesimilarity between 0. volvulus infection rates and lar-val distribution among parous flies in the village andcoffee area indicated that a joint analysis of samplingsites was appropriate. Determination of infection lev-els of S. ochraceum with onchocercal larvae in Gua-temala often has been reported for the parous portionof the fly population, so results were comparable.Because the main effect of ivermectin is the long-
term suppression of microfilariae in the skin of thehuman reservoir, mass administration of this drugshould affect the L[ uptake by the simuliid population(Plaisier et al. 1990). Consequently, entomologicalevaluations of ivermectin control programs shouldconsider all larval stages of0 volvulus and not just theinfective La larvae. In addition to better reflectinglevels of coverage and compliance, the considerationof all larval stages increases the chances of detectinginfections, thereby facilitating statistical analyses andthe monitoring of program success.The relationship between prevalence (proportion
of infected flies) and mean density (mean larval bur-den) depends on the underlying distribution of thenumber ofindividuals (larvae) per sampling unit (fly).As in the case ofmany other macroparasitic infections,larval distribution among parous S. ochraceum for eachindependent sample was strongly aggregated, with a
YMR > (Anderson and Gordon 1982). For eachsample the negative binomial model was fitted and theclumping parameter k estimated by maximum likeli-hood; only those samples with enough larval catego-ries to permit application of the chi-square goodness-of-fit test were selected for further analyses. The

752
THI.I<- ^GiiHl.-mnl" <<
Locality
Sta. Anita
SibajaSta. EmiliaLos TarralesProvidenciaSibajaRosarioMonica IvonePena BlancaE1 JardinFanimaquibLos TarralesSta. IsabelMon’IosLos AndesSta. EmiliaTarrales TZ"Colondrina.s
Los AndesLos AndesSta. EmiliaSta. EmiliaTarrales TZ"Tan-ales TZ"ColondrinasOoloiidriiiasGoloiidnnasGolondrinasMorelosMorelMorelosN\ ReformAsraria
>siirvry
M-fr- (Pr<--trl)
country
GCGGCGGGGGGCGMGCGM
GGGGGGMMMMMMMM
f //.
an<l
JOURNAL OF
rfilrnliis natural iiafl<-r (Post-Ctrl^
Year
197-1197419-41976-19-1976-19-1976-197-1976-19-197--19-81977-19781979-19801979-19801979-19801979-19801979-19811988198819881990-1991
19S919901989199019S919901991-19921993199419951993199419951995
MEDICAL
ifpction in par’Tniannual ad
Iverrnestatus
Pre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrlPre-ctrl
Post-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-CtrlPost-Ctrl
ENTO
ctin
(yr)
(1)(2)(1)(2)(1)(2)(1)(2)(3)(4)(3)(4)(5)(5)
MOLOCY
biting S. orhriniiiinislration of ivprr
Proportion
Parous
0.3860.3070.4690.3610.4730.5720.7860.2470.4460.4620.4270.4230.5070.3460.3960.4500,40-
0.419
0.3960.3960.4.500,450
0.4070.4070.4550.4340.5840.6610,340
0.5900.5200.530
S.I. po|nilnt
iicctin
of flies
Infected
0.03110.10490.166-0.04790.01770.125S0.17360.00690.04450.00900.04020.06000.08570.02780.04600,0500
0.09100.0168
0.00400.00500.01100.01300.03300.01900.00520.00340.00080.00190.00030.00250.00280.0028
^ 1\(""’--M^,,
’k
^^Carnisl9^^\Gamis 197.5Garnis 19-3Collins 1979Garnis and (uGarnis and Oc.i^l’Gai-msandak’lfOchoa 1982 ^1.Ochoa 19S2Porter et al. Kn,Porter e.al,i^Porter etal,^Poner et al. 19"OTte^ et al. ]^Cupp et a|. 19^’Cupp et al. 199.^Cupp et al. 199^Rodnguez-Per,,,
1995 and hisCupp et al. [y^^Cupp et al. 199;Cupp et al. 1992Cupp et al. 1993Cupp et al. 1993Cupp et al. 1992Rodriguez-Pere,
This workUnpublished dadUnpublished dat,Unpublished dataUnpublished data
"Tarrales TZ: Tarrales transmission zone; includes the villages of Santa Isabel, El Vesubio and Los Tarrales (see Cupp et al. 199.2)
negative binomial adequately has described the dis-tribution oflarvae in other Onchocerca-Simiiliiiin com-binations (Cheke et al. 1982, Garms and Cheke 1985Renz 1987).
In the case ofO. wkiiliis larvae in populations ofS.ochraceum s.l. from southern Mexico, a plot of themaximum likelihood estimate ofk against mean num-ber of larvae per parous fly showed that the degree oflarval aggregation (inversely measured by the value ofk) decreased as larval burden increased. As a conse-quence, a common ^, value could not be calculated. Asignificant decrease in parasite aggregation with in-creasing infection intensity has been observed in otherhost-parasite systems (Lwambo et al. 1992, Medley etal. 1993). Because in the study presented here therelationship between k and the mean was describedadequately by a model of the form k{p.) k^p., anexplicit expression to estimate the mean larval densityfrom the observed proportion of positive flies wasderived in equation 1.The relationship between prevalence and intensity
is usually strongly nonlinear, reflecting both the de-gree of parasite overdispersion and the fact that theproportion of infected hosts cannot be >1 (Andersonand May 1985). Because the fraction of naturally in-fected vectors and the mean larval loads are generally
low, a plot of the field values for the mean lan.burden against infection rate fell within the iniulinear portion of the relationship (Dye 1994). Adoaing the A-, value estimated from the data shown onFiand using equation 1, a reasonable prediction,,
larval densities was generated not only for the Mercan data, from which this aggregation parameter^estimated, but also for the Guatemalan data, for whythis degree and pattern of aggregation were assureto hold. We used presence-absence data to estinufemean larval loads from onchocerciasis infections!simuliid vectors. Our approach could be adaptedother Oncliocercn-SimiiInim combinations, other filia-vector systems (Das et al. 1995, Nicolas et a!. 1996and other vector-borne infections such as malaii(Medley et al. 1993, Billingsley et al. 1994).
VVilson and Gerrard (1971) had adopted presence.absence criteria for rapidly estimating populationdsities ofdiprionid flies infecting plantations ofyouapines in North America. They found that k valueremained constant among sample sets making it vali:to obtain a common k^. estimate for the whole seriaOther attempts to furnish density estimates from prev-alence data in the Helds of agricultural and medid.entomology have concluded that the negative bino^mial was not a satisfactory distribution model

nu.-r, jy^
IM)aI"i 1
19-791979197S
SSS8S8S8192
1,’z et al.
worl-.1-2)"7
l2929292
latal.it.ilatalata----32).
- ’^i1’initi^^cloP’-
fill-nonf"-,-.ctio.1le Nl^
^^sl"11;esti" l,,,ctio^ .[f^-\\^’" l.ii’1’1n">1-
,r^-’, ::;.tio’">r, f>11^l-11^ ^1"
I1lu .-I-*1
;;^"", i,.-’
I1
’^,1 ’’t’1 ’^val^1"’6"1 a" algorithm by which estiinates of^ of infection and confidence limits for
OTDengoc. BASANEZ OT AL.TSA
|--]-B
i^nW^^ CO
----~-------------Proportion ofpirom Infected
mm,-----------------------------~~~IBJ
-:~.
’^. ----.-.V^-------nl--’ n
A. ^--"Q --3--n-^--
’10
Proportion ofparoue Hiee infected
^ 5. The number of pai-ous S. oclirciceuin flies to be’"^ed for 0. whiilii.’i infections for tolerable levels of^"sion ranging from D 0,10 to 0.30 and realistic infection
^ Open squares (before ivermectin); closed squares (un-’’’I’ermectin control programs). 5A: From literature surveyw"f^. 5B; From this work (Table 4).
.^onn egg populations, Gerrard and Chiang 1970),
^ common ^. value was not appropriate (Aedrs.^Pwovitrap, Mogi et al. 1990]), orthat the degreelegation changed with the age of the organism
-^ density was estimated (culicid larvae in rice
^.’’^airn et al. [19141). A more empirical ap-^to the problem of obtaining density estimates
\,^ ^quency of occurrence has been proposed by
If-o"^ ^’"o (1958) and Cerrard and Chiang
’"TM ^Kh is not dependent on well-known ag-
^d ^quency distributions. Their procedure’Iv i^115^’1 ’n Kuno 1986) was not adopted in our
"’tir.r ^we the efficiency of estimation falls off
n’< ."^Ptiy when the proportion ofoccupied sam-
"’\ fl"’15 tloes "ot lie between 0.5 and 0.8 (Gerrard.""’"’g^TO).
’"’’^ i’""116 Sftomological evaluations ofivermec-
>^ to0 p’’ogra"ns "sing 0. rodiiiu.s-specific DNA.’.’lio^j est Pools of flies, presence-absence of in-
’’""w 1- vectors may be used to derive estimates
.’"’’tiili ^’" ^ad (a variable not furnished by these
^’’^av ques)- Tms Presupposes that PCR-
^ofK;1’ are eapable of providing accurate esti-
^"^’ion6 proPortion of infected flies in the vector
"("’^PonS rlot Just minimum infection rates (a’L "^1 n1ay ^ntain >1 infected fly). Katholi et
MFLES
P0
c
e
PP
c
r
-75T’
population samples can be obtained from the number
of negative pools in a total of pools being tested eachcontaining a given number of flies. However, it is
essential that the assay is sensitive enough to detect a
single infected insect in a pooled sample and that the
pool size is chosen carefully to produce only a few
positive pools. Oskam et al. (1996) have developed an
assay that permits detection of 0. fo/L-;!(S-infected
S. ochraceum in a pool of 79 uninfected flies.
Polvmerase chain reaction-based approaches are
meant to obviate the necessity to dissect flies at all.
However, because ofthe reasons discussed above, our
results are based on the parous fly population, imply-
ing that prior assessment of reproductive status is
required. The method we propose would still be ad-
vantageous if parity can be ascertained relatively
quicklv by examining the ovaries of the flies (abdo-mens only) during the remaining half-hr of each
hourly collection (N. J. Villamizar. personal commu-nication ), and specific identification of filarial larvae is
important. Parous flies can be preserved in 100% eth-
anol for later DNA studies in the laboratory (Post et
al. 1993). Assays performed on flies classified as nul-
lipars could be used to help refine the criteria applied
for age-grading in the field.The charts in Fig. 5 could be used as rough guides
for sample size estimation as control with ivermectin
progresses and the proportion of infected S. ochraceum
decreases. While under the assumption of a common
k the sample size increases with the degree of overd-
ispersion. the sample size derived here is not explicitly
dependent on the value of fc|. This only applies to the
particular case explored in this study, where the re-
lationship between k and the mean is linear and goes
through the origin. Three levels of precision are de-
picted, D 10%, 20% or 30% SE percentage values of
the mean larval density. Even accepting a 30% error
level, the number ofparous flies to be examined for 0.
wkulus infections may become very large and deter-
mining infection rates for pools of parous flies rather
than individual flies clearly would be an advantage.
Obviously, the amount of sampling days necessary to
collect such amount of parous flies will depend on the
daily biting and parity rates at the particular locality.
The host-seeking density of S, ochraccum s.l. may be
verv high; in fact. meso- to hyperendemic onchocer-
ciasis transmission by this species can result only from
high biting rates (Wada 1982, Basanez 1996). These
would be the areas where control with ivermectin is
a priority. In Las Golondrinas, where after 9 biannual
treatments with extensive coverage and compliance,
infection levels have dropped to <0.2%, a minimum
number between 6,000 and 13,000 parous flies would
have to be collected for a precision D between 0.20
and 0.30. With a mean daily biting rate (housing and
coffee areas analyzed together) of^76 parous flies per
person per dav (collection periods equal to 30 min/h,
11 h/d), this represents between 80 and 170 collecting
davs to be distributed among the highest transmission
months (January to April). The sampling regime de^scribed here would require 2 baits, in the village and
the other in the coffee plantation to work for 10-20

754 JOURNAL OF MEDICAL ENTOMOLOGYVol.^d/ ino to collect the required number ofparous flies.
Alternatively, 4 baits could work for 5-10 d/moduring the 4-mo dry season. The practicalities of thesampling regime for each particular locality willhave to depend on the corresponding host-seekingparous density of S. ochraceum and on logistic andeconomic factors.However, because the true proportion of infected
flies often will be an unknown quantity, equation 2 canbe used, alternatively, to calculate a target number ofinfected Hies to be collected for a desired level ofprecision rather than a total number of parous flies.Equation 2 does in fact approximate the expression: n^l/Zy’TT. If TT (unknown) is written as n,/n^, wheren, is the number of infected parous flies in the totalparous sample, n^, for a level D of precision, it followsthati, D2. ForD 0.20 or 0.30, this translates intoa target of 2.5 or 11 infected flies respectively. There-fore, possible sampling scheme would be to continuesampling until the number of infected flies detectedreaches approximately such values. With the applica-tion ofDNA probes to pools of parous flies, it is con-ceivable that each monthly collection in the Heldcould be followed by a period of fly analysis in thelaboratory, the results of which would determine themagnitude of the sampling effort required for thefollowing month. Field collections would then stoponce the target number of infected flies has beenachieved.
In the current article, the referred levels of preci-sion relate to the estimation ofthe mean larval burdenfrom the proportion ofparous flies infected. However,regarding the field application of the methods herediscussed, or of any other sampling schemes, accept-able levels of precision would have to depend onwhether the control program in question is aiming ateradication of infection or elimination of morbidity.For instance, other approaches to the problem ofsam-ple sizes in onchocerciasis entomological evaluationswould include the estimation ofthe number of flies tobe processed to detect, with a desired level of signif-icance, a given drop in infection rates (rather than ininfection intensity). Given the linearity between in-fection rate and infection intensity described hereinfor S. ochraceum, relationships between intensity ofinfection in the flies and various epidemiological orclinical indices would be expected to hold also forthe fraction of infected flies (Dye 1994). Prelimi-nary analyses (data not shown) suggest that it maybe possible to infer the proportion of infective fliesfrom the proportion infected in S. ochraceum sam-ples. This would permit estimation of annual infec-tive biting rates. National onchocerciasis controlprograms might benefit from exploring the relation-ship between annual infective biting rates and on-chocerciasis epidemiological patterns as they havefrom establishing that between annual transmissionpotentials and onchocercal infection and disease inthe human host (Duke et al. 1972, 1975; Thylefors etal. 1978; Renz et al. 1987).
f
AcknowledgmentsE. W. Cupp kindly provided the original Gual
set from his TDR-funded study. At Oxford Ch ^iand David R. Cox provided general statistical ^ii’1latter in particular commented on possihl \’schemes. Man-Suen Chan and John Williams (W^r^’lthanked for helpful discussions and Brvan Gre f^bridge) for the use of his Pascal program for th" ^’likelihood estimation of k. We also acknowled^t^our Beld workers in Chiapas (A. Aguin-e. A. Avi- ^V. Donringuez, D. Gomez, S. Marti’nez, R. Rod^-lkSandoval, N. Valdivieso, R. Vasquez, andE. Villafu’18’1^out whom this work would not have been possibi erIe^’supported by the Wellcome Trust inter-centgrant for the development of onchocerciasis matF11^models. M.A.R.P. is the recipient of a doctoral scho]^the University of Salford awarded by Consejo Na ^SPloy./.l’n "^lOrhtCienciayTecnoIogfa (CONACyT) of Mexico (gran1?’1’Work in Mexico took place at El Colegio de la Frn 2S’(ECOSUR, San Cristobal de Las Casas) and Cent Hvestigacion del Paludismo (CIP, Tapachula) withe0’1’*support from CONACyT (grants 3187-M and 3320pnlar"’
Appendix
Derivation of the Expression for the RelatioBetween Mean Larval Load per Parous Fly an3Proportion ofInfected Parous Flies. For an under) ^negative binomial distribution in which k dependP-, the prevalence, TT, is a function of the intenseinfection 4’
l-t(fl)p-TT= + k(^))
Because it was assumed that k IJL was a linear In,,tion of the form k(p.) fcp + k^ but the estiina"intercept k^ was not statistically different from ^the expression k(ii) k^ was substituted in ^previous equation to give
-tin7T= 1 +
This equation can be rearranged in order to obtaiithe estimated mean intensity of larval infection, mfrom the observed proportion, p, of parous flies willlarvae
ln(l p)
^[^ k-JDerivation of the Expression for the Sample Sia
Given a Specified Degree of Precision. D is given hithe general formula (Elliot 1977, Mogi et al. 1990)
mm)D
According to Kuno (1986), the derivation oftheexpression for the variance and the bias of the mean(respectively V(m) and B(m)), depends on whetherthe mean population density will be estimated froffl.direct larval counts or from presence-absence data.Because this article focuses on the latter, V(m)

---DASANEZ-I-Spnibenyya aAMFQTOCCEa’CTr-i.inirw
BIm) need to be derived from first principles forprevalence data. Following Kuno (in the work cited)
V(m) [{/(p) -/(ir)}2] {f’WYV(p)
B(m) [{/(p) -/(7r)}] ^f"(TT)V(p)*here/(p) is the function relating mean larval load, m,;o the proportion of infected flies, p (equation 1); wishe true prevalence of infection among wild flies;’’=(ir) and/’(Tr) are, respectively, the 1st and the 2ndPrivative with respect to TT, and V(p) is the error
iriance ofp, which is (Armitage and Berry 1994)
TT(1V(p)
For k depending linearly on p. as described above,’e variance and the bias of the mean are
V(m)
n(l ir) fciln + r-
B(m)
^1 ^(1 .)J^"^CL about the mean can be calculated as m +/-
H’"’1’ ^e ""biased estimate of the mean
;,’ dbe obtained by subtracting the result of B(m)
^’ trlat of equation 1, although this is a very minor,
"J ""necessary, refinement. From the expres--,- ^""’"g precision, it is obvious that V(m) D231, ^ore, rearranging the equation for V(m)
’<e to btain n for a fixed precision D, the
t sampling units, n^, becomes
[2]D\\ TT)[ ln(l TT)]2’
References Cited
’l"stn ^yi- Determinants of infection in human
^lli^0""35"’ PP- 279-300. In A. F. Mahmoud [ed.],^’""ase c ca^ tropical medicine and communicable%, p
01 2, 2nd ed. Bailliere Tindall, London.
icin^ (, -I. "’d D. M. Gordon. 1982. Processes influ-
"Mtti ^"tion of parasite numbers within host(j’10" tiort’1? wlt^ ^"al emphasis on parasite-induced%>,i "’"" Parasitology 85: 373-398.
’^"lan"lndR-M.May. 1985. Helminth infections
’r^ ^onhr ."^^"’atical models, population dynamics,"S, p Adv- Parasitol. 24: 1-101.
^al rpc- Kerrv. 1994. Statistical methods in^ M. c^*’ 3rd e(l- BIackwell, Oxford, England.
’"’""ssion r?’" *:>ensity-dependent processes in the
^t to0 ""man onchocerciasis with particular ref-
^I’^^ion Te ^^’’^"nulium interaction. Ph.D.^icirie ^’^Perial College ofScience, Technologyand^.G ^^Yo! London.Ifc ’""lar’r """Kinesq, J. Prod’hon, H. Frontado, N. J.i,i^"y-dep^ Medley, and R. M. Anderson. 1994.
""^Oce, .t Presses in the transmission ofhu-clas1s: intensity of microfilariae in the skin
and their uptake by the simuliid host. Parasitology 108:115-127.
Basanez, M. G., J.H.F. Remme, E. S. Alley, 0. Bain, A. J.Shelley, G. F. Medley, and R. M. Anderson. 1995. Den-sity-dependent processes in the transmission of humanonchocerciasis: relationship between the numbers of mi-crofilariae ingested and successful larval development in
the simuliid vector. Parasitology 110: 409-427.Basanez, M. G.,H. Townson, J. R. Williams, H. Frontado, N. J.
Villamizar, and R. M. Anderson. 1996. Density-depen-dent processes in the transmission of human onchocer-ciasis; relationship between microfilarial intake and mor-
tality of the simuliid vector. Parasitology 113: 331-355.Billingsley, P. F., G. F. Medley, D. Charlwood, and R. E.
Sinden. 1994. Relationship between prevalence and in-
tensity of Plosmodium falciparum infection in naturalpopulations of Anopheles mosquitoes. Am. J. Trop. Med.Hyg. 51: 260-270.
Bliss, C. I., and A.R.G. Owen. 1958. Negative binomial dis-tributions with common k. Biometrika 45: 37-58.
Cheke,R.A.,B.Garms,andM.Kerner. 1982. The fecundityof Simulium damnosum S.I. in northern Togo and infec-tions with Onchocerca spp. Ann. Trop. Med. Parasitol. 76:
561-568.Collins, R. C. 1979. Onchocerciasis transmission potentials
of four species of Guatemalan Simuliidae. Am. J. Trop.Med. Hyg. 28: 72-75.
Collins, R. C.,J. 0. Ochoa, E. W. Cupp, C. Gonzalez-Peralta,and C. H. Porter. 1992. Microepidemiology of on-
chocerciasis in Guatemala: dispersal and survival of Sim-tilium ochraceum. Am. J. Trop. Med. Hyg. 47: 147-155.
Cupp, E. W. 1992. Treatment of onchocerciasis with iver-
mectin in Central America. Parasitol. Today 8: 212-214.
Cupp, E. W., J. 0. Ochoa, R. C. Collins, F. R. Ramberg, andG. Zea-Flores. 1989. The effect of multiple ivermectin
treatments on infection of Simulium ochraceum with On-chocerca volvulus. Am. J. Trop. Med. Hyg. 40: 501-506.
Cupp, E. W., J. 0. Ochoa, R. C. Collins, M. S. Cupp, C.Gonzalez-Peralta, J. Castro, and G. Zea-Flores. 1992.
The effects of repetitive community-wide ivermectin
treatment on transmission of Onchocerca volvulus in Gua-temala. Am. J. Trop. Med. Hyg. 47: 170-180.
Das, P. K., S. Subramanian, A. Manoharan, K. D. Ramaiah, P.Vanamail, B. T. Grenfell, D.A.P. Bundy, and E. Michael.1995. Frequency distribution of Wuchereria bancrofti in-
fection in the vector host in relation to the human host:evidence for density dependence. Acta Trop. 60:159-165.
Duke,B.O.L. 1968. Studies on factors influencing the trans-
mission ofonchocerciasis. IV. The biting-cycles, infective
biting density and transmission potential of ’forest’ Sim-
ulium damnosum. Ann. Trop. Med. Parasitol. 62: 95-106.
Duke, B.O.L-, P. J.Moore, and J. Anderson. 1972. Studies on
factors influencing the transmission of onchocerciasis.VII. A comparison of the Onchocerca volvulus transmis-
sion potentials of the Simulium damnosum populations in
four Cameroon rain-forest villages, and the pattern ofonchocerciasis associated therewith. Ann. Trop. Med.Parasitol. 66: 219-234.
Duke, B.O.L., J. Anderson, and H. Fuglsang. 1975. The On-chocerca volvulus transmission potentials and associatedpatterns of onchocerciasis at four Cameroon Sudan-sa-vanna villages. Tropenmed. Parasitol. 26: 143-154.
Dye, C. 1994. The epidemiological context of vector con-
trol. In Approaches to vector control: new and trusted.Trans. R. Soc. Trop. Med. Hyg. 88: 147-149.
Elliot, J. M. 1977. Some methods for the statistical analysisof samples of benthic invertebrates, 2nd ed. Freshwater

756 JOURNAL OF MEDICAL ENTOMOLOGY Vol. 35.
Biological Association, Scientific Publication 25. Titus
Wilson, Cumbria, England.ranns R 1975 Observations filanal infections and par-
ous’rates ofanthropophilic blackflies in Guatemala, with
reference to the transmission of Onclwcerca volvulus.
Tropenmed. Parasitol. 26: 169-182.
1985. Morphological differentiation from Onchocerca vol-vulus of first stage larvae of unknown Blarial speciescommonly found in Simulium sanctipauli S.I. in Liberia.Trop. Med. Parasitol. 36: 255-236.
1987. Infection rates and parasitic loads of Onclwcerca vol-vulus, and other filariae, in Simuliiim sanctipauli S.I. and S.yahense in a rain-forest area of Liberia. Trop. Med. Fara-sitol. 38: 201-204.
Garms, R., and R. A. Cheke. 1985. Infections with On-chocerca volvulus in different members of the Simuliumdamnosum complex in Togo and Benin. Z. Angew. Zool.72: 479-495.
Garms, R., and J. 0. Ochoa. 1979. Further studies on therelative importance of Guatemalan blackfly species asvectors of Onchocerca volvulus. Tropenmed. Parasitol. 30:120-128.
Gerrard, D. J., and H. C. Chiang. 1970. Density estimationof corn rootworm egg populations based upon frequencyof occurrence. Ecology 51: 237-245.
Gleason, H. A. 1920. Some applications of the quadratmethod. Bull. Torr. Bot. Club 47: 21-33.
Grenfell, B. T., P. K. Das, P. K. Rajagopalan, and D.A.P.Bundy. 1990. Frequency distribution of lymphatic filariasis microfilariae in human populations; population pro-
and statistical estimation. Parasitologv 101: 417-427.
Katholi, C. R., L. Toe, A. Merriweather, and T. R. Unnasch.1995. Determining the prevalence of Onchocerca volvu-lus infection in vector populations by polymerase chainreaction screening of pools of black flies. J. Infect. Dis.172: 1414-1417.
Kono, T., and T. Sugino. 1958. On the estimation of thedensity of rice stem infested by the rice stem borer. Jpn.J. Appl. Entomol. Zool. 2: 184-188 (In Japanese withEnglish summary).
Kuno, E. 1986. Evaluation of statistical precision and designof efficient sampling for the population estimation basedon frequency of occurrence. Res. Popul. Ecol. 28: 305-319.
LudwigJ. A., and J.F.Reynolds. 1988. Statistical ecology. Aprimer on methods and computing. Wiley, New York.
Lwambo, N.J.S., D.A.P. Bundy, and G.F.H. Medley. 1992. Anew approach to morbidity risk assessment in hookwormendemic communities. Epidemic!. Infect. 108: 469-481.
McCrady, M. H. 1915. The numerical interpretation of fer-mentation-tube results. J. Infect. Dis. 17: 183-212.
Meredith, S.E.O., G. Lando, A. A. Gbakima, P. A. Zimmer-man, and T. R. Unnasch. 1991. Onclwcerca volvulus: ap-plication of the polymerase chain reaction to identifica-tion and strain differentiation of the parasite. Exp.Parasitol. 73: 335-344.
Medley, G. F., R. E. Sinden, S. Fleck, P. F. Billingsley, N.Tirawanchai,andM. H. Rodriguez. 1993. Heterogeneityin patterns of malarial oocyst infections in the mosquitovector. Parasitology 106; 441-449.
Mogi, M., W. Choochote, C. Khamboonruang, and P. Suwan-panit. 1990. Applicability of presence-absence and se-
quential sampling for ovitrap surveillance of Aedes(Diptera: Culicidae) in Chiang Mai, northern Thailand.
J. Med. Entomol. 27: 509-514.Nicolas, L., P. Luquiaud, F. Lardeux, and D. R. Mercer. 1996.
A polymerase chain reaction assay to determine infection
f
of Aedes polynesiensis by Wuchereria bancroff-Soc. Trop. Med. Hyg. 90: 136-139. \.
Ochoa, J. 0. 1982. Studies on the anthropophil-species in Guatemala, with special reference ~\imission of onchocerciasis in the southeastern *\area. Jpn. J. Sanit. Zool. 33: 129-138. ^Omar, M. S., and R. Garms. 1975. The fate and mimicrofilariae of a Guatemalan strain of Onct ^^,vulus in Simulium ochraceum and S. metaHicu "^\,role of the buccopharyngeal armature in the dp atl<^of microfilariae. Tropenmed. Parasitol. 26: litt,5] ^Ortega, M., M. Oliver, and A. Rami’rez. 1992. Emde la oncocercosis en el Soconusco, Chiapas. VI p’Scuantitativos de la transmisi6n de Onchocerca ^^.por tres especies de simulidos en una comunidailSendemia. Rev. Fac. Med. UNAM 35: 95-103. S
Oskam, L., G. J. Schoone, C. C. Kroon, R. LujanDavies. 1996. Polymerase chain reaction for rj "IOnchocerca volvulus in pools of blackflies. Trop lu1^’Health 1: 522-527. ^Pagano, M., and K. Gauvreau. 1993. Principles of hitics. Wadsworth, Duxbury, Belmont, CA.
Pielou, D. P. 1960. Contagious distributions in thepean red mite, Panonycus ulmi (Koch) and a meriigrading population densities from a count ofnutleaves. Can. J. Zool. 38: 645-653.
Pitcaim, M. J., L. T. Wilson, R. K. Washino, and E. Rp,kova. 1994. Spatial patterns of Anopheles freebomCulex tarsalis (Diptera: Culicidae) larvae in Calif’’’rice fields. J. Med. Entomol. 31: 545-553. ^Plaisier, A. P., G. J. Oortmarssen, J.D.F. Habbeni.Remme, and E. S. Alley. 1990. ONCHOSIM: Aand computer simulation program for the transniKand control of onchocerciasis. Comput. Methods I?grams Biomed. 31: 43-56.
Porter, C. H., and R. C. Collins. 1984. Descriptive staticfor the larval stages of Onchocerca volvulus in host-sding Simu/ium ochraceum. Am. J. Trop. Med. Hye.252-260.
1988. Seasonality of adult blackflies and Onchocercavulus transmission in Guatemala. Am. J. Trop. Med Ha38: 153-167.
Porter, C. H., R. C. Collins, and A. D. Brandling-Bennei1988. Vector density, parasite prevalence, and transim.sion of Onchocerca volvulus in Guatemala. Am. J. Tin;Med. Hyg. 39: 567-574.
Post, R. J., P. K. Flook, and A. L. Millest. 1993. Methods f,the preservation ofinsects forDNA studies. Biochem.SiiEcol. 21: 85-92.
Renz, A. 1987. Studies on the dynamics of transmissiononchocercia5is in Sudan-savanna area of North Cameroon. III. Infection rates of the Simulium vectors miOnclwcerca volvulus transmission potentials. Ann. TrmMed. Parasitol. 81: 239-252.
Renz, A., P. Wenk, J. Anderson, and H. Fuglsang. 19KStudies the dynamics oftransmission ofonchocerciasiin a Sudan-savanna area of North Cameroon. V. Whalistolerable level of Annual Transmission Potential? Am
Trop. Med. Parasitol. 81: 263-274.Rodriguez-Perez, M. A., and F. Reyes-Villanueva. 19h
Efecto de la ivermectina sobre la transmisi6n de On-chocerca volvulus en el Sur de Mexico. Salud Publ. Mn36:281-290.
Rodriguez-Perez, M. A., M. H. Rodriguez, H. M. MargAPerez, and A. R. Rivas-AlcaIa. 1995. Effect of semiamnultreatments ofivennectin on the prevalence and intensit;of Onchocerca volvulus skin infection, ocular lesions,

rlWoT- --BASANEZ El AL.: MMror
infectivitv of Simulium ochraceum populations in south-
ern Mexico. Am. J. Trop. Med. Hyg. 52: 429-434.
St^el, S. 1956. Nonparametric statistics for the behavioralsciences. International student edition. McGraw-HillKogakusha, Japan.
Hylefors,B.,B.Philippon,andA.Prost. 1978. Transmission
potentials of Onchoarca volvulus and the associated in-
tensity of onchocerciasis in a Sudan-savanna area. Tro-
penmed, Parasitol. 29: 346-354.Toe, L, A. Merriweather, and T. R. Unnasch. 1994. DNA
probe-based classification of Simithum damnosum s.l.-bome and human-derived filarial parasites in the On-chocerciasis Control Program area. Am. J. Trop. Med.Hyg. 51: 676-683.
Wada, Y. 1982. Theoretical approach to the epidemiology ofonchocerciasis in Guatemala. Jpn. J. Med. Sci. Biol. 35:
183-196.Walsh, J. F., J. B. Davies, R. Le Berre, and K. Garms. 1978.
Standardization of criteria for assessing the effects ofSimulium control in onchocerciasis control programmes.Trans. R. Soc. Trop. Med. Hyg. 72: 675-676.
Wilson, L. F., and D. J. Gerrard. 1971. A new procedure forrapidly estimating European pine sawfly (Hymenoptera:Diprionidae) population levels in young pine plantations.Can. Entomol. 103: 1315-1322.
Received/or publication 11 August 1997; accepted 9 March1998.